Submitted:
10 July 2025
Posted:
11 July 2025
You are already at the latest version
Abstract
Proteus mirabilis is a major uropathogen with growing concern over its presence in animal products and the associated zoonotic transmission risks. As a gut commensal in both humans and animals, it is increasingly detected in wild, farm, and companion animals, as well as in animal-derived foods and related environments. This review summarizes current evidence on its distribution across these sources and explores potential transmission routes to humans. Special attention is given to reported genomic similarities and shared antimicrobial resistance patterns between animal and human isolates. The role of P. mirabilis in exacerbating intestinal inflammation further highlights its relevance beyond urinary infections. By revealing the epidemiology, pathogenic traits, and resistance profiles of animal-associated isolates, this review underscores the zoonotic potential of P. mirabilis and emphasizes the need for enhanced surveillance and research from a One Health perspective.
Keywords:
Proteus mirabilis
; animals
; transmission
; antibiotic resistance
; One Health
1. Introduction
Proteus mirabilis, a Gram-negative facultative anaerobic bacterium, belongs to the genus Proteus within the family Morganellaceae. Traditionally classified in the Enterobacteriaceae family, the taxonomy of P. mirabilis was reassessed with advancements in genomic analysis, leading to its reclassification under Morganellaceae by Adeolu et al. in 2024 based on phylogenetic studies [1]. As of December 9, 2024, the LPSN database lists P. mirabilis as a formally recognized species within this family, alongside other species such as Proteus vulgaris, Proteus cibi, and Proteus faecis [2,3]. The genus Proteus includes ten officially named species, with additional unnamed genomic species (genotypes 4, 5, and 6) [2,3].
Morphologically, P. mirabilis displays remarkable features, with a cell size ranging from 1.0 to 3.0 µm in length and 0.4 to 0.8 µm in width. The bacterium is polymorphic, appearing as short rods, spheres, and filaments, and lacks both a capsule and spore structure [4,5]. It is motile due to the presence of flagella and exhibits a characteristic swarming behavior on agar surfaces. This strong motility leads to the formation of concentric rings and visible demarcation lines—known as Dienes’ lines—which are used for strain differentiation [6,7]. The bacterium also possesses pili, enabling adhesion to epithelial cells of plants and fungi [8]. P. mirabilis is a major uropathogen, contributing to more than 40% of catheter-associated urinary tract infections (CAUTIs) cases [9,10,11,12]. Its virulence factors, including urease, flagella, pili, hemolysins, and metalloproteases, play crucial roles in its ability to colonize the host, damage tissues, and evade immune responses [13,14]. Although typically a commensal in the intestinal tract, recent studies have linked P. mirabilis to inflammatory bowel diseases such as Crohn’s disease [15]. It has been shown to exacerbate colitis by disrupting the intestinal mucus barrier and modulating host immune responses [16].
Although the epidemiology and pathogenicity of P. mirabilis have been widely studied, its significance as a zoonotic pathogen remains poorly documented. This review summarizes current findings on the distribution of P. mirabilis in wildlife, farm animals, companion animals, and associated environments and products. We also aim to elucidate the potential transmission routes of this zoonotic pathogen between humans and animals, with a focus on reported genomic similarities and shared antibiotic resistance patterns. By highlighting the zoonotic risks associated with P. mirabilis, this review seeks to provide a basis for future research on its pathogenic mechanisms and its impact on human and animal health.
2. The Detection of P. mirabilis in Animals
As a commensal bacterium in the gastrointestinal tract of both humans and animals, P. mirabilis is widely distributed across diverse animal species, including wildlife, farm animals, and companion animals. This broad host range suggests potential transmission routes between domestic and wild environments, as well as between animals and humans. As shown in the Table 1, the frequent isolation of P. mirabilis from various animal sources underscores its widespread presence.
Wildlife has been identified as a significant reservoir for P. mirabilis, with the bacterium isolated from a wide range of species and their environments, underscoring its zoonotic potential. In undisturbed natural settings, Suárez-Pérez et al. detected P. mirabilis in 36.2% (17/47) of cloacal swabs from Egyptian vulture chicks in the Canary Islands, Spain [17]. Gao et al. isolated a single P. mirabilis strain (KUST-1312) from 1 out of 37 migratory bird fecal samples collected at Dianchi Lake, China, representing a detection rate of 2.7% (1/37) [18]. These findings underscore the potential role of wild birds in the transmission of P. mirabilis. In addition to vertebrate hosts, ectoparasites may also contribute to the transmission of P. mirabilis among wildlife populations. For example, Ergunay et al. reported a high isolation rate 48.5% (17/35) from ticks collected from wild animals in Kenya—including black rhinos, buffaloes, elephants, giraffes, Grevy’s zebras, lions, and leopards—as well as from domestic cattle [19].
In wildlife held in controlled environments such as zoos or farms, P. mirabilis has also been successfully isolated. Liu et al. employed multiplex PCR to detect P. mirabilis in 100 fecal samples from giant pandas in Sichuan, China, reporting an isolation rate of 30% (35/100) [20]. Furthermore, in an investigation of 32 red panda deaths, three positive cases of P. mirabilis were found in kidney, liver, and urine samples, yielding an isolation rate of 9.38% (3/32), in the same center [21]. Lv et al. isolated 53 strains of P. mirabilis from the feces of foxes, raccoons, and minks, as well as from the surrounding soil in farming environments. The isolation rates were 41.51% (22/53) for foxes, 33.96% (18/53) for raccoons, and 24.53% (13/53) for minks [22]. Under controlled laboratory conditions, P. mirabilis has been isolated from various wildlife species. Yu et al. reported its detection in the feces of diarrheal rhesus macaques 9.5% (7/74) and ferrets 30% (4/12) [23]. As an emerging laboratory animal in biomedical research, the tree shrew harbors Proteus species as dominant members of its gut microbiota, with P. mirabilis being the most frequently isolated. Gu et al. reported that P. mirabilis was isolated from 34 out of 36 fecal samples, yielding a prevalence rate of 94.4% (34/36) [24].
Farm animals serve as important reservoirs for P. mirabilis, with the presence of this bacterium raising concerns for both animal health and public safety. As essential sources of food for humans, farm animals, particularly livestock and poultry, act as major reservoirs for this bacterium. Chinnam et al. found a 15.95% (26/163) isolation rate of P. mirabilis from rectal swabs of pigs in Krishna District, Andhra Pradesh, India, highlighting its widespread presence in the digestive systems of livestock [25]. In Zhejiang, China, Qu et al. reported a 5.55% (30/541) isolation rate from pigs across three cities, with the highest contamination found in the Jinhua region (8.91%) [26]. The bacterium’s presence is not limited to the digestive tract, as it has also been detected in boar semen samples from an artificial insemination center in Rome, Italy. Here, P. mirabilis was shown to negatively impact sperm motility, which could influence reproductive success and breeding efficiency [27]. Besides pigs, Chinnam et al. reported that in Krishna District, Andhra Pradesh, India, P. mirabilis was isolated from cloacal swabs of healthy chickens at a rate of 21.36% (47/220), as well as from rectal swabs of cattle and sheep, with isolation rates of 33.33% (20/60) and 31.91% (15/47), respectively [25]. A study conducted in Shandong, China revealed an isolation rate of 7.07% (50/707) in broilers [28], while Ramatla et al. reported a lower isolation rate of 5.4% (26/480) in chicken manure from farms in the Ngaka Modiri Molema district, South Africa [29]. These findings underscore the role of farm animals as key players in the ecology of P. mirabilis and highlight the potential risks to animal health and food safety.
However, the association of P. mirabilis with disease in farm animals presents a more concerning aspect. For example, in Guangxi, China, Ge et al. found an isolation rate of 21.42% (21/98) in fecal and tissue samples from diseased pigs, highlighting the potential pathogenic role of this bacterium in compromised animal health [30]. Similarly, Sun et al. reported a higher isolation rate of 22.5% (18/80) in diarrheal poultry and 23.26% (20/86) in cattle from northeastern China, indicating that P. mirabilis may exacerbate disease in affected animals [31]. In ducks, Algammal et al. found an isolation rate of 14.6% (35/240) in both healthy and diseased individuals in Port-Said Province, Egypt, further suggesting the bacterium’s diverse role in animal health [32].
Companion animals, which have frequent and close interactions with humans, serve as a potential source for the transmission of zoonotic pathogens, including P. mirabilis. This bacterium has been increasingly identified in pets, underlining its significance in the epidemiology of zoonotic diseases. Marques et al. reported that in households with companion animals, P. mirabilis was isolated from the feces of both humans and dogs. The isolation rate in humans was 12.5% (3/24), while in dogs, it was significantly higher at 44.4% (8/18), with no detection in cats [33]. Liu et al. isolated 75 strains of P. mirabilis from fecal samples of both household and stray dogs in Sichuan, China, with an overall isolation rate of 31.12% (75/241). Notably, the isolation rate 36.17% (34/94) in stray dogs was significantly higher than 27.89% (41/147) in household dogs [34]. Interestingly, exotic pets have also been identified as carriers of P. mirabilis. Pathirana et al. isolated the bacterium from pet turtles bought from pet shops and online stores in South Korea, with an isolation rate of 28.8% (15/52) [35].
