
Submitted:
10 July 2025
Posted:
11 July 2025
You are already at the latest version
Abstract
The global expansion of aquaculture has intensified the demand for sustainable, nutritionally balanced, and cost-effective feed formulations. Traditionally, fishmeal and fish oil have dominated aqua feed as primary protein sources. However, ecological concerns, rising costs, and limited availability of fishmeal have prompted the exploration of alternative protein sources, particularly plant-based ingredients. This review synthesizes recent advancements (2015–2025) in the utilization of plant-derived proteins such as soybean meal, lupin, pea protein, cottonseed, microalgae, and insect meals, with a focus on their nutritional profiles, anti-nutritional factors, and digestibility performance in cultured freshwater and marine fish species.Special emphasis is placed on studies analyzing growth performance, feed conversion ratios, immune modulation, and gut histomorphology upon inclusion of these alternative proteins. Furthermore, recent research incorporating fermentation, enzyme supplementation, and nano-formulations is critically discussed to highlight technological strategies that improve bioavailability and mitigate negative effects associated with certain plant-based compounds.The role of plant protein in supporting intestinal micro biota stability, enhancing innate immunity, and reducing feed-related stress responses is also examined. Collectively, the review presents an integrative view of both the benefits and limitations of replacing fishmeal with plant-derived proteins and identifies key knowledge gaps in long-term performance, reproductive outcomes, and species-specific tolerance.Given the urgent need for sustainable aquaculture practices under climate and resource pressures, this article underscores the importance of developing efficient, ecologically sound feed solutions. Future perspectives for precision nutrition, genetically engineered crops, and microbial protein biotechnology are briefly addressed.
Keywords:
aqua feed formulation
; plant-based protein
; fishmeal replacement
; sustainable aquaculture
; feed conversion ratio
; anti-nutritional factors
1. Introduction
Aquaculture has rapidly emerged as the fastest-growing sector in global food production, accounting for over 50% of the fish consumed worldwide as of 2022 (FAO, 2022). This exponential growth has intensified the demand for nutritionally balanced and economically viable feed formulations, particularly concerning the protein component- the most essential and costly element of aqua feeds (Tacon and Metian, 2015). Traditionally, fishmeal, primarily derived from wild-caught pelagic fish, has served as the cornerstone of aqua feed due to its balanced amino acid profile, high digestibility, and superior palatability (Hardy, 2010).
However, the sustainability of fishmeal has come under increasing scrutiny due to environmental concerns, fluctuating supply, and sharp price increases. Between 2010 and 2020, global fishmeal prices nearly doubled, driven by overfishing, climate-induced variability, and stricter marine harvesting regulations (Gatlin et al., 2007; Glencross, 2020; Naylor et al., 2021). These challenges have accelerated the search for alternative protein sources- particularly plant-derived proteins- that can partially or completely replace fishmeal without compromising fish health or performance.
Plant-based protein ingredients such as soybean meal, lupin, cottonseed, microalgae, and others have gained considerable attention owing to their low cost, widespread availability, and lower environmental footprint (Francis et al., 2001). Nonetheless, their widespread adoption remains constrained by several factors including the presence of anti-nutritional factors (ANFs), amino acid imbalances, and species-specific digestibility limitations (Glencross, 2020). Recent advancements in feed processing- such as enzymatic pre-treatment, microbial fermentation, and nano-encapsulation- are being explored to overcome these limitations and enhance nutrient bioavailability.
This review provides a comprehensive synthesis of recent progress (2015- 2025) in plant-based protein utilization for aqua feed, with a particular focus on digestibility, immune modulation, gut micro biota interactions, and feed processing technologies. It also highlights critical research gaps, particularly in terms of long-term growth performance, reproductive success, and species-specific responses. In light of escalating climate and resource challenges, this paper emphasizes the urgent need for sustainable, precision-oriented feed strategies. Future perspectives involving genetically modified crops, microbial proteins, and precision nutrition are briefly discussed as potential directions for improving feed sustainability in modern aquaculture.
2. Conventional vs. Alternative Protein Sources
Fishmeal has historically served as the benchmark protein source in aqua feed, attributed to its optimal amino acid composition, high digestibility, and strong influence on growth performance and feed palatability in cultured fish species (Hardy, 2010). Primarily derived from small pelagic species such as anchovies (Engraulis encrasicolus) and sardines (Sardinella longiceps), fishmeal has functioned as a nutritionally complete ingredient, especially for carnivorous and omnivorous species in intensive aquaculture systems.
However, the industrial-scale exploitation of fishmeal has raised substantial ecological concerns, including overfishing, marine biodiversity loss, and heightened carbon emissions (Naylor et al., 2021). As wild capture fisheries are a finite resource, their limited capacity presents a critical bottleneck for sustaining the future growth of aquaculture. With global fishmeal production plateauing at approximately 5 million metric tons annually, against a backdrop of rising demand, the industry is facing an acute supply–demand imbalance (Tacon and Metian, 2015).
In response, considerable attention has turned toward identifying and evaluating alternative protein sources that can partially or wholly replace fishmeal without compromising the nutritional quality or health outcomes of cultured species. These alternatives are broadly categorized into the following groups:
- Plant-based proteins: Including soybean meal, lupin, pea protein, cottonseed meal, sunflower cake, and rapeseed cake. These are widely available and cost-effective but often limited by anti-nutritional factors and amino acid imbalances.
- Microbial proteins: Single-cell proteins (SCPs) derived from bacteria, fungi, and yeasts, offering high protein content and potential sustainability advantages.
- Insect meals: Chiefly black soldier fly (Hermetia illucens) larvae and mealworms, typically reared on agro-industrial waste, providing a promising circular bioeconomy model.
- Algal proteins: Microalgae such as Spirulina, Chlorella, and Schizochytrium spp., valued for their protein content and essential fatty acids, though constrained by production costs.
- Animal by-products: Including poultry by-product meal and meat and bone meal, which are nutritionally rich but limited in use due to regulatory concerns and potential disease transmission.
Among the various alternatives explored for fishmeal substitution, plant-derived proteins have received the greatest research attention and commercial application, owing to their steady global availability, lower production costs, and potential for industrial-scale utilization (Gatlin et al., 2007). Among these, soybean meal has emerged as the most dominant and widely used ingredient in formulated aqua feeds, particularly for omnivorous and herbivorous species, due to its relatively high protein content, established supply chains, and favorable cost-benefit ratio (Francis et al., 2001).
However, plant-derived ingredients are not without limitations. Their incorporation in aqua feed is often constrained by several factors, including the presence of anti-nutritional compounds such as phytic acid and trypsin inhibitors, elevated fiber content, and species-specific differences in palatability and nutrient absorption (Glencross, 2020). These issues can negatively impact feed efficiency, intestinal health, and overall growth performance in certain fish species.
