
Submitted:
03 July 2025
Posted:
03 July 2025
You are already at the latest version
Abstract
Background: Biofilms consist of complex microbial communities embedded in an extra-cellular matrix which confer resistance to the most used antimicrobial agents. Chronic wounds are often associated with burns, trauma, surgery, diabetes and peripheral vascu-lar disease. They are characterized by a marked delay in wound healing favoring the de-velopment of microbial biofilms, which in turn further delay tissue regeneration. Staphy-lococcus aureus, Staphylococcus epidermidis, and methicillin-resistant staphylococci biofilms are found in chronic wounds, seriously hindering wound treatment. Vitreoscilla filiformis, a Gram-negative non-pathogenic filamentous bacterium, has been shown to improve atopic dermatitis by reducing S. aureus colonization and inducing antioxidant responses in the skin. Methods: The effect of V. filiformis supernatant (VFS) on bacteria growth was assessed by microbial growth kinetics and on biofilm formation and dispersal. Antioxi-dant potential was determined by DPPH scavenging ability and reduction of intracellular reactive oxygen species (ROS). The regenerative properties were assessed by scratch assay. Results: V. filiformis VFS holds strong anti-biofilm activity against S. aureus, S. epidermidis and methicillin resistant S. aureus (MRSA), acting during both biofilm formation and dis-persion. The decrease in biofilm mass is accompanied by a significant increase in the planktonic form compared to the untreated cells. Moreover, VFS is characterized by an in-teresting antioxidant activity, as demonstrated by a cell-free DPPH assay and a neutro-phil-based in vitro assay. In addition, VFS can stimulate tissue regeneration in human dermal fibroblasts. Conclusions: The demonstrated anti-biofilm, antioxidant and regener-ative properties of V. filiformis supernatant could be exploited for the treatment of biofilm-associated wound infections.
Keywords:
Vitreoscilla filiformis supernatant
; anti-biofilm
; regenerative properties
; wound infections treatment
1. Introduction
Human skin, covering roughly 30 m2 on average, provides a vast habitat for a diverse community of microorganisms. Sampling shows that the number of these microorganisms can range from 103 to 106 per site [1]. Notably, its bacterial population is mainly derived from four key phyla Actinomycetota, Bacillota, Pseudomonadota and Bacteroidota, with genera such as Staphylococcus, Cutibacterium, Corynebacterium, Micrococcus, Streptococcus and Acinetobacter being particularly prevalent [2].
The human skin is colonized by millions of bacteria who are primarily acquired during birth and continuously subjected to deep modifications during life. Under “steady state” conditions, host factors and interactions between microorganisms define a relative stability of the microbial communities preventing the colonization by pathogens [3]. These bacterial communities form the skin microbiota, a critical contributor to host health by enhancing immune defences and promoting tissue regeneration [4]. However, when dysbiosis occurs, bacteria that normally reside in the skin as commensals may become pathogenic and colonization by pathogens might be favoured.
Changes in the composition of the skin microbiota have been demonstrated to occur in many cutaneous diseases. In atopic dermatitis (AD), the proportion of Staphylococcus epidermidis and Staphylococcus aureus significantly increases during disease flares [5]; the proliferation of staphylococci is partly attributed to their ability to bind to a compromised skin barrier, such as fibronectin in the upper epidermis, which enhances S. aureus colonization in AD patients skin [6]. Furthermore it has been demonstrated that severity of AD correlate whit presence of different Staphylococcus species [7]. Saheb Kashaf S. et al. characterized the predominance of staphylococci during AD at three defined time points: flare (skin disease exacerbation without recent therapies), post-flare and baseline, in a cohort of 83 AD subjects and 15 healthy controls. The results showed that among the four most prominent genera (Staphylococcus, Corynebacterium, Streptococcus and Cutibacterium) present on the skin at affected disease sites, only Staphylococcus showed a significant increase. A more detailed metagenomic analysis conducted from the skin swab samples describes how, at the infection site, most subjects (66%) are predominantly colonized by S. aureus or the closely related Staphylococcus argenteus, while 29% are predominantly colonized by S. epidermidis [8]. Moreover, it has been shown that the severity of AD correlates with the presence of S. aureus – biofilm producers, and that biofilm-growing cultures from AD isolates display a greater tolerance to antibiotics than planktonic-growing cultures, which could account for the bacterial re-colonization few weeks after antibiotic treatment [9].
Another relevant skin disease is psoriasis, in which the skin microbiota affects the course of the disease. It is observed that the skin microbiota of individuals affected by psoriasis is substantially different from that of healthy subjects [10], with a marked reduction in its variability among those suffering from the condition compared to healthy individuals [11,12]. Both, lesional and non-lesional plaques, are enriched for S. aureus compared to healthy skin [13].
Polymicrobial colonization is a hallmark of both acute and chronic wounds, which may arise from burns, trauma, surgery, diabetes and peripheral vascular disease [14]. The analysis of swabs obtained from chronic wound infections in 163 patients showed that in 156 of them the culture test was positive. Pseudomonas aeruginosa was the dominant species (75 cases), with co-detections of Klebsiella pneumoniae (21 cases), Staphylococcus aureus (14 cases) and Proteus mirabilis (13 cases) [15]. Overall, chronic topical wounds commonly harbor concurrent infections by S. aureus and P. aeruginosa [16]; co-infection by these two bacterial species has been shown to impede wound healing, often extending closure times significantly compared to infections with only one species [17].
It has been demonstrated that S. aureus infection compromises wound healing by significantly prolonging the re-epithelialization phase. A deeper molecular analysis showed that S. aureus exudate hinders keratinocyte migration in vitro through the upregulation of Connexin-43 (Cx43), an effect that disappears when Cx43 is silenced. Further analysis in in vivo model identified activation of the PI3K/Akt pathway as a key mechanism behind Cx43 upregulation in S. aureus-infected wounds. Inhibiting this pathway reduced Cx43 levels and restored normal wound healing in vivo [18].
Chronic wounds are often associated with the development of microbial biofilms, which contribute to the delay in wound healing typical of chronic wounds [19,20]. Evidence indicates that the prevalence of biofilm in wounds varies significantly depending on their condition: biofilm is present in 60% of chronic wounds, while it drops to 6% in acute wounds [21]. A pooled analysis of eight prospective cohort studies (185 chronic wounds) and one case report found that 78.2% of unhealed chronic wounds in humans contained biofilms [22].
Chronic wound biofilms form resilient mixed-species communities that hinder both epithelial and granulation tissue development and sustain a persistent low-level inflammatory state, all of which delay healing. Once established, this hidden microbial ecosystem can progressively undermine normal repair processes [23]. Bacterial biofilm can act by modulating the host’s immune response. Biofilms release planktonic bacteria, LPS, quorum-sensing signals, exotoxins and DNA, triggering neutrophil recruitment [24,25]. Recruited neutrophils are unable to carry out their function because the ROS produced cannot penetrate the EPS (Extracellular Polymeric Substances); at the same time, the presence of a biofilm hinders the normal clearance of neutrophils [26,27]. This leads to abnormal release of high levels of proteases that interfere with and delay the healing process [28].
Biofilms are microbial sessile communities in which microorganisms live attached to abiotic or biotic surfaces and communicate with each other via quorum sensing mechanisms. The extracellular matrix embedding cells is composed of proteins, lipids, nucleic acids and polysaccharides and confers resistance to antimicrobial agents and immune responses, rendering the microbial biofilm particularly difficult to treat compared to the planktonic counterpart [19]. The commensal S. epidermidis is one of the most commonly cultured bacteria in clinical microbiology laboratories among coagulase negative staphylococci (CoNS), probably due to the presence of infection-associated genetic elements correlating with biofilm formation and methicillin resistance [29]; moreover, because S. epidermidis is ubiquitous on human skin, it often contaminates and thereby infects medical devices [30]. S. aureus wound infection represents the main cause of difficult to treat complications in Foot Diabetic Ulcers [31,32], particularly due to its activity in delaying wound healing [33]. The prevalence of methicillin-resistant S. aureus (MRSA) in foot ulcers is 15-30 %, [34]. A recent study characterizing wound infections in 266 subjects found that 34.58% (92/266) were colonized by S. aureus; of those 28.3% (26/92) were MRSA [35].
The development of alternative biofilm eradication strategies is of pivotal importance for the treatment of skin infections.
Vitreoscilla filiformis is a non-pathogenic Gram-negative bacterium belonging to the order of Beggiatoales. This bacterium has been identified as “beneficial” for its positive effects on skin health. In clinical studies, V. filiformis biomass lysate could improve clinical signs in AD and reduce S. aureus colonization of the skin [36,37] . The protective effects on skin health could be due to the proven activation of skin major endogenous inducible free radical scavenger MnSOD (Manganese superoxide dismutase), the stimulation of antimicrobial β-defensins production [38,39], and the induction of tolerogenic dendritic cells and Treg (regulatory T) cells able to suppress cutaneous inflammation [40].