The involvement of P. mirabilis in diseased companion animals has also been documented. As a well-known urinary tract infection (UTI) pathogen in humans, P. mirabilis similarly contributes to UTIs in dogs and cats. Fonseca et al. identified P. mirabilis in 22.7% (145/637) of canine urine samples, whereas its detection in feline samples was markedly lower at 2.2% (4/171), based on specimens submitted to NationWide Laboratories in the United Kingdom [36]. Similarly, Moyaert et al. found P. mirabilis in 11.0% (48/437) of dog and 1.1% (2/179) of cat urine samples across Europe, while Escherichia coli being the most common pathogen for UTI [37]. In Chiang Mai, Thailand, Amphaiphan et al. detected P. mirabilis in 13.6% (27/198) of dog and 16.7%.
3. Detection of P. mirabilis in Animal-Derived Foods
Animal-derived foods have become a major focus of public health concern due to their contamination with P. mirabilis. The prevalence of P. mirabilis in animal products varies significantly across different countries and regions, reflecting differences in hygiene practices and environmental conditions (as shown in table). In Andhra Pradesh, India, beef samples exhibited the highest contamination rate at 32.73% (17/55), while chicken and pork samples showed relatively lower detection rates of 19.49% (38/195) and 14.38% (23/160), respectively [25]. In Londrina-PR regions, Brazil, chicken meat displayed the contamination rate of chicken meat was the highest at 100% (200/200), that of beef was much lower at 27.8% (100/360) [41]. In Al Qalyubia Governorate, Egypt, chicken and milk samples had contamination rates of 1.51% (1/66) and 3.45% (2/58) [42]. In Ghent, Belgium, Yu et al. reported P. mirabilis in 36.25% (29/80) broiler carcasses [43], while Liu et al. reported a much higher contamination rate of 66% (66/100) in fresh chicken at Heibei, China [44]. A study by Ma et al. in wet market in Chengdu, China, isolated 89 strains of P. mirabilis from 347 samples of chicken, pork, and aquatic products, with an overall contamination rate of 25.65% (89/347). Among these, chicken showed the highest rate at 54.39% (62/114), followed by pork 14.18% (20/141) and aquatic products 7.61% (7/92) [45]. Lan et al. found P. mirabilis in 490 of 579 fresh meat samples (84.63%) from five wet markets in Zhongshan, China, with chicken 78.95% (180/228), duck 67.90% (84/124), and pork 65.61% (149/227) being the most contaminated [46]. These findings indicate that poor hygiene at poultry and meat stalls may result in significant contamination and cross-contamination, particularly affecting poultry meat. Flies associated with animal-derived food also appear to serve as potential vectors of transmission. Zaher et al. detected P. mirabilis from flies collected on pig carcasses [47].
Currently, P. mirabilis has been detected in food derived from farm animals, and its epidemiology in these animals has been well-documented. However, detection of P. mirabilis in game meat from wildlife has not yet been reported. This gap in the research highlights an area for further investigation, as the potential for P. mirabilis to be present in game meat from wildlife and contribute to zoonotic transmission remains unexplored.
4. Genomic Similarity Between Animal- and Human-Derived P. mirabilis
High genomic similarity has been observed between P. mirabilis isolates from hu-mans, animals, and associated products and environments (as shown in the figure). Notably, P. mirabilis isolates from companion animals have been shown to share high genomic similarity with human isolates, with some strain pairs originating from indi-viduals and pets within the same household. Marques et al. collected urine samples from 76 human patients and 107 companion animals with UTIs, and subsequently con-structed a phylogenetic tree based on PFGE genotyping to compare the genetic relat-edness of the isolates [48]. Among 39 clusters, 17 contained both human and animal isolates, with genomic similarity ranging from 80% to 100%. One canine isolate of P. mirabilis showed 100% similarity with a human isolate, while a feline isolate shared over 90% similarity with a human strain [48]. In a separate study, Marques et al. identified a human-dog pair harboring genetically related P. mirabilis strains, exhibiting 82.5% sim-ilarity to the animal-derived clinical strain FMV4938/07, which was isolated from a dog with a urinary tract infection [33]. One fecal P. mirabilis isolate from a dog in a separate household clustered with two human community-acquired UTI isolates, showing 80.9% and 88.9% genomic similarity, respectively [33]. Wang et al. isolated P. mirabilis (CC16012 strain) from a diarrheal dog, which was closely related to the human-derived Crl143 strain from the United States [39]. Pathirana et al. found that the mrpA gene se-quence of P. mirabilis from pet turtles showed 96.4% and 94.9% similarity to human isolates from UTI and respiratory infections, respectively [35]. In experimental animals, Yu et al. isolated P. mirabilis from diarrheal primate feces, which exhibited 99.6% simi-larity to the UTI patient-derived HI4320 strain, demonstrating a high level of genomic similarity [23].
Figure 1.
Transmission routes of Proteus mirabilis between animals, products, environments, and humans. Line for similar genome: Similarity of genomes between isolates from both sources has been reported; Line for similar antimicrobial resistance: Similar patterns of antibiotic resistance have been reported; Line for potential route: Potential connection between sources, but not yet studied.
Figure 1.
Transmission routes of Proteus mirabilis between animals, products, environments, and humans. Line for similar genome: Similarity of genomes between isolates from both sources has been reported; Line for similar antimicrobial resistance: Similar patterns of antibiotic resistance have been reported; Line for potential route: Potential connection between sources, but not yet studied.
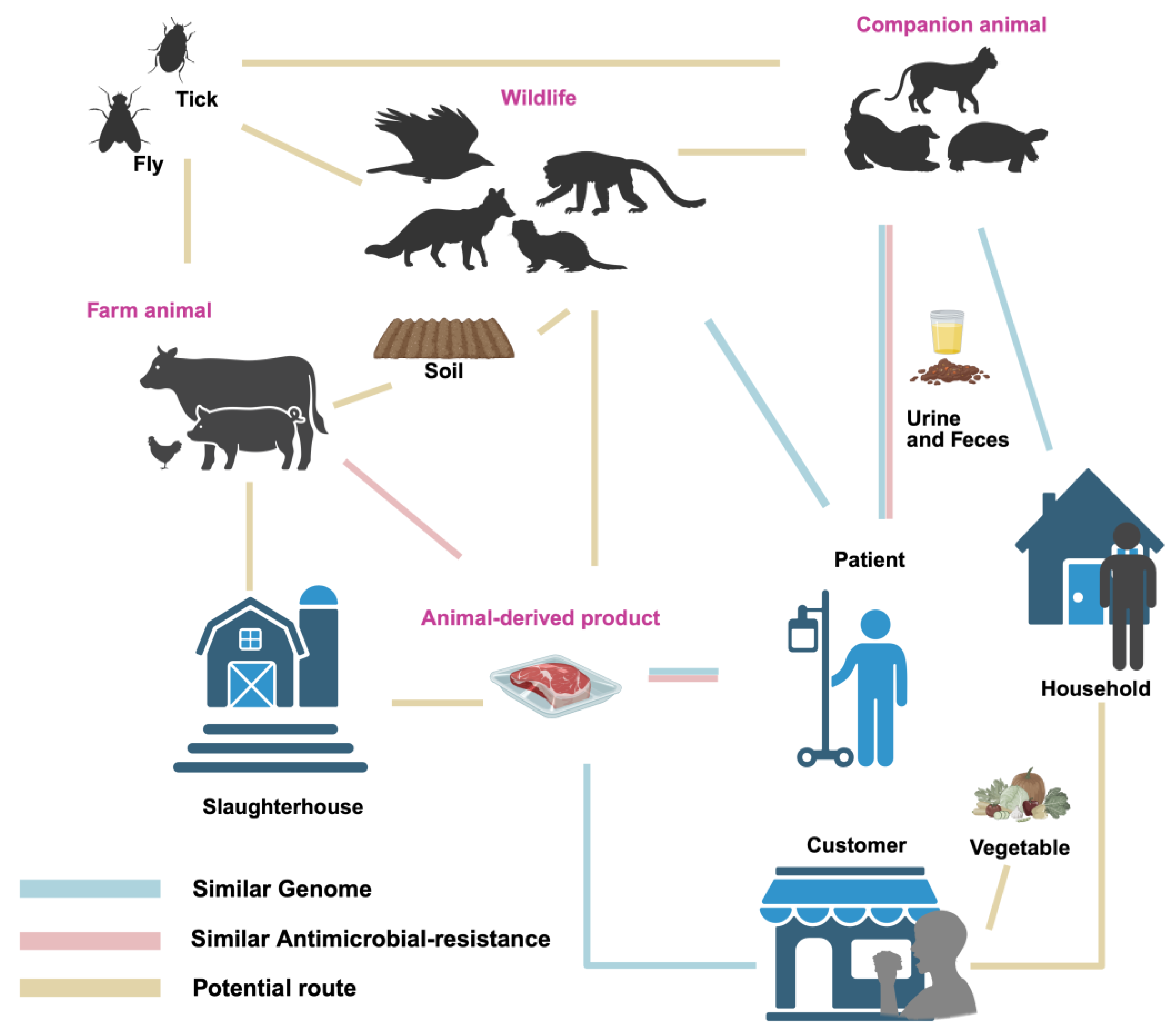
Isolates from animal-derived products have shown high genomic similarity with those from human sources. Yu et al. reported that, based on PFGE analysis, the highest similarity observed between a broiler carcass isolate and a human stool isolate was 83.3%, differing by only two bands [43]. Similarly, Sanches et al. confirmed clonal rela-tionships exclusively between chicken-derived isolates and those causing communi-ty-acquired urinary tract infections (UTI-CA), particularly within cluster C01. Notably, two strains within this cluster—one from chicken meat and the other from a UTI-CA patient—exhibited 100% genomic similarity as determined by PFGE [49]. Furthermore, the blaNDM-1 gene in strain JZ109 showed 100% nucleotide identity (with only a single base difference) to SGI1-1NDM, which was the first reported clinical P. mirabilis strain from China [45].
