Submitted:
23 June 2025
Posted:
24 June 2025
You are already at the latest version
Abstract
Background: Diabetes represents a significant global health challenge and poses a serious threat to human wellbeing. 1,25(OH)2D3 (vitamin D, VD) has been recognized for its anti-diabetic effects. Nevertheless, it is still unknown how 1,25-dihydroxyvitamin D3 (1,25D) prevents Type 2 diabetes mellitus (T2DM).MethodsCuCl2 was used to induce the pancreatic β-cell lines MIN6 and EndoC-βH1. To accomplish knockdown, these β-cells were then transfected with siRNA that targeted FDX1. The amount of insulin released was measured using an enzyme-linked immunosorbent assay (ELISA). To assess the cuprotosis process, concentrations of copper (Cu), pyruvate acid (PA), and α-ketoglutarate dehydrogenase (α-KG) were evaluated. Cell viability was also evaluated using an assay called the Cell Counting Kit 8 (CCK-8). The expression of FDX1, vitamin D receptor (VDR), and histone lactylation was measured using western blotting. The enrichment of histone lactylation on FDX1 and FDX1 promoter activity was determined using chromatin immunoprecipitation (ChIP) and luciferase reporter gene assays. ResultsInduction of cuprotosis in pancreatic β-cells significantly suppressed cell viability and insulin secretion, simultaneously reduced α-KG and PA levels, and elevated RNA level of FDX1. VD3 treatment promotes viability and represses cuprotosis in pancreatic β-cells. VD3 treatment up-regulated insulin secretion in MIN6 cells, whereas VDR knockdown abolished this effect. VD3 notably upregulated VDR and downregulated FDX1, which was abolished by VDR knockout (VDR-KO). VD3 reduced histone lactylation of FDX1 and its promoter activity, consequently suppressing FDX1 expression.Conclusion The VD3/VDR axis can alleviate the cuprotosis of pancreatic β-cells by regulating the histone lactylation of FDX1.
Keywords:
Diabetes
; 1
; 25(OH)2D3
; Type 2 diabetes mellitus
; Pancreatic β-cells
; Cuprotosis
Introduction
Metabolic disorders such as obesity and diabetes are frequently associated with the dysregulation of various metal ions [1,2]. Among these, copper plays a crucial role in numerous biological functions, including mitochondrial respiration, anti-aging mechanisms, and the synthesis of biomolecules, and it is closely linked to the severity and progression of diabetes [3,4,5]. Studies indicate that serum copper levels are markedly elevated in individuals with diabetes and in streptozotocin-induced diabetic rat models [6].
The tiny molecules known as copper ionophores, which help move copper into cells, have attracted attention in this regard. In contrast to the actions of small-molecule chaperones, studies have shown that the buildup of copper within cells is the primary mechanism by which these ionophores cause cell death [7]. Notably, investigations revealed that a range of structurally diverse compounds that bind copper exhibit similar cytotoxic profiles across numerous cell lines. Further structure-function analyses indicated that modifications that disrupt the copper-binding capability of these compounds result in diminished cell lethality, while copper chelation abolishes their cytotoxic effects [9,10].
These findings led to the conclusion that cell death induced by copper ionophores is largely driven by the buildup of intracellular copper. The Golub group has coined the term “Cuprotosis” to describe this novel form of cell death associated with copper ionophores [11].
Acetylation, methylation, phosphorylation, ubiquitination, and SUMOylation are among the many changes that occur to histones [12,13,14]. Recently, Zhang et al. identified a novel epigenetic modification known as lactylation, which is derived from lactate and can activate gene transcription [15]. Notably, lactic acid is produced during glycolysis under low-oxygen conditions or in response to bacterial invasion, leading to histone lactylation and the subsequent activation of downstream gene expression [16]. In many clinical diseases, this alteration is an important epigenetic regulator [17,18,19]. For example, in response to microbial signals and other harmful stimuli, macrophages can more easily shift from an inflammatory to a repair state when histone lactylation is elevated at the locus of genes linked to macrophage repair [20]. Furthermore, histone lactylation at pluripotency-related gene loci, induced by Glis1, promotes somatic rearrangements. However, the specific impact of histone lactylation on β-cell function in the context of diabetes is still not well understood [21].
Vitamin D is essential for immunological modulation, cellular differentiation, and proliferation in addition to its functions in controlling calcium, phosphorus, and bone metabolism. By interacting with the vitamin D receptor (VDR), 1,25(OH)2D3 (VD3), the active form of vitamin D, regulates the metabolism of calcium, phosphate, and bone. By forming heterodimers with retinoid X receptors, VDR controls nuclear-level gene expression. In this work, we examined how VD3/VDR affects β-cell cuprotosis.
