
Submitted:
04 July 2025
Posted:
07 July 2025
You are already at the latest version
Abstract
Fungi, from saprophytes to pathogens, face predictable daily fluctuations in light, temperature, humidity, and nutrient availability. To cope, they have evolved an internal circadian clock that confers a major adaptive advantage. This review argues that the fungal clock is not a passive timekeeper but a proactive regulatory hub orchestrating stress responses, metabolism, and pathogenesis. We summarize current knowledge of the core oscillator—centered on the FREQUENCY/WHITE COLLAR (FRQ/WCC) tran-scription-translation feedback loop in Neurospora crassa—and examine non-canonical timekeeping mechanisms in yeasts like Saccharomyces cerevisiae. We propose that the clock "gates" cellular defenses against oxidative, osmotic, and nutritional stress, ena-bling fungi to anticipate and withstand environmental insults. A central focus is the emerging concept of chrono-pathogenesis, in which the circadian regulation of viru-lence factors aligns fungal attacks with host vulnerability, as shown in Botrytis cinerea and Fusarium oxysporum. We explore the downstream pathways translating temporal signals into physiological outputs, including rhythmic gene expression, protein modi-fications, and chromatin remodeling. Finally, we highlight critical knowledge gaps and propose future research directions, including the development of "chrono-fungicides"—novel therapeutics that disrupt fungal clocks—as a promising strategy for managing fungal infections in agriculture and medicine.
Keywords:
circadian rhythm
; fungi
; stress response
; oxidative stress
; osmotic stress
; pathogenesis
; chronobiology
; Neurospora crassa
; clock-controlled genes
; chrono-fungicides
1. Introduction: The Fungal Clock as a Proactive Survival Mechanism
Endogenous circadian clocks, the internal timekeeping mechanisms found in nearly all domains of life, represent one of biology's most elegant solutions to a fundamental challenge: life on a rotating planet [1]. These oscillators generate self-sustaining rhythms with a period of approximately 24 hours, allowing organisms to synchronize their internal physiology with the predictable daily cycles of the external world [2]. However, the true power of the circadian clock lies not in its ability to simply keep time, but in its capacity to anticipate change. By generating an internal representation of the 24-hour day, the clock enables organisms to prepare for forthcoming environmental opportunities and threats, such as the rising sun, falling temperatures, or the activity patterns of hosts and competitors [3]. This proactive scheduling of biological processes, from metabolism to behavior, confers a significant fitness advantage over organisms that can only react to environmental stimuli after they occur [3]. The traditional view of stress response is often framed as a reactive process: a stimulus, such as high temperature or UV radiation, is detected and triggers a corresponding defensive pathway. The circadian clock introduces a fundamentally different paradigm—one of proactive defense. Instead of waiting for cellular damage to occur and then initiating a costly repair process, a clock-equipped organism can pre-emptively upregulate protective mechanisms just before the stressor is expected to arrive. For example, it can synthesize antioxidant enzymes before dawn to counter the oxidative burst caused by morning light or accumulate compatible solutes to prepare for the desiccation that accompanies midday heat. This anticipatory strategy is far more efficient, conserving energy and minimizing the cellular damage that would otherwise accumulate. This review will explore how fungi have harnessed this anticipatory power to master their environments. The fungal kingdom is characterized by extraordinary diversity in lifestyle and habitat, but a common thread is the constant exposure to a multitude of environmental stressors. For a filamentous fungus growing on a plant surface, the daily cycle presents a predictable gauntlet of challenges. Dawn brings intense light and ultraviolet (UV) radiation, which can cause direct DNA damage and generate harmful reactive oxygen species (ROS). The rising temperature throughout the day increases the risk of thermal damage and desiccation, imposing severe osmotic stress [4]. Concurrently, nutrient availability may fluctuate, and the fungus must compete with other microbes for limited resources [5]. For pathogenic fungi, the challenge is compounded by the host's own rhythms. The host immune system is not static; its defensive capacity waxes and wanes over the 24-hour cycle, creating windows of vulnerability that a savvy pathogen can exploit [6]. From saprophytes decomposing organic matter to pathogens invading a living host, fungi must navigate these rhythmic abiotic and biotic pressures to survive, grow, and reproduce [7]. This review synthesizes the rapidly accumulating evidence positioning the fungal circadian clock as a central regulatory hub that integrates diverse environmental signals with the internal metabolic state to orchestrate a coherent, anticipatory survival strategy. We will argue that the clock's influence extends far beyond simple developmental rhythms to encompass the direct control of cellular defense pathways, metabolic programming, and, critically, the temporal regulation of virulence in pathogenic species. We begin by examining the molecular architecture of the core clock machinery, contrasting the canonical model from Neurospora crassa with the divergent systems found in other fungi, and exploring how these clocks are synchronized with the environment. We then delve into the clock's role in gating specific stress responses, including oxidative, osmotic, and nutritional challenges. A major focus will be the emerging field of "chrono-pathogenesis," where we will dissect how fungal pathogens use their internal clocks to time their attack for maximum effect. Finally, we will explore the molecular output pathways that translate temporal information from the core oscillator into concrete physiological action, and we will conclude by identifying key knowledge gaps and proposing future research directions that could leverage the fungal clock as a novel target for disease control, with a special focus on the concept of "chrono-fungicides." As summarized in Figure 1, the fungal circadian clock (center) coordinates three major stress-response modules—oxidative, osmotic, and nutritional—by temporally gating their activation. This integrated model provides a conceptual framework for how timing enhances stress resilience under daily and environmental fluctuations.
2. The Core Oscillator: A Conserved Engine with Species-Specific Adaptations
The molecular machinery that generates circadian rhythmicity in fungi is a testament to both deep evolutionary conservation and remarkable niche-specific adaptation. The transcription-translation feedback loop (TTFL) first elucidated in the filamentous fungus Neurospora crassa has served as a powerful paradigm, yet studies in other fungi reveal a fascinating diversity of timekeeping strategies.
2.1. The Neurospora crassa Paradigm: The FRQ-WCC Oscillator
For decades, the bread mold Neurospora crassa has been the preeminent model for dissecting the eukaryotic circadian clock, revealing a mechanism centered on a negative feedback loop [8]. This canonical system, often referred to as the FREQUENCY/WHITE COLLAR (FRQ/WCC) oscillator, is composed of distinct positive and negative elements.
- ● Positive Arm: The primary positive-acting component is the White Collar Complex (WCC), a heterodimer of two GATA-type zinc-finger transcription factors, WHITE COLLAR-1 (WC-1) and WHITE COLLAR-2 (WC-2) [8]. WC-1 contains a Light-Oxygen-Voltage (LOV) domain, which functions as a blue-light photoreceptor, directly linking the clock to its most dominant environmental cue [8]. The WCC binds to specific promoter elements (Clock boxes or C-boxes) in its target genes to activate their transcription [9].
- ● Negative Arm: The masterstroke of the oscillator is that the WCC drives the transcription of its own inhibitor, the frequency (frq) gene [8]. The FRQ protein, upon translation in the cytoplasm, forms a complex with the FRQ-interacting RNA helicase (FRH) and casein kinases [3]. This complex then enters the nucleus and physically interacts with the WCC, repressing its transcriptional activity and thereby shutting down its own expression [8].
- ● Setting the Pace: The ~24-hour periodicity of the clock is not determined by the simple on/off switch but by a crucial delay mechanism. FRQ undergoes progressive, time-dependent phosphorylation by several kinases. This series of phosphorylation events governs its stability and its ability to inhibit the WCC. Once FRQ becomes hyperphosphorylated, it is targeted for degradation, which releases the WCC from inhibition and allows a new cycle of frq transcription to begin [3]. This elegant, phosphorylation-based time delay is the key to generating a robust, near-24-hour rhythm.