5. Pathogenicity of P. mirabilis in Humans and Animals
The pathogenicity of P. mirabilis has been reported in various tissues and organs of both humans and animals, with severe cases leading to death. One of the most notable associations of P. mirabilis is as a major pathogen in UTIs in both humans and companion animals [37,50]. Studies indicate that P. mirabilis accounts for 1-10% of all UTI cases in humans [51]. In nearly 3,000 confirmed UTI cases in North America, infections caused by P. mirabilis represented 4% of all cases. Furthermore, the incidence of catheter-associated urinary tract infections (CAUTIs) caused by P. mirabilis is as high as 45% or more [52]. The incidence of P. mirabilis-induced UTI is significantly higher in women and the elderly population [51,52,53]. In animals, P. mirabilis has been linked to recurrent urinary stones in dogs with urinary system disorders, as evidenced by Song et al. [54]. Herout et al. also reported a high infection rate of P. mirabilis in a murine CAUTI model, further highlighting the bacterium’s role in urinary infections across species [55]. These findings highlight the role of P. mirabilis in UTIs in both humans and animals, with important implications for medical treatment and public health.
P. mirabilis has also been implicated in food poisoning incidents. Between 2016 and 2017, 3.61% of reported food poisoning cases in the Datong, China were caused by P. mirabilis, with symptoms such as abdominal pain, diarrhea, nausea, and dizziness [56]. Wang et al. reported a food poisoning incident in Beijing in 2018, where P. mirabilis contamination in braised meatballs led to illness among customers, with the bacterium detected on the hands of the chef and waitstaff [57]. Furthermore, between August and December 2018, Fan et al. isolated P. mirabilis from 49 out of 486 diarrheal pediatric samples, yielding a detection rate of 10.1% [58]. Zhang et al. compared the feces and inflamed colon samples from Crohn’s disease (CD) patients and healthy individuals, finding a significant increase in the abundance of P. mirabilis in CD patients. The experimental results showed signs of colon shortening, and liver and spleen enlargement, indicating that P. mirabilis plays a critical role in inducing CD inflammation [15]. Additionally, Kitamoto et al. suggested that oral inflammation could exacerbate intestinal inflammation, and the use of proton pump inhibitors may promote the proliferation of P. mirabilis and other microbes in the intestines, thus triggering intestinal inflammation [59].
In animals, P. mirabilis has been associated with various cases of gastrointestinal diseases. In 2018, a bamboo rat farm in Guangdong, China reported the deaths of 400 bamboo rats due to vomiting and diarrhea, with P. mirabilis identified as the causative agent [60]. Yu et al. reported similar symptoms in 74 rhesus monkeys infected with P. mirabilis, including diarrhea and bloody stools [23]. In a rabbit farm in Henan, China, P. mirabilis (strain HN001) infection resulted in lethargy, yellow watery diarrhea, and mass fatalities, accompanied by multi-organ tissue damage [61]. In Lhasa, China, a breeding farm experienced deaths of breeding rabbits due to P. mirabilis (strain T2018) infection, presenting with diarrhea and subsequent fatality [62]. Moreover, Dong et al. identified P. mirabilis (strain 17f) as the primary pathogen responsible for diarrhea in lambs in the Hotan area of Xinjiang, China [63]. These reports underscore the significant role of P. mirabilis as a gastrointestinal pathogen in both humans and animals, with implications for public health, food safety, and animal welfare.
In addition to affecting the urinary and gastrointestinal systems, P. mirabilis has been implicated in a wide range of infections across various organs in both animals and humans. In human medicine, Mistry et al. reported the isolation of P. mirabilis from skin abscesses with an isolation rate of 21.6%, second only to methicillin-sensitive Staphylococcus aureus (24.3%) [64]. In pigs, Qin et al. and Chen et al. identified P. mirabilis (GX-PM1 and GX-Y9251 strains) as a cause of respiratory symptoms such as fever and difficulty breathing. They also noted that P. mirabilis could cross the placental barrier, resulting in fetal death [65,66]. Similarly in animals, Li et al. found that P. mirabilis caused multi-organ lesions and systemic inflammation in pigs, which progressed to septicemia and death [67]. In Northern Paraná, Brazil, Sanches et al. isolated P. mirabilis strains (LBUEL-A33 and LBUEL-A34) from broiler chickens, where the bacterium induced caseous exudates and hemorrhaging in the subcutaneous tissue, leading to condemnation in poultry industry. Histopathological analysis revealed edema, congestion, and necrosis in the pectoral muscles, along with cellulitis and infiltration of inflammatory cells [68].
Furthermore, P. mirabilis has been reported to cause severe infections in other animal species. Abdollahi et al. and Ghahremani et al. were the first to document P. mirabilis-induced pyoderma and purulent pericarditis in sheep [69,70]. Sacristán et al. identified P. mirabilis as a significant causative agent of neck abscesses and bacteremia in sea lions [71]. Pattanayak et al. observed P. mirabilis-induced hemorrhaging in the glomeruli and localized necrosis with mononuclear cell infiltration in the kidneys of infected Indian carp [72].
6. Antibiotic Resistance of P. mirabilis from Animals and Animal-Derived Products
Antibiotic resistance in P. mirabilis isolates from animal and food sources has been widely studied due to the growing concerns about public health (Figure & Supplementary Table S1). Among farmed wildlife, Lv et al. reported differences in antibiotic resistance rates of P. mirabilis isolates from foxes, raccoons, and minks. Fox-derived strains showed high resistance to cefotaxime (94.4%), gentamicin (83.3%), and ampicillin (88.9%), significantly higher than those from raccoons and minks. Mink-derived strains showed narrower resistance profiles, maintaining sensitivity to cefotaxime, ceftazidime and ofloxacin. Notably, most isolates were resistant to imipenem (71.70%), a carbapenem [22]. In traditionally farmed livestock, P. mirabilis isolates from pigs, as reported by Ge et al. and Qin et al., were found to be multi-drug resistant (MDR), exhibiting resistance to at least three classes of antibiotics [30,65]. These MDR strains exhibited 100% resistance to tetracyclines, ampicillin, and sulfonamides, with resistance rates for imipenem and kanamycin at 85.7%. Resistance to gentamicin and amikacin was 9.5%, and ciprofloxacin resistance was 38.1%, with fosfomycin resistance at 28.6% [30,65]. Qu et al. found that all 30 P. mirabilis strains isolated from pigs were MDR, with 100% (30/30) resistance to tetracyclines, chloramphenicol, and macrolides. Resistance to ampicillin and amoxicillin/clavulanate reached 90% (27/30), while all are sensetive to meropenem. Among quinolones, ciprofloxacin had the highest resistance 76.67% (23/30), and ofloxacin had the lowest 43.33% (13/30). For aminoglycosides, gentamicin and amikacin resistance rates were 56.67% (17/30) and 20% (6/30), respectively, with rifampicin resistance at 40% (12/30) [26].
