
Submitted:
08 July 2025
Posted:
09 July 2025
You are already at the latest version
Abstract
Background: The placenta plays a central role in the metabolic adaptations required for a healthy pregnancy and its dysfunction contributes to Gestational Diabetes Mellitus (GDM) development. We explored the placental metabolic profile, particularly metabolites of one-carbon metabolism, in GDM pregnancies and its association with maternal dietary methyl donor intake and neonatal outcomes.
Methods: Dietary records were obtained from a prospective cohort with pregnant women. Groups were divided into control (n=13) and GDM (n=11). A subset of placental samples collected at delivery (control n=6; GDM n=5) was used for MS and NMR-based metabolomics.
Results: Dietary intake was similar between the groups as well as the sum of dietary methyl donors (choline, betaine and methionine). Importantly, 100% of women presented with inadequate dietary choline. Placentas from pregnancies affected by GDM presented lower levels of choline, serine, glycine and SAM, despite good glycemic control and the absence of adverse neonatal outcomes.
Conclusion: While the placenta demonstrated adaptive mechanisms that possibly mitigated placental stress, the long-term implications of these adaptations on offspring health remain uncertain. Given the critical role of methyl donors in epigenetic regulation and fetal programming, further research is needed to refine dietary guidelines, potentially guiding more tailored nutritional interventions focusing on choline supplementation during pregnancy.
Keywords:
gestational diabetes mellitus
; metabolomics
; one-carbon metabolism
; placental metabolism
1. Introduction
Pregnancy is a pro-diabetogenic state characterized by maternal insulin resistance, which raises blood glucose levels and ensures an adequate supply of nutrients and energy for the fetus. The placenta plays a central role in the metabolic adaptations required for a healthy pregnancy and its dysfunction may contribute to the development of Gestational Diabetes Mellitus (GDM). Also, GDM itself can further disrupt placental function, creating a cycle of metabolic dysregulation [1,2].
GDM is characterized by the inability to compensate for the physiological insulin resistance, leading to a glucose intolerance of varying severity first detected during gestation [3,4]. GDM results in adverse outcomes not only for the mother and child but also for future generations, particularly an increased risk of obesity and metabolic disorders [5,6,7]. In Brazil and globally the prevalence of GDM is 18%, depending on the diagnostic criteria used [8]. Obesity and being overweight are well established risk factors for GDM and their increasing prevalence worldwide represents a significant public health concern [9].
GDM affects the expression of placental genes associated with signaling pathways involved in energy metabolism, oxidative stress and inflammation [10,11,12,13]. These alterations can compromise both the structure and function of the placenta [14]. Our group has shown that placental metabolism is altered in GDM pregnancies, including lipid metabolism, and is associated with mitochondrial and endoplasmic reticulum stress [15,16,17].
One-carbon metabolism is a complex network of interlinked pathways in which single-carbon units are transferred between intermediates. It plays a crucial role in placental development and regulates the resources available for fetal growth [1]. In addition to generating S-adenosyl methionine (SAM), the universal methyl donor, this metabolic network integrates lipid and glucose metabolism and supports essential physiological functions such as cell proliferation, nucleic acid synthesis, redox balance and the establishment of epigenetic marks making it particularly critical during pregnancy [18,19]. The pathway integrates the folate and methionine cycles, as well as the homocysteine transsulfuration pathway. These processes depend on key metabolites mostly supplied by the diet that serve as methyl donors and cofactors, such as folate, vitamins B6 and B12, choline, betaine, and methionine (20; Figure 1).
Figure 1.
Schematic representation of the 1C metabolism pathway which integrates folate and methionine metabolism. This pathway regulates the production of metabolites involved in purine and pyrimidine synthesis, methylation reactions and glutathione synthesis in which cytosolic and mitochondrial enzymes. 5,10-methylene-tethrahydrofolate (5,10-meTHF), 10 formyl-tetrahydrofolate (10-formylTHF), dimethylglycine (DMG); tetrahydrofolate (THF); dihydrofolate (DHF) S-adenosylmethionine (SAM); S- adenosylhomocysteine (SAH). Dietary compounds are shown in red. To include the enzymes.
Figure 1.
Schematic representation of the 1C metabolism pathway which integrates folate and methionine metabolism. This pathway regulates the production of metabolites involved in purine and pyrimidine synthesis, methylation reactions and glutathione synthesis in which cytosolic and mitochondrial enzymes. 5,10-methylene-tethrahydrofolate (5,10-meTHF), 10 formyl-tetrahydrofolate (10-formylTHF), dimethylglycine (DMG); tetrahydrofolate (THF); dihydrofolate (DHF) S-adenosylmethionine (SAM); S- adenosylhomocysteine (SAH). Dietary compounds are shown in red. To include the enzymes.
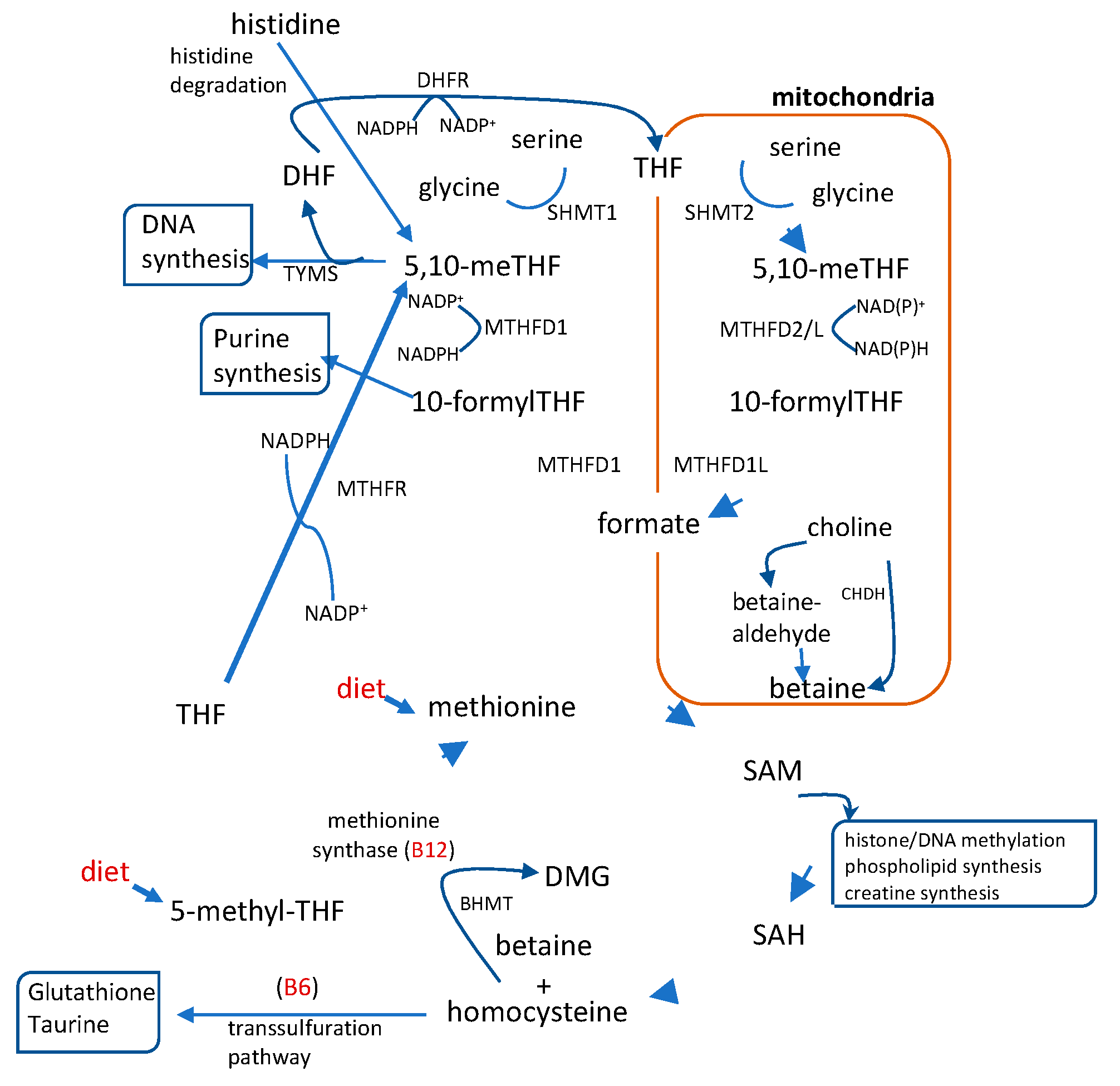
Evidence from both mouse and human studies highlights a connection between the intake of methyl donors during pregnancy, glucose homeostasis and fetal and neonatal outcomes. Supplementation of folic acid to pregnant mice resulted in improved glucose homeostasis in the offspring when exposed to LPS [21]. This effect may be related to the upregulation of genes such as PPARy and IGF2, which play key roles in insulin sensitivity and fetal growth. Conversely, excessive maternal folic acid intake has been shown to disrupt one-carbon metabolism and lipid metabolism in both mother and fetus, contributing to glucose intolerance in mouse models [22,23]. Additionally, supplementation of choline to pregnant mice appears to regulate placental nutrient transport, contributing to adequate fetal growth in both diet-induced obese [24] and lean models [25,26].
Dysregulation of one-carbon metabolism during human pregnancy has been associated with higher maternal BMI and dyslipidemia [27], and with fetal growth restriction [28,29]. Similarly, higher maternal betaine supplementation and serum folate levels were associated with lower IGF2 DNA methylation in cord blood and the placenta, and increased betaine intake was inversely correlated with birthweight [31]. Higher maternal intake of folate and choline has also been linked with lower infant bodyweight at two years [30].
Although the importance of one-carbon metabolism-related nutrients during pregnancy to adequate maternal and neonatal health seems to be well documented, the specific impact of their dietary insufficiency on placental metabolism and function in GDM remains unclear [32]. Therefore, this study aims to explore the placental metabolic profile, with particular emphasis on metabolites of one-carbon metabolism, in GDM pregnancies and its association with maternal dietary methyl donor intake and neonatal outcomes.