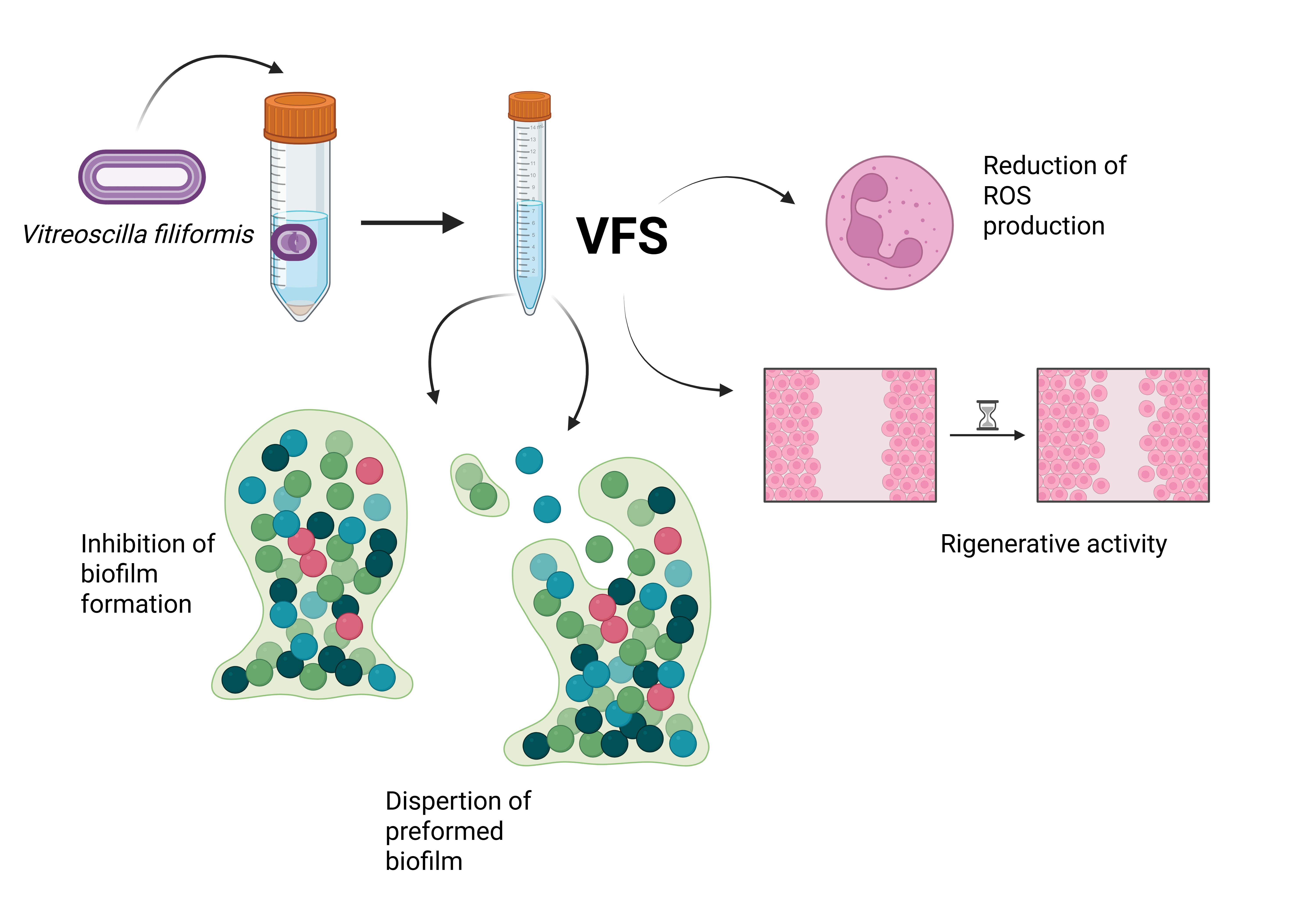
The development of antibiotic resistance in cutaneous infections restricts the possibilities of treatment favouring the incidence of serious health complications. The aim of this study was to evaluate the beneficial biological activities of V. filiformis (VF)-conditioned medium (supernatant, VFS) on biofilm formation by bacteria involved in wound infections and cutaneous diseases, as P. aeruginosa, S. aureus, S. epidermidis and the clinical isolate methicillin resistant strain MRSA. We also analysed the regenerative properties of the VFS on human dermal fibroblasts and its antioxidant activity.
2. Materials and Methods
2.1. Microbial Strains and Growth Conditions
Staphylococcus aureus (ATCC 29213, American Type Culture Collection, Manassas, VA, USA), Staphylococcus epidermidis (ATCC 35984), P. aeruginosa (PAO-1, ATCC15692) and the clinical isolate methicillin resistant S. aureus MRSA (collected during the routine clinical laboratory activity at the University Hospital of Perugia, Italy) were initially streaked from -80°C glicerol stock and maintained in tryptic soy agar (TSA, Sigma Aldrich). The day before the test, one colony was inoculated in tryptic soy broth (TSB, Sigma Aldrich) and incubated for 24 h at 37°C. Microbial cells were harvested by centrifugation, washed in PBS (Phosphate Buffered Saline), counted by spectrophotometric analysis at 600 nm and resuspended to the desired concentration in the appropriate culture medium.
2.2. Vitreoscilla filiformis and Culture Supernatant Preparation
Vitreoscilla filiformis (ATCC® 15551™) was grown in Beggiatoa (ATCC® medium: 138 Beggiatoa medium) at 37°C. 0.1ml were transferred in new media every 7 days. The day prior the experiment, 7 ml TSB were inoculated with 100 µl of an overnight culture (dilution 1:70) and incubated for 18-24h at 37°C. The culture (1–1.5 x 108 CFU/ml) was centrifuged at 3000 rpm for 10 minutes, the culture medium was filtered through a 0.45 µm filter (GVS Filter Technology) and used for downstream experiments.
2.3. Microbial Growth Kinetics
200 µl of staphylococci and P. aeruginosa suspensions (105 CFU/ml) were incubated in TSB at 37°C in the presence of different percentages of V. filiformis supernatant (VFS - 1%, 5%, 25%, 75%, 90%, v/v) or gentamicin as control in a microplate reader (Infinite M200 pro, TECAN). Absorbance at 600 nm was recorded every 60 minutes for a total of 24 hours of incubation. Each experiment was performed at least twice, and each sample was analysed in triplicate.
2.4. Effect of V. filiformis Supernatant (VFS) on S. aureus, S. epidermidis and MRSA Biofilm Formation and Dispersal
The in vitro static biofilm assay was performed as previously described with some modifications [41]. Briefly, overnight S. aureus, S. epidermidis and MRSA cultures were diluted 1:100 in TSB supplemented with 2% sucrose. 100 µl of standardized bacterial cultures (1x106 CFU/ml) were incubated in a flat bottom 96 well plate at 37°C for 24h under static conditions in the presence or absence of different percentages (5-25-75 % v/v) of VFS or gentamicin as a control. To assess the capacity of VFS to disperse established biofilms, bacteria were cultured in TSB 2% sucrose for 24h at 37°C as previously described. Biofilms were treated with different percentages (1-5-25-75-90-100 % v/v) of VFS for 4h or 24h at 37°C. After incubation, the planktonic cultures were transferred to a new plate and absorbance was read at 600 nm. For analysing the biofilm mass, wells were washed twice with 200 µl of distilled water. 50 µl of 0.4% Crystal Violet were added to each well for a minimum of 20 minutes. Wells were then washed twice with distilled water and the Crystal Violet was resuspended by adding 100 µl of 90% ethanol and incubated at room temperature for 15 minutes. Absorbance was read at 570 nm. Bioassays were performed in triplicate in at least three separate experiments.
2.5. DPPH Radical Scavenging Activity
The antioxidant activity of VFS was evaluated by using the 2,2-diphenyl-1-picrylhydrazyl (DPPH) free radical scavenging assay as described by Dutra et al. [42] with some modifications. DPPH is a stable free radical at room temperature, which can be reduced by the transfer of a hydrogen atom from an antioxidant agent, causing a change in colour of DPPH from deep violet to yellow evaluable by spectrophotometry analysis. Because TSB showed some antioxidant activity V. filiformis was cultured in 7 ml of RPMI 1640 (Roswell Park Memorial Institute 1640, Gibco-BRL). The day after, the culture was centrifuged at 3000 rpm for 10 minutes and the supernatant was filtered. Different percentages of Vitreoscilla supernatant were prepared (1-25-75 % v/v) in RPMI 1640. The reaction was carried out in a flat bottom 96 well plate in a final volume of 200 µl. The reaction mixture consisted of sample and DPPH in ethanol (25 µg/ml). RPMI 1640 has been used as negative sample control. The control solution was prepared by mixing ethanol and DPPH. A mixture of ethanol and sample was used as blank. The absorbance was read at 517 nm in a spectrophotometer plate reader (TECAN). Ascorbic acid (100 µg/ml) was used as positive control. Experiments were performed in triplicate.
