
Submitted:
02 July 2025
Posted:
02 July 2025
You are already at the latest version
Abstract
Natural products have contributed significantly to anti-inflammatory drug development, but the therapeutic potential of fish visceral extracts has yet to be fully investigated. In this study, we investigated the anti-inflammatory properties of a crude spleen extract from rainbow trout (Oncorhynchus mykiss). The extract significantly reduced nitric oxide (NO) production and downregulated iNOS and COX-2 expression at both mRNA and protein levels. Among four fractions derived from the extract, the OSB fraction showed the strongest and most consistent activity. OSB suppressed LPS-induced expression of iNOS, COX-2, and pro-inflammatory cytokines, while enhancing anti-inflammatory cytokine expression. Mechanistic studies revealed that OSB inhibited NF-κB activation by blocking nuclear translocation of the p65 subunit. Additionally, OSB reduced M1 macrophage markers and LPS-induced ROS generation, indicating inhibition of M1 polarization. These findings suggest that OSB exerts anti-inflammatory effects by modulating the NF-κB pathway and highlight its potential as a natural therapeutic agent for inflammatory diseases.
Keywords:
Rainbow trout (Oncorhynchus mykiss)
; spleen crude extract
; fractionation
; inflammation
; NF-κB signaling pathway
1. Introduction
Inflammation is a crucial component of the innate immune response, triggered by microbial infections, tissue injury, or antigenic invasion. It is typically characterized by symptoms such as pain, swelling, itching, and fever, which accompany the elimination of pathogens and damaged cells from affected tissues [1]. Although the inflammatory response is generally self-limiting, its dysregulation or persistence can lead to chronic inflammatory conditions and contribute to the development and progression of various diseases, including cancer, diabetes, inflammatory bowel disease, asthma, and other autoimmune disorders [2]. Conventional anti-inflammatory agents, which primarily act by modulating inflammation-related cytokine expression, remain the standard treatment for these conditions [3]. However, their prolonged use is frequently associated with adverse effects such as gastrointestinal ulceration, mucosal perforation, and renal toxicity. These limitations underscore the urgent need for safer and more effective therapeutic alternatives with a reduced risk of drug resistance [4,5].
Macrophages, as key effectors of the innate immune system, play a central role in orchestrating inflammatory responses via activation of the Toll-like receptor 4 (TLR4)/nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling pathway upon lipopolysaccharide (LPS) stimulation [6,7]. Dysregulation of this pathway is implicated in numerous inflammatory diseases, including rheumatoid arthritis, pneumonia, and inflammatory bowel disease, making it a critical therapeutic target [8,9,10,11].
In recent years, natural product-derived compounds have gained significant attention for their ability to modulate inflammation-related signaling pathways [12,13]. The development of anti-inflammatory agents from natural sources is considered a promising alternative to synthetic drugs, owing to their favorable safety profiles and cost-effectiveness [14]. Notably, approximately 25% of anti-inflammatory drugs approved by the U.S. Food and Drug Administration (FDA) are derived from natural products, highlighting their therapeutic potential [15]. Fish processing by-products, such as heads, viscera, and bones—often comprising up to 50% of the total body mass depending on the species—are typically discarded through landfilling or incineration, contributing to environmental pollution including greenhouse gas emissions, water contamination, and ecosystem disruption [16,17,18]. Repurposing these by-products offers not only a sustainable waste management strategy but also a valuable opportunity to discover novel bioactive compounds from marine sources [19].
Previous studies have reported anti-inflammatory effects from various fish by-products. For example, cutlassfish head peptone was shown to modulate the mitogen-activated protein kinase (MAPK) signaling pathway in macrophages [20], and extracts from olive flounder heads suppressed iNOS and COX-2 expression [21]. Similarly, salmon bone extracts inhibited nitric oxide production, suggesting anti-inflammatory potential [22]. Despite these findings, the anti-inflammatory properties of fish visceral organ extracts remain poorly characterized and underexplored.
Among fish species, rainbow trout (Oncorhynchus mykiss) has gained attention for its high content of polydeoxyribonucleotide (PDRN), a DNA-derived compound with molecular weights ranging from 50 kDa to 1500 kDa [23]. PDRN has been shown to attenuate LPS-induced acute lung injury in rats by inhibiting both NF-κB and MAPK pathways [24], and to promote wound healing through the upregulation of vascular endothelial growth factor (VEGF) in diabetic mouse models [25]. Additionally, PDRN enhances osteoblast proliferation [26], increases collagen synthesis in incisional wound models [27], and exerts anti-inflammatory effects by modulating cytokine expression in osteoarthritic cell systems [28]. These findings collectively highlight the therapeutic potential of PDRN in treating inflammatory conditions [29]. However, commercial PDRN production typically relies on sperm- or milt-derived extraction from rainbow trout, which is limited by high production costs and the seasonal availability of spermatozoa [30].
In this study, we aimed to elucidate the molecular mechanisms underlying the anti-inflammatory effects of visceral extracts derived from rainbow trout using an LPS-stimulated macrophage model. Our findings provide novel insights into the bioactivity of fish by-product-derived visceral organ extracts and their potential as sustainable sources of anti-inflammatory therapeutics.
2. Materials and Methods
2.1. Preparation of Crude Extract and Fractionation Samples
Thirteen-month-old female rainbow trout were obtained from Woori Trout Farm (Chuncheon, Gangwon-do, Republic of Korea). Following dissection, the liver, gallbladder, spleen, and kidney were harvested and extracted with 70% ethanol at 60°C for 16 hours. The resulting crude extracts were lyophilized, reconstituted in 70% ethanol, and subsequently diluted in culture medium for use in in vitro experiments.
2.2. Cell Culture
RAW264.7 murine macrophage cells were cultured at 37°C in a humidified atmosphere containing 5% CO₂. The cells were maintained in DMEM supplemented with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin (P/S). Cells were gently detached using a cell scraper without trypsinization. Freeze-dried extracts from rainbow trout liver, gallbladder, spleen, and kidney were dissolved in 70% ethanol and diluted in the culture medium to the desired concentrations. Lipopolysaccharide (LPS) was dissolved in triple-distilled water and diluted to a final concentration of 1 μg/mL in DMEM.
2.3. Nitric Oxide Assay
RAW264.7 cells were seeded in 96-well plates and incubated for 24 hours. The cells were then treated with crude extracts from the liver, gallbladder, spleen, or kidney of rainbow trout in combination with 1 μg/mL LPS. After 24 hours, the culture supernatant was collected and transferred to a new 96-well plate for nitric oxide measurement using the NO Plus Detection Kit (iNtRON Biotechnology, Seongnam, Korea) according to the manufacturer’s protocol.