Materials and Methods
Cell Culture and Treatment
For our studies, we employed the mouse β-cell line MIN6 and the human β-cell line EndoC-βH1. Both cell lines were cultivated in Dulbecco's Modified Eagle's Medium (DMEM, Hyclone, USA) supplemented with 10% fetal bovine serum (Hyclone, USA) and 1% penicillin-streptomycin. The cultures were maintained in a humidified incubator at 37°C with 5% CO2. To induce cuprotosis, the cells were pretreated for 12 hours with elesclomol (5 µM) and CuCl2 (40 µM).
Cell Transfection
Small interfering RNAs (siRNAs) targeting VDR (VDR-KO) were purchased from GenePharma (Shanghai, China). Cells were seeded in 6-well plates to reach 70% confluence. siRNA was then mixed with the transfection reagent Lipofectamine 2000 (Invitrogen, USA) and added to the cell culture medium. Cells were gathered for further research after 48 hours of incubation.
Cell Counting Kit 8 (CCK-8)
The Cell Counting Kit-8 (CCK-8; Solarbio, China) was used to measure the vitality of the cells. After being plated at a density of 1 × 10^5 cells/mL in 96-well plates, the cells were exposed to CuCl2 (40 µM) and VD3 (50 µM) for a whole day. Following the treatment time, each well received 10 µL of CCK-8 reagent, which was then incubated for two hours. A microplate reader (Thermo Fisher Scientific) was then used to measure the absorbance at 450 nm.
Insulin Level Detection
The culture medium of the β-cells was collected and analyzed by enzyme-linked immunosorbent assay (ELISA) using human and mouse insulin ELISA kits (Beyotime, China).
Copper Ion Detection
Intracellular levels of Cu (II) were detected using a Cu detection kit (Elabscience, China) following the manufacturer’s protocol.
Measurement of a-Ketoglutarate Dehydrogenase (α-KG) and Pyruvate Acid (PA)
Using a PA detection kit (Abbkine, China) and an Alpha Ketoglutarate (alpha KG) Assay Kit (Abcam, USA), respectively, the production of PA and α-KG was assessed in accordance with the manufacturer's instructions.
RNA Extraction and qPCR
Cells were homogenized with TRIzol reagent (Invitrogen, Carlsbad, CA, USA) to isolate total RNA. Following this, the Prime Script RT Reagent Kit (Takara, Japan) was utilized to quantify the extracted RNA and reverse-transcribe it into complementary DNA (cDNA), adhering strictly to the manufacturer’s guidelines. The expression level of FDX1 RNA was determined through quantitative PCR (qPCR) using the SYBR Mixture Kit (Takara, Japan). Gene expression data were normalized to the internal control, β-actin, and analyzed using the 2−ΔΔCt method for accurate quantification.
Western Blotting Assay
Total protein was extracted from cells utilizing the radioimmunoprecipitation assay (RIPA) lysis buffer. To ensure optimal results, protease and phosphatase inhibitor cocktails (Thermo, USA) were added to the RIPA lysis buffer (Beyotime, China). A BCA Protein Assay Kit (Beyotime, China) was then used to measure the protein concentration. Polyvinylidene fluoride (PVDF) membranes (Merck Millipore, USA) were then loaded with equal amounts of total protein for examination using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The blots were then blocked for one hour using 10% skim milk, and then they were incubated for a whole night at 4°C with primary antibodies against VDR, H3k18la, FDX1, lipid-DLAT, DLAT, lipid-DLST, DLST, H3, and β-actin. Following a PBST wash the following day, the blots were incubated for one hour at room temperature with secondary anti-mouse or anti-rabbit antibodies coupled with horseradish peroxidase. The protein bands were reacted with the ECL reagent (Millipore, USA) and visualized using an imaging system.
Chromatin Immunoprecipitation (ChIP) Assay
As directed by the manufacturer, the ChIP assay was carried out using a ChIP assay kit (Abcam, ab500). To put it briefly, cells were homogenized using lysis buffer, cross-linked in 1% formaldehyde, and then sonicated to produce roughly 500 bp DNA fragments. Protein G beads and anti-H3K18la antibodies were used to immunoprecipitate cell lysates at 4°C for an entire night. After reverse-cross-linking the protein/DNA precipitates, qPCR was used to determine the relative gene expression of FDX1.
Luciferase Reporter Gene Assay
The promoter sequence of FDX1 was synthesized and cloned into the pGL3-basic plasmid. The plasmid was cotransfected with the reference vector pTK plasmid into MIN6 and VDR-knockdown MIN6 cells. Following a 24-hour transfection, the cells were lysed, and the luciferase activity was measured using a Dual Luciferase Reporter system (Promega, USA).
Statistical Analayis
The findings, which came from at least three separate investigations, are shown as mean ± standard deviation (SD). SPSS software (version 19.0; SPSS Inc., Chicago, IL, USA) was used to conduct the statistical analysis. We used the Student's t-test and one-way analysis of variance (ANOVA) to evaluate differences between two or more groups. To ascertain statistical relevance, a significance threshold of P < 0.05 was established.