In poultry, Li et al. found that all 50 P. mirabilis isolates from meat chickens in Shandong exhibited MDR. Resistance rates to β-lactams (cefazolin, ceftriaxone, and cefuroxime) and aminoglycosides (tobramycin and gentamicin) exceeded 50%, with no resistance to aztreonam. Ciprofloxacin, chloramphenicol, and trimethoprim-sulfamethoxazole resistance rates were all 98% [28]. Ramatla et al. found that 30.7% (8/26) of P. mirabilis strains from broilers exhibited MDR, with ciprofloxacin resistance at 61.5% (16/26), nalidixic acid resistance at 30.8% (8/26), and gentamicin resistance at 38.5% (10/26). Resistance to amoxicillin-clavulanate was 46.2% (12/26), while imipenem and ertapenem resistance was low at 7.7% (2/26) [29]. Moreover, Algammal et al. reported that P. mirabilis isolates from ducks showed 100% (35/35) resistance to amoxicillin, penicillin, trimethoprim-sulfamethoxazole, and doxycycline, with 31.4% (11/35) of the strains classified as extensively drug-resistant (XDR), and 8.6% (3/35) as pan-drug-resistant (PDR) [32]. In the β-lactam class, the resistance rates to piperacillin, amoxicillin-clavulanate, and ampicillin-sulbactam were 77.1% (27/35), while meropenem and imipenem resistance was 8.6% (3/35). Among the quinolones, nalidixic acid resistance reached 62.8% (22/35), while norfloxacin resistance was 8.6% (3/35). Resistance rates to erythromycin and polymyxins were 62.8% (22/35) and 40% (14/35), respectively [32]. Moreover, In the environment, Wang et al. reported the isolation of P. mirabilis carrying the blaNDM-1 gene from the surface of houseflies at a sheep farm in Hubei, China [73]. Similarly, Odetoyin et al. found two Proteus spp. isolates from captured houseflies, both of which were multidrug-resistant and exhibited 100% (2/2) resistance to streptomycin, trimethoprim-sulfamethoxazole, and amoxicillin [74].
For companion animals, Liu et al. isolated 75 strains of P. mirabilis from dog feces in Chengdu, China, including 41 from pet dogs and 33 from stray dogs. Of these, 53.33% (40/75) were MDR, with pet dogs exhibiting a significantly higher proportion of MDR strains (75%) compared to stray dogs (25%). Pet dog strains showed resistance to β-lactam antibiotics, with 31.71% (13/41) resistance to cefotaxime, 36.59% (15/41) to ciprofloxacin, 21.95% (9/41) to gentamicin, and 51.22% (21/41) to chloramphenicol. All strains were resistant to tetracyclines 100% (75/75). In contrast, isolates from stray dogs remained susceptible to these antibiotics [34]. This contrast suggests that the use of antimicrobials in companion animal clinics may play a role in the emergence and exacerbation of antibiotic resistance.
In diseased animals, Marques et al. reported P. mirabilis strains from companion animals with UTIs in Portugal had significant resistance to β-lactams such as ceftazidime 5.6% (6/107), cefotaxime 9.4% (10/107), cefoxitin 9.4% (10/107), and cefovecin 9.4% (10/107), as well as to chloramphenicol 28% (30/107) [48]. Sun et al. reported that 60.22% (106/176) of P. mirabilis strains isolated from diarrheal animals (dogs, minks, cattle, and fowl) from a northeast farm, China, were MDR, with 16.48% (29/176) classified as XDR. High resistance rates were observed to β-lactams like ampicillin 59.09% (104/176), quinolones such as ciprofloxacin 57.39% (101/176), aminoglycosides like streptomycin 55.68% (98/176), tetracyclines such as doxycycline 63.64% (112/176), and tetracycline 55.12% (97/176). imipenem resistance was the lowest at 20.45% (36/176). Additionally, resistance was observed to sulfamethoxazole 43.19% (76/176), nitrofurantoin 42.61% (75/176), and polymyxin B 39.2% (69/176) [31]. El-Tarabili et al. found P. mirabilis isolates from diarrheal dogs in Egypt exhibited 100% (25/25) resistance to β-lactams like penicillin and amoxicillin, and sulfonamides such as trimethoprim-sulfamethoxazole. Resistance to cefotaxime 36% (9/25), ceftazidime 36% (9/25), amoxicillin-clavulanate 32% (8/25), aztreonam 28% (7/25), gentamicin 12% (3/25), nalidixic acid 12% (3/25), and tetracycline 32% (8/25) was also observed [75]. Pattanayak et al. reported that P. mirabilis isolates from diseased carp exhibited MDR and XDR properties, showing resistance to eight classes of antibiotics, including β-lactams (Penicillin G, etc.), tetracyclines (tetracycline), aminoglycosides (tobramycin), sulfonamides, erythromycin, lincomycin, polymyxin, and vancomycin [72].
Since P. mirabilis isolates from farm animals exhibit widespread antibiotic resistance, the presence of resistant strains in animal-derived food poses an undeniable public health concern. Sanches et al. reported high rates of MDR in P. mirabilis strains isolated from chicken 76.5% (153/200), pork 46% (38/83), and beef 6% (6/100). Chicken-derived strains exhibited the highest resistance to most antibiotics, except for chloramphenicol and florfenicol, with resistance levels surpassing those in beef and pork. Pork-derived strains showed higher resistance to quinolones (nalidixic acid, enrofloxacin), sulfamethoxazole-trimethoprim, florfenicol, and chloramphenicol compared to beef-derived strains [49]. In another study, Lan et al. isolated 490 P. mirabilis strains from meat in the Zhongshan market and found that chicken-derived strains had the highest resistance rates to doxycydine 83.33%, β-lactams 82.87%, aminoglycosides 89.81%, sulfonamides 91.67%, and quinolones 54.17%. Pork-derived strains showed resistance only to erythromycin 100%, while duck meat strains exhibited intermediate resistance profiles. Notably, 14.9% of the strains were PDR, highlighting the severe issue of antibiotic misuse [46]. Ma et al. found that 91.01% (81/89) of P. mirabilis strains isolated from meat and aquatic products were MDR, with resistance to β-lactams 83.15%, quinolones 79.78%, chloramphenicol 86.52%, and trimethoprim-sulfamethoxazole 94.38%. Among aminoglycosides, resistance to streptomycin 93.26% was high, while amikacin resistance was only 5.62% [45].
Interestingly, non-animal-derived foods have also shown P. mirabilis resistant strains. Li et al. found that P. mirabilis from vegetables exhibited multi-drug resistance, with resistance to at least four antibiotic classes, including β-lactams (cefotaxime, etc.), quinolones (ciprofloxacin, etc.), aminoglycosides (streptomycin, etc.), and tetracyclines. This suggests the possibility of cross-contamination between animal-derived and other food sources [76].
In addition to genomic similarities, antibiotic resistance profiles of human- and animal-derived P. mirabilis strains also show notable similarities. Marques et al. reported that P. mirabilis strains isolated from companion animals and human with UTIs in Portugal harboured common antimicrobial resistance [48]. Both companion animal and human strains were sensitive to carbapenems like imipenem, meropenem, and ertapenem. Similarly, Yu et al. found higher resistance rates in poultry-derived P. mirabilis strains compared to human patient isolates for quinolones (ciprofloxacin) and penicillins (ampicillin), with poultry strains showing 48% (25/52) resistance to ciprofloxacin and 62% (32/52) resistance to ampicillin, compared to 35% (17/48) and 44% (21/48) in human isolates. Moreover, both groups exhibited two MDR profiles, suggesting the potential for cross-transmission of resistance genes [43]. These findings indicate a risk of resistance transmission between clinical and food sources.
Furthermore, similar antibiotic resistance patterns were observed between isolates from animals and those from animal-derived food. Chinnam et al. found that 72 P. mirabilis strains (31.03%) from 232 animal and food sources were positive for β-lactamase production, including 60 strains confirmed to produce extended-spectrum β-lactamases (ESBLs), which were resistant to ceftazidime and cefotaxime but could be inhibited by β-lactamase inhibitors. About 42% of the sub-clusters contained strains from different hosts, indicating the potential for cross-contamination in slaughterhouse environments [25].
Antibiotic resistance genes (ARGs) contribute significantly to the resistance phenotypes of P. mirabilis isolates and have been identified across various animal sources and related products (see Supplementary Table S1). For β-lactam antibiotics, resistance genes blaOXA-1, blaPSE, blaTEM, blaCTX-M, and blaNDM have been found in isolates from farmed foxes, raccoons, and minks [22]. In farm animals such as pigs, chickens, and ducks, a wider range of resistance genes was detected, including norA, acrB, blaOXA, blaTEM, blaCTX-M, blaNDM, blaDHA, and blaKPC [25,26,28,29,30,32,66]. Isolates from companion animals (dogs and cats) carried blaOXA-1, blaTEM, blaCTX-M, and blaDHA [34,48,75], while foodborne isolates harbored blaCTX-M, blaOXA, blaDHA, blaCMY-2, blaNDM, blaTEM, blaSHV, blaFOX, blaCIT, blaEBC, and bleMBL [25,45,49,76].