2. Results
2.1. Mothers Characteristics
In this study, 13 women with no pregnancy complications (control group) and 11 diagnosed with GDM (GDM group) were enrolled. Maternal age, pre-gestational body mass index (BMI) and total gestational weight gain were all similar between the groups (Table 1). In total, 62 % and 82 % of women were overweight or obese in the control and GDM group, respectively (Table 1). In the GDM group, blood glucose levels were within the normal range (33) and diabetes management was achieved by diet alone (27% of women, n=3) and diet + insulin (73% of women, n=8). This corresponded to a clinically-controlled cohort as fasting and post-prandial glucose levels, measured during the last 2-3 months of pregnancy, were within the normal range [33] (Table 1). According to the charts for weight gain during pregnancy for Brazilian women (34), 46% and 64% of women from control and GDM groups, respectively, had adequate weight gain, with no significant differences between groups, while 31% had excessive weight gain in the control group as opposed to 9% in the GDM group.

Table 1.
Characteristics of the women participating in the study.
| Variables | Median (p25-p75) a or % frequency (n)b | ||
| Control (n=13) | Gestational diabetes mellitus (n=11) | p value | |
| Age (years) a | 28 (23.5-32.5) | 31 (27-37) | 0.26 |
| Pre-gestational BMI (kg/m²) a Normal weight b Overweight b Obesity b |
25.7 (22.5-29.7) 38 (5) 38 (5) 24 (3) |
28.3 (25.1-34.7) 18 (2) 36 (4) 46 (5) |
0.15 |
| Total weight gain (kg) a Adequate b Insufficient b Excessive b |
14.2 (6.9-18.8) 46 (6) 23 (3) 31 (4) |
8.8 (6.6-12.9) 64 (7) 27 (3) 9 (1) |
0.13 |
| Insulin therapy b Blood glucose (mg/dL) a Fasting Post-prandial |
n/a - - |
73 (8) 89 (87.5-97.0) 112 (94.5-123.1) |
n/a n/a n/a |
a Mann–Whitney test; b Fisher exact test. N.A.= not applicable. BMI: Body Mass Index. Classification of weight gain according to Kac et al., 2021.
2.2. Dietary Data
There were no significant differences between the control and GDM groups regarding the intake of energy, total sugar and ultra-processed food. By contrast, carbohydrate, lipid and fiber intakes were higher in the GDM group (Supplementary Material, Table S1). The average intake of energy and proteins was adequate in both groups according to reference values [35]. However, carbohydrate and lipid intake was inadequate in 46 % and 23% of the control group respectively. Concerning fiber, inadequate intake was observed in all women from the control group and in 82% of the GDM group.
Table 2.
Dietary methyl donors and B vitamins that are related to one-carbon metabolism pathway.
| Dietary intake1 |
Prevalence of Inadequacy % (n) |
Reference value2 |
p value* |
|||
| Control (n=13) | GDM (n=11) | Control (n=13) | GDM (n=11) | |||
|
Vitamins Folic Acid (μg) |
200.2 ± 74.2 |
257.4 ± 81.7 |
100 (13) |
100 (11) |
520 µg |
0.08 |
|
Vitamin B12 (μg) Vitamin B6 (mg) Methyl Donors Choline (mg) Betaine (mg) Methionine (mg) Cystine (mg) ∑ Methyl donors (mg) |
4.6 ± 1.6 1.5 ± 0.4 281.8 ± 0.4 69.9 ± 0.2 1.7 ± 0.1 0.8 ± 0.1 353.5 ± 0.4 |
4.9 ± 1.2 1.6 ± 0.2 281.6 ± 0.5 69.8 ± 0.4 1.7 ± 0.2 0.8 ± 0.1 353.0 ±1.0 |
8 (1) 46 (6) 100 (13) n/a n/a n/a n/a |
9 (1) 45 (5) 100 (11) n/a n/a n/a n/a |
2.2 µg 1.6 mg 450 mg - - - - |
0.67 0.55 0.15 0.52 0.38 0.33 0.16 |
1Intake (average ± SD) was calculated by at least three 24h dietary recalls between 13th and 34-36th gestational weeks, estimated by the Multiple Source Method (36); 2 estimated average requirement was used for folic acid, vitamin B12 and B6 values and adequate intake for choline [37] reference values; n/a: not applicable since no reference value is established. *Student’s t-test.
Regarding the intake of dietary sources of methyl groups related to one-carbon metabolism we evaluated folic acid, vitamin B12, vitamin B6, choline, betaine, methionine and cystine (Table 2). Similar intake was observed between the groups and the sum of dietary methyl donors (choline, betaine and methionine) was also similar between the groups. While inadequacy of vitamin B12 intake was relatively low, vitamin B6 had a prevalence of inadequacy of approximately 45 % in both groups. Importantly, 100% of women participating in this study presented with inadequate dietary intake of folic acid and choline. Of note, all women received folic acid supplementation during gestation as part of the pre-natal care at the Maternidade Escola/UFRJ. No other nutrient/food supplementation was prescribed or used by the participants.
2.3. Labor and Newborn Characteristics
Considering that all women participating in this study had insufficient choline intake we analyzed placental metabolites that are directly or indirectly involved in one-carbon metabolism. For this purpose, a subset of 11 placentas from the larger study were analyzed (control n=6; GDM=5), in which gestational age at birth, delivery mode (vaginal or C-section), fetal sex, newborn birth weight and length were similar between the groups (Table 3). Birth weight and length were within the normal range [38]. The prevalence of C-section appeared higher in the GDM group (80%), although the difference did not reach statistical significance (Table 3). Taken together, the similarities between the groups in gestational age at birth and mode of delivery are an important strength of the present study, since they indicate that the placental samples were subjected to the same conditions, except for the development of GDM.
Table 3.
Labor and newborn characteristics.
| Labor | Median (p25-p75) a or % frequency (n) b | |||
| Control (n=6) | Gestational diabetes mellitus (n=5) | p value | ||
| Gestational age at birth (weeks) a | 39.8 (38.6-41.2) | 38.7 (38.4- 39.5) | 0.23 | |
| Delivery mode b Vaginal C-section |
50 (3) 50 (3) |
20 (1) 80 (4) |
0.10 | |
| Newborn | ||||
| Sex b Female Male |
50 (3) 50 (3) |
80(4) 20 (1) |
0.55 | |
| Birth weight (g) a | 3433 (3139-3955) | 3240 (2970-3275) | 0.43 | |
| Birth length (cm) a | 50.3 (48.3-51.0) | 47.5 (45.5-49.5) | 0.24 | |
a Mann–Whitney test; b Fisher exact test.
2.4. Semi-Targeted Metabolomics in Placental Tissue
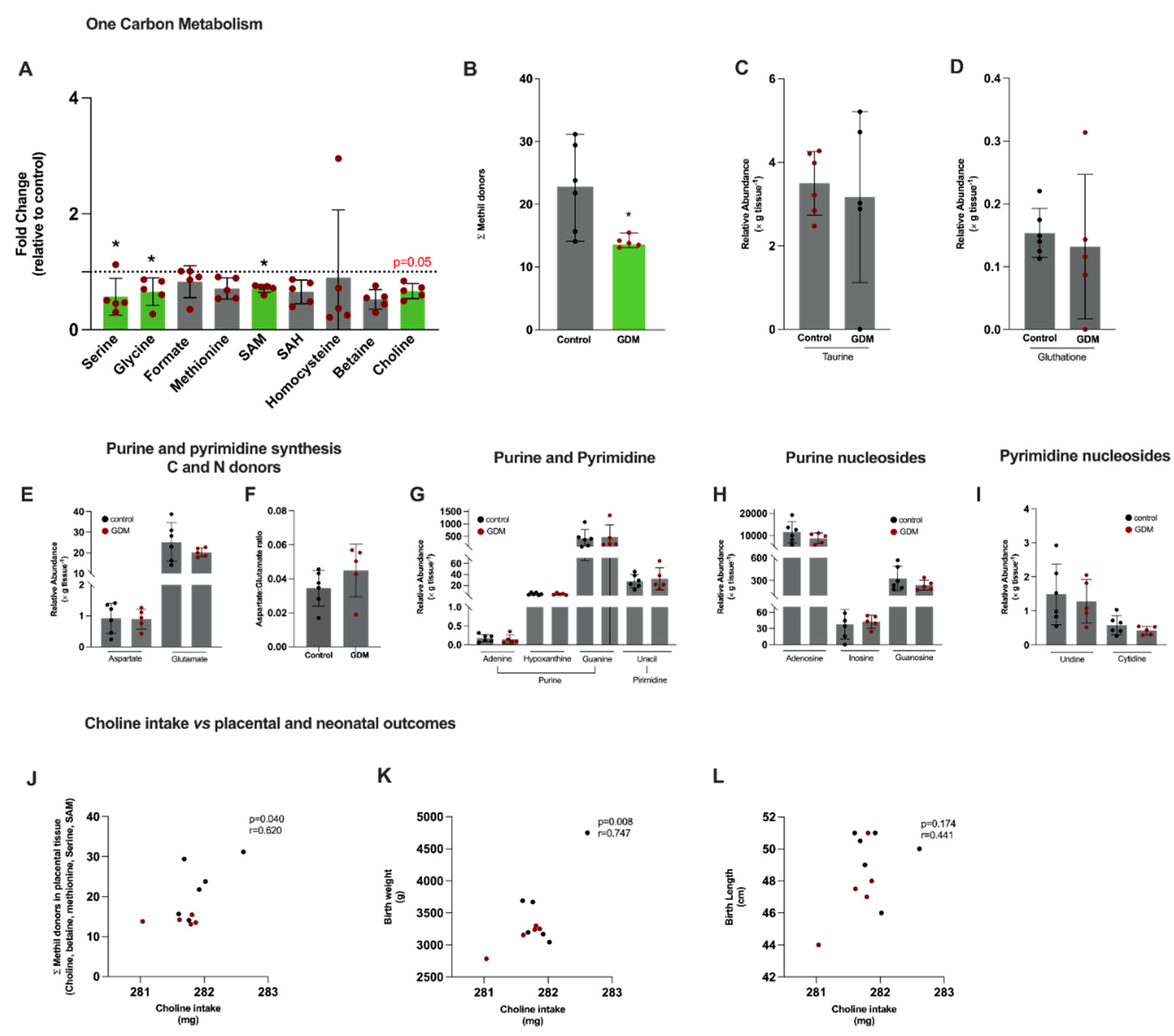
We first evaluated metabolites directly involved in one-carbon metabolism and also pathways that depend upon its intermediates (Figure 2). Using semi-targeted nuclear magnetic resonance (NMR) and mass spectrometry (MS) metabolomics, we detected serine, glycine, formate, methionine, S-adenosylmethionine (SAM), S-adenosylhomocysteine (SAH), homocysteine, betaine, and choline (Figure 2A). A significantly lower content of serine, glycine, SAM and a marginally lower content of choline (p=0.05) were observed in the GDM group (Figure 2A). In agreement, the sum of placental methyl donors (choline, methionine, betaine, SAM) was significantly lower in GDM (Figure 2B). The levels of taurine and glutathione, antioxidant molecules that are synthesized from intermediates derived from one-carbon metabolism, did not differ between the groups (Figure 2C,D, respectively).
