
Submitted:
22 July 2025
Posted:
22 July 2025
You are already at the latest version
Abstract
Medicinal plants have been used since ancient times for the treatment and prevention of various diseases. Plant-derived compounds offer a promising alternative to synthetic an-ti-inflammatory drugs, with potent activity and reduced toxicity. The seagrass Halodule uninervis exhibit several pharmacological properties due to the presence of various bioac-tive metabolites. However, its anti-inflammatory activity is not yet explored. In this study, we investigated the anti-inflammatory potential of an ethanolic extract of H. uninervis (HUE) in lipopolysaccharide (LPS)-stimulated RAW 264.7 macrophages by assessing its effect on the expression and activation of different inflammatory mediators. Our results revealed that HUE inhibited the LPS-stimulated inflammatory response in macrophages by downregulating the expression of inducible nitric oxide synthase (iNOS) and cycloox-ygenase-2 (COX-2) and reduced the expression of pro-inflammatory cytokines, interleu-kin-6 (IL-6) and tumor necrosis factor-alpha (TNF-α). Moreover, the anti-inflammatory potential of HUE was associated with reduced cellular migration and the suppression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB), signal transducer and activator of transcription 3 (STAT3), and mitogen-activated protein kinases (MAPKs) pathways. Altogether, H. uninervis could serve as a potent therapeutic agent in preventing chronic inflammatory diseases.
Keywords:
Halodule uninervis
; anti-inflammatory
; NF-κB
; STAT3
; MAPKs
1. Introduction
Inflammation is a physiological response mechanism of the immune system to protect the body against harmful stimuli, such as pathogens, toxins, and tissue damage [1]. Clinically, inflammation is recognized by five primary signs, including heat, pain, redness, swelling, and loss of function [2]. The inflammatory process is a complex cascade of biochemical and cellular reactions essential for maintaining physiological homeostasis. It is characterized by the recruitment and activation of immune cells that aim to eliminate the cause of infection or injury and promote tissue repair [3].
The initiation of inflammation involves a multifaceted interaction between signaling pathways and inflammatory mediators. When tissues are infected or injured, innate immune cells, such as macrophages, recognize pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs), leading to the activation of intracellular signaling pathways and production of pro-inflammatory mediators [4]. The activation of transcription factor nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) serves as a central mediator of inflammation, which induces the expression of pro-inflammatory cytokines, chemokines, and other inflammatory mediators [5]. Pro-inflammatory cytokines, such as interleukin-1 (IL-1), interleukin-6 (IL-6), and tumor necrosis factor-alpha (TNF-α), play a role in amplifying the immune response by promoting the recruitment of additional immune cells to the site of inflammation [6,7]. Concomitantly, inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) are upregulated during the inflammatory response and catalyze the synthesis of nitric oxide (NO) and prostaglandin E2 (PGE2), respectively, two crucial mediators of inflammation [8,9,10]. In addition to NF-κB, the signal transducer and activator of transcription 3 (STAT3) and mitogen-activated protein kinases (MAPKs) signaling pathways play and important role in mediating the inflammatory response. Their activation enhances the production of pro-inflammatory cytokines and other inflammatory mediators [11]. This coordinated release of inflammatory mediators forms a complex network that regulates the immune response and tissue repair to ultimately restore homeostasis.
Under normal conditions, inflammation is acute, rapidly clearing the cause of infection or injury and protecting the body from harmful stimuli [12]. However, chronic inflammation is a persistent process arising from dysregulated immune responses. Chronic inflammation contributes to the pathogenesis and progression of a wide range of diseases, including atherosclerosis, autoimmune disorders, cardiovascular diseases, and cancer [13]. Due to its severity, there is an ongoing effort to find more effective treatments for chronic inflammation. Conventional treatments of chronic inflammation typically include the administration of steroids and non-steroidal anti-inflammatory drugs (NSAIDs), which are associated with notable side effects [14,15]. Therefore, this necessitates the development of alternative treatment strategies with potent anti-inflammatory activity and high safety profile.
As a source of herbal medicine, plants have been widely used for the prevention and treatment of many disorders, including inflammatory diseases. The use of plant-derived compounds helps mitigate chronic inflammation and overcome adverse reactions due to their phytochemical diversity and natural origin [16,17]. Seagrasses are the only flowering plants to thrive underwater in marine habitats. They play fundamental ecological roles by supporting intricate food webs, providing a habitat for many marine organisms, and contributing to water quality [18,19,20]. Like other plants, seagrasses are rich in a wide variety of bioactive metabolites, which act as defense mechanisms in high stress conditions and account for different pharmacological properties observed in seagrasses [21]. Halodule uninervis, a seagrass species from the family Cymodoceaceae is abundant in tropical and subtropical coastal areas globally. The presence of diverse array of bioactive metabolites in H. uninervis extracts accounts for the plant’s pharmacological potential, exhibiting antidiabetic [22,23], antimicrobial [24], antioxidant [25,26,27,28], and anticancer properties [25,26,29]. However, research into the potential anti-inflammatory activity of H, uninervis remains unexplored.
In this study, we investigated the anti-inflammatory potential of Halodule uninervis ethanolic extract (HUE) against lipopolysaccharide (LPS)-stimulated RAW 264.7 macrophages. In particular, we assessed the effect of HUE on the expression of iNOS, COX-2, and pro-inflammatory cytokines. To understand its mechanism of action, we also explored the effect of HUE on NF-kB, STAT3, and MAPK signaling pathways, which are key regulators of the inflammatory response.
2. Materials and Methods
2.1. Collection of Halodule uninervis Leaves and Powder Preparation
Fresh leaves of Halodule uninervis were collected from the Gulf of Aqaba, Jordan. A voucher specimen of H. uninervis was deposited in the Post Herbarium, Department of Biology, American University of Beirut (AUB) under the identification number JO 2023-02. The leaves were thoroughly washed in distilled water and air-dried in the shade at room temperature. The dried leaves were then ground into fine powder using a blender and stored in the dark at room temperature for further experiments.
2.2. Preparation of Halodule uninervis Ethanolic Extract (HUE)
10 grams of dried powder was suspended in 80% ethanol. The mixture was placed on a shaker at 50 rpm and 25 °C for 72 h. The solution was then filtered using a filter paper. Afterwards, the extract was concentrated under reduced pressure in a rotary vacuum evaporator, lyophilized, and stored in the dark at 4 °C.
2.3. Cell Culture
RAW 264.7 macrophages from mice (Mus musculus) were obtained from the American Tissue Culture Collection (ATCC, Manassas, VA). Cells were cultured in DMEM high-glucose medium (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% fetal bovine serum (FBS) (Sigma-Aldrich, St. Louis, MO, USA) and 1% penicillin/streptomycin (Lonza, Switzerland) and maintained at 37°C and 5% CO2 in a humidified incubator.
2.4. Cell Viability Assay
RAW 264.7 macrophages were seeded in 96-well plates at a density of 5x103 cells per well and incubated for 24 h until they reached 30-40% confluency. Cells were then treated with different concentrations of HUE (0,10, 25, 50, 100, and 200 μg/mL) and incubated for 24 h. Cell viability was measured using 3-(4,5- dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sigma-Aldrich, St. Louis, MO, USA) reduction assay. Cell growth was calculated as the proportional viability of the HUE-treated cells in comparison with the vehicle (ethanol)-treated ones, where the viability was assumed to be 100%. The assay was performed in triplicate and repeated three times. Data are presented as mean values ± SEM.
2.5. Whole-Cell Extracts and Western Blotting
RAW 264.7 macrophages were seeded in 6-well plates at a density of 5x105 cells per well and incubated for 24 h. LPS stimulation was performed at 1 μg/mL for 30 min prior to treatment with non-toxic concentrations of HUE for 18 h. To collect whole-cell lysates, RAW 264.7 macrophages were washed with PBS and lysed using a lysis buffer (pH 6.7) containing 2% SDS, 60 mM Tris. The lysate was then centrifuged at 15,000 g for 10 minutes. Proteins concentration in the supernatants was determined by the Bradford protein assay kit (Bio-Rad, Hercules, CA, United States). Subsequently, 25–30 μg of protein extracts were resolved by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then transferred onto a polyvinylidene difluoride membrane (Immobilon PVDF; Bio-Rad, Hercules, CA, United States). The membrane was blocked with 5% nonfat dry milk in TBST (TBS and 0.05% Tween) for 1 h at room temperature. Immunodetection involved incubating the membrane with the specified primary antibodies, followed by the corresponding secondary antibody. Immunoreactive bands were visualized using the Clarity Western ECL substrate kit (Bio-Rad, Hercules, CA, United States), following the manufacturer’s instructions. The primary and secondary antibodies utilized were obtained from Cell Signaling (Cell Signaling Technology, Inc., Danvers, MA, United States). Experiments were repeated three times and data are presented as mean values ± SEM.
2.6. Real-Time PCR
RAW 264.7 macrophages were seeded in 6-well plates (5x105 cells per well) and incubated for 24 h. Cells were pre-treated with (1 μg/mL) for 30 min and were then incubated with non-toxic concentrations of HUE for 18 h. RAW 264.7 macrophages were directly homogenized in TRIzol (Sigma-Aldrich, St. Louis, MO, USA) to lyse the cells. RNA was then extracted using chloroform and precipitated with isopropanol. A nanodrop spectrophotometer (Thermo Scientific, Waltham, MA, US) was used to quantify the extracted RNA samples. The ReadyScript cDNA Synthesis Mix (Sigma-Aldrich, St. Louis, MO, USA) was used for the reverse transcription of the RNA samples into cDNA. Real-time PCR was performed in presence of SYBR green (Sigma-Aldrich, St. Louis, MO, USA) using the thermocycler CFX 96 Touch Real Time PCR (BIO RAD, Hercules, CA, US). Experiments were repeated three times and data are presented as mean values ± SEM. Primers were obtained from TIB Molbiol (Berlin, Germany). Primers used were:

Table 1.