2.4. Quantitative Reverse Transcription PCR (qRT-PCR)
RAW264.7 cells were seeded in culture dishes and incubated for 24 hours, followed by treatment with OSH, OSE, OSB, or OSW in the presence of 1 μg/mL LPS for an additional 24 hours. Total RNA was isolated using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), and cDNA synthesis was performed using oligodT primers and M-MLV reverse transcriptase (Promega, Madison, WI, USA). qRT-PCR was conducted using the TOPreal™ qPCR 2X PreMIX (Enzynomics, Daejeon, Republic of Korea). Relative gene expression levels were calculated using the 2^−ΔΔCt method, with normalization to GAPDH. Primer sequences are listed below:
| Gene | The Sequences of the Primers |
| iNOS | F-CCAAGCCCTCACCTACTTCC |
| R-CTCTGAGGGCTGACACAAGG | |
| COX-2 | F-CATCCCCTTCCTGCGAAGTT |
| R-CATGGGAGTTGGGCAGTCAT | |
| IL-6 | F-AGTCCTTCCTACCCCAATTTCC |
| R-TAACGCACTAGGTTTGCCGA | |
| TNF-α | F-ACCGTCAGCCGATTTGCTAT |
| R-TTGGGCAGATTGACCTCAGC | |
| IL-12p40 | F-AGACCCTGCCCATTGAACTG |
| R-CAGGAGTCAGGGTACTCCCA | |
| IL-23p19 | F-CAGCAGCTCTCTCGGAATCT |
| R-CAGACCTTGGCGGATCCTTT | |
| IL-13 | F-GTATGGAGTGTGGACCTGGC |
| R-ATTTTGGTATCGGGGAGGCTG | |
| TGF-β | F-CTGCTGACCCCCACTGATAC |
| R-GGGGCTGATCCCGTTGATTT | |
| CD40 | F-GCTATGGGGCTGCTTGTTGA |
| R-GGTGGCATTGGGTCTTCTCA | |
| CD86 | F-ATGGACCCCAGATGCACCA |
| R-TGTGCCCAAATAGTGCTCGT | |
| GAPDH | F-CTCATGACCACAGTCCATGC |
| R-CACATTGGGGGTAGGAACAC |
2.5. Western Blotting
RAW264.7 cells were seeded into 100 mm dishes and treated 24 hours later according to experimental conditions. After treatment, cells were harvested using a scraper, and total protein was extracted using RIPA lysis buffer (10 mM Tris-HCl pH 8.0, 1 mM EDTA, 140 mM NaCl, 1% Triton X-100, 0.1% sodium deoxycholate, 0.1% SDS) containing cOmplete™ Protease Inhibitor Cocktail (Roche, Basel, Switzerland). Protein concentration was measured using the Bradford assay (Thermo Fisher Scientific, Waltham, MA, USA). Proteins were separated by SDS-PAGE and transferred to 0.45 μm PVDF membranes. Membranes were blocked with 5% skim milk for 30 minutes at room temperature, then incubated overnight at 4°C with primary antibodies diluted in 1× TBST containing 1% BSA. After washing, membranes were incubated with HRP-conjugated secondary antibodies diluted in 5% skim milk for 2 hours at room temperature. Protein bands were visualized using an ECL detection kit (GE Healthcare, Chicago, IL, USA). iNOS (13120S), COX-2 (12282S), IκBα (9242S), p-p65 (3033S), and CD86 (19589S) antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA), and p65 (ab32536) and Lamin B1 (ab16048) antibodies were obtained from Abcam (Cambridge, UK).
2.6. Immunofluorescence
RAW264.7 cells were seeded onto coverslips in 6-well plates and treated 24 hours after seeding. Cells were fixed with 4% paraformaldehyde for 10 minutes, permeabilized with 0.1% Triton X-100 in PBS for 3 minutes, and blocked with 5% skim milk in PBS for 1 hour at room temperature. Cells were incubated with primary antibodies (1:200 dilution) for 2 hours, followed by Alexa Fluor 488-conjugated goat anti-mouse IgG (1:200; Abcam, Boston, MA, USA) for 1 hour. Nuclei were stained with DAPI (1:2000 in PBS). Fluorescence images were acquired using a confocal microscope (Nikon, Tokyo, Japan).
2.7. Reactive Oxygen Species (ROS) Analysis
RAW264.7 cells were seeded in 60 mm dishes and treated according to experimental protocols 24 hours after seeding. Cells were incubated with 10 μM carboxy-H₂DCFDA (Invitrogen, USA) at 37°C for 30 minutes. After washing twice with PBS, intracellular ROS levels were measured using a flow cytometer (FACSymphony, Bergen County, NJ, USA) with excitation at 488 nm and emission at 525 nm.
2.8. Statistical Analysis
Statistical analyses were performed using GraphPad Prism 8.0 and ImageJ v1.54d. Data are presented as the mean ± standard deviation (SD) from at least three independent experiments. Comparisons between two groups were evaluated using Student’s t-test. A p-value < 0.05 was considered statistically significant, with the following notation: p ≤ 0.05 (*), p ≤ 0.01 (**).
3. Results
3.1. Anti-Inflammatory Effect of Rainbow Trout Spleen Crude Extract in LPS-Stimulated RAW264.7 Cells
To investigate the bioactivity of organ-derived extracts from rainbow trout, crude extracts were prepared from the liver, gallbladder, spleen, and kidney using 70% ethanol (Figure 1A). The anti-inflammatory potential of each extract was evaluated by treating RAW264.7 macrophages with LPS in the presence of each crude extract. NO production, a key marker of inflammation, was measured to assess the inflammatory response. LPS stimulation induced a nearly 10-fold increase in NO production compared to untreated controls. Importantly, none of the crude extracts alone triggered an inflammatory response. Among the organ extracts tested, only the spleen crude extract markedly reduced LPS-induced NO production in a concentration-dependent manner, whereas the liver, gallbladder, and kidney extracts had no significant effect (Figure 1B). To further examine the anti-inflammatory activity of the spleen extract, we analyzed the expression levels of inflammation-related genes. Quantitative real-time PCR demonstrated that spleen extract significantly downregulated the LPS-induced expression of iNOS and COX-2 mRNA in a dose-dependent manner (Figure 1C). Consistent with the mRNA findings, Western blot analysis revealed that spleen extract treatment also reduced the protein expression levels of iNOS and COX-2 induced by LPS (Figure 1D). Collectively, these results indicate that the spleen crude extract of rainbow trout exerts potent anti-inflammatory effects by suppressing NO production and downregulating the expression of key pro-inflammatory mediators in LPS-stimulated RAW264.7 macrophages.
3.2. Fractionation of Rainbow Trout Spleen Crude Extract and Anti-Inflammatory Effects of Each Fraction
To identify the bioactive anti-inflammatory components within the spleen crude extract, Solvent partitioning based on polarity was employed to obtain four fractions: OSH (n-hexane layer), OSE (ethyl acetate layer), OSB (n-butanol layer), and OSW (aqueous layer) (Figure 2A). The chemical composition of each fraction was analyzed by ultra-performance liquid chromatography-mass spectrometry (UPLC-MS), revealing distinct profiles among the fractions (Figure 2B). To evaluate and compare the anti-inflammatory activity of each fraction, RAW264.7 cells were treated with 1 μg/mL LPS in the presence of either the crude spleen extract or individual fractions (OSH, OSE, OSB, or OSW) at a concentration of 200 μg/mL for 24 hours. Quantitative PCR analysis was performed to assess the mRNA expression of iNOS, COX-2, and the pro-inflammatory cytokines IL-6 and TNF-α. LPS stimulation significantly increased the expression of all four genes. Co-treatment with the spleen crude extract, OSH, OSE, or OSB attenuated this upregulation to varying degrees. Among the tested fractions, OSH, OSE, and OSB exhibited stronger inhibitory effects on inflammatory gene expression than the unfractionated crude extract. However, microscopic examination revealed that OSH and OSE induced cytotoxic effects, as indicated by the presence of floating and detached cells. In contrast, OSB treatment significantly suppressed LPS-induced expression of iNOS, COX-2, IL-6, and TNF-α without causing observable cytotoxicity (Figure 2C,D). These results suggest that the principal anti-inflammatory activity of the spleen crude extract is enriched in the OSB fraction, which exerts potent anti-inflammatory effects while maintaining cell viability.