Since one-carbon metabolism functions as a hub for several other pathways essential for tissue function, including biosynthetic pathways and proliferation, we next evaluated metabolites related to nucleic acids and nucleotide metabolism. Aspartate and glutamate are essential for nucleotide metabolism as they donate carbon and nitrogen for purine and pyrimidine synthesis. Levels were similar between the groups (Figure 2E). The aspartate-to-glutamate ratio provides a proxy measure of glutamate-oxaloacetate transaminase activity that synthesizes aspartate from glutamate in the mitochondria. The value was equivalent between the groups (Figure 2F). Metabolites related to purine synthesis, adenine, hypoxanthine and guanine, and their corresponding nucleosides, adenosine, guanosine and inosine, were also similar between the groups, as were the pyrimidine metabolite uracil and the nucleosides cytidine and uridine (Figure 2G,H,I). The alterations in one-carbon metabolism suggest that placental methyl-donors supply might be compromised, despite the fact that purine and pyrimidine synthesis seemed not to be affected.
Correlation analyses were performed to investigate associations between dietary intake to placental methyl donors and birth weight and length. A positive correlation between choline intake and the sum of placental methyl donors metabolites (choline, betaine, methionine, serine and SAM) was observed (Figure 2J). Also, choline intake correlated positively with birth weight, but not with birth length (Figure 2K,L, respectively).
Figure 2.
Placental semi-targeted metabolomics show alterations in 1C metabolism pathway in gestational diabetes mellitus pregnancies. Control, n=6 (black circles) and GDM, n=5 (red circles). (A) 1C metabolism intermediates: serine, glycine, formate, methionine, S-adenosylmethionine (SAM), S- adenosylhomocysteine (SAH), homocysteine, betaine and choline; (B): Sum of serine, methionine, SAM, betaine and choline; (C) Taurine; (D) Glutathione; (E) Aspartate and glutamate; (F) Aspartate-to-glutamate ratio; (G) Purine and pyrimidine metabolites: adenine, hypoxanthine, guanine and uracil; (H) Purine nucleosides: adenosine, inosine and guanosine; (I) Pyrimidine nucleosides: uridine and cytidine (J) Pearson’s correlation between choline intake and methyl donors sum (choline, betaine, methionine, serine and SAM); (K) Pearson’s correlation between choline intake and neonate birth weight; (L) Pearson’s correlation between choline intake and neonate birth length. *Significant differences compared to the control group, Mann-Whitney test.
Figure 2.
Placental semi-targeted metabolomics show alterations in 1C metabolism pathway in gestational diabetes mellitus pregnancies. Control, n=6 (black circles) and GDM, n=5 (red circles). (A) 1C metabolism intermediates: serine, glycine, formate, methionine, S-adenosylmethionine (SAM), S- adenosylhomocysteine (SAH), homocysteine, betaine and choline; (B): Sum of serine, methionine, SAM, betaine and choline; (C) Taurine; (D) Glutathione; (E) Aspartate and glutamate; (F) Aspartate-to-glutamate ratio; (G) Purine and pyrimidine metabolites: adenine, hypoxanthine, guanine and uracil; (H) Purine nucleosides: adenosine, inosine and guanosine; (I) Pyrimidine nucleosides: uridine and cytidine (J) Pearson’s correlation between choline intake and methyl donors sum (choline, betaine, methionine, serine and SAM); (K) Pearson’s correlation between choline intake and neonate birth weight; (L) Pearson’s correlation between choline intake and neonate birth length. *Significant differences compared to the control group, Mann-Whitney test.
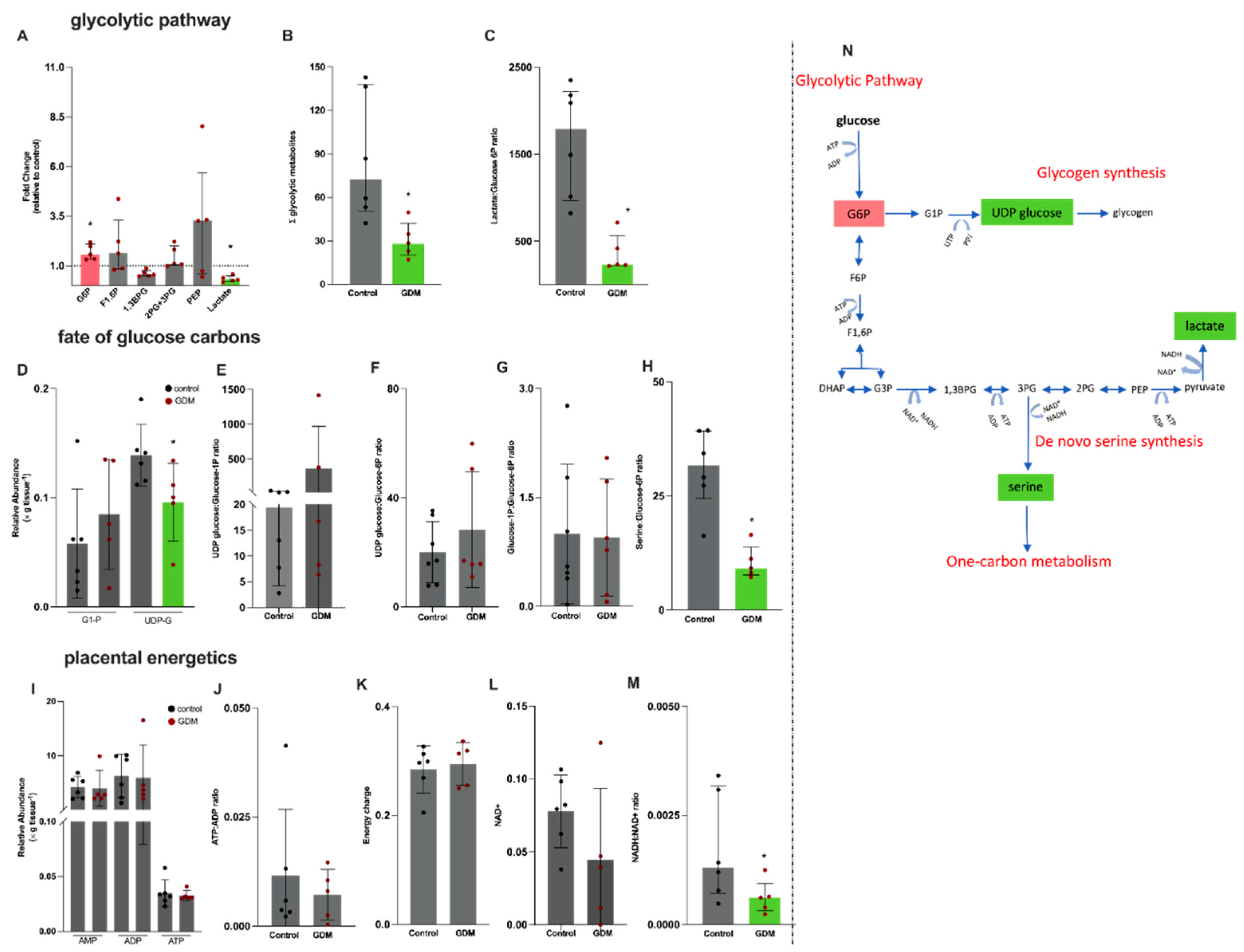
To gain insight into the alterations in one-carbon metabolism, we next investigated placental metabolites related to glucose metabolism (glycolytic pathway and glycogen synthesis) and placental energetics (Figure 3). Glycolytic intermediates detected in placental tissue were glucose-6-phosphate (G6P), fructose 1,6 biphosphate (F1,6BP), 1,3 biphosphoglycerate (1,3BPG), 2-phosphoglycerate and 3-phosphoglycerate (which could not be separated in the chromatographic analysis of MS metabolomics) and were analyzed in combination (2PG+3PG), phosphoenolpyruvate (PEP) and lactate. Detected metabolites related to glycogen metabolism were glucose-1P (G1P) and UDP-glucose.
A significant 50% higher content of placental G6P and a 1.7-fold lower content of lactate were found in the GDM group compared to controls (Figure 3A). In addition, the sum of the glycolytic intermediates (Figure 3B) and the lactate-to-G6P ratio were lower in the GDM group (Figure 3C). To investigate the potential fate of glucose carbons in placental metabolism, we analyzed glucose-1P (G1P) and uridine diphosphate glucose (UDP-glucose) (Figure 3D). Our results showed a significantly lower UDP-glucose content in GDM placentas compared to controls, while G1P levels remained similar between both groups (Figure 3D). Additionally, the UDP-glucose-to-G1P, UDP-glucose-to-G6P and G1P-to-G6P ratios were similar between groups (Figure 3E–G).
