
Submitted:
01 July 2025
Posted:
02 July 2025
You are already at the latest version
Abstract
The tomato leafminer Phthorimaea absoluta is an important pest of solanaceous plants and is considered a key pest of tomato plants. This pest is rapidly dispersed, difficult to con-trol, and resistant to the main synthetic insecticides. The objective was to determine the lethal and sublethal effects of three essential oils (Eos) of Melaleuca alternifolia and Euca-lyptus staigeriana against P. absoluta. We evaluated the effects of the EOs during the life cy-cle of the pest using an age-stage, two-sex life table. The treatments were solubilized in ac-etone and applied to the back of the caterpillars. The EOs of M. alternifolia and E. staigeriana reduced P. absoluta longevity to 22.2 and 15.9 days and negatively impacted demographic parameters. Melaleuca alternifolia notably decreased the intrinsic rate of increase. Chroma-tographic findings revealed the presence of 19 compounds in the M. alternifolia EO, with terpinen-4-ol, γ-terpinene and α-terpinene being the most abundant. On the other hand, E. staigeriana presented 25 constituents, with the compounds limonene, geranial and neral being the most abundant. The results obtained demonstrate the insecticidal potential of M. alternifolia and E. staigeriana EOs in the control of P. absoluta.
Keywords:
South American tomato pinworm
; Botanical insecticides
; IPM
1. Introduction
The South American tomato pinworm, Phthorimaea absoluta (Meyrick, 1917) (Lepidoptera: Gelechiidae), is a highly invasive pest of Solanaceae and is recognized as one of the most significant threats to tomato (Solanum lycopersicum L.) production globally, having been reported in more than 90 countries [1,2]. In Asia, its presence has already been confirmed in Korea, with the first detection reported in Jeonbuk Province [3]. Owing to its high reproductive potential, rapid generational turnover, and ability to complete several generations annually [4,5], infestations can result in substantial yield losses, ranging from 50% to total crop failure [6].
The rapid expansion of P. absoluta in Solanaceae worldwide, which led to a significant increase in the use of synthetic insecticides [8]. The indiscriminate use of synthetic chemical insecticides led to the rapid selection of P. absoluta populations resistant to the main chemical classes, such as organophosphates, pyrethroids and diamides [9,10,11]. In addition, insecticides can cause negative impacts on natural enemies and the contamination of the environment and enable the resurgence of secondary pests [12,13,14]. Due to the adverse effects of synthetic insecticides, studies that seek alternatives to control P. absoluta should be encouraged, and in this context, there is a growing demand for the development of botanical insecticides to control this pest [15,16,17].
Botanical insecticides are plant products that have activity against insect pests and may originate from dried and ground plant structures, crude extracts or isolated components of the secondary metabolism of plants [17]. Botanical products can cause lethal (mortality) and sublethal effects, such as effects on behavior, development, and reproduction and interfering with metabolic pathways in arthropod pests [17,18,19,20]. Natural insecticides are most often easily degradable and are considered environmentally friendly products [21,22]. In addition, the raw materials are available in large quantities and at low cost, and many plant derivatives have several mechanisms of action still unexplored [23]. Among botanical insecticides, essential oils (EOs) exhibit bioactivity against several groups of insect pests and have been gaining prominence among biopesticides [17].
The use of plant EOs represents a promising strategy within integrated pest management (IPM) programs, as they are rich in bioactive compounds and exhibit insecticidal, repellent, and oviposition-deterring properties [24,25,26]. In addition to their efficacy, EOs generally pose low environmental risks due to their volatility, rapid degradation, and minimal soil persistence, reducing the likelihood of resistance development in pest populations owing to their multiple modes of action [25]. These oils are typically complex mixtures of terpenoids, whose biological effects often result from synergistic or additive interactions among constituents [26]. In P. absoluta, various EOs have been shown to induce adult and larval mortality [27], exhibit repellency [28], and inhibit oviposition behavior [29]. Despite these benefits, EOs derived from Myrtaceae species remain underexplored in the management of tomato leafminers [15]. Recent studies have further confirmed the fumigant toxicity [30] and behavioral effects of selected plant EOs on P. absoluta, as well as their safety to natural predators such as Macrolophus pygmaeus (Rambor, 1839) (Hemiptera: Miridae) [31].
The EOs of plants of the Myrtaceae family represent a class of volatile secondary metabolites containing terpenes and aromatic compounds as the main components [32]. These plants are gaining increasing interest in IPM programs due to their ovicidal, larvicidal, and adulticidal action; repellent activity; and fumigant and contact toxicity [33]. The Eucalyptus staigeriana F. Muell. ex Bailey and Melaleuca alternifolia Cheel are among the species of the family Myrtaceae with insecticidal activity. The EOs from the leaves of plants of the genus Eucalyptus are commercially used in the pharmaceutical, cosmetic and food industries [34]. The EO bioactivity of E. staigeriana has been reported for different insect orders. For example, in insects of the order Coleoptera, a harmful effect was observed on the oviposition and emergence of Zabrotes subfasciatus (Both, 1833) (Bruchidae) and Calosobruchus maculatus (Fabricius, 1775) (Chrysomelidae) [35] as well as repellency to C. maculatus [36] and Sitophilus zeamais (Mots, 1885) (Curculionidae) [37]. In addition, the survival of Lutzomyia longipalpis (Lutz and Neiva, 1912) (Diptera: Psychodidae) [38] and the reproductive parameters of Spodoptera frugiperda (Smith, 1797) (Lepidoptera: Noctuidae) were affected [39]. On the other hand, EOs from the stems and leaves of M. alternifolia, which are rich in terpenes, have natural pharmacological properties [40]. In addition, in insect pests, these EOs have an inhibitory effect on feeding [41,42], a lethal effect, and on behavior [43], as well as potential in the control of stored grain pests, due to their fumigant action [41]. However, studies evaluating the potential of these EOs in controlling P. absoluta are still scarce.
In this study, we assessed the insecticidal potential of E. staigeriana and M. alternifolia EOs against P. absoluta. Initially, both oils were chemically characterized to determine their main constituents. Then, their effect was inferred from the dose‒response curves and life history parameters of P. absoluta. The results offer valuable insights into the lethal and sublethal effects of these EOs on P. absoluta, contributing to the understanding of their impact on pest population dynamics. This study provides relevant information on the applicability of the tested EOs to control a pest of global importance.
2. Materials and Methods
2.1. Biological Material and Experimental Conditions
The rearing was established by using eggs and caterpillars of P. absoluta collected from tomato plants grown on the Campus of the Federal University of Lavras (UFLA) and in the field of the company Agroteste LTDA (21°12′ S, 45°03′ W), with no history of application of synthetic chemical pesticides. After collection, the insects were transferred to acrylic cages (60 × 30 × 30 cm) containing tomato (S. lycopersicum cv. Santa Clara) branches for feeding.
After emergence, approximately 500 adults were transferred to a new cage containing tomato shoots that served as oviposition substrates. The adults were offered an aqueous solution of honey (1:1) soaked in moistened cotton wool as a food source. The branches containing the eggs were removed every four days and placed in acrylic cages for maintenance in the laboratory, aiming to provide insects for the bioassays. From the second generation onwards, second-instar and same-generation caterpillars were used in the bioassays. The rearing of P. absoluta and the bioassays were conducted under controlled temperature (25 ± 2°C), relative humidity (70 ± 10%), and photophase (12h) conditions.
2.2. Obtention and Chemical Characterization of EOs
The EOs of M. alternifolia and E. staigeriana were purchased from Empresa Ferquima Indústria e Comércio Ltd.a., Vargem Grande Paulista, São Paulo - Brazil. The chromatographic parameters and equipment used to analyze the EOs were the same as those described in the established methodology [44]. All analyses were performed in triplicate. The analyte concentrations were expressed as the mean relative area percentage of the chromatographic peaks ± standard deviation (n = 3).
2.3. Acute Toxicity of EOs in a Topical Application Against P. absoluta
The EOs of M. alternifolia and E. staigeriana were previously diluted in acetone and applied topically to the dorsum of second-instar P. absoluta caterpillars at a dose of 120 µL EO.caterpillar-1. Each caterpillar received 1 µL of the solution on its back using a microsyringe (Hamilton® 25 µL). Untreated control insects were treated with acetone alone. A completely randomized design was used, with 60 caterpillars per treatment, each replicate consisting of a single treated caterpillar placed in a 5 cm diameter Petri dish containing a leaflet of tomato plant S. lycopersicum cv. Santa Clara for feeding the insects. A piece of filter paper moistened with distilled water was placed under the leaflet to maintain its turgidity. The experiment was repeated twice on different days, totaling 120 replicates per treatment. The mortality evaluations of the caterpillars were performed at 6, 12, 24, 36, 48, and 72 h after the application of the treatments using a stereoscopic microscope (40x). A dead caterpillar was considered one that did not respond to touch with a soft-bristled brush, remaining still.
2.4. Dose–Response and Time–Response Bioassays of EOs
The EO from M. alternifolia was applied to P. absoluta caterpillars at doses of 30, 40, 55, 74 and 100 µg EO.caterpillar-1, and E. staigeriana EO was applied at doses of 50, 62, 78, 97 and 120 µg EO.caterpillar-1. These doses were determined by means of previous tests and by arithmetic progression, aiming to obtain average percentages of mortality between 20 and 90% [45]. The negative control treatment was acetone. All treatments were applied topically to the back of second-instar P. absoluta caterpillars as described in subitem 2.3. A completely randomized design was used, with 60 replicates per treatment, each represented by a treated caterpillar kept in a Petri dish containing one tomato leaflet. The assays were repeated twice.
2.5. Effects of Sublethal Doses of EOs on the Life History Parameters of P. absoluta
For the life table bioassay, approximately 200 adult couples of P. absoluta 72h after emergence were kept in acrylic cages (60 × 30 × 30 cm) containing tomato plants cv. Santa Clara (15 cm tall) for 48h in order to oviposition. The plants were observed daily to verify the appearance of 2nd instar caterpillars when they were removed with the aid of a brush and subjected to the application of the treatments. The EOs were used at doses equivalent to the previously estimated LD50 (lethal dose capable of killing 50% of the insect population) (subitem 2.4). The treatments consisted of acetone (negative control), M. alternifolia (76.5 µL EO.caterpillar-1) and E. staigeriana (78.5 µL EO.caterpillar-1), which were applied to the dorsum of the caterpillars with the aid of a microsyringe (subitem 2.3). Then, the caterpillars were individualized in Petri dishes (2 cm high × 5 cm in diameter) and fed every 72h with leaflets of tomato plants that contained their petioles wrapped in cotton moistened with distilled water and inserted into a microtube. All plates were sealed with perforated PVC film to allow gas exchange and moisture stabilization. The experimental design used was completely randomized, with 3 treatments and 100 replicates, each consisting of a Petri dish with a treated second-instar caterpillar. The duration of the instars, larval and pupal survival, and duration of the larval and pupal stages of the insects were evaluated daily after the topical application of the EOs.