For quinolones antibiotics, reistance genes qnrA and qnrC were detected in isolates from farm-rasied wild animals [22], and additional genes such as qnrS, parC, qnrD, and oqxA were reported in farm animal isolates [26,28,30]. qnrA and qnrD were also found in isolates from companion animals [34,48,75], while only qnrD was identified in food-derived isolates [45,49]. Aminoglycosides resistance genes including aac(6′)-Ib-cr, aadA, aadB, aphA6, and aaC2 were identified in farmed wild animal isolates [22]. Farm animals carried aac(6′)-Ib-cr, aph(3)-IIa, rmtB, aaC1, and aaC2[26,28,30,66], while companion animals harbored aphAI-IAB, aac(3′)-IV, aac(6′)-Ib, and aadA1[34,48,75]. In foodborne isolates, aac(6′)-Ib-cr, aph(4)-Ia, aadA1, aadA2, aac(3′)-Ia, aac(3)-IV, and aac(3)-IVa were detected [45,49,76].
Tetracycline resistance genes such as tetO, catI, tet(J), tetA (48), tetA, tetB, tet (C), and tetM were found in isolates from farm animals, companion animals, and food sources, but not in wild animal isolates [26,32,34,45,65,66,75,76]. Sulfonamide resistance genes (sul1, sul2, sul3, dfrIa) and chloramphenicol resistance genes (floR, catB3, cml, cmlA, stcM, cat, cat1, cat2) were widely present in isolates from farm animals (pigs, poultry)[26,28,32,65,66], companion animals (dogs, cats)[48,75], wild animals (foxes, raccoons, minks)[22], and food sources (chicken, pork, beef, vegetables, aquatic products)[45,49,76].
Resistance genes to macrolides, including mphE, ermB, mefA, and mrsD, were only reported in isolates from pigs and chickens [26,28]. The polymyxin resistance gene mcr-1 and glycopeptide resistance gene mecA were exclusively found in chicken-derived isolates [29]. fos and fosA3, related to fosfomycin resistance, were identified in isolates from pigs and chickens, as well as food sources such as chicken, pork, and aquatic products [26,45,49].
Lincosamide resistance genes cfr and lnu(F) were detected by Ma et al. and Li et al. in food sources including chicken, pork, aquatic products, and vegetables. The same teams also identified the tigecycline resistance gene tmexCD3-toprJ1 and the rifampin resistance gene arr-3, respectively [45,76]. In addition, the rifampin resistance gene arr-3 and the disinfectant resistance gene qacH were found in pig-derived isolates [26]. The co-occurrence of arr-3 and qacH may enhance resistance to rifampin and reduce sensitivity to common disinfectants, potentially compromising hygiene measures and increasing the risk of cross-contamination.
7. Perspectives
The emergence and spread of P. mirabilis across diverse animal hosts and food products, coupled with its multidrug resistance and virulence, underscore its growing importance as a zoonotic and foodborne pathogen. The widespread detection of clinically relevant ARGs such as blaNDM, mcr-1, and tmexCD3-toprJ1 in isolates from farm animals, companion animals, wildlife, and animal-derived food highlight the urgent need for integrated surveillance systems. Moreover, the identification of resistance genes in vectors such as houseflies and in contaminated slaughterhouse environments suggests overlooked transmission routes that warrant further investigation.
Moving forward, a One Health approach should be emphasized, integrating data from human, animal, food, and environmental sectors to better understand the ecology and evolution of P. mirabilis. Molecular epidemiology tools such as whole-genome sequencing, resistome and virulome profiling, and comparative genomics will be invaluable for tracking its transmission and adaptation mechanisms. In addition, studies exploring biofilm formation, quorum sensing, and host-pathogen interactions may offer new targets for intervention. As P. mirabilis continues to adapt and spread under antimicrobial selective pressure, coordinated international efforts are needed to mitigate its threat to public health and food safety.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Antibiotic resistance of Proteus mirabilis from animals and animal-derived products.
Author Contributions
Conceptualization, X.L. and Z.Y.; validation, X.L. and Z.Y.; resources, X.L.; data curation, X.L.; writing—original draft preparation, X.L. and Z.Y.; writing—review and editing, Z.Y.; visualization, X.L. and Z.Y.; supervision, Z.Y.; project administration, Z.Y. All authors have read and agreed to the published version of the manuscript.”
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
During the preparation of this manuscript/study, the author(s) used ChatGPT-4o for the purposes of improving readability and language. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| P. mirabilis | Proteus mirabilis |
| NDRJ | multi-drug resistant |
| XDR | extensively drug-resistant |
| PDR | pan-drug-resistant |
References
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-Based Phylogeny and Taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales Ord. Nov. Divided into the Families Enterobacteriaceae, Erwiniaceae Fam. Nov., Pectobacteriaceae Fam. Nov., Yersiniaceae Fam. Nov., Hafniaceae Fam. Nov., Morganellaceae Fam. Nov., and Budviciaceae Fam. Nov. Int J Syst Evol Microbiol. 2016, 66, 5575–5599.
- Liu, H.; Xia, N.; Suksawat, F.; Tengjaroenkul, B.; Hu, Y.; Zhou, X.; Li, X.; Huang, C.; Bao, Y.; Wu, Q.; et al. Prevalence and Characterization of IncQ1α-Mediated Multi-Drug Resistance in Proteus Mirabilis Isolated from Pigs in Kunming, Yunnan, China. Front Microbiol 2024, 15, 1483633. [Google Scholar] [CrossRef]
- Mohr O’hara, C.; Brenner, F.W.; Miller, J.M. Classification, Identification, and Clinical Significance of Proteus, Providencia, and Morganella. Clin Microbiol Rev 2000, 13, 534–546. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chai, J. Research Progress on Proteus mirabilis. Chin Vet Sci 2017, 37, 196–200. [Google Scholar]
- Wang, J.; Zahng, Z.; Wang, S.; Liu,Y. Mixed infection and drug sensitivity analysis of Escherichia coli, Proteus mirabilis and coccidia from rabbits. Acta Vet. Zootech. Sin. 2024, 55, 4204–4212.
- Ali, S.; Karaynir, A.; Salih, H.; Öncü, S.; Bozdoğan, B. Characterization, Genome Analysis and Antibiofilm Efficacy of Lytic Proteus Phages RP6 and RP7 Isolated from University Hospital Sewage. Virus Res 2023, 326, 199049. [Google Scholar] [CrossRef]
- Dienes, L. Reproductive Processes in Proteus Cultures. Proc. Soc. Exp. Biol. 1946, 63, 265–270. [Google Scholar] [CrossRef]
- Cao, Y, Isolation. Identification, and Analysis of Drug Resistance and Virulence Genes of ESBLs-Producing Proteus mirabilis from Fur - Bearing Animals. Master’s Professional Degree Thesis, Shandong Agricultural University, Tai’an, June 2021.
- Cobbley, H.K.; Evans, S.I.; Brown, H.M.F.; Eberhard, B.; Eberhard, N.; Kim, M.; Moe, H.M.; Schaeffer, D.; Sharma, R.; Thompson, D.W.; et al. Complete Genome Sequences of Six Chi-Like Bacteriophages That Infect Proteus and Klebsiella. Microbiol Resour Announc 2022, 11, e01215–21. [Google Scholar] [CrossRef]
- González, M.J.; Navarro, N.; Cruz, E.; Sánchez, S.; Morales, J.O.; Zunino, P.; Robino, L.; Lima, A.; Scavone, P. First Report on the Physicochemical and Proteomic Characterization of Proteus Mirabilis Outer Membrane Vesicles under Urine-Mimicking Growth Conditions: Comparative Analysis with Escherichia Coli. Front Microbiol 2024, 15, 1493859. [Google Scholar] [CrossRef]
- Drzewiecka, D. Significance and Roles of Proteus Spp. Bacteria in Natural Environments. Microb Ecol 2016, 72, 741–758. [Google Scholar] [CrossRef]
- da Silva, J.D.; Bens, L.; Santos, A.J. do C.; Lavigne, R.; Soares, J.; Melo, L.D.R.; Vallino, M.; Dias, R.S.; Drulis-Kawa, Z.; de Paula, S.O.; et al. Isolation and Characterization of the Acadevirus Members BigMira and MidiMira Infecting a Highly Pathogenic Proteus Mirabilis Strain. Microorganisms 2023, 11, 2141.