2.2. The Neurospora crassa Paradigm: The FRQ-WCC Oscillator
The FRQ-WCC model, while foundational, is not universal across the fungal kingdom. The presence, absence, or modification of this system across different fungal lineages provides profound clues about their distinct ecological strategies and evolutionary histories. The architecture of a fungus's clock appears to be a direct reflection of the most critical rhythmic variable in its environment. For fungi living on surfaces exposed to the sun, the primary entrainment cue is light. For those in fermentative environments, it is the feast-or-famine cycle of nutrient availability. For symbiotic fungi living underground, it is the rhythmic metabolic dialogue with their photosynthetic host. This demonstrates that the clock is not an arbitrary, conserved relic but a highly adapted tool for survival.
- ● Filamentous Ascomycetes: The FRQ-WCC system is remarkably well-conserved among many filamentous ascomycetes, particularly those with lifestyles exposed to daily environmental cycles. Crucially, this includes major plant pathogens. In Botrytis cinerea, the gray mold fungus, a functional clock with clear homologs of frq, wc-1, and wc-2 is essential for regulating virulence [10]. Similarly, the vascular wilt pathogen Fusarium oxysporum possesses multiple frq homologs and a WCC that are indispensable for its pathogenicity [11]. The conservation of this light-responsive clock in these pathogens underscores its fundamental importance for coordinating their infectious cycle with the external environment and the physiology of their plant hosts.
- ● The Enigma of the Yeast Clock: In stark contrast, the budding yeast Saccharomyces cerevisiae, a powerhouse of genetic research, lacks an obvious homolog of the core negative element frq [12]. For many years, this led to the assumption that yeast lacked a true circadian clock. However, pioneering work using continuous cultures (chemostats) revealed that yeast populations exhibit robust, temperature-compensated metabolic oscillations, notably in oxygen consumption, known as Yeast Respiratory Oscillations (YROs) [13]. These rhythms can be entrained by temperature cycles but damp out quickly in constant conditions, suggesting a less self-sustained oscillator compared to the Neurospora model [13]. This points to the existence of a non-canonical, FRQ-independent timekeeping mechanism in yeast, likely rooted in metabolic feedback loops rather than a dedicated TTFL. This architecture is well-suited to its typical fermentative lifestyle, where sporadic nutrient availability is a more pressing rhythmic challenge than light.
- ● Emerging Clocks in Symbionts: Perhaps the most intriguing recent discovery is the presence of the complete FRQ-WCC gene set in arbuscular mycorrhizal fungi (AMF), such as Rhizoglomus irregulare [14]. These fungi are obligate symbionts that live in the relatively dark and stable soil environment, forming intimate connections with plant roots. The presence of a "light-responsive" clock machinery in a non-photosynthetic, subterranean organism seems paradoxical. However, this strongly suggests that the clock is not entrained by light but by rhythmic signals from the host plant, namely the daily flux of photosynthates and other metabolites. The AMF clock likely serves to coordinate its metabolic activity—absorbing nutrients from the soil and exchanging them for carbon from the plant—with the host's own robust circadian rhythm, ensuring maximal efficiency for the symbiosis [15].
2.3. Entrainment: Synchronizing with the External World
For an internal clock to be useful, it must be synchronized, or entrained, to the external 24-hour day. Fungi use several environmental cues, known as zeitgebers (German for "time givers"), to achieve this.
- ● Light: As the most reliable environmental signal of the day-night cycle, light is the dominant entrainment cue for most surface-dwelling fungi. In the Neurospora model, the WCC's function as a direct blue-light photoreceptor provides an elegant mechanism for entrainment. A pulse of light rapidly induces frq transcription, and the effect on the clock's phase depends on when the pulse is received: a light pulse in the subjective evening delays the clock, while one in the late subjective night advances it, effectively aligning the internal rhythm with the external light cycle [2].
- ● Temperature: Temperature is another critical zeitgeber. Fungal clocks exhibit temperature compensation, a hallmark property of circadian systems, meaning the period of the rhythm remains relatively constant across a range of physiological temperatures [16]. This prevents the clock from running faster on warm days and slower on cool days, which would render it useless as a timekeeper. However, the clock is still sensitive to changes in temperature. Temperature cycles can entrain the rhythm, a feature that is likely crucial for subterranean or other fungi where light cues are weak or absent [17]. The molecular mechanisms for temperature compensation are complex but are thought to involve temperature-dependent changes in protein translation and localization that counterbalance each other to maintain a stable period [16]. Table 1 provides a comparative overview of the core clock machinery, highlighting the conserved paradigm and key evolutionary divergences that reflect niche-specific adaptations.
3. Circadian Gating of Cellular Defense: Preparing for the Inevitable
One of the most critical functions of the fungal circadian clock is the temporal regulation of stress response pathways. Rather than maintaining a constant state of high alert, which is metabolically expensive, the clock "gates" the expression and activity of defensive systems, deploying them only when they are most likely to be needed. This represents a profound optimization of cellular economy, a form of temporal budgeting where the clock creates a schedule of expected defensive expenditures, and other pathways, such as those sensing nutrients, act as an accounting check to ensure resources are available. This anticipatory defense is a key contributor to fungal fitness. The molecular underpinnings of circadian gating in stress responses are illustrated in Figure 2, with subpanels A–C detailing specific cascades: (A) activation of antioxidant genes via ROS-timed PP2A/WCC signaling, (B) anticipatory HOG MAPK phosphorylation preparing for osmotic stress, and (C) nutrient-deficiency adaptation through clock-modulated kinase/phosphatase networks.
3.1. Anticipating Oxidative Threats
Exposure to sunlight and metabolic activity inevitably generates reactive oxygen species (ROS), such as superoxide anions (O2−) and hydrogen peroxide (H2O2), which can damage DNA, proteins, and lipids [19]. The circadian clock plays a central role in managing this threat. In Neurospora, cellular ROS levels themselves have been shown to oscillate in a circadian manner [20]. This rhythm is actively managed by the clock through the timed expression of key antioxidant enzymes. The gene encoding catalase-1 (cat-1), an enzyme that neutralizes H2O2, is a well-characterized clock-controlled gene (ccg) in Neurospora [21]. Its transcript levels peak in anticipation of dawn, preparing the cell to detoxify the ROS that will be generated by light exposure. This principle of temporal defense is conserved across eukaryotes; the activities of other crucial antioxidant enzymes, including superoxide dismutase (SOD) and glutathione peroxidase (GPx), are also under circadian control in many organisms, highlighting a universal strategy of preparing for predictable oxidative insults [19].
3.2. Managing Osmotic and Desiccation Stress
For terrestrial fungi, the daily cycle of temperature and humidity imposes a predictable rhythm of osmotic stress. The primary defense against this is the High-Osmolarity Glycerol (HOG) pathway, a highly conserved mitogen-activated protein kinase (MAPK) cascade that leads to the intracellular accumulation of glycerol to counteract water loss [22]. In Saccharomyces cerevisiae, this pathway is activated by hyperosmotic shock [23]. In Neurospora, the clock has co-opted this pathway for anticipatory defense. The terminal MAPK of the cascade, OS-2, undergoes robust rhythmic phosphorylation that peaks around subjective dawn, even in the complete absence of any osmotic stress [21]. This clock-driven "pre-activation" of the HOG pathway primes the fungus for the desiccation stress that will accompany the rising sun and temperatures of the day [24]. This rhythmic phosphorylation of OS-2 in turn drives the rhythmic expression of downstream target genes, such as ccg-1, ensuring that the full defensive program is in place before it is critically needed [24].