The percentage of antioxidant activity (AA%) was calculated according to the following formula: AA%= 100 - [[(Abs sample – Abs blank) x 100] / Abs control]
2.6. Polymorphonuclear Cells (PMN) Isolation
Human heparinized venous blood was obtained from buffy coat provided by the Blood Bank of Ospedale della Misericordia of Perugia. Donors signed the consensus form (MOSIT 06) approved by Ethics Committee CEAS (Comitato Etico Aziende Sanitarie) (Rev. 3 Ottobre 2014) in which they authorize the use of their sample for research studies. The blood was diluted with RPMI 1640 (Gibco-BRL) and subjected to density gradient centrifugation over Ficoll-Hypaque Plus (Pharmacia Biotech). PMN were recovered and washed twice. The pellet was treated with hypotonic saline solution to lyse contaminant erythrocytes and PMN were suspended in RPMI 1640, washed twice, counted and diluted to the appropriate concentration.
2.7. Evaluation of ROS Production by Luminol Assay
100 µl of cellular suspension (1.25x106cells/ml) were stimulated with different percentages of VF culture supernatant (1-5-25 % v/v) grown in RPMI 1640 in the presence of 50 µl of Luminol (final concentration 0.28 mM) and the mixture was incubated for 3 minutes at 37°C. The cells were then stimulated with 50 µl of PMA (phorbol-12-myristate-13-acetate), final concentration 100 ng/ml) and chemiluminescence was monitored for 20 min in a luminometer reader (TECAN). The light output was recorded as RLU (relative luminescence units). Each experiment was performed in triplicate.
2.8. Effect of Vitreoscilla filiformis Supernatant on the Regenerative Capacity of Human Dermal Fibroblasts
Human dermal fibroblasts (HDF, Sigma Aldrich 106-05A) were seeded in Micro-Insert 4 Well µ-Dish 35 mm (Ibidi, 80406) at a concentration of 2x105/ml (final volume 15 µl). At confluence, the insert was removed, and cells were treated with different concentrations of VFS (1-25-75 % v/v). The growth was monitored by measuring the distance between the two sides of the monolayers at 24h, 48h and 72h under an inverted microscope (Nikon Eclipse TF2000-S microscope; Nikon, Milan, Italy) at 20x magnification.
2.9. Statistical Analysis
Results are expressed as mean ± standard deviation (SD). Significance was tested by means of Student’s two-tailed t-test. P<0.05 was considered significant.
3. Results
3.1. V. filiformis Supernatant Effect on Growth Kinetics of Bacteria
The effect of V. filiformis supernatant (VFS) on the growth of Gram-positive bacteria S. aureus, the clinical isolate strain MRSA, S. epidermidis and the Gram-negative bacteria P. aeruginosa was evaluated by spectrophotometry in the presence of different percentages of VFS (Figure 1).
As shown in Figure 1, VFS at the concentrations of 75 % and 90 % could significantly reduce the growth of the Gram-positive bacteria, while no effect was observed on P. aeruginosa. Lower percentages of the VFS did not affect the growth of the bacteria. This result is in line with the work of Gueniche showing a reduction of S. aureus colonization of the skin upon V. filiformis lysate treatment [36] .
3.2. Effect of V. filiformis Supernatant on Biofilm Formation and Dispersal
To analyse the anti-biofilm properties of VFS, we tested both the capacity of the supernatants to interfere with staphylococci biofilm formation on abiotic surface and their activity on preformed biofilms. Given the absence of antimicrobial activity of VFS on P. aeruginosa, the anti-biofilm activity has not been tested. For biofilm formation, bacteria were seeded in 96 well plates in the presence of different concentrations of VFS (5%, 25%, 75%) and cultured under static conditions for 24h at 37°C; while for biofilm dispersal, bacteria were first cultured for 24h to allow the formation of biofilms and then treated with VFS. The biofilm mass was assessed by crystal violet staining. VFS was able to inhibit biofilm formation of all tested bacteria at the concentrations of 25% and 75%; moreover, the concentration of 5% was effective in inhibiting S. epidermidis biofilm formation (Figure 2).
While the results observed for VFS 75% are not surprising because VFS 75 % is able to decrease the growth of Staphylococcus cells, VFS 25% showed a good anti-biofilm activity. Moreover, the VFS 5 % was able to reduce the biofilm mass of S. epidermidis. To confirm that the observed results were due to the anti-biofilm properties of the VFS and not to the inhibition of bacterial growth, the planktonic bacterial biomass was evaluated by optical density (O.D. 600nm). Interestingly, the decrease in biofilm mass was accompanied by a significant increase in the planktonic form compared to the untreated cells. Of interest, the anti-biofilm effect of VFS was also detected on drug resistant MRSA often isolated in clinical wounds [43]. Gentamicin inhibited the biofilm formation and bacterial growth. As expected, the results underline the effect of VFS on the mechanism of biofilm development, in particular the increase in the planktonic biomass suggests that VFS could have anti-adhesive activity and does not interfere with bacterial growth.
VFS was able to significantly reduce the mass of preformed biofilms of S. aureus, S. epidermidis and MRSA both after 4h and after 24h of treatment (Figure 3).
At 24h, a parallel increase in the planktonic mass was observed for all tested bacteria. VFS also induced an increase in the planktonic form of S. aureus after 4h of treatment (Figure 3 A). These results highlight that the anti-biofilm activity of VFS is not due the inhibition of bacterial growth but to its activity on the extracellular matrix characterizing mature biofilms.
3.3. Antioxidant Activity
The antioxidant capacity of V. filiformis supernatant was tested both in terms of neutralization of the free DPPH radical and reduction of total ROS produced from PMA-stimulated human neutrophils. Results shown in Figure 4 A highlight that VFS at a concentration of 25% and 75% holds a scavenging activity against DPPH.
The antioxidant activity was tested in human neutrophils by means of a luminol-based chemiluminescent assay after stimulation in the presence of PMA. As shown in Figure 4B, VFS was able to reduce ROS production in a dose dependent manner. Mahé et al. demonstrated the induction of intracellular MnSOD (Manganese Superoxide Dismutase), a major inducible free-radical scavenger of the skin, in normal human dermal fibroblasts and epidermal keratinocytes following V. filiformis treatment [39]. Immune cells characterize the wound microenvironment and play an important role in the initiation of healing. However, chronic wounds are constantly infiltrated by neutrophils, which release excessive amounts of ROS exerting deleterious effects on wound healing, by degrading extra-cellular matrix proteins and impairing the function of dermal fibroblasts and keratinocytes [44]. Our results on human neutrophils further confirm the antioxidant properties of V. filiformis, shedding light on the mechanisms underlying its beneficial effects on skin diseases.
