
Submitted:
17 June 2025
Posted:
18 June 2025
You are already at the latest version
Abstract
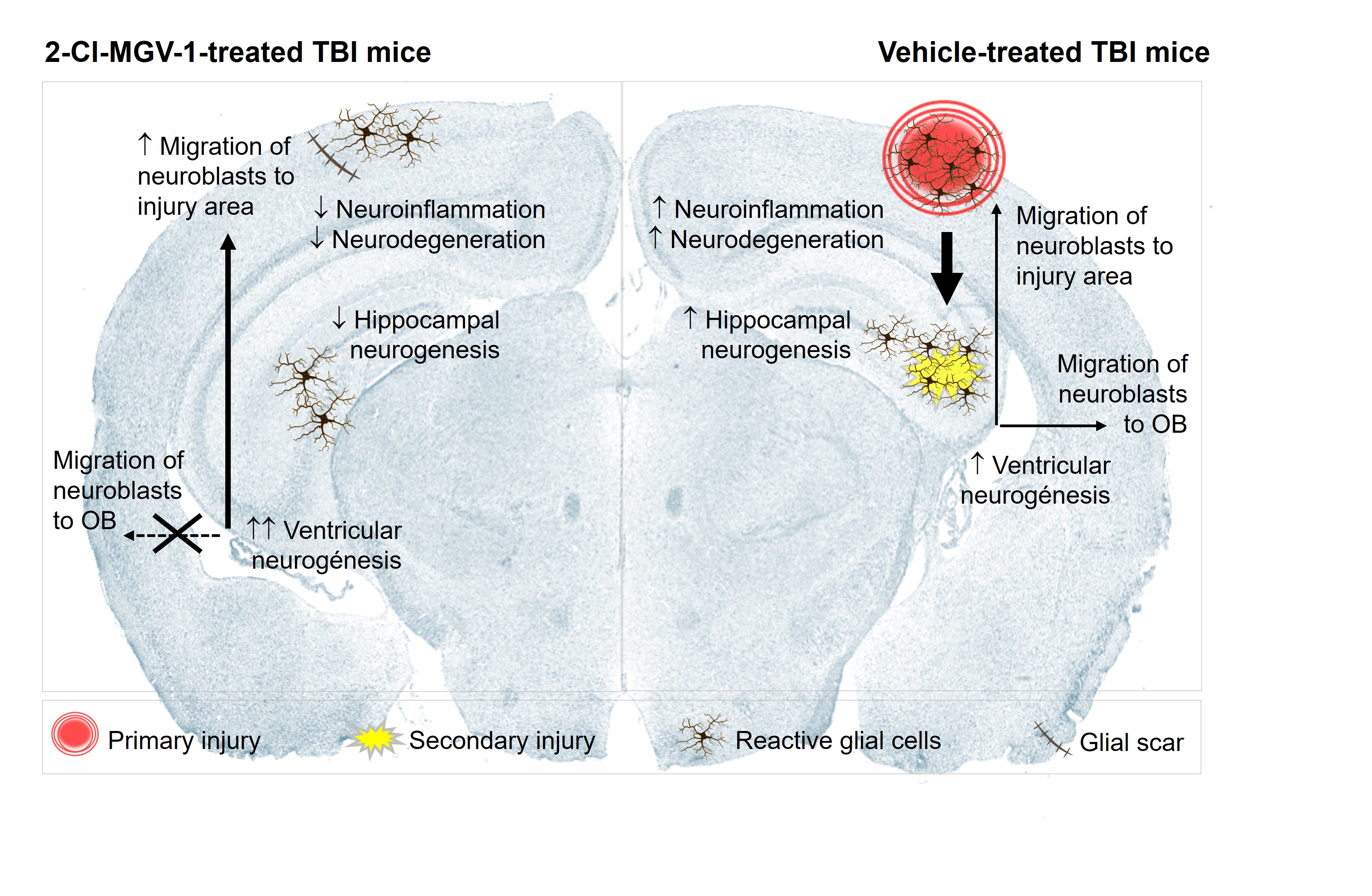
We evaluated the effects of the TSPO ligand 2-Cl-MGV-1 on neurogenesis in a murine model with traumatic brain injury (TBI). Mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days post-TBI showed reduced neuroinflammation and neurodegeneration in the cortex and hippocampus, as indicated by decreased TSPO expression. Notably, 2-Cl-MGV-1 modulated neurogenesis in a niche-specific manner. In the subventricular zone (SVZ), it enhanced neurogenic activity by increasing the expression of neural stem (Nestin) and progenitor (NeuroD1) cell markers, and by redirecting migration of neuroblasts from their canonical path to the olfactory bulb toward the injured cortex. This shift suggests an adaptive reparative response that facilitates brain recovery. In contrast, hippocampal neurogenesis was not enhanced; expression of Nestin, NeuroD1, and β-III-Tubulin in the dentate gyrus was reduced by 2-Cl-MGV-1. Nonetheless, hippocampal neuroprotection was observed, likely due to anti-inflammatory and anti-neurodegenerative effects of the treatment. These results reveal a niche-specific modulation of neurogenesis by 2-Cl-MGV-1, promoting regenerative activity by the ventricular neurogenesis while pre-serving hippocampal integrity through non-neurogenic mechanisms. Altogether, our findings support the therapeutic potential of 2-Cl-MGV-1 to improve brain recovery after TBI by reducing chronic responses post-injury and enhancing endogenous repair mecha-nisms in a neurogenic niche-specific manner.
Keywords:
Neurogenesis
; Neurodegeneration
; Neuroinflammation
; Traumatic Brain Injury
; The 18-kDa mitochondrial translocator protein
; 2-Cl-MGV-1
1. Introduction
TBI remains the leading cause of mortality in individuals under the age of 45, with an estimated incidence of 69 million new cases reported annually worldwide [1,2]. TBI is a serious medical condition resulting from an external force that damages the brain, leading to a broad spectrum of cognitive, physical, behavioral, and emotional impairments. The primary causes of TBI include falls, motor vehicle accidents, sports-related head injuries, and blast-wave exposures [3]. The deleterious effects of TBI extend beyond the primary injury site [3]. The initial mechanical insult -comprising contusion, vascular damage, brain edema, lacerations, hemorrhage, and potential axonal shearing- triggers a cascade of secondary pathological events at the molecular, cellular, and metabolic levels. These secondary injuries exacerbate neurological dysfunction and tissue loss and involved mechanism of excitotoxicity, oxidative stress, inflammation, apoptosis, necrosis, diffuse axonal injury, and progressive neurodegeneration [3]. Notably, a chronic neuroinflammation can perpetuate brain damage, impair recovery, and increase the risk of long-term complications [1]. Indeed, many TBI patients suffer from persistent cognitive deficits, post-traumatic stress disorder and long-term disability [3,4]. Furthermore, TBI significantly increases the risk of developing neurodegenerative diseases [4,5].