3.3. Adapting to Nutritional Fluctuations
The circadian clock and cellular metabolism are deeply and bidirectionally intertwined. In Neurospora, global transcriptomic analyses have revealed that as much as 40% of the genome can be expressed under circadian control, with a profound enrichment for genes involved in metabolic pathways [3]. The clock appears to partition metabolism across the day, favoring catabolism and energy production during the active "day" phase and biosynthesis and growth during the "night" phase [3]. A critical molecular link that allows the clock to adjust to the cell's nutritional status has recently been uncovered. The evolutionarily conserved GCN2 signaling pathway, which is activated by amino acid starvation, is essential for maintaining a robust circadian rhythm under nutrient-poor conditions [9]. The mechanism involves the GCN2-regulated transcription factor CPC-1, which, under starvation, helps recruit the SAGA histone acetyltransferase complex to the frq promoter. This modulates histone H3 acetylation, ensuring that the promoter remains accessible for rhythmic binding by the WCC.20 This elegant mechanism directly couples the cell's real-time nutritional state to the function of the core oscillator, allowing the clock to run robustly when resources are available but potentially altering its output when the cell is starving.
3.4. Responding to Chemical Stress
The relationship between the circadian clock and the response to xenobiotic chemicals, such as fungicides, is an emerging area of research with significant practical implications. Studies in the medicinal fungus Cordyceps militaris have shown that exposure to various fungicides, including amphotericin B and terbinafine, can alter the expression level of the core clock protein CmFRQ [25]. This suggests that chemical stress can directly perturb the timekeeping machinery. Conversely, it is highly probable that the clock regulates detoxification pathways, such as those involving cytochrome P450 enzymes or efflux pumps, to anticipate exposure to naturally occurring toxins in the environment. This temporal regulation could have a significant impact on the efficacy of agricultural and clinical antifungal treatments.
4. The Chrono-Pathogenesis of Fungal Infections
The interaction between a pathogenic fungus and its host is a dynamic and intricate battle. Success for the pathogen depends not only on its arsenal of virulence factors but also on the timing of its attack. A growing body of evidence reveals that fungal pathogens possess their own circadian clocks that rhythmically control their virulence, a concept termed "chrono-pathogenesis" [26]. This internal timing allows the pathogen to coordinate its infection programs with the daily rhythms of its host, particularly the host's immune system, which itself exhibits strong circadian fluctuations [26]. This temporal coordination suggests a co-evolutionary arms race fought on the dimension of time, opening up novel avenues for disease control by disrupting this pathogenic timing.
4.1. A Timed Attack: Coordinating Virulence with Host Vulnerability
The outcome of a host-pathogen interaction is not a fixed property but varies dramatically depending on the time of day the infection occurs [10]. This is because both the pathogen's capacity to cause disease and the host's ability to defend itself are under circadian control. A successful pathogen has evolved to launch its attack not at random, but during a "window of opportunity" when its own virulence is at its peak and the host's defenses are at their nadir [26]. This temporal strategy maximizes the probability of establishing a successful infection. The discovery that the pathogen's clock is a key driver of this process has shifted the paradigm from viewing virulence as a static trait to seeing it as a rhythmic, carefully timed process.
4.2. Case Study: Botrytis cinerea on Arabidopsis thaliana
The interaction between the necrotrophic fungus Botrytis cinerea and its host plant Arabidopsis thaliana provides one of the most compelling demonstrations of chrono-pathogenesis. B. cinerea has a functional FRQ-WCC circadian clock that is essential for its rhythmic virulence [10]. Experiments have shown that the size of the necrotic lesions produced by the fungus on Arabidopsis leaves varies depending on the time of inoculation, with the most severe disease occurring when the fungus is at its subjective dawn (the peak of its activity cycle) [10]. Remarkably, elegant experiments using out-of-phase light cycles and various clock-null mutants of both the fungus and the plant have unequivocally demonstrated that it is the fungal clock, not the host clock, that is the primary determinant of this rhythm [10]. When wild-type fungi infect clock-deficient plants, the rhythm of virulence persists. Conversely, when clock-deficient fungi (e.g., a bcfrq1 mutant or fungi grown in constant light to abolish rhythmicity) infect wild-type plants, the rhythmic pattern of lesion size is completely abrogated, and the overall level of infection is significantly reduced [10]. This proves that a functional, oscillating clock in the pathogen is required to mount a fully virulent, timed attack.
4.3. Case Study: Fusarium oxysporum on Tomato
Similar principles govern the pathogenesis of the soil-borne fungus Fusarium oxysporum, a devastating vascular wilt pathogen. Its ability to cause disease in tomato plants is also under strict circadian control, with infections initiated at dawn resulting in the most severe symptoms [11]. As with Botrytis, the disruption of core clock genes, such as Fowc1, Fowc2, or the primary Fofrq1, renders the fungus almost completely nonpathogenic [11]. Time-series transcriptomics have revealed how the Fusarium clock orchestrates this timed attack. The clock rhythmically controls the expression of a suite of downstream transcription factors that, in turn, regulate specific virulence programs required to overcome host defenses [11]:
- ● Overcoming Zinc Starvation: During infection, plants actively sequester essential micronutrients like zinc to starve the invading pathogen. The Fusarium clock anticipates this defense by rhythmically driving the expression of the transcription factor FoZafA. FoZafA is essential for the fungus to adapt to the zinc-limited environment within the host plant, and its timed expression ensures this adaptation is active when needed most [11].
- ● Deploying Chemical Weapons: The clock also controls the production of phytotoxins. It rhythmically regulates the transcription factor FoCzf1, which governs the entire biosynthetic gene cluster for fusaric acid, a potent toxin that contributes to wilt symptoms. By timing the production of this chemical weapon, the fungus can deploy it for maximal impact [11].
4.4. The Host-Pathogen Temporal Dialogue
The success of chrono-pathogenesis relies on the fungus exploiting the host's own rhythms. The mammalian immune system, for example, is profoundly regulated by the circadian clock. Key processes such as the metabolism of tryptophan into the immunomodulatory kynurenine pathway, and the trafficking and activity of innate immune cells like neutrophils, exhibit robust daily oscillations [26]. This creates predictable times of day when the immune response is stronger or weaker. A pathogen whose clock has co-evolved with its host can time the expression of its virulence factors—such as those that evade immune detection or suppress immune responses—to coincide with these periods of host immunosuppression. This temporal interplay is a critical, yet often overlooked, dimension of the host-pathogen arms race. Understanding this dialogue is key to understanding disease outcomes and developing new therapeutic strategies. For instance, a "chrono-fungicide" that does not kill the pathogen but instead targets and disrupts its clock could be a revolutionary approach. By desynchronizing the pathogen from its host, it would be rendered "bad at its job," attacking at the wrong times when the host is fully prepared to defend itself. This strategy of sabotage, rather than direct assault, may be less susceptible to the development of resistance and could enhance the efficacy of existing antifungal agents.
5. Molecular Output Pathways: From Oscillation to Action
The core circadian oscillator, whether the canonical TTFL or a non-canonical variant, is ultimately a timekeeping device. For this timekeeping to have a physiological impact, its temporal information must be translated into rhythmic changes in cellular function. Fungi have evolved a sophisticated, multi-layered system of output pathways to achieve this, involving transcriptional cascades, post-translational modifications, and epigenetic regulation. This hierarchy of control ensures that the clock's output is both robust and flexible, allowing for the precise timing of diverse processes while also integrating feedback from other cellular states, such as nutrient availability.