3.4. Regenerative Activity
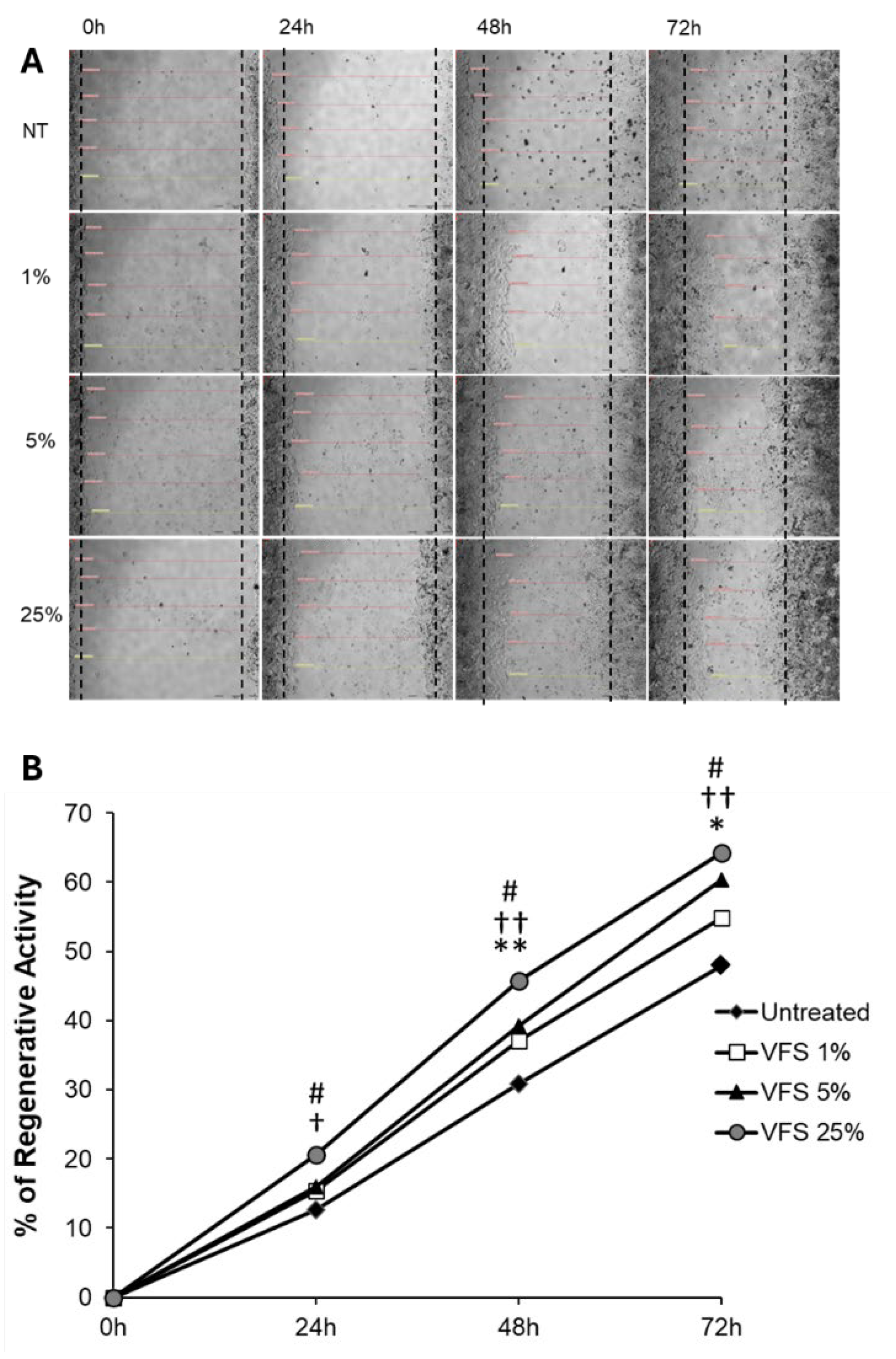
To test the regenerative properties of VFS, HDF cells were cultured in Micro-Insert 4 Well µ-Dish 35 mm until confluence is reached, after removal of the insert, the growth of cells in the presence of different percentages of VFS was monitored by measuring the distance between the two sides of the monolayers up to 72h. Figure 5 shows that the growth of HDF cells was stimulated by the addition of VFS in the culture medium in a dose dependent manner, with a maximum increase of growth in the presence of VFS 25% (+16.24% compared to untreated cells).
Current approaches for the treatment of chronic wounds include negative pressure wound therapy (NPWT) and hyperbaric oxygen therapy (HBO), aimed at clearing the wound of exudate, increasing tissue perfusion and promoting granulation tissue formation, as well as PDGF, Tβ4 and angiotensin, which are used to promote angiogenesis and stimulate cell proliferation [45]. Our results show that VFS can stimulate cell proliferation and facilitate wound closure in a in vitro model of dermal fibroblast regeneration; this effect could therefore be helpful for the treatment of chronic wounds.
4. Discussion
V. filiformis cell-free supernatant is able to inhibit cell growth of S. aureus, MRSA and S. epidermidis at high concentrations and to dampen bacterial biofilm formation; low concentrations were also effective in inducing the dispersal of mature biofilms even after 4h of treatment in all tested bacteria. The significant increase of bacterial concentrations during both biofilm formation and dispersal observed upon VFS treatment is particularly interesting. Indeed, enhancing the transition of biofilms to the planktonic state could revert the resistance of bacteria embedded in biofilm to the most used antimicrobials. Excessive amounts of ROS have been shown to characterize chronic wounds and to play a central role in delaying wound healing. VFS was able to reduce total ROS production in a cell human neutrophils-based assay in a dose-dependent manner. The direct antioxidant capacity of VFS was also confirmed by cell free DPPH assay. Clinical studies from Gueniche and Seite have clearly highlighted the beneficial effects (lysate including membrane and cytosol) of Vitreoscilla filiformis on skin diseases. Moreover, topically applied Vitreoscilla filiformis extract has demonstrated probiotic activity by restoring skin barrier function, stimulating the immune response, and reducing infection rates [46,47]. However, while their work involved the use of bacterial lysates and biomass respectively, we only focused on cell free bacterial conditioned medium. Our results on VFS and antioxidant activity, concomitantly with its ability to improve the regeneration of HDF monolayers, evidence its possible role for the prevention and treatment of chronic wounds, a skin condition where both microbial biofilm control and the stimulation of tissue regeneration are needed.
Author Contributions
Conceptualization, D.P. and M.P.; methodology, M.P.; validation, G.C.; M.P. and D.P.; formal analysis, A.G. and D.P.; investigation, M.P. and G.C.; resources, D.P.; data curation, M.P and A.G.; writing—original draft preparation, M.P. and G.C.; writing—review and editing, D.P. and G.C; supervision, D.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committee of University of Perugia (MOSIT 06 and date 3 October 2014).” for studies involving humans.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
All data to support the findings of this study are available from the corresponding author upon request.
Acknowledgments
The authors thank Professor Rita Pagiotti for sharing her knowledge on natural products.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| VFS | V. filiformis supernatant |
| MRSA | Methicillin-resistant Staphylococcus aureus |
| ROS | Reactive oxygen species |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| AD | Atopic Dermatitis |
References
- Skowron, K.; Bauza-Kaszewska, J.; Kraszewska, Z.; Wiktorczyk-Kapischke, N.; Grudlewska-Buda, K.; Kwiecińska-Piróg, J.; Wałecka-Zacharska, E.; Radtke, L.; Gospodarek-Komkowska, E. Human Skin Microbiome: Impact of Intrinsic and Extrinsic Factors on Skin Microbiota. Microorganisms 2021, 9, 543. [CrossRef]
- Cundell, A.M. Microbial Ecology of the Human Skin. Microb Ecol 2018, 76, 113–120. [CrossRef]
- Buffie, C.G.; Pamer, E.G. Microbiota-Mediated Colonization Resistance against Intestinal Pathogens. Nat Rev Immunol 2013, 13, 790–801. [CrossRef]
- Flowers, L.; Grice, E.A. The Skin Microbiota: Balancing Risk and Reward. Cell Host Microbe 2020, 28, 190–200. [CrossRef]
- Kong, H.H.; Oh, J.; Deming, C.; Conlan, S.; Grice, E.A.; Beatson, M.A.; Nomicos, E.; Polley, E.C.; Komarow, H.D.; NISC Comparative Sequence Program; et al. Temporal Shifts in the Skin Microbiome Associated with Disease Flares and Treatment in Children with Atopic Dermatitis. Genome Res 2012, 22, 850–859. [CrossRef]
- Miajlovic, H.; Fallon, P.G.; Irvine, A.D.; Foster, T.J. Effect of Filaggrin Breakdown Products on Growth of and Protein Expression by Staphylococcus Aureus. J Allergy Clin Immunol 2010, 126, 1184-1190.e3. [CrossRef]
- Byrd, A.L.; Deming, C.; Cassidy, S.K.B.; Harrison, O.J.; Ng, W.-I.; Conlan, S.; NISC Comparative Sequencing Program; Belkaid, Y.; Segre, J.A.; Kong, H.H. Staphylococcus Aureus and Staphylococcus Epidermidis Strain Diversity Underlying Pediatric Atopic Dermatitis. Sci Transl Med 2017, 9, eaal4651. [CrossRef]