In view of the observed alterations in the glycolytic pathway and the high energy demand of the placenta, we also evaluated placental energy efficiency by analyzing AMP, ADP and ATP contents (Figure 3I), the ATP-to-ADP ratio (Figure 3J) and the energy charge (ATP+0.5*ADP/(AMP+ADP+ATP); Figure 3K). These parameters were similar between the groups. The NADH:NAD+ ratio (Figure 3M) was significantly reduced in placentas from GDM pregnancies compared to controls, while oxidized NAD+ content remained similar between the groups (Figure 3L).
The lower contents of lactate and UDP-glucose, and the lower lactate-to-G6P ratio, indicate decreased utilization of glucose carbons and prompted us to calculate the serine-to-glucose-6P ratio, which was significantly lower in placentas from GDM pregnancies compared to controls (Figure 3H). An overview of the interactions between the glycolytic pathway, glycogen and the de novo serine synthesis is shown in Figure 3N.
We identified 19 amino acids and derived metabolites in placental tissue, including 9 essential (lysine, histidine, threonine, valine, methionine, isoleucine, leucine, phenylalanine and tryptophan) and 10 nonessentials (arginine, methylhistidine, taurine, alanine, citrulline, methylarginine, proline, tyrosine, asparagine and glutamine) (Supplementary Figure S1). With the exception of tryptophan and methylarginine, there was a general trend towards a lower amino acid content in the GDM group, corresponding to approximately 40% reduction.
Figure 3.
Placental semi-targeted metabolomics shows alterations in the glycolytic pathway in gestational diabetes mellitus pregnancies. Control, n=6 (black circles) and GDM, n=5 (red circles). (A) Glycolytic intermediates: glucose-6-phosphate (G6P), fructose 1,6 biphosphate (F1,6BP), 1,3 biphosphogliceric acid (1,3BPG), 2-Phosphoglyceric and + 3- Phosphoglyceric acid (analyzed in combination (2PG+3PG), phosphoenolpyruvate (PEP) and lactate; (B) Sum of the glycolytic intermediates; (C) lactate-to-glucose-6-phosphate ratio, as a proxy of the glycolytic flux; (D) glucose-1P and UDP glucose; (E) UDP-glucose-to-G1P ratio; (F) UDP-glucose-to-G1P ratio; (G) glucose-1P-to-glucose-6P ratio; (H) serine-to-glucose6P ratio; (I) adenosine monophosphate (AMP), adenosine diphosphate (ADP) and adenosine triphosphate (ATP); (J) ATP-to-ADP ratio; (K) energy charge calculated as: ATP+0.5*ADP/(AMP+ADP+ATP); (L) NAD+; (M) NADH-to-NAD+ ratio; (N) Schematic representation of the glycolytic pathway, indicating the deviation of glucose-6P to glycogen synthesis and 3-phosphoglycerate to the de novo serine synthesis. Metabolites in green presented lower contents and in red higher content in the GDM group compared to the control group. *Significant differences compared to the control group, Mann-Whitney test.
Figure 3.
Placental semi-targeted metabolomics shows alterations in the glycolytic pathway in gestational diabetes mellitus pregnancies. Control, n=6 (black circles) and GDM, n=5 (red circles). (A) Glycolytic intermediates: glucose-6-phosphate (G6P), fructose 1,6 biphosphate (F1,6BP), 1,3 biphosphogliceric acid (1,3BPG), 2-Phosphoglyceric and + 3- Phosphoglyceric acid (analyzed in combination (2PG+3PG), phosphoenolpyruvate (PEP) and lactate; (B) Sum of the glycolytic intermediates; (C) lactate-to-glucose-6-phosphate ratio, as a proxy of the glycolytic flux; (D) glucose-1P and UDP glucose; (E) UDP-glucose-to-G1P ratio; (F) UDP-glucose-to-G1P ratio; (G) glucose-1P-to-glucose-6P ratio; (H) serine-to-glucose6P ratio; (I) adenosine monophosphate (AMP), adenosine diphosphate (ADP) and adenosine triphosphate (ATP); (J) ATP-to-ADP ratio; (K) energy charge calculated as: ATP+0.5*ADP/(AMP+ADP+ATP); (L) NAD+; (M) NADH-to-NAD+ ratio; (N) Schematic representation of the glycolytic pathway, indicating the deviation of glucose-6P to glycogen synthesis and 3-phosphoglycerate to the de novo serine synthesis. Metabolites in green presented lower contents and in red higher content in the GDM group compared to the control group. *Significant differences compared to the control group, Mann-Whitney test.
3. Discussion
This is the first study to provide an overview of the placental metabolic profile in GDM pregnancies in relation to dietary intake of methyl donors. We observed important metabolic disruptions in GDM related to one-carbon metabolism, despite the fact there were no clinical complications during gestation and delivery apart from GDM development. In addition, birth weight and length were within the normal range.
The clinical characteristics of the participants confirmed that this was a clinically well-controlled cohort, and blood glucose levels remained within the normal range among all women diagnosed with GDM [33]. However, the majority of participants, particularly in the GDM group, were classified as overweight or obese, which was expected as pre-gestational obesity is an important risk factor for the development of GDM [39]. Interestingly, we found a higher frequency of excessive gestational weight gain among women from the control group. This may be explained, in part, as a consequence of the individualized nutritional counselling provided to women with GDM, which possibly improved their dietary intake compared with the pre-pregnancy period.
Regarding maternal diet, the intake of fiber, total sugar and ultra-processed products were comparable between the control and GDM groups. However, it is important to highlight that fiber intake was insufficient in both groups. It is known that fiber intake modulates the microbiota which interferes directly with many metabolic pathways, including glycemic control. Therefore, it could negatively interfere with many aspects of the pathophysiology of GDM, such as increased insulin resistance [40].
Inadequate intake of nutrients involved in one-carbon metabolism, including folic acid, cobalamin, choline and other methyl donors, has been associated with elevated levels of homocysteine as well as adverse maternal and neonatal outcomes [41,42,43]. Indeed, in this study, 100% of women presented inadequate choline intake. Also, our results showed that placentas from GDM pregnancies contained significantly lower amounts of serine, glycine, SAM and a trend to a lower amount of choline (p=0.05) if compared to controls.
Choline plays a multifaceted role in maternal and fetal health. Choline is required for liver and brain function, lipid metabolism, and cell membrane synthesis, and is an important methyl donor for methylation reactions [44]. Also, it has been demonstrated that maternal intake of methyl donors is associated with improved neurocognitive development in children and a lower risk of neural tube defects [43,45]. Van Lee et al. found a correlation between maternal choline status, neonatal BMI z-score and total body fat, suggesting increased adiposity at birth and reinforcing the role of choline in early growth [46]. Animal studies suggest that choline intake may support key placental functions, including angiogenesis [47,48], inflammation [49] and nutrient transport [50]. Moreover, a connection between choline and nutrient metabolism associated with rapamycin (mTOR) signaling pathway in the placenta [24] and downregulation of lipogenic gene expression in GDM mouse embryos [50] have been described. Indeed, in the present study, we observed a significant positive correlation between maternal choline intake and birth weight, and also between placental methyl donors metabolites (choline, betaine, methionine, serine and SAM).
Choline in the maternal circulation is likely the primary source for the fetus given the low level of expression in both the fetal liver and placenta of phosphatidylethanolamine N-methyltransferase (PEMT), a key regulatory enzyme in the choline synthesis pathway [51]. The placenta expresses specific choline transporters, and in term pregnancies fetal plasma choline concentrations are typically higher than those in maternal plasma [52], suggesting that placental choline transport and metabolism are essential for the fetal supply. Therefore, the inadequacy in choline intake, particularly in women that developed GDM might have contributed to these alterations in metabolites from the one-carbon metabolism, decreasing the availability of methyl donors for both placental and fetal requirements.
Deficiency in dietary choline induces hepatic steatosis in rat models and was associated with increased oxidative stress [53]. Related, low-grade placental endoplasmic reticulum stress in GDM pregnancies, characterized by dilation of endoplasmic reticulum cisternae and increased phosphorylation of eukaryotic initiation factor 2 subunit alpha, has been reported by our group [15]. It was also observed in the trophoblast-like cell line BeWo when exposed to a high-glucose insult, and could be prevented by addition of antioxidants vitamin C and E to the culture medium. Despite placental homocysteine content being similar between the groups, the dysregulation in placental one-carbon metabolism and also the inadequate intake of choline might have contributed to the increased placental oxidative stress in GDM pregnancies.
Like choline, glycine is a conditionally essential amino acid during pregnancy required for adequate maternal protein synthesis [54]. Also, the fetus requires large amounts of glycine to meet structural and metabolic demands. Placental supply of glycine may be limited both via its direct transfer [55] and also through serine conversion due to the low activity of the enzyme serine hydroxymethyltransferase [56]. Although not conclusively shown in humans, it seems that fetal glycine demand may be met by the fetal conversion of serine. Therefore, the lower content of serine in placentas from GDM pregnancies may compromise the fetal supply of glycine. Placental release of glycine to the maternal circulation also seems important for placental uptake of essential amino acids via L-system transporters [57]. Therefore, the lower content of glycine in placentas from GDM pregnancies may compromise placental protein synthesis and other functions. Indeed, the lower overall amino acid content in GDM placentas corroborates with this aspect and with low-grade ER stress [15].