3.3. Regulatory Effect of OSB on iNOS and COX-2 Expression
Among the spleen-derived fractions, OSB exhibited the most potent and consistent anti-inflammatory activity, suggesting that the key bioactive constituents are predominantly enriched in this fraction. To further elucidate its mechanism of action, we examined the effects of OSB on iNOS and COX-2 protein expression in LPS-stimulated RAW264.7 cells. Western blot analysis revealed that OSB treatment markedly suppressed the LPS-induced upregulation of iNOS and COX-2 protein levels in a dose-dependent manner (Figure 3A). In addition, a time-course experiment demonstrated that OSB effectively inhibited the time-dependent induction of both proteins following LPS stimulation (Figure 3B). Consistent with these results, immunofluorescence analysis showed a marked decrease in cytoplasmic fluorescence signals for iNOS and COX-2 following OSB and LPS co-treatment. In contrast, cells treated with LPS alone exhibited strong fluorescence intensity, indicating robust protein expression (Figure 3C, D). Therefore, these results demonstrate that OSB effectively attenuates LPS-induced inflammatory responses by suppressing the expression of key pro-inflammatory enzymes, iNOS and COX-2, in RAW264.7 macrophages.
3.4. Regulatory Effect of OSB on the NF-κB Signaling Pathway
To investigate the mechanism by which OSB regulates inflammatory responses, we examined its effects on the NF-κB signaling pathway, a key upstream regulator of iNOS and COX-2 expression that is activated by LPS stimulation. In RAW264.7 macrophages, OSB treatment inhibited LPS-induced phosphorylation of IκBα, thereby preventing its degradation. This inhibition subsequently suppressed the phosphorylation of the NF-κB subunit p65, which is dependent on IκBα activation (Figure 4A). Western blot analysis further demonstrated that OSB-mediated suppression of p65 phosphorylation resulted in reduced nuclear translocation of p65 following LPS stimulation (Figure 4B). Supporting these findings, immunofluorescence analysis showed that p65 was translocated to the nucleus upon LPS stimulation, while OSB co-treatment blocked this translocation (Figure 4C). These results indicate that OSB attenuates LPS-induced inflammatory signaling by modulating the NF-κB pathway, thereby inhibiting the activation and nuclear translocation of p65.
3.5. Regulatory Effect of OSB on M1 Macrophage Polarization
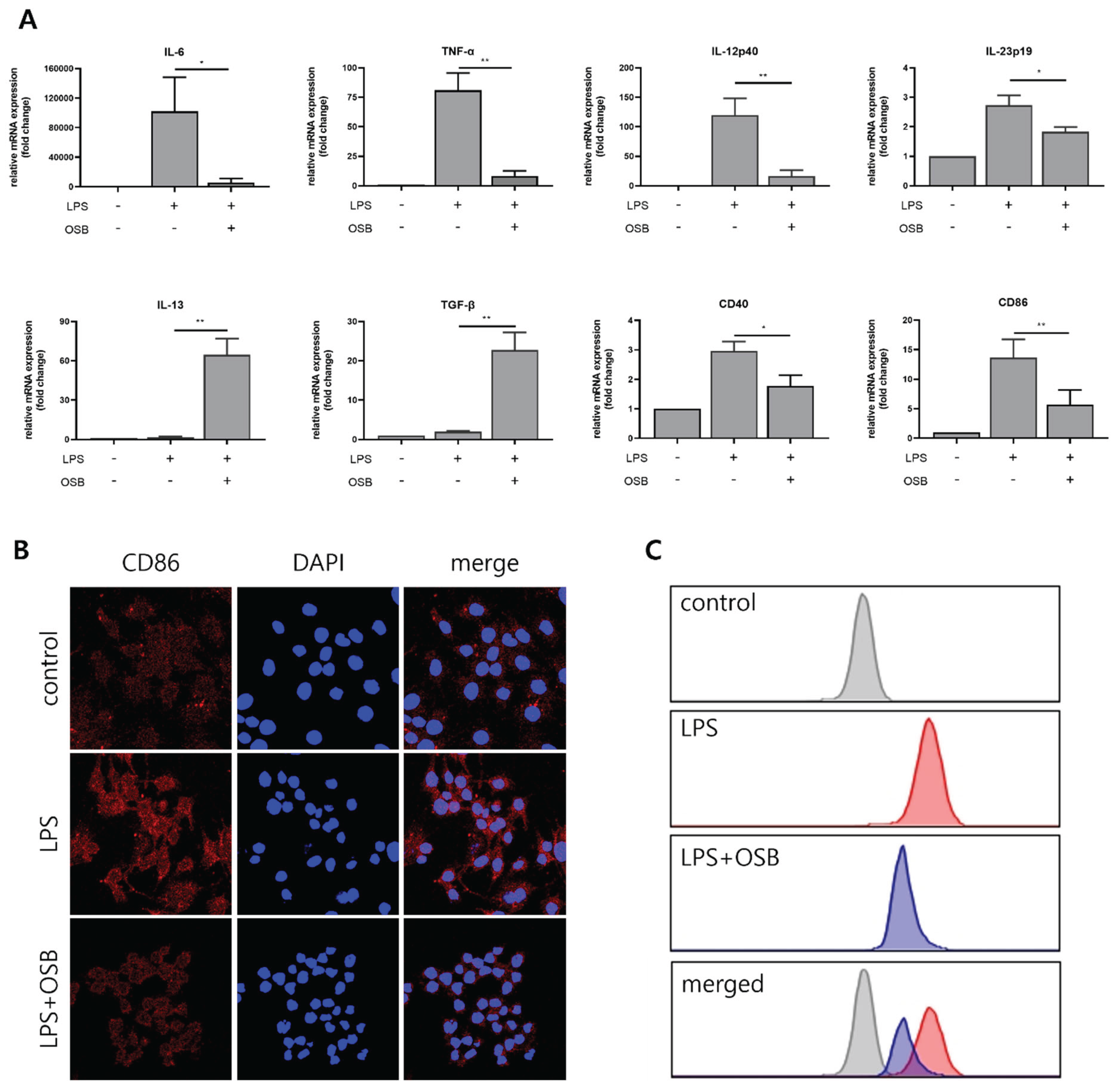
The effects of OSB on macrophage polarization were assessed by analyzing mRNA expression levels of pro-inflammatory cytokines (IL-6, TNF-α, IL-23p19, and IL-12p40) and anti-inflammatory cytokines (IL-13 and TGF-β) using quantitative PCR. OSB treatment significantly suppressed LPS-induced upregulation of the pro-inflammatory cytokines, while simultaneously enhancing the expression of the anti-inflammatory cytokines IL-13 and TGF-β. Furthermore, the expression of M1 macrophage surface markers CD40 and CD86 was evaluated. LPS-induced increases in CD40 and CD86 mRNA levels were notably attenuated by OSB treatment (Figure 5A). Protein-level changes in CD86 expression were confirmed by immunofluorescence staining, which demonstrated marked upregulation of CD86 on RAW264.7 cell surfaces following LPS stimulation; this increase was effectively inhibited by OSB (Figure 5B). Additionally, intracellular reactive oxygen species (ROS) levels were measured using the fluorescent probe H2DCFDA. OSB treatment dose-dependently reduced LPS-induced ROS production in RAW264.7 cells (Figure 5C). Collectively, these results indicate that OSB effectively suppresses LPS-induced M1 macrophage polarization by modulating cytokine expression, surface marker levels, and oxidative stress.