Results
Cuprotosis in Pancreatic β Cells
First, we evaluated the cuprotosis status of MIN6 and EndoC-βH1 cells. Cells were induced with CuCl2, and the viability of MIN6 and EndoC-βH1 cells was significantly suppressed (Figure 1A), along with reduced insulin secretion (Figure 1B). Consistent with the cuprotosis signature, CuCl2 induced a notable elevation of Cu and FDX1 in cells and reduced α-KG and PA levels (Figure 1C-F). These data indicate that the cuprotosis of pancreatic β-cells is correlated with insulin production.
VD3/VDR Regulates Pancreatic β Cells Cuprotosis
To determine the regulatory role of the VD3/VDR axis in cell cuprotosis, we knocked out the VDR (VDR-KO) and evaluated cell cuprotosis. We observed that treatment with VD3 notably enhanced the viability of parent cells, but not VDR-KO cells (Figure 2A). VD3 treatment upregulated insulin secretion in MIN6 cells, whereas VDR knockdown abolished this effect (Figure 2B). Moreover, VD3 treatment caused a reduction in Cu (Figure 2C) and an increase in α-KG and PA (Figure 2D and E) in MIN6 cells, but not in VDR-KO cells. We also observed notable upregulation of VDR and downregulation of FDX1 under VD3 treatment, which was abolished by VDR-KO (Figure 2F). These data indicate that VDR mediates β-cell cuprotosis following VD3 treatment.
VD3/VDR Axis Regulates FDX1 Expression in Pancreatic β Cells Via Regulating H3K18la Modification
Lactylation is a novel epigenetic regulator that is involved in gene expression. We studied the lactylation of histones in the MIN6 cells. As shown in Figure 3A, treatment with VD3 suppressed H3K18la levels in MIN6 cells and FDX1, lipid-DLAT, and lipid-DLST levels (Figure 3A). Next, we performed a chromatin immunoprecipitation (ChIP) assay to evaluate the lactylation of FDX1. As shown in Figure 3B, the enrichment of H3K18la modification in FDX1 was notably reduced by VD3 treatment, which was not observed in VDR-KO cells (Figure 3B). In addition, VD3 treatment suppressed the activity of the FDX1 promoter in MIN6 cells but not in VDR-KO cells (Figure 3C). These findings indicate that VD3 regulates the lactylation of FDX1 via the VDR.
VD3 Affects Pancreatic β Cell Function Via the Regulating Cuprotosis
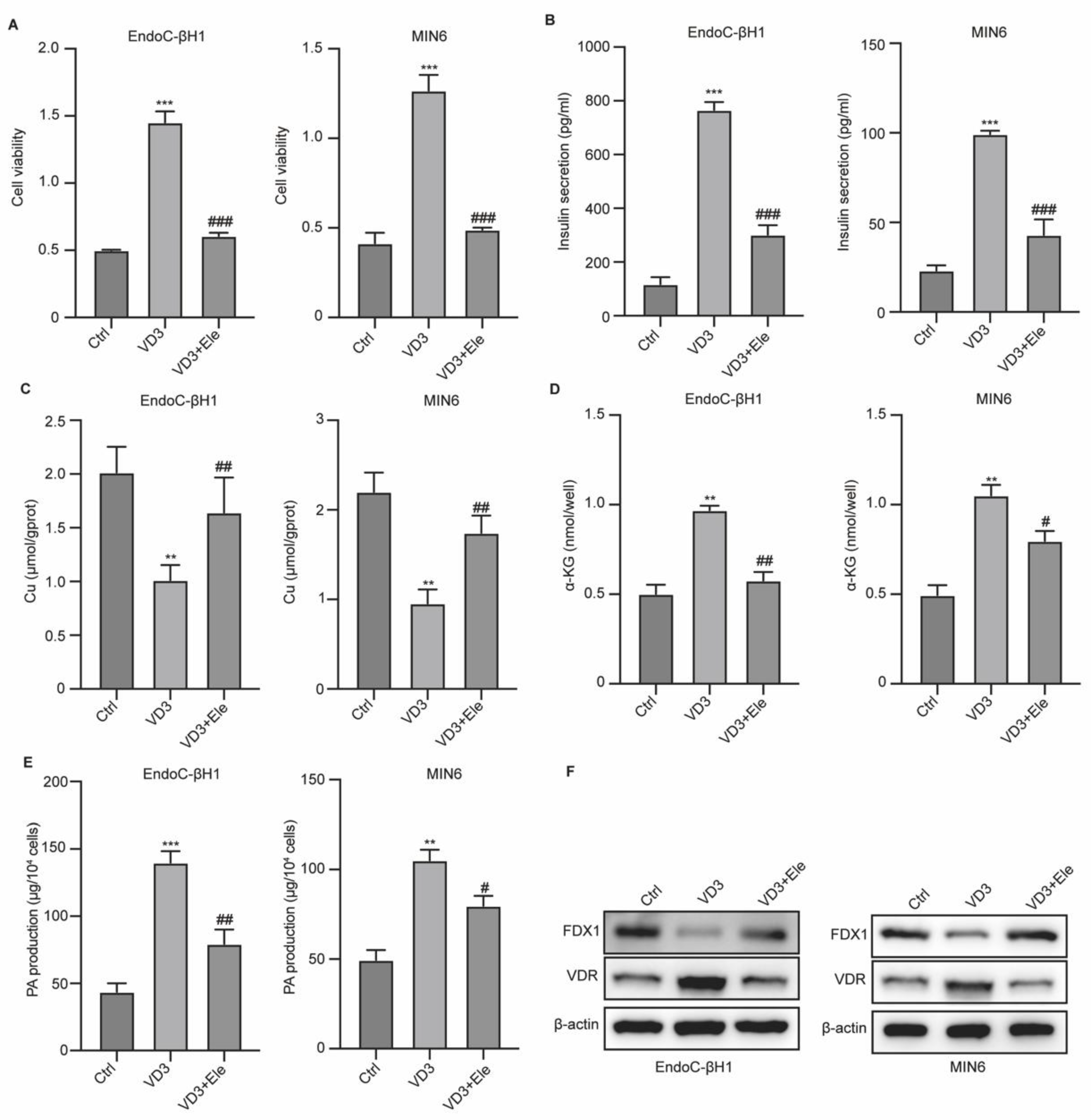
Subsequently, we measured the correlation between VD3, cuprotosis, and β-cell function. We administered VD3 treatment with elesclomol (Ele), a copper chelator, to pancreatic β-cells and evaluated cell viability and cuprotosis. As shown in Figure 4A and B, VD3 treatment notably enhanced the viability and insulin secretion of β-cells, whereas Ele treatment abolished these effects. The Cu level was reduced by VD3 treatment but was reversed by Ele treatment (Figure 4C). Moreover, enhanced production of α-KG and PA after VD3 treatment was abolished by Ele treatment (Figure 4D and E). The decrease in FDX1 and increase in VDR by VD3 in β-cells were also reversed by Ele treatment (Figure 4F). These data indicated that VD3 affects pancreatic β-cell function by regulating cuprotosis.
Discussion
In the current study, we investigated the effects of VD3 on pancreatic β-cell cuprotosis and its underlying mechanisms. We observed increased insulin secretion and reduced Cu levels in pancreatic β cells, as well as accumulated PA and α-KG under VD3 treatment, which indicated that VD3 alleviated cuprotosis. Knockdown of VDR, the receptor for VD3, abolished the effects of VD3 on cuprotosis. Further mechanistic studies revealed reduced H3K18la modification of the FDX1 gene after VD3 treatment, and treatment with the copper chelator Ele blocked the anti-cuprotosis effects of VD3.