The 18-kDa translocator protein (TSPO), located in the outer mitochondrial membrane, has emerged as a valuable biomarker for monitoring neuroinflammation following brain injury [1,6,7,8,9,10,11]. TSPO is implicated in several critical cellular processes, including the generation of reactive oxygen species (ROS), regulation of intracellular calcium (Ca²⁺) homeostasis, ATP production, immune and inflammatory responses, and the regulation of cell death and proliferation [6,7,12,13,14]. A noteworthy role of TSPO in the regulation of nuclear gene expression has also been described [13], which may help explain its broad functional spectrum. Under physiological conditions, TSPO expression in the central nervous system (CNS) is low and mainly restricted to glial cells [15]. However, under pathological conditions involving neurodegeneration or neuroinflammation, TSPO levels are significantly upregulated in glial cells [1,6,7,14]. Consequently, TSPO is widely used in clinical practice as a biomarker for assessing neuroinflammation through positron emission tomography (PET) imaging in a variety of neurological and psychiatric disorders, including TBI [14]. In addition to glial cells, damaged neurons also can overexpress TSPO, particularly in brain regions directly affected by injury or subjected to cellular stress [16,17]. Regarding TBI, clinical studies have also detected elevated TSPO levels in the peripheral blood of patients with TBI, which correlates with increased systemic inflammatory markers such as interleukin-6 (IL-6), tumor necrosis factor-alpha (TNF-α), and C-reactive protein, along with decreased neurological function [10]. Moreover, Bao et al. also reported a strong association between increased TSPO expression in microglial cells and unfavorable clinical outcomes, including severe disability, persistent vegetative state, or death in TBI patients [1].
Accordingly, various TSPO-binding ligands have demonstrated neuroprotective effects by mitigating cell death, reducing inflammatory responses, and attenuating excitotoxicity in both in vitro and in vivo models under diverse toxic and neurodegenerative conditions, including exposure to glutamate, kainic acid, and ammonium chloride [7,13,14,18,19]. Among these, ligands such as PK 11195, Ro5-4864, etifoxine, and 2-Cl-MGV-1 have shown particularly beneficial effects in the context of TBI [1,5,18,20,21]. The neuroprotective actions of 2-Cl-MGV-1 extend beyond TBI models. For instance, 2-Cl-MGV-1 has been reported to promote neuronal recovery in rats subjected to hippocampal damage induced by kainic acid [18]. Its administration has also mitigated neurodegeneration associated with focal infarctions in the rat cortex, contributing to improved cognitive outcomes following injury [22]. In this context, the regenerative potential of the adult brain -particularly its neurogenic capacity- plays a critical role in the recovery of nervous tissue under conditions of neurodegeneration and chronic neuroinflammation [23]. In mammals, two canonical regions of the adult brain are known to generate new neurons: (a) the subventricular zone (SVZ), located along the lateral ventricles, and (b) the subgranular zone (SGZ) of the hippocampal dentate gyrus [24]. Interestingly, several TSPO ligands can induce neuronal differentiation in vitro. For instance, PK 11195 promotes neuronal differentiation in PC12, N1E-115, and P19 cell lines [13,25,26]. Similarly, the TSPO ligand MGV-1 has been shown to induce neuronal differentiation in PC12 cells [18], suggesting a potential role of TSPO modulation in enhancing neuroregeneration.
Taken together, these findings underscore the diagnostic relevance of TSPO as a biomarker of neuroinflammation and highlight the therapeutic potential of its ligands as promising tools to mitigate brain damage and support functional recovery following TBI. In recent preclinical studies using animal models of TBI, early post-injury administration of the TSPO ligand 2-Cl-MGV-1 significantly attenuated neurodegeneration and microgliosis, while enhancing neuronal survival in the injured cortical region. Consequently, treatment with 2-Cl-MGV-1 led to substantial neural tissue regeneration within the injured cortical regions of TBI-affected mice [21]. Building on these findings, the present study aims to determine whether the TSPO ligand 2-Cl-MGV-1 stimulates neurogenesis within the primary neurogenic niches of the injured mouse brain, thereby contributing to neuronal recovery after TBI. By exploring its ability to activate endogenous regenerative pathways, this work seeks to further support the therapeutic value of 2-Cl-MGV-1 in promoting brain repair in the context of TBI.
2. Results
2.1. The TSPO Ligand 2-Cl-MGV-1 Promotes Neural Tissue Regeneration Following TBI
TBI mice treated with vehicle (DMSO) exhibited substantial loss of neural tissue in the injured brain cortex (Figure 1A). Specifically, the primary injury resulted in marked neuronal loss within the gray matter of the left parietal cortex. In contrast, TBI mice treated with 2-Cl-MGV-1 exhibited marked preservation of neural tissue integrity, supporting gray matter recovery in the injured cortex and reducing the extent of damage to a minor scar (Figure 1B). Additionally, TBI affected multiple hippocampal subregions, including the pyramidal cell layers of the CA2 and CA3 areas (Figure 1A), which appeared structurally restored in mice treated with 2-Cl-MGV-1 (Figure 1B). The neuroprotective effect of 2-Cl-MGV-1 was further confirmed in the CA3 region (Figure 1C, D), where neuronal integrity was best preserved, in contrast to the severely compromised CA2 region and injured cortical area. Accordingly, quantitative analysis revealed that neuronal nuclei antigen (NeuN) levels in the hippocampal CA3 region were significantly higher in TBI mice treated with 2-Cl-MGV-1 compared to vehicle-treated mice (p < 0.001; Figure 1E), indicating enhanced neuronal survival. These findings support a robust neuroprotective effect of 2-Cl-MGV-1 in the context of TBI.
2.2. The TSPO Ligand 2-Cl-MGV-1 Reduces Neuroinflammation Following TBI
TBI mice treated with vehicle (DMSO) exhibited severe neuroinflammation, driven by the activation of reactive glia in the damaged brain area following TBI. Consequently, these animals showed high TSPO levels in the injured brain cortex (Figure 2A, C). In contrast, TBI animals treated with 2-Cl-MGV-1 (Figure 2B, D) displayed reduced neuroinflammation, with a significant decrease (p < 0.001) in TSPO levels in the injured cortex compared to vehicle-treated mice (Figure 2E).
2.3. The TSPO Ligand 2-Cl-MGV-1 Enhances Neurogenesis in the SVZ Following TBI
We evaluated different neurogenesis markers (TSPO, Nestin, NeuroD1 and β-Tubulin III) in the SVZ of TBI mice (Figure 3).
Besides being an important marker of reactive glia under conditions of neuroinflammation [1,9,10,11], TSPO is also expressed in pluripotent/multipotent stem cells [27,28,29] and it is considered a repressor of neuronal differentiation [26]. Interestingly, TSPO levels significantly decreased (p < 0.001) in the SVZ of TBI mice treated with 2-Cl-MGV-1 compared to vehicle-treated mice (Figure 3A-C). However, expression of the neural stem cells marker nestin [30], was significantly increased (p < 0.001) in the SVZ of TBI mice treated with 2-Cl-MGV-1 compared to vehicle-treated mice (Figure 3D-F). Similarly, the neurogenic transcription factor NeuroD1, a marker of neural precursor cells or neuroblasts [30], was significantly upregulated (p < 0.001) in the SVZ of 2-Cl-MGV-1-treated TBI mice compared to vehicle-treated mice (Figure 3G-I). The cytoskeletal protein β-Tubulin III, which is expressed early in neuronal precursors [31] and maintained in both postmitotic immature and mature neurons, it also is widely recognized as an early marker of newly generated neurons [30]. β-Tubulin III levels in the SVZ of TBI mice did not show significant differences between 2-Cl-MGV-1-treated and vehicle-treated TBI mice (Figure 3J-L). These findings suggest that 2-Cl-MGV-1 enhances neurogenesis in the SVZ following TBI.