To evaluate the effects of the oils on the reproduction and longevity of adults from treated caterpillars, couples were formed with newly emerged insects (< 48 h of age). Each pair was placed in a Petri dish (2 cm high × 15 cm diameter) covered with PVC plastic film with small holes made with an entomological pin to prevent the escape of insects and enable gas exchange. Each Petri dish contained a piece of cotton soaked in a 1:1 aqueous solution of a tomato plant (five 5 leaflets) with its stem wrapped in moistened cotton and placed in a microtube, which served as a substrate for oviposition. The oviposition period, survival, and longevity of males and females, as well as fecundity, were recorded.
2.6. Statistical Analyses
Data on insect survival over time were analyzed using the nonparametric Kaplan–Meier estimator and subjected to the log-rank test using the survival package [45]. The survival curves were compared using the pairwise multiple comparison test. The median lethal time (LT50), i.e., the time needed to cause 50% mortality in the population was also estimated for each treatment. To determine the median lethal dose (LD50), the data were subjected to logit analysis using the drc package [45]. These analyses were performed using the statistical program R® [47].
The processing of the data analysis for the preparation of the life tables was performed using the TWOSEX-MSCHART program [48]. The means, variances and standard errors of the parameters were compared in pairs between treatments by the bootstrap method with 100,000 replicates [48]. The life table considers the means of survival parameters, life expectancy and fertility until age x and stage j are reached. Differences between treatments were analyzed using the paired bootstrap test with a significance level of 5%.
3. Results
3.1. Chemical Characterization of EOs
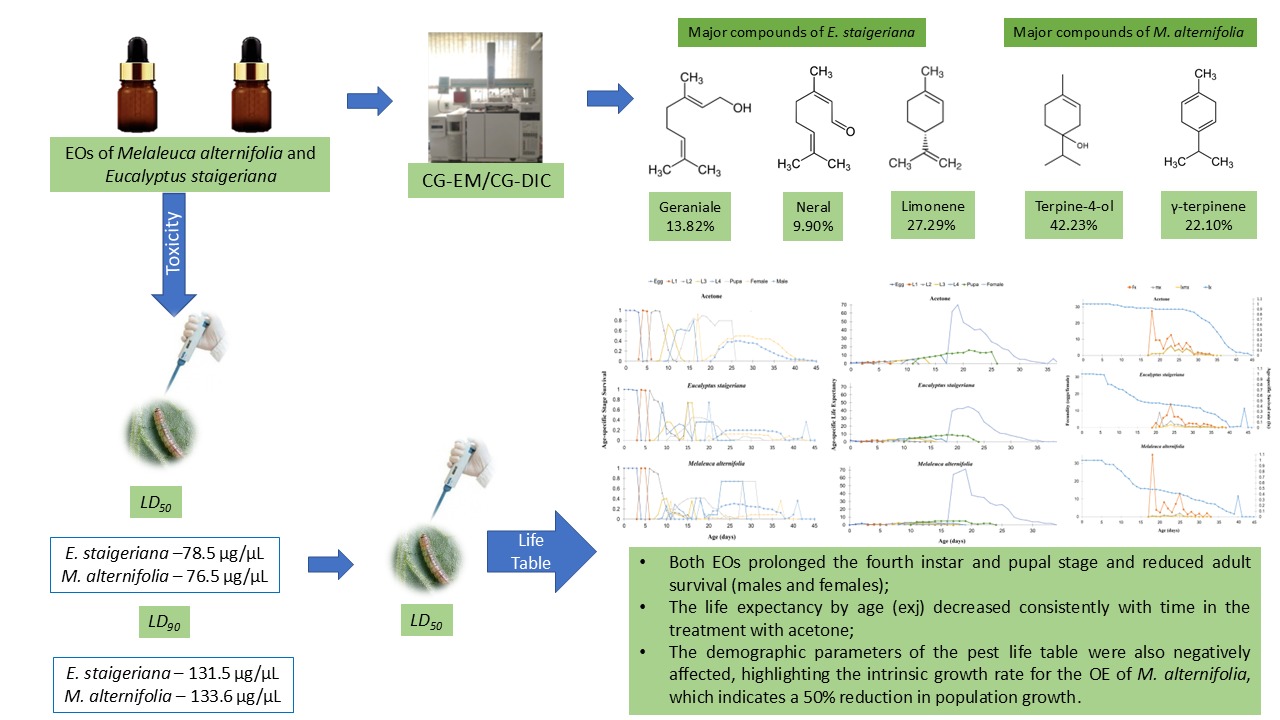
The chemical analysis of the EO of E. staigeriana indicated the presence of 25 constituents, and the main compounds were limonene, neral, and geraniale, with amounts varying from 9.90% to 27.29%. The EO of M. alternifolia presented 19 chemical compounds; 4-terpineol, γ-terpinene and α-terpinene were the major compounds with areas of 42.23%, 22.10% and 10.44%, respectively (Table 1).
3.2. Acute Toxicity of EOs in a Topical Application Against P. absoluta
The EOs of M. alternifolia and E. staigeriana caused 100% mortality of P. absoluta caterpillars at a dose of 120 µL EO.caterpillar-1 36 h after application (ꭓ2 = 183; df = 2; p = < 0.001; Figure 1). Notably, after 6 h, both EOs caused mortality in 50% of the insects.
3.3. Determination of Dose‒Response and Time-Response Curves of EOs
After the application of the EOs on second-instar P. absoluta caterpillars, it was found that the EOs of E. staigeriana and M. alternifolia presented similar LD50 and LD90 values (Table 2).
An increase in the mortality of P. absoluta caterpillars was observed with the increase in the tested doses of E. staigeriana EOs (ꭓ2 = 242; df= 5; p <0.001 and M. alternifolia (ꭓ2 = 223; df=5; p <0.001). The lowest dose needed to reduce the probability of survival was 50 µg for E. staigeriana EO and 40 µg M. alternifolia EO. For the E. staigeriana EO, the dose of 97 µg.µL-1 caused a probability of survival of 56.7% at the end of the evaluation period; while for M. alternifolia EO the dose of 100 µg.µL-1 was enough to cause a probability of survival of 6.67% (Figure 2).
3.4. Effects of Sublethal Doses of EOs on the Life History Parameters of P. absoluta
It was observed that M. alternifolia EO reduced the development time of the third and fourth instars when compared to the other treatments and increased the duration of the pupal stage compared to the acetone control. The EOs of E. staigeriana and M. alternifolia reduced the longevity of adults. The EO of M. alternifolia decreased the life cycle of females, while that of E. staigeriana prolonged this biological parameter (Table 3).
Except for the oviposition period that increased in the treatment with E. staigeriana EO, the reproductive parameters of P. absoluta were not affected whose second-instar caterpillars were treated with the EOs of M. alternifolia and E. staigeriana, as there was no difference compared with treatment with acetone (Table 4).
The age-specific survival rate (sxj) was affected by the treatments. Both EOs prolonged the fourth instar and the pupal stage and reduced adult survival (males and females) (Figure 3).
It was observed that life expectancy by age (exj) decreased consistently with time in the treatment with acetone. However, there were fluctuations throughout the cycle in the other treatments (Figure 4).
The age-specific survival rate (lx) decreased from the 6th to the 9th day of life of the insects in the treatments with EOs. However, in the treatment with acetone, this decrease was observed later on the 28th day of the life cycle. There was a delay and a lower age-specific fecundity peak (fxj) in the treatment with E. staigeriana EO, with approximately 15 eggs on the 24th day, while in the other treatments, the fecundity peak occurred on the 19th day of the gestation cycle, with approximately 30 eggs (Figure 5).
When analyzing the reproductive value by specific age (vxj), it was found that in the treatment with the EO of M. alternifolia, the females reached a lower peak, with approximately 40 eggs, while in the other treatments, the values reached approximately 70 eggs (Figure 6).
The intrinsic rate of increase (r) was reduced by 50% when insects were treated with M. alternifolia EO. Both EOs caused lower values of the finite rate of increase (λ) and net reproductive rate (R0). Regarding the crude reproductive rate (GRR), only the treatment with M. alternifolia EO differed from that with acetone (Table 5).
4. Discussion
The EOs of aromatic plants and their main constituents are considered an alternative to conventional pesticides for the population control of many arthropod pests [15,16]. The EOs of M. alternifolia and E. staigeriana showed insecticidal activity against important insect pests, such as Lucilia cuprina (Wiedemann, 1830) (Diptera: Calliphoridae), C. maculatus, Paropsisterna tigrina (Olivier, 1807) (Coleoptera: Chrysomelidae) and Faex sp. (Coleoptera: Chrysomelidae), S. frugiperda and S. zeamais [36,37,39,49,50]. Although, to the best of the authors’ knowledge, this is the first report of the lethal and sublethal effects of these EOs on P. absoluta larvae, other studies have previously reported the toxicity of Myrtaceae-derived oils against this pest [30,31].
The estimated LD50 for the EOs of M. alternifolia and E. staigeriana for P. absoluta was 76.5 µg of EO.caterpillar-1 and 78.5 µg of EO. caterpillar-1, respectively, values higher than the LD50 of 50.28 µg.caterpillar-1 of the EO of M. alternifolia for Helicoverpa armigera (Hübner, 1805) (Lepidoptera: Noctuidae), which also caused food deterrence [41]. The achievement of high mortality rates in insect pest populations should not be the main objective of the use of botanical insecticides [17,18,19]. Generally, high mortality rates result from the use of high concentrations and large amounts of oil feedstock. Thus, sublethal effects on the life cycle, oviposition, reproduction and demographic parameters are also desirable in the management of insect pest populations.
The investigation of the chemical composition of natural products is very important during the process of developing new compounds, especially considering the variety of flora worldwide [17,51]. In this sense, the present study identified the constituents of the EOs of M. alternifolia and E. staigeriana using GC/MS, and it was found that compound terpinene-4-ol (42.23%) was the most abundant in the EO of M. alternifolia, revealing the chemotype. Several studies have shown that terpinene-4-ol is a monoterpene present in the EOs of many aromatic plants [52,53,54]. The principal components of the EO of M. alternifolia were similar to those reported by other authors, with small variations in the percentages of α-terpinene (11%), γ-terpinene (21%) and terpinene-4-ol (40%) [39,41].