5.1. Transcriptional Cascades: The WCC as a Master Regulator
The most direct output from the core FRQ-WCC oscillator is transcriptional. The WCC, as the primary clock-controlled transcription factor, binds to the promoters of a large set of clock-controlled genes (ccgs) to drive their rhythmic expression [3]. In Neurospora, this can encompass a substantial portion of the expressed genome [3]. A key feature of this network is that many of the direct targets of the WCC are themselves transcription factors [11]. This architecture creates downstream transcriptional cascades. The central clock rhythmically activates a secondary tier of regulatory proteins, which in turn control the expression of specific functional gene sets related to metabolism, development, or stress response [8]. This hierarchical structure allows a single central oscillator to generate complex and diverse phase relationships in the expression of thousands of downstream genes, providing a mechanism for the temporal orchestration of the entire cellular program.
5.2. Post-Translational Rhythms: Modulating Protein Activity
The clock's influence is not limited to the level of transcription. It can also impose rhythmicity on post-translational events, allowing for rapid modulation of protein activity without the delay of new gene expression and translation. The rhythmic activation of the OS-2 MAPK in Neurospora is a prime example [21]. The clock directs the rhythmic phosphorylation of OS-2, meaning that the activity of this key stress-signaling kinase oscillates throughout the day, even in constant conditions. This rhythmic signal can then propagate further down the pathway. It has been shown that the OS-2 pathway leads to the phosphorylation of eukaryotic elongation factor-2 (eEF-2), a critical regulator of the elongation step in protein synthesis [27]. As a result, the overall translational activity of the cell exhibits a circadian rhythm, peaking at a different time than transcription. This demonstrates a powerful layer of clock control that gates the final step of gene expression—the production of proteins—ensuring that even constitutively transcribed mRNAs may only be translated at specific times of day [27].
5.3. Chromatin and Epigenetic Regulation
A third layer of control involves the dynamic, rhythmic regulation of the chromatin environment. The accessibility of gene promoters to transcription factors is a key determinant of gene expression, and this accessibility is itself under circadian control. As discussed previously, the link between the GCN2 nutrient-sensing pathway and the circadian clock in Neurospora is mediated by chromatin modification [9]. Under amino acid starvation, the CPC-1 transcription factor recruits the SAGA complex to the frq promoter to ensure rhythmic histone H3 acetylation [9]. This epigenetic mark keeps the chromatin in an "open" state, allowing the WCC to bind rhythmically and maintain the clock's oscillation. This reveals that the clock's own operation is subject to epigenetic control that integrates information about the cell's metabolic state. It is highly likely that this principle extends to the clock's output, with the clock rhythmically directing chromatin modifications at the promoters of ccgs to fine-tune their expression. This epigenetic layer adds a crucial dimension of flexibility and robustness to the clock's control over the genome. This multi-layered system—transcriptional, post-translational, and epigenetic—explains both the clock's robustness and its flexibility. The redundancy ensures that if one layer is perturbed, others can compensate. The integration of signals like nutrient status at the epigenetic level allows the entire system's output to be modulated without re-engineering the core timekeeping loop, transforming the clock from a simple on/off switch into a sophisticated rheostat. Table 2 systematically connects specific stresses and pathogenic functions to the known molecular players regulated by the circadian clock, serving as a central reference for the key output pathways.
6. Discussion
The evidence synthesized in this review paints a clear picture of the fungal circadian clock as a master regulatory system, fundamental to survival and success. It is far more than a simple mechanism for timing developmental events like sporulation. The clock is a central coordinator that integrates external environmental cues with the internal physiological state of the cell to generate a proactive, anticipatory program for life. By rhythmically gating cellular defenses, the clock prepares the fungus for predictable daily onslaughts of oxidative, thermal, and osmotic stress, optimizing resource allocation and minimizing cellular damage. By partitioning metabolic activities across the 24-hour day, it ensures maximal efficiency in nutrient utilization and growth. And in pathogenic species, the clock has been weaponized, transforming into a tool for chrono-pathogenesis that times virulence programs to exploit temporal weaknesses in host defenses. The multi-layered output pathways—from transcriptional cascades to post-translational and epigenetic modifications—provide the clock with a robust and flexible means to exert its influence over virtually every aspect of fungal biology. In any complex ecosystem, organisms compete for resources in space. Figure 3 depicts the ecological impact of circadian coherence: fungi with synchronized clocks exhibit improved tolerance to heat, drought, and salinity, while clock disruption (e.g., due to climate stressors) predicts reduced resilience. Comparative adaptations in fungal clock genes across altitudes support this model. The circadian clock allows them to also compete in, and partition, the dimension of time. By scheduling key activities for specific times of day, fungi can occupy a unique temporal niche. For example, the clock-controlled release of spores by Neurospora just before dawn, when humidity is high and temperatures are cool, maximizes their dispersal and viability. A fungus that forages for nutrients at night may avoid competition from diurnal organisms. This temporal partitioning reduces direct competitive pressure and minimizes exposure to predictable threats, such as diurnal predators or the peak UV radiation of midday. In the context of a holobiont, such as the mycorrhizal symbiosis, the clock's role in temporal coordination becomes even more critical, ensuring that the metabolic activities of the fungus and its host plant are synchronized for their mutual benefit. The ability to claim and defend a temporal niche is a powerful ecological strategy that has undoubtedly contributed to the evolutionary success of the fungal kingdom. Our understanding of the fungal circadian clock has advanced significantly, moving from the initial characterization of the core oscillator in Neurospora crassa to a broader appreciation of its central role in fungal biology. It is now established that the clock is a proactive, anticipatory system that provides a major fitness advantage. Its role in gating cellular responses to abiotic stresses like oxidative and osmotic shock is well-documented in model systems.
Furthermore, the paradigm of chrono-pathogenesis, where the fungal clock rhythmically controls virulence to exploit host rhythms, is firmly established in key plant pathogens. The molecular output pathways are beginning to be mapped, revealing a complex hierarchy of transcriptional, post-translational, and epigenetic control. Despite this progress, many fundamental questions remain unanswered, highlighting exciting frontiers for future research. The molecular basis of the clock is understood in only a handful of filamentous ascomycetes. The mechanisms of timekeeping in the vast majority of the fungal kingdom, including the entire phylum Basidiomycota (mushrooms, rusts, smuts), remain a black box. While genomic surveys confirm the presence of core clock gene homologs (wc-1, wc-2, frq) in many basidiomycetes, their functionality is largely unproven [28]. Intriguing evidence of rhythmic behaviors, such as the clock-controlled bioluminescence in the mushroom Neonothopanus gardneri and sporulation rhythms in Pellicularia filamentosa, strongly suggests that functional clocks are present [29]. Elucidating the molecular machinery in these economically and ecologically important fungi is a major priority. In animals and cyanobacteria, robust circadian rhythms in the oxidation state of peroxiredoxin proteins have been identified, which can persist even without a functional TTFL. These NTOs represent a more ancient form of timekeeping. Whether similar TTFL-independent oscillators exist in fungi and how they might interact with or be coupled to the canonical FRQ-WCC clock is a major unanswered question [30]. The respiratory oscillations in yeast, which lack core TTFL components, may be a manifestation of such a system and share features with mammalian rhythms, suggesting a deeply conserved ancestral mechanism [11]. While transcriptomic studies have identified hundreds of clock-controlled genes, this is only the first step. The complete network of clock outputs, including the full complement of rhythmically modified proteins (the "chrono-proteome") and metabolites (the "chrono-metabolome"), is far from mapped. Understanding this complete network is essential to grasp the full physiological impact of the clock. The discovery of clock genes in non-photosynthetic, subterranean arbuscular mycorrhizal fungi (AMF) is tantalizing [14]. It strongly implies that the fungal clock is entrained by rhythmic signals from its host plant, likely related to the daily flux of photosynthates [31]. However, the molecular nature of this temporal communication—the specific signals sent by the plant and the receptors used by the fungus—is completely unknown. Deciphering this symbiotic dialogue, which governs the temporal dynamics of nutrient exchange, is a key challenge for understanding these critical ecological partnerships [31]. Addressing these knowledge gaps will require a multi-pronged approach leveraging cutting-edge technologies:
- Comparative Genomics and Transcriptomics: Applying long-read sequencing and time-series RNA-seq to a much broader phylogenetic diversity of fungi, particularly within Basidiomycota and early-diverging lineages, will be crucial for identifying novel clock components and uncovering new clock architectures.