2.4. The TSPO Ligand 2-Cl-MGV-1 Promotes Neuroblasts Migration from the SVZ to the Injured Brain Cortex Following TBI
Neuroblasts are newly generated but still immature neurons that express NeuroD1 and possess migratory capacity, though they lack proliferative ability [32]. We observed high levels of NeuroD1-positive cells in the injured brain cortex of TBI mice treated with vehicle (Figure 4A). However, treatment with 2-Cl-MGV-1 significantly increased (p < 0.01) NeuroD1 levels in the injured cortex compared to vehicle-treated mice (Figure 4B, C). Therefore, 2-Cl-MGV-1 appears to promote the migration of neuroblasts from the SVZ toward the injured cortical region following TBI.
2.5. The TSPO Ligand 2-Cl-MGV-1 Reduces Neuroblasts Migration from the SVZ to the Olfatory Bulb Following TBI
Neuroblasts generated in the SVZ typically migrate to the olfactory bulb in both rodents and humans [32] to give rise to interneurons, primarily in the granule and glomerular cell layers [33,34]. Therefore, we also evaluated the presence of NeuroD1-positive cells in the olfactory bulb of TBI mice (Figure 5).
NeuroD1-positive cells were clearly detected in the mitral and granule cell layers of the olfactory bulb in TBI mice treated with vehicle (Figure 5A), whereas their presence was markedly reduced in TBI mice treated with 2-Cl-MGV-1 (Figure 5B). In both experimental groups, only scarce NeuroD1-positive cells were observed in the glomerular cell layer of the olfactory bulb (Figure 5C, D). Quantitative analysis confirmed a significant decrease in NeuroD1 expression in the mitral and granule cell layers of 2-Cl-MGV-1-treated mice compared to vehicle-treated controls (p < 0.001; Figure 5E). These findings indicate that, in 2-Cl-MGV-1-treated mice, neuroblasts originating from the SVZ are redirected toward the injured cortex rather than migrating to the olfactory bulb.
2.6. The TSPO Ligand 2-Cl-MGV-1 Reduces Hippocampal Neurogenesis Following TBI
We also evaluated different neurogenesis markers (TSPO, Nestin, NeuroD1, and β-Tubulin III) in the dentate gyrus of the hippocampus, including the neurogenic SGZ, in TBI mice (Figure 6).
TSPO levels were significantly decreased (p < 0.001) in the dentate gyrus of 2-Cl-MGV-1-treated mice compared to vehicle-treated mice (Figure 6A–C). Notably, TSPO expression was not restricted to the neurogenic SGZ but was also detected in the granule neuron layer of the dentate gyrus. This widespread distribution likely reflects the intense neuroinflammation present in the hippocampus, which was substantially reduced following 2-Cl-MGV-1 treatment. Similarly, Nestin (Figure 6D-F), NeuroD1 (Figure 6G-I), and β-Tubulin III (Figure 6 J-L) levels significantly decreased (p < 0.001) in the dentate gyrus of TBI mice treated with 2-Cl-MGV-1 compared to vehicle-treated mice. Given that these markers are associated with neural stem/progenitor cells and the neuronal differentiation process [30], their reduction suggests that 2-Cl-MGV-1 may suppress hippocampal neurogenesis.
3. Discussion
Microglial activation drives neuroinflammation and plays a central role in mediating secondary damage following TBI [1,2,3,5,20]. TSPO is a reliable maker of reactive microglia and neuroinflammation that even can activate mitochondrial cell death [6,7,8,14]. Elevated TSPO expression after TBI has been detected in both brain tissue and peripheral blood, and is strongly associated with poor clinical outcomes, including severe disability, vegetative state, or death [1,10]. TSPO ligands are commonly used to quantitatively monitor neuroinflammation after TBI, with increased TSPO expression detectable as early as 2 days post-injury, peaking around day 6, and returning to baseline by day 28 [9,35,36]. Recent studies have reported microglial activation in the brains of mice following TBI [21]. In the present study, we confirmed sustained neuroinflammation and elevated TSPO expression up to 60 days post-injury, which was significantly reduced by treatment with 2-Cl-MGV-1. The potent anti-inflammatory properties of 2-Cl-MGV-1 have already been demonstrated in different studies [37,38], showing even greater efficacy than the classical high-affinity TSPO ligand PK11195 [19,39].
Recent evidence in TBI mice models also revealed key signs of neurodegeneration, including chromatolysis, dark neurons, and neuronal cell death in the injured cortex [21]. In our study, NeuN immunostaining confirmed significant neuronal loss in cortical areas following TBI, along with marked tissue damage extending into the CA2 and CA3 hippocampal regions. Consistent with its anti-inflammatory effects, treatment with 2-Cl-MGV-1 significantly reduced TBI-induced neurodegeneration in these mice [21]. Our recent findings consistently confirm the potent anti-inflammatory and neuroprotective effects of 2-Cl-MGV-1, highlighting its ability to promote recovery of the injured cortex compared to vehicle-treated TBI mice. Notably, 2-Cl-MGV-1 promoted complete neural tissue recovery in the hippocampus of TBI mice, with clear signs of neuronal regeneration reversing the damage observed in vehicle-treated mice. However, recovery of the injured brain cortex was accompanied by the formation of a small but distinct glial scar, marked by elevated TSPO staining. While glial scar formation is known to hinder neuroregeneration [33,40,41], 2-Cl-MGV-1-treated mice still exhibited improved motor performance [21], indicating both structural and functional neural recovery despite the presence of the scar.
Interestingly, as previously reported [16], we also observed increased TSPO expression in injured cortical neurons near the lesion site. This may reflect a cellular stress response, where heightened neuronal activity and mitochondrial demand drive TSPO upregulation [17]. Such elevation is consistent with ongoing neuroinflammation and progressive neurodegeneration persisting up to two months post-injury. In this context, elevated TSPO levels may indicate glial activation, mitochondrial dysfunction and neuronal loss. Importantly, the marked reduction in TSPO expression observed following 2-Cl-MGV-1 treatment suggests that this TSPO ligand may attenuate chronic inflammation and metabolic stress associated with TBI, ultimately promoting neuronal survival and tissue repair. In line with this, Chen and colleagues demonstrated the anti-apoptotic and anti-inflammatory effects of 2-Cl-MGV-1 in the thalamus and hippocampus of rats with focal cortical infarction [22], along with its capacity to improve cognitive function and enhance angiogenesis, particularly in the thalamus [42]. Beyond its anti-inflammatory properties, MGV-1 has been shown to mimic key developmental processes involved in brain repair and regeneration [18]. In a rat model of kainic acid-induced neurotoxicity, MGV-1 prevented seizures, reduced cerebral edema, limited neurodegeneration, and restored normal behavior [18]. Moreover, in the R6/2 mouse model of Huntington’s disease, derivatives of 2-Cl-MGV-1 extended lifespan by over 20% and significantly reduced motor abnormalities [8,18]. Taken together, the present findings support the neuroprotective potential of 2-Cl-MGV-1, acting through modulation of neuroinflammation and neurodegeneration following brain injury.