The chronic metabolic disease known as type 2 diabetes mellitus (T2DM) has a major impact on people's health. Insulin resistance, insufficient insulin production, and pancreatic islet dysfunction are among the multifactorial elements that are fundamental to its etiology [22]. Numerous studies have found a relationship between vitamin D levels and pancreatic β-cell activity, and higher vitamin D levels have been associated with better glucose metabolism and insulin secretion [23]. Vitamin D (VD) and its derivatives have garnered attention for their potential therapeutic effects on diabetes, owing to their effects on pancreatic β-cell function, insulin sensitivity, pancreatic islet cell integrity, and immune regulation [24,25,26]. VD3 is an active form of VD that binds to VDR expressed on a variety of target cells, including β cells in the pancreas, to produce its biological effects. When VDR and the retinoid X receptor (RXR) bind together, they produce a complex that moves into the nucleus and functions as a transcription factor that alters the expression of target genes involved in different cellular functions [27]. Existing evidence suggests that VD exerts beneficial effects on various pathological aspects of diabetes, including bone loss, cardiac dysfunction, and impaired osteogenesis [28,29,30]. Numerous mechanisms, including autophagy control, critical signaling pathway regulation, and contact with certain receptors, underlie these effects [28,29,30]. Nevertheless, it is still unknown how VD3 prevents pancreatic β-cell cuprotosis in diabetes. Cuprotosis is a novel copper-dependent form of cell death that has been observed in multiple diseases such as neurological diseases, cardiovascular diseases, and cancers, and is regulated by FDX1 [31,32,33,34]. For instance, elesclomol-induced cuprotosis markedly inhibited thyroid cancer cell growth in vitro and in vivo while concurrently increasing Cu levels and FDX1 expression; these effects were eliminated when FDX1 was depleted [35]. Wang et al. reported that a signature containing six cuprotosis-related genes could accurately predict the prognosis of clear cell carcinoma [32]. In the present study, we demonstrated that elesclomol treatment could induce pancreatic β-cell cuprotosis and that VD3 treatment repressed β-cell cuprotosis. Further studies indicated that VD3 reduced lactylation of the FDX1 gene, which mediated pancreatic β-cell cuprotosis. Lactylation is a recently identified epigenetic regulation of gene expression involved in multiple diseases. For example, histone lactylation acts as a novel epigenetic code that primes cells toward a malignant state [36]. By encouraging reparative gene transcription, histone lactylation regulates the dual activity of monocyte-macrophages, balancing their pro-angiogenic and anti-inflammatory properties. After myocardial infarction, our research shown that histone lactylation improves heart function and fosters a healing environment [37]. Our study identified the direct histone lactylation of FDX1. VD3 treatment reduced histone lactylation and promoter activity of FDX1 in pancreatic β-cells, and VDR knockdown interrupted these effects.