The EOs of Eucalyptus spp. are complex mixtures of various volatile organic components whose composition and proportion vary with plant species [32,34,35,36,38,55,56,57,58]. The GC‒MS results found for the EO of E. staigeriana corroborate those found in the literature [34]. There were small variations in the percentages of limonene (28.7%), geranial (15.2%) and neral (12.16%). However, qualitative and quantitative variations may occur in the chemical composition of the EO of E. staigeriana. For example, limonene, (E)-citral and (Z)-citral were present in the respective amounts of 28.82%, 14.16% and 10.77%, respectively [57], while in another study, chemical analysis showed that the EO of E. staigeriana was formed by limonene (72.9%), cineole (9.5%) and o-cymene (4.59%) [58]. Differences in the chemical composition of EOs are usually due to the location of the plant, climate, soil type, fertility regime, method used to dry the plant material and method of oil extraction [56,57,58].
The constituents of EOs may have different mechanisms of action in target arthropods. The EO of M. alternifolia inhibited the activities of the enzymes acetylcholinesterase (AChE) and glutathione-S-transferase (GST) in H. armigera [41]. On the other hand, terpinene-4-ol strongly inhibited Na+/K+-ATPase activity in Musca domestica (Linnaeus, 1758) (Diptera: Muscidae) [59], whereas the same compound reduced the activity of the enzymes GST, catalase (CAT), and AChE and the sodium/potassium pump (Na+/K+-ATPase) in Plutella xylostella (Linnaeus, 1758) (Lepidoptera: Plutellidae) [60].
Among the major constituents identified in the EO of E. staigeriana, limonene, a monoterpene, is known for its insecticidal activity via multiple routes of exposure, including cuticular penetration (contact toxicity), respiratory absorption (fumigant action), and ingestion [39]. Its toxic effects are frequently attributed to AChE inhibition, leading to the accumulation of acetylcholine (ACh) at synaptic junctions, which results in hyperexcitability, paralysis, and ultimately insect death [61,62,63]. The AChE plays a key role in the hydrolysis of ACh, thereby restoring the resting potential of the neuronal membrane and terminating nerve impulses [61,62,63]. The physiological consequences of this disruption include ataxia and neuromuscular failure [64,65]. In addition, Na+/K+-ATPase, an essential ion pump involved in the maintenance of electrochemical gradients in nerve cells - has also been identified as a molecular target for botanical insecticides, due to its central role in neuronal signaling and homeostasis [65,66].
Caterpillars of S. frugiperda, when subjected to limonene, by topical contact and in high doses, exhibited a state of hyperexcitation, followed by paralysis and death [41], similar to the mode of action of synthetic insecticides belonging to the organophosphate and carbamate chemical groups, which are commonly used to control of P. absoluta. The limonene also causes adverse effects on the nutrition of S. frugiperda, with reduced amounts of lipids, proteins, carbohydrates and total sugars. This occurs in addition to the induction of apoptosis, which affects the reproduction of the insect and essential parameters for its survival and establishment in agricultural crops [41].
In the present study, it was observed that insects exposed to the LD50 of the EOs of M. alternifolia and E. staigeriana had reduced parameters that indicate the population growth of P. absoluta, such as the intrinsic growth rate, finite growth rate and reproduction rate. Toxicological analyses that consider population parameters are more efficient in evaluating the impact of the compound on the insect over prolonged periods when compared to studies of the lethal effect alone [67]. Therefore, the use of life tables by developmental stage for both sexes makes it possible to more accurately determine the population changes of the arthropod pest, as this incorporates the dynamic rates of development over time and the differentiation of the individual growth stages [48,68]. Thus, the results provide evidence supporting the integration of EOs from E. staigeriana and M. alternifolia into IPM strategies targeting P. absoluta.
5. Conclusions
The EO of M. alternifolia has nineteen chemical compounds the majority ones being terpinene-4-ol, γ-terpinene and α-terpinene, and that of E. staigeriana contains twenty-five constituents, the majority compounds being limonene, geranial and neral. The EOs of M. alternifolia and E. staigeriana are toxic to P. absoluta and negatively affected the duration of the pupal stage and longevity of the insects. The demographic parameters of the pest life table were also negatively affected, highlighting the intrinsic growth rate for the OE of M. alternifolia, which indicates a 50% reduction in population growth. Thus, the results found in the present study provide insights for the use of EOs from E. staigeriana and M. alternifolia in integrated management programs for P. absoluta.
Author Contributions
Conceptualization, B.C.F.B., G.A.C. and D.S.A.; methodology, B.C.F.B., A.F.L., K.G.F, J.A.C.O., S.K.V.B., G.A.C. and D.S.A; software, B.C.F.B., K.G.F.; validation, S.K.V.B., G.A.C. and D.S.A.; investigation, B.C.F.B., A.F.L., K.G.F, J.A.C.O. and V.C.C.; data curation, B.C.F.B., A.F.L., K.G.F, J.A.C.O and D.S.A.; writing – preparation of original draft, B.C.F.B., J.A.C.O., G.A.C. and D.S.A.; writing – review and editing, B.C.F.B., J.A.C.O., V.C.C.; G.A.C. and D.S.A.; supervision, G.A.C. and D.S.A.; project administration, G.A.C. and D.S.A.
All authors read and agreed to the published version of the manuscript.
Acknowledgments
The authors thank the National Council for Scientific and Technological Development (CNPq), Minas Gerais State Foundation (FAPEMIG), and the Coordination for the Improvement of Higher Education Personnel (CAPES), Brazil the support.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| EOs | Essential Oils |
| EO | Essential Oil |
References
- EPPO. Tuta absoluta (GNORAB). Available online: https://gd.eppo.int/taxon/GNORAB/distribution (accessed on 15 May 2023).
- Simoglou, K.Β.; Stavrakaki, M.; Alipranti, K.; Mylona, K.; Roditakis, E. Understanding Greenhouse Tomato (Solanum lycopersicum L.) Growers’ Perceptions for Optimal Phthorimaea absoluta (Meyrick) Management—A Survey in Greece. Agriculture 2024, 14, 2291. [Google Scholar] [CrossRef]
- Lee, M.H.; Jeong, D.; Lee, G.-S.; Paik, C. First report of Phthorimaea absoluta (Lepidoptera: Gelechiidae) in Korea. J. Integr. Pest Manag. 2024, 15, 36. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Picanço, M.C. The tomato borer Tuta absoluta in South Aamerica: Pest status, management and insecticide resistance. EPPO Bull 2011–2016. 2012. [Google Scholar] [CrossRef]
- Biondi, A.; Guedes, R.N.C.; Wan, F.H.; Desneux, N. Ecology, worldwide spread, and management of the invasive South American tomato pinworm, Tuta absoluta: Past, present, and future. Annu. Rev. Entomol 2018, 63, 239–258. [Google Scholar] [CrossRef]
- Biratu, W. Review on the effect of climate change on tomato (Solanum lycopersicon) production in Africa and mitigation strategies. J. Nat. Sci. Res. 2018, 8, 62–70. [Google Scholar]
- Mahlangu, L.; Sibisi, P.; Nofemela, R.S.; Ngmenzuma, T.; Ntushelo, K. The differential effects of Tuta absoluta infestations on the physiological processes and growth of tomato, potato, and eggplant. Insects 2022, 13, 754. [Google Scholar] [CrossRef]
- Haddi, K.; Berger, M.; Bielza, P.; Rapisarda, C.; Williamson, M.S.; Moores, G.; et al. Mutation in the ace-1 gene of the tomato leaf miner (Tuta absoluta) associated with organophosphates resistance. J. Appl. Entomol 2017, 141, 612–619. [Google Scholar] [CrossRef]
- Haddi, K.; Berger, M.; Bielza, P.; Cifuentes, D.; Field, L.M.; Gorman, K.; et al. Identification of mutations associated with pyrethroid resistance in the voltage-gated sodium channel of the tomato leaf miner (Tuta absoluta). Insect Biochem. Mol. Biol 2012, 42, 506–513. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Roditakis, E.; Campos, M.R.; Haddi, K.; Bielza, P.; Siqueira, H.A.A.; et al. Insecticide resistance in the tomato pinworm Tuta absoluta: Patterns, spread, mechanisms, management and outlook. J. Pest Sci. 2019, 92, 1329–1342. [Google Scholar] [CrossRef]
- Silva, G.A.; Picanço, M.C.; Bacci, L.; Crespo, A.L.B.; Rosado, J.F.; Guedes, R.N.C. Control failure likelihood and spatial dependence of insecticide resistance in the tomato pinworm, Tuta absoluta. Pest Manag. Sci. 2011, 67, 913–920. [Google Scholar] [CrossRef]
- Biondi, A.; Zappalà, L.; Stark, J.D.; Desneux, N. Do biopesticides affect the demographic traits of a parasitoid wasp and its biocontrol services through sublethal effects? PLoS ONE 2013, 8, 76548. [Google Scholar] [CrossRef]