- Chrono-Proteomics and -Metabolomics: The use of high-resolution mass spectrometry to perform time-series analyses of the entire proteome, phosphoproteome (to identify rhythmic kinase activity), and metabolome of fungi grown in constant conditions will be essential for mapping the full extent of the clock's output pathways.
- Structural Biology: Determining the high-resolution 3D structures of key clock protein complexes, such as the WCC and the FRQ-FRH complex, using techniques like cryo-electron microscopy. This will provide deep mechanistic insights into their function and reveal potential pockets for the rational design of small-molecule inhibitors ("chrono-fungicides").
- In Vivo Infection Models: Developing and employing advanced imaging techniques, such as live-cell microscopy with dual-reporter systems (e.g., fluorescent reporters for both fungal clock phase and host immune gene activation). This will allow researchers to dissect the temporal dynamics of the host-pathogen interaction in real-time and to precisely determine the contribution of each partner's clock to the outcome of the infection.
By pursuing these avenues, the scientific community can move toward a comprehensive understanding of this fundamental aspect of fungal biology, paving the way for innovative solutions to long-standing challenges in agriculture and human health.
7. Conclusions
The fungal circadian clock has emerged as a central regulator of stress responses across diverse environmental conditions. Far from functioning solely as a developmental timer, it operates as a proactive system that anticipates and mitigates predictable stressors, including oxidative, osmotic, thermal, and nutritional challenges. By rhythmically gating defense pathways, modulating cell wall dynamics, and temporally organizing gene expression, the clock significantly enhances fungal resilience. In an era of accelerating climate variability, the alignment between internal circadian rhythms and external environmental cycles can determine whether a fungus thrives or fails. This temporal synchronization optimizes resource use, minimizes cellular damage, and confers a competitive ecological advantage. In pathogenic fungi, the clock has further evolved into a tool for chrono-pathogenesis, coordinating the timing of virulence factor expression to exploit host vulnerabilities. This concept reframes our understanding of host-pathogen dynamics under stress and points to new therapeutic avenues. Overall, the circadian clock represents a powerful molecular framework that links environmental perception with stress adaptation. Its study not only deepens our understanding of fungal biology but also offers novel strategies to enhance stress tolerance and disease control in both agricultural and clinical settings.
Author Contributions
“Conceptualization, V.C-R.; methodology, V.C-R.; formal analysis, V.C-R..; investigation, V.C-R..; resources, V.C-R. and D.B-R. .; data curation, V.C-R.; writing—original draft preparation, V.C-R.; writing—review and editing, V.C-R..; visualization, V.C-R. and D.B-R.; supervision, V.C-R..; project administration, V.C-R.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
All the research data can be found in the text.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| FRQ | FREQUENCY |
| WCC | WHITE COLLAR COMPLEX |
| TTFL | Transcription-Translation Feedback Loop |
| ROS | Reactive Oxygen Species |
| YROs | Yeast Respiratory Oscillations |
| AMF | Arbuscular Mycorrhizal Fungi |
| MAPK | Mitogen-Activated Protein Kinase |
| PP2A | Protein Phosphatase 2A |
| eEF-2 | Eukaryotic Elongation Factor 2 |
| ccg | Clock-Controlled Genes |
| NTOs | Non-Transcriptional Oscillators |
| GCN2 | General Control Nonderepressible 2 |
| CPC-1 | Cross-Pathway Control 1 |
| SAGA | Spt-Ada-Gcn5 Acetyltransferase (complex) |
References
- Konakchieva, R.; Mladenov, M.; Konaktchieva, M.; Sazdova, I.; Gagov, H.; Nikolaev, G. Circadian Clock Deregulation and Metabolic Reprogramming: A System Biology Approach to Tissue-Specific Redox Signaling and Disease Development. International Journal of Molecular Sciences 2025, 26, 6267. [Google Scholar] [CrossRef]
- Bell-Pedersen, D.; Garceau, N.; Loros, J. Circadian Rhythms in Fungi. Journal of Genetics 1996, 75, 387–401. [Google Scholar] [CrossRef]
- Hurley, J.M.; Dasgupta, A.; Emerson, J.M.; Zhou, X.; Ringelberg, C.S.; Knabe, N.; Lipzen, A.M.; Lindquist, E.A.; Daum, C.G.; Barry, K.W.; et al. Analysis of Clock-Regulated Genes in Neurospora Reveals Widespread Posttranscriptional Control of Metabolic Potential. Proceedings of the National Academy of Sciences 2014, 111, 16995–17002. [Google Scholar] [CrossRef]
- Chaffai, R.; Ganesan, M.; Cherif, A. Mechanisms of Plant Response to Heat Stress: Recent Insights. In Plant Adaptation to Abiotic Stress: From Signaling Pathways and Microbiomes to Molecular Mechanisms; Chaffai, R., Ganesan, M., Cherif, A., Eds.; Springer Nature: Singapore, 2024; pp. 83–105 ISBN 978-981-97-0672-3.