Neuroinflammation and neurogenesis are closely linked after brain injury. While acute inflammatory responses can stimulate the activation, differentiation, and migration of neural stem cells, chronic inflammation disrupts the neurogenic niche, impairing neuronal maturation and survival [43,44]. Thus, the timing and balance of inflammatory signals are critical for effective brain regeneration [43,44]. Neurogenesis is increasingly recognized as a promising therapeutic strategy for brain repair after TBI [4,40,45,46,47,48]. Newly generated neurons may replace lost cells, restore disrupted neural circuits, support cognitive functions, and improve the injured microenvironment by releasing neurotrophic factors and modulating inflammation [49,50]. The neurogenesis markers analyzed in this study—TSPO, Nestin, NeuroD1, and βIII-Tubulin—indicated activation of neurogenesis in the SVZ of vehicle-treated TBI mice. The presence of NeuroD1-positive cells in the injured cortex and surrounding areas suggests that neuronal precursors migrate from the SVZ toward the lesion site, potentially contributing to brain recovery. This is consistent with previous reports showing that TBI can activate endogenous repair mechanisms, including neurogenesis in the SVZ and subcortical structures such as the striatum and optic tract, which support neuronal regeneration and axonal remyelination [41]. Notably, 2-Cl-MGV-1 treatment significantly enhanced neurogenesis in the SVZ of TBI mice, as evidenced by decreased TSPO expression and increased levels of Nestin and NeuroD1 compared to vehicle-treated controls. While TSPO is known to be expressed in neural stem cells and has been implicated in repressing neuronal differentiation [26,27,28,29], Nestin and NeuroD1 are well-established markers of neural stem and progenitor cells, respectively [30]. Moreover, 2-Cl-MGV-1-treated mice exhibited a greater number of NeuroD1-positive cells within the injured cortex and perilesional regions than vehicle-treated mice. These findings suggest that, beyond its anti-inflammatory properties, 2-Cl-MGV-1 may enhance brain repair by promoting neural stem cell proliferation, neuronal differentiation, and directed migration of precursors to the site of injury, thus supporting more effective regeneration of injured cortical tissue following TBI.
In rodents, neuronal precursors originating from the SVZ typically migrate along the rostral migratory stream to the olfactory bulb, where they differentiate into GABAergic interneurons that support olfactory neurogenesis [32,33,34]. However, following brain injury, this migratory path can be redirected toward damaged cortical areas [32,33,43,51], a process influenced by inflammatory signals released from reactive microglia [43]. In the present study, we detected NeuroD1-positive cells throughout several regions of the olfactory bulb in vehicle-treated mice, with the highest density observed in the mitral and granule cell layers, and to a lesser extent in the periglomerular zone. Strikingly, TBI mice treated with 2-Cl-MGV-1 showed a significant reduction in the number of these neuronal precursor cells in the olfactory bulb. These findings suggest that 2-Cl-MGV-1 facilitates the redirection of newly generated neuronal precursors away from their canonical route toward the olfactory bulb, instead promoting their migration to the injured cortex. This rerouting likely enhances neuroblast recruitment to the lesion site, potentially contributing to neuronal replacement and the restoration of disrupted cortical circuitry following TBI.
Our findings align with previous studies showing that TSPO ligands can modulate cell fate and promote differentiation. Compounds such as diazepam, Ro5-4864, PK 11195 and MGV-1 have been shown to induce cell proliferation and differentiation in vitro [13,18,25,26,28]. These findings suggest that TSPO may function as a regulatory node in signaling pathways that control stem cell commitment and neuronal differentiation, extending beyond its conventional role as a biomarker of neuroinflammation. Recent evidence indicates that TSPO ligands such as PK 11195 can influence nuclear transcription through mitochondria-to-nucleus signaling pathways, modulated by fluctuations in ROS, Ca²⁺, and ATP levels [13]. Other ligands, such as PIGA and XBD173, modulate steroidogenesis [52,53], which may aid neuroregeneration and impact gene networks involved in neurogenesis and plasticity [54,55,56]. For example, TSPO activation can promote oligodendrocyte maturation via neurosteroid biosynthesis [57]. Altogether, this data point to TSPO plays an active role in cell differentiation and gene regulation during brain repair. Accordingly, 2-Cl-MGV-1 may relieve TSPO-linked transcriptional repression, facilitating neurogenic gene expression, as evidenced by increased Nestin and NeuroD1 levels and enhanced migration of neuronal precursors to the injured cortex. This underscores the neurogenic and regenerative potential of 2-Cl-MGV-1 following TBI.
Several studies have shown that TBI can also stimulate hippocampal neurogenesis [58,59,60,61,62,63], with increased cell proliferation and differentiation evident within one week post-injury [60], and a marked rise in newly generated granule cells in the dentate gyrus persisting for up to two months [64]. In line with this, vehicle-treated TBI mice in our study exhibited hippocampal neurogenesis, evidenced by elevated expression of neural stem/progenitor cell markers and high β-III Tubulin levels in the dentate gyrus, including the neurogenic SGZ. In contrast, hippocampal neurogenesis was reduced in 2-Cl-MGV-1-treated TBI mice. Compared to vehicle-treated mice, neurogenic markers (TSPO, Nestin, NeuroD1, and β-III-Tubulin) were significantly downregulated after treatment with 2-Cl-MGV-1. These data suggest that the observed hippocampal recovery occurred largely independently of endogenous neurogenic mechanisms. Importantly, the extent of hippocampal neurogenesis following TBI can be influenced by injury severity. While mild TBI has minimal effect, moderate TBI increases stem cell proliferation without promoting differentiation, whereas severe TBI leads to a robust neurogenic response [62]. In our model, the injury extended to the CA2 and CA3 regions and was associated with high TSPO expression in the dentate gyrus of vehicle-treated mice, reflecting ongoing inflammation and degeneration, both of which were significantly mitigated by 2-Cl-MGV-1 treatment. Similar neuroprotective effects following TBI have been observed with other TSPO ligands, including Etifoxine, Ro5-4864, and PK 11195 [1,16,20,65]. Their beneficial actions are attributed to attenuation of cell death, neuroinflammation, and excitotoxicity, as demonstrated in diverse experimental models [1,7,13,18,19,20,66,67]. Accordingly, the hippocampal effects of 2-Cl-MGV-1 likely reflect its anti-inflammatory and neuroprotective actions, rather than stimulation of neurogenesis. This interpretation is consistent with the findings of Gao et al., who reported persistent neuroinflammation in the hippocampus after TBI, with neural stem/progenitor cells preferentially differentiating into astrocytes rather than neurons [68].