- Coker, C.; Poore, C.A.; Li, X.; Mobley, H.L.T. Pathogenesis of Proteus Mirabilis Urinary Tract Infection. Microbes Infect 2000, 2, 1497–1505. [Google Scholar] [CrossRef]
- Yang, A.; Tian, Y.; Li, X. Unveiling the Hidden Arsenal: New Insights into Proteus Mirabilis Virulence in UTIs. Front Cell Infect Microbiol 2024, 14, 1465460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hoedt, E.C.; Liu, Q.; Berendsen, E.; Teh, J.J.; Hamilton, A.; O’ Brien, A.W.; Ching, J.Y.L.; Wei, H.; Yang, K.; et al. Elucidation of Proteus Mirabilis as a Key Bacterium in Crohn’s Disease Inflammation. Gastroenterology 2021, 160, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Li, P.; Qiu, K.; Liao, Y.; Chen, X.; Xuan, J.; Wang, F.; Ma, H.; Wang, Y.; Zhu, M. Proteus Mirabilis Exacerbates Ulcerative Colitis by Inhibiting Mucin Production. Front Microbiol 2025, 16, 1556953. [Google Scholar] [CrossRef]
- Suárez-Pérez, A.; Corbera, J.A.; González-Martín, M.; Tejedor-Junco, M.T. Antimicrobial Resistance Patterns of Bacteria Isolated from Chicks of Canarian Egyptian Vultures (Neophron Percnopterus Majorensis): A “One Health” Problem? Comp Immunol Microbiol Infect Dis 2023, 92, 101925. [Google Scholar] [CrossRef]
- Gao, J.; Liu, S.; Bano, S.; Xia, X.; Baloch, Z. First Report of Complete Genome Analysis of Multiple Drug Resistance Proteus Mirabilis KUST-1312 Isolate from Migratory Birds in China: A Public Health Threat. Transbound Emerg Dis 2024, 2024, 8102506. [Google Scholar] [CrossRef]
- Ergunay, K.; Mutinda, M.; Bourke, B.; Justi, S.A.; Caicedo-Quiroga, L.; Kamau, J.; Mutura, S.; Akunda, I.K.; Cook, E.; Gakuya, F.; et al. Metagenomic Investigation of Ticks From Kenyan Wildlife Reveals Diverse Microbial Pathogens and New Country Pathogen Records. Front Microbiol 2022, 13, 932224. [Google Scholar] [CrossRef]
- Liu, S.; Zheng, W.; Huang, H.; Liu, S.; Yang, M.; Yan, X.; Li, Y.; Yue, C.; Hou, R.; Zhang, D. Establishment and application of multiplex PCR detection method for Proteus mirabilis,Klebsiella pneumoniae and Escherichia coli from giant panda. Chin J Prev Vet Med 2023, 45, 598–603. [Google Scholar]
- Liu, S.; Li, Y.; Zhang, D.; Su, X.; Yue, C.; E.Ayala, J.; Yan, X.; Hou, R.; Li, L.; Xie, Y.; et al. Mortality Analysis of Captive Red Panda Cubs within Chengdu, China. BMC Vet Res 2022, 18, 68.
- Lv, P.; Hao, G.; Cao, Y.; Cui, L.; Wang, G.; Sun, S. Detection of Carbapenem Resistance of Proteus Mirabilis Strains Isolated from Foxes, Raccoons and Minks in China. Biology (Basel) 2022, 11, 292. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; He, Z.; Huang, F. Multidrug-Resistant Proteus Mirabilis Isolated from Newly Weaned Infant Rhesus Monkeys and Ferrets. Jundishapur J Microbiol 2015, 8, e16822. [Google Scholar] [CrossRef]
- Gu, W.; Wang, W.; Tong, P.; Liu, C.; Jia, J.; Lu, C.; Han, Y.; Sun, X.; Kuang, D.; Li, N.; et al. Comparative Genomic Analysis of Proteus Spp. Isolated from Tree Shrews Indicated Unexpectedly High Genetic Diversity. PLoS One 2020, 15, e0229125. [Google Scholar] [CrossRef]
- Chinnam, B.K.; Nelapati, S.; Tumati, S.R.; Bobbadi, S.; Peddada, V.C.; Bodempudi, B. Detection of β-Lactamase-Producing Proteus Mirabilis Strains of Animal Origin in Andhra Pradesh, India and Their Genetic Diversity. J Food Prot 2021, 84, 1374–1379. [Google Scholar] [CrossRef]
- Qu, X.; Zhou, J.; Huang, H.; Wang, W.; Xiao, Y.; Tang, B.; Liu, H.; Xu, C.; Xiao, X. Genomic Investigation of Proteus Mirabilis Isolates Recovered From Pig Farms in Zhejiang Province, China. Front Microbiol 2022, 13, 952982. [Google Scholar] [CrossRef] [PubMed]
- Costinar, L.; Herman, V.; Pitoiu, E.; Iancu, I.; Degi, J.; Hulea, A.; Pascu, C. Boar Semen Contamination: Identification of Gram-Negative Bacteria and Antimicrobial Resistance Profile. Animals 2022, 12, 43. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Peng, C.; Zhang, G.; Shen, Y.; Zhang, Y.; Liu, C.; Liu, M.; Wang, F. Prevalence and Characteristics of Multidrug-Resistant Proteus Mirabilis from Broiler Farms in Shandong Province, China. Poult Sci 2022, 101, 101710. [Google Scholar] [CrossRef]
- Ramatla, T.; Ramaili, T.; Lekota, K.; Mileng, K.; Ndou, R.; Mphuthi, M.; Khasapane, N.; Syakalima, M.; Thekisoe, O. Antibiotic Resistance and Virulence Profiles of Proteus Mirabilis Isolated from Broiler Chickens at Abattoir in South Africa. Vet Med Sci 2024, 10, e1371. [Google Scholar] [CrossRef]
- Ge, Q.; Ma, D.; Zhou, Y.; Chen, H.; Yuan, J.; Li, X.; Wang, X. Isolation, identification and characteristics analysis of Proteus mirabilis from swine. China Anim Husb Vet Med 2021, 48, 1804–1815. [Google Scholar]
- Sun, Y.; Wen, S.; Zhao, L.; Xia, Q.; Pan, Y.; Liu, H.; Wei, C.; Chen, H.; Ge, J.; Wang, H. Association among Biofilm Formation, Virulence Gene Expression, and Antibiotic Resistance in Proteus Mirabilis Isolates from Diarrhetic Animals in Northeast China. BMC Vet Res 2020, 16, 1–10. [Google Scholar] [CrossRef]
- Algammal, A.M.; Hashem, H.R.; Alfifi, K.J.; Hetta, H.F.; Sheraba, N.S.; Ramadan, H.; El-Tarabili, R.M. AtpD Gene Sequencing, Multidrug Resistance Traits, Virulence-Determinants, and Antimicrobial Resistance Genes of Emerging XDR and MDR-Proteus Mirabilis. Sci Rep 2021, 11, 9476. [Google Scholar] [CrossRef] [PubMed]
- Marques, C.; Belas, A.; Menezes, J.; da Silva, J.M.; Cavaco-Silva, P.; Trigueiro, G.; Gama, L.T.; Pomba, C. Human and Companion Animal Proteus Mirabilis Sharing. Microbiol Res (Pavia) 2022, 13, 38–48. [Google Scholar] [CrossRef]
- Liu, L.; Dong, Z.; Ai, S.; Chen, S.; Dong, M.; Li, Q.; Zhou, Z.; Liu, H.; Zhong, Z.; Ma, X.; et al. Virulence-Related Factors and Antimicrobial Resistance in Proteus Mirabilis Isolated from Domestic and Stray Dogs. Front Microbiol 2023, 14, 1141418. [Google Scholar] [CrossRef] [PubMed]
- Pathirana, H.N.K.S.; De Silva, B.C.J.; Wimalasena, S.H.M.P.; Hossain, S.; Heo, G.J. Comparison of Virulence Genes in Proteus Species Isolated from Human and Pet Turtle. Iran. J. Vet. Res. 2018, 68, 72–76+154. [Google Scholar]
- Fonseca, J.D.; Mavrides, D.E.; Graham, P.A.; McHugh, T.D. Results of Urinary Bacterial Cultures and Antibiotic Susceptibility Testing of Dogs and Cats in the UK. J. Small Anim. Pract. 2021, 62, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Moyaert, H.; Morrissey, I.; De Jong, A.; El Garch, F.; Klein, U.; Ludwig, C.; Thiry, J.; Youala, M. Antimicrobial Susceptibility Monitoring of Bacterial Pathogens Isolated from Urinary Tract Infections in Dogs and Cats across Europe: ComPath Results. Microb. Drug Resist. 2017, 23, 391–403. [Google Scholar] [CrossRef]
- Amphaiphan, C.; Yano, T.; Som-in, M.; Kungwong, P.; Wongsawan, K.; Pusoonthornthum, R.; Salman, M.D.; Tangtrongsup, S. Antimicrobial Drug Resistance Profile of Isolated Bacteria in Dogs and Cats with Urologic Problems at Chiang Mai University Veterinary Teaching Hospital, Thailand (2012–2016). Zoonoses Public Health 2021, 68, 452–463. [Google Scholar] [CrossRef]