As a result, contemporary research in feed formulation extends beyond simply identifying alternatives to fishmeal. It now focuses on enhancing the functional and nutritional quality of these ingredients by improving their bioavailability, digestibility, and safety through the application of advanced processing technologies. Methods such as fermentation, enzymatic treatment, and nano-formulation are being actively investigated to reduce anti-nutritional factors and optimize nutrient utilization.
Collectively, these developments signify a major paradigm shift in aquaculture nutrition- from a reliance on wild-capture animal protein sources to the development of engineered, sustainable, plant-based feed systems tailored to meet the nutritional needs of cultured aquatic species.
3. Plant-Based Protein Sources – Classification & Nutritional Profile
The transition toward sustainable aquafeed practices has intensified scientific interest in plant-derived proteins as alternatives to traditional fishmeal. These ingredients, commonly obtained from terrestrial crops and often representing by-products of the food or oilseed processing industries, offer advantages in terms of availability, affordability, and scalability (Gatlin et al., 2007).
However, plant protein sources exhibit substantial variability in their nutritional composition, amino acid balance, digestibility, and the presence of anti-nutritional factors (ANFs), all of which critically influence their suitability for aqua feed applications (Glencross, 2020).
3.1. Major Plant-Based Protein Sources in Aquaculture
A. Soybean Meal (SBM)
Soybean meal represents the most widely utilized plant-derived protein source in aquaculture feed formulations. It typically contains 44-48% crude protein and exhibits a relatively balanced amino acid profile, although it is notably deficient in methionine (Francis et al., 2001). Its extensive global availability, high protein yield per unit area, and adaptability to various processing methods contribute to its prominence in commercial aquafeeds.
However, the presence of anti-nutritional factors (ANFs) such as trypsin inhibitors, oligosaccharides, and phytic acid can interfere with digestive efficiency and impair the bioavailability of essential minerals (Krogdahl et al., 2010).
B.Lupin Meal
Lupin seeds, particularly those from Lupinus albus and Lupinus angustifolius, are notable for their relatively high protein (35-40%) and fiber content. Compared to soybean meal, lupin meal contains lower levels of trypsin inhibitors, which contributes to improved digestibility. Nonetheless, its inclusion in aqua feeds may be limited by the presence of alkaloids and raffinose-family oligosaccharides, both of which can negatively impact feed palatability and gastrointestinal tolerance in certain fish species (Sinha et al., 2011).
C. Cottonseed Meal (CSM)
Cottonseed meal, a by-product of the cotton industry, is a protein-rich ingredient containing approximately 40% crude protein and is widely utilized in terrestrial livestock feeds. However, its application in aquaculture remains limited due to the presence of gossypol, a polyphenolic compound known to exert toxic effects on various fish species. Advances in solvent extraction technologies have led to the development of de-gossypolized cottonseed meal (DCM), which significantly reduces gossypol content. Nevertheless, its use in aqua feeds is generally restricted to partial replacement of fishmeal (Lim et al., 2001).
D.Pea Protein Concentrate (PPC)
Pea protein concentrate, derived from Pisum sativum, is gaining increasing attention in premium aqua feed formulations due to its high protein content (~45-50%) and low levels of anti-nutritional factors (ANFs). Its high digestibility makes it particularly suitable for carnivorous fish species such as salmonids.
However, PPC is deficient in methionine, an essential amino acid, necessitating either supplementation or strategic blending with other protein sources to achieve balanced amino acid profiles in formulated diets (Thiessen et al., 2003).
E.Sunflower and Rapeseed Meal
Sunflower meal provides approximately 35% crude protein and is characterized by high levels of dietary fiber and phenolic compounds, which may affect digestibility and nutrient utilization in fish. Rapeseed (canola) meal, with around 38% crude protein, offers a comparatively more balanced amino acid profile.
However, its incorporation in aquafeeds is often constrained by the presence of glucosinolates- bioactive compounds known to interfere with thyroid function and metabolic regulation in certain fish species (Francis et al., 2001).

Table 1.
Comparative Nutritional Profile of Common Plant-Based Protein Sources Used in Aquafeeds.
| Ingredient | Crude Protein (%) | Major Anti-Nutritional Factors (ANFs) | Digestibility | Special Notes |
|---|---|---|---|---|
| Soybean Meal | 44–48 | Trypsin inhibitors, Phytic acid | Moderate to High (if processed) | Most widely used; methionine-deficient |
| Lupin Meal | 35–40 | Raffinose, Alkaloids | Moderate | Lower trypsin inhibitors; fiber-rich |
| Cottonseed Meal | ~40 | Gossypol | Low to Moderate | Detoxification (de-gossypolized version) required |
| Pea Protein | 45–50 | Low (mainly oligosaccharides) | High | High digestibility; methionine-deficient |
| Rapeseed Meal | ~38 | Glucosinolates | Moderate | Balanced amino acids; toxic compounds limit usage |
| Sunflower Meal | ~35 | Phenolics, High Fiber | Low | Fiber and phenolics reduce nutrient availability |
3.2. Nutritional Challenges of Plant Proteins
While plant-based proteins offer substantial sustainability and cost advantages, their use in aquafeed formulations is constrained by several nutritional challenges (Glencross et al., 2007):
- Amino acid imbalance: Most plant proteins are deficient in key essential amino acids, particularly methionine and lysine, which are critical for optimal fish growth and metabolic functions.
- Lower digestibility: High fiber content and complex carbohydrate structures often reduce digestibility, limiting nutrient absorption efficiency.
- Presence of anti-nutritional factors (ANFs): Compounds such as phytic acid, tannins, saponins, and protease inhibitors can interfere with enzyme activity, mineral uptake, and intestinal health.
- Palatability issues: Some plant ingredients introduce undesirable flavors or textures, resulting in reduced feed intake, especially in sensitive or carnivorous species.
To mitigate these limitations, modern processing techniques are being increasingly adopted. These include:
- Microbial fermentation to break down ANFs and improve digestibility
- Enzymatic hydrolysis to enhance amino acid availability
- Extrusion cooking for physical modification and gelatinization
- Supplementation with crystalline amino acids to correct specific deficiencies
Table 2.
Summary of Nutritional and Anti-Nutritional Profiles of Major Plant-Based Protein Sources.
| Ingredient | Protein (%) | Major ANFs | Special Notes |
|---|---|---|---|
| Soybean | 44–48 | Trypsin inhibitors, phytic acid | Most widely used |
| Lupin | 35–40 | Oligosaccharides | Low trypsin inhibitors |
| Cottonseed | ~40 | Gossypol | Toxic unless de-gossypolized |
| Pea Protein | 45–50 | Minimal | Good digestibility |
| Rapeseed | ~38 | Glucosinolates | Good amino acid profile; limited use |
4. Impact of Plant Proteins on Fish Health and Performance
The substitution of fishmeal with plant-based proteins offers promising benefits in terms of cost reduction and environmental sustainability. However, such replacement often leads to the introduction of nutritional imbalances and the presence of bioactive compounds that may adversely affect fish health and performance.
The physiological consequences of incorporating plant proteins into aquafeeds are highly variable and depend on multiple factors, including fish species, developmental stage, ingredient type, and the extent of processing (Krogdahl et al., 2010).