4. Discussion
The development of therapeutics derived from natural products for inflammatory diseases has garnered increasing attention in recent years [31,32,33]. A critical aspect of anti-inflammatory drug development involves elucidating the regulatory mechanisms that govern inflammation-associated signaling pathways [34]. Among these, the TLR4/NF-κB pathway has been identified as a central target in the treatment of inflammatory disorders [35]. While several studies have explored the anti-inflammatory potential of fish by-product extracts, including those from heads, bones, and fins, the biological activity of viscera-derived extracts remains largely underexplored [20,21,22]. Notably, extracts from the viscera of marine organisms such as Turbo cornutus and abalone have demonstrated anti-inflammatory activity by suppressing MAPK signaling components, including JNK and p38 phosphorylation [34,36].
Rainbow trout viscera have traditionally been considered industrial waste; however, recent efforts have focused on repurposing these by-products as valuable sources of bioactive compounds [37]. In this study, we investigated the anti-inflammatory effects of crude extracts from the liver, gallbladder, spleen, and kidney of rainbow trout, with a focus on identifying active constituents with therapeutic potential. Initial evaluation was conducted via NO assays, as macrophages produce NO as a hallmark of LPS-induced inflammatory responses. Among the organ extracts tested, the spleen extract exhibited a dose-dependent inhibition of NO production. Furthermore, this extract significantly reduced both the mRNA and protein expression levels of iNOS as well as COX-2, another key inflammatory mediator. These results suggest that the spleen extract exerts a more pronounced anti-inflammatory effect relative to other tissue-derived extracts.
To isolate the bioactive constituents responsible for these effects, the spleen crude extract underwent solvent fractionation via liquid–liquid extraction based on differential polarity. This process yielded four fractions: OSH, OSE, OSB, and OSW. Each fraction, along with the crude extract, was evaluated for its ability to modulate the mRNA expression of inflammatory mediators (iNOS, COX-2, IL-6, and TNF-α). The OSH, OSE, and OSB fractions exhibited greater inhibitory activity than the crude extract. However, cytotoxic effects, including cell detachment and lysis, were observed following treatment with OSH and OSE. In contrast, the OSB fraction demonstrated strong and consistent anti-inflammatory activity without cytotoxicity, suggesting it contains enriched bioactive compounds from the rainbow trout spleen.
Subsequent analysis confirmed that OSB treatment suppressed LPS-induced expression of iNOS and COX-2 in a dose-dependent manner at the protein level. Time-course experiments revealed that although OSB suppressed these inflammatory proteins, their levels gradually increased over time, indicating partial rather than complete inhibition. These effects were validated through both Western blotting and immunofluorescence microscopy. Notably, OSB treatment reduced the cytoplasmic fluorescence intensity of iNOS and COX-2 induced by LPS, consistent with the biochemical results.
Given the pivotal role of NF-κB signaling in regulating the transcription of iNOS, COX-2, and other pro-inflammatory genes, we further investigated the upstream mechanisms modulated by OSB. LPS stimulation leads to IκBα phosphorylation and degradation, facilitating the nuclear translocation of the NF-κB p65 subunit. OSB treatment inhibited both IκBα phosphorylation/degradation and p65 activation, thereby preventing its translocation to the nucleus. Western blotting and immunofluorescence analyses confirmed this suppression, demonstrating a reduction in nuclear p65 localization upon OSB treatment. These findings indicate that OSB interferes with NF-κB activation and nuclear translocation, thereby attenuating transcriptional activation of pro-inflammatory genes and subsequent inflammatory responses.
Macrophage polarization is another critical determinant of inflammation. Upon LPS exposure, macrophages undergo M1 polarization, characterized by enhanced expression of surface markers (CD40, CD86), pro-inflammatory cytokines, and ROS production. Our results demonstrated that OSB significantly downregulated LPS-induced mRNA expression of IL-6, TNF-α, IL-12p40, and IL-23p19. In parallel, it upregulated anti-inflammatory cytokines such as IL-13 and TGF-β. OSB also suppressed the expression of M1 surface markers CD40 and CD86 at both the mRNA and protein levels. Immunofluorescence analysis further confirmed reduced CD86 surface expression. Moreover, OSB effectively diminished LPS-induced ROS generation, suggesting inhibition of oxidative stress—a hallmark of M1 polarization. These results collectively support the notion that OSB suppresses M1 polarization in LPS-stimulated RAW264.7 macrophages.
In summary, our study identifies the OSB fraction of rainbow trout spleen extract as a novel, potent, and non-cytotoxic anti-inflammatory agent. OSB mitigated LPS-induced expression of iNOS and COX-2 through modulation of the NF-κB signaling cascade and suppressed M1 macrophage polarization by regulating cytokine expression, surface markers, and intracellular ROS levels. This study provides mechanistic evidence supporting the anti-inflammatory potential of bioactive compounds derived from fish viscera and highlights the OSB fraction of rainbow trout spleen extract as a promising source for natural anti-inflammatory drug development.
References
- Wallach, D.; Kang, T.-B.; Kovalenko, A. Concepts of tissue injury and cell death in inflammation: a historical perspective. Nature Reviews Immunology 2014, 14, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Leuti, A.; Fazio, D.; Fava, M.; Piccoli, A.; Oddi, S.; Maccarrone, M. Bioactive lipids, inflammation and chronic diseases. Adv Drug Deliv Rev 2020, 159, 133–169. [Google Scholar] [CrossRef]
- Opal, S.M.; DePalo, V.A. Anti-Inflammatory Cytokines. Chest 2000, 117, 1162–1172. [Google Scholar] [CrossRef] [PubMed]
- James, D.S. The multisystem adverse effects of NSAID therapy. J Am Osteopath Assoc 1999, 99, S1–s7. [Google Scholar] [CrossRef]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem Pharmacol 2020, 180, 114147. [Google Scholar] [CrossRef]
- Rossol, M.; Heine, H.; Meusch, U.; Quandt, D.; Klein, C.; Sweet, M.J.; Hauschildt, S. LPS-induced cytokine production in human monocytes and macrophages. Crit Rev Immunol 2011, 31, 379–446. [Google Scholar] [CrossRef]
- Li, P.; Wu, Y.H.; Zhu, Y.T.; Li, M.X.; Pei, H.H. Requirement of Rab21 in LPS-induced TLR4 signaling and pro-inflammatory responses in macrophages and monocytes. Biochem Biophys Res Commun 2019, 508, 169–176. [Google Scholar] [CrossRef]