- Wang, D.; Kong, L.; Dong, W.; Liu, S.; Gao, Y.; Ma, H.; Luan, W. Isolation identification and biological characteristics of Proteus mirabilis of canine. Heilongjiang Anim Sci Vet Med 2020, 68, 72–76+154. [Google Scholar]
- Sui, Z. Isolation,Identifi cation and Drug Susceptibility of Proteus mirabilis Isolates from Dogs in Weifang City. Chin. J. Anim. Infect. Dis. 2022, 30, 59–63. [Google Scholar]
- Sanches, M.S.; Rodrigues da Silva, C.; Silva, L.C.; Montini, V.H.; Lopes Barboza, M.G.; Migliorini Guidone, G.H.; Dias de Oliva, B.H.; Nishio, E.K.; Faccin Galhardi, L.C.; Vespero, E.C.; et al. Proteus Mirabilis from Community-Acquired Urinary Tract Infections (UTI-CA) Shares Genetic Similarity and Virulence Factors with Isolates from Chicken, Beef and Pork Meat. Microb Pathog 2021, 158, 105098. [Google Scholar] [CrossRef]
- Edris, S.N.; Hamad, A.; Awad, D.A.B.; Sabeq, I.I. Prevalence, Antibiotic Resistance Patterns, and Biofilm Formation Ability of Enterobacterales Recovered from Food of Animal Origin in Egypt. Vet World 2023, 16, 403–413. [Google Scholar] [CrossRef]
- Yu, Z.; Joossens, M.; Van den Abeele, A.M.; Kerkhof, P.J.; Houf, K. Isolation, Characterization and Antibiotic Resistance of Proteus Mirabilis from Belgian Broiler Carcasses at Retail and Human Stool. Food Microbiol 2021, 96, 103724. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, L.; Li, H.; Liu, N.; Liu, Y.; Du, J. Quantitative PCR detection and antimicrobial resistance analysis of Proteus mirabilis in fresh chicken meat. Farm Prod Process 2022, 24, 63–67+71. [Google Scholar]
- Ma, W.Q.; Han, Y.Y.; Zhou, L.; Peng, W.Q.; Mao, L.Y.; Yang, X.; Wang, Q.; Zhang, T.J.; Wang, H.N.; Lei, C.W. Contamination of Proteus Mirabilis Harbouring Various Clinically Important Antimicrobial Resistance Genes in Retail Meat and Aquatic Products from Food Markets in China. Front Microbiol 2022, 13, 1086800. [Google Scholar] [CrossRef] [PubMed]
- Lan, C.; Hu, Y.; Wang, M.; Li, C.; Bi, S. Prevalence and Antibiotics Resistance of Proteus Mirabilis in Raw Meat. Sci. Technol. Food Ind. 2023, 44, 257–263. [Google Scholar]
- Zaher, A.H.; Kabadaia, M.M.; Hammad, K.M.; Mekky, A.E.; Salem, S.S. Forensic Flies as Carries of Pathogenic Bacteria Associated with a Pig Carcass in Egypt. Al-Azhar Bulletin of Science 2023, 34, 5. [Google Scholar] [CrossRef]
- Marques, C.; Belas, A.; Aboim, C.; Trigueiro, G.; Cavaco-Silva, P.; Gama, L.T.; Pomba, C. Clonal Relatedness of Proteus Mirabilis Strains Causing Urinary Tract Infections in Companion Animals and Humans. Vet Microbiol 2019, 228, 77–82. [Google Scholar] [CrossRef]
- Sanches, M.S.; Silva, L.C.; Silva, C.R. da; Montini, V.H.; Oliva, B.H.D. de; Guidone, G.H.M.; Nogueira, M.C.L.; Menck-Costa, M.F.; Kobayashi, R.K.T.; Vespero, E.C.; et al. Prevalence of Antimicrobial Resistance and Clonal Relationship in ESBL/AmpC-Producing Proteus Mirabilis Isolated from Meat Products and Community-Acquired Urinary Tract Infection (UTI-CA) in Southern Brazil. Antibiotics 2023, 12, 370. [Google Scholar] [CrossRef]
- Costa, T.; Linhares, I.; Ferreira, R.; Neves, J.; Almeida, A. Frequency and Antibiotic Resistance of Bacteria Implicated in Community Urinary Tract Infections in North Aveiro between 2011 and 2014. Microb. Drug Resist. 2018, 24, 493–504. [Google Scholar] [CrossRef]
- Schaffer, J.N.; Pearson, M.M. Proteus Mirabilis and Urinary Tract Infections. Microbiol Spectr 2017, 3, 383–433. [Google Scholar]
- Chakkour, M.; Hammoud, Z.; Farhat, S.; El Roz, A.; Ezzeddine, Z.; Ghssein, G. Overview of Proteus Mirabilis Pathogenicity and Virulence. Insights into the Role of Metals. Front Microbiol 2024, 15, 1383618. [Google Scholar]
- Hafiz, T.A.; Alghamdi, G.S.; Alkudmani, Z.S.; Alyami, A.S.; Almazyed, A.; Alhumaidan, O.S.; Mubaraki, M.A.; Alotaibi, F.E. Multidrug-Resistant Proteus Mirabilis Infections and Clinical Outcome at Tertiary Hospital in Riyadh, Saudi Arabia. Infect Drug Resist 2024, 17, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Ruan, H.; Yang, H.; Mu, H.; Jin, Y.; Wang, Z.; Ge, Y.; Zheng, J. Study on identification, biological characteristic of Proteus mirabilis isolated from canine and its ability to induced calculus formation. Heilongjiang Anim Sci Vet Med 2021, 68, 80–84+163. [Google Scholar]
- Herout, R.; Khoddami, S.; Moskalev, I.; Reicherz, A.; Chew, B.H.; Armbruster, C.E.; Lange, D. Role of Bacterial Surface Components in the Pathogenicity of Proteus Mirabilis in a Murine Model of Catheter-Associated Urinary Tract Infection. Pathogens 2023, 12, 509. [Google Scholar] [CrossRef]
- Gong, Z.; Shi, X.; Bai, F.; He, X.; Zhang, H.; Li, Y.; Wan, Y.; Lin, Y.; Qiu, Y.; Chen, Q.; et al. Characterization of a Novel Diarrheagenic Strain of Proteus Mirabilis Associated With Food Poisoning in China. Front Microbiol 2019, 10, 2810. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, S.; Yu, J.; Zhang, H.; Yuan, Z.; Sun, Y.; Zhang, L.; Zhu, Y.; Song, H. An Outbreak of Proteus Mirabilis Food Poisoning Associated with Eating Stewed Pork Balls in Brown Sauce, Beijing. Food Control 2010, 21, 302–305. [Google Scholar] [CrossRef]
- Fan, C.; Li, X.; Wu, X.; Ni, K. Etiological characteristics and clinical features analysis of 486 pediatric diarrhea cases caused by Proteus mirabilis. Chin J Rural Med Pharm 2021, 28, 52–53. [Google Scholar]
- Kitamoto, S.; Nagao-Kitamoto, H.; Jiao, Y.; Gillilland, M.G.; Hayashi, A.; Imai, J.; Sugihara, K.; Miyoshi, M.; Brazil, J.C.; Kuffa, P.; et al. The Intermucosal Connection between the Mouth and Gut in Commensal Pathobiont-Driven Colitis. Cell 2020, 182, 447–462. [Google Scholar] [CrossRef]
- Zhai, Q.; Zhai, S.; Lv, Y.; Wen, X.; Wu, G.; Huo, W.; Tu, D.; Wei, W.; Jia, C.; Zhou, X. Isolation, Identification and Drug Resistance of Proteus Mirabilis from Bamboo Rats. Chin J Anim Infect Dis 2021, 29, 94–97. [Google Scholar]
- Wang, J.; Li, S.; Zhang, Z.; Wang, S.; Wei, L.; Zhu, X.; Liu,Y. Isolation, identification and pathogenicity analysis of Proteus mirabilis from rabbits. Chin J Prevent Vet Med 2024, 46, 32–38.
- Wang, J.; Xu, J.; Jin, H.; Hou, S.; Feng, J. Isolation,Identification and Drug Resistance Analysis of Rabbit Proteus mirabilis. China Anim Husb Vet Med 2019, 46, 1509–1515. [Google Scholar]
- Dong, G.; Yang, G.; Jiang, X.; Zhang,S.; Wang, Y.; Hr, J.; Tao D. Isolation and Identification of Proteus Mirabilis from Diarrhea Lamb in Xinjiang and Its Pathogenicity in Mice. J Anim Sci Vet Med 2022, 54, 91–95.
- Mistry, R.D.; Scott, H.F.; Alpern, E.R.; Zaoutis, T.E. Prevalence of Proteus Mirabilis in Skin Abscesses of the Axilla. Journal of Pediatrics 2010, 156, 850–851. [Google Scholar] [CrossRef]
- Qin, S.; He, C.; Li, C.; Zeng, W.; Ma, L.; Liu, J.; Bai, A.; Yang, L.; Wu, J. Isolation and Identifi cation of Biofilm-forming Protrus mirabils from Diseased Pigs and Its Pathogenicity. Chin. J. Anim. Infect. Dis. 2023, 31, 113–121. [Google Scholar]
- Chen, H.; ma, L.; Qin, S.; Chen, Y.; Song, R.; Sun, Q.; Qin, S.; Liu, J.; Chen,F.; Wu, J. Isolation,identification and biological characteristics of swine-sourced Proteus mirabilis producing AmpC enzyme. Anim Husb Vet Med 2023, 55, 46–52.