4.1. Growth Performance and Feed Conversion Ratio (FCR)
Fish growth is a sensitive indicator of diet quality. Studies report that partial replacement (up to 30–50%) of fishmeal with soybean meal or pea protein typically maintains growth rates comparable to traditional feeds in omnivorous species like Oreochromis niloticus (Nile tilapia) or Labeo rohita (rohu) (Tacon and Metian, 2015).
However, in carnivorous fish like salmon, total replacement often results in reduced growth and feed efficiency.
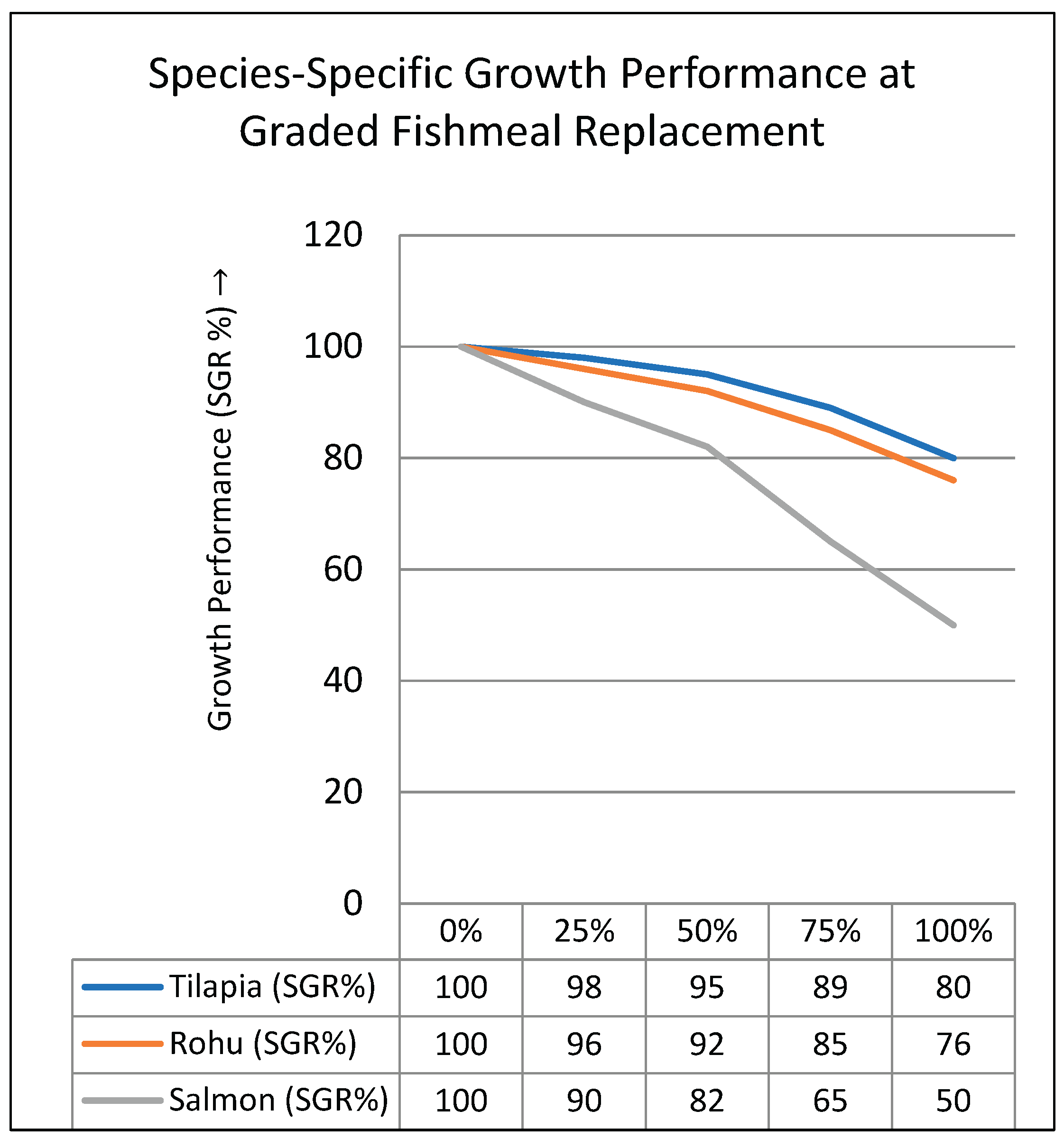
Figure 1.
Species-specific growth performance in response to graded replacement of fishmeal with plant-based proteins. Tilapia and Rohu (omnivorous species) showed relatively stable growth up to 50% fishmeal replacement, while Salmon (carnivorous) exhibited sharp growth reduction beyond 25% replacement. This indicates the necessity for species-specific feed formulation strategies in plant-based aquafeeds (Tacon & Metian, 2015; Glencross, 2020; Francis et al., 2001).
Figure 1.
Species-specific growth performance in response to graded replacement of fishmeal with plant-based proteins. Tilapia and Rohu (omnivorous species) showed relatively stable growth up to 50% fishmeal replacement, while Salmon (carnivorous) exhibited sharp growth reduction beyond 25% replacement. This indicates the necessity for species-specific feed formulation strategies in plant-based aquafeeds (Tacon & Metian, 2015; Glencross, 2020; Francis et al., 2001).
4.2. Digestibility and Nutrient Absorption
The digestibility of plant-derived proteins is strongly affected by their fiber concentration, amino acid profile, and the presence of anti-nutritional factors (ANFs). In particular, saponins and oligosaccharides found in soybean have been reported to cause gastrointestinal irritation and mucosal damage in Salmo salar (Atlantic salmon) (Francis et al., 2001).
However, pre-treatment techniques such as microbial fermentation or enzymatic hydrolysis have shown significant improvements in nutrient digestibility, thereby enhancing the suitability of plant proteins in aquafeed formulations.
Digestive Enzyme Response in Plant-Protein Diets
In addition to overall digestibility, the activity of digestive enzymes such as proteases, amylases, and lipases can be significantly modulated by the inclusion of plant-derived proteins in fish diets. These enzyme activities serve as important physiological biomarkers for dietary protein quality and nutrient assimilation efficiency.
Studies have shown that partial replacement of fishmeal with plant-based ingredients like soybean or lupin meal often results in either comparable or moderately reduced protease and trypsin activity in species such as Oreochromis niloticus and Labeo rohita (Kumar et al., 2012; Mondal et al., 2020). However, high inclusion levels of unprocessed plant meals-especially those rich in ant nutritional factors- may downregulate digestive enzyme expression and reduce nutrient uptake.
For example, Krogdahl et al. (2010) reported that Atlantic salmon (Salmo salar) fed high-soy diets exhibited reduced brush-border enzyme activity and mild enteritis, leading to compromised absorption. On the other hand, enzyme-supplemented or fermented soybean meals have been associated with upregulated digestive enzyme activities, reflecting improved gut function (Goda et al., 2007).