- Roditakis, E.; Skarmoutsou, C.; Staurakaki, M. Toxicity of insecticides to populations of tomato borer Tuta absoluta (Meyrick) from Greece. Pest Manag. Sci. 2013, 69, 834–840. [Google Scholar] [CrossRef]
- Sola, P.; Mvumi, B.M.; Ogendo, J.O.; Mponda, O.; Kamanula, J.F.; Nyirenda, S.P.; et al. Botanical pesticide production, trade and regulatory mechanisms in Sub-Saharan Africa: Making a case for plant-based pesticidal products. Food Secur. 2014, 6, 369–384. [Google Scholar] [CrossRef]
- Soares, M.A.; Campos, M.R.; Passos, L.C.; Carvalho, G.A.; Haro, M.M.; Lavoir, A.V.; et al. Botanical insecticide and natural enemies: A potential combination for pest management against Tuta absoluta. J. Pest Sci. 2019, 92, 1433–1443. [Google Scholar] [CrossRef]
- Salazar, A.M.; Arismendi, N.; López, M.D.; Vargas, M.; Schoebitz, M.; Palacio, D.A.; et al. Stability of the oil-based nanoemulsion of Laureliopsis philippiana (Looser) and its insecticidal activity against tomato borer (Tuta absoluta Meyrick). Ind. Crops Prod. 2022, 188, 115635. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides in the twenty-first century-fulfilling their promise? Annu. Rev. Entomol. 2020, 65, 233–249. [Google Scholar] [CrossRef]
- Tak, J.H.; Isman, M.B. Metabolism of citral, the major constituent of lemongrass oil, in the cabbage looper, Trichoplusia ni, and effects of enzyme inhibitors on toxicity and metabolism. Pestic. Biochem. Physiol. 2016, 133, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.F.; do Prado, R.L.; Gonçalves, G.L.P.; Maimone, N.M.; Gissi, D.S. de Lira et al. Searching for bioactive compounds from solanaceae: Lethal and sublethal toxicity to Spodoptera frugiperda and untargeted metabolomics approaches. J. Pest Sci. 2022, 95, 1317–1329. [Google Scholar] [CrossRef]
- Lima, A.F.; Ribeiro, L.P.; Lira, S.P.; Carvalho, G.A.; Vendramim, J.D. Growth inhibitory activities and feeding deterrence of solanaceae-based derivatives on fall armyworm. Agriculture. 2023, 13, 420. [Google Scholar] [CrossRef]
- Mossa, A.-T.H. Green pesticides: Essential oils as biopesticides in insect-pest management. J. Environ. Sci. Technol. 2016, 9, 354–378. [Google Scholar] [CrossRef]
- Nollet, L.M.; Rathore, H.S. Green pesticides handbook: Essential oils for pest control; CRC Press: Boca Raton, 2017. [Google Scholar]
- Acheuk, F.; Basiouni, S.; Shehata, A.A.; Dick, K.; Hajri, H.; Lasram, S.; et al. Status and Prospects of Botanical Biopesticides in Europe and Mediterranean Countries. Biomolecules 2022, 12, 311. [Google Scholar] [CrossRef]
- Bibiano, C.S.; Alves, D.S.; Freire, B.C.; Bertolucci, S.K.V.; Carvalho, G.A. Toxicity of essential oils and pure compounds of lamiaceae species against Spodoptera frugiperda (Lepidoptera: Noctuidae) and their safety for the nontarget organism Trichogramma pretiosum (Hymenoptera: Trichogrammatidae). Crop Prot. 2022, 158, 106011. [Google Scholar] [CrossRef]
- Chaudhari, A.K.; Singh, V.K.; Kedia, A.; Das, S.; Dubey, N.K. Essential oils and their bioactive compounds as eco-friendly novel green pesticides for management of storage insect pests: Prospects and retrospects. Environ. Sci. Pollut. Res. 2021, 15, 18918–18940. [Google Scholar] [CrossRef] [PubMed]
- Tak, J.-H.; Isman, M.B. Enhanced cuticular penetration as the mechanism for synergy of insecticidal constituents of rosemary essential oil in Trichoplusia ni. Sci. Rep. 2015, 5, 12690. [Google Scholar] [CrossRef]
- Ngongang, M.D.T.; Eke, P.; Sameza, M.L.; Ngo, M.N.L.; Lordon, C.D.; et al. Chemical constituents of essential oils from Thymus vulgaris and Cymbopogon citratus and their insecticidal potential against the tomato borer, Tuta absoluta (Lepidoptera: Gelechiidae). Int. J. Trop. Insect Sci. 2022, 42, 31–43. [Google Scholar] [CrossRef]
- Essoung, F.R.E.; Tadjong, A.T.; Chhabra, S.C.; Mohamed, S.A.; Hassanali, A. Repellence and fumigant toxicity of essential oils of Ocimum gratissimum and Ocimum kilimandscharicum on Tuta absoluta (Lepidoptera: Gelechiidae). Environ. Sci. Pollut. Res. 2020, 27, 37963–37976. [Google Scholar] [CrossRef]
- Yarou, B.B.; Bawin, T.; Boullis, A.; Heukin, S.; Lognay, G.; Verheggen, F.J.; et al. Oviposition deterrent activity of basil plants and their essentials oils against Tuta absoluta (Lepidoptera: Gelechiidae). Environ. Sci. Pollut. Res. 2018, 25, 29880–29888. [Google Scholar] [CrossRef] [PubMed]
- Cherif, A.; Mansour, R.; Ncibi, S.; WHached, W.; Grissa-Lebdi, K. Chemical composition and fumigant toxicity of five essential oils toward Tuta absoluta and its mirid predator Macrolophus pygmaeus. J. Plant Dis. Prot. 2025, 132, 34. [Google Scholar] [CrossRef]
- Lo Pinto, M.; Vella, L.; Agrò, A. Oviposition deterrence and repellent activities of selected essential oils against Tuta absoluta Meyrick (Lepidoptera: Gelechiidae): Laboratory and greenhouse investigations. Int. J. Trop. Insect Sci. 2022, 42, 3455–3464. [Google Scholar] [CrossRef]
- Nouri-Ganbalani, G.; Ebadollahi, A.; Nouri, A. Chemical composition of the essential oil of Eucalyptus procera Dehnh. and its insecticidal effects against two stored product insects. J. Essent. Oil Bear. Plants. 2016, 19, 1234–1242. [Google Scholar] [CrossRef]
- Ebadollahi, A. Essential Oils Isolated from Myrtaceae Family as Natural Insecticides. Annu. Res. Rev. Biol. 2013, 3, 148–175. [Google Scholar]
- Pedrotti, C.; Trentin, T.R.; Cavião, H.C.; Vilasboa, J.; Scariot, F.J.; Echeverrigaray, S.; et al. Eucalyptus staigeriana essential oil in the control of postharvest fungal rots and on the sensory analysis of grapes. Pesqui. Agropecuária Bras. 2022, 57, 02782. [Google Scholar] [CrossRef]
- Brito, J.P.; Baptistussi, R.C.; Funichello, M.; Oliveira, J.E.M.; de Bortoli, S.A. Efeito de óleos essenciais de Eucalyptus spp. sobre Zabrotes subfasciatus (Boh., 1833) (Coleoptera: Bruchidae) e Callosobruchus maculatus (Fabr., 1775) (Coleoptera: Bruchidae) em duas espécies de feijões. Boletín Sanid. Veg. 2006, 32, 573–580. [Google Scholar]
- Gusmão, N.M.S.; de Oliveira, J.V.; Navarro, D.M.A.F.; Dutra, K.A.; da Silva, W.A.; Wanderley, M.J.A. Contact and fumigant toxicity and repellency of Eucalyptus citriodora Hook., Eucalyptus staigeriana F., Cymbopogon winterianus Jowitt and Foeniculum vulgare Mill. essential oils in the management of Callosobruchus maculatus. J. Stored Prod. Res. 2013, 54, 41–47. [Google Scholar] [CrossRef]
- Araújo, A.M.N.; Oliveira, J.V.; França, S.M.; Navarro, D.M.A.F.; Barbosa, D.R.S.; Dutra, K.A. Toxicity and repellency of essential oils in the management of Sitophilus zeamais. Rev. Bras. Eng. Agrícola e Ambient. 2019, 23, 372–377. [Google Scholar] [CrossRef]
- Maciel, M.V.; Morais, S.M.; Bevilaqua, C.M.L.; Silva, R.A.; Barros, R.S.; Sousa, R.N.; et al. Chemical composition of Eucalyptus spp. essential oils and their insecticidal effects on Lutzomyia longipalpis. Vet. Parasitol. 2010, 167, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Cruz, G.S.; Wanderley-Teixeira, V.; Oliveira, J.V.; D’Assunção, C.G.; Cunha, F.M.; Teixeira, Á.A.C.; et al. Effect of trans-anethole, limonene and your combination in nutritional components and their reflection on reproductive parameters and testicular apoptosis in Spodoptera frugiperda (Lepidoptera: Noctuidae). Chem. Biol. Interact. 2017, 263, 74–80. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, S.; Lin, G.; Chen, J.; Li, H.; Xiao, Y.; Chen, X.; et al. The chromosome-level Melaleuca alternifolia genome provides insights into the molecular mechanisms underlying terpenoids biosynthesis. Ind. Crops Prod. 2022, 189, 115819. [Google Scholar] [CrossRef]
- Liao, M.; Xiao, J.-J.; Zhou, L.-J.; Yao, X.; Tang, F.; Hua, R.-M.; et al. Chemical composition, insecticidal and biochemical effects of Melaleuca alternifolia essential oil on the Helicoverpa armigera. J. Appl. Entomol. 2017, 141, 721–728. [Google Scholar] [CrossRef]
- Ismail, S.M.; Hassan, N.A.; Wahba, T.F.; Shaker, N. Chemical composition and bioactivities of Melaleuca alternifolia essential oil and its main constituents against Spodoptera littoralis (Boisaduval, 1833). Bull. Natl. Res. Cent. 2022, 46, 157. [Google Scholar] [CrossRef]
- Chohan, T.; Chohan, T.; Zhou, L.; Yang, Q.; Min, L.; Cao, H. Repellency, toxicity, gene expression profiling and in silico studies to explore insecticidal potential of Melaleuca alternifolia essential oil against Myzus persicae. Toxins (Basel). 2018, 10, 425. [Google Scholar] [CrossRef]
- Oliveira, J.A.C.; Ferreira, L.S.; Garcia, I.P.; Santos, H.L.; Ferreira, G.S.; Rocha, J.P.M.; et al. Eugenia uniflora, Melaleuca armillaris, and Schinus molle essential oils to manage larvae of the filarial vector Culex quinquefasciatus (Diptera: Culicidae). Environ. Sci. Pollut. Res. 2022, 29, 34749–34758. [Google Scholar] [CrossRef]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, 0146021. [Google Scholar] [CrossRef] [PubMed]
- Therneau, T. A package for survival analysis in R. 2023. [Google Scholar]
- R CoreTeam R: A language and environment for statistical computing. 2023.