- Ullah, F.; Ali, S.; Siraj, M.; Akhtar, M.S.; Zaman, W. Plant Microbiomes Alleviate Abiotic Stress-Associated Damage in Crops and Enhance Climate-Resilient Agriculture. Plants 2025, 14, 1890. [Google Scholar] [CrossRef]
- Shchaslyvyi, A.Y.; Antonenko, S.V.; Telegeev, G.D. Comprehensive Review of Chronic Stress Pathways and the Efficacy of Behavioral Stress Reduction Programs (BSRPs) in Managing Diseases. Int J Environ Res Public Health 2024, 21, 1077. [Google Scholar] [CrossRef] [PubMed]
- Shelake, R.M.; Wagh, S.G.; Patil, A.M.; Červený, J.; Waghunde, R.R.; Kim, J.-Y. Heat Stress and Plant–Biotic Interactions: Advances and Perspectives. Plants 2024, 13, 2022. [Google Scholar] [CrossRef]
- Dunlap, J.C.; Loros, J.J. Making Time: Conservation of Biological Clocks from Fungi to Animals. Microbiology Spectrum 2017, 5, 10.1128/microbiolspec.funk-0039–2016. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.; Shen, Q.; Yu, Z. Fungi Employ the GCN2 Pathway to Maintain the Circadian Clock under Amino Acid Starvation. TIL 2023, 1, 100026–2. [Google Scholar] [CrossRef]
- Hevia, M.A.; Canessa, P.; Müller-Esparza, H.; Larrondo, L.F. A Circadian Oscillator in the Fungus Botrytis Cinerea Regulates Virulence When Infecting Arabidopsis Thaliana. Proc Natl Acad Sci U S A 2015, 112, 8744–8749. [Google Scholar] [CrossRef]
- Lu, Q.; Yu, M.; Sun, X.; Zhou, X.; Zhang, R.; Zhang, Y.; Liu, X.-L.; Li, Z.; Cai, L.; Liu, H.; et al. Circadian Clock Is Critical for Fungal Pathogenesis by Regulating Zinc Starvation Response and Secondary Metabolism. Sci Adv 11, eads1341. [CrossRef]
- Merrow, M.; and Raven, M. Finding Time: A Daily Clock in Yeast. Cell Cycle 2010, 9, 1671–1672. [Google Scholar] [CrossRef]
- Eelderink-Chen, Z.; Mazzotta, G.; Sturre, M.; Bosman, J.; Roenneberg, T.; Merrow, M. A Circadian Clock in Saccharomyces Cerevisiae. Proceedings of the National Academy of Sciences 2010, 107, 2043–2047. [Google Scholar] [CrossRef]
- Simon, N.M.L.; Dodd, A.N. A New Link between Plant Metabolism and Circadian Rhythms? Plant, Cell & Environment 2017, 40, 995–996. [Google Scholar] [CrossRef]
- Lee, S.-J.; Morse, D.; Hijri, M. Holobiont Chronobiology: Mycorrhiza May Be a Key to Linking Aboveground and Underground Rhythms. Mycorrhiza 2019, 29, 403–412. [Google Scholar] [CrossRef]
- Tseng, Y.-Y.; Hunt, S.M.; Heintzen, C.; Crosthwaite, S.K.; Schwartz, J.-M. Comprehensive Modelling of the Neurospora Circadian Clock and Its Temperature Compensation. PLOS Computational Biology 2012, 8, e1002437. [Google Scholar] [CrossRef] [PubMed]
- Sulzman, F.M.; Ellman, D.; Fuller, C.A.; Moore-Ede, M.C.; Wassmer, G. Neurospora Circadian Rhythms in Space: A Reexamination of the Endogenous-Exogenous Question. Science 1984, 225, 232–234. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.L.; Loros, J.J.; Dunlap, J.C. The Circadian Clock of Neurospora Crassa. FEMS Microbiol Rev 2012, 36, 95–110. [Google Scholar] [CrossRef]
- Wilking, M.; Ndiaye, M.; Mukhtar, H.; Ahmad, N. Circadian Rhythm Connections to Oxidative Stress: Implications for Human Health. Antioxid Redox Signal 2013, 19, 192–208. [Google Scholar] [CrossRef]
- Yoshida, Y.; Iigusa, H.; Wang, N.; Hasunuma, K. Cross-Talk between the Cellular Redox State and the Circadian System in Neurospora. PLOS ONE 2011, 6, e28227. [Google Scholar] [CrossRef]
- Lamb, T.M.; Finch, K.E.; Bell-Pedersen, D. The Neurospora Crassa OS MAPK Pathway-Activated Transcription Factor ASL-1 Contributes to Circadian Rhythms in Pathway Responsive Clock-Controlled Genes. Fungal Genet Biol 2012, 49, 180–188. [Google Scholar] [CrossRef]
- Gonzalez, R.; Morales, P.; Tronchoni, J.; Cordero-Bueso, G.; Vaudano, E.; Quirós, M.; Novo, M.; Torres-Pérez, R.; Valero, E. New Genes Involved in Osmotic Stress Tolerance in Saccharomyces Cerevisiae. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Rep, M.; Reiser, V.; Gartner, U.; Thevelein, J.M.; Hohmann, S.; Ammerer, G.; Ruis, H. Osmotic Stress-Induced Gene Expression in Saccharomyces Cerevisiae Requires Msn1p and the Novel Nuclear Factor Hot1p. Mol Cell Biol 1999, 19, 5474–5485. [Google Scholar] [CrossRef] [PubMed]
- Vitalini, M.; De Paula, R.; Goldsmith, C.; Jones, C.; Borkovich, K.; Bell-Pedersen, D. Circadian Rhythmicity Mediated by Temporal Regulation of the Activity of P38 MAPK. Proceedings of the National Academy of Sciences of the United States of America 2007, 104, 18223–18228. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.-M.; Zhang, D.-D.; Huang, Z.-Y.; Fu, M.-J. The Stress of Fungicides Changes the Expression of Clock Protein CmFRQ and the Morphology of Fruiting Bodies of Cordyceps Militaris. Journal of Fungi 2024, 10, 150. [Google Scholar] [CrossRef]
- Costantini, C.; Renga, G.; Sellitto, F.; Borghi, M.; Stincardini, C.; Pariano, M.; Zelante, T.; Chiarotti, F.; Bartoli, A.; Mosci, P.; et al. Microbes in the Era of Circadian Medicine. Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef]
- Caster, S.Z.; Castillo, K.; Sachs, M.S.; Bell-Pedersen, D. Circadian Clock Regulation of mRNA Translation through Eukaryotic Elongation Factor eEF-2. Proc Natl Acad Sci U S A 2016, 113, 9605–9610. [Google Scholar] [CrossRef] [PubMed]
- Valim, H.F.; Dal Grande, F.; Otte, J.; Singh, G.; Merges, D.; Schmitt, I. Identification and Expression of Functionally Conserved Circadian Clock Genes in Lichen-Forming Fungi. Sci Rep 2022, 12, 15884. [Google Scholar] [CrossRef]
- Kalangutkar, J.; Kamat, N. Some Studies on Circadian Rhythm in the Culture of Omphalina Quelet Sp. (Basidiomycota, Agaricales) A Mycobiont of an Unidentified Basidiolichen. Nat Prec 2010, 1–1. [CrossRef]
- Causton, H.C.; Feeney, K.A.; Ziegler, C.A.; O’Neill, J.S. Metabolic Cycles in Yeast Share Features Conserved among Circadian Rhythms. Curr Biol 2015, 25, 1056–1062. [Google Scholar] [CrossRef]
- Mycorrhizal symbioses. Available online: https://www.periodicos.capes.gov.br/index.php/acervo/buscador.html?task=detalhes&id=W4246537531 (accessed on 1 July 2025).
Figure 1.
Comprehensive conceptual model of the clock and stress. This figure illustrates the central interconnection between the fungal circadian clock (represented by the FRQ–WC1–WC2 complex) and the main stress response pathways. The clock acts as a master regulator that temporally coordinates defenses against (A) oxidative stress (ROS), (B) osmotic stress (mediated by the HOG-MAPK pathway), and nutritional stress (such as zinc deficiency). This model provides a visual map of how circadian synchronization is fundamental to the fungus's resilience and adaptation to a fluctuating environment.
Figure 1.
Comprehensive conceptual model of the clock and stress. This figure illustrates the central interconnection between the fungal circadian clock (represented by the FRQ–WC1–WC2 complex) and the main stress response pathways. The clock acts as a master regulator that temporally coordinates defenses against (A) oxidative stress (ROS), (B) osmotic stress (mediated by the HOG-MAPK pathway), and nutritional stress (such as zinc deficiency). This model provides a visual map of how circadian synchronization is fundamental to the fungus's resilience and adaptation to a fluctuating environment.