Alterations in glucose metabolism, evidenced by the higher G6P content, the lower sum of glycolytic intermediates and the lower lactate-to-G6P ratio might have compromised the de novo synthesis of serine, contributing to the observed alterations in the one-carbon metabolism. Indeed, the lower serine-to-G6P ratio confirmed the decreased flow of glucose carbons towards serine synthesis. The lower NADH-to-NAD+ ratio in placentas from GDM pregnancies further supports the decreased glycolytic flux. Decreased glycolysis has been reported in trophoblasts from GDM pregnancies [4] as well as increased content of placental G6P [58]. The molecular aspects involved in alterations in glycolysis in placentas from GDM pregnancies might be related to miR-143 [59], and miR-9 and miR-12 [60]. Although altered glycolysis certainly contributed to the lower content of one-carbon metabolites and possibly to their fetal supply, placental energetics seemed not to be affected. Also, the lower lactate content in placentas from GDM pregnancies might suggest adequate glycemic [61] and ketogenesis [62] control possibly associated with the use of insulin, which was administered to the majority of women with GDM. Decreased content of placental UDP-glucose in GDM suggests decreased glycogen synthesis, which may also be associated with adequate glycemic control and overall decreased placental stress [63], as already proposed in our previous study considering the lower content of F2-Isoprostanes in placentas from GDM pregnancies [17]. Altogether, these alterations suggest, to some extent, placental compensatory responses that maintained birthweight and length.
Limitations and Future Recommendations
A major limitation of this study is the small sample size which impacts on the statistical power. We could not address placental sex dimorphism, as groups were unbalanced in sex and sample size was limited. This would be an important aspect to address as placental metabolism seems to differ according to fetal sex in gestational diabetes [64]. In addition, these data would benefit from mechanistic studies, which would allow specific metabolic steps involved in the overall alterations in placental one-carbon metabolism to be investigated.
4. Materials and Methods
4.1. Study Design and Participants
Our study comprises data from a prospective cohort of pregnant women that was conducted between July 2017 to March 2019, at the Maternidade Escola, Federal University of Rio de Janeiro, a referral hospital in pre-natal and delivery care in Rio de Janeiro. “Women” is used to describe the participants as only cisgender women participated in this study. The cohort was composed of four follow-up waves: at recruitment up to 24 weeks, between 24-28 weeks (2nd trimester), between 34-35 weeks (3rd trimester) and at delivery. Socio-demographic data were collected at recruitment using a structured questionnaire and labor and neonatal outcomes from the medical charts. All women were followed up by the research group through telephone contact and in person during the routine prenatal care visits.
Inclusion criteria were: age between 18 and 45 years; beginning of follow-up at ME-UFRJ with up to 24 weeks of gestation; BMI > 18.5 kg/m2; free of infectious and non-transmissible chronic diseases (except obesity); singleton pregnancy; intention to deliver at the referral maternity hospital.
GDM diagnosis was performed between 24th and 28th weeks of gestation according to the American Diabetes Association criteria: plasma glucose values >92 mg/dL (fasting) or after 75-g oral glucose tolerance test >180 mg/dL (1 h) or >153 mg/dL (2h). GDM management included nutritional counselling for all, plus insulin therapy if needed. Gestational weight gain was classified accordingly [34] and birthweight and length were classified according to the INTERGROWTH-21st Project Curves [38]. Pregnant women as well as placental samples were grouped according to GDM.
For this study, maternal pre-gestational overweight and obesity and GDM were the only associated pregnancy complications. We prioritized the homogeneity of placental samples and pregnancies with complications were not included, resulting in a final sample size of 24 women for the dietary data. Placental sample size was n=11: n=6 for women that did not develop GDM (control group) and n=5 for women who developed GDM (GDM group).
4.2. Ethical Approval
This study was conducted according to the guidelines laid down in the Declaration of Helsinki. All procedures involving human subjects/patients were approved by the Research Ethics Committee of the Maternidade Escola-UFRJ and registered at the Research Ethics National Council Certificate of presentation for ethic appreciation 66949217.0.0000.5257 and registered at clinicaltrials.org NCT05174728 and at the Brazilian Registry of Clinical Trials - REBEC (RBR-3xbpwqy). All participants who agreed to take part in the study signed a two-copy form of a free and informed consent.
4.3. Assessment of Dietary Data
Dietary data was obtained by trained dietitians. Two 24-h recall (24hR) were collected in the 2nd and 3rd trimesters of pregnancy using the multiple-pass method [65]: one face-to-face and another up to 2 days later by a phone interview [66]. This method was validated for obtaining dietary recalls of pregnant women in Brazil [67]. Trained dietitians systematized the portions and home measures, as well as the insertion of dietary data into the database. Nutrient intake was analyzed with the Dietbox® v.4.0 software Rio de Janeiro, Brazil, using the United States department of Agriculture (USDA), Brazilian institute of geography and statistics (IBGE) and the Brazilian table of food composition (TACO) database. Ultra-processed food was identified using the NOVA Classification [68]. The consumption of ultra-processed food was assessed in relation to daily energy intake. Dietary intake was analyzed by the Multiple Source Method (MSM) which is a statistical method for estimating usual dietary intake that considers intra- and inter-individual variation over the same period using at least two repeated measurements per participant (e.g., dietary recalls and/or food frequency questionnaire) [36]. MSM ability to estimate usual nutrient or food intake for occasional consumers is one of the strengths of this method, which may reduce the need for large number of subjects to assess dietary intake. The MSM has been shown to accurately estimate usual nutrient intake during pregnancy [67].
4.4. Placental Sampling and Tissue Extraction for Metabolomics
Placental samples were collected at delivery from 13 women according to a standardized protocol [69]. Briefly, villous samples (1-2 cm³) were taken at four random sites, representative of the whole tissue, from the maternal surface after the removal of the basal plate and decidua. Villi were washed twice in ice-cold phosphate-buffered saline at 4 °C, cut into smaller pieces (50 mg), placed into cryovials (all four sites combined) and immediately frozen in liquid nitrogen. All samples were stored at −80 °C until analysis.
We used a modified Folch protocol for placental metabolites’ extraction. Briefly, 0.35 mL of cold methanol and 0.35 mL of chloroform were added to plastic tubes containing 45±5 mg of frozen placental fragments and one metallic bead. Placental tissue was disrupted in a tissue lyser (Tissue Lyser-II, Qiagen) under a 2 × 2.5 min, 17 s−1. Subsequently, 0.35 ml of chloroform and 0.15 mL of ultrapure water were added and samples were thoroughly vortexed for 30 seconds and centrifugated (17,000 × g, 10 min). This optimized extraction protocol ensures consistency and reproducibility in NMR metabolite profiling.
The upper hydrophilic phase was aliquoted (~700 μl) into a glass vial, dried under a gentle stream of nitrogen and stored at −80 °C until further analysis.
4.4.1. Semi-Targeted Mass Spectrometry Metabolomics
Dried placental samples were subjected to two solvent reconstitutions due to two different methods with UHPLC+series/MS-TSQ Quantiva/ESI (+3500V and – 2500V mode at the same time): (a) normal phase column (BEH Amide (150 X 2,1 mm 1,7 μm) and (b) reversed phase column ACE Excel 2 C18 PFP (100A. 150 x 2.1mm 5μ) both at 30o C. Samples were reconstituted in (a) 0.1 mL of acetonitrile:ammonium carbonate water solution (10 mM) (7:3, v/v) containing a mixture of 10 internal standards (representative of all assigned metabolites) at the concentration of 10 μM (proline-d3, valine-d8, leucine-d10, U-13C-lysine, U-13C glutamic acid, phenilanine-d5, succinic acid-d3, serotonine-d4, adenosine- 13C10, 15N5 5′-monophosphate and adenosine- 15N5 5′-triphosphate and (b) 0.1 mL of an aqueous solution containing ammonium acetate (10 mM). The Xcalibur software was used for data acquisition. Putative recognition of all detected metabolites was performed using a targeted MS/MS analysis and intensities were normalized by their respective homologous compound or to their closest class. Prior to statistical analyses all intensities were normalized for wet weight of each placenta sample [70]. Molecular mass and mass fragments of the assigned metabolites are reported in Table 2.
4.4.2. 1H-Nuclear Magnetic Resonance-Based Metabolomics
Before NMR-based metabolomics, samples were reconstituted in 600 μL of sodium phosphate buffer (50 mM, pH 7.4) prepared in 10% deuterium oxide (v/v) with 100 μL trimethylsilypropanoic acid (TSP) as an internal chemical shift reference.
NMR spectra were acquired on a Bruker Advance III at 500.13 MHz at 300 K, coupled with a cooled automatic sample case at 280 K. 1D-1H NMR spectra were acquired using excitation sculpting to suppress the solvent signal [71] as well as a CPMG (Carr-Purcell- Meiboom-Gill) T2 filter [72] with 32 loop counters and a delay of 0.001 s. 65,536 complex data points were acquired per transient, for a total of 1024 transients. The spectral width was set to 19.99 ppm, resulting in an acquisition time of 3.27 s per FID. The relaxation delay was set to 1.74 s. Spectra data were pre-processed in the MetaboLab [73] software v. 2022.0726.1733. Prior to the Fourier transform, the FIDs were apodised using an exponential window function with 0.3 Hz line-broadening and then zero-filled to 131,072 data points. After Fourier transform, each spectrum was manually phase corrected, followed by a spline baseline correction. The Human Metabolome Database (HMDB) [74] and Chenomx NMR Suite 12® program (Chenomx Inc., Edmonton, AB, Canada) were used for the assignment of metabolites.
Supplementary Table S3 presents the 1H NMR assignment information for the metabolites of interest.
5. Conclusions
In conclusion, we provide an overview of the placental metabolic profile in pregnancies complicated with gestational diabetes mellitus and its relation to the intake of dietary methyl donors. We highlight significant metabolic alterations in placentas from pregnancies affected by GDM, despite good glycemic control and the absence of adverse neonatal outcomes. The observed disruptions in one-carbon metabolism, notably reduced levels of choline, serine and SAM, underscore the alterations in placental metabolic state in GDM.
Importantly, the study emphasizes the critical role of maternal nutrition, particularly the intake of methyl donors like choline, in maintaining placental metabolic balance. The consistent inadequacy of choline intake across both study groups raises concerns about current dietary recommendations during pregnancy. The positive correlation between maternal choline intake, placental methyl donors and neonatal birth weight further reinforces the importance of dietary choline during pregnancy.