Primers used in this paper.
| COX-2 Forward | 5′-GATACTCAGGCAGAGATGATCTACCC-3′ |
| COX-2 Reverse | 5′-AGACCAGGCACCAGACCAAAGA-3′ |
| TNF-α Forward | 5′-GTAGCCCACGTCGTAGCAAACCAC-3′ |
| TNF-α Reverse | 5′-GGTACAACCCATCGGCTGGCAC-3′ |
| IL-6 Forward | 5′-CCTCTCTGCAAGAGACTTCCATCCA-3′ |
| IL-6 Reverse | 5′-TCCTCTGTGAAGTCTCCTCTCCGG-3′ |
2.7. Trans-Well Migration Assay
Trans-well inserts (8 μm pore size; BD Biosciences, Bedford, MA, USA) were used to assess the migratory abilities of RAW 264.7 macrophages. 2.5 × 105 cells were seeded into the upper chamber of the insert. Cells were then pre-treated with (1 μg/mL) for 30 min and were then incubated with non-toxic concentrations of HUE for 24 h. DMEM medium supplemented with 10% FBS was used as a chemoattractant in the lower chamber. Cells were then incubated at 37 °C and allowed to migrate. Cells that stayed on the upper surface of the insert were removed with a sterile cotton swab. And cells that migrated to the lower surface of the insert were fixed with 4% formaldehyde, stained with DAPI (1 μg/mL), and viewed under a fluorescence microscope at 4× magnification for quantification. The assay was repeated three times, and data are presented as mean values ± SEM.
2.8. Statistical Analysis
Statistical analysis involved evaluating the data using Student’s t-test, one-way ANOVA (with Dunnett’s post hoc test), or two-way ANOVA (with Tukey–Kramer’s post hoc test). Data were expressed as mean ± SEM. A p-value below 0.05 was considered statistically significant.
3. Results
3.1. Effects of HUE on RAW 264.7 Cell Viability
The effect of H. uninervis ethanolic extract on the viability of RAW264.7 macrophages was assessed using the MTT colorimetric assay. Cells were treated with various concentrations of HUE (0, 10, 25, 50, 100, and 200 μg/mL) for 24 h. The results indicated that HUE at concentrations 10, 25, and 50 μg/mL had no significant cytotoxic effects on the viability of RAW264.7 macrophages (Figure 1). Therefore, 10 and 25 μg/mL of HUE were determined as the optimal concentrations, i.e., having anti-inflammatory effects with minimum cytotoxicity, and used in further experiments.
3.2. HUE Decreases Levels of COX-2 and iNOS in LPS-Stimulated RAW 264.7 Cells
COX-2 is a crucial enzyme in the inflammatory response due to its role in producing prostaglandin E2 (PGE2), a potent pro-inflammatory mediator [30]. The effect of HUE on the mRNA expression levels and the protein levels of COX-2 in LPS-stimulated RAW 264.7 macrophages was evaluated using RT-PCR and western blotting, respectively. As shown in Figure 2, LPS-stimulation induced the mRNA expression of COX-2, while HUE treatment decreased COX-2 expression in a concentration-dependent manner, achieving a significant decrease at 25 μg/mL (0.46 ± 0.02-fold change in the LPS-treated control). Furthermore, COX-2 protein levels were significantly increased in LPS-stimulated cells. Consistent with its effect on mRNA expression levels, results showed that treatment with HUE reduced the protein levels of COX-2 in a concentration-dependent manner, achieving a significant decrease at 25 μg/mL (1.26 ± 0.26-fold change in the LPS-treated control).
iNOS is another pro-inflammatory enzyme that is upregulated in macrophages in response to inflammatory signals. iNOS catalyzes the production of nitric oxide (NO), a signaling molecule that modulates the inflammatory response [31]. The protein levels of iNOS were assessed in LPS-stimulated RAW 264.7 macrophages with and without HUE treatment. Results showed that LPS increased the expression of iNOS, and 10 or 25 μg/mL HUE significantly reduced the protein levels of iNOS by 0.90 ± 0.11- and 0.77 ± 0.11-fold, respectively, compared to the LPS-treated control (Figure 2).
3.3. HUE Reduces TNF-α and IL-6 Cytokines mRNA Expression Levels in LPS-Stimulated RAW 264.7 Cells
Other essential mediators of inflammation include pro-inflammatory cytokines. Upon their production in response to inflammatory stimuli, these signaling molecules play crucial roles in the initiation, amplification, and regulation of inflammatory responses. TNF-α and IL-6 are among the first molecules released in response to infection or tissue injury, signaling the onset of the inflammatory cascade [32,33]. Therefore, targeting them is critical to help reduce the overall inflammatory burden. Treating LPS-stimulated RAW 264.7 macrophages with HUE suppressed TNF-α expression in a concentration-dependent manner, achieving a significant decrease at 25 μg/mL (0.37 ± 0.06-fold change in the LPS-treated control). While HUE at concentrations 10 and 25 μg/mL significantly suppressed the upregulation of IL-6 (0.39 ± 0.05- and 0.22 ± 0.04-fold reductions, respectively) mRNA expression levels (Figure 3). These results suggest that HUE exerts its anti-inflammatory effect possibly by inhibiting the expression of pro-inflammatory cytokines at the transcription level.
3.4. HUE Inhibits the Phosphorylation of NF-κB in LPS-Stimulated RAW 264.7 Cells
NF-κB is a key mediator in the network of inflammatory pathways. It is a transcription factor that is activated through phosphorylation in response to various stimuli, including oxidative stress, pathogens, and cytokines. Upon activation, NF-κB translocates into the nucleus and promotes the transcription of pro-inflammatory genes, such as enzymes (iNOS and COX-2) and cytokines (IL-1, IL-6, and TNF-α) [5]. To investigate if HUE mediates its anti-inflammatory effect via targeting NF-κB, we assessed the protein levels of phosphorylated NF-κB in HUE-treated RAW 264.7 macrophages. Our results revealed that HUE treatment at 10 and 25 μg/mL significantly downregulated the levels of phosphorylated NF-κB by 1.35 ± 0.003- and 1.19 ± 0.07-fold, respectively, relative to the LPS-treated control (Figure 4). Altogether, these data suggest that HUE mediates its inhibitory effects on pro-inflammatory enzymes and cytokines by impeding the activation of NF-κB.
3.5. HUE Inhibits the STAT3 Pathway in LPS-Stimulated RAW 264.7 Cells
Another major mediator of the inflammatory response is the STAT3 signaling pathway. Once activated by phosphorylation, STAT3 promotes the expression of genes encoding pro-inflammatory cytokines [34]. The protein levels of phosphorylated STAT3 were assessed in HUE-treated RAW 264.7 macrophages. Results showed that treatment with LPS increased the levels of phosphorylated STAT3, whereas treatment with 10 and 25 μg/mL of HUE significantly its phosphorylation by 1.56 ± 0.06- and 1.36 ± 0.08-fold, respectively, compared to the LPS-treated control (Figure 5). Taken together, these results indicate that HUE exhibits its anti-inflammatory activity by targeting the STAT3 signaling pathway.
3.6. HUE Inhibits the MAPKs Pathway in LPS-Stimulated RAW 264.7 Cells
MAPKs, a family of serine/threonine protein kinases, play a role in regulating fundamental biological processes, including proliferation, cell survival, cell motility, differentiation, as well as inflammation [35]. In response to external inflammatory stimuli, MAPKs, in particular ERK and p38, promote the inflammatory response by activating several pro-inflammatory mediators, such as the NF-κB signaling pathway [36]. To determine the effect of HUE on the activation of MAPKs, the protein levels of phosphorylated ERK and p38 were assessed in LPS-stimulated RAW 264.7 macrophages. Results showed that treatment with 25 μg/mL of HUE significantly HUE inhibited the LPS-mediated increase in the levels of phosphorylated ERK and p38 by 1.46 ± 0.3-fold and 1.46 ± 0.03-fold, respectively, compared to the LPS-treated control (Figure 6). Overall, these results reveal an anti-inflammatory activity of HUE by targeting the MAPKs signaling pathway.