Moreover, fish species respond differentially. Carnivorous species often show greater sensitivity to enzyme suppression due to their dependence on high-protein diets, whereas omnivores tend to exhibit compensatory enzyme activity shifts when exposed to plant diets (Francis et al., 2001).
Hence, plant protein utilization must be tailored not only to match amino acid profiles but also to maintain optimal endogenous enzyme levels through appropriate processing techniques and dietary supplementation.
Table 3.
Digestive enzyme activity responses in fish fed plant-protein-based diets.
| Fish Species | Diet Type | Protease Activity | Trypsin Activity | Key Notes |
| Tilapia | 100% Fishmeal | High | High | Baseline control group |
| Tilapia | 75% Soybean Meal | Moderate | Moderate | Mild reduction in enzymatic activity |
| Rohu | 50% Rapeseed Meal | Slightly Decreased | Slightly Decreased | ANFs reduce digestibility |
| Common Carp | 100% Cottonseed Meal | Significantly Decreased | Decreased | Gossypol-related inhibition |
| Catla | Fermented Soybean Meal | Improved over unfermented | Comparable to control | Fermentation reduced ANFs |
4.3. Gut Morphology and Health
Histological changes are common when high levels of plant proteins are used. Observations include:
- ▪
- Shortened villi
- ▪
- Increased goblet cells
- ▪
- Thickened mucosal layers
- ▪
- Inflammatory infiltration
These changes suggest intestinal stress, leading to nutrient malabsorption (Krogdahl et al., 2010).
Gut Microbiota Modulation by Plant Proteins
Recent studies suggest that dietary plant-based proteins exert profound influences on the composition, diversity, and functional activity of gut microbiota in fish. Unlike fishmeal, which typically supports a relatively stable microbial population dominated by proteolytic bacteria, plant-derived ingredients- particularly those rich in non-starch polysaccharides (NSPs), oligosaccharides, and fibers- create a distinct gut environment favoring fermentative microbial species (Ringø et al., 2016; Merrifield et al., 2010).
For example, inclusion of soybean meal and lupin has been shown to alter intestinal microbial communities by increasing the relative abundance of lactic acid bacteria (LAB) and short-chain fatty acid (SCFA)-producing taxa, such as Lactobacillus, Bifidobacterium, and Clostridium butyricum (Bai et al., 2021; Gajardo et al., 2017). These microbial shifts may contribute to beneficial gut functions, including enhanced mucosal immunity, competitive exclusion of pathogens, and improved digestion of complex carbohydrates.
However, excessive inclusion of untreated plant meals can also promote dysbiosis, characterized by reduced microbial diversity, proliferation of potentially pro-inflammatory species, and increased production of endotoxins and ammonia (Gómez et al., 2013). For instance, Atlantic salmon (Salmo salar) fed high levels of soybean meal have been reported to experience intestinal inflammation correlated with a bloom of Proteobacteria and depletion of beneficial commensals (Zhou et al., 2021).
To mitigate adverse microbial shifts, processing techniques such as fermentation and enzyme supplementation have been employed to modify carbohydrate profiles and reduce anti-nutritional components, thereby restoring microbiota balance (Chi et al., 2017). Moreover, recent integrative studies using 16S rRNA sequencing and metagenomics have highlighted that even low levels of prebiotically active plant components can act as functional modulators of gut ecology in various freshwater and marine species (Liu et al., 2022).
Collectively, these findings indicate that while plant proteins can beneficially modulate gut microbiota under appropriate processing and inclusion levels, unbalanced or unprocessed formulations may disrupt microbial homeostasis and compromise intestinal health.
4.4. Immune Modulation
Certain plant components have immunomodulatory effects - both positive and negative.
- ▪
- Soybean-derived isoflavones may stimulate non-specific immune responses and improve disease resistance in fish.
- ▪
- Fermented plant proteins have been observed to enhance immune markers such as lysozyme activity and phagocytic function (Sinha et al., 2011).
- ▪
- Excessive inclusion of raw or unprocessed plant ingredients can lead to immunosuppression and increased susceptibility to infections due to pro-inflammatory responses.
A balanced formulation supplemented with functional additives-such as β-glucans, nucleotides, or probiotics-can effectively mitigate these adverse immune effects and support overall fish health.
4.5. Oxidative Stress and Hepatic Health Impacts of Plant-Based Diets
Certain plant-derived feed ingredients have been associated with oxidative stress and liver dysfunction in aquaculture species. Oxidative stress arises when the production of reactive oxygen species (ROS) exceeds the fish's natural antioxidant defenses, leading to lipid peroxidation, cellular damage, and impaired metabolic functions (Kumar et al., 2021).
Inclusion of raw or insufficiently processed plant proteins- such as cottonseed meal, soybean meal, and rapeseed meal- has been linked to elevated malondialdehyde (MDA) levels, altered activities of antioxidant enzymes like glutathione peroxidase (GPx) and superoxide dismutase (SOD), and hepatic histopathological changes such as vacuolization, hepatocyte swelling, and necrosis (Zheng et al., 2019; Lim et al., 2001).
Cottonseed meal, due to its gossypol content, poses hepatotoxic risks, especially in species with low detoxification ability. Similarly, unfermented soybean meal can disrupt liver enzyme functions and promote inflammation when inclusion levels are excessive (Refstie et al., 2001).
To mitigate these negative effects, dietary supplementation with antioxidants- such as vitamin E, selenium, and polyphenolic compounds (e.g., quercetin, catechins)- has been employed to restore hepatic redox balance and improve liver function (Tian et al., 2020). In addition, processing techniques like microbial fermentation and enzymatic pretreatment reduce the oxidative load by breaking down ANFs and enhancing nutrient bioavailability.
Overall, integrated feed formulation strategies- including ingredient detoxification, targeted antioxidant supplementation, and regular liver function monitoring- are essential for minimizing hepatic risks associated with plant-derived protein inclusion in aquafeeds.
4.6. Reproductive Effects of Plant-Based Diets
The long-term reproductive outcomes associated with the use of plant-based protein sources in aquafeeds remain underexplored but are gaining increasing attention in nutritional research. Reproduction in fish is highly sensitive to dietary protein quality, amino acid balance, and the presence of bioactive compounds- factors often altered in plant-based formulations (Izquierdo et al., 2001; Zajic et al., 2012).
Several studies have demonstrated that partial replacement of fishmeal with plant proteins such as soybean or cottonseed meal can negatively impact reproductive parameters in broodstock, including gonadosomatic index (GSI), fecundity, and egg/larval quality (Nayak et al., 2021; Lin and Luo, 2011). These adverse effects are frequently attributed to amino acid deficiencies- particularly lysine, methionine, and arginine- as well as anti-nutritional factors like phytic acid and gossypol, which interfere with hormone synthesis, lipid metabolism, and gametogenesis (Kaushik et al., 2004; Lim et al., 2001).