While the placenta demonstrated adaptive mechanisms that possibly mitigated placental stress, the long-term implications of these adaptations on offspring health remain uncertain. Given the critical role of methyl donors in epigenetic regulation and fetal programming, this study advocates for further research to refine dietary guidelines, potentially guiding more tailored nutritional interventions with a specific focus on choline supplementation during pregnancy. Such strategies could improve maternal and neonatal health, mitigating the intergerational impact of GDM and promoting optimal pregnancy outcomes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Table S1. Intake of energy, macronutrients, fiber, added sugar and ultra-processed food. Supplementary Table S2: Molecular mass and mass fragments of the assigned metabolites. Supplementary table 3: Assignment table of metabolites of interested in NMR metabolomics. Supplementary Figure S1. Placental semi-targeted metabolomics shows an overall lower amino acids concentration in gestational diabetes mellitus pregnancies
Author Contributions
Conceptualization, T.E.-B. and G.J.B; methodology, V.A.G., D.L., A.M., C.L., C.L., T.E.-B.; formal analysis, V.A.G., D.L., L.R., M.T.N.R., C.S.F., G.D.A.P., A.M., C.L., A.G.T. and T.E.-B.; investigation, V.A.G., D.L., L.R., M.T.N.R., C.S.F., G.D.A.P., A.M., C.L., and T.E.-B.; resources, T.E.-B., G.J.B., J.L.G.; data curation, A.M., C.L., C.L., and J.L.G.; writing—original draft preparation, V.A.G. and T.E.-B.; writing—review and editing, all authors; visualization, all authors; supervision, T.E.-B.; project administration, T.E.-B.; funding acquisition, T.E.-B., A.G.T., G.J.B., J.L.G.. All authors contributed to data interpretation and performed final editing checks and approved the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro for T.E.-B. (E-026/203.254/2017 and E-26/200.930/2022), Brazilian National Council for Scientific Development (CNPq) fellowship for T.E.-B. (314971/2021-1, PQ2), Isaac Newton Trust research grant for T.E.-B., Global Challenge Research Fund (ref. 102642/A19819) for T.E., A.G.T., and G.J.B., and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior–CAPES-Brasil (Finance code 001), Brazil. V.A.G., D. L., L.R. were recipients of CAPES scholarships. M.T.N.R. was recipient of CNPq undergraduate scholarship. C.S.F. is a post-doctoral fellow recipient of the CNPq scholarship (PDJ). G.D.A.P. is a post-doctoral fellow recipient of the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro scholarship PDR-Nota 10. The APC was covered partially by FAPERJ and the University of Birmingham.
Institutional Review Board Statement
This study was conducted according to the guidelines laid down in the Declaration of Helsinki. All procedures involving human subjects/patients were approved by the Research Ethics Committee of the Maternidade Escola-UFRJ and registered at the Research Ethics National Council Certificate of presentation for ethic appreciation 66949217.0.0000.5257 on July 2017 and registered at clinicaltrials.org NCT05174728 and at the Brazilian Registry of Clinical Trials - REBEC (RBR-3xbpwqy). All participants who agreed to take part in the study signed a two-copy form of a free and informed consent.
Acknowledgments
The authors acknowledge the contribution of the women that participated in this study, without any financial compensation.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ADP – Adenosine diphosphate AMP – Adenosine monophosphate ATP – Adenosine triphosphate 1,3BPG - 1,3 biphosphoglycerate BMI – Body mass index DHF - Dihydrofolate DMG - Dimethylglycine DNA – Deoxyribonucleic acid DOHaD – Developmental origins of Health and Disease ER – Endoplasmic reticulum F1,6P - Fructose 1,6 biphosphate G1P – Glucose-1-phosphate G6P – Glucose-6-phosphate GDM – Gestational diabetes mellitus GOT - glutamate-oxaloacetate transaminase GSH – Glutathione IGF2 – Insulin-like Growth Factor 2 IOM – Institute of Medicine LPS – lipopolysaccharide MS – Mass spectrometry MS/MS – Tandem mass spectrometry MSM – Multiple source method NAD+ - Oxidised Nicotinamide adenine dinucleotide NADH – Reduced Nicotinamide adenine dinucleotide NMR – Nuclear magnetic resonance PPARy – Proliferator-Activated Receptor Gamma 2PG+3PG - 2-Phosphoglycerate and 3-Phosphoglycerate 3PS – 3-phosphoserine 3PG - 3-phosphoglyceric acid PEP – phosphoenolpyruvate PEMT – phosphatidylethanolamine N-methyltransferase R24h – 24-hour dietary recordings SAH – S-adenosylhomocysteine SAM - S-adenosyl methionine SHMT – Serine hydroxymethyltransferase 5,10-meTHF – 5,10 methyltetrahydrofolate 10-formylTHF – 10 formyl tetrahydrofolate THF - Tetrahydrofolate UDP-glucose – Uridine diphosphate glucose |
References
- Parrettini, S.; Caroli, A.; Torlone, E. Nutrition and Metabolic Adaptations in Physiological and Complicated Pregnancy: Focus on Obesity and Gestational Diabetes. Front Endocrinol (Lausanne) 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.J.; Parra, H.; Santeliz, R.; Bautista, J.; Luzardo, E.; Villasmil, N.; Martínez, M.S.; Chacín, M.; Cano, C.; Checa-Ros, A.; D’Marco, L.; Bermúdez, V.; De Sanctis, J.B. The Placental Role in Gestational Diabetes Mellitus: A Molecular Perspective. touchREVIEWS in Endocrinology 2024, 20, 10. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, H.D.; Catalano, P.; Zhang, C.; Desoye, G.; Mathiesen, E.R.; Damm, P. Gestational Diabetes Mellitus. Nat Rev Dis Primers 2019, 5. [Google Scholar] [CrossRef] [PubMed]
- Valent, A.M.; Choi, H.; Kolahi, K.S.; Thornburg, K.L. Hyperglycemia and Gestational Diabetes Suppress Placental Glycolysis and Mitochondrial Function and Alter Lipid Processing. FASEB Journal. 2021. [CrossRef]
- Lizárraga, D.; García-Gasca, A. The Placenta as a Target of Epigenetic Alterations in Women with Gestational Diabetes Mellitus and Potential Implications for the Offspring. Epigenomes 2021, 5, 13. [Google Scholar] [CrossRef]
- He, Y.; Yang, X.; Wu, N. TGF β 1, SNAIL2, and PAPP-A Expression in Placenta of Gestational Diabetes Mellitus Patients. J Diabetes Res 2024, 2024. [Google Scholar] [CrossRef]
- Yi, Y.; Wang, T.; Xu, W.; Zhang, S.-H. Epigenetic Modifications of Placenta in Women with Gestational Diabetes Mellitus and Their Offspring. World J Diabetes 2024, 15, 378–391. [Google Scholar] [CrossRef]
- GLOBAL REPORT ON DIABETES WHO Library Cataloguing-in-Publication Data. ISBN 978, 92–94.