3.7. HUE Inhibits the Cellular Migration of LPS-Stimulated RAW 264.7 Cells
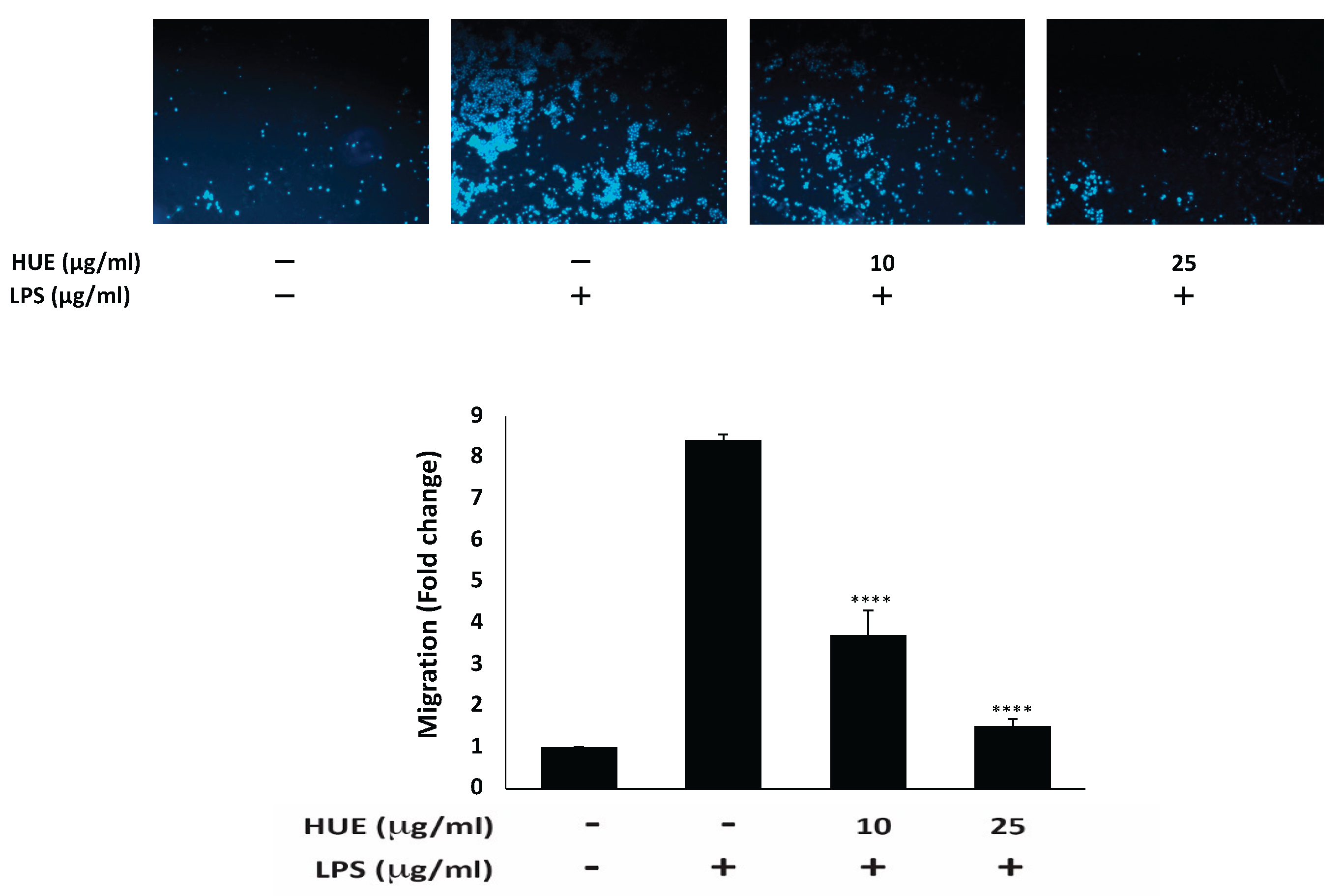
While cell migration is a crucial mechanism in immune surveillance, tissue repair, and wound healing, aberrant cell migration underlies pathological conditions such as chronic inflammation. Indeed, the sustained migration of macrophages in chronic inflammation exacerbates tissue damage and prolongs inflammatory responses, ultimately leading to progressive chronic diseases [3]. The effect of HUE on the migration of RAW 264.7 macrophages was assessed using trans-well migration assay. Results indicate that HUE significantly reduced the migration ability of LPS-stimulated RAW 264.7 macrophages. Particularly, Figure 7 shows that treatment with 10 and 25 μg/mL of HUE caused a significant decrease in the number of cells crossing from the upper chamber to the lower one by 3.7 ± 0.6- and 1.5 ± 0.2-fold, respectively, compared to the LPS-treated control.
4. Discussion
Inflammation is a fundamental biological response to harmful stimulus, including infection and tissue injury. Chronic inflammation develops when the immune system fails to rapidly eliminate the initial cause of infection or injury. As its persistence is implicated in the development and pathogenesis of many diseases, managing inflammation is essential to prevent chronic diseases and maintain overall health. Despite their efficacy as anti-inflammatory drugs, steroids and NSAIDs are associated with adverse effects when administered for a prolonged period of time. Therefore, there is a growing interest in utilizing plant-derived compounds for the development of novel anti-inflammatory agents.
Plants are rich in a plethora of bioactive metabolites, which are known to exhibit antioxidant, antimicrobial, anticancer, and anti-inflammatory activities [37,38]. These compounds include alkaloids, flavonoids, polyphenols, quinones, and tannins, among others [39,40,41]. Consequently, modern research continues to explore the utilization of plant-derived compounds as therapeutic agents. Although several plants have been known for their anti-inflammatory activity, the anti-inflammatory potential of the seagrass H. unienrvis remains poorly investigated. In this study, we evaluated the anti-inflammatory potential of the ethanolic extract of H. uninervis (HUE) on LPS-stimulated RAW 264.7 macrophages by assessing its effect on the production of well-known pro-inflammatory cytokines and mediators.
LPS, a component of the outer membrane of gram-negative bacteria, is a potent activator of the immune system [42,43]. It is used as a pro-inflammatory stimulus to activate macrophages and trigger the immune response in vitro. Binding of LPS to toll-like receptor 4 (TLR4) on the surface of macrophages initiates intracellular signaling cascades, leading to the production and release of pro-inflammatory mediators and cytokines, such as NO, iNOS, COX-2, TNF-α, and ILs [44].
iNOS is an enzyme that plays a crucial role in mediating immune responses. It is induced in activated macrophages and catalyzes the synthesis of NO, a pro-inflammatory signaling molecule. NO is involved in promoting vasodilation, increasing vascular permeability, and recruiting more immune cells to the site of infection or injury [31]. Our results showed that LPS induced the activation of iNOS in macrophages; however, treatment with HUE downregulated iNOS protein levels. These results suggest that the inhibition of iNOS could contribute to the anti-inflammatory potential of HUE possibly by inhibiting the production of NO.
Another enzyme induced by inflammatory stimuli is COX-2, which catalyzes the synthesis of PGE2 from arachidonic acid. PGE2, a lipid mediator, is predominantly expressed in LPS-stimulated macrophages and contributes to the inflammatory response [45,46]. It induces vasodilation, enhances vascular permeability, and sensitizes nerve endings to pain, leading to redness, swelling, and pain, which are classic features of inflammation [30]. Therefore, targeting COX-2 has emerged as a promising strategy in the development of anti-inflammatory drugs, such as NSAIDs. HUE suppressed the mRNA expression levels of COX-2, which could in turn inhibit formation of PGE2 in LPS-stimulated macrophages. In accordance with our results, several plant extracts, including Morus alba [47], Matricaria chamomilla [48], Lasia spinosa [49], and the seagrass Halodule pinifolia [50], among others, were shown to exert their anti-inflammatory activity by targeting the production of NO and PGE2 through the downregulation of both iNOS and COX-2, respectively.
When exposed to inflammatory stimuli, macrophages secrete cytokines, such as TNF-α and IL-6. While essential for initiating the inflammatory response to eliminate infections and repair tissue injury, the dysregulated production of pro-inflammatory cytokines can lead to chronic inflammation, contributing to the pathogenesis of many diseases. These potent signaling molecules play a role in amplifying the immune response by activating endothelial cells, recruiting inflammatory cells, and stimulating the production of other pro-inflammatory mediators [51]. In our study, LPS stimulation upregulated the mRNA expression of both TNF-α and IL-6. This increase was suppressed by the treatment with HUE, which is in line with the reported anti-inflammatory activity of Halodule pinifolia, a seagrass species within the same family of H. uninervis. The ethyl acetate extract of H. pinifolia was found to effectively attenuate the secretion of pro-inflammatory cytokines in LPS-, carrageenan-, and crystal-induced inflammation in both in vitro and in vivo models [50]. These results are also in accordance with other studies reporting the anti-inflammatory activity of different terrestrial plant extracts by targeting TNF-α and IL-6 cytokines [47,49,52,53].
NF-κB is a crucial transcription factor that regulates the expression of genes encoding pro-inflammatory cytokines (TNF-α and IL-6) and enzymes (iNOS and COX-2), thus amplifying and sustaining the inflammatory response. LPS binds to the kappa B inhibiting protein (IκB), freeing and activating NF-κB in the cytoplasm, which then translocates to the nucleus and promotes the activation of pro-inflammatory mediators [54,55]. Here, HUE treatment decreased the phosphorylation of the p65 subunit of NF-κB in LPS-stimulated macrophages. Indeed, phosphorylation of the p65 subunit has been shown to enhance the transcriptional activity of NF-κB, promoting the expression of pro-inflammatory genes [56,57,58]. Therefore, inhibiting the phosphorylation of the p65 subunit could serves as a promising therapeutic strategy to help mitigate chronic inflammation. Given that NF-κB activation is a hallmark of the inflammatory response, the ability of HUE to target NF-κB further supports its anti-inflammatory potential. In accordance, the suppression of NF-κB activation by inhibiting the phosphorylation of p65 subunit has been the target of several plant extracts as documented in other studies [48,52,53,59]. Overall, these results suggest that HUE downregulates the expression levels of iNOS, COX-2, TNF-α, and IL-6 possibly by the inhibition of NF-κB activation.