4. Materials and Methods
4.1. Animals and TBI Model
This study included male C57BL/6N mice (Harlan, Jerusalem), aged 6-15 weeks (6-7 weeks at the start of the study), whose weight-range was 20–24 g at the time of TBI induction. Mice were housed in the animal facility of the Israel Institute of Technology (Technion) under a 12:12-hour light/dark cycle, in a temperature-controlled environment (20–23 °C). Animals had tap water and standard pellet diet ad libitum. All procedures were conducted in accordance with institutional guidelines and were approved by the Animal Experimentation Ethics Committee at the Technion (IL-034-03-2015).
Blunt TBI was induced in mice using the Shohami´s Weight Drop device, which mimics human TBI pathophysiology and variability among individuals [3,69,70]. Immediately before the procedure, mice were anesthetized with 0.5–1.5% isoflurane (Piramal Critical Care, Bethlehem, PA) supplemented with oxygen (0.8 L/min) applied by a Midmark VME2 Wall Mount System anesthesia machine (UXR, Pointe-Claire, QC, Canada). A focal injury was delivered to the left cerebral hemisphere by placing a 2 mm diameter Teflon-tipped cone on the exposed skull, positioned 1 mm lateral to the midline and 1 mm caudal to bregma. A 95 g weight was then dropped from a height of 6 cm through a Plexiglas tube onto the cone. Animals were carefully monitored throughout the procedure, and if necessary, mechanically stimulated to ensure continuous breathing.
4.2. Treatments
Mice were treated daily for 60 days with subcutaneous injections of either vehicle (DMSO; N = 6) or 2-Cl-MGV-1 [2-(2-chlorophenyl) quinazolin-4-yl dimethylcarbamate] at a dose of 7.5 mg/kg (N = 6). Injections were administered in the dorsal cervical region (back of the neck) within a post-injury time window of 7 to 11 hours following TBI induction. The dosage of 2-Cl-MGV-1 was selected based on previously published studies demonstrating its pharmacological efficacy and safety [18,21,22].
4.3. Histological Procedures
At the end of the treatment period, animals were deeply anesthetized with a ketamine/xylazine mixture (10:1 mg/kg, i.p.) and transcardially perfused through the left ventricle at a rate of 15 mL/min, initially with phosphate-buffered saline (PBS, pH 7.4, 10 mL), followed by 4% paraformaldehyde in PBS for 10 minutes. The brains fixed in paraformaldehyde were sent to the University of Oviedo, where they were dehydrated through a graded ethanol series (70%, 80%, 96%, 100%), followed by immersion in 100% isopropanol. Subsequently, brains were embedded in paraffin. Coronal brain sections (8–10μm thick) were obtained using a rotary microtome (Leica RM2155, Leica Biosystems, Nussloch, Germany). Sections encompassing the injured areas were mounted on Superfrost® Plus microscope slides (Thermo Scientific, Waltham, MA, USA).
4.4. Immunohistochemistry
After deparaffinization, histological sections were rehydrated and incubated for 10 minutes in TBS buffer (5 mM Tris, 136 mM NaCl, pH 7.4) containing 0.01% Triton X-100. Sections were then washed in TBS (3 × 5 min). For antigen retrieval, the sections were incubated in a pre-warmed 0.01 M citrate buffer (pH 6.0) in a water bath and subsequently subjected to five heating cycles in a microwave oven at 600 W for 3 minutes each, taking care to prevent evaporation. After heating, the sections were allowed to cool at room temperature for 10 minutes in the citrate buffer, followed by a further 10-minute incubation in TBS.
Endogenous peroxidase activity was quenched by incubating the sections in 3% H₂O₂ in methanol for 20 minutes. After washing in TBS (3 × 5 min), non-specific binding was blocked for 40 minutes using 50µg of BSA and pure rabbit serum diluted 1:40 in TBS. Sections were then incubated overnight at 4 °C in a humidified, dark chamber with specific primary antibodies (Table 1), diluted 1:100 in TBS.
Following another series of washes (3 × 5 min in TBS), sections were incubated for 90 minutes at room temperature with HRP-conjugated anti-IgG antibodies (Sigma), diluted 1:1000 in TBS, also in a humidified, light-protected chamber. This was followed by a 1-hour incubation at room temperature with peroxidase anti-peroxidase complexes (Sigma), diluted 1:200 in TBS. After a final round of washes (3 × 5 min in TBS), immunoreactivity was visualized by incubating the sections for 10 minutes with 3,3′-diaminobenzidine tablets (SIGMAFAST™, Sigma). Counterstaining was performed using 0.1% hematoxylin for 5 minutes, followed by dehydration and mounting in an aqueous medium (Eukitt).
4.5. Images Analysis
Histological analyses were performed using a Nikon Eclipse E400 light microscope equipped with a DS-Fi1 camera. Images were acquired with the Toup View 3.7 software and subsequently analyzed using FIJI (Image J). The DAB and hematoxylin signals were quantified independently in each image. For each experimental condition (vehicle or 2-Cl-MGV-1) and histological staining (NeuN, TSPO, Nestin, NeuroD1, β-Tubulin III), a total of 10 images per animal were analyzed at 400x magnifications. Results were expressed as the percentage of DAB intensity relative to the number of nuclei, normalized to vehicle-treated mice (set as 100%).
4.6. Statistical Analysis
Statistical analyses were performed using GraphPad Prism 6. Data are presented as mean ± S.E.M., calculated from at least three independent experiments. Normality was assessed using the Kolmogorov–Smirnov test. Comparisons between vehicle-treated and 2-Cl-MGV-1-treated mice were made using an unpaired Student’s t-test. A p-value < 0.05 was considered statistically significant.
5. Conclusions
TBI induced strong neuroinflammation and neurodegeneration in the cortex and hippocampus of vehicle-treated mice. These injuries elicited a neurogenic response in both primary neurogenic niches, the SVZ and SGZ, likely as an attempt to restore brain homeostasis and promote recovery. Treatment with 2-Cl-MGV-1 attenuated neuroinflammation and neuronal damage in the brains of mice subjected to TBI, contributing to improved outcomes relative to vehicle-treated mice. Specifically, 2-Cl-MGV-1 promoted substantial structural recovery in the hippocampus and reduced cortical injury to a small glial scar. Interestingly, the effects of 2-Cl-MGV-1 on neurogenesis were region-specific. In the SVZ, it significantly enhanced neurogenic activity and redirected neuroblasts -normally destined for the olfactory bulb- toward the cortical lesion site, thereby supporting regeneration at the primary site of injury. In contrast, hippocampal neurogenesis was diminished following treatment with 2-Cl-MGV-1, suggesting that recovery in this region was driven by non-neurogenic mechanisms. Here, the protective actions of 2-Cl-MGV-1 appear to rely primarily on anti-inflammatory effects and the preservation of neuronal integrity. Together, these results highlight the differential modulation of neurogenesis by 2-Cl-MGV-1 across distinct brain regions and support its therapeutic potential in enhancing brain repair after TBI (see Graphical Abstract).