- Giannakou, K.; Evangelou, E.; Yiallouros, P.; Christophi, C.A.; Middleton, N.; Papatheodorou, E.; Papatheodorou, S.I. Risk Factors for Gestational Diabetes: An Umbrella Review of Meta-Analyses of Observational Studies. PLoS One 2019, 14. [Google Scholar] [CrossRef]
- Xie, Y.P.; Lin, S.; Xie, B.Y.; Zhao, H.F. Recent Progress in Metabolic Reprogramming in Gestational Diabetes Mellitus: A Review. Front Endocrinol (Lausanne) 2024, 14. [Google Scholar] [CrossRef]
- Saucedo, R.; Ortega-Camarillo, C.; Ferreira-Hermosillo, A.; Díaz-Velázquez, M.F.; Meixueiro-Calderón, C.; Valencia-Ortega, J. Role of Oxidative Stress and Inflammation in Gestational Diabetes Mellitus. Antioxidants 2023, 12, 1812. [Google Scholar] [CrossRef]
- Zgutka, K.; Tkacz, M.; Tomasiak, P.; Piotrowska, K.; Ustianowski, P.; Pawlik, A.; Tarnowski, M. Gestational Diabetes Mellitus-Induced Inflammation in the Placenta via IL-1β and Toll-like Receptor Pathways. Int J Mol Sci 2024, 25, 11409. [Google Scholar] [CrossRef] [PubMed]
- Radaelli, T.; Varastehpour, A.; Catalano, P.; Mouzon, S.H. Gestational Diabetes Induces Placental Genes for Chronic Stress and Inflammatory Pathways GDM ALTERS EXPRESSION PROFILE OF PLACENTAL GENES. Diabetes 2003, 52, 2951–2958. [Google Scholar] [CrossRef] [PubMed]
- Phoswa, W.N.; Khaliq, O.P. The Role of Oxidative Stress in Hypertensive Disorders of Pregnancy (Preeclampsia, Gestational Hypertension) and Metabolic Disorder of Pregnancy (Gestational Diabetes Mellitus). Oxid Med Cell Longev 2021, 2021. [Google Scholar] [CrossRef] [PubMed]
- Yung, H. wa; Alnæs-Katjavivi, P.; Jones, C.J.P.; El-Bacha, T.; Golic, M.; Staff, A.C.; Burton, G.J. Placental Endoplasmic Reticulum Stress in Gestational Diabetes: The Potential for Therapeutic Intervention with Chemical Chaperones and Antioxidants. Diabetologia 2016. [CrossRef]
- Pinto, G.D.A.; Murgia, A.; Lai, C.; Ferreira, C.S.; Goes, V.A.; De A., B. Guimarães, D.; Ranquine, L.G.; Reis, D.L.; Struchiner, C.J.; Griffin, J.L.; Burton, G.J.; Torres, A.G.; El-Bacha, T. Sphingolipids and Acylcarnitines Are Altered in Placentas from Women with Gestational Diabetes Mellitus. Br J Nutr 2023, 130, 921–932. [Google Scholar] [CrossRef]
- Ferreira, C.S.; Pinto, G.D.A.; Reis, D.L.; Vigor, C.; Goes, V.A.; Guimarães, D. de A. B.; Mucci, D.B.; Belcastro, L.; Saraiva, M.A.; Oger, C.; Galano, J.M.; Sardinha, F.L.C.; Torres, A.G.; Durand, T.; Burton, G.J.; El-Bacha, T. Placental F4-Neuroprostanes and F2-Isoprostanes Are Altered in Gestational Diabetes Mellitus and Maternal Obesity. Prostaglandins Leukot Essent Fatty Acids 2023, 189. [Google Scholar] [CrossRef]
- Cai, S.; Quan, S.; Yang, G.; Ye, Q.; Chen, M.; Yu, H.; Wang, G.; Wang, Y.; Zeng, X.; Qiao, S. One Carbon Metabolism and Mammalian Pregnancy Outcomes. Mol Nutr Food Res 2021, 65. [Google Scholar] [CrossRef]
- Nema, J.; Joshi, N.; Sundrani, D.; Joshi, S. Influence of Maternal One Carbon Metabolites on Placental Programming and Long Term Health. Placenta 2022, 125, 20–28. [Google Scholar] [CrossRef]
- Cochrane, K.M.; Williams, B.A.; Elango, R.; Barr, S.I.; Karakochuk, C.D. Pregnancy-Induced Alterations of 1-Carbon Metabolism and Significance for Maternal Nutrition Requirements. Nutr Rev 2022, 80, 1985–2001. [Google Scholar] [CrossRef]
- Sun, W.X.; Shu, Y.P.; Yang, X.Y.; Huang, W.; Chen, J.; Yu, N.N.; Zhao, M. Effects of Folic Acid Supplementation in Pregnant Mice on Glucose Metabolism Disorders in Male Offspring Induced by Lipopolysaccharide Exposure during Pregnancy. Sci Rep 2023, 13. [Google Scholar] [CrossRef]
- Kintaka, Y.; Wada, N.; Shioda, S.; Nakamura, S.; Yamazaki, Y.; Mochizuki, K. Excessive Folic Acid Supplementation in Pregnant Mice Impairs Insulin Secretion and Induces the Expression of Genes Associated with Fatty Liver in Their Offspring. Heliyon 2020, 6, e03597. [Google Scholar] [CrossRef] [PubMed]
- Williamson, J.M.; Arthurs, A.L.; Smith, M.D.; Roberts, C.T.; Jankovic-Karasoulos, T. High Folate, Perturbed One-Carbon Metabolism and Gestational Diabetes Mellitus. Nutrients 2022, 14, 3930. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.; Greenwald, E.; Jack-Roberts, C.; Ajeeb, T.T.; Malysheva, O.V.; Caudill, M.A.; Axen, K.; Saxena, A.; Semernina, E.; Nanobashvili, K.; Jiang, X. Choline Prevents Fetal Overgrowth and Normalizes Placental Fatty Acid and Glucose Metabolism in a Mouse Model of Maternal Obesity. J Nutr Biochem 2017, 49, 80–88. [Google Scholar] [CrossRef] [PubMed]
- King, J.H.; Kwan, S.T.C.; Yan, J.; Klatt, K.C.; Jiang, X.; Roberson, M.S.; Caudill, M.A. Maternal Choline Supplementation Alters Fetal Growth Patterns in a Mouse Model of Placental Insufficiency. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- King, J.H.; Kwan, S.T.C.; Yan, J.; Jiang, X.; Fomin, V.G.; Levine, S.P.; Wei, E.; Roberson, M.S.; Caudill, M.A. Maternal Choline Supplementation Modulates Placental Markers of Inflammation, Angiogenesis, and Apoptosis in a Mouse Model of Placental Insufficiency. Nutrients 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Adaikalakoteswari, A.; Wood, C.; Mina, T.H.; Webster, C.; Goljan, I.; Weldeselassie, Y.; Reynolds, R.M.; Saravanan, P. Vitamin B12 Deficiency and Altered One-Carbon Metabolites in Early Pregnancy Is Associated with Maternal Obesity and Dyslipidaemia. Sci Rep 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Li, X.; Du, Y.; Yang, J.; Li, X.; Li, J.; Zhao, Y.; Wei, Y. Relationship between One-Carbon Metabolism and Fetal Growth in Twins: A Cohort Study. Food Sci Nutr 2023, 11, 6626–6633. [Google Scholar] [CrossRef]
- Du, Y.F.; Wei, Y.; Yang, J.; Cheng, Z.Y.; Zuo, X.F.; Wu, T.C.; Shi, H.F.; Wang, X.L. Maternal Betaine Status, but Not That of Choline or Methionine, Is Inversely Associated with Infant Birth Weight. Br J Nutr 2019, 121, 1279–1286. [Google Scholar] [CrossRef]
- Kadam, I.; Dalloul, M.; Hausser, J.; Vaday, D.; Gilboa, E.; Wang, L.; Hittelman, J.; Hoepner, L.; Fordjour, L.; Chitamanni, P.; Saxena, A.; Jiang, X. Role of One-Carbon Nutrient Intake and Diabetes during Pregnancy in Children’s Growth and Neurodevelopment: A 2-Year Follow-up Study of a Prospective Cohort. Clin Nutr 2024, 43, 1216–1223. [Google Scholar] [CrossRef]
- Kadam, I.; Dalloul, M.; Hausser, J.; Huntley, M.; Hoepner, L.; Fordjour, L.; Hittelman, J.; Saxena, A.; Liu, J.; Futterman, I.D.; Minkoff, H.; Jiang, X. Associations between Nutrients in One-Carbon Metabolism and Fetal DNA Methylation in Pregnancies with or without Gestational Diabetes Mellitus. Clin Epigenetics 2023, 15, 137. [Google Scholar] [CrossRef]
- Barzilay, E.; Moon, A.; Plumptre, L.; Masih, S.P.; Sohn, K.J.; Visentin, C.E.; Ly, A.; Malysheva, O.; Croxford, R.; Caudill, M.A.; O’Connor, D.L.; Kim, Y.I.; Berger, H. Fetal One-Carbon Nutrient Concentrations May Be Affected by Gestational Diabetes. Nutr Res 2018, 55, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Tests, D.; Diabetes, F.O.R. 2. Classification and Diagnosis of Diabetes. Diabetes Care 2016, 39, S13–S22. [Google Scholar] [CrossRef]
- Kac, G.; Carrilho, T.R.B.; Rasmussen, K.M.; Reichenheim, M.E.; Farias, D.R.; Hutcheon, J.A. Gestational Weight Gain Charts: Results from the Brazilian Maternal and Child Nutrition Consortium. Am J Clin Nutr 2021, 113, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Medicine, I. of. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids (Macronutrients) 2002, 1–1331. [CrossRef]
- Harttig, U.; Haubrock, J.; Knüppel, S.; Boeing, H. The MSM Program: Web-Based Statistics Package for Estimating Usual Dietary Intake Using the Multiple Source Method. Eur J Clin Nutr 2011, 65 (Suppl. 1), S87–S91. [Google Scholar] [CrossRef]
- Medicine, I. of Dietary Reference Intakes: The Essential Guide to Nutrient Requirements. 2006. [CrossRef]
- Villar, J.; Ismail, L.C.; Victora, C.G.; Ohuma, E.O.; Bertino, E.; Altman, D.G.; Lambert, A.; Papageorghiou, A.T.; Carvalho, M.; Jaffer, Y.A.; et al. International Standards for Newborn Weight, Length, and Head Circumference by Gestational Age and Sex: The Newborn Cross-Sectional Study of the INTERGROWTH-21st Project. Lancet 2014, 384, 857–868. [Google Scholar] [CrossRef]
- Li, Y.; Ren, X.; He, L.; Li, J.; Zhang, S.; Chen, W. Maternal Age and the Risk of Gestational Diabetes Mellitus: A Systematic Review and Meta-Analysis of over 120 Million Participants. Diabetes Res Clin Pract 2020, 162. [Google Scholar] [CrossRef]
- Pheiffer, C.; Riedel, S.; Dias, S.; Adam, S. Gestational Diabetes and the Gut Microbiota: Fibre and Polyphenol Supplementation as a Therapeutic Strategy. Microorganisms 2024, Vol. 12, Page 633 2024, 12, 633. [Google Scholar] [CrossRef]
- Finkelstein, J.D. Pathways and Regulation of Homocysteine Metabolism in Mammals. Semin Thromb Hemost 2000, 26, 219–225. [Google Scholar] [CrossRef]