- Saheb Kashaf, S.; Harkins, C.P.; Deming, C.; Joglekar, P.; Conlan, S.; Holmes, C.J.; Almeida, A.; Finn, R.D.; Segre, J.A.; Kong, H.H. Staphylococcal Diversity in Atopic Dermatitis from an Individual to a Global Scale. Cell Host & Microbe 2023, 31, 578-592.e6. [CrossRef]
- Di Domenico, E.G.; Cavallo, I.; Bordignon, V.; Prignano, G.; Sperduti, I.; Gurtner, A.; Trento, E.; Toma, L.; Pimpinelli, F.; Capitanio, B.; et al. Inflammatory Cytokines and Biofilm Production Sustain Staphylococcus Aureus Outgrowth and Persistence: A Pivotal Interplay in the Pathogenesis of Atopic Dermatitis. Sci Rep 2018, 8, 9573. [CrossRef]
- Rademaker, M.; Agnew, K.; Anagnostou, N.; Andrews, M.; Armour, K.; Baker, C.; Foley, P.; Gebauer, K.; Gupta, M.; Marshman, G.; et al. Psoriasis and Infection. A Clinical Practice Narrative. Australas J Dermatol 2019, 60, 91–98. [CrossRef]
- Alekseyenko, A.V.; Perez-Perez, G.I.; De Souza, A.; Strober, B.; Gao, Z.; Bihan, M.; Li, K.; Methé, B.A.; Blaser, M.J. Community Differentiation of the Cutaneous Microbiota in Psoriasis. Microbiome 2013, 1, 31. [CrossRef]
- Weyrich, L.S.; Dixit, S.; Farrer, A.G.; Cooper, A.J.; Cooper, A.J. The Skin Microbiome: Associations between Altered Microbial Communities and Disease. Australas J Dermatol 2015, 56, 268–274. [CrossRef]
- Chang, H.-W.; Yan, D.; Singh, R.; Liu, J.; Lu, X.; Ucmak, D.; Lee, K.; Afifi, L.; Fadrosh, D.; Leech, J.; et al. Alteration of the Cutaneous Microbiome in Psoriasis and Potential Role in Th17 Polarization. Microbiome 2018, 6, 154. [CrossRef]
- Sorg, H.; Tilkorn, D.J.; Hager, S.; Hauser, J.; Mirastschijski, U. Skin Wound Healing: An Update on the Current Knowledge and Concepts. Eur Surg Res 2017, 58, 81–94. [CrossRef]
- Sandmann, S.; Nunes, J.V.; Grobusch, M.P.; Sesay, M.; Kriegel, M.A.; Varghese, J.; Schaumburg, F. Network Analysis of Polymicrobial Chronic Wound Infections in Masanga, Sierra Leone. BMC Infectious Diseases 2023, 23, 250. [CrossRef]
- Yung, D.B.Y.; Sircombe, K.J.; Pletzer, D. Friends or Enemies? The Complicated Relationship between Pseudomonas Aeruginosa and Staphylococcus Aureus. Mol Microbiol 2021, 116, 1–15. [CrossRef]
- Pastar, I.; Nusbaum, A.G.; Gil, J.; Patel, S.B.; Chen, J.; Valdes, J.; Stojadinovic, O.; Plano, L.R.; Tomic-Canic, M.; Davis, S.C. Interactions of Methicillin Resistant Staphylococcus Aureus USA300 and Pseudomonas Aeruginosa in Polymicrobial Wound Infection. PLoS One 2013, 8, e56846. [CrossRef]
- Xu, W.; Dielubanza, E.; Maisel, A.; Leung, K.; Mustoe, T.; Hong, S.; Galiano, R. Staphylococcus Aureus Impairs Cutaneous Wound Healing by Activating the Expression of a Gap Junction Protein, Connexin-43 in Keratinocytes. Cell Mol Life Sci 2021, 78, 935–947. [CrossRef]
- Wu, Y.-K.; Cheng, N.-C.; Cheng, C.-M. Biofilms in Chronic Wounds: Pathogenesis and Diagnosis. Trends Biotechnol 2019, 37, 505–517. [CrossRef]
- Han, G.; Ceilley, R. Chronic Wound Healing: A Review of Current Management and Treatments. Adv Ther 2017, 34, 599–610. [CrossRef]
- James, G.A.; Swogger, E.; Wolcott, R.; Pulcini, E. deLancey; Secor, P.; Sestrich, J.; Costerton, J.W.; Stewart, P.S. Biofilms in Chronic Wounds. Wound Repair Regen 2008, 16, 37–44. [CrossRef]
- Malone, M.; Bjarnsholt, T.; McBain, A.J.; James, G.A.; Stoodley, P.; Leaper, D.; Tachi, M.; Schultz, G.; Swanson, T.; Wolcott, R.D. The Prevalence of Biofilms in Chronic Wounds: A Systematic Review and Meta-Analysis of Published Data. J Wound Care 2017, 26, 20–25. [CrossRef]
- Metcalf, D.G.; Bowler, P.G. Biofilm Delays Wound Healing: A Review of the Evidence. Burns Trauma 2015, 1, 5–12. [CrossRef]
- Hirschfeld, J. Dynamic Interactions of Neutrophils and Biofilms. J Oral Microbiol 2014, 6, 10.3402/jom.v6.26102. [CrossRef]
- Karlsson, T.; Musse, F.; Magnusson, K.-E.; Vikström, E. N-Acylhomoserine Lactones Are Potent Neutrophil Chemoattractants That Act via Calcium Mobilization and Actin Remodeling. J Leukoc Biol 2012, 91, 15–26. [CrossRef]
- Wolcott, R.D.; Rhoads, D.D.; Dowd, S.E. Biofilms and Chronic Wound Inflammation. J Wound Care 2008, 17, 333–341. [CrossRef]
- Prince, L.R.; Bianchi, S.M.; Vaughan, K.M.; Bewley, M.A.; Marriott, H.M.; Walmsley, S.R.; Taylor, G.W.; Buttle, D.J.; Sabroe, I.; Dockrell, D.H.; et al. Subversion of a Lysosomal Pathway Regulating Neutrophil Apoptosis by a Major Bacterial Toxin, Pyocyanin. J Immunol 2008, 180, 3502–3511. [CrossRef]
- Rohde, H.; Burdelski, C.; Bartscht, K.; Hussain, M.; Buck, F.; Horstkotte, M.A.; Knobloch, J.K.-M.; Heilmann, C.; Herrmann, M.; Mack, D. Induction of Staphylococcus Epidermidis Biofilm Formation via Proteolytic Processing of the Accumulation-Associated Protein by Staphylococcal and Host Proteases. Molecular Microbiology 2005, 55, 1883–1895. [CrossRef]
- Méric, G.; Mageiros, L.; Pensar, J.; Laabei, M.; Yahara, K.; Pascoe, B.; Kittiwan, N.; Tadee, P.; Post, V.; Lamble, S.; et al. Disease-Associated Genotypes of the Commensal Skin Bacterium Staphylococcus Epidermidis. Nat Commun 2018, 9, 5034. [CrossRef]
- Uçkay, I.; Pittet, D.; Vaudaux, P.; Sax, H.; Lew, D.; Waldvogel, F. Foreign Body Infections Due to Staphylococcus Epidermidis. Ann Med 2009, 41, 109–119. [CrossRef]