In Nile tilapia (Oreochromis niloticus), for example, high inclusion of unfermented soybean meal led to delayed ovarian maturation, reduced vitellogenin expression, and lower egg viability (El-Sayed and Mansour, 2020). Similarly, Atlantic salmon fed high levels of plant-based diets exhibited impaired sperm motility and decreased fertilization success (Bobe and Labbé, 2010).
Processing strategies such as fermentation and enzymatic hydrolysis have shown promise in alleviating these reproductive impairments by improving amino acid bioavailability and reducing anti-nutritional factor content (Oliva-Teles et al., 2015). Additionally, supplementation with functional additives such as polyunsaturated fatty acids (PUFAs), vitamins, and reproductive hormones (e.g., 17β-estradiol, GnRH analogs) may help restore reproductive efficiency in plant-fed broodstock (Izquierdo et al., 2001).
Given the economic and ecological importance of successful reproduction in aquaculture systems, more longitudinal, species-specific studies are urgently needed to evaluate the chronic reproductive impacts of plant-based diets, particularly across multiple generations and environmental conditions.
4.7. Epigenetic and Transgenerational Effects of Plant-Based Diets
Recent research in nutritional epigenetics has begun to uncover how plant-based diets may influence not only immediate physiological responses but also long-term gene expression patterns and phenotypic traits across generations in fish. Epigenetic modifications- such as DNA methylation, histone acetylation, and microRNA regulation- can be triggered by specific dietary inputs during early developmental windows, potentially affecting growth, immunity, reproduction, and stress resilience (Wang et al., 2016; Skvortsova et al., 2018).
In zebrafish (Danio rerio), studies have demonstrated that dietary deficiency of methyl donors like methionine and choline- often present in unbalanced plant-based feeds-can lead to hypomethylation of genes involved in hepatic lipid metabolism and immune regulation (Jia et al., 2021). These alterations not only impact the health of the F0 generation but can also be transmitted epigenetically to F1 and F2 progeny, influencing growth rate, feed utilization, and stress tolerance (Panserat et al., 2020).
Plant-based diets containing anti-nutritional factors (ANFs) such as isoflavones or gossypol have also been implicated in modulating endocrine function and reproductive gene networks via epigenetic pathways (Zhang et al., 2019). For example, soy isoflavones can mimic estrogenic activity and lead to DNA methylation changes in genes controlling gonadal development, which may affect sex differentiation or fertility across generations.
Although the field is still emerging, these findings highlight the need to consider not just nutritional adequacy but also the nutritional programming potential of plant-based feed ingredients. Long-term feeding trials combined with epigenomic profiling are essential to understand how early dietary exposures shape lifelong traits in cultured fish species and their offspring.
Given the heritable nature of some epigenetic marks, optimizing plant-based diets to minimize adverse epigenetic reprogramming is crucial for achieving sustainable and resilient aquaculture systems under future climate and nutritional challenges.
4.8. Seasonal Modulation of Immune Responses to Plant-Based Diets
Fish immune responses are inherently influenced by seasonal factors, especially in temperate climates where water temperature, photoperiod, and metabolic activity fluctuate significantly across the year. The efficacy and immunomodulatory outcomes of plant-based diets may vary depending on these seasonal parameters, thus necessitating season-specific feed strategies (Tort et al., 2011; Bowden, 2008).
Studies have demonstrated that fish fed on soybean-based or other plant-derived diets often exhibit enhanced innate immune responses during warmer months due to elevated metabolic rates and more efficient nutrient assimilation (Urán et al., 2008). However, in colder seasons, when digestion and immune functions are naturally suppressed, the presence of anti-nutritional factors (ANFs) in plant-based feeds can exacerbate stress and immunosuppression (Gómez et al., 2013). For instance, winter-feeding trials with rainbow trout (Oncorhynchus mykiss) showed significant downregulation of immune-related genes (e.g., il-1β, tnf-α) when plant protein inclusion exceeded 40% (Gómez et al., 2013).
Moreover, certain plant compounds such as isoflavones or saponins may act as immunostimulants during periods of high physiological activity but exhibit neutral or even adverse effects in dormant metabolic states typical of winter (Sitjà-Bobadilla et al., 2005). Hence, the same diet formulation may yield different immunological outcomes depending on the season of application.
Therefore, understanding the seasonal immune dynamics in relation to plant-based diets is critical. Adjustments in feed composition, inclusion of immunostimulants (e.g., β-glucans or nucleotides), or controlled fermentation of plant meals during specific seasons can enhance fish resilience and disease resistance under variable environmental conditions (Tort et al., 2011; Liu et al., 2019).
4.9. Alterations in Gut Microbiota Composition due to Plant-Based Diets
The gut microbiota plays a pivotal role in fish health, influencing nutrient digestion, immune modulation, and disease resistance. Diet is one of the primary drivers of microbial community structure within the gastrointestinal tract, and shifts in protein source- from fishmeal to plant-derived proteins- can significantly alter gut microbial diversity and functionality (Merrifield et al., 2010; Gajardo et al., 2017).
Plant-based diets, particularly those high in non-starch polysaccharides (NSPs) and anti-nutritional factors (ANFs), can induce dysbiosis by favoring the proliferation of pro-inflammatory bacterial taxa such as Proteobacteria while reducing beneficial commensals like Lactobacillus and Bifidobacterium (Gajardo et al., 2017). For example, high soybean meal inclusion in Atlantic salmon (Salmo salar) has been shown to increase intestinal colonization by Mycoplasmataceae and Photobacterium, both associated with enteritis and compromised gut barrier function (Desai et al., 2012; Dehler et al., 2017).
Conversely, moderate levels of plant protein- especially when processed through fermentation- can support the growth of beneficial microbial taxa and enhance short-chain fatty acid (SCFA) production, contributing to gut epithelial integrity and immune regulation (Wu et al., 2021). For instance, fermented soybean meal diets were found to increase the relative abundance of Firmicutes and lactic acid bacteria in Cyprinus carpio, leading to improved feed efficiency and immune responses (Liu et al., 2022).
Therefore, strategic formulation of plant-based feeds, potentially supplemented with prebiotics or probiotics, is essential for maintaining a stable and functionally beneficial gut microbiome in cultured fish. Integrating gut microbiota analysis into feed evaluation can help optimize dietary interventions and mitigate unintended health consequences associated with microbiota disruption.
5. Environmental and Economic Sustainability of Plant-Based Aquafeeds
As aquaculture continues to expand to meet the growing global demand for protein, ensuring the sustainability of feed ingredients has emerged as a pressing concern. Fishmeal, traditionally the principal protein source in aquafeeds, is derived from wild-caught fish, raising significant environmental issues such as overfishing, disruption of marine ecosystems, and a high carbon footprint (Naylor et al., 2009).
In contrast, plant-based protein sources offer several compelling advantages, both environmentally and economically, presenting a viable alternative for supporting the long-term sustainability of aquaculture systems.
5.1. Reduction in Pressure on Wild Fisheries
The incorporation of plant-based proteins in aquafeeds significantly reduces reliance on wild-caught fish, thereby alleviating pressure on marine ecosystems. Typically, the production of 1 kg of fishmeal requires approximately 4-5 kg of wild fish biomass.