- Li, N.; Wang, B.; Shu, Z.; Zeng, Z.; Huang, T.; Tang, D.; Wu,X. Isolation and identification of swine-sourced Proteus mirabilis, and detection of its drug resistance and virulence genes. J Anim Sci Vet Med 2021, 53, 89–94.
- Sanches, M.S.; Baptista, A.A.S.; de Souza, M.; Menck-Costa, M.F.; Justino, L.; Nishio, E.K.; Oba, A.; Bracarense, A.P.F.R.L.; Rocha, S.P.D. Proteus Mirabilis Causing Cellulitis in Broiler Chickens. Braz. J. Microbiol. 2020, 51, 1353–1362. [Google Scholar] [CrossRef]
- Abdollahi, M.; Javan, A.J.; Shokrpoor, S.; Beidokhtinezhad, M.; Tamai, I.A. Pyoderma Caused by Proteus Mirabilis in Sheep. Vet Med Sci 2022, 8, 2562–2567. [Google Scholar] [CrossRef]
- Najd Ghahremani, A.; Abdollahi, M.; Shokrpoor, S.; Ashrafi Tamai, I. Pericarditis Caused by Proteus Mirabilis in Sheep. Vet Med Sci 2023, 9, 1737–1741. [Google Scholar] [CrossRef]
- Sacristán, C.; Costa-Silva, S.; Reisfeld, L.; Navas-Suárez, P.E.; Ewbank, A.C.; Duarte-Benvenuto, A.; Coelho Couto de Azevedo Fernandes, N.; Albergaria Ressio, R.; Antonelli, M.; Rocha Lorenço, J.; et al. Novel Alphaherpesvirus in a Wild South American Sea Lion (Otaria Byronia) with Pulmonary Tuberculosis. Braz. J. Microbiol. 2021, 52, 2489–2498. [Google Scholar] [CrossRef]
- Pattanayak, S.; Kumar, P.R.; Sahoo, M.K.; Paul, A.; Sahoo, P.K. First Field-Based Evidence of Association of Proteus Mirabilis Causing Large Scale Mortality in Indian Major Carp Farming. Aquaculture 2018, 495, 435–442. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, H.; Chen, G.; Dong, N. Isolation of Four Carbapenem-Resistant Gram-Negative Species from a Single Fly. Animal Diseases 2024, 4, 4. [Google Scholar] [CrossRef]
- Odetoyin, B.; Adeola, ; Babatunde; Olaniran, Olarinde. Frequency and Antimicrobial Resistance Patterns of Bacterial Species Isolated from the Body Surface of the Housefly (Musca Domestica) in Akure, Ondo State, Nigeria. J. Arthropod-Borne Dis. 2020, 14, 88.
- El-Tarabili, R.M.; Ahmed, E.M.; Alharbi, N.K.; Alharbi, M.A.; AlRokban, A.H.; Naguib, D.; Alhag, S.K.; El Feky, T.M.; Ahmed, A.E.; Mahmoud, A.E. Prevalence, Antibiotic Profile, Virulence Determinants, ESBLs, and Non-β-Lactam Encoding Genes of MDR Proteus Spp. Isolated from Infected Dogs. Front Genet 2022, 13, 952689. [Google Scholar] [CrossRef]
- Li, C.A.; Guo, C.H.; Yang, T.Y.; Li, F.Y.; Song, F.J.; Liu, B.T. Whole-Genome Analysis of BlaNDM-Bearing Proteus Mirabilis Isolates and Mcr-1-Positive Escherichia Coli Isolates Carrying BlaNDM from the Same Fresh Vegetables in China. Foods 2023, 12, 492. [Google Scholar] [CrossRef]

Table 1.
Isoalation of Proteus mirabilis from animals and animal-drived foods.
| Category | Host | Region (Year) | Isolation Rate | Virulence Gene | Reference |
| Wildlife | Migratory Birds | China (2024) | 2.7% (1/37) | NA | [18] |
| Canarian Egyptian Vultures | Spain (2022) | 36.2% (17/47) | NA | [17] | |
| Tick from wildlife | Kenya (2022) | 48.5% (17/35) | NA | [19] | |
| Panda | China (2023) | 30% (30/100) | NA | [20] | |
| Red panda | China (2022) | 9.38% (3/32) | NA | [21] | |
| Fox | China (2022) | 41.51% (22/53) | ureC, zapA, pmfA, atfA, mrpA, atfC , hmpA, rsmA, rsbA, ucaA | [22] | |
| Raccoon | China (2022) | 33.96% (18/53) | ureC, zapA, pmfA, atfA, mrpA, atfC , hmpA, rsmA, rsbA, ucaA | [22] | |
| Ferrets | China (2015) | 30% (4/12) | NA | [23] | |
| Mink | China (2020, 2022) | 24.53% (13/53)-28.7% (62/216) | ureC, zapA, pmfA, atfA, mrpA, atfC , hmpA, rsmA, rsbA, ucaA, FliL | [22,31] | |
| Rhesus Monkeys | China (2015) | 9.5% (7/74) | NA | [23] | |
| Tree shrews | China (2020) | 94.4% (34/36) | NA | [24] | |
| Farm animals | Pig | China (2021, 2022), Rome (2021), India (2021) | 5.55% (30/541)-21.43% (21/98) | ureC, hpmA, zapA, pmfA, rsbA, ucaA, mrpA, atfA, ireA , ptA | [25,26,27,30] |
| Broiler | China (2020, 2022), India (2021), South Africa (2024) | 5.4% (26/480)-22.5% (18/80) | ureC, rsmA, hmpA, FliL , ireA, ptA , zapA, ucaA, pmfA, atfA , mrpA, hlyA , hpmA | [25,28,29,31] | |
| Duck | Egypt (2021) | 14.6% (35/240) | atpD, ureC, rsbA , zapA | [32] | |
| Cattle | China (2020), India (2021) | 23.26% (20/86)-33.33% (20/60) | ureC, zapA, rsmA, hmpA, mrpA, atfA, pmfA, FliL, ucaA | [25,31] | |
| Sheep | India (2021) | 31.91% (15/47) | NA | [25] | |
| Companion animals | Dog | China (2020, 2022, 2023), Egypt (2022), UK (2021), Thailand (2019), European countries (2016), Portugal (2018, 2021) | 11.0% (48/437)-87.85% (94/107) | ureC, FliL, ireA, zapA, ptA, hpmA, hpmB, pmfA, rsbA, mrpA, ucaA, rsmA, atfA | [31,33,34,36,37,38,39,40,48,75] |
| Cat | UK (2021), Thailand (2020), Europe (2017), Portugal (2019, 2022) | 0-16.7% (4/24) | hmpA/hmpB, mrpA, pmfA, ucaA | [33,36,37,38,48] | |
| Pet turtle | South Korea (2018) | 28.8% (15/52) | ureC, rsbA, zapA , mrpA | [35] | |
| Animal-derived foods | Pork | China (2022, 2023), Brazil (2021), India (2021) | 14.38% (23/160)- 65.61% (149/227) | mrpA, pmfA, ucaA, atfA, hpmA, zapA, ptA, ireA | [25,41,45,46] |
| Beef | Brazil (2021) | 27.8% (100/360)- 32.73% (17/55) | mrpA, pmfA, ucaA, atfA, hpmA, zapA, ptA, ireA | [25,41] | |
| Mutton | India (2021) | 25.51% (25/98) | NA | [25] | |
| Chicken | China (2022, 2023), Belgium (2020), Brazil (2021), India (2021), Egypt (2023) | 1.51% (1/66)-100% (200/200) | mrpA, pmfA, ucaA, atfA, hpmA, zapA, ptA, ireA | [25,41,42,43,45,46] | |
| Duck meat | China (2023) | 67.9% (84/124) | NA | [46] | |
| Milk/Dairy Products | India (2021), Egypt (2023) | 3.45% (2/58)-22.11% (21/95) | NA | [25,42] | |
| Other source | Aquatic products | China (2022) | 7.61% (7/92) | NA | [45] |
| Vegetables | China (2023) | 62.5% (5/8) | hpmA, mrpA, ptA, ireA, zapA, pmfA , atfA | [76] |
(4/24) of cat urine samples [38]. Beyond UTIs, P. mirabilis has been isolated from companion animals with gastrointestinal symptoms. Wang et al. reported a 19.35% (12/62) isolation rate from fecal samples of dogs with diarrhea in Jilin, China [39]. Furthermore, Sui et al. found P. mirabilis in 47.37% (9/19) of dogs infected with canine parvovirus and 10% (2/20) of dogs suffering from canine distemper [40].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