Conclusion
In summary, we identified that the VD3/VDR axis could alleviate the cuprotosis of pancreatic β-cells by regulating the histone lactylation of FDX1. Our findings provide novel insights into the pathogenesis of diabetes and identify the correlation between VD3 and FDX1 lactylation and cuprotosis in diabetes.
Funding
Huai 'an Science and Technology Plan Project (Municipal Basic Research Plan) Project No. HAB202302. Xuzhou Medical University Affiliated Hospital Science and Technology Development Fund project (face project) Project number XYFM202310.
References
- Chen, T., et al., Ferroptosis and cuproptposis in kidney Diseases: dysfunction of cell metabolism. Apoptosis, 2024. 29(3-4): p. 289-302. [CrossRef]
- Fu, Z. and S. Xi, The effects of heavy metals on human metabolism. Toxicol Mech Methods, 2020. 30(3): p. 167-176. [CrossRef]
- Kornblatt, A.P., V.G. Nicoletti, and A. Travaglia, The neglected role of copper ions in wound healing. J Inorg Biochem, 2016. 161: p. 1-8. [CrossRef]
- Zhang, Z., et al., Copper incorporated biomaterial-based technologies for multifunctional wound repair. Theranostics, 2024. 14(2): p. 547-570. [CrossRef]
- Cui, L., et al., Mitochondrial copper depletion suppresses triple-negative breast cancer in mice. Nat Biotechnol, 2021. 39(3): p. 357-367. [CrossRef]
- Qiu, Q., et al., Copper in Diabetes Mellitus: a Meta-Analysis and Systematic Review of Plasma and Serum Studies. Biol Trace Elem Res, 2017. 177(1): p. 53-63. [CrossRef]
- O'Brien, H., T. Davoodian, and M.D.L. Johnson, The promise of copper ionophores as antimicrobials. Curr Opin Microbiol, 2023. 75: p. 102355. [CrossRef]
- Huo, S., et al., ATF3/SPI1/SLC31A1 Signaling Promotes Cuproptosis Induced by Advanced Glycosylation End Products in Diabetic Myocardial Injury. Int J Mol Sci, 2023. 24(2). [CrossRef]
- Xie, J., et al., Cuproptosis: mechanisms and links with cancers. Mol Cancer, 2023. 22(1): p. 46. [CrossRef]
- Guo, B., et al., Cuproptosis Induced by ROS Responsive Nanoparticles with Elesclomol and Copper Combined with αPD-L1 for Enhanced Cancer Immunotherapy. Adv Mater, 2023. 35(22): p. e2212267. [CrossRef]
- Tsvetkov, P., et al., Copper induces cell death by targeting lipoylated TCA cycle proteins. Science, 2022. 375(6586): p. 1254-1261. [CrossRef]
- He, Y., et al., Melatonin ameliorates histone modification disorders in mammalian aged oocytes by neutralizing the alkylation of HDAC1. Free Radic Biol Med, 2023. 208: p. 361-370. [CrossRef]
- Millán-Zambrano, G., et al., Histone post-translational modifications - cause and consequence of genome function. Nat Rev Genet, 2022. 23(9): p. 563-580. [CrossRef]
- Zaib, S., N. Rana, and I. Khan, Histone Modifications and their Role in Epigenetics of Cancer. Curr Med Chem, 2022. 29(14): p. 2399-2411. 10.2174/0929867328666211108105214.
- Zhang, D., et al., Metabolic regulation of gene expression by histone lactylation. Nature, 2019. 574(7779): p. 575-580. [CrossRef]
- Li, X., et al., Lactate metabolism in human health and disease. Signal Transduct Target Ther, 2022. 7(1): p. 305. [CrossRef]
- Chen, A.N., et al., Lactylation, a Novel Metabolic Reprogramming Code: Current Status and Prospects. Front Immunol, 2021. 12: p. 688910. [CrossRef]
- Pan, R.Y., et al., Positive feedback regulation of microglial glucose metabolism by histone H4 lysine 12 lactylation in Alzheimer's disease. Cell Metab, 2022. 34(4): p. 634-648.e6. [CrossRef]
- Yu, J., et al., Histone lactylation drives oncogenesis by facilitating m(6)A reader protein YTHDF2 expression in ocular melanoma. Genome Biol, 2021. 22(1): p. 85. [CrossRef]
- Shi, W., et al., Lactic acid induces transcriptional repression of macrophage inflammatory response via histone acetylation. Cell Rep, 2024. 43(2): p. 113746. [CrossRef]
- Li, L., et al., Glis1 facilitates induction of pluripotency via an epigenome-metabolome-epigenome signalling cascade. Nat Metab, 2020. 2(9): p. 882-892. [CrossRef]
- Wu, Y., et al., Risk factors contributing to type 2 diabetes and recent advances in the treatment and prevention. Int J Med Sci, 2014. 11(11): p. 1185-200. [CrossRef]
- Habibian, N., et al., Role of vitamin D and vitamin D receptor gene polymorphisms on residual beta cell function in children with type 1 diabetes mellitus. Pharmacol Rep, 2019. 71(2): p. 282-288. [CrossRef]
- Lips, P., et al., Vitamin D and type 2 diabetes. J Steroid Biochem Mol Biol, 2017. 173: p. 280-285.