- Chi, H.; Güncan, A.; Kavousi, A.; Gharakhani, G.; Atlihan, R.; Özgökçe, M.S.; et al. R. TWOSEX-MSChart: The key tool for life table research and education. Entomol. Gen. 2022, 42, 845–849. [Google Scholar] [CrossRef]
- Bustos-Segura, C.; Fornoni, J.; Núñez-Farfán, J. Evolutionary changes in plant tolerance against herbivory through a resurrection experiment. J. Evol. Biol. 2014, 27, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Callander, J.T.; James, P.J. Insecticidal and repellent effects of tea tree (Melaleuca alternifolia) oil against Lucilia cuprina. Vet. Parasitol. 2012, 184, 271–278. [Google Scholar] [CrossRef]
- Isman, M.B.; Miresmailli, S.; MacHial, C. Commercial opportunities for pesticides based on plant essential oils in agriculture, industry and consumer products. Phytochem. Rev. 2011, 10, 197–204. [Google Scholar] [CrossRef]
- Nóbrega, F.F.F.; Salvadori, M.G.S.S.; Masson, C.J.; Mello, C.F.; Nascimento, T.S.; Leal-Cardoso, J.H.; et al. Monoterpenoid terpinen-4-ol exhibits anticonvulsant activity in behavioural and electrophysiological studies. Oxid. Med. Cell. Longev. 2014, 1–9. [Google Scholar] [CrossRef]
- Raina, A.P.; Abraham, Z. Chemical composition of essential oils obtained from plant parts of Alpinia calcarata Rosc. (Lesser Galangal) germplasm from South India. J. Essent. Oil Res. 2015, 27, 238–243. [Google Scholar] [CrossRef]
- Baldissera, M.D.; Grando, T.H.; Souza, C.F.; Gressler, L.T.; Stefani, L.M.; da Silva, A.S.; et al. In vitro and in vivo action of terpinen-4-ol, γ-terpinene, and α-terpinene against Trypanosoma evansi. Exp. Parasitol. 2016, 162, 43–48. [Google Scholar] [CrossRef]
- Dhakad, A.K.; Pandey, V.V.; Beg, S.; Rawat, J.M.; Singh, A. Biological, medicinal and toxicological significance of Eucalyptus leaf essential oil: A Review. J. Sci. Food Agric. 2018, 98, 833–848. [Google Scholar] [CrossRef]
- Khammassi, M.; Polito, F.; Amri, I.; Khedhri, S.; Hamrouni, L.; Nazzaro, F.; et al. Chemical composition and phytotoxic, antibacterial and antibiofilm activity of the essential oils of Eucalyptus occidentalis, E. striaticalyx and E. stricklandii. Molecules 2022, 27, 5820. [Google Scholar] [CrossRef] [PubMed]
- Macedo, I.T.F.; Bevilaqua, C.M.L.; de Oliveira, L.M.B.; Camurça-Vasconcelos, A.L.F.; Vieira, L.S.; Oliveira, F.R.; et al. Anthelmintic effect of Eucalyptus staigeriana essential oil against goat gastrointestinal nematodes. Vet. Parasitol. 2010, 173, 93–98. [Google Scholar] [CrossRef]
- Ribeiro, W.L.C.; Macedo, I.T.F.; dos Santos, J.M.L.; de Oliveira, E.F.; Camurça-Vasconcelos, A.L.F.; de Paula, H.C.B.; et al. Activity of chitosan-encapsulated Eucalyptus staigeriana essential oil on Haemonchus contortus. Exp. Parasitol. 2013, 135, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Zhi-Bo, G.; Zhi-Qing, M.; Jun-Tao, F.; Xing, Z. Inhibition of Na+,K+-ATPase in housefly (Musca domestica L.) by terpinen- 4-ol and its ester derivatives. Agric. Sci. China. 2009, 8, 1492–1497. [Google Scholar] [CrossRef]
- Huang, X.; Du, L.; Liu, T.; Ma, R.; Liu, X.; Yuan, H.; Liu, S. Insecticidal activity of a component, (-)-4-terpineol, isolated from the essential oil of Artemisia lavandulaefolia DC. against Plutella xylostella (L.). Insects 2022, 13, 1126. [Google Scholar] [CrossRef] [PubMed]
- Rajashekar, Y.; Raghavendra, A.; Bakthavatsalam, N. Acetylcholinesterase inhibition by biofumigant (coumaran) from leaves of Lantana camara in stored grain and household insect pests. Biomed Res. Int. 2014, 1–6. [Google Scholar] [CrossRef]
- Bezerra, C.S.; Pott, A.; Elifio-Esposito, S.; Dalarmi, L.; Fialho, K.N.; Burci, L.M.; et al. Effect of donepezil, tacrine, galantamine and rivastigmine on acetylcholinesterase inhibition in Dugesia tigrina. Molecules. 2016, 21, 53. [Google Scholar] [CrossRef]
- Johnson, T.O.; Abolaji, A.O.; Omale, S.; Longdet, I.Y.; Kutshik, R.J.; Oyetayo, B.O.; et al. Benzo[a]Pyrene and Benzo[a]Pyrene-7,8-Dihydrodiol-9,10-Epoxide induced locomotor and reproductive senescence and altered biochemical parameters of oxidative damage in canton-S Drosophila melanogaster. Toxicol. Reports 2021, 8, 571–580. [Google Scholar] [CrossRef]
- Lu, Y.H.; He, Y.P.; Gao, X.W. Comparative studies on acetylcholinesterase characteristics between the aphids, Sitobion avenae and Rhopalosiphum padi. J. Insect Sci. 2013, 13, 1–9. [Google Scholar] [CrossRef]
- Oboh, G.; Ademosun, A.O.; Olumuyiwa, T.A.; Olasehinde, T.A.; Ademiluyi, A.O.; Adeyemo, A.C. Insecticidal activity of essential oil from orange peels (Citrus sinensis) against Tribolium confusum, Callosobruchus maculatus and Sitophilus oryzae and its inhibitory effects on acetylcholinesterase and Na+/K+-ATPase activities. Phytoparasitica 2017, 45, 501–508. [Google Scholar] [CrossRef]
- Cheng, D.; Feng, M.; Ji, Y.; Wu, W.; Hu, Z. Effects of Celangulin IV and V from Celastrus angulatus Maxim on Na + /K + -ATPase Activities of the Oriental Armyworm (Lepidoptera: Noctuidae). J. Insect Sci. 2016, 16, 59. [Google Scholar] [CrossRef] [PubMed]
- Stark, J.D.; Banks, J.E. Population-level effects of pesticides and other toxicants on arthropods. Annu. Rev. Entomol. 2003, 48, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
Figure 1.
Mortality of Phthorimaea absoluta caterpillars, over time, treated topically with the essential oils of Eucalyptus staigeriana and Melaleuca alternifolia at a dose of 120 µg of essential oil.caterpillar-1.
Figure 1.
Mortality of Phthorimaea absoluta caterpillars, over time, treated topically with the essential oils of Eucalyptus staigeriana and Melaleuca alternifolia at a dose of 120 µg of essential oil.caterpillar-1.
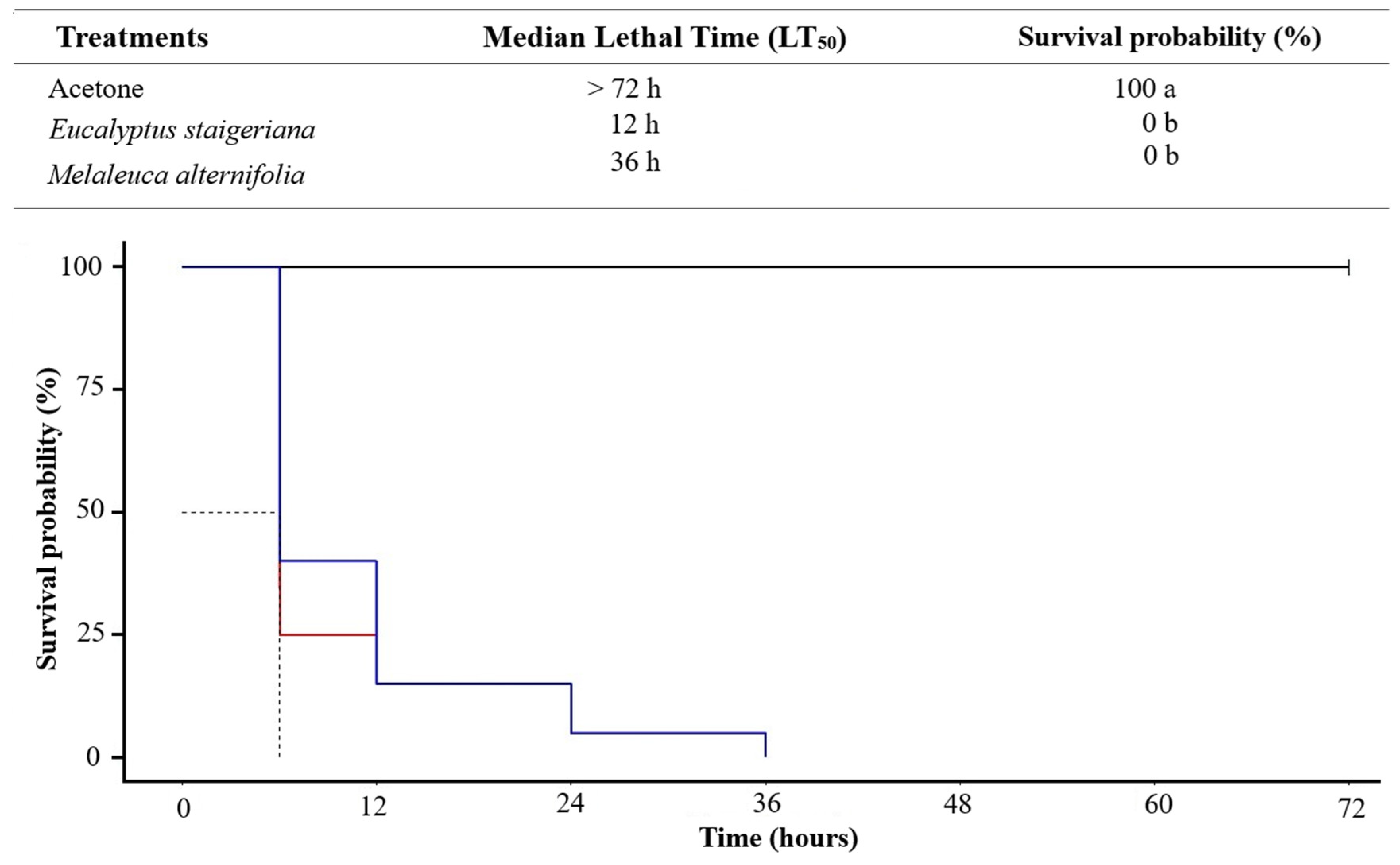
Figure 2.
Survival analysis, over time, of Phthorimaea absoluta caterpillars that were topically treated with different doses of essential oils of a) Eucalyptus staigeriana and b) Melaleuca alternifolia. EO: essential oil.
Figure 2.
Survival analysis, over time, of Phthorimaea absoluta caterpillars that were topically treated with different doses of essential oils of a) Eucalyptus staigeriana and b) Melaleuca alternifolia. EO: essential oil.