Besides NF-κB, the STAT3 signaling pathway plays a central role in promoting the inflammatory response. Upon activation by inflammatory stimuli, STAT3 translocates to the nucleus and regulates the expression of genes encoding pro-inflammatory mediators [11,60]. Suppressing the phosphorylation of STAT3 by different plant extracts was shown to attenuate LPS-induced inflammation by inhibiting the production of NO, PGE2, TNF-α, and IL-6, as well as the activation of NF-κB [61,62,63,64]. In line with those studies, the anti-inflammatory activity of HUE in LPS induced macrophages could be mediated via the downregulation of the STAT3 pathway and its targeted pro-inflammatory genes. Notably, STAT3 and NF-κB are highly interconnected. Studies have demonstrated that the inhibition of STAT3 and NF-κB pathways mitigates inflammation by inhibiting the production of pro-inflammatory factors [65,66,67,68]. Therefore, the development of anti-inflammatory drugs co-targeting STAT3 and NF-κB represents a promising strategy in treating chronic inflammation.
MAPK signaling pathway is also involved in regulating and mediating the inflammatory response in macrophages. MAPKs include ERK (extracellular signal- regulated kinase), JNK (c-Jun N-terminal kinase), and p38 MAPK [35]. Upon activation by inflammatory stimuli, MAPKs promote the production of pro-inflammatory mediators through the activation of STAT3 and NF-kB signaling pathways [69,70]. In our study, HUE was shown to downregulate the phosphorylated levels of ERK1/2 and p38 MAPK in LPS-stimulated macrophages, suggesting that both MAPKs could inhibit the activation of NF-kB and STAT3, leading to a reduction in pro-inflammatory mediators. Several studies reported the anti-inflammatory effects of various plant extracts by the inhibition of MAPK, STAT3, and NF-kB pathways [71,72,73]. Consistent with these findings, our results indicate that the anti-inflammatory activity of HUE is likely mediated by its impact on the NF-kB, STAT3, and MAPK signaling pathways.
In addition to targeting inflammatory mediators and signaling pathways, HUE demonstrated a significant impact on macrophage migration, a key event in the progression of inflammation. While macrophage recruitment to damaged tissues is essential for initiating tissue repair and restoring of homeostasis, uncontrolled or prolonged migration can exacerbate inflammation, leading to chronic diseases, such as atherosclerosis, cancer, and pulmonary fibrosis [3,74]. Therefore, the ability of HUE to suppress macrophage migration under inflammatory conditions may help prevent the amplification and persistence of inflammation. Several studies showed that plant extracts impede the inflammatory response by inhibiting the migratory potential of macrophages [75,76,77]. In accordance, HUE may contribute to resolving inflammatory responses by attenuating macrophage migration, underscoring its therapeutic potential in inflammatory disorders.
The ability of HUE to suppress the LPS-induced inflammatory response in macrophages could be attributed to the presence of a diversity of phytochemical compounds. We have previously identified the presence of 18 bioactive metabolites in HUE, most of which are phenols and flavonoids that exhibit several pharmacological activities, such as antioxidant, anticancer, antimicrobial, as well as anti-inflammatory [25]. Some of the compounds in HUE known for their anti-inflammatory activity include apigenin, coumaric acid, kaempferol, and vanillic acid. For example, apigenin is an abundantly occurring flavonoid found in many fruits and vegetables. The anti-inflammatory activity of apigenin has been well documented in both in vitro and in vivo models. For instance, apigenin has been reported to exert its anti-inflammatory activity by inhibiting the production of pro-inflammatory cytokines and mediators, mainly through targeting major pro-inflammatory signaling pathways involved in inflammation, such as NF-κB, MAPKs, and STAT3 [78,79,80,81,82]. Kaempferol is another example of natural flavonoids found in fruits and vegetables with anti-inflammatory activity. Several studies have demonstrated that kaempferol exerts its anti-inflammatory activity by inhibiting NF-κB signaling pathway, which mediates the expression of many pro-inflammatory mediators, including iNOS and COX-2 [40,83,84]. Vanillic acid, a derivative of vanillin, is a naturally occurring phenolic acid primarily found in vanilla bean. Vanillic acid has been also shown to possess anti-inflammatory properties by inhibiting NF-κB-mediated inflammatory response [85,86,87,88]. And the HPLC analysis of an ethyl acetate extract of the seagrass Halodule pinifolia revealed the presence of high amounts of vanillic acid, which attributed to the anti-inflammatory activity of the extract in vitro and in vivo [50]. Additionally, coumaric acid, another example of phenolic acids, exhibits anti-inflammatory activity in several in vitro and in vivo models of inflammation. Studies have reported that coumaric acid mitigates inflammation by suppressing the expression of pro-inflammatory cytokines, such as TNF-α and IL-6, possibly by targeting the NF-κB signaling pathway [89,90,91,92].
5. Conclusions
To the best of our knowledge, this is the first study to investigate the anti-inflammatory activity of the ethanolic extract of H. uninervis (HUE) in LPS-stimulated macrophages. Our results indicate an anti-inflammatory potential of HUE by targeting key players of inflammation. HUE at non-cytotoxic concentrations suppressed the expression of iNOS and COX-2, inhibiting the production of NO and PFE2, respectively. Moreover, HUE treatment reduced the secretion of pro-inflammatory cytokines, TNF-α and IL-6. The anti-inflammatory activity of HUE could be potentially mediated by targeting NF-κB, STAT3, and MAPKs signaling pathways. The pharmacological activities of HUE, including anti-inflammation, are attributed to the presence of a wide array of bioactive metabolites. Further research will be carried out to purify and isolate potential anti-inflammatory compounds present in HUE. Moreover, the anti-inflammatory activity of HUE in an in vitro model of inflammation will also be investigated. Taken together, our findings provide evidence for the anti-inflammatory activity of H. uninervis, highlighting its potential use in the development of anti-inflammatory agents for the treatment of chronic diseases.
Author Contributions
Conceptualization, N.W. and E.B.; methodology, N.W, R.A., J.E.M., and E.B.; validation, M.M., and E.B.; formal analysis, N.W., and R.A.; resources, M.M., E.B., and A.B.; data curation, N.W., R.A, and E.B.; writing—original draft preparation, N.W.; writing—review and editing, N.W., R.A., and J.E.M.; supervision, E.B.; project administration, E.B.; funding acquisition, E.B. and A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by a grant from the University Research Board of the American University of Beirut, Lebanon, to E.B. and the University of Petra, Amman, Jordan, to A.B.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The authors declare that the data supporting the findings of this study are available within the paper. Should any raw data files be needed in another format, they are available from the corresponding author upon reasonable request.
Acknowledgments
The authors acknowledge the University Research Board (URB) of the American University of Beirut and the University of Petra for their generous support provided.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Netea, M.G. , et al., A guiding map for inflammation. Nature immunology, 2017. 18(8): p. 826-831.
- Rather, L.J. , Disturbance of function (functio laesa): the legendary fifth cardinal sign of inflammation, added by Galen to the four cardinal signs of Celsus. Bulletin of the New York Academy of Medicine, 1971. 47(3): p. 303.
- Luster, A.D., R. Alon, and U.H. von Andrian, Immune cell migration in inflammation: present and future therapeutic targets. Nature immunology, 2005. 6(12): p. 1182-1190.
- Roh, J.S. and D.H. Sohn, Damage-associated molecular patterns in inflammatory diseases. Immune network, 2018. 18(4).
- Liu, T. , et al., NF-κB signaling in inflammation. Signal transduction and targeted therapy, 2017. 2(1): p. 1-9.
- Zhang, J.-M. and J. An, Cytokines, inflammation, and pain. International anesthesiology clinics, 2007. 45(2): p. 27-37.
- Feghali, C.A. and T.M. Wright, Cytokines in acute and chronic inflammation. Front Biosci, 1997. 2(1): p. d12-d26.
- Pan, M.-H. , et al., Acacetin suppressed LPS-induced up-expression of iNOS and COX-2 in murine macrophages and TPA-induced tumor promotion in mice. Biochemical pharmacology, 2006. 72(10): p. 1293-1303.
- Zamora, R., Y. Vodovotz, and T.R. Billiar, Inducible nitric oxide synthase and inflammatory diseases. Molecular medicine, 2000. 6: p. 347-373.
- Simon, L.S. , Role and regulation of cyclooxygenase-2 during inflammation. The American journal of medicine, 1999. 106(5): p. 37S-42S.
- Xia, T. , et al., Advances in the role of STAT3 in macrophage polarization. Frontiers in immunology, 2023. 14: p. 1160719.
- Hannoodee, S. and D.N. Nasuruddin, Acute inflammatory response. 2020.
- Pahwa, R., A. Goyal, and I. Jialal, Chronic inflammation. 2018.
- Bindu, S., S. Mazumder, and U. Bandyopadhyay, Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochemical pharmacology, 2020. 180: p. 114147.
- Patrignani, P. and C. Patrono, Cyclooxygenase inhibitors: from pharmacology to clinical read-outs. Biochimica et Biophysica Acta (BBA)-Molecular and Cell Biology of Lipids, 2015. 1851(4): p. 422-432.
- Nunes, C.d.R. , et al., Plants as Sources of Anti-Inflammatory Agents. Molecules, 2020. 25(16): p. 3726.
- Tasneem, S. , et al., Molecular pharmacology of inflammation: Medicinal plants as anti-inflammatory agents. Pharmacological research, 2019. 139: p. 126-140.
- Potouroglou, M. , et al., Measuring the role of seagrasses in regulating sediment surface elevation. Scientific reports, 2017. 7(1): p. 11917.
- Ondiviela, B. , et al., The role of seagrasses in coastal protection in a changing climate. Coastal Engineering, 2014. 87: p. 158-168.
- Nagelkerken, I. , et al., Importance of mangroves, seagrass beds and the shallow coral reef as a nursery for important coral reef fishes, using a visual census technique. Estuarine, coastal and shelf science, 2000. 51(1): p. 31-44.