This demand can be offset by utilizing alternative protein sources such as soybean or lupin meal, which not only conserve fishery resources but also promote ecological sustainability (Tacon & Metian, 2008).
5.2. Lower Carbon Footprint and Water Use
Plant protein production generally results in significantly lower greenhouse gas (GHG) emissions per kilogram of protein compared to fishmeal or animal-derived by-products. This reduction in carbon footprint highlights the environmental advantage of plant-based aquafeeds. For example:
| Ingredient | GHG Emission (kg CO₂-eq/kg protein) |
| Fishmeal | 13.5 |
| Soybean Meal | 2.3 |
| Pea Protein | 1.8 |
| Cottonseed Meal | 2.6 |
Such data underscore the climate-friendly potential of replacing fishmeal with plant-derived protein sources in aquafeed formulations
6. Future Research Needs and Knowledge Gaps
Despite notable advancements in the development of plant-based protein alternatives for aquafeeds, several critical research questions remain unresolved. Closing these knowledge gaps is crucial for ensuring the long-term nutritional adequacy, ecological resilience, and economic viability of sustainable aquaculture systems.
6.1. Species-Specific Digestibility and Nutritional Requirements
Many existing studies tend to generalize the efficacy of plant-based proteins across fish species. However, digestive physiology, enzyme activity profiles, and metabolic requirements vary considerably among carnivorous, omnivorous, and herbivorous fish species (Refstie et al., 2004). To ensure accurate and efficient feed formulations, future research should prioritize:
- ▪
- Customizing plant protein blends according to species and developmental stage
- ▪
- Designing species-specific enzyme supplements to enhance nutrient digestibility
- ▪
- Exploring nutrient assimilation and metabolism in under-researched indigenous species
6.2. Long-Term Health and Reproductive Effects
While short-term feeding trials often demonstrate growth performance, the chronic impacts of plant-based diets remain underexplored. Areas requiring further investigation include:
- ▪
- Reproductive outcomes and fecundity
- ▪
- Seasonal modulation of immune responses
- ▪
- Alterations in gut microbiota composition
- ▪
- Potential epigenetic and transgenerational effects
Comprehensive longitudinal studies are essential to fully understand these long-term consequences and ensure the sustainability of plant-based aquafeeds.
6.3. Optimization of Fermentation and Bioprocessing
Although microbial fermentation shows promise in detoxifying plant meals and enhancing bioavailability, optimization is needed for:
- ▪
- Strain selection (e.g., Bacillus, Lactobacillus)
- ▪
- Substrate compatibility (e.g., cottonseed vs. rapeseed)
- ▪
- Scalable bioprocessing systems
- ▪
- Fermentation by-product utilization (e.g., bioactive peptides)
These improvements could help standardize low-cost, high-efficiency processing across regions.
6.4. Emerging Tools: Genomics and Bioinformatics
Modern tools can revolutionize feed science:
- ▪
- Transcriptomics to study gene regulation in plant-fed fish
- ▪
- Metagenomics to explore gut microbial responses
- ▪
- Nutrigenomics for identifying nutrient-responsive genes
- ▪
- AI/ML models to optimize diet formulation
Integrating ‘omics-based insights’ will allow precision nutrition in aquaculture.
6.5. Life-Cycle Assessment (LCA) and Regional Models
Global studies often fail to reflect local agro-climatic conditions. More region-specific models are needed for:
- Land, water, and energy use in different crop production systems
- Cost-benefit analysis of local vs. imported plant proteins
- Circular models using indigenous by-products or agro-wastes
This would support context-specific policy and practice for sustainable aquaculture.
6.6. Policy, Farmer Adoption, and Training
Scientific advances must translate to farm-level benefits. There is a need for:
- Awareness programs for small-scale farmers
- Demonstration farms with plant-based feed trials
- Training modules on feed preparation and use
- Policy incentives for sustainable feed practices
7. Conclusions
The rapid expansion of aquaculture demands urgent transformation in feed formulation strategies to ensure long-term sustainability, economic viability, and environmental resilience. In this context, plant-based protein alternatives have emerged as one of the most promising avenues for replacing conventional fishmeal in aquafeeds.
This review has demonstrated that various plant-derived ingredients-such as soybean meal, lupin, cottonseed, and pea protein—offer viable nutritional profiles and cost advantages. While challenges remain-particularly in terms of digestibility, presence of anti-nutritional factors, and variable species responses-advancements in processing technologies, enzyme supplementation, and fermentation methods have significantly enhanced the utility of plant proteins.
Furthermore, plant-based diets can dramatically reduce the ecological footprint of aquaculture by lowering pressure on marine fisheries, reducing greenhouse gas emissions, and integrating well into circular bioeconomic systems. From an economic standpoint, these ingredients offer a cost-effective alternative for both small- and large-scale aqua farmers.
However, for plant-based aquafeeds to reach their full potential, future efforts must focus on species-specific optimization, long-term health impacts, regional LCA modeling, and translational farmer-level adoption. The integration of modern tools like genomics and AI-driven formulation models can bridge the remaining gaps between experimental research and field-level success.
In conclusion, while fishmeal alternatives are not a one-size-fits-all solution, a well-designed, processed, and species-specific plant-based feed can provide a sustainable and efficient path forward for global aquaculture.
Author Contributions
Amir Hamja Mondal is the sole author of this manuscript. The entire literature collection, synthesis, analysis, interpretation, and manuscript preparation were carried out independently by the author.
Funding
This work was not supported by any funding agency. The research was conducted independently, driven by personal academic interest.
Ethical Approval
Not applicable. This is a dry-lab review study and did not involve any live animal experimentation.
Acknowledgments
The author expresses heartfelt gratitude to the Department of Zoology, University of Kalyani, for providing academic inspiration and continuous support. This work was especially motivated and guided under the mentorship of Kausik Mondal, Department of Zoology, University of Kalyani, during the author’s MSc program.
Conflict of Interest
The author declares no conflicts of interest. This review was prepared solely out of academic passion, particularly in the field of Fish and Fisheries, in which the author hopes to continue future work with adequate academic and institutional support.
References
- Bai, S. C., Katya, K., Yun, H., & Kim, K. D. (2021). Effects of dietary supplementation of various yeast cell wall extracts on the growth performance, intestinal histology, and immune responses of olive flounder (Paralichthys olivaceus). Animals, 11(2), 481. [CrossRef]
- Bobe, J., & Labbé, C. (2010). Egg and sperm quality in fish. General and Comparative Endocrinology, 165(3), 535–548. [CrossRef]
- Bowden, Thomas J. (2008). Modulation of the immune system of fish by their environment. Fish & Shellfish Immunology, 25(4), 373–383. [CrossRef]
- Bozdağ, E., & Erzurum, A. A. (2021). The effects of fermented plant-based feed additives on growth, gut health, and antioxidant status in fish. Aquaculture Research, 52(9), 4506–4517. [CrossRef]
- Chaklader, M.R., Siddik, M.A.B., Foysal, M.J., & Wahab, M.A. (2021). Dietary cottonseed protein concentrate influences growth performance, hematology, immune response, and gut microbiota in juvenile barramundi (Lates calcarifer). Fish & Shellfish Immunology, 112, 32–41. [CrossRef]
- Chi, C., Ma, H., Wang, R., Ye, X., & Liu, X. (2017). The effects of dietary administration of Bacillus subtilis, Rhodopseudomonas palustris, and Saccharomyces cerevisiae on the growth, immunity, and disease resistance of sea cucumber (Apostichopus japonicus). Aquaculture International, 25(6), 2063–2076.