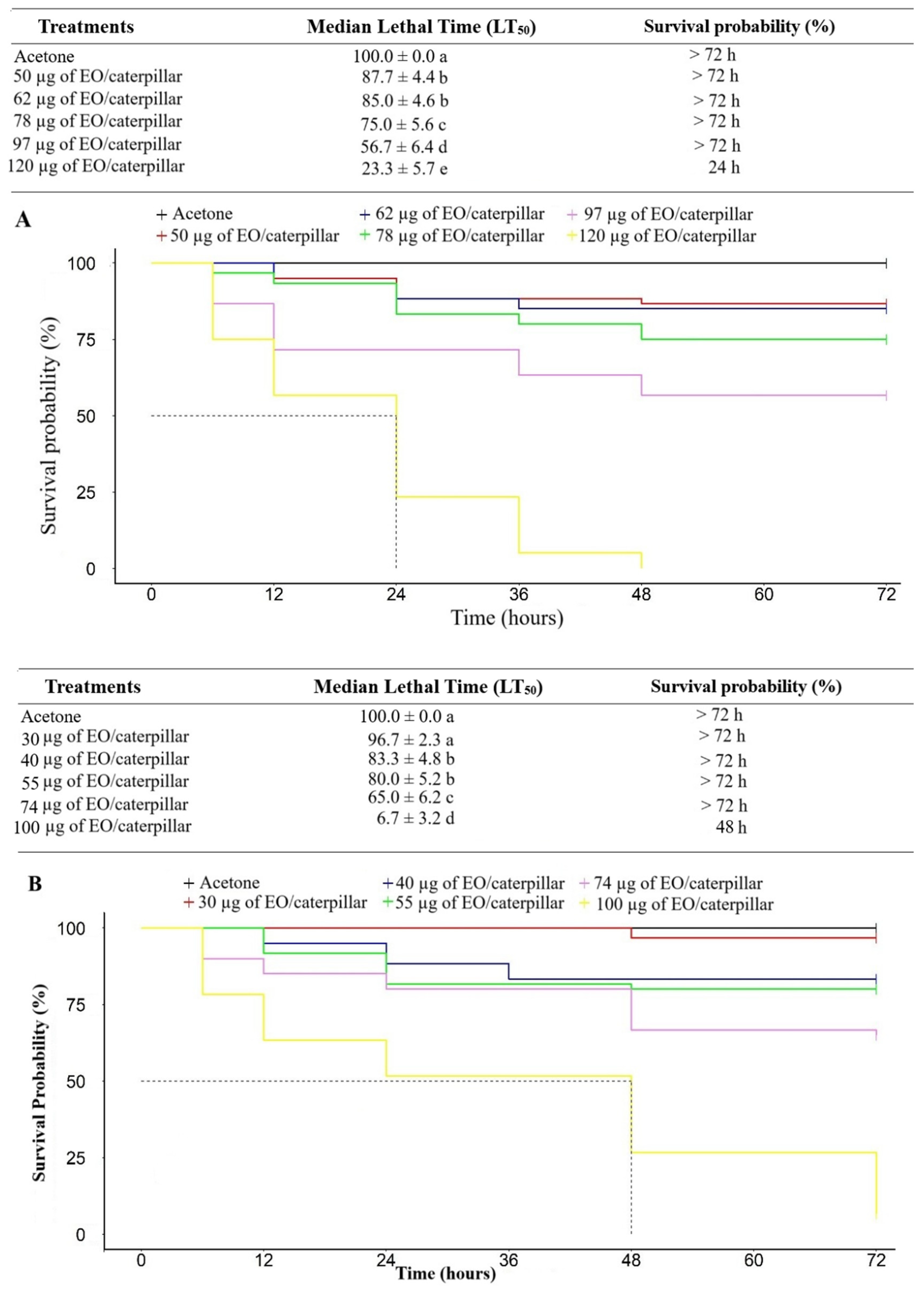
Figure 3.
Age-specific survival rate (sxj) of Phthorimaea absoluta in the acetone treatments, Melaleuca alternifolia (76.5 µg of essential oil.caterpillar-1) and Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1). L1 = 1st instar caterpillar, L2 = 2nd instar caterpillar, L3 = 3rd instar and L4 = 4th instar caterpillar.
Figure 3.
Age-specific survival rate (sxj) of Phthorimaea absoluta in the acetone treatments, Melaleuca alternifolia (76.5 µg of essential oil.caterpillar-1) and Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1). L1 = 1st instar caterpillar, L2 = 2nd instar caterpillar, L3 = 3rd instar and L4 = 4th instar caterpillar.
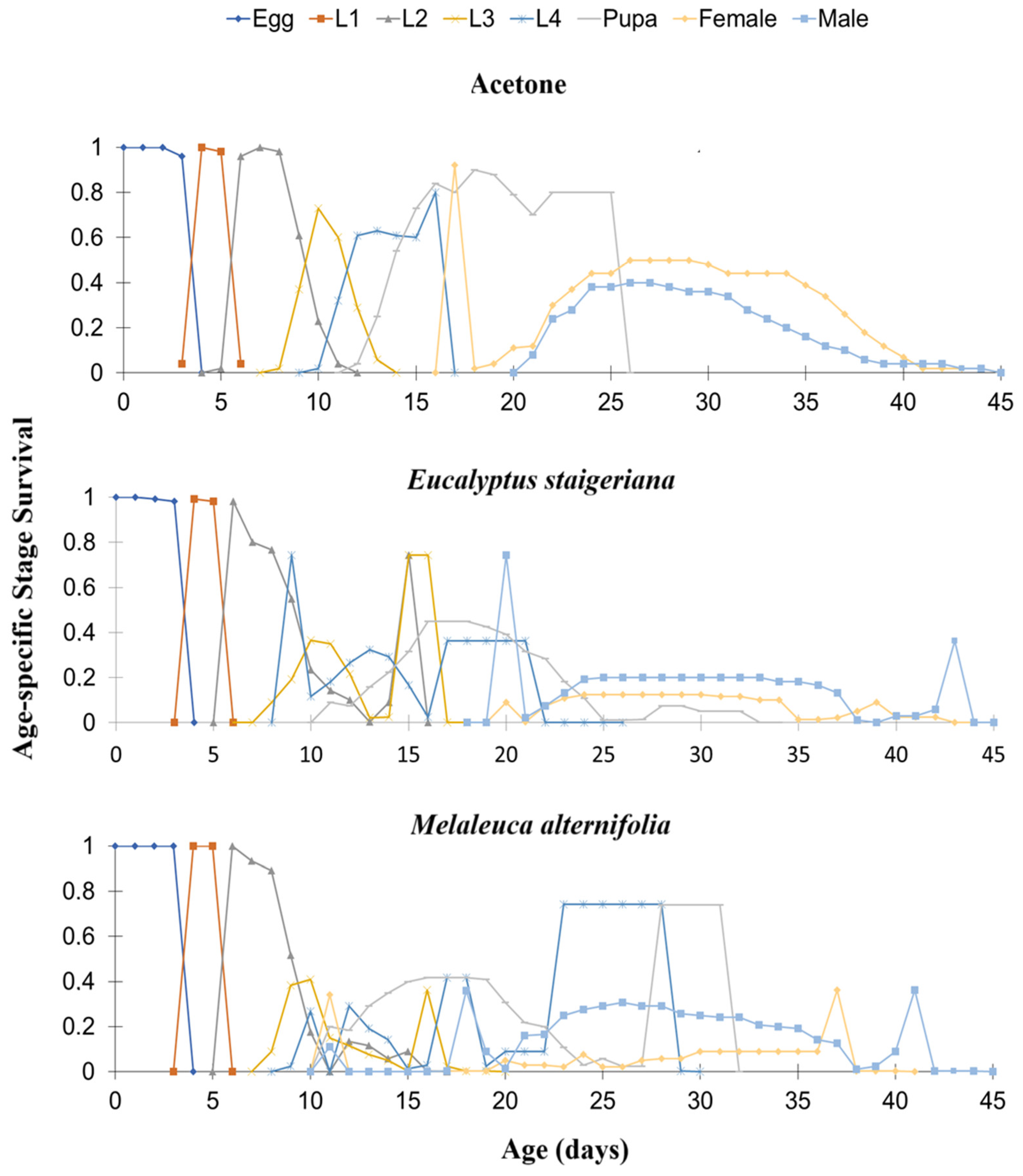
Figure 4.
Age-specific life expectancy (exj) of Phthorimaea absoluta in the acetone, Melaleuca alternifolia (76.5 µg of essential oil.caterpillar-1) and Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1) treatments. L1 = 1st instar caterpillar, L2 = 2nd instar caterpillar, L3 = 3rd instar and L4 = 4th instar caterpillar.
Figure 4.
Age-specific life expectancy (exj) of Phthorimaea absoluta in the acetone, Melaleuca alternifolia (76.5 µg of essential oil.caterpillar-1) and Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1) treatments. L1 = 1st instar caterpillar, L2 = 2nd instar caterpillar, L3 = 3rd instar and L4 = 4th instar caterpillar.
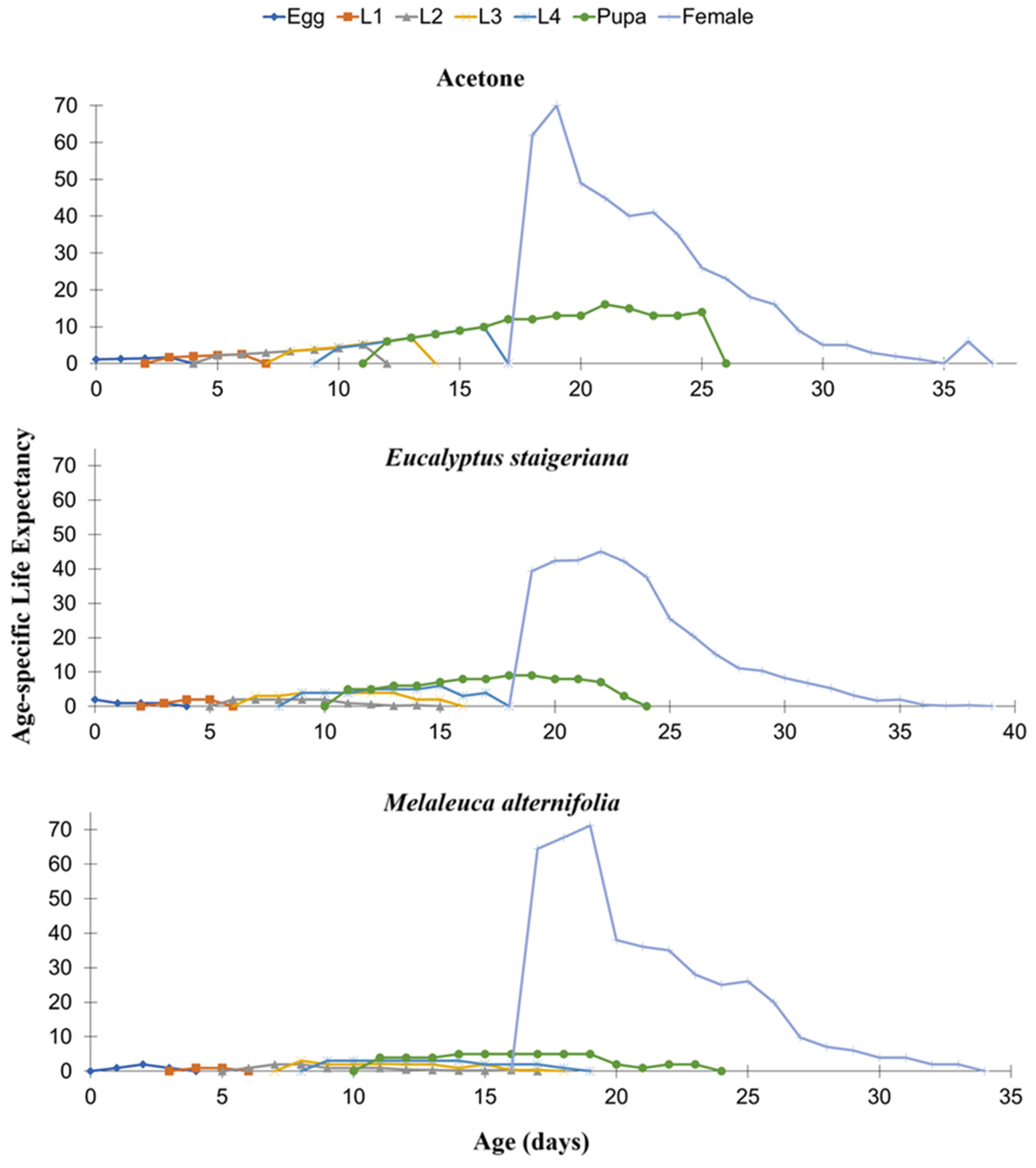
Figure 5.