- Yin, J.; Huang, J.; Zhou, P.; Li, L.; Zheng, Q.; Fu, H. The role of TLR4/NF-kB signaling axis in pneumonia: from molecular mechanisms to regulation by phytochemicals. Naunyn Schmiedebergs Arch Pharmacol 2025. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Wang, D.; Yang, Z.; Wang, T. Pharmacological Effects of Polyphenol Phytochemicals on the Intestinal Inflammation via Targeting TLR4/NF-κB Signaling Pathway. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Dejban, P.; Nikravangolsefid, N.; Chamanara, M.; Dehpour, A.; Rashidian, A. The role of medicinal products in the treatment of inflammatory bowel diseases (IBD) through inhibition of TLR4/NF-kappaB pathway. Phytother Res 2021, 35, 835–845. [Google Scholar] [CrossRef]
- O'Neill, L.A. Primer: Toll-like receptor signaling pathways--what do rheumatologists need to know? Nat Clin Pract Rheumatol 2008, 4, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Winand, L.; Sester, A.; Nett, M. Bioengineering of Anti-Inflammatory Natural Products. ChemMedChem 2021, 16, 767–776. [Google Scholar] [CrossRef]
- Wang, R.-X.; Zhou, M.; Ma, H.-L.; Qiao, Y.-B.; Li, Q.-S. The Role of Chronic Inflammation in Various Diseases and Anti-inflammatory Therapies Containing Natural Products. ChemMedChem 2021, 16, 1576–1592. [Google Scholar] [CrossRef] [PubMed]
- Gautam, R.; Jachak, S.M. Recent developments in anti-inflammatory natural products. Med Res Rev 2009, 29, 767–820. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J Nat Prod 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Varatharajan, V.; Peng, H.; Senadheera, R. Utilization of marine by-products for the recovery of value-added products. Journal of Food Bioactives 2019, 6, 10–61. [Google Scholar] [CrossRef]
- Coppola, D.; Lauritano, C.; Palma Esposito, F.; Riccio, G.; Rizzo, C.; de Pascale, D. Fish Waste: From Problem to Valuable Resource. Marine Drugs 2021, 19. [Google Scholar] [CrossRef]
- Ketnawa, S.; Liceaga, A.M. Effect of microwave treatments on antioxidant activity and antigenicity of fish frame protein hydrolysates. Food and bioprocess technology 2017, 10, 582–591. [Google Scholar] [CrossRef]
- Halim, N.R.A.; Yusof, H.M.; Sarbon, N.M. Functional and bioactive properties of fish protein hydolysates and peptides: A comprehensive review. Trends in Food Science & Technology 2016, 51, 24–33. [Google Scholar] [CrossRef]
- Lee, S.-J.; Im, J.; Marasinghe, S.D.; Jo, E.; Bandara, M.S.; Lee, Y.; Lee, J.; Park, G.-H.; Oh, C. Antioxidant and Anti-Inflammatory Activities of Cutlassfish Head Peptone in RAW 264.7 Macrophages. Antioxidants 2025, 14. [Google Scholar] [CrossRef]
- Jayawardhana, H.H.A.C.K.; Liyanage, N.M.; Nagahawatta, D.P.; Lee, H.-G.; Jeon, Y.-J.; Kang, S.I. Pepsin Hydrolysate from Surimi Industry-Related Olive Flounder Head Byproducts Attenuates LPS-Induced Inflammation and Oxidative Stress in RAW 264.7 Macrophages and In Vivo Zebrafish Model. Marine Drugs 2024, 22. [Google Scholar] [CrossRef] [PubMed]
- Saisavoey, T.; Sangtanoo, P.; Reamtong, O.; Karnchanatat, A. Free radical scavenging and anti-inflammatory potential of a protein hydrolysate derived from salmon bones on RAW 264.7 macrophage cells. Journal of the Science of Food and Agriculture 2019, 99, 5112–5121. [Google Scholar] [CrossRef] [PubMed]
- Belmontesi, M. Polydeoxyribonucleotide for the improvement of a hypertrophic retracting scar—An interesting case report. Journal of Cosmetic Dermatology 2020, 19, 2982–2986. [Google Scholar] [CrossRef] [PubMed]
- Ko, I.-G.; Hwang, J.J.; Chang, B.S.; Kim, S.-H.; Jin, J.-J.; Hwang, L.; Kim, C.-J.; Choi, C.W. Polydeoxyribonucleotide ameliorates lipopolysaccharide-induced acute lung injury via modulation of the MAPK/NF-κB signaling pathway in rats. International Immunopharmacology 2020, 83, 106444. [Google Scholar] [CrossRef]
- Galeano, M.; Bitto, A.; Altavilla, D.; Minutoli, L.; Polito, F.; Calò, M.; Lo Cascio, P.; Stagno d'Alcontres, F.; Squadrito, F. Polydeoxyribonucleotide stimulates angiogenesis and wound healing in the genetically diabetic mouse. Wound Repair and Regeneration 2008, 16, 208–217. [Google Scholar] [CrossRef]
- Guizzardi, S.; Galli, C.; Govoni, P.; Boratto, R.; Cattarini, G.; Martini, D.; Belletti, S.; Scandroglio, R. Polydeoxyribonucleotide (PDRN) promotes human osteoblast proliferation: A new proposal for bone tissue repair. Life Sciences 2003, 73, 1973–1983. [Google Scholar] [CrossRef]
- Jeong, W.; Yang, C.E.; Roh, T.S.; Kim, J.H.; Lee, J.H.; Lee, W.J. Scar Prevention and Enhanced Wound Healing Induced by Polydeoxyribonucleotide in a Rat Incisional Wound-Healing Model. International Journal of Molecular Sciences 2017, 18. [Google Scholar] [CrossRef]
- Baek, A.; Kim, M.; Kim, S.H.; Cho, S.-R.; Kim, H.J. Anti-inflammatory Effect of DNA Polymeric Molecules in a Cell Model of Osteoarthritis. Inflammation 2018, 41, 677–688. [Google Scholar] [CrossRef]
- Kim, T.H.; Heo, S.Y.; Oh, G.W.; Heo, S.J.; Jung, W.K. Applications of Marine Organism-Derived Polydeoxyribonucleotide: Its Potential in Biomedical Engineering. Mar Drugs 2021, 19. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Kim, M.-J.; Kweon, D.-K.; Lim, S.-T.; Lee, S.-J. Polydeoxyribonucleotide Activates Mitochondrial Biogenesis but Reduces MMP-1 Activity and Melanin Biosynthesis in Cultured Skin Cells. Applied Biochemistry and Biotechnology 2020, 191, 540–554. [Google Scholar] [CrossRef]
- Fernandes, A.; Rodrigues, P.M.; Pintado, M.; Tavaria, F.K. A systematic review of natural products for skin applications: Targeting inflammation, wound healing, and photo-aging. Phytomedicine 2023, 115, 154824. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, W.; Chen, Y.; Wan, X.; Wang, J. Fucoxanthin: A promising compound for human inflammation-related diseases. Life Sci 2020, 255, 117850. [Google Scholar] [CrossRef] [PubMed]
- Almutary, A.G.; Begum, M.Y.; Kyada, A.K.; Gupta, S.; Jyothi, S.R.; Chaudhary, K.; Sharma, S.; Sinha, A.; Abomughaid, M.M.; Imran, M.; et al. Inflammatory signaling pathways in Alzheimer's disease: Mechanistic insights and possible therapeutic interventions. Ageing Res Rev 2025, 104, 102548. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.A.; Kang, N.; Kim, J.; Yang, H.W.; Ahn, G.; Heo, S.J. Anti-Inflammatory Effect of Turbo cornutus Viscera Ethanolic Extract against Lipopolysaccharide-Stimulated Inflammatory Response via the Regulation of the JNK/NF-kB Signaling Pathway in Murine Macrophage RAW 264.7 Cells and a Zebrafish Model: A Preliminary Study. Foods 2022, 11. [Google Scholar] [CrossRef]
- Cao, J.; Li, Q.; Shen, X.; Yao, Y.; Li, L.; Ma, H. Dehydroepiandrosterone attenuates LPS-induced inflammatory responses via activation of Nrf2 in RAW264.7 macrophages. Molecular Immunology 2021, 131, 97–111. [Google Scholar] [CrossRef]
- Qian, Z.-J.; Ryu, B.; Sun Park, W.; Choi, I.L.W.; Jung, W.-K. Inhibitory Effects and Molecular Mechanism of an Anti-inflammatory Peptide Isolated from Intestine of Abalone, Haliotis Discus Hannai on LPS-Induced Cytokine Production via the p-p38/p-JNK Pathways in RAW264.7 Macrophages. Journal of Food and Nutrition Research 2025, 4, 690–698. [Google Scholar]
- Nikoo, M.; Benjakul, S.; Yasemi, M.; Ahmadi Gavlighi, H.; Xu, X. Hydrolysates from rainbow trout (Oncorhynchus mykiss) processing by-product with different pretreatments: Antioxidant activity and their effect on lipid and protein oxidation of raw fish emulsion. LWT 2019, 108, 120–128. [Google Scholar] [CrossRef]
Figure 1.