- Kim, D.H. , et al., Nutritional and bioactive potential of seagrasses: A review. South African Journal of Botany, 2021. 137: p. 216-227.
- Baehaki, A. , et al., Antidiabetic Activity with N-Hexane, Ethyl-Acetate and Ethanol Extract of Halodule uninervis Seagrass. Pharmacognosy Journal, 2020. 12(4).
- Karthikeyan, R. and M. Sundarapandian, Antidiabetic activity of methanolic extract of Halodule uninervis in streptozotocin-induced diabetic mice. Journal of Pharmaceutical Sciences and Research, 2017. 9(10): p. 1864-1868.
- Supriadi, A., A. Baehaki, and M.C. Pratama, Antibacterial activity of methanol extract from seagrass of Halodule uninervis in the coastal of Lampung. Pharm Lett, 2016. 8: p. 77-79.
- Wehbe, N. , et al., The Antioxidant Potential and Anticancer Activity of Halodule uninervis Ethanolic Extract against Triple-Negative Breast Cancer Cells. Antioxidants, 2024. 13(6): p. 726.
- Ghandourah, M. , et al., Fatty Acids and Other Chemical Compositions of Some Seagrasses Collected from the Saudi Red Sea with Potential of Antioxidant and Anticancer Agents. Thalassas: An International Journal of Marine Sciences, 2021. 37: p. 13-22.
- Baehaki, A., A. Supriadi, and M.C. Pratama, Antioxidant Activity of Methanol Extract of Halodule uninervis Seagrass from the Coastal of Lampung, Indonesia. RESEARCH JOURNAL OF PHARMACEUTICAL BIOLOGICAL AND CHEMICAL SCIENCES, 2016. 7(3): p. 1173-1177.
- Ramah, S. , et al., Prophylactic antioxidants and phenolics of seagrass and seaweed species: A seasonal variation study in a Southern Indian Ocean Island, Mauritius. Internet Journal of Medical Update-EJOURNAL, 2014. 9(1): p. 27-37.
- Parthasarathi, P. , et al., Phytochemical screening and in-vitro anticancer activity of ethyl acetate fraction of Seagrass Halodule uninervis from Mandapam Coastal Region Rameswaram Gulf of Mannar India. International Journal of Pharmaceutical Sciences and Drug Research, 2021. 13(6): p. 677-684.
- Kawahara, K. , et al., Prostaglandin E2-induced inflammation: Relevance of prostaglandin E receptors. Biochimica et Biophysica Acta (BBA)-Molecular and Cell Biology of Lipids, 2015. 1851(4): p. 414-421.
- Korhonen, R. , et al., Nitric oxide production and signaling in inflammation. Current Drug Targets-Inflammation & Allergy, 2005. 4(4): p. 471-479.
- Parameswaran, N. and S. Patial, Tumor necrosis factor-α signaling in macrophages. Critical Reviews™ in Eukaryotic Gene Expression, 2010. 20(2).
- Tanaka, T., M. Narazaki, and T. Kishimoto, IL-6 in inflammation, immunity, and disease. Cold Spring Harbor perspectives in biology, 2014. 6(10): p. a016295.
- Kasembeli, M.M. , et al., Contribution of STAT3 to inflammatory and fibrotic diseases and prospects for its targeting for treatment. International journal of molecular sciences, 2018. 19(8): p. 2299.
- Cargnello, M. and P.P. Roux, Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiology and molecular biology reviews, 2011. 75(1): p. 50-83.
- Kaminska, B. , MAPK signalling pathways as molecular targets for anti-inflammatory therapy—from molecular mechanisms to therapeutic benefits. Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics, 2005. 1754(1-2): p. 253-262.
- Choudhari, A.S. , et al., Phytochemicals in cancer treatment: From preclinical studies to clinical practice. Frontiers in pharmacology, 2020. 10: p. 1614.
- Nisar, A. , et al., Phytochemicals in the treatment of inflammation-associated diseases: the journey from preclinical trials to clinical practice. Frontiers in pharmacology, 2023. 14: p. 1177050.
- Ginwala, R. , et al., Potential role of flavonoids in treating chronic inflammatory diseases with a special focus on the anti-inflammatory activity of apigenin. Antioxidants, 2019. 8(2): p. 35.
- Kadioglu, O. , et al., Kaempferol is an anti-inflammatory compound with activity towards NF-κB pathway proteins. Anticancer research, 2015. 35(5): p. 2645-2650.
- Wang, X.-L. , et al., The effects of resveratrol on inflammation and oxidative stress in a rat model of chronic obstructive pulmonary disease. Molecules, 2017. 22(9): p. 1529.
- Page, M.J., D. B. Kell, and E. Pretorius, The role of lipopolysaccharide-induced cell signalling in chronic inflammation. Chronic Stress, 2022. 6: p. 24705470221076390.
- Fujihara, M. , et al., Molecular mechanisms of macrophage activation and deactivation by lipopolysaccharide: roles of the receptor complex. Pharmacology & therapeutics, 2003. 100(2): p. 171-194.
- Kawai, T. and S. Akira, The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nature immunology, 2010. 11(5): p. 373-384.
- Sheppe, A.E. , et al., PGE2 augments inflammasome activation and M1 polarization in macrophages infected with Salmonella Typhimurium and Yersinia enterocolitica. Frontiers in microbiology, 2018. 9: p. 2447.
- Williams, J.A. and E. Shacter, Regulation of macrophage cytokine production by prostaglandin E2: distinct roles of cyclooxygenase-1 and-2. Journal of Biological Chemistry, 1997. 272(41): p. 25693-25699.
- Suriyaprom, S. , et al., Antioxidant and anti-inflammatory activity on LPS-stimulated RAW 264.7 macrophage cells of white mulberry (Morus alba L.) leaf extracts. Molecules, 2023. 28(11): p. 4395.
- Duan, X. , et al., Chemical component and in vitro protective effects of Matricaria chamomilla (L.) against lipopolysaccharide insult. Journal of Ethnopharmacology, 2022. 296: p. 115471.
- Nguyen, T.Q. , et al., Anti-inflammatory effects of Lasia spinosa leaf extract in lipopolysaccharide-induced RAW 264.7 macrophages. International Journal of Molecular Sciences, 2020. 21(10): p. 3439.
- Begum, S.A. , et al., Halodule pinifolia (Seagrass) attenuated lipopolysaccharide-, carrageenan-, and crystal-induced secretion of pro-inflammatory cytokines: mechanism and chemistry. Inflammopharmacology, 2021. 29: p. 253-267.
- Kany, S., J. T. Vollrath, and B. Relja, Cytokines in inflammatory disease. International journal of molecular sciences, 2019. 20(23): p. 6008.
- Okuda-Hanafusa, C. , et al., Turmeronol A and turmeronol B from Curcuma longa prevent inflammatory mediator production by lipopolysaccharide-stimulated RAW264. 7 macrophages, partially via reduced NF-κB signaling. Food & function, 2019. 10(9): p. 5779-5788.
- Yu, M.-H. , et al., Suppression of LPS-induced inflammatory activities by Rosmarinus officinalis L. Food Chemistry, 2013. 136(2): p. 1047-1054.
- Sharif, O. , et al., Transcriptional profiling of the LPS induced NF-κB response in macrophages. BMC immunology, 2007. 8: p. 1-17.
- Tak, P.P. and G.S. Firestein, NF-κB: a key role in inflammatory diseases. The Journal of clinical investigation, 2001. 107(1): p. 7-11.
- Christian, F., E. L. Smith, and R.J. Carmody, The regulation of NF-κB subunits by phosphorylation. Cells, 2016. 5(1): p. 12.
- Naumann, M. and C. Scheidereit, Activation of NF-kappa B in vivo is regulated by multiple phosphorylations. The EMBO journal, 1994. 13(19): p. 4597-4607.
- Moreno, R. , et al., Specification of the NF-κB transcriptional response by p65 phosphorylation and TNF-induced nuclear translocation of IKKε. Nucleic acids research, 2010. 38(18): p. 6029-6044.
- Vasarri, M. , et al., Anti-inflammatory properties of the marine plant Posidonia oceanica (L.) Delile. Journal of ethnopharmacology, 2020. 247: p. 112252.
- Zhao, J. , et al., Protective effect of suppressing STAT3 activity in LPS-induced acute lung injury. American Journal of Physiology-Lung Cellular and Molecular Physiology, 2016. 311(5): p. L868-L880.
- Yaqin, Z. , et al., Resveratrol alleviates inflammatory bowel disease by inhibiting JAK2/STAT3 pathway activity via the reduction of O-GlcNAcylation of STAT3 in intestinal epithelial cells. Toxicology and Applied Pharmacology, 2024. 484: p. 116882.
- Liu, Y. , et al., 6-Gingerol attenuates microglia-mediated neuroinflammation and ischemic brain injuries through Akt-mTOR-STAT3 signaling pathway. European Journal of Pharmacology, 2020. 883: p. 173294.
- Li, L. , et al., Echinacoside alleviated LPS-induced cell apoptosis and inflammation in rat intestine epithelial cells by inhibiting the mTOR/STAT3 pathway. Biomedicine & Pharmacotherapy, 2018. 104: p. 622-628.
- Liu, L. , et al., Curcumin ameliorates dextran sulfate sodium-induced experimental colitis by blocking STAT3 signaling pathway. International immunopharmacology, 2013. 17(2): p. 314-320.