- Wu, M., et al., Vitamin D protects against high glucose-induced pancreatic β-cell dysfunction via AMPK-NLRP3 inflammasome pathway. Mol Cell Endocrinol, 2022. 547: p. 111596. [CrossRef]
- Yan, X., et al., Combination therapy with saxagliptin and vitamin D for the preservation of β-cell function in adult-onset type 1 diabetes: a multi-center, randomized, controlled trial. Signal Transduct Target Ther, 2023. 8(1): p. 158.
- Hu, X., et al., 1,25-(OH)2D3 protects pancreatic beta cells against H2O2-induced apoptosis through inhibiting the PERK-ATF4-CHOP pathway. Acta Biochim Biophys Sin (Shanghai), 2021. 53(1): p. 46-53. [CrossRef]
- Jiang, Y., et al., 1α,25-Dihydroxyvitamin D3 ameliorates diabetes-induced bone loss by attenuating FoxO1-mediated autophagy. J Biol Chem, 2021. 296: p. 100287. [CrossRef]
- Guo, X., et al., 1,25-Dihydroxyvitamin D attenuates diabetic cardiac autophagy and damage by vitamin D receptor-mediated suppression of FoxO1 translocation. J Nutr Biochem, 2020. 80: p. 108380. [CrossRef]
- Luo, W., et al., 1ɑ,25-Dihydroxyvitamin D(3) promotes osteogenesis by down-regulating FGF23 in diabetic mice. J Cell Mol Med, 2021. 25(8): p. 4148-4156.
- Wang, Q., Five cuprotosis-related lncRNA signatures for prognosis prediction in acute myeloid leukaemia. Hematology, 2023. 28(1): p. 2231737. [CrossRef]
- Zhang, F., et al., Cuprotosis-related signature predicts overall survival in clear cell renal cell carcinoma. Front Cell Dev Biol, 2022. 10: p. 922995. [CrossRef]
- Zhao, S., et al., Machine learning-based characterization of cuprotosis-related biomarkers and immune infiltration in Parkinson's disease. Front Genet, 2022. 13: p. 1010361. [CrossRef]
- Zhu, W., et al., Cuprotosis clusters predict prognosis and immunotherapy response in low-grade glioma. Apoptosis, 2024. 29(1-2): p. 169-190. [CrossRef]
- Chen, G., et al., FDX1 inhibits thyroid cancer malignant progression by inducing cuprotosis. Heliyon, 2023. 9(8): p. e18655. [CrossRef]
- Lv, X., Y. Lv, and X. Dai, Lactate, histone lactylation and cancer hallmarks. Expert Rev Mol Med, 2023. 25: p. e7. [CrossRef]
- Wang, N., et al., Histone Lactylation Boosts Reparative Gene Activation Post-Myocardial Infarction. Circ Res, 2022. 131(11): p. 893-908. [CrossRef]
Figure 1.
Cuprotosis in pancreatic β cells. (A) The viability was measured by CCK-8. (B) Insulin level in cells. (C-E) The levels of cuprotosis features Cu, α-KG, and PA. (F) The RNA level of FDX1 in cells. **p<0.01 and ***p<0.001 vs Ctrl group.
Figure 1.
Cuprotosis in pancreatic β cells. (A) The viability was measured by CCK-8. (B) Insulin level in cells. (C-E) The levels of cuprotosis features Cu, α-KG, and PA. (F) The RNA level of FDX1 in cells. **p<0.01 and ***p<0.001 vs Ctrl group.