Author Contributions
Conceptualization, A.C.M, B.C., L.V., N.Y., A.W. and M.G; Methodology, N.Y., E.A., C.C.V, C.G.G, P.F.G. and N.M.C; Formal analysis, B.C., Y.P. J.A.B. and I.V.N; Investigation, B.C., J.A.B., E.A., C.C.V., C.G.G, P.F.G., N.M.C., I.V.N., Y.P., L.V. and N.Y. ; Resources, A.C.M, M.G. and A.W.; Data curation, Y.P., I.V.N., J.A.B., E.A., C.C.V., C.G.G, P.F.G. and N.M.C.; Writing—original draft; B.C. and A.C. M.; Writing—review & editing, Y.P., I.V:N., M.G, L.V. and A.W.; Supervision, B.C., A.C.M., L.V., A.W. and M.G.; Project administration, A.C.M, M.G., A.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Sekuens Agency (Government of the Principality of Asturias), gran number IDE/2024/000719; Instituto de Salud Carlos III, grant number PI21/01596; and the Israel Innovation Authority: Kamin Incentive Program, grant num. 2024874. EA thanks his pre-doctoral fellowship FI22/00286 to Instituto de Salud Carlos III. CCV thanks her pre-doctoral fellowship PA-23-BP22-073 to the Government of the Principality of Asturias.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board of the Technion (protocol code IL-034-03-2015 from March 2015) for studies involving animals.
Informed Consent Statement
Not applicable
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed at the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 2-Cl-MGV-1 | 2-(2-chlorophenyl) quinazolin-4-yl dimethylcarbamate |
| DMSO | Dimethyl sulfoxide |
| GCL | Granule cell layer |
| GL | Glomerular cell layer |
| Nestin | Neuroectodermal stem cell protein |
| NeuN | Neuronal nuclear antigen |
| NeuroD1 | Neurogenic differentiation factor 1 |
| SGZ | Subgranular zone |
| SVZ | Subventricular zone |
| TBI | Traumatic brain injury |
| TSPO | The 18-kDa translocator protein |
References
- Bao, Q.; Yuan, X.; Bian, X.; Wei, W.; Jin, P.; Jiang, W. Prognostic Significance of Translocator Protein in Brain Tissue Following Traumatic Brain Injury. Turkish neurosurgery 2023, 33, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Mohan, M.C.; Anjana, A.S.; Hilmi Jaufer, T.A.; Deepti, A.; Krishnakumar, I.M.; Baby Chakrapani, P.S. Co-delivery of curcumin-resveratrol-carnosic acid complex promotes neurogenesis and cognitive recovery in a rodent model of repeated mild traumatic brain injury. Biomedicine & pharmacotherapy 2025, 183, 117818. [Google Scholar] [CrossRef]
- Ma, X.; Aravind, A.; Pfister, B.J.; Chandra, N.; Haorah, J. Animal Models of Traumatic Brain Injury and Assessment of Injury Severity. Molecular neurobiology 2019, 56, 5332–5345. [Google Scholar] [CrossRef] [PubMed]
- Khazaal, A.Q.; Ismaeel, H.M.; Cheah, P.S.; Nordin, N. Cellular Stem Cell Therapy for Treating Traumatic Brain Injury: Strategies for Enhancement of Therapeutic Efficacy. Molecular neurobiology, 2025. [Google Scholar] [CrossRef]
- Donat, C.K.; Scott, G.; Gentleman, S.M.; Sastre, M. Microglial Activation in Traumatic Brain Injury. Frontiers in aging neuroscience 2017, 9, 208. [Google Scholar] [CrossRef]
- Caballero, B.; Veenman, L.; Gavish, M. Role of mitochondrial translocator protein (18 kDa) on mitochondrial- related cell death processes. Recent patents on endocrine, metabolic & immune drug discovery 2013, 7, 86–101. [Google Scholar] [CrossRef]
- Caballero, B.; Veenman, L.; Bode, J.; Leschiner, S.; Gavish, M. Concentration-dependent bimodal effect of specific 18 kDa translocator protein (TSPO) ligands on cell death processes induced by ammonium chloride: potential implications for neuropathological effects due to hyperammonemia. CNS & neurological disorders drug targets 2014, 13, 574–592. [Google Scholar] [CrossRef]
- Veenman, L.; Vainshtein, A.; Gavish, M. TSPO as a target for treatments of diseases, including neuropathological disorders. Cell death & disease 2015, 6, e1911. [Google Scholar] [CrossRef]
- McNeela, A.M.; Bernick, C.; Hines, R.M.; Hines, D.J. TSPO regulation in reactive gliotic diseases. Journal of neuroscience research 2018, 96, 978–988. [Google Scholar] [CrossRef]
- Luo, L.F.; Weng, J.F.; Cen, M.; Dong, X.Q.; Yu, W.H.; Du, Q.; Yang, D.B.; Zheng, Y.K.; Hu, W.; Yu, L. , et al. Prognostic significance of serum translocator protein in patients with traumatic brain injury. Clinica chimica acta; international journal of clinical chemistry 2019, 488, 25–30. [Google Scholar] [CrossRef]
- Missault, S.; Anckaerts, C.; Blockx, I.; Deleye, S.; Van Dam, D.; Barriche, N.; De Pauw, G.; Aertgeerts, S.; Valkenburg, F.; De Deyn, P.P. , et al. Neuroimaging of Subacute Brain Inflammation and Microstructural Changes Predicts Long-Term Functional Outcome after Experimental Traumatic Brain Injury. Journal of neurotrauma 2019, 36, 768–788. [Google Scholar] [CrossRef]
- Bode, J.; Veenman, L.; Caballero, B.; Lakomek, M.; Kugler, W.; Gavish, M. The 18 kDa translocator protein influences angiogenesis, as well as aggressiveness, adhesion, migration, and proliferation of glioblastoma cells. Pharmacogenetics and genomics 2012, 22, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Yasin, N.; Veenman, L.; Singh, S.; Azrad, M.; Bode, J.; Vainshtein, A.; Caballero, B.; Marek, I.; Gavish, M. Classical and Novel TSPO Ligands for the Mitochondrial TSPO Can Modulate Nuclear Gene Expression: Implications for Mitochondrial Retrograde Signaling. International journal of molecular sciences 2017, 18. [Google Scholar] [CrossRef]
- Dimitrova-Shumkovska, J.; Krstanoski, L.; Veenman, L. Diagnostic and Therapeutic Potential of TSPO Studies Regarding Neurodegenerative Diseases, Psychiatric Disorders, Alcohol Use Disorders, Traumatic Brain Injury, and Stroke: An Update. Cells 2020, 9. [Google Scholar] [CrossRef]