- An, H.-J. , et al., STAT3/NF-κB decoy oligodeoxynucleotides inhibit atherosclerosis through regulation of the STAT/NF-κB signaling pathway in a mouse model of atherosclerosis. International Journal of Molecular Medicine, 2023. 51(5): p. 1-11.
- Liu, X. , et al., LPS-induced proinflammatory cytokine expression in human airway epithelial cells and macrophages via NF-κB, STAT3 or AP-1 activation. Molecular medicine reports, 2018. 17(4): p. 5484-5491.
- Basu, A. , et al., STAT3 and NF-κB are common targets for kaempferol-mediated attenuation of COX-2 expression in IL-6-induced macrophages and carrageenan-induced mouse paw edema. Biochemistry and biophysics reports, 2017. 12: p. 54-61.
- Seo, Y.-J. , et al., Isocyperol, isolated from the rhizomes of Cyperus rotundus, inhibits LPS-induced inflammatory responses via suppression of the NF-κB and STAT3 pathways and ROS stress in LPS-stimulated RAW 264.7 cells. International immunopharmacology, 2016. 38: p. 61-69.
- Li, X. , et al., Sorafenib inhibits LPS-induced inflammation by regulating Lyn-MAPK-NF-kB/AP-1 pathway and TLR4 expression. Cell Death Discovery, 2022. 8(1): p. 281.
- Han, Y.-H. , et al., Anti-inflammatory effect of hispidin on LPS induced macrophage inflammation through MAPK and JAK1/STAT3 signaling pathways. Applied Biological Chemistry, 2020. 63: p. 1-9.
- Linghu, K.-G. , et al., Comprehensive comparison on the anti-inflammatory effects of three species of Sigesbeckia plants based on NF-κB and MAPKs signal pathways in vitro. Journal of ethnopharmacology, 2020. 250: p. 112530.
- Li, R., P. Hong, and X. Zheng, β-Carotene attenuates lipopolysaccharide-induced inflammation via inhibition of the NF-κB, JAK2/STAT3 and JNK/p38 MAPK signaling pathways in macrophages. Animal Science Journal, 2019. 90(1): p. 140-148.
- Yu, Q. , et al., Resokaempferol-mediated anti-inflammatory effects on activated macrophages via the inhibition of JAK2/STAT3, NF-κB and JNK/p38 MAPK signaling pathways. International immunopharmacology, 2016. 38: p. 104-114.
- Guak, H. and C.M. Krawczyk, Implications of cellular metabolism for immune cell migration. Immunology, 2020. 161(3): p. 200-208.
- Cui, S. , et al., Quercetin inhibits LPS-induced macrophage migration by suppressing the iNOS/FAK/paxillin pathway and modulating the cytoskeleton. Cell Adhesion & Migration, 2019. 13(1): p. 1-12.
- Chen, Y.-C. , et al., Morus alba and active compound oxyresveratrol exert anti-inflammatory activity via inhibition of leukocyte migration involving MEK/ERK signaling. BMC Complementary and Alternative Medicine, 2013. 13: p. 1-10.
- Tripathi, S., D. Bruch, and D.S. Kittur, Ginger extract inhibits LPS induced macrophage activation and function. BMC complementary and Alternative Medicine, 2008. 8: p. 1-7.
- Charalabopoulos, A. , et al., Apigenin exerts anti-inflammatory effects in an experimental model of acute pancreatitis by down-regulating TNF-α. in vivo, 2019. 33(4): p. 1133-1141.
- Ai, X.-Y. , et al., Apigenin inhibits colonic inflammation and tumorigenesis by suppressing STAT3-NF-κB signaling. Oncotarget, 2017. 8(59): p. 100216.
- Malik, S. , et al., Apigenin ameliorates streptozotocin-induced diabetic nephropathy in rats via MAPK-NF-κB-TNF-α and TGF-β1-MAPK-fibronectin pathways. American Journal of Physiology-Renal Physiology, 2017. 313(2): p. F414-F422.
- Wang, J. , et al., Anti-inflammatory effects of apigenin in lipopolysaccharide-induced inflammatory in acute lung injury by suppressing COX-2 and NF-kB pathway. Inflammation, 2014. 37: p. 2085-2090.
- Zhang, X. , et al., Flavonoid apigenin inhibits lipopolysaccharide-induced inflammatory response through multiple mechanisms in macrophages. PloS one, 2014. 9(9): p. e107072.
- Hämäläinen, M. , et al., Anti-inflammatory effects of flavonoids: Genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-κB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-κB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediators of inflammation, 2007. 2007(1): p. 045673.
- García-Mediavilla, V. , et al., The anti-inflammatory flavones quercetin and kaempferol cause inhibition of inducible nitric oxide synthase, cyclooxygenase-2 and reactive C-protein, and down-regulation of the nuclear factor kappaB pathway in Chang Liver cells. European journal of pharmacology, 2007. 557(2-3): p. 221-229.
- Singh, B. , et al., Protective effect of vanillic acid against diabetes and diabetic nephropathy by attenuating oxidative stress and upregulation of NF-κB, TNF-α and COX-2 proteins in rats. Phytotherapy Research, 2022. 36(3): p. 1338-1352.
- Huang, X. , et al., Vanillic acid attenuates cartilage degeneration by regulating the MAPK and PI3K/AKT/NF-κB pathways. European journal of pharmacology, 2019. 859: p. 172481.
- Calixto-Campos, C. , et al., Vanillic acid inhibits inflammatory pain by inhibiting neutrophil recruitment, oxidative stress, cytokine production, and NFκB activation in mice. Journal of natural products, 2015. 78(8): p. 1799-1808.
- Kim, M.-C. , et al., Vanillic acid inhibits inflammatory mediators by suppressing NF-κB in lipopolysaccharide-stimulated mouse peritoneal macrophages. Immunopharmacology and immunotoxicology, 2011. 33(3): p. 525-532.
- Ferreira, J.C. , et al., Baccharin and p-coumaric acid from green propolis mitigate inflammation by modulating the production of cytokines and eicosanoids. Journal of ethnopharmacology, 2021. 278: p. 114255.
- Kheiry, M. , et al., p-Coumaric acid attenuates lipopolysaccharide-induced lung inflammation in rats by scavenging ROS production: an in vivo and in vitro study. Inflammation, 2019. 42: p. 1939-1950.
- Zhu, H. , et al., Anti-inflammatory effects of p-coumaric acid, a natural compound of Oldenlandia diffusa, on arthritis model rats. Evidence-Based Complementary and Alternative Medicine, 2018. 2018(1): p. 5198594.
- Pragasam, S.J., V. Venkatesan, and M. Rasool, Immunomodulatory and anti-inflammatory effect of p-coumaric acid, a common dietary polyphenol on experimental inflammation in rats. Inflammation, 2013. 36: p. 169-176.
Figure 1.
Effect of Halodule uninervis ethanolic extract on the viability of RAW264.7 cells. Cells were treated with the vehicle control or with the indicated concentrations of HUE for 24 h, and cell viability was examined using the metabolic-dye-based MTT assay. Data represent the mean ± SEM of three independent experiments (n = 3) and are expressed as a percentage of the corresponding control cells. (*p < 0.05).
Figure 1.
Effect of Halodule uninervis ethanolic extract on the viability of RAW264.7 cells. Cells were treated with the vehicle control or with the indicated concentrations of HUE for 24 h, and cell viability was examined using the metabolic-dye-based MTT assay. Data represent the mean ± SEM of three independent experiments (n = 3) and are expressed as a percentage of the corresponding control cells. (*p < 0.05).
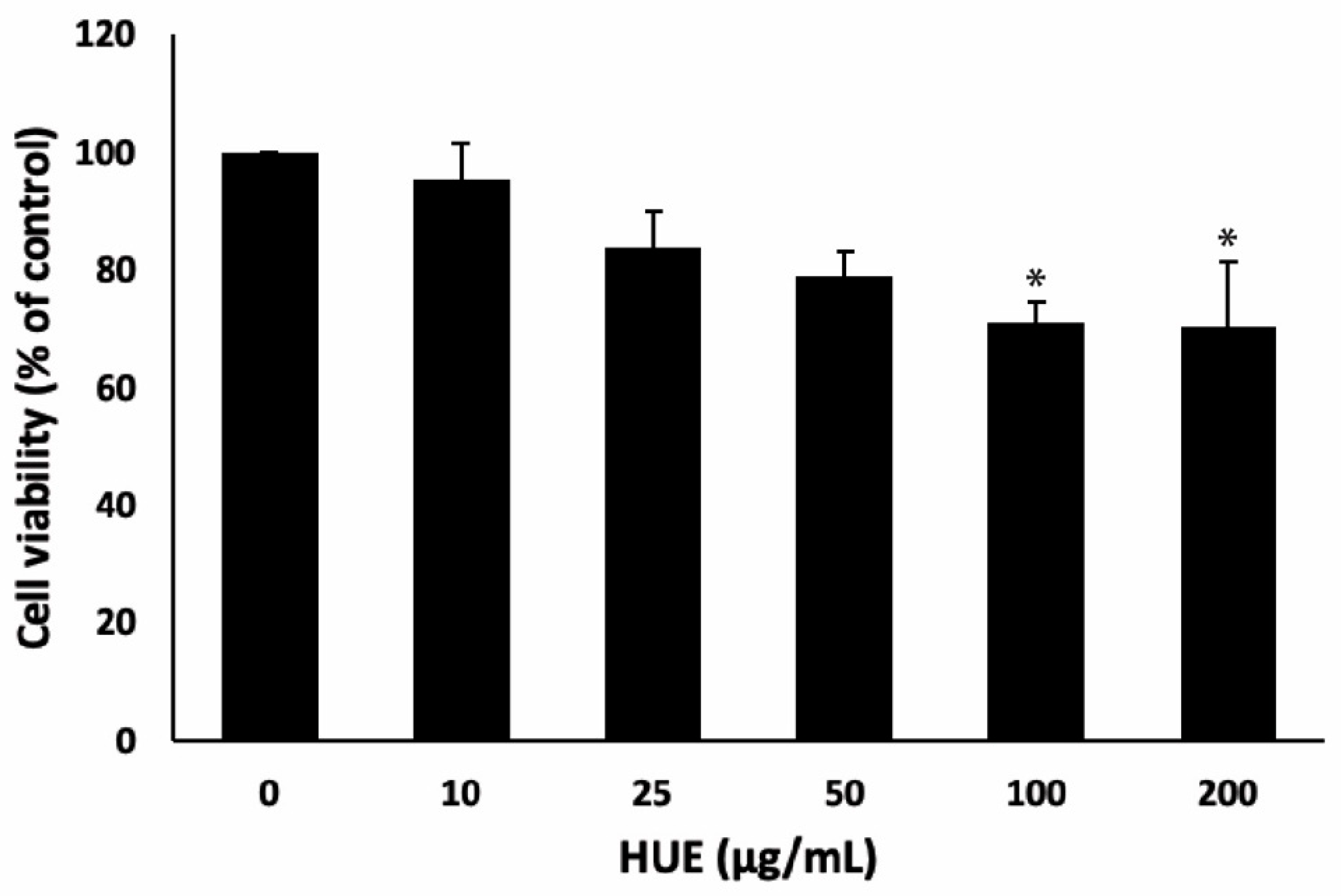
Figure 2.