- Saltoglu, N.; Ergonul, O.; Tulek, N.; Yemisen, M.; Kadanali, A.; Karagoz, G.; Batirel, A.; Ak, O.; Sonmezer, C.; Eraksoy, H.; et al. Influence of Multidrug Resistant Organisms on the Outcome of Diabetic Foot Infection. International Journal of Infectious Diseases 2018, 70, 10–14. [CrossRef]
- Giurato, L.; Meloni, M.; Izzo, V.; Uccioli, L. Osteomyelitis in Diabetic Foot: A Comprehensive Overview. World J Diabetes 2017, 8, 135–142. [CrossRef]
- Ortines, R.V.; Liu, H.; Cheng, L.I.; Cohen, T.S.; Lawlor, H.; Gami, A.; Wang, Y.; Dillen, C.A.; Archer, N.K.; Miller, R.J.; et al. Neutralizing Alpha-Toxin Accelerates Healing of Staphylococcus Aureus-Infected Wounds in Nondiabetic and Diabetic Mice. Antimicrob Agents Chemother 2018, 62, e02288-17. [CrossRef]
- Eleftheriadou, I.; Tentolouris, N.; Argiana, V.; Jude, E.; Boulton, A.J. Methicillin-Resistant Staphylococcus Aureus in Diabetic Foot Infections. Drugs 2010, 70, 1785–1797. [CrossRef]
- Tsige, Y.; Tadesse, S.; G/Eyesus, T.; Tefera, M.M.; Amsalu, A.; Menberu, M.A.; Gelaw, B. Prevalence of Methicillin-Resistant Staphylococcus Aureus and Associated Risk Factors among Patients with Wound Infection at Referral Hospital, Northeast Ethiopia. J Pathog 2020, 2020, 3168325. [CrossRef]
- Gueniche, A.; Knaudt, B.; Schuck, E.; Volz, T.; Bastien, P.; Martin, R.; Röcken, M.; Breton, L.; Biedermann, T. Effects of Nonpathogenic Gram-Negative Bacterium Vitreoscilla Filiformis Lysate on Atopic Dermatitis: A Prospective, Randomized, Double-Blind, Placebo-Controlled Clinical Study. Br J Dermatol 2008, 159, 1357–1363. [CrossRef]
- Seité, S.; Zelenkova, H.; Martin, R. Clinical Efficacy of Emollients in Atopic Dermatitis Patients – Relationship with the Skin Microbiota Modification. Clin Cosmet Investig Dermatol 2017, 10, 25–33. [CrossRef]
- Mahe, Y.F.; Perez, M.-J.; Tacheau, C.; Fanchon, C.; Martin, R.; Rousset, F.; Seite, S. A New Vitreoscilla Filiformis Extract Grown on Spa Water-Enriched Medium Activates Endogenous Cutaneous Antioxidant and Antimicrobial Defenses through a Potential Toll-like Receptor 2/Protein Kinase C, Zeta Transduction Pathway. Clin Cosmet Investig Dermatol 2013, 6, 191–196. [CrossRef]
- Mahé, Y.F.; Martin, R.; Aubert, L.; Billoni, N.; Collin, C.; Pruche, F.; Bastien, P.; Drost, S.S.; Lane, A.T.; Meybeck, A. Induction of the Skin Endogenous Protective Mitochondrial MnSOD by Vitreoscilla Filiformis Extract. International Journal of Cosmetic Science 2006, 28, 277–287. [CrossRef]
- Volz, T.; Skabytska, Y.; Guenova, E.; Chen, K.-M.; Frick, J.-S.; Kirschning, C.J.; Kaesler, S.; Röcken, M.; Biedermann, T. Nonpathogenic Bacteria Alleviating Atopic Dermatitis Inflammation Induce IL-10-Producing Dendritic Cells and Regulatory Tr1 Cells. J Invest Dermatol 2014, 134, 96–104. [CrossRef]
- Iwase, T.; Uehara, Y.; Shinji, H.; Tajima, A.; Seo, H.; Takada, K.; Agata, T.; Mizunoe, Y. Staphylococcus Epidermidis Esp Inhibits Staphylococcus Aureus Biofilm Formation and Nasal Colonization. Nature 2010, 465, 346–349. [CrossRef]
- Dutra, R.P.; Abreu, B.V. de B.; Cunha, M.S.; Batista, M.C.A.; Torres, L.M.B.; Nascimento, F.R.F.; Ribeiro, M.N.S.; Guerra, R.N.M. Phenolic Acids, Hydrolyzable Tannins, and Antioxidant Activity of Geopropolis from the Stingless Bee Melipona Fasciculata Smith. J Agric Food Chem 2014, 62, 2549–2557. [CrossRef]
- Branski, L.K.; Al-Mousawi, A.; Rivero, H.; Jeschke, M.G.; Sanford, A.P.; Herndon, D.N. Emerging Infections in Burns. Surg Infect (Larchmt) 2009, 10, 389–397. [CrossRef]
- Dunnill, C.; Patton, T.; Brennan, J.; Barrett, J.; Dryden, M.; Cooke, J.; Leaper, D.; Georgopoulos, N.T. Reactive Oxygen Species (ROS) and Wound Healing: The Functional Role of ROS and Emerging ROS-Modulating Technologies for Augmentation of the Healing Process. Int Wound J 2017, 14, 89–96. [CrossRef]
- Zielins, E.R.; Brett, E.A.; Luan, A.; Hu, M.S.; Walmsley, G.G.; Paik, K.; Senarath-Yapa, K.; Atashroo, D.A.; Wearda, T.; Lorenz, H.P.; et al. Emerging Drugs for the Treatment of Wound Healing. Expert Opin Emerg Drugs 2015, 20, 235–246. [CrossRef]
- Gueniche, A.; Liboutet, M.; Cheilian, S.; Fagot, D.; Juchaux, F.; Breton, L. Vitreoscilla Filiformis Extract for Topical Skin Care: A Review. Front Cell Infect Microbiol 2021, 11, 747663. [CrossRef]
- Gueniche, A.; Valois, A.; Kerob, D.; Rasmont, V.; Nielsen, M. A Combination of Vitreoscilla Filiformis Extract and Vichy Volcanic Mineralizing Water Strengthens the Skin Defenses and Skin Barrier. J Eur Acad Dermatol Venereol 2022, 36 Suppl 2, 16–25. [CrossRef]
Figure 1.
Kinetics of S. aureus (A), MRSA (B) S. epidermidis (C), P. aeruginosa (D) and in the presence or absence of the indicated percentages of VFS. The bacterial growth of was evaluated by measuring the O.D. at 600 nm every 60 minutes for 24 hours at 37°C (TECAN). Gentamicin (250 µg/ml) was used as positive control. Data represent the mean of two independent experiments performed in triplicate. * P<0.01, ** P<0.001 (VFS-treated versus untreated).
Figure 1.
Kinetics of S. aureus (A), MRSA (B) S. epidermidis (C), P. aeruginosa (D) and in the presence or absence of the indicated percentages of VFS. The bacterial growth of was evaluated by measuring the O.D. at 600 nm every 60 minutes for 24 hours at 37°C (TECAN). Gentamicin (250 µg/ml) was used as positive control. Data represent the mean of two independent experiments performed in triplicate. * P<0.01, ** P<0.001 (VFS-treated versus untreated).
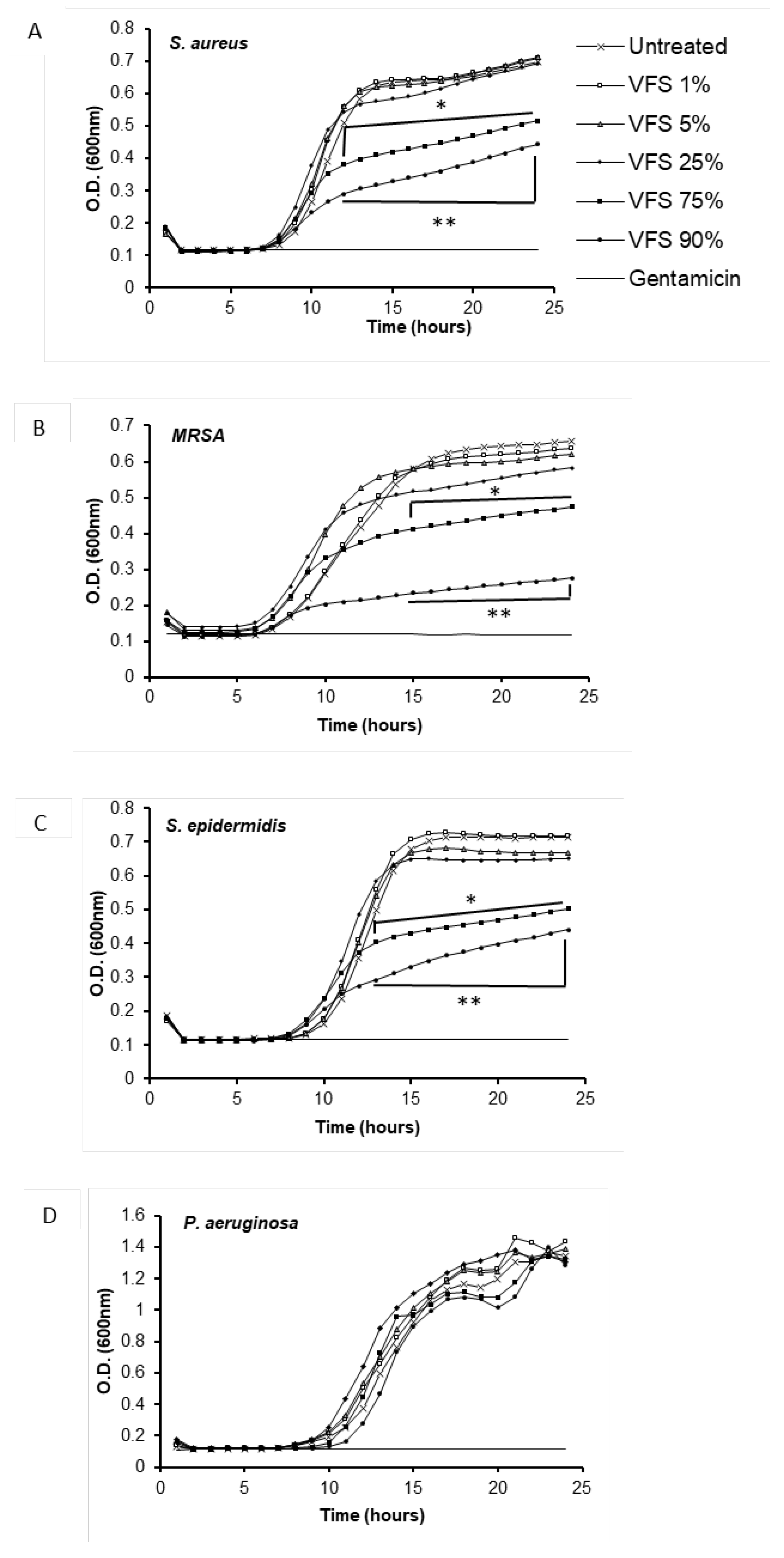
Figure 2.