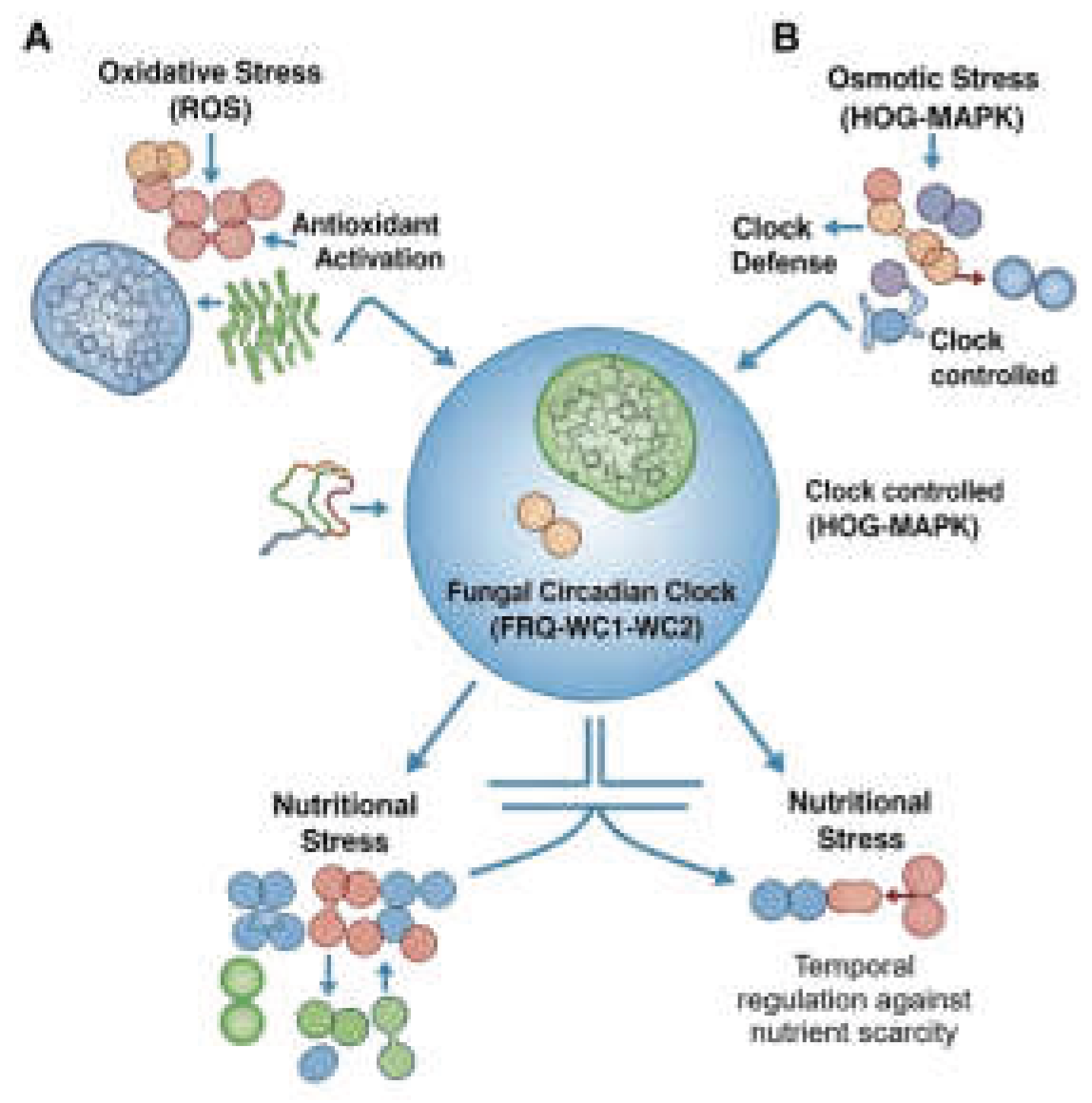
Figure 2.
Stress-specific molecular cascades. This figure details the molecular mechanisms connecting the circadian clock to specific stress responses in three subpanels. (A) Oxidative stress: It shows how reactive oxygen species (ROS) influence the central oscillator to activate antioxidant genes. (B) Osmotic stress: It illustrates the regulation of the HOG-MAPK pathway by the clock to manage cell wall integrity. (C) Nutritional/Pathogen stress: It represents how the clock temporally controls nutrient acquisition pathways and virulence factors, synchronizing these functions with external or host conditions.
Figure 2.
Stress-specific molecular cascades. This figure details the molecular mechanisms connecting the circadian clock to specific stress responses in three subpanels. (A) Oxidative stress: It shows how reactive oxygen species (ROS) influence the central oscillator to activate antioxidant genes. (B) Osmotic stress: It illustrates the regulation of the HOG-MAPK pathway by the clock to manage cell wall integrity. (C) Nutritional/Pathogen stress: It represents how the clock temporally controls nutrient acquisition pathways and virulence factors, synchronizing these functions with external or host conditions.
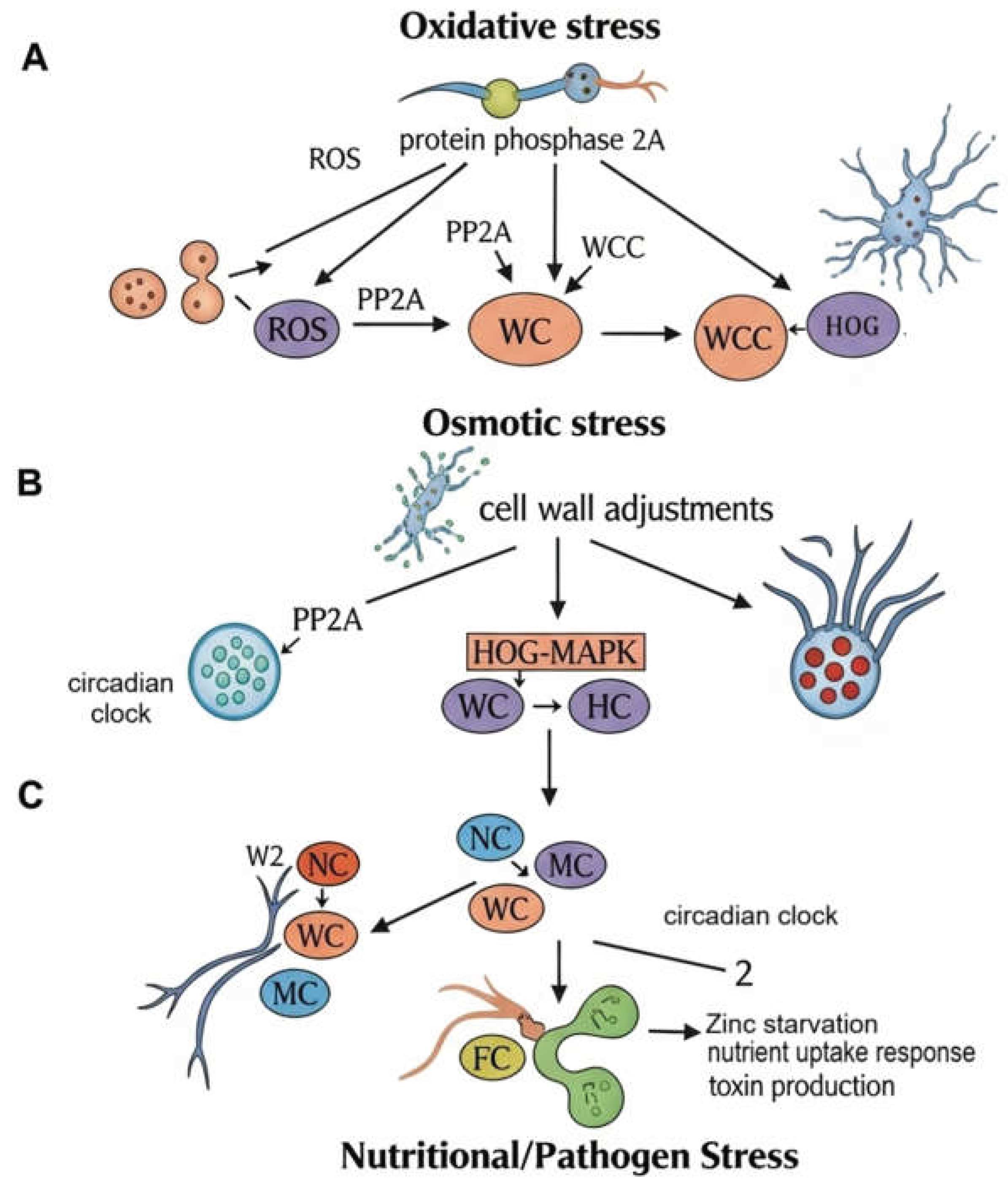
Figure 3.