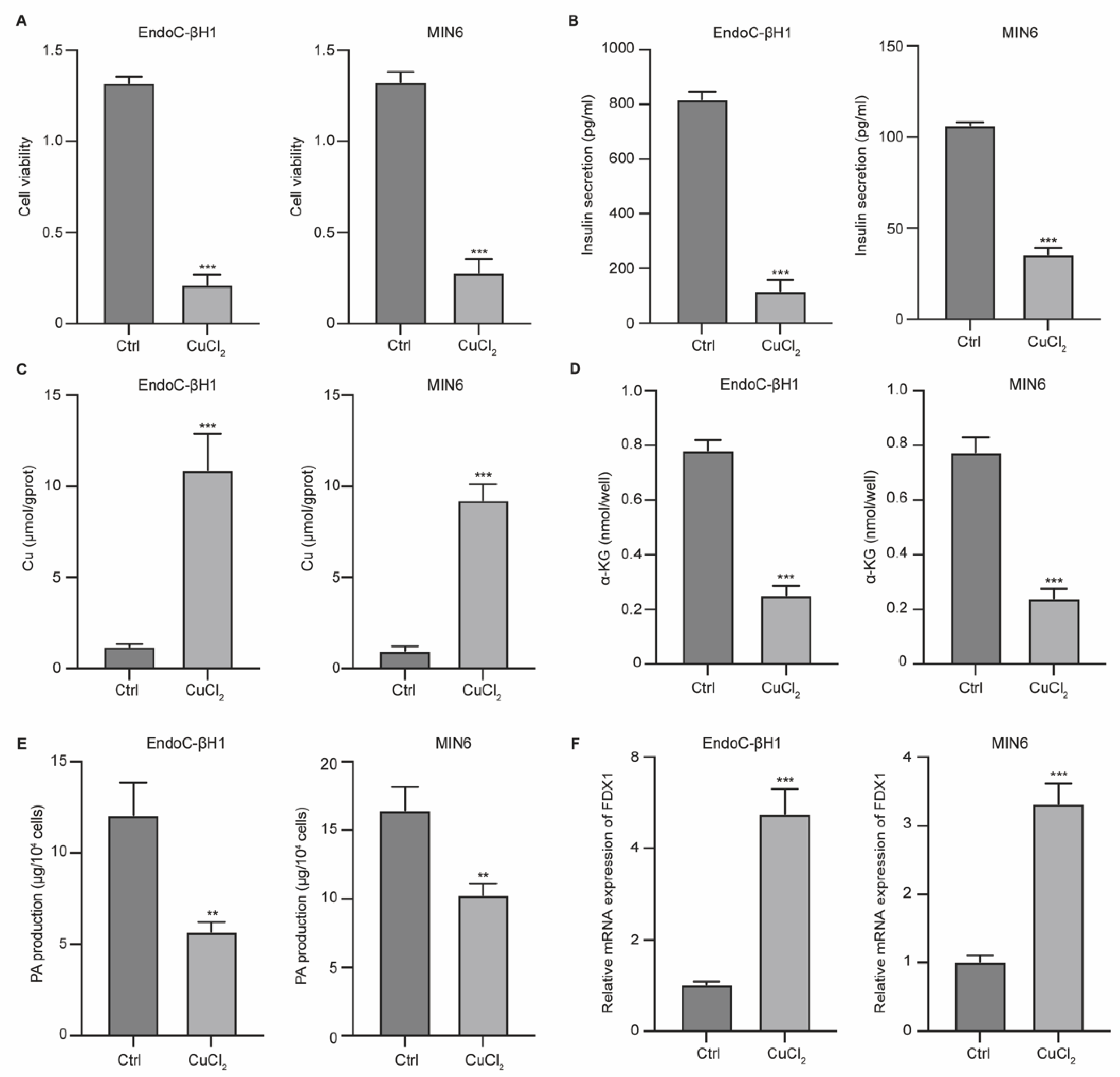
Figure 2.
VD3/VDR regulates pancreatic β cells cuprotosis. MIN6 cells were transfected with siVDR. (A) Cell viability was measured by CCK-8. (B) Production of insulin was measured by ELISA. (C-E) The levels of cuprotosis features Cu, α-KG, and PA in MIN6 and VDR-KO MIN6 cells. (F) Protein levels of FDX1 and VDR in cells was measured by western blotting assay. **p<0.01 and ***p<0.001 vs Ctrl group.
Figure 2.
VD3/VDR regulates pancreatic β cells cuprotosis. MIN6 cells were transfected with siVDR. (A) Cell viability was measured by CCK-8. (B) Production of insulin was measured by ELISA. (C-E) The levels of cuprotosis features Cu, α-KG, and PA in MIN6 and VDR-KO MIN6 cells. (F) Protein levels of FDX1 and VDR in cells was measured by western blotting assay. **p<0.01 and ***p<0.001 vs Ctrl group.