Age-specific survival rate (lx), age-specific fertility and stage of development (fx), age-specific fertility (mx) and age-specific maternity (lxmx) of Phthorimaea absoluta in the acetone, Melaleuca alternifolia (76.5 µg of essential oil.caterpillar-1) and Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1) treatments.
Figure 5.
Age-specific survival rate (lx), age-specific fertility and stage of development (fx), age-specific fertility (mx) and age-specific maternity (lxmx) of Phthorimaea absoluta in the acetone, Melaleuca alternifolia (76.5 µg of essential oil.caterpillar-1) and Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1) treatments.
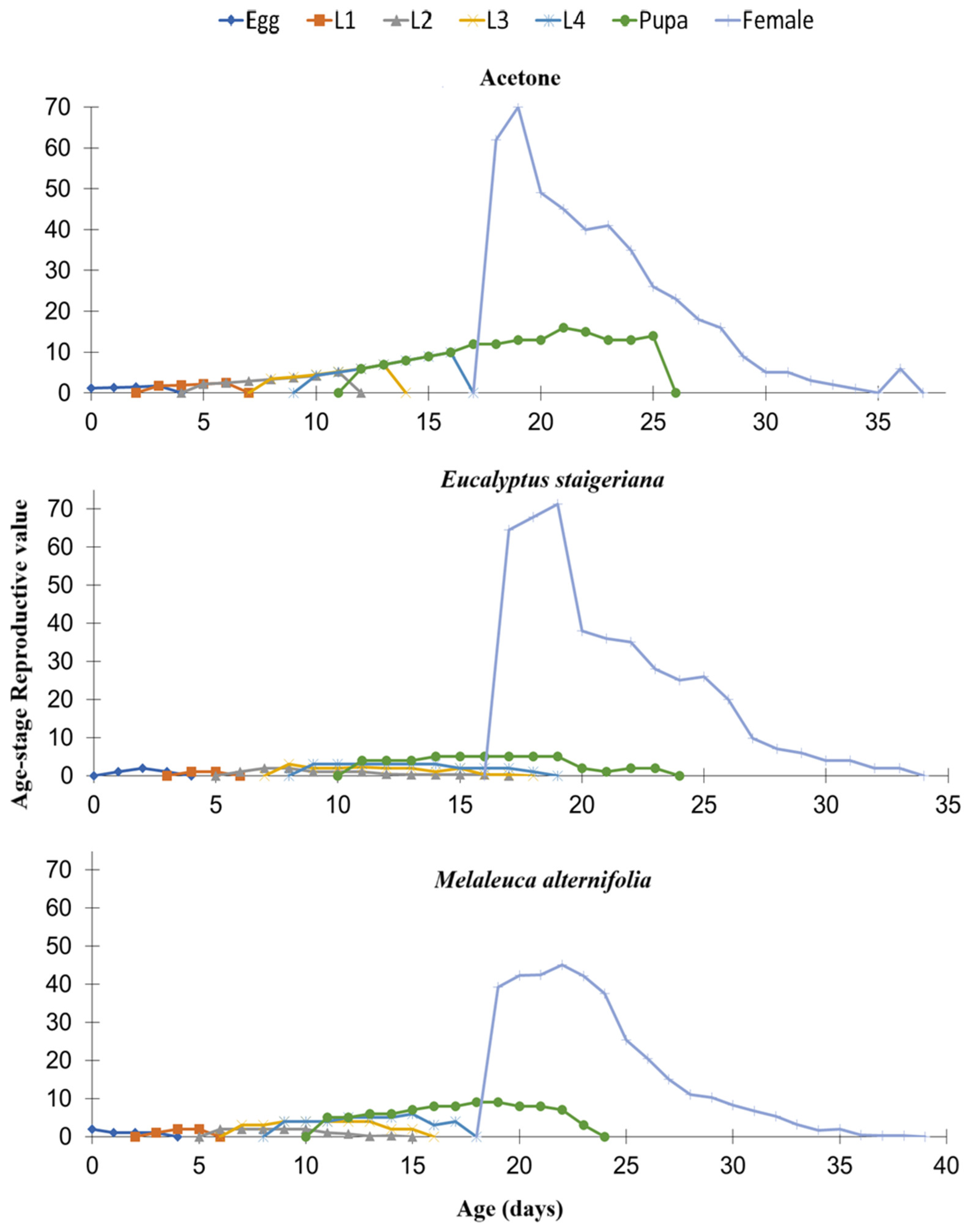
Figure 6.
Reproductive value by age (vxd) of Phthorimaea absoluta in the acetone, Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1) and Melaleuca alternifolia (76.5 µg essential oil.caterpillar-1) treatments. L1 = 1st instar caterpillar, L2 = 2nd instar caterpillar, L3 = 3rd instar and L4 = 4th instar caterpillar.
Figure 6.
Reproductive value by age (vxd) of Phthorimaea absoluta in the acetone, Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1) and Melaleuca alternifolia (76.5 µg essential oil.caterpillar-1) treatments. L1 = 1st instar caterpillar, L2 = 2nd instar caterpillar, L3 = 3rd instar and L4 = 4th instar caterpillar.
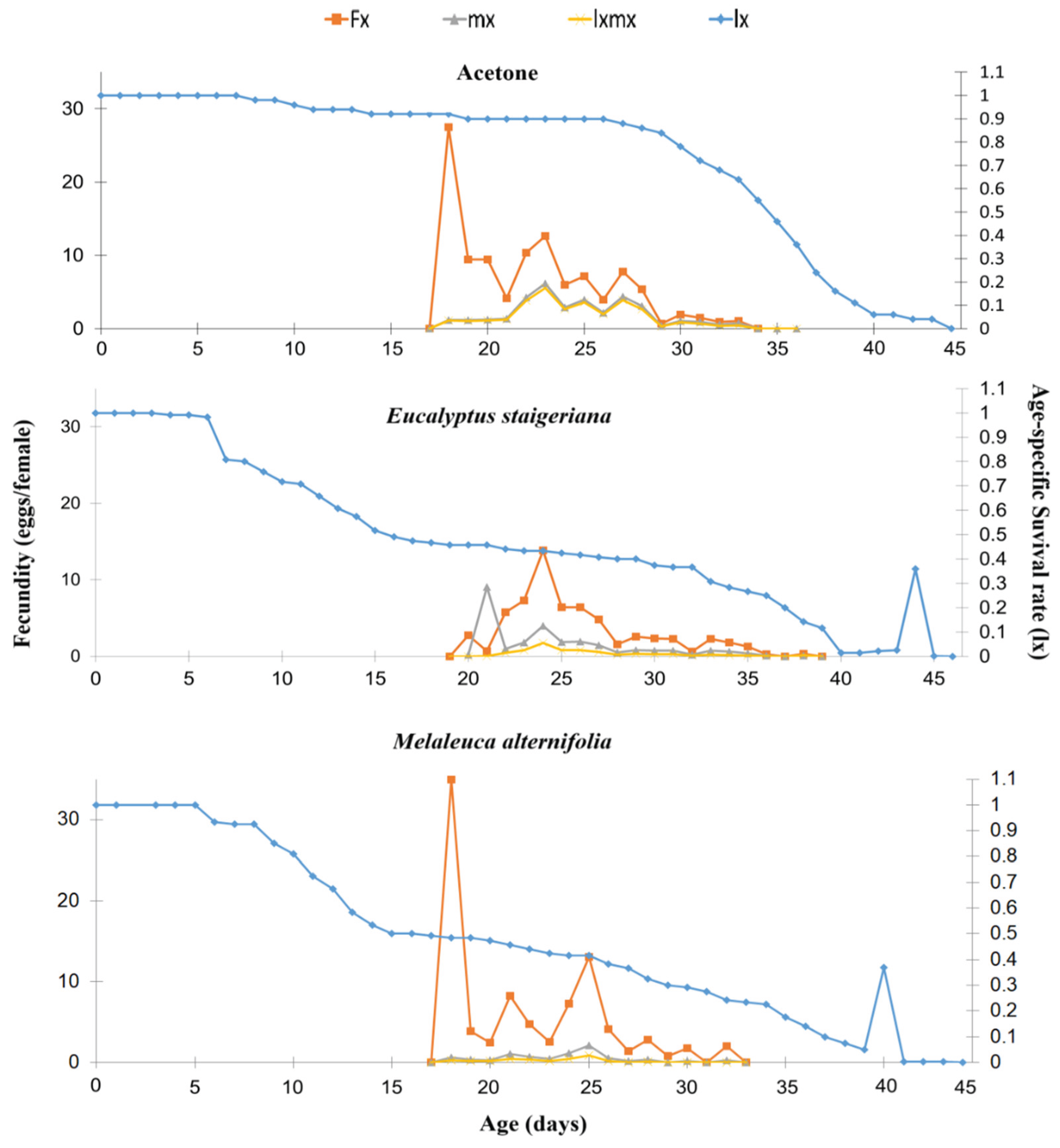

Table 1.
Chemical composition of the essential oils of Eucalyptus staigeriana and Melaleuca alternifolia.
Table 1.