Inhibition of biofilm formation by VFS. S. aureus, S. epidermidis and MRSA overnight cultures were diluted 1:100 in TSB 2% sucrose and incubated in in 96 well plates for 24h at 37°C in the presence or absence of the indicated concentrations of VFS. The biofilm mass was quantified by crystal violet staining (absorbance 570nm), while the planktonic mass was determined by absorbance at 600 nm. Gentamicin (250 µg/ml) was used as positive control. Data are presented as % of increase or decrease compared to untreated bacteria (100). Data represent the Mean ± SD of at least three independent experiments performed in triplicate. * P<0.05, ** P<0.001 (biofilm mass, VFS – treated bacteria versus untreated); # P<0.05, ## P<0.001 (planktonic bacteria, VFS – treated bacteria versus untreated).
Figure 2.
Inhibition of biofilm formation by VFS. S. aureus, S. epidermidis and MRSA overnight cultures were diluted 1:100 in TSB 2% sucrose and incubated in in 96 well plates for 24h at 37°C in the presence or absence of the indicated concentrations of VFS. The biofilm mass was quantified by crystal violet staining (absorbance 570nm), while the planktonic mass was determined by absorbance at 600 nm. Gentamicin (250 µg/ml) was used as positive control. Data are presented as % of increase or decrease compared to untreated bacteria (100). Data represent the Mean ± SD of at least three independent experiments performed in triplicate. * P<0.05, ** P<0.001 (biofilm mass, VFS – treated bacteria versus untreated); # P<0.05, ## P<0.001 (planktonic bacteria, VFS – treated bacteria versus untreated).
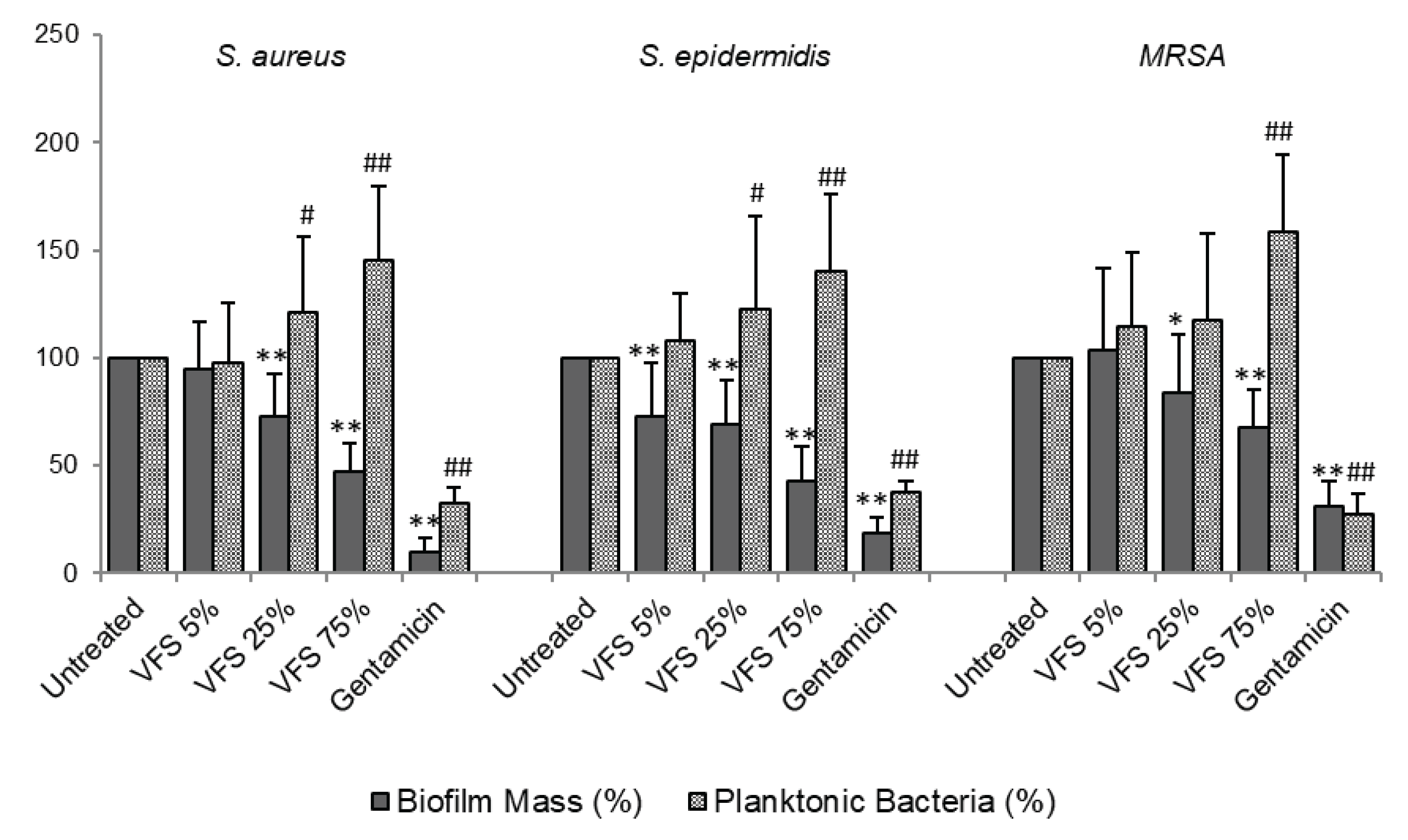
Figure 3.
Dispersal activity of VFS on pre-formed biofilms. S. aureus (A), S. epidermidis (B) and MRSA (C) overnight cultures were diluted 1:100 in TSB 2% sucrose and incubated in in 96 well plates at 37°C. After 24h, the preformed biofilms were treated with different concentrations of VFS and incubated for further 4h and 24h. The biofilm mass was quantified by crystal violet staining (absorbance 570nm), while the planktonic mass was determined by absorbance at 600nm. Gentamicin (250 µg/ml) was used as positive control. Data are presented as % of increase or decrease compared to untreated bacteria. Data represent the Mean ± SD of three independent experiments performed in triplicate. * P<0.05, ** P<0.001 (biofilm mass, VFS – treated bacteria versus untreated); # P<0.05, ## P<0.001 (planktonic bacteria, VFS – treated bacteria versus untreated).
Figure 3.
Dispersal activity of VFS on pre-formed biofilms. S. aureus (A), S. epidermidis (B) and MRSA (C) overnight cultures were diluted 1:100 in TSB 2% sucrose and incubated in in 96 well plates at 37°C. After 24h, the preformed biofilms were treated with different concentrations of VFS and incubated for further 4h and 24h. The biofilm mass was quantified by crystal violet staining (absorbance 570nm), while the planktonic mass was determined by absorbance at 600nm. Gentamicin (250 µg/ml) was used as positive control. Data are presented as % of increase or decrease compared to untreated bacteria. Data represent the Mean ± SD of three independent experiments performed in triplicate. * P<0.05, ** P<0.001 (biofilm mass, VFS – treated bacteria versus untreated); # P<0.05, ## P<0.001 (planktonic bacteria, VFS – treated bacteria versus untreated).