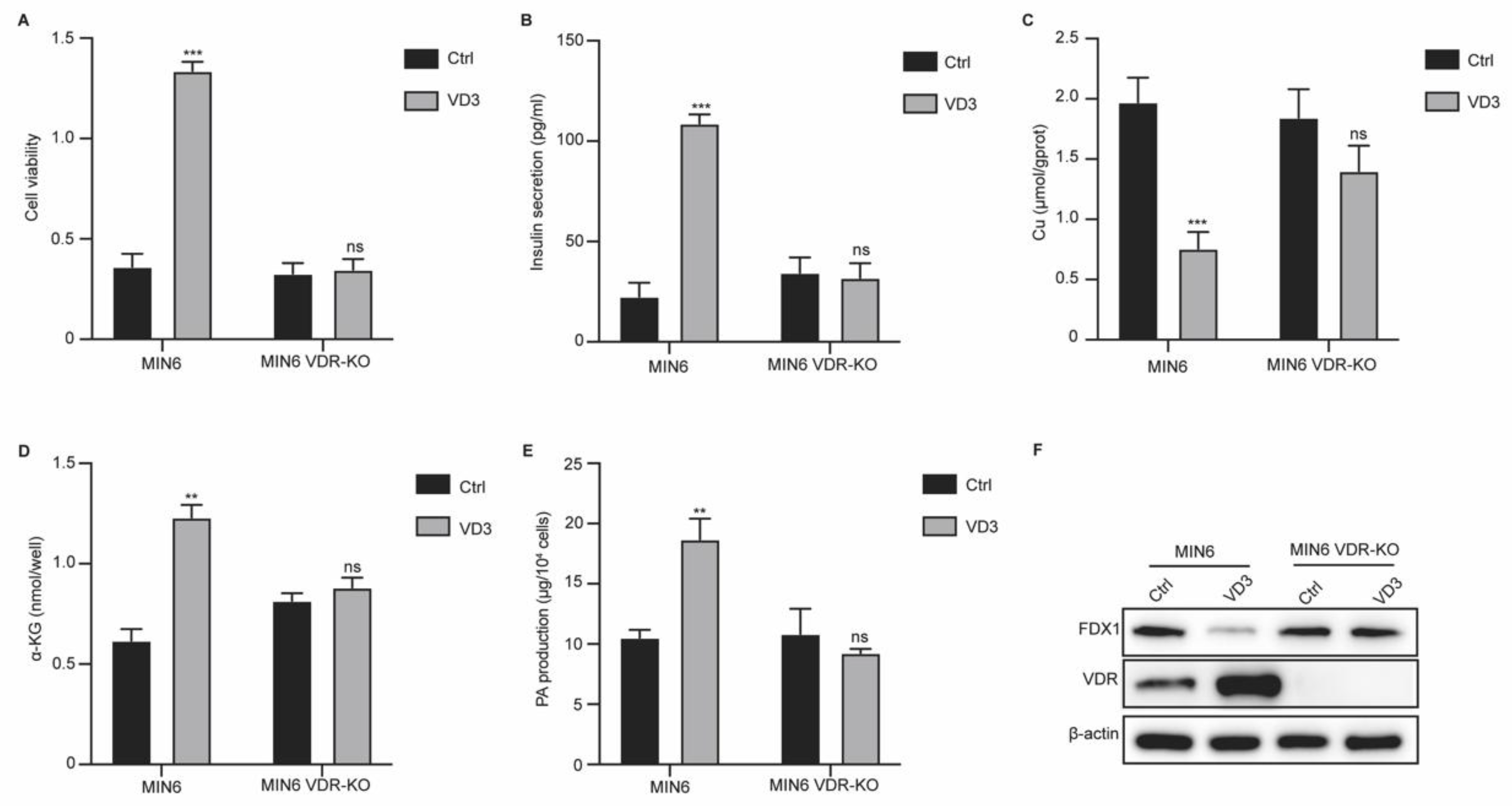
Figure 3.
VD3/VDR axis regulates FDX1 expression in pancreatic β cells via regulating H3K18la modification. (A) Protein levels of H3K18la, FDX1, lipid-DLAT, lipid-DLST, DLAT, and DLST were measured by western blotting assay. (B) Enrichment of H3K18la on FDX1 gene was measured by ChIP assay. (C) The activity of FDX1 promoter was measured by luciferase reporter gene assay.
Figure 3.
VD3/VDR axis regulates FDX1 expression in pancreatic β cells via regulating H3K18la modification. (A) Protein levels of H3K18la, FDX1, lipid-DLAT, lipid-DLST, DLAT, and DLST were measured by western blotting assay. (B) Enrichment of H3K18la on FDX1 gene was measured by ChIP assay. (C) The activity of FDX1 promoter was measured by luciferase reporter gene assay.
Figure 4.
VD3 affects pancreatic β cell function via the regulating cuprotosis. Cells were treated with VD3 and elesclomol (Ele). (A) Cell viability was measured by CCK-8. (B) Insulin level in cells. (C-E) The levels of cuprotosis features Cu, α-KG, and PA. (F) The protein levels of FDX1 and VDR in cells. **p<0.01 and ***p<0.001 vs Ctrl group, #p<0.05 and ##p<0.01 vs VD3 group.
Figure 4.
VD3 affects pancreatic β cell function via the regulating cuprotosis. Cells were treated with VD3 and elesclomol (Ele). (A) Cell viability was measured by CCK-8. (B) Insulin level in cells. (C-E) The levels of cuprotosis features Cu, α-KG, and PA. (F) The protein levels of FDX1 and VDR in cells. **p<0.01 and ***p<0.001 vs Ctrl group, #p<0.05 and ##p<0.01 vs VD3 group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