Halodule uninervis ethanolic extract inhibits iNOS and COX-2. RAW264.7 cells were stimulated with LPS (1 μg/mL) for 30 min and the treated either with vehicle-containing control or HUE (100 or 200 μg/mL) for 18 h. (A) The protein levels of iNOS were assessed via Western blotting, using β-actin as a loading control. (B) The protein levels and mRBA levels of COX-2 were assessed via Western blotting, using β-actin as a loading control, and RT-PCR, respectively. Data represent the mean ± SEM of three independent experiments (n = 3). (* p < 0.05 and ** p < 0.005).
Figure 2.
Halodule uninervis ethanolic extract inhibits iNOS and COX-2. RAW264.7 cells were stimulated with LPS (1 μg/mL) for 30 min and the treated either with vehicle-containing control or HUE (100 or 200 μg/mL) for 18 h. (A) The protein levels of iNOS were assessed via Western blotting, using β-actin as a loading control. (B) The protein levels and mRBA levels of COX-2 were assessed via Western blotting, using β-actin as a loading control, and RT-PCR, respectively. Data represent the mean ± SEM of three independent experiments (n = 3). (* p < 0.05 and ** p < 0.005).

Figure 3.
Halodule uninervis ethanolic extract inhibits the mRNA expression levels of (A) TNF-α and (B) IL-6. RAW264.7 cells were stimulated with LPS (1 μg/mL) for 30 min and the treated either with vehicle-containing control or HUE (100 or 200 μg/mL) for 18 h. The mRNA levels were measured by RT-PCR. Data represent the mean ± SEM of three independent experiments (n = 3). (*** p < 0.001 and **** p < 0.0001).
Figure 3.
Halodule uninervis ethanolic extract inhibits the mRNA expression levels of (A) TNF-α and (B) IL-6. RAW264.7 cells were stimulated with LPS (1 μg/mL) for 30 min and the treated either with vehicle-containing control or HUE (100 or 200 μg/mL) for 18 h. The mRNA levels were measured by RT-PCR. Data represent the mean ± SEM of three independent experiments (n = 3). (*** p < 0.001 and **** p < 0.0001).

Figure 4.
Halodule uninervis ethanolic extract inhibits the NF-κB signaling pathway. RAW264.7 cells were stimulated with LPS (1 μg/mL) for 30 min and the treated either with vehicle-containing control or HUE (100 or 200 μg/mL) for 18 h. The protein levels of the phosphorylated NF-κB were assessed via Western blotting, using β-actin as a loading control. Data represent the mean ± SEM of three independent experiments (n = 3). (* p < 0.05 and ** p < 0.005).
Figure 4.
Halodule uninervis ethanolic extract inhibits the NF-κB signaling pathway. RAW264.7 cells were stimulated with LPS (1 μg/mL) for 30 min and the treated either with vehicle-containing control or HUE (100 or 200 μg/mL) for 18 h. The protein levels of the phosphorylated NF-κB were assessed via Western blotting, using β-actin as a loading control. Data represent the mean ± SEM of three independent experiments (n = 3). (* p < 0.05 and ** p < 0.005).
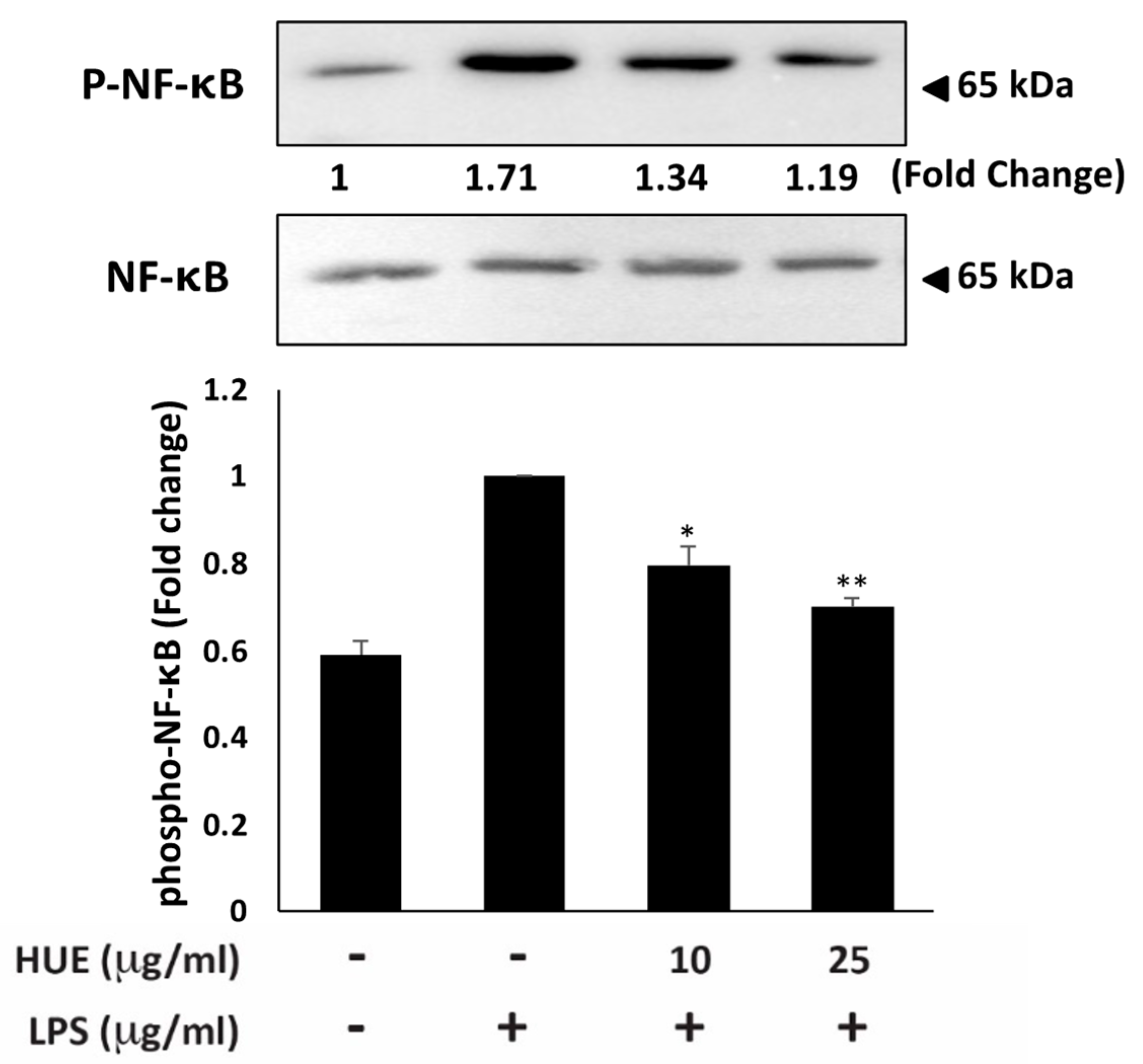
Figure 5.
Halodule uninervis ethanolic extract inhibits the STAT3 signaling pathway. RAW264.7 cells were stimulated with LPS (1 μg/mL) for 30 min and the treated either with vehicle-containing control or HUE (100 or 200 μg/mL) for 18 h. The protein levels of the phosphorylated STAT3 were assessed via Western blotting, using β-actin as a loading control. Data represent the mean ± SEM of three independent experiments (n = 3). (* p < 0.05 and ** p < 0.005).
Figure 5.
Halodule uninervis ethanolic extract inhibits the STAT3 signaling pathway. RAW264.7 cells were stimulated with LPS (1 μg/mL) for 30 min and the treated either with vehicle-containing control or HUE (100 or 200 μg/mL) for 18 h. The protein levels of the phosphorylated STAT3 were assessed via Western blotting, using β-actin as a loading control. Data represent the mean ± SEM of three independent experiments (n = 3). (* p < 0.05 and ** p < 0.005).