Anti-inflammatory effects of rainbow trout spleen crude extract in LPS-stimulated RAW264.7 cells. (A) Crude extracts were prepared from the liver, gallbladder, spleen, and kidney of rainbow trout by 70% ethanol extraction at 60°C for 16 hours. (B) RAW264.7 cells were treated with each extract (10–200 μg/mL) in the presence of 1 μg/mL LPS, and nitric oxide (NO) production was quantified. (C) The mRNA expression levels of iNOS and COX-2 were assessed by qRT-PCR following treatment with spleen crude extract (100–400 μg/mL). (D) Protein expression of iNOS and COX-2 was analyzed by Western blotting under the same treatment conditions. All treatments were performed for 24 hours. Data are presented as mean ± SD (n = 3). Statistical analysis was conducted using one-way ANOVA followed by multiple comparisons.
Figure 1.
Anti-inflammatory effects of rainbow trout spleen crude extract in LPS-stimulated RAW264.7 cells. (A) Crude extracts were prepared from the liver, gallbladder, spleen, and kidney of rainbow trout by 70% ethanol extraction at 60°C for 16 hours. (B) RAW264.7 cells were treated with each extract (10–200 μg/mL) in the presence of 1 μg/mL LPS, and nitric oxide (NO) production was quantified. (C) The mRNA expression levels of iNOS and COX-2 were assessed by qRT-PCR following treatment with spleen crude extract (100–400 μg/mL). (D) Protein expression of iNOS and COX-2 was analyzed by Western blotting under the same treatment conditions. All treatments were performed for 24 hours. Data are presented as mean ± SD (n = 3). Statistical analysis was conducted using one-way ANOVA followed by multiple comparisons.
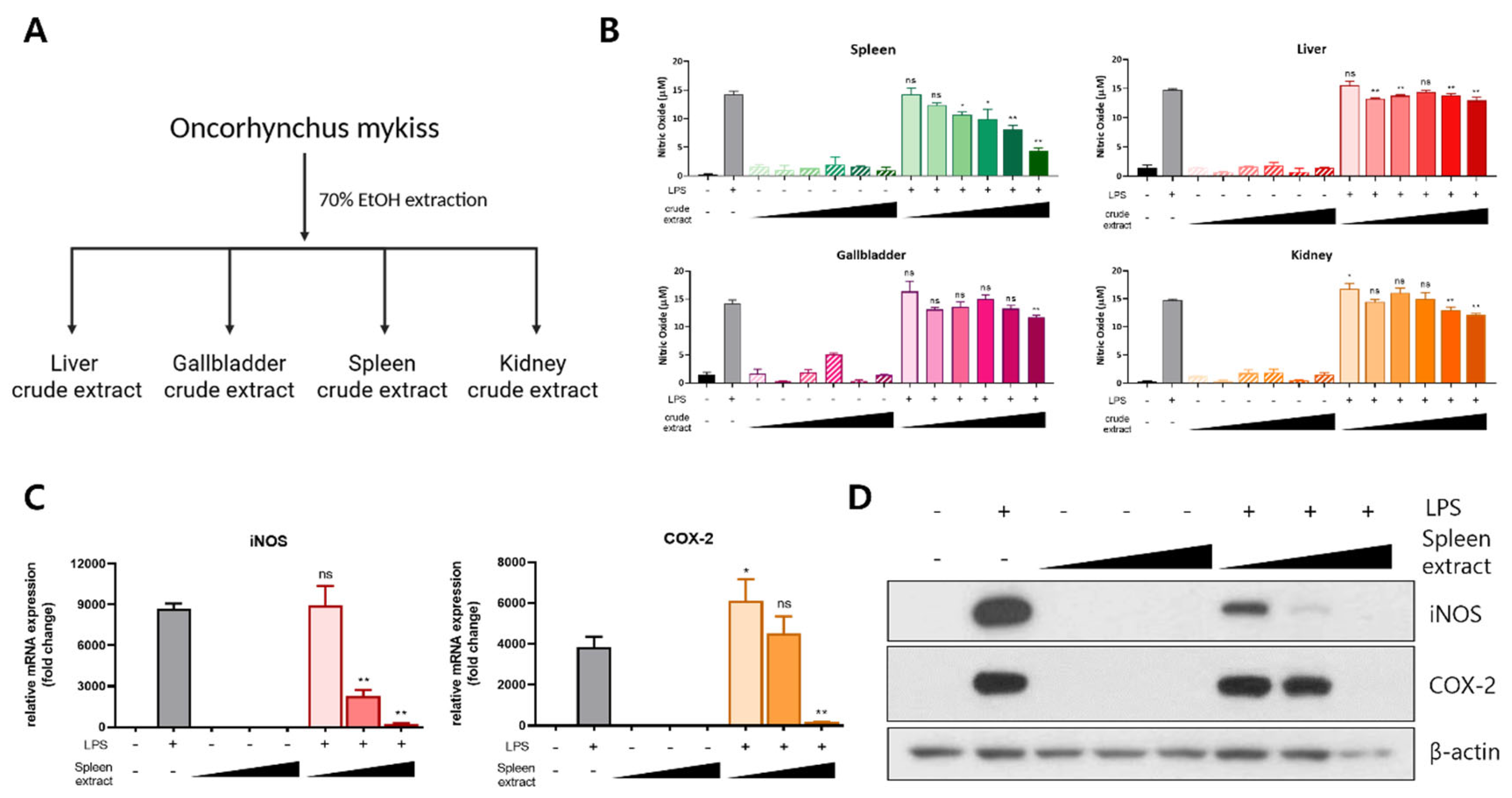
Figure 2.
Fractionation of rainbow trout spleen extract and evaluation of anti-inflammatory activity of each fraction. (A) The spleen crude extract was fractionated into four solvent layers (OSH, OSE, OSB, OSW) by polarity-based liquid–liquid extraction and lyophilized. (B) Major constituents of each fraction were characterized via UPLC-MS analysis. (C) RAW264.7 cells were treated with the crude extract or each fraction (200 μg/mL) in the presence of 1 μg/mL LPS, and mRNA expression of iNOS, COX-2, IL-6, and TNF-α was quantified by qRT-PCR. (D) Morphological changes in RAW264.7 cells were observed by phase-contrast microscopy following treatment. Data are presented as mean ± SD (n = 3). Statistical significance was determined using one-way ANOVA with multiple comparisons.
Figure 2.
Fractionation of rainbow trout spleen extract and evaluation of anti-inflammatory activity of each fraction. (A) The spleen crude extract was fractionated into four solvent layers (OSH, OSE, OSB, OSW) by polarity-based liquid–liquid extraction and lyophilized. (B) Major constituents of each fraction were characterized via UPLC-MS analysis. (C) RAW264.7 cells were treated with the crude extract or each fraction (200 μg/mL) in the presence of 1 μg/mL LPS, and mRNA expression of iNOS, COX-2, IL-6, and TNF-α was quantified by qRT-PCR. (D) Morphological changes in RAW264.7 cells were observed by phase-contrast microscopy following treatment. Data are presented as mean ± SD (n = 3). Statistical significance was determined using one-way ANOVA with multiple comparisons.