- Varga, B.; Marko, K.; Hadinger, N.; Jelitai, M.; Demeter, K.; Tihanyi, K.; Vas, A.; Madarasz, E. Translocator protein (TSPO 18kDa) is expressed by neural stem and neuronal precursor cells. Neuroscience letters 2009, 462, 257–262. [Google Scholar] [CrossRef]
- Palzur, E.; Sharon, A.; Shehadeh, M.; Soustiel, J.F. Investigation of the mechanisms of neuroprotection mediated by Ro5-4864 in brain injury. Neuroscience 2016, 329, 162–170. [Google Scholar] [CrossRef]
- Notter, T.; Schalbetter, S.M.; Clifton, N.E.; Mattei, D.; Richetto, J.; Thomas, K.; Meyer, U.; Hall, J. Neuronal activity increases translocator protein (TSPO) levels. Molecular psychiatry 2021, 26, 2025–2037. [Google Scholar] [CrossRef] [PubMed]
- Vainshtein, A.; Veenman, L.; Shterenberg, A.; Singh, S.; Masarwa, A.; Dutta, B.; Island, B.; Tsoglin, E.; Levin, E.; Leschiner, S. , et al. Quinazoline-based tricyclic compounds that regulate programmed cell death, induce neuronal differentiation, and are curative in animal models for excitotoxicity and hereditary brain disease. Cell death discovery 2015, 1, 15027. [Google Scholar] [CrossRef] [PubMed]
- Azrad, M.; Zeineh, N.; Weizman, A.; Veenman, L.; Gavish, M. The TSPO Ligands 2-Cl-MGV-1, MGV-1, and PK11195 Differentially Suppress the Inflammatory Response of BV-2 Microglial Cell to LPS. International journal of molecular sciences 2019, 20. [Google Scholar] [CrossRef]
- Shehadeh, M.; Palzur, E.; Apel, L.; Soustiel, J.F. Reduction of Traumatic Brain Damage by Tspo Ligand Etifoxine. International journal of molecular sciences 2019, 20. [Google Scholar] [CrossRef]
- Yasin N., V. L., Caballero B., Zeineh N., Gonzalez-Blanco L., Weizman A., Gavish G. TSPO ligand 2-Cl-MGV-1 mitigates trumatic brain INjury (TBI) in a mouse model. International Journal of Molecular Science 2025, 26. [Google Scholar] [CrossRef]
- Chen, Y.; Veenman, L.; Singh, S.; Ouyang, F.; Liang, J.; Huang, W.; Marek, I.; Zeng, J.; Gavish, M. 2-Cl-MGV-1 Ameliorates Apoptosis in the Thalamus and Hippocampus and Cognitive Deficits After Cortical Infarct in Rats. Stroke 2017, 48, 3366–3374. [Google Scholar] [CrossRef] [PubMed]
- Cachan-Vega, C.; Vega-Naredo, I.; Potes, Y.; Bermejo-Millo, J.C.; Rubio-Gonzalez, A.; Garcia-Gonzalez, C.; Antuna, E.; Bermudez, M.; Gutierrez-Rodriguez, J.; Boga, J.A. , et al. Chronic Treatment with Melatonin Improves Hippocampal Neurogenesis in the Aged Brain and Under Neurodegeneration. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Gross, C.G. Neurogenesis in the adult brain: death of a dogma. Nature reviews. Neuroscience 2000, 1, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Krestinina, O.V.M., S. N.; Baburina, Y; Fadeev, R.S.; Azarashavili, T.S.; Akatov, V.S. The effects of isoquinoline carboxamide and melatonin on the differentation of N1E-115 mouse neuroblastoma cells (clone C-1300) and on the expression of the the TSPO translocation protein and 2´,3´-cyclonucleotide-3´-phosphodiesterase in these cells. Neurochem J 2017, 11. [Google Scholar] [CrossRef]
- Gonzalez-Blanco, L.; Bermejo-Millo, J.C.; Oliveira, G.; Potes, Y.; Antuna, E.; Menendez-Valle, I.; Vega-Naredo, I.; Coto-Montes, A.; Caballero, B. Neurogenic Potential of the 18-kDa Mitochondrial Translocator Protein (TSPO) in Pluripotent P19 Stem Cells. Cells 2021, 10. [Google Scholar] [CrossRef]
- Betlazar, C.; Harrison-Brown, M.; Middleton, R.J.; Banati, R.; Liu, G.J. Cellular Sources and Regional Variations in the Expression of the Neuroinflammatory Marker Translocator Protein (TSPO) in the Normal Brain. International journal of molecular sciences 2018, 19. [Google Scholar] [CrossRef]
- Lee, D.H.; Kang, S.K.; Lee, R.H.; Ryu, J.M.; Park, H.Y.; Choi, H.S.; Bae, Y.C.; Suh, K.T.; Kim, Y.K.; Jung, J.S. Effects of peripheral benzodiazepine receptor ligands on proliferation and differentiation of human mesenchymal stem cells. Journal of cellular physiology 2004, 198, 91–99. [Google Scholar] [CrossRef]
- Tanimoto, Y.; Yamasaki, T.; Nagoshi, N.; Nishiyama, Y.; Nori, S.; Nishimura, S.; Iida, T.; Ozaki, M.; Tsuji, O.; Ji, B. , et al. In vivo monitoring of remnant undifferentiated neural cells following human induced pluripotent stem cell-derived neural stem/progenitor cells transplantation. Stem cells translational medicine 2020, 9, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jiao, J. Molecular Biomarkers for Embryonic and Adult Neural Stem Cell and Neurogenesis. BioMed research international 2015, 2015, 727542. [Google Scholar] [CrossRef]
- Memberg, S.P.; Hall, A.K. Dividing neuron precursors express neuron-specific tubulin. Journal of neurobiology 1995, 27, 26–43. [Google Scholar] [CrossRef]
- Akter, M.; Kaneko, N.; Sawamoto, K. Neurogenesis and neuronal migration in the postnatal ventricular-subventricular zone: Similarities and dissimilarities between rodents and primates. Neuroscience research 2021, 167, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Navarro Quiroz, E.; Navarro Quiroz, R.; Ahmad, M.; Gomez Escorcia, L.; Villarreal, J.L.; Fernandez Ponce, C.; Aroca Martinez, G. Cell Signaling in Neuronal Stem Cells. Cells 2018, 7. [Google Scholar] [CrossRef]
- Defterali, C.; Moreno-Estelles, M.; Crespo, C.; Diaz-Guerra, E.; Diaz-Moreno, M.; Vergano-Vera, E.; Nieto-Estevez, V.; Hurtado-Chong, A.; Consiglio, A.; Mira, H. , et al. Neural stem cells in the adult olfactory bulb core generate mature neurons in vivo. Stem cells 2021, 39, 1253–1269. [Google Scholar] [CrossRef]
- Wang, Y.; Yue, X.; Kiesewetter, D.O.; Niu, G.; Teng, G.; Chen, X. PET imaging of neuroinflammation in a rat traumatic brain injury model with radiolabeled TSPO ligand DPA-714. European journal of nuclear medicine and molecular imaging 2014, 41, 1440–1449. [Google Scholar] [CrossRef] [PubMed]