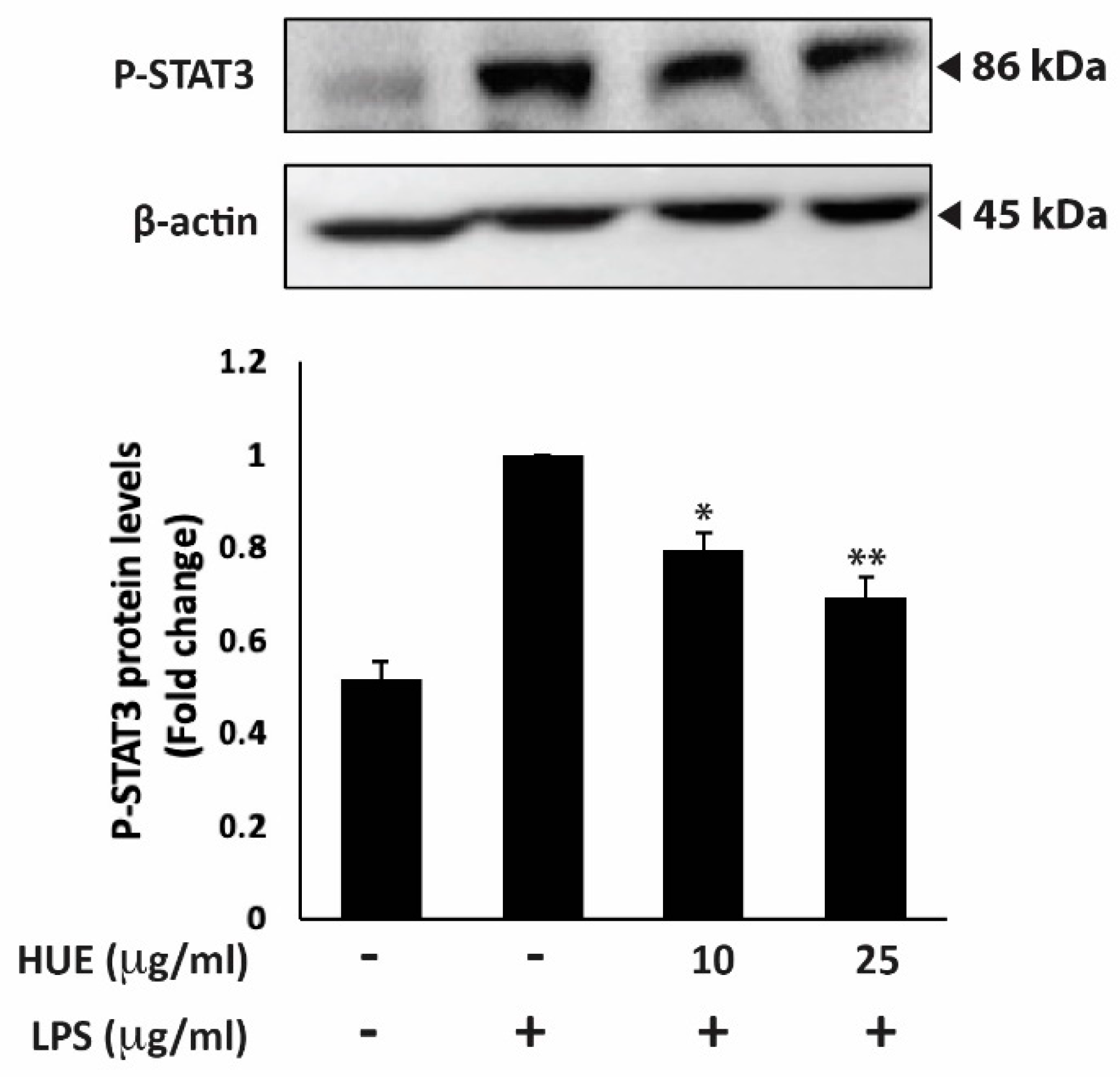
Figure 6.
Halodule uninervis ethanolic extract inhibits the MAPKs signaling pathway. RAW264.7 cells were stimulated with LPS (1 μg/mL) for 30 min and the treated either with vehicle-containing control or HUE (100 or 200 μg/mL) for 18 h. The protein levels of the phosphorylated ERK1/2 and p38 were assessed via Western blotting, using β-actin as a loading control. Data represent the mean ± SEM of three independent experiments (n = 3). (** p < 0.005).
Figure 6.
Halodule uninervis ethanolic extract inhibits the MAPKs signaling pathway. RAW264.7 cells were stimulated with LPS (1 μg/mL) for 30 min and the treated either with vehicle-containing control or HUE (100 or 200 μg/mL) for 18 h. The protein levels of the phosphorylated ERK1/2 and p38 were assessed via Western blotting, using β-actin as a loading control. Data represent the mean ± SEM of three independent experiments (n = 3). (** p < 0.005).
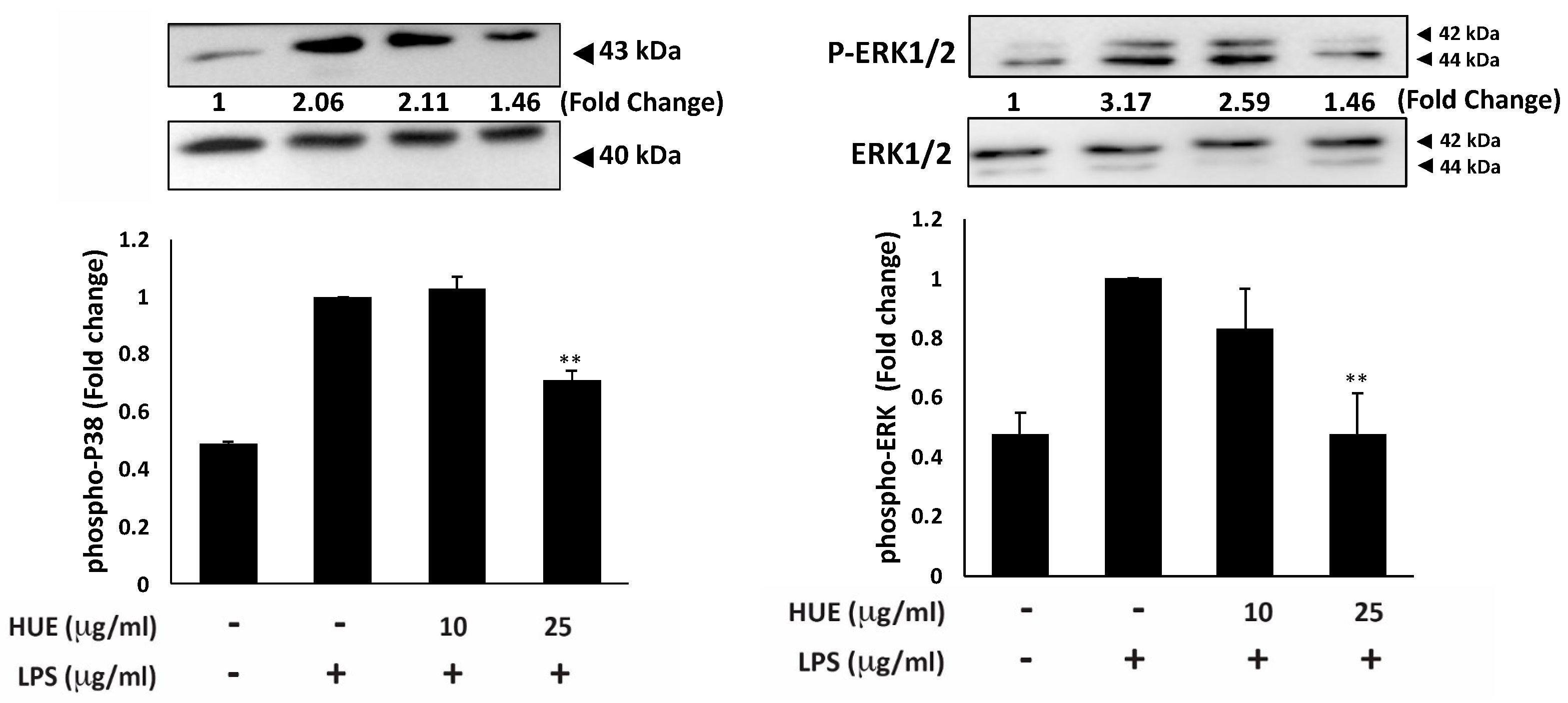
Figure 7.
Halodule uninervis ethanolic extract inhibits the migration of RAW 264.7 macrophages. RAW264.7 cells were stimulated with LPS (1 μg/mL) for 30 min and the treated either with vehicle-containing control or HUE (100 or 200 μg/mL) for 24 h in Boyden chamber trans-well inserts. Cells that migrated to the lower surface of the chamber were DAPI-stained, photographed and 4× magnification, and analyzed. Data represent the mean ± SEM of three independent experiments (n = 3). (**** p < 0.0001).
Figure 7.
Halodule uninervis ethanolic extract inhibits the migration of RAW 264.7 macrophages. RAW264.7 cells were stimulated with LPS (1 μg/mL) for 30 min and the treated either with vehicle-containing control or HUE (100 or 200 μg/mL) for 24 h in Boyden chamber trans-well inserts. Cells that migrated to the lower surface of the chamber were DAPI-stained, photographed and 4× magnification, and analyzed. Data represent the mean ± SEM of three independent experiments (n = 3). (**** p < 0.0001).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