Environmental impact and resilience to climate change. This scenario diagram illustrates the ecological implications of the circadian clock's function. Two paths are compared: on the left (Resilience), a synchronized clock allows fungi to anticipate environmental changes (e.g., temperature), adapting their phenology to survive and thrive. On the right (Vulnerability), the disruption of the clock by climate change causes desynchronization, which increases the fungus's vulnerability to stress and threatens its function in the ecosystem, with potential repercussions for biodiversity and agriculture.
Figure 3.
Environmental impact and resilience to climate change. This scenario diagram illustrates the ecological implications of the circadian clock's function. Two paths are compared: on the left (Resilience), a synchronized clock allows fungi to anticipate environmental changes (e.g., temperature), adapting their phenology to survive and thrive. On the right (Vulnerability), the disruption of the clock by climate change causes desynchronization, which increases the fungus's vulnerability to stress and threatens its function in the ecosystem, with potential repercussions for biodiversity and agriculture.
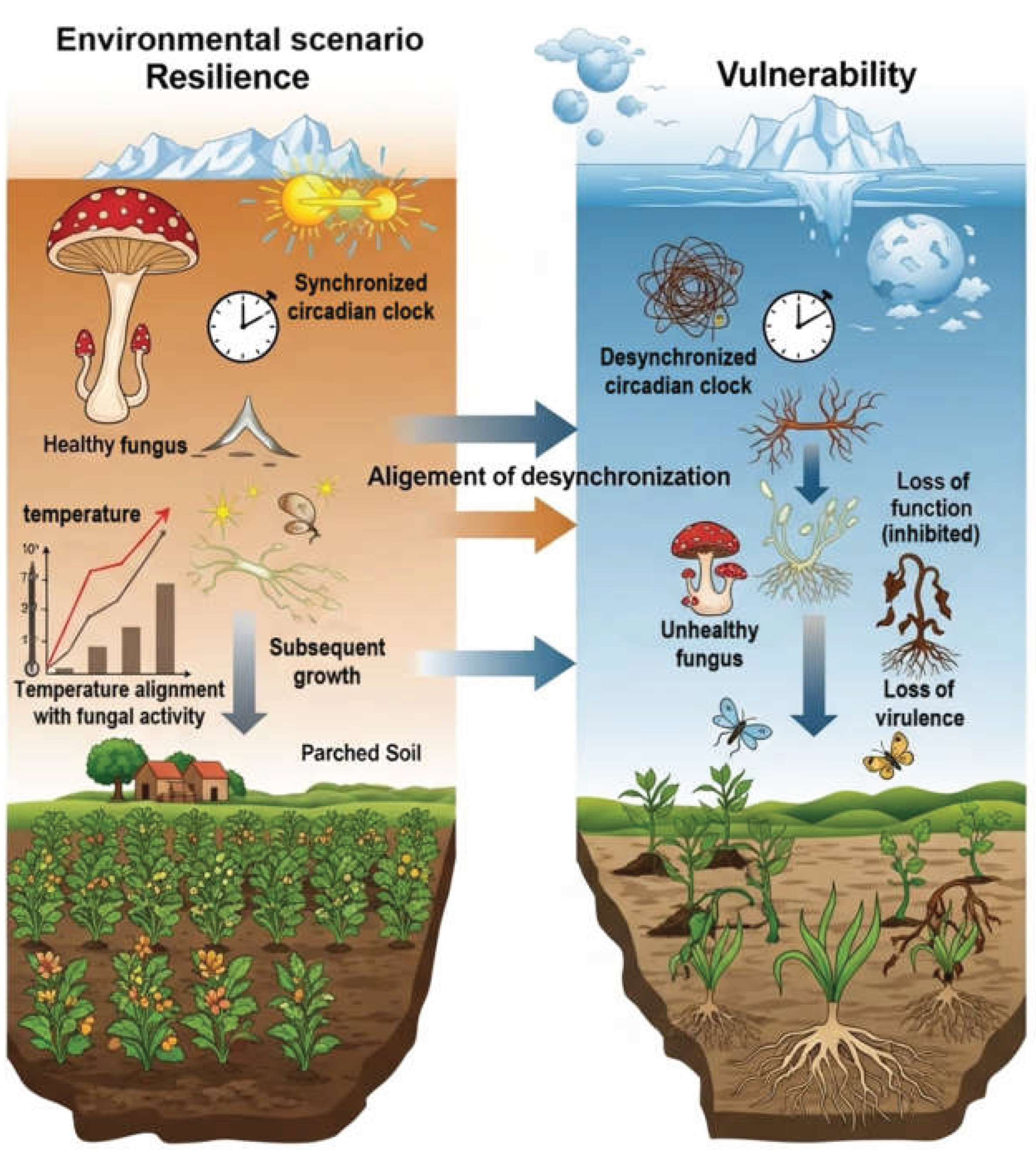

Table 1.
Conservation and function of core circadian clock components across fungal species.
| Fungus | WC-1/WC-2 Homologs (Presence/Function) | FRQ Homolog(s) (Presence/Function) | Primary Entrainment Cue(s) | Key Rhythmic Output |
|---|---|---|---|---|
| Neurospora crassa | Present / Positive element, photoreceptor [8] | Present / Negative element [8] | Light, Temperature [8] | Asexual sporulation (conidiation), Stress response [18] |
| Botrytis cinerea | Present / Positive element homologs [10] | Present / Negative element, virulence regulator [10] | Light [10] | Virulence, Pathogenesis [10] |
| Fusarium oxysporum | Present / Positive element homologs essential for virulence [11] | Present / Primary negative element essential for virulence [11] | Light, Host signals (inferred) | Virulence, Toxin production, Zinc homeostasis [11] |
| Saccharomyces cerevisiae | Absent [12] | Absent [12] | Temperature, Metabolic cycles [12] | Respiratory oscillations (YROs), Gene expression [13] |
| Rhizoglomus irregulare | Present / Expressed in pre- and post-symbiotic stages [14] | Present / Expressed in pre- and post-symbiotic stages [14] | Host metabolic signals (hypothesized) [15] | Coordination with host plant physiology (hypothesized) [15] |
Table 2.
Clock-Regulated Stress Response Pathways and Virulence Factors in Fungi.
| Stress/Function | Key Clock-Controlled Gene/Protein | Mechanism of Regulation | Fungal Species | Reference(s) |
|---|---|---|---|---|
| Oxidative Stress | Catalase-1 (cat-1) | Transcriptional regulation by WCC | Neurospora crassa | [21] |
| Osmotic Stress | OS-2 (MAPK) | Rhythmic phosphorylation (activation) | Neurospora crassa | [21] |
| Osmotic Stress | ccg-1 (osmotic-responsive gene) | Transcriptional regulation downstream of rhythmic OS-2 activation | Neurospora crassa | [24] |
| Nutritional Stress | frequency (frq) | Epigenetic (histone acetylation) via GCN2/CPC-1/SAGA pathway | Neurospora crassa | [9] |
| Virulence | bcfrq1 (clock core component) | Required for rhythmic virulence | Botrytis cinerea | [10] |
| Zinc Starvation | FoZafA (Transcription Factor) | Rhythmic transcription regulated by the clock | Fusarium oxysporum | [11] |
| Toxin Production | FoCzf1 (Transcription Factor) | Rhythmic transcription regulated by the clock | Fusarium oxysporum | [11] |
| Translation | eEF-2 (Elongation Factor) | Rhythmic phosphorylation downstream of rhythmic OS-2 activation | Neurospora crassa | [27] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