- Zeisel, S.H. Choline: Critical Role during Fetal Development and Dietary Requirements in Adults. Annu Rev Nutr 2006, 26, 229–250. [Google Scholar] [CrossRef]
- Shaw, G.M.; Carmichael, S.L.; Yang, W.; Selvin, S.; Schaffer, D.M. Periconceptional Dietary Intake of Choline and Betaine and Neural Tube Defects in Offspring. Am J Epidemiol 2004, 160, 102–109. [Google Scholar] [CrossRef]
- Li, Z.; Vance, D.E. Phosphatidylcholine and Choline Homeostasis. J. Lipid Res. 2008, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Obeid, R.; Schön, C.; Derbyshire, E.; Jiang, X.; Mellott, T.J.; Blusztajn, J.K.; Zeisel, S.H. A Narrative Review on Maternal Choline Intake and Liver Function of the Fetus and the Infant; Implications for Research, Policy, and Practice. Nutrients 2024, 16, 260. [Google Scholar] [CrossRef] [PubMed]
- Van Lee, L.; Crozier, S.R.; Aris, I.M.; Tint, M.T.; Sadananthan, S.A.; Michael, N.; Quah, P.L.; Robinson, S.M.; Inskip, H.M.; Harvey, N.C.; et al. Prospective Associations of Maternal Choline Status with Offspring Body Composition in the First 5 Years of Life in Two Large Mother-Offspring Cohorts: The Southampton Women’s Survey Cohort and the Growing Up in Singapore Towards Healthy Outcomes Cohort. Int J Epidemiol 2019, 48, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Bar, H.Y.; Yan, J.; Jones, S.; Brannon, P.M.; West, A.A.; Perry, C.A.; Ganti, A.; Pressman, E.; Devapatla, S.; Vermeylen, F.; Wells, M.T.; Caudill, M.A. A Higher Maternal Choline Intake among Third-Trimester Pregnant Women Lowers Placental and Circulating Concentrations of the Antiangiogenic Factor Fms-like Tyrosine Kinase-1 (SFLT1). FASEB J 2013, 27, 1245–1253. [Google Scholar] [CrossRef]
- Jiang, X.; Jones, S.; Andrew, B.Y.; Ganti, A.; Malysheva, O.V.; Giallourou, N.; Brannon, P.M.; Roberson, M.S.; Caudill, M.A. Choline Inadequacy Impairs Trophoblast Function and Vascularization in Cultured Human Placental Trophoblasts. J Cell Physiol 2014, 229, 1016–1027. [Google Scholar] [CrossRef]
- King, J.H.; Kwan, S.T.C.; Yan, J.; Jiang, X.; Fomin, V.G.; Levine, S.P.; Wei, E.; Roberson, M.S.; Caudill, M.A. Maternal Choline Supplementation Modulates Placental Markers of Inflammation, Angiogenesis, and Apoptosis in a Mouse Model of Placental Insufficiency. Nutrients 2019, 11, 374. [Google Scholar] [CrossRef]
- Kwan, S.T. (Cecilia); King, J. H.; Yan, J.; Wang, Z.; Jiang, X.; Hutzler, J.S.; Klein, H.R.; Brenna, J.T.; Roberson, M.S.; Caudill, M.A. Maternal Choline Supplementation Modulates Placental Nutrient Transport and Metabolism in Late Gestation of Mouse Pregnancy. J Nutr 2017, 147, 2083–2092. [Google Scholar] [CrossRef]
- Caudill, M.A. Pre- and Postnatal Health: Evidence of Increased Choline Needs. J Am Diet Assoc 2010, 110, 1198–1206. [Google Scholar] [CrossRef]
- Molloy, A.M.; Mills, J.L.; Cox, C.; Daly, S.F.; Conley, M.; Brody, L.C.; Kirke, P.N.; Scott, J.M.; Ueland, P.M. Choline and Homocysteine Interrelations in Umbilical Cord and Maternal Plasma at Delivery. Am J Clin Nutr 2005, 82, 836–842. [Google Scholar] [CrossRef]
- Oliveira, C.P.M.S.; Da Costa Gayotto, L.C.; Tatai, C.; Della Bina, B.I.; Janiszewski, M.; Lima, E.S.; Abdalla, D.S.P.; Lopasso, F.P.; Laurindo, F.R.M.; Laudanna, A.A. Oxidative Stress in the Pathogenesis of Nonalcoholic Fatty Liver Disease, in Rats Fed with a Choline-Deficient Diet. J Cell Mol Med 2002, 6, 399–406. [Google Scholar] [CrossRef]
- Rasmussen, B.F.; Ennis, M.A.; Dyer, R.A.; Lim, K.; Elango, R. Glycine, a Dispensable Amino Acid, Is Conditionally Indispensable in Late Stages of Human Pregnancy. J Nutr 2020, 151, 361. [Google Scholar] [CrossRef] [PubMed]
- Cetin, I.; Marconi, A.M.; Baggiani, A.M.; Buscaglia, M.; Pardi, G.; Fennessey, P.V.; Battaglia, F.C. In Vivo Placental Transport of Glycine and Leucine in Human Pregnancies. Pediatric Research 1995, 37, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.M.; Godfrey, K.M.; Jackson, A.A.; Cameron, I.T.; Hanson, M.A. Low Serine Hydroxymethyltransferase Activity in the Human Placenta Has Important Implications for Fetal Glycine Supply. J Clin Endocrinol Metab 2005, 90, 1594–1598. [Google Scholar] [CrossRef] [PubMed]
- Holm, M.B.; Bastani, N.E.; Holme, A.M.; Zucknick, M.; Jansson, T.; Refsum, H.; Mørkrid, L.; Blomhoff, R.; Henriksen, T.; Michelsen, T.M. Uptake and Release of Amino Acids in the Fetal-Placental Unit in Human Pregnancies. PLoS One 2017, 12. [Google Scholar] [CrossRef]
- Wang, S.; Ning, J.; Huai, J.; Yang, H. Hyperglycemia in Pregnancy-Associated Oxidative Stress Augments Altered Placental Glucose Transporter 1 Trafficking via AMPKα/P38MAPK Signaling Cascade. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Muralimanoharan, S.; Maloyan, A.; Myatt, L. Mitochondrial Function and Glucose Metabolism in the Placenta with Gestational Diabetes Mellitus: Role of MiR-143. Clin Sci (Lond) 2016, 130, 931–941. [Google Scholar] [CrossRef]
- Song, T.R.; Su, G.D.; Chi, Y.L.; Wu, T.; Xu, Y.; Chen, C.C. Dysregulated MiRNAs Contribute to Altered Placental Glucose Metabolism in Patients with Gestational Diabetes via Targeting GLUT1 and HK2. Placenta 2021, 105, 14–22. [Google Scholar] [CrossRef]
- Hauguel, S.; Desmaizieres, V.; Challier, J.C. Glucose Uptake, Utilization, and Transfer by the Human Placenta as Functions of Maternal Glucose Concentration. Pediatr Res 1986, 20, 269–273. [Google Scholar] [CrossRef]
- Whatley, E.G.; Truong, T.T.; Wilhelm, D.; Harvey, A.J.; Gardner, D.K. β-Hydroxybutyrate Reduces Blastocyst Viability via Trophectoderm-Mediated Metabolic Aberrations in Mice. Hum Reprod 2022, 37, 1994–2011. [Google Scholar] [CrossRef]
- Tunster, S.J.; Watson, E.D.; Fowden, A.L.; Burton, G.J. REPRODUCTION REVIEW Placental Glycogen Stores and Fetal Growth: Insights from Genetic Mouse Models. 2020. [CrossRef]
- Kadam, L.; Veličković, M.; Stratton, K.; Nicora, C.D.; Kyle, J.E.; Wang, E.; Monroe, M.E.; Bramer, L.M.; Myatt, L.; Burnum-Johnson, K.E. Sexual Dimorphism in Lipidomic Changes in Maternal Blood and Placenta Associated with Obesity and Gestational Diabetes: A Discovery Study. Placenta 2025, 159, 76–83. [Google Scholar] [CrossRef]
- Johnson, R.K.; Soultanakis, R.P.; Matthews, D.E. Literacy and Body Fatness Are Associated with Underreporting of Energy Intake in US Low-Income Women Using the Multiple-Pass 24-Hour Recall: A Doubly Labeled Water Study. J Am Diet Assoc 1998, 98, 1136–1140. [Google Scholar] [CrossRef] [PubMed]
- Tran, K.; Johnson, R.K.; Soultanakis, R.P.; Matthews, D.E. In-Person vs Telephone-Administered Multiple-Pass 24-Hour Recalls in Women: Validation with Doubly Labeled Water. J Am Diet Assoc 2000, 100, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Sartorelli, D.S.; Crivellenti, L.C.; Manochio-Pina, M.G.; Baroni, N.F.; Carvalho, M.R.; Diez-Garcia, R.W.; Franco, L.J. Study Protocol Effectiveness of a Nutritional Intervention Based on Encouraging the Consumption of Unprocessed and Minimally Processed Foods and the Practice of Physical Activities for Appropriate Weight Gain in Overweight, Adult, Pregnant Women: A Randomized Controlled Trial. BMC Pregnancy Childbirth 2020, 20, 24. [Google Scholar] [CrossRef]
- Monteiro, C.A.; Cannon, G.; Moubarac, J.C.; Levy, R.B.; Louzada, M.L.C.; Jaime, P.C. The UN Decade of Nutrition, the NOVA Food Classification and the Trouble with Ultra-Processing. Public Health Nutr 2018, 21, 5–17. [Google Scholar] [CrossRef]
- Burton, G.J.; Sebire, N.J.; Myatt, L.; Tannetta, D.; Wang, Y.L.; Sadovsky, Y.; Staff, A.C.; Redman, C.W. Optimising Sample Collection for Placental Research. Placenta 2014, 35, 9–22. [Google Scholar] [CrossRef]
- Forcisi, S.; Moritz, F.; Kanawati, B.; Tziotis, D.; Lehmann, R.; Schmitt-Kopplin, P. Liquid Chromatography–Mass Spectrometry in Metabolomics Research: Mass Analyzers in Ultra High Pressure Liquid Chromatography Coupling. J Chromatogr A 2013, 1292, 51–65. [Google Scholar] [CrossRef]
- Hwang, T.L.; Shaka, A.J. Water Suppression That Works. Excitation Sculpting Using Arbitrary Wave-Forms and Pulsed-Field Gradients. J Magn Reson A 1995, 112, 275–279. [Google Scholar] [CrossRef]
- Carr, H.Y.; Purcell, E.M. Effects of Diffusion on Free Precession in Nuclear Magnetic Resonance Experiments. Physical Review 1954, 94, 630. [Google Scholar] [CrossRef]
- Ludwig, C.; Günther, U.L. MetaboLab--Advanced NMR Data Processing and Analysis for Metabolomics. BMC Bioinformatics 2011, 12. [Google Scholar] [CrossRef]
- Wishart DS, Guo A, Oler E, Wang F, Anjum A, Peters H et al. HMDB 5.0: the Human Metabolome Database for 2022. Nucleic Acids Research 2022, 50, D622–D631. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