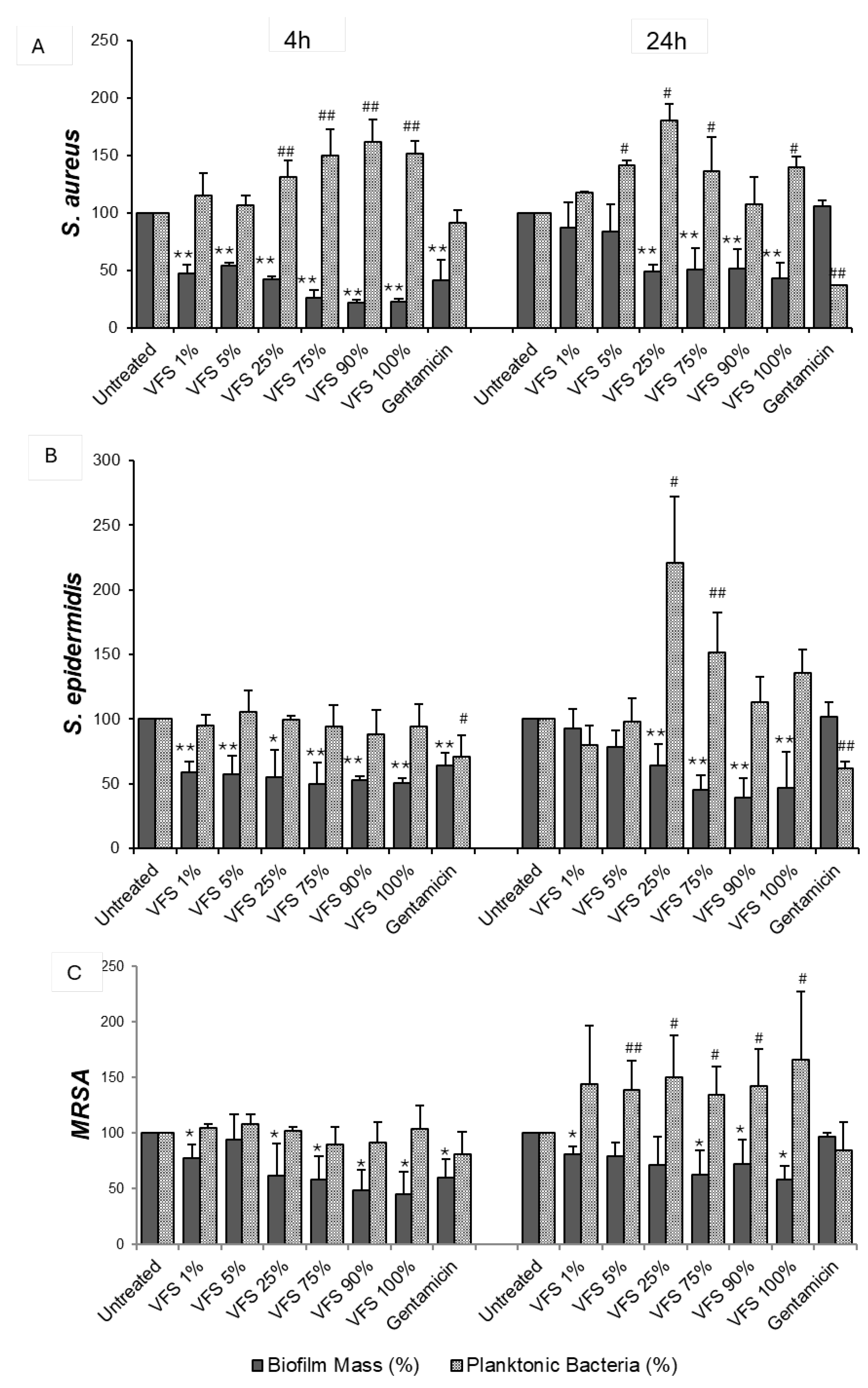
Figure 4.
Antioxidant activity of VFS. (A) The antioxidant activity of 1%, 5%, 25% and 75% VFS was tested by DPPH assay. Results are expressed % of DPPH scavenging activity. Ascorbic acid (100 µg/ml) was used as positive control. Data represent the Mean ± SD of two independent experiments performed in triplicate. (B) The effect of the indicated concentrations of VFS on ROS production by human neutrophils was evaluated by luminol chemiluminescence assay after PMA stimulation. Data are expressed as RLU (Relative Luminescence Units) and are representative of two independent experiments with similar results performed in triplicate. * P<0.005 (1%, 5%, 25%. 75% VFS versus 1%, 5%, 25%, 75% RPMI).
Figure 4.
Antioxidant activity of VFS. (A) The antioxidant activity of 1%, 5%, 25% and 75% VFS was tested by DPPH assay. Results are expressed % of DPPH scavenging activity. Ascorbic acid (100 µg/ml) was used as positive control. Data represent the Mean ± SD of two independent experiments performed in triplicate. (B) The effect of the indicated concentrations of VFS on ROS production by human neutrophils was evaluated by luminol chemiluminescence assay after PMA stimulation. Data are expressed as RLU (Relative Luminescence Units) and are representative of two independent experiments with similar results performed in triplicate. * P<0.005 (1%, 5%, 25%. 75% VFS versus 1%, 5%, 25%, 75% RPMI).
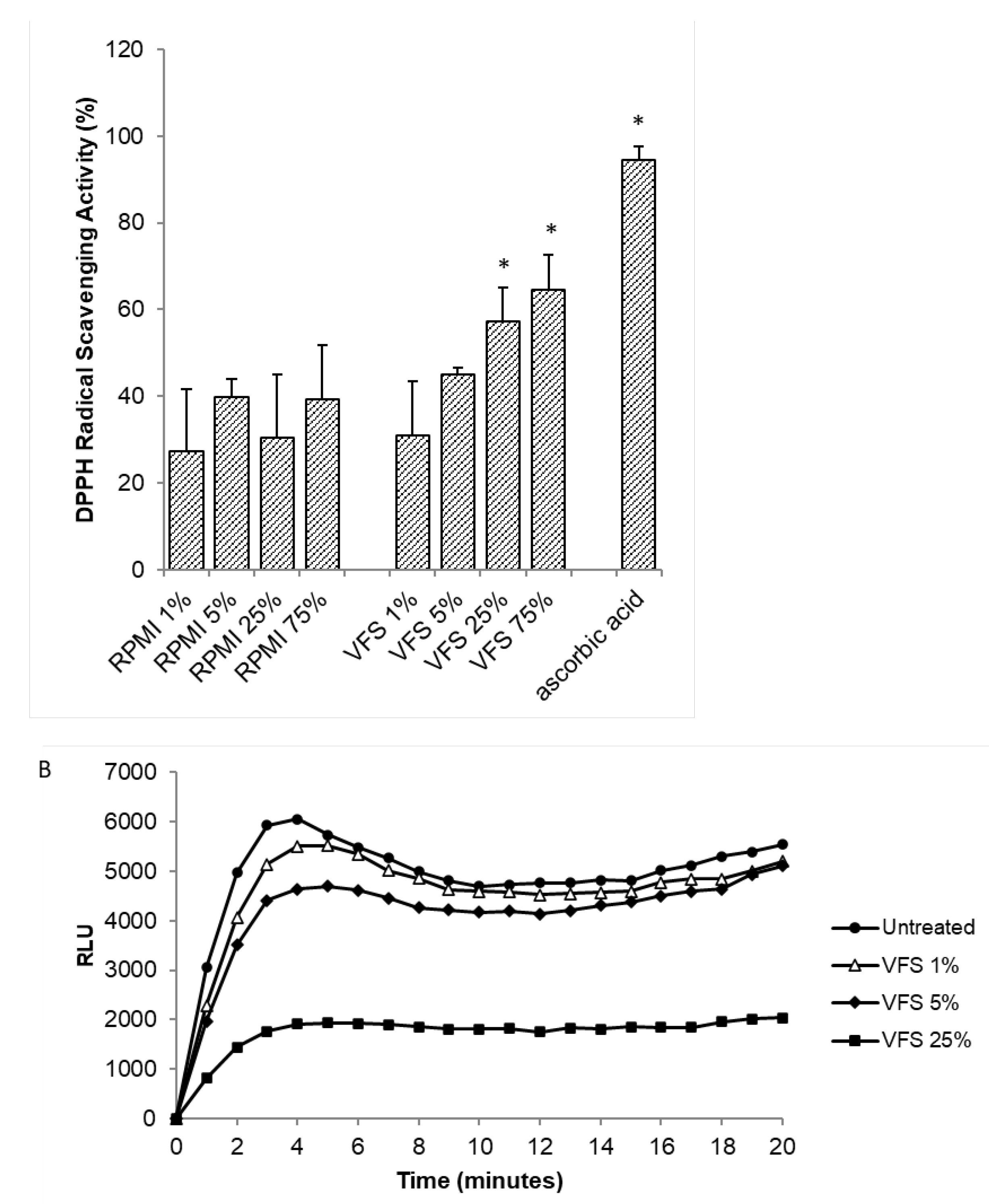
Figure 5.
Regenerative activity of VFS. HDF cells (3x103) were seeded in Micro-Insert 4 Well µ-Dishes (35 mm) and cultured until confluency. The insert was removed with sterile tweezers. Cells were treated with 1%, 5% and 25% VFS and the growth was monitored at 24h, 48h and 72h in an phase-contrast microscope. (A) Photo micrographs of HDF cells at different time points cultured in the presence/absence of different percentages of VFS. (B) % of regenerative activity of VFS on HDF cells. Data are expressed as % of regenerative activity of VFS-treated cells compared to untreated cells and are presented as Mean ± SD of two independent experiments. * P<0.05, **P<0.01 (1%VFS-treated cells versus untreated cells), † P<0.01, †† P<0.001 (5%VFS-treated cells versus untreated cells).
Figure 5.
Regenerative activity of VFS. HDF cells (3x103) were seeded in Micro-Insert 4 Well µ-Dishes (35 mm) and cultured until confluency. The insert was removed with sterile tweezers. Cells were treated with 1%, 5% and 25% VFS and the growth was monitored at 24h, 48h and 72h in an phase-contrast microscope. (A) Photo micrographs of HDF cells at different time points cultured in the presence/absence of different percentages of VFS. (B) % of regenerative activity of VFS on HDF cells. Data are expressed as % of regenerative activity of VFS-treated cells compared to untreated cells and are presented as Mean ± SD of two independent experiments. * P<0.05, **P<0.01 (1%VFS-treated cells versus untreated cells), † P<0.01, †† P<0.001 (5%VFS-treated cells versus untreated cells).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