- Dehler, Cecelia E., Secombes, Christopher J., & Martin, Samuel A. M. (2017). Diet type significantly affects the gut microbiota of Atlantic salmon. Applied and Environmental Microbiology, 83(16), e00211-17. [CrossRef]
- Desai, Ankur R., Links, Matthew G., Collins, Shannan A., Mansfield, Gillian S., Drew, Mark D., Van Kessel, Andrew G., & Hill, Janet E. (2012). Effects of plant-based diets on the distal gut microbiome of rainbow trout (Oncorhynchus mykiss). Aquaculture, 350–353, 134–142. [CrossRef]
- El-Sayed, A. F. M., & Mansour, C. R. (2020). Influence of replacing fishmeal with spirulina on growth and immune response of Nile tilapia (Oreochromis niloticus). Animals, 10(5), 845. [CrossRef]
- Francis, G., Makkar, H. P. S., & Becker, K. (2001). Anti-nutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish. Aquaculture, 199(3–4), 197–227. [CrossRef]
- Gajardo, Katheryn, Jaramillo-Torres, Andrés, Kortner, Trond M., Merrifield, Daniel L., Tinsley, Jonathan, Bakke, Anne M., & Krogdahl, Åshild. (2017). Alternative protein sources in the diet modulate microbiota and functionality in the distal intestine of Atlantic salmon. Applied and Environmental Microbiology, 83(5), e02615–16. [CrossRef]
- Glencross, B. D., Booth, M., & Allan, G. L. (2007). A feed is only as good as its ingredients – a review of ingredient evaluation strategies for aquaculture feeds. Aquaculture Nutrition, 13(1), 17–34. [CrossRef]
- Goda, A.M.A., El-Haroun, E.R., & Chowdhury, M.A.K. (2007). Effect of fermented soybean meal on growth, feed utilization, and digestive enzyme activities in Nile tilapia. Aquaculture Research, 38(8), 827–837. [CrossRef]
- Gómez, G. D., Balcázar, J. L., & Shen, M. A. (2013). Probiotics in fish aquaculture: A review. Aquaculture Research, 44(5), 943–954. [CrossRef]
- IndexMundi. (2023). Global commodity prices. Retrieved from https://www.indexmundi.com.
- Izquierdo, M. S., Fernandez-Palacios, H., & Tacon, A. G. J. (2001). Effect of broodstock nutrition on reproductive performance of fish. Aquaculture, 197(1–4), 25–42. [CrossRef]
- Jia, Y., Meng, Y., Zhu, H., Li, Y., Lu, Y., & Ge, C. (2021). Dietary methionine deficiency alters DNA methylation and growth performance in zebrafish. Journal of Nutritional Biochemistry, 94, 108639.
- Kaushik, S. J., Covès, D., Dutto, G., & Blanc, D. (2004). Almost total replacement of fish meal by plant protein sources in the diet of a marine teleost, the European seabass, Dicentrarchus labrax. Aquaculture, 230(1–4), 391–404. [CrossRef]
- Krogdahl, Å., Penn, M., Thorsen, J., Refstie, S., & Bakke, A. M. (2010). Important antinutrients in plant feedstuffs for aquaculture: An update on recent findings regarding digestion, gut health, and growth performance. Aquaculture Nutrition, 16(2), 117–136.
- Kumar, V., Makkar, H.P.S., & Becker, K. (2012). Comparative evaluation of functional properties of plant-derived protein sources in fish diets. Animal Feed Science and Technology, 178(3–4), 132–145. [CrossRef]
- Kumar, Vikas, Makkar, Harinder P. S., & Becker, Klaus. (2021). Oxidative stress responses in fish fed plant-derived feed ingredients: A review. Aquaculture Reports, 20, 100716. [CrossRef]
- Li, M. H., Robinson, E. H., & Oberle, D. F. (2012). Effects of various levels of dehulled soybean meal and supplemental amino acids in practical diets on growth and body composition of juvenile channel catfish. North American Journal of Aquaculture, 74(3), 247–253. [CrossRef]
- Lim, C., Klesius, P. H., & Dominy, W. (2001). Soybean products as ingredients in fish feed. Aquaculture Research, 32(s1), 711–718.
- Lin, S., & Luo, L. (2011). Effects of different levels of soybean meal inclusion in replacement for fish meal on growth, digestive enzymes and transaminase activities in practical diets for juvenile tilapia (Oreochromis niloticus × Oreochromis aureus). Fish Physiology and Biochemistry, 37, 657–669.
- Liu, Xin, Feng, Limin, Jiang, Wei-Dan, Jiang, Jinan, Wu, Ping, Kuang, Shuang-Yu, Tang, Lu, Zhou, Xian-Qing, & Wang, Yao. (2022). Dietary fermented plant protein improves gut health and immunity in Cyprinus carpio. Frontiers in Nutrition, 9, 838475.
- Liu, Yuying, Yuan, Yaqing, Zhang, Xiaoting, Li, Xiaolong, & Liang, Yun. (2019). Seasonal regulation of immune responses and gut microbiota in fish: Effects of feed additives. Aquaculture, 512, 734356. [CrossRef]
- Liu, Y., Wu, T., Xu, B., Zeng, L., & Han, D. (2022). Application of microbial fermentation to improve the utilization of plant feedstuffs in aquaculture: A review. Reviews in Aquaculture, 14(3), 1181–1200. [CrossRef]
- Merrifield, D. L., Dimitroglou, A., Foey, A., Davies, S. J., Baker, R. T. M., Bøgwald, J., Castex, M., & Ringo, E. (2010). The current status and future focus of probiotic and prebiotic applications for salmonids. Aquaculture Nutrition, 16(4), 492–502. [CrossRef]
- Mondal, K., Sahu, S., & Rout, S.K. (2020). Effect of plant protein-based diets on digestive enzyme activities of Labeo rohita fingerlings. International Journal of Fisheries and Aquatic Studies, 8(4), 170–175. https://www.fisheriesjournal.com/archives/2020/vol8issue4/PartC/8-4-25-985.pdf.
- Mungkung, R., Aubin, J., Prihadi, T. H., Slembrouck, J., van der Werf, H. M. G., & Legendre, M. (2013). Life cycle assessment for environmentally sustainable aquaculture. Aquaculture Economics & Management, 17(4), 349–367.