Chemical composition of the essential oils of Eucalyptus staigeriana and Melaleuca alternifolia.
| N | Compound | RI* | RIL | Area (≥0.1%±SD | |
|---|---|---|---|---|---|
| E. staigeriana | M. alternifolia | ||||
| 1 | α-thujene | 910 | 924 | 0.279 ± 0.003 | 0.710 ± 0.002 |
| 2 | α-pinene | 913 | 932 | 2.981 ± 0.050 | 3.074 ± 0.013 |
| 3 | α-phelandrene | 937 | 1002 | Nd | 0.635 ± 0.002 |
| 4 | α-terpinene | 950 | 1014 | 0.179 ± 0.023 | 10.44 ± 0.091 |
| 5 | sylvestre | 957 | 1025 | Nd | 2.109 ± 0.005 |
| 6 | β-pinene | 974 | 974 | 1.454 ± 0.022 | 0.618 ± 0.002 |
| 7 | γ-terpinene | 975 | 1054 | Nd | 22.102 ± 0.129 |
| 8 | NI | 990 | - | 1.085 ± 0.003 | Nd |
| 9 | α-phellandrene | 1004 | 1002 | 2.257 ± 0.025 | Nd |
| 10 | o-cymene | 1022 | 1022 | 2.150 ± 0.001 | 3.378 ± 0.157 |
| 11 | limonene | 1027 | 1024 | 27.298 ± 0.330 | Nd |
| 12 | 1.8-cineol | 1030 | 1026 | 4.133 ± 0.055 | 2.241 ± 0.005 |
| 13 | (Z)-β-ocimene | 1035 | 1032 | 0.227 ± 0.001 | Nd |
| 14 | (E)-β-ocimene | 1045 | 1044 | 0.435 ± 0.002 | Nd |
| 15 | γ-terpinene | 1055 | 1054 | 1.882 ± 0.016 | Nd |
| 16 | terpinolene | 1087 | 1086 | 8.919 ± 0.076 | 3.353 ± 0.010 |
| 17 | linalool | 1100 | 1095 | 1.562 ± 0.009 | Nd |
| 18 | NI | 1170 | - | 0.847 ± 0.315 | Nd |
| 19 | terpinen-4-ol | 1176 | 1174 | 0.887 ± 0.316 | 42.235 ± 0.100 |
| 20 | α-terpineol | 1190 | 1186 | 1.089 ± 0.016 | 3.386 ± 0.139 |
| 21 | nerol | 1228 | 1227 | 2.063 ± 0.019 | Nd |
| 22 | neral | 1241 | 1235 | 9.905 ± 0.024 | Nd |
| 23 | geraniol | 1255 | 1249 | 6.386 ± 0.048 | Nd |
| 24 | geranial | 1272 | 1264 | 13.825 ± 0.079 | Nd |
| 25 | methyl geraniate | 1324 | 1322 | 3.787 ± 0.021 | Nd |
| 26 | neryl acetate | 1366 | 1359 | 1.166 ± 0.012 | Nd |
| 27 | geranyl acetate | 1385 | 1379 | 3.246 ± 0.046 | Nd |
| 28 | α-gurjunene | 1406 | 1409 | Nd | 0.293 ± 0.001 |
| 29 | E-caryophyllene | 1415 | 1417 | 0.146 ± 0.001 | 0.224 ± 0.001 |
| 30 | aromadendrene | 1435 | 1439 | Nd | 1.238 ± 0.005 |
| 31 | allo -aromadendrene | 1456 | 1458 | Nd | 0.364 ± 0.001 |
| 32 | trans-cadina-1(6),4-dieno | 1470 | 1475 | Nd | 0.169 ± 0.001 |
| 33 | viridiflorene | 1492 | 1496 | Nd | 1.277 ± 0.004 |
| 34 | δ-cadinene | 1521 | 1522 | Nd | 0.900 ± 0.004 |
| Total | 98.188 | 99.220 | |||
*Retention index relative to the n-alkane series (C8-C20) in the HP-5 MS column in elution order; 1 Retention rate according to the literature [49]; 2Area (≥ 0.1%): mean of the relative area of the chromatographic peaks above de 0.1%. SD: standard deviation (n = 3); Nd: not detected or peaks with a relative area of less than 1%; NI: not identified; N = number of compounds.
Table 2.
Dose-resposta of Melaleuca alternifolia and Eucalyptus staigeriana for Phthorimaea absoluta.
Table 2.
Dose-resposta of Melaleuca alternifolia and Eucalyptus staigeriana for Phthorimaea absoluta.
| Treatment | n | X2 | P | *b | *e | DL50 (μg.μL-1) (LS –LI) | DL90 (μg.μL-1) (LS –LI) |
|---|---|---|---|---|---|---|---|
| E. staigeriana | 60 | 186 | 0.56 | -4.259 | 78.514 | 78.5 (72.0 – 85.0) | 131.5 (111.2– 151.8) |
| M. alternifolia | 60 | 285.9 | 0.68 | -3.947 | 76.571 | 76.5 (70.3 – 82.8) | 133.6 (110.0– 157.3) |
*Doses in μg.μL-1. SL= upper limit; LI – lower limit. b is proportional to the slope at the LD50 χ2 value, and the p values correspond to the fit test ** “b” = coefficients of the equation f(x)=1/1+exp(b(log(x)-log(e)))). n = number of specimens at each developmental stage.
Table 3.
Mean (± SE) of the development time of the life stages and longevity (days) of Phthorimaea absoluta subjected to treatments with sublethal doses of essential oils of Melaleuca alternifolia (76.5 µg of essential oil.caterpillar-1) and Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1).
Table 3.
Mean (± SE) of the development time of the life stages and longevity (days) of Phthorimaea absoluta subjected to treatments with sublethal doses of essential oils of Melaleuca alternifolia (76.5 µg of essential oil.caterpillar-1) and Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1).
| Parameter | Stage | Acetone | Melaleuca alternifolia | Eucalyptus staigeriana | |||
|---|---|---|---|---|---|---|---|
| N | Mean ± SE | N | Mean ± SE | N | Mean ± SE | ||
| Development time (days) Longevity (days) |
Egg | 100 | 3.96 ± 0.02 a | 120 | 3.00±0.00a | 100 | 3.00±0.00a |
| L1 | 100 | 2.06 ± 0.03 a | 120 | 2.00±0.00a | 100 | 2.00±0.00a | |
| L2 | 96 | 3.83 ± 0.09 a | 92 | 6.86±0.71b | 51 | 7.39±0.38b | |
| L3 | 96 | 2.16 ± 0.04 a | 84 | 8.41±0.59b | 48 | 4.33±0.23c | |
| L4 | 92 | 2.36 ± 0.07 a | 55 | 3.74±0.27ab | 48 | 4.12±0.24b | |
| Pupa | 90 | 8.00 ± 0.12 b | 49 | 5.63±0.23a | 46 | 5.63±0.15a | |
| Egg - Pupa | 90 | 22.44 ± 0.18 a | 49 | 12.14±0.47a | 35 | 11.83±0.37ab | |
| Adult | 90 | 34.02 ± 0.79 a | 49 | 39.00±0.00a | 35 | 37.63±0.58b | |
| Life cycle (days)* |
Female | 50 | 37.36 ± 0.47 a | 9 | 32.22 ± 1.99 b | 16 | 37.73 ± 1.03 a |
| Male | 40 | 35.00 ± 0.66 b | 40 | 34.85 ± 0.82 b | 23 | 38.48 ± 0.93 a | |
| Egg - Adult | 90 | 36.31 ± 0.41 b | 49 | 34.37 ± 0.78 c | 39 | 38.2 ± 0.69 a | |
The means on the same line followed by different letters differed from each other (p <0.05). Differences between treatments were obtained using the paired bootstrap test with 100,000 replicates. N = number of specimens at each developmental stage. * Mean total life cycle for males and females (days) only for insects that became adults; L1 = 1st instar caterpillar, L2 = 2nd instar caterpillar, L3 = 3rd instar and L4 = 4th instar caterpillar.
Table 4.
Reproductive parameters of Phthorimaea absoluta adults derived from second-instar caterpillars that survived exposure to Melaleuca alternifolia (76.5 µg of essential oil.caterpillar-1) and Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1).
Table 4.
Reproductive parameters of Phthorimaea absoluta adults derived from second-instar caterpillars that survived exposure to Melaleuca alternifolia (76.5 µg of essential oil.caterpillar-1) and Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1).
| Parameter | Acetone | Melaleuca alternifolia | Eucalyptus staigeriana | |||
|---|---|---|---|---|---|---|
| N | Mean ± SE | N | Mean ± SE | N | Mean ± SE | |
| Total fecundity (O/F) | 50 | 62.76 ± 4.28 a | 9 | 46.78 ± 9.97 a | 15 | 56.13 ± 6.23 a |
| Fertility (O/F)* | 44 | 71.32 ± 3.08 a | 8 | 60.14 ± 6.03 a | 14 | 60.14 ± 5.12 a |
| Oviposition (days) | 44 | 4.57 ± 0.26 b | 8 | 4.00 ± 0.22 b | 14 | 5.50 ± 0.36 a |
| PPOA (days) | 44 | 1.73 ± 0.17 a | 8 | 2.29± 0.64 a | 14 | 1.71 ± 0.22 a |
| PPOT (days) | 44 | 24.02 ± 0.39 a | 8 | 22.86 ± 0.96 a | 14 | 23.29 ± 0.42 a |
| TFM (O/F) | - | 107 | - | 74 | - | 93 |
| DFM (O/F) | - | 70 | - | 50 | - | 39 |
Legend: Maximum daily fertility (DMF), maximum total fertility (FMT), adult preoviposition period (PPOA), (O/F) = eggs per female, N= number of specimens for each parameter, and total preoviposition period (PPOT). * Total number of females that laid eggs. The means in the same row followed by different letters are significantly different at p <0.05. Differences between treatments were obtained using the paired bootstrap test with 100.000 replicates.
Table 5.
Demographic parameters of Phthorimaea absoluta treated with the EO of Melaleuca alternifolia (76.5 µg of essential oil.caterpillar-1) and Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1).
Table 5.
Demographic parameters of Phthorimaea absoluta treated with the EO of Melaleuca alternifolia (76.5 µg of essential oil.caterpillar-1) and Eucalyptus staigeriana (78.5 µg of essential oil.caterpillar-1).
| Demographic parameter | Acetone | M. alternifolia | E. staigeriana |
|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | |
| Intrinsic growth rate (r) | 0.13 ± 0.005 a | 0.05 ± 0.017 b | 0.07 ± 0.010 b |
| Finite growth rate (λ) | 1.14 ± 0.006 a | 1.05 ± 0.018 b | 1.08 ± 0.011 b |
| Net reproductive rate (R0) | 31.38 ± 3.800 a | 3.51 ± 1.324 b | 7.02 ± 1.847 b |
| Average generation time (T) | 25.81 ± 0.422a | 25.03 ± 1.011 a | 26.66 ± 0.425 a |
| Gross reproductive rate (GRR) | 35.80 ± 4.073 a | 8.40 ± 3.056 b | 17.42 ± 4.233 ab |
| Intrinsic growth rate (r) | 0.13 ± 0.005 a | 0.05 ± 0.017 b | 0.07 ± 0.010 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