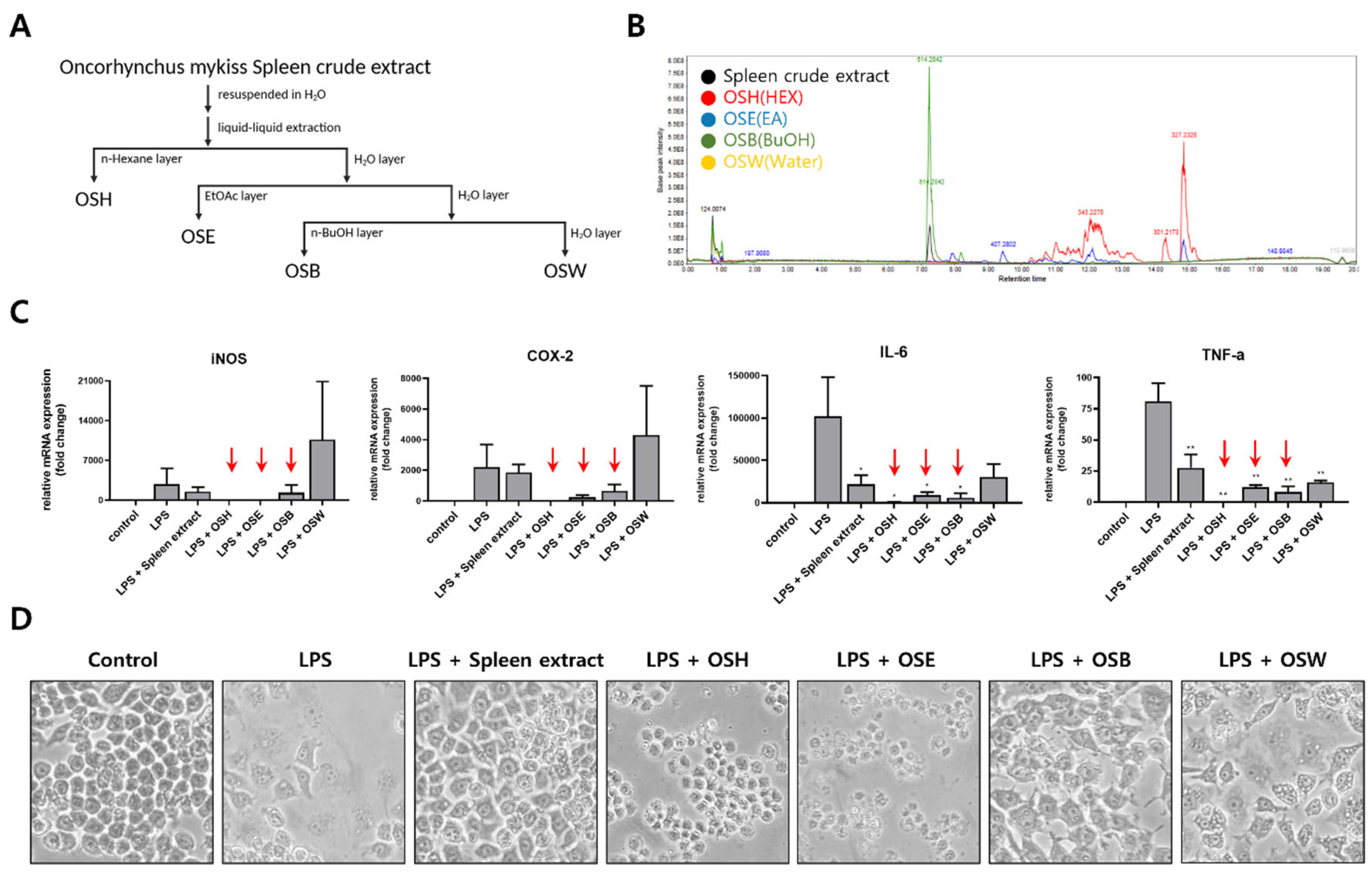
Figure 3.
Regulatory effects of the OSB fraction on iNOS and COX-2 expression in LPS-stimulated RAW264.7 cells. (A) Dose-dependent effects of OSB (100–400 μg/mL) on iNOS and COX-2 protein expression were analyzed by Western blotting. (B) Time-course analysis of iNOS and COX-2 expression following treatment with OSB (200 μg/mL) for 6–24 hours in the presence of LPS. (C, D) Immunofluorescence analysis of iNOS and COX-2 protein expression after treatment with OSB (200 μg/mL) for 24 hours. All experiments were conducted with LPS stimulation (1 μg/mL, 24 hours).
Figure 3.
Regulatory effects of the OSB fraction on iNOS and COX-2 expression in LPS-stimulated RAW264.7 cells. (A) Dose-dependent effects of OSB (100–400 μg/mL) on iNOS and COX-2 protein expression were analyzed by Western blotting. (B) Time-course analysis of iNOS and COX-2 expression following treatment with OSB (200 μg/mL) for 6–24 hours in the presence of LPS. (C, D) Immunofluorescence analysis of iNOS and COX-2 protein expression after treatment with OSB (200 μg/mL) for 24 hours. All experiments were conducted with LPS stimulation (1 μg/mL, 24 hours).
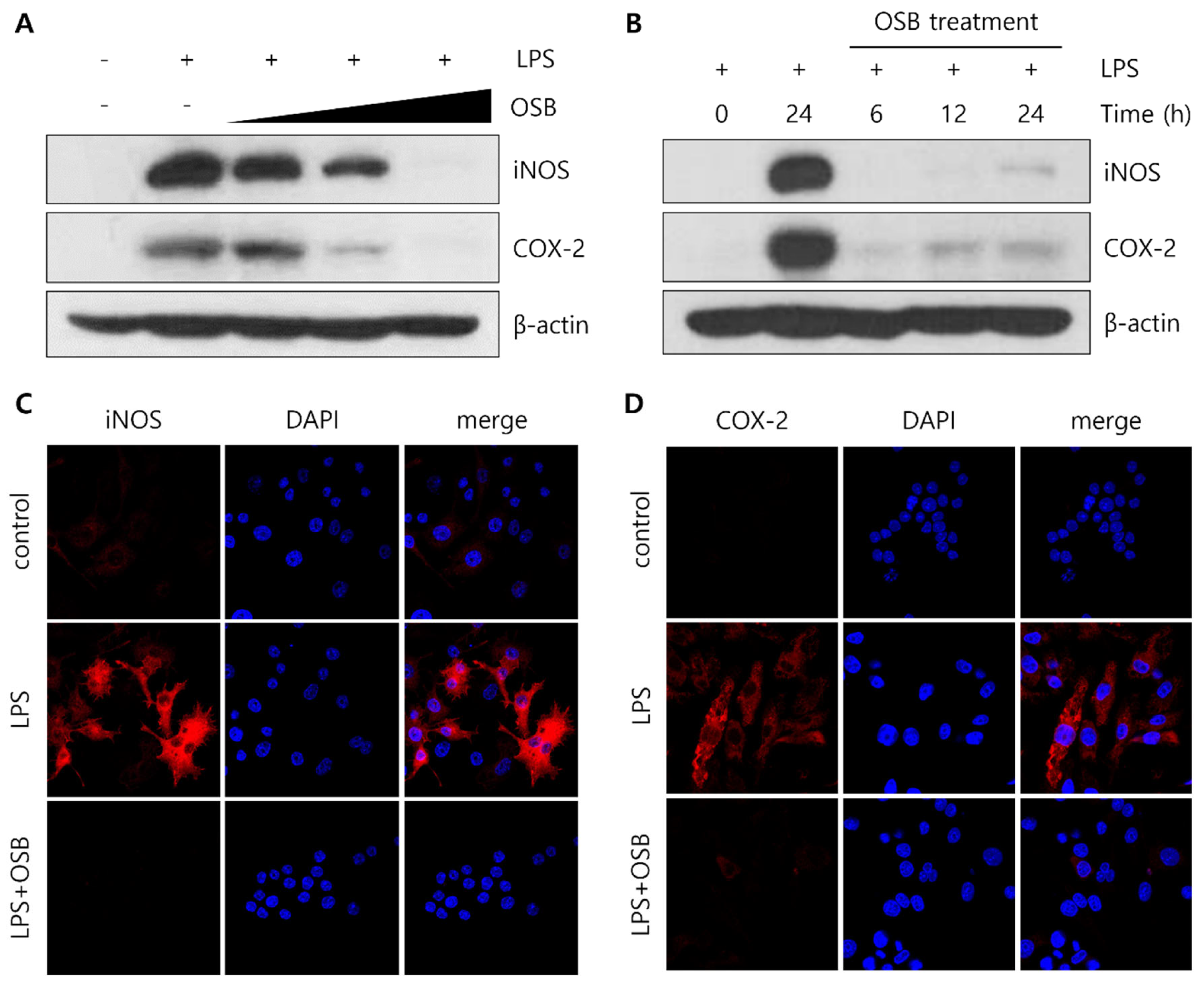
Figure 4.