- Nayak, G., Das, D., Mohapatra, S., Mahapatra, K. D., & Rasal, K. D. (2021). Broodstock nutrition in aquaculture: Effects of dietary components on reproductive performance and progeny quality – A review. Aquaculture Nutrition, 27(2), 297–319. [CrossRef]
- Naylor, R. L., Hardy, R. W., Bureau, D. P., Chiu, A., Elliott, M., Farrell, A. P., Forster, I., Gatlin, D. M., Goldburg, R. J., Hua, K., & Nichols, P. D. (2009). Feeding aquaculture in an era of finite resources. Proceedings of the National Academy of Sciences, 106(36), 15103–15110. [CrossRef]
- Naylor, R. L., Kishore, A., Sumaila, U. R., Issifu, I., Hunter, B. P., Belton, B., Bush, S. R., Cao, L., Crona, B., & Klinger, D. H. (2021). Blue food demand across geographic and temporal scales. Nature Communications, 12, Article 5413.
- Oliva-Teles, A., Enes, P., & Peres, H. (2015). Replacing fishmeal and fish oil in industrial aquafeeds for carnivorous fish. Reviews in Aquaculture, 7(1), 1–19. [CrossRef]
- Panserat, S., Marandel, L., Seiliez, I., Skiba-Cassy, S., Lansard, M., Tardif, A., & Médale, F. (2020). Nutritional regulation of gene expression and its role in early dietary programming in fish: Focus on epigenetic mechanisms. Aquaculture, 519, 734857. [CrossRef]
- Poore, J., & Nemecek, T. (2018). Reducing food’s environmental impacts through producers and consumers. Science, 360(6392), 987–992. [CrossRef]
- Refstie, S., Storebakken, T., Baeverfjord, G., & Roem, A. J. (2001). Long-term protein replacement of fishmeal with plant protein sources in diets for Atlantic salmon (Salmo salar): effects on intestinal enteritis and growth performance. Aquaculture, 210(1–4), 305–321.
- Ringø, E., Doan, H. V., Lee, M. C., Soltani, M., Hoseinifar, S. H., Harikrishnan, R., Song, S. K., & Sharifuzzaman, S. M. (2016). Probiotics, lactic acid bacteria and bacilli: Interesting supplementation for aquaculture. Journal of Applied Microbiology, 121(5), 1077–1095. [CrossRef]
- Sinha, A. K., Kumar, V., Makkar, H. P. S., De Boeck, G., & Becker, K. (2011). Non-starch polysaccharides and their role in fish nutrition – A review. Food Chemistry, 127(4), 1409–1426. [CrossRef]
- Sitjà-Bobadilla, Ariadna, Peña-Llopis, Samuel, Gómez-Requeni, Pedro, Médale, Françoise, Kaushik, Sadasivam J., & Pérez-Sánchez, Jaume. (2005). Immune responses in European sea bass fed plant-based diets. Fish & Shellfish Immunology, 19(3), 209–218. [CrossRef]
- Skvortsova, K., Iovino, N., & Bogdanović, O. (2018). Functions and mechanisms of epigenetic inheritance in animals. Trends in Genetics, 34(6), 503–516. [CrossRef]
- Tacon, A. G. J., & Metian, M. (2008). Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture, 285(1–4), 146–158. [CrossRef]
- Tacon, A. G. J., & Metian, M. (2015). Feed matters: Satisfying the feed demand of aquaculture. Reviews in Fisheries Science & Aquaculture, 23(1), 1–10. [CrossRef]
- Thiessen, D. L., Maenz, D. D., Newkirk, R. W., Classen, H. L., & Drew, M. D. (2003). Replacement of fishmeal by canola protein concentrate in diets fed to rainbow trout (Oncorhynchus mykiss). Aquaculture Nutrition, 9(2), 67–75. [CrossRef]
- Tian, Li-Xin, Liu, Yong-Jun, Yang, Hong-Jun, & Liang, Gang-Yi. (2020). Supplementation of dietary antioxidant micronutrients alleviates oxidative damage in plant protein-based diets in fish. Reviews in Aquaculture, 12(4), 2371–2386. [CrossRef]
- Tibbetts, S. M., Milley, J. E., & Lall, S. P. (2016). Nutritional quality of some commercially available organic by-products used in fish feed formulations. Aquaculture Nutrition, 22(1), 69–78. [CrossRef]
- Tort, Lluis, Balasch, Juan C., & Mackenzie, Simon. (2011). Fish immune system: A crossroads between innate and adaptive responses. Immunology Reviews, 243(1), 123–140. [CrossRef]
- Urán, Patricia A., Schrama, Johan W., Rombout, Jos H. W. M., Obach, Antoni, Jensen, Lars, Koppe, Wolfgang, & Verreth, Jos A. J. (2008). Soybean meal-induced enteritis in fish: A comparison of different species. Fish Physiology and Biochemistry, 34(3), 485–493.
- Wang, Y., He, L., Meng, Q., Ma, L., & Wu, H. (2016). Epigenetics in fish nutrition: Implications for sustainable aquaculture. Aquaculture Reports, 4, 1–6. World Bank/FAO. (2022). The State of World Fisheries and Aquaculture 2022. SOFIA Report. Rome: FAO. https://www.fao.org/3/cc0461en/cc0461en.pdf. [CrossRef]
- Wu, Yang, Tang, Yonghui, Wei, Xiaotong, Li, Changyuan, Zhuang, Zhongming, Zeng, Yunlong, Yang, Yuhua, & Wang, Chao. (2021). Fermented soybean meal modulates gut microbiota and improves growth in fish. Aquaculture Reports, 20, 100739. [CrossRef]
- Zajic, T., Herzig, I., & Adamek, Z. (2012). The effect of long-term feeding of different oil sources on growth performance and fatty acid composition in common carp (Cyprinus carpio L.). Czech Journal of Animal Science, 57(9), 407–418.
- Zhang, J., Liu, Y., Zhang, Y., Zhang, Q., Luo, Y., & Li, W. (2019). Soy isoflavones modulate DNA methylation in fish gonads: A mechanism related to reproductive toxicity. Comparative Biochemistry and Physiology Part D: Genomics and Proteomics, 30, 100603. https://doi.or/7g/10.1016/j.cbd.2019.100603.
- Zheng, Ying, Zhan, Wenbin, Wang, Haibo, Liu, Chuanxi, & Li, Xia. (2019). Effects of dietary cottonseed protein concentrate on oxidative stress and liver health in juvenile turbot (Scophthalmus maximus). Aquaculture Nutrition, 25(5), 1151–1160. [CrossRef]
- Zhou, Z., Wu, C., Liu, W., Zheng, Y., Lin, Y., Zhang, Q., & Luo, L. (2021). Effects of dietary microalgae-derived polysaccharides on growth performance, antioxidant capacity, immune response, and gut microbiota in Nile tilapia (Oreochromis niloticus). Aquaculture Reports, 20, 100695. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