Inhibitory effects of OSB on NF-κB signaling pathway activation. (A) Western blot analysis of IκBα and p65 phosphorylation in RAW264.7 cells treated with OSB (200 μg/mL) in the presence of LPS. (B) Nuclear translocation of p65 was examined after treatment with OSB (100–400 μg/mL) for 2 hours. (C) Immunofluorescence analysis of p65 localization following OSB treatment (400 μg/mL, 2 hours). Nuclear p65 fluorescence intensity was quantified using ImageJ. All experiments included LPS stimulation (1 μg/mL).
Figure 4.
Inhibitory effects of OSB on NF-κB signaling pathway activation. (A) Western blot analysis of IκBα and p65 phosphorylation in RAW264.7 cells treated with OSB (200 μg/mL) in the presence of LPS. (B) Nuclear translocation of p65 was examined after treatment with OSB (100–400 μg/mL) for 2 hours. (C) Immunofluorescence analysis of p65 localization following OSB treatment (400 μg/mL, 2 hours). Nuclear p65 fluorescence intensity was quantified using ImageJ. All experiments included LPS stimulation (1 μg/mL).
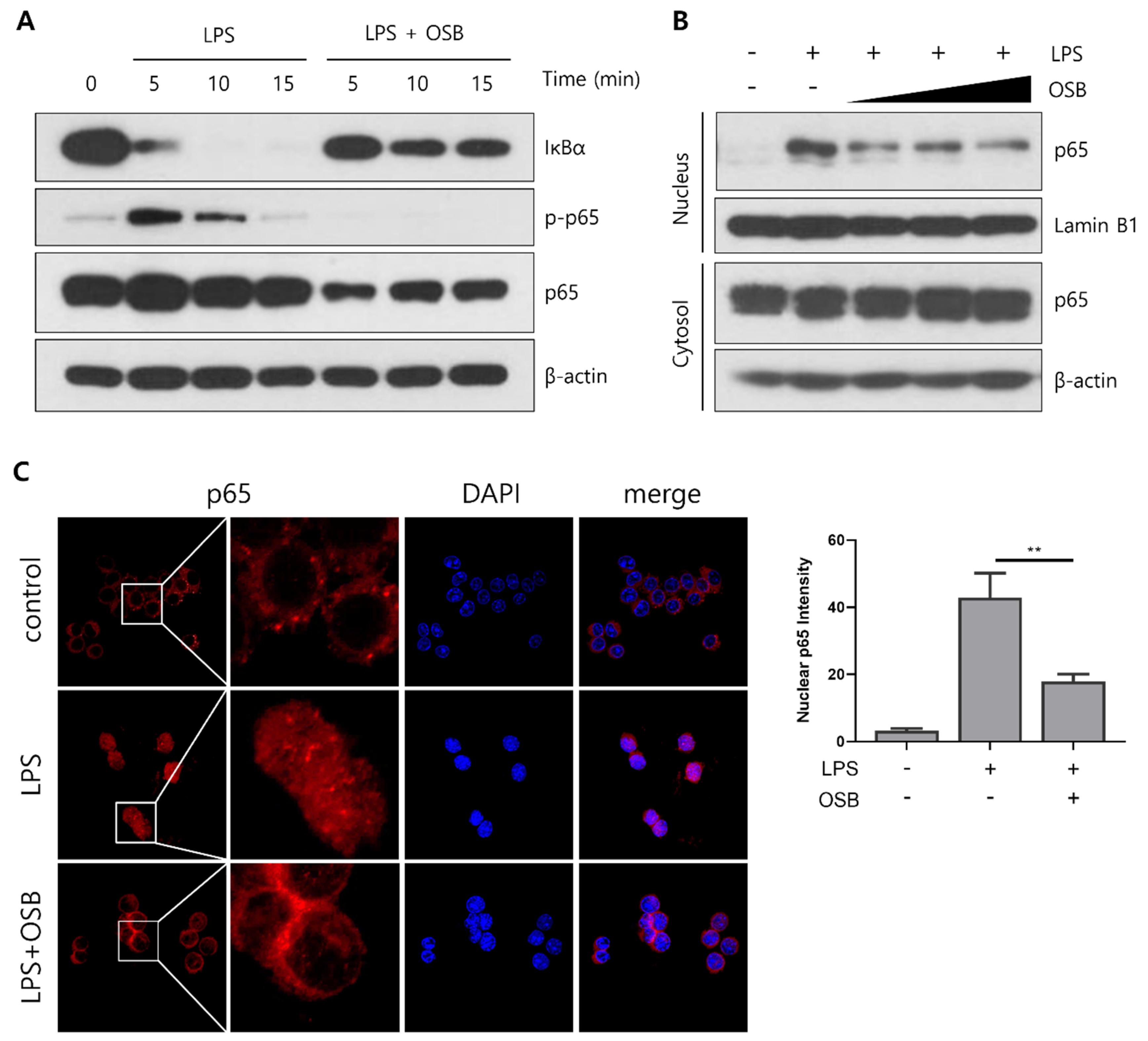
Figure 5.
Effects of OSB on LPS-induced M1 macrophage polarization. (A) qRT-PCR analysis of pro-inflammatory cytokines (IL-6, TNF-α, IL-12p40, IL-23p19), anti-inflammatory cytokines (IL-13, TGF-β), and M1 surface markers (CD40, CD86) following OSB treatment (200 μg/mL, 24 hours). (B) Immunofluorescence analysis of CD86 expression in RAW264.7 cells treated with OSB (200 μg/mL). (C) Intracellular reactive oxygen species (ROS) levels were measured using H₂DCFDA after OSB treatment (400 μg/mL, 24 hours). All cells were stimulated with 1 μg/mL LPS. Data are expressed as mean ± SD (n = 3), and statistical significance was assessed by one-way ANOVA with multiple comparisons.
Figure 5.
Effects of OSB on LPS-induced M1 macrophage polarization. (A) qRT-PCR analysis of pro-inflammatory cytokines (IL-6, TNF-α, IL-12p40, IL-23p19), anti-inflammatory cytokines (IL-13, TGF-β), and M1 surface markers (CD40, CD86) following OSB treatment (200 μg/mL, 24 hours). (B) Immunofluorescence analysis of CD86 expression in RAW264.7 cells treated with OSB (200 μg/mL). (C) Intracellular reactive oxygen species (ROS) levels were measured using H₂DCFDA after OSB treatment (400 μg/mL, 24 hours). All cells were stimulated with 1 μg/mL LPS. Data are expressed as mean ± SD (n = 3), and statistical significance was assessed by one-way ANOVA with multiple comparisons.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