- Israel, I.; Ohsiek, A.; Al-Momani, E.; Albert-Weissenberger, C.; Stetter, C.; Mencl, S.; Buck, A.K.; Kleinschnitz, C.; Samnick, S.; Siren, A.L. Combined [(18)F]DPA-714 micro-positron emission tomography and autoradiography imaging of microglia activation after closed head injury in mice. Journal of neuroinflammation 2016, 13, 140. [Google Scholar] [CrossRef]
- Monga, S.; Nagler, R.; Amara, R.; Weizman, A.; Gavish, M. Inhibitory Effects of the Two Novel TSPO Ligands 2-Cl-MGV-1 and MGV-1 on LPS-induced Microglial Activation. Cells 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Obeid, F.; Kahana, M.; Dahle, B.; Monga, S.; Zohar, Y.; Weizman, A.; Gavish, M. Novel TSPO Ligand 2-Cl-MGV-1 Can Counteract Lipopolysaccharide Induced Inflammatory Response in Murine RAW264.7 Macrophage Cell Line and Lung Models. Cells 2024, 13. [Google Scholar] [CrossRef]
- Monga, S.; Weizman, A.; Gavish, M. The Efficacy of the Novel TSPO Ligands 2-Cl-MGV-1 and 2,4-Di-Cl-MGV-1 Compared to the Classical TSPO Ligand PK 11195 to Counteract the Release of Chemokines from LPS-Stimulated BV-2 Microglial Cells. Biology 2020, 9. [Google Scholar] [CrossRef]
- Wang, Z.; Zheng, Y.; Zheng, M.; Zhong, J.; Ma, F.; Zhou, B.; Zhu, J. Neurogenic Niche Conversion Strategy Induces Migration and Functional Neuronal Differentiation of Neural Precursor Cells Following Brain Injury. Stem cells and development 2020, 29, 235–248. [Google Scholar] [CrossRef]
- Astakhova, O.; Ivanova, A.; Komoltsev, I.; Gulyaeva, N.; Enikolopov, G.; Lazutkin, A. Traumatic Brain Injury Promotes Neurogenesis and Oligodendrogenesis in Subcortical Brain Regions of Mice. Cells 2025, 14. [Google Scholar] [CrossRef]
- Chen, Y.; Veenman, L.; Liao, M.; Huang, W.; Yu, J.; Zeng, J. Enhanced angiogenesis in the thalamus induced by a novel TSPO ligand ameliorates cognitive deficits after focal cortical infarction. Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism 2024, 44, 477–490. [Google Scholar] [CrossRef] [PubMed]
- Amanollahi, M.; Jameie, M.; Heidari, A.; Rezaei, N. The Dialogue Between Neuroinflammation and Adult Neurogenesis: Mechanisms Involved and Alterations in Neurological Diseases. Molecular neurobiology 2023, 60, 923–959. [Google Scholar] [CrossRef]
- Castro, E.S.J.H.; Pieropan, F.; Rivera, A.D.; Butt, A.M.; Costa, S.L. Agathisflavone Modulates Reactive Gliosis After Trauma and Increases the Neuroblast Population at the Subventricular Zone. Nutrients 2024, 16. [Google Scholar] [CrossRef]
- Badner, A.; Reinhardt, E.K.; Nguyen, T.V.; Midani, N.; Marshall, A.T.; Lepe, C.A.; Echeverria, K.; Lepe, J.J.; Torrecampo, V.; Bertan, S.H. , et al. Freshly Thawed Cryobanked Human Neural Stem Cells Engraft within Endogenous Neurogenic Niches and Restore Cognitive Function after Chronic Traumatic Brain Injury. Journal of neurotrauma 2021, 38, 2731–2746. [Google Scholar] [CrossRef] [PubMed]
- Badner, A.; Cummings, B.J. The endogenous progenitor response following traumatic brain injury: a target for cell therapy paradigms. Neural regeneration research 2022, 17, 2351–2354. [Google Scholar] [CrossRef] [PubMed]
- Hajinejad, M.; Ebrahimzadeh, M.H.; Ebrahimzadeh-Bideskan, A.; Rajabian, A.; Gorji, A.; Sahab Negah, S. Exosomes and Nano-SDF Scaffold as a Cell-Free-Based Treatment Strategy Improve Traumatic Brain Injury Mechanisms by Decreasing Oxidative Stress, Neuroinflammation, and Increasing Neurogenesis. Stem cell reviews and reports 2023, 19, 1001–1018. [Google Scholar] [CrossRef]
- Choi, B.Y.; Hong, D.K.; Kang, B.S.; Lee, S.H.; Choi, S.; Kim, H.J.; Lee, S.M.; Suh, S.W. Engineered Mesenchymal Stem Cells Over-Expressing BDNF Protect the Brain from Traumatic Brain Injury-Induced Neuronal Death, Neurological Deficits, and Cognitive Impairments. Pharmaceuticals 2023, 16. [Google Scholar] [CrossRef]
- Galgano, M.; Toshkezi, G.; Qiu, X.; Russell, T.; Chin, L.; Zhao, L.R. Traumatic Brain Injury: Current Treatment Strategies and Future Endeavors. Cell transplantation 2017, 26, 1118–1130. [Google Scholar] [CrossRef]
- Schepici, G.; Silvestro, S.; Bramanti, P.; Mazzon, E. Traumatic Brain Injury and Stem Cells: An Overview of Clinical Trials, the Current Treatments and Future Therapeutic Approaches. Medicina 2020, 56. [Google Scholar] [CrossRef]
- Deshpande, S.S.; Malik, S.C.; Conforti, P.; Lin, J.D.; Chu, Y.H.; Nath, S.; Greulich, F.; Dumbach, M.A.; Uhlenhaut, N.H.; Schachtrup, C. P75 neurotrophin receptor controls subventricular zone neural stem cell migration after stroke. Cell and tissue research 2022, 387, 415–431. [Google Scholar] [CrossRef]
- Schule, C.; Nothdurfter, C.; Rupprecht, R. The role of allopregnanolone in depression and anxiety. Progress in neurobiology 2014, 113, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Da Pozzo, E.; Giacomelli, C.; Costa, B.; Cavallini, C.; Taliani, S.; Barresi, E.; Da Settimo, F.; Martini, C. TSPO PIGA Ligands Promote Neurosteroidogenesis and Human Astrocyte Well-Being. International journal of molecular sciences 2016, 17. [Google Scholar] [CrossRef]
- Charalampopoulos, I.; Remboutsika, E.; Margioris, A.N.; Gravanis, A. Neurosteroids as modulators of neurogenesis and neuronal survival. Trends in endocrinology and metabolism: TEM 2008, 19, 300–307. [Google Scholar] [CrossRef]
- Hojo, Y.; Kawato, S. Neurosteroids in Adult Hippocampus of Male and Female Rodents: Biosynthesis and Actions of Sex Steroids. Frontiers in endocrinology 2018, 9, 183. [Google Scholar] [CrossRef] [PubMed]
- Sahab-Negah, S.; Hajali, V.; Moradi, H.R.; Gorji, A. The Impact of Estradiol on Neurogenesis and Cognitive Functions in Alzheimer's Disease. Cellular and molecular neurobiology 2020, 40, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Tremolanti, C.; Angeloni, E.; Da Pozzo, E.; Germelli, L.; Giacomelli, C.; Scalzi, E.; Taliani, S.; Da Settimo, F.; Mensah-Nyagan, A.G.; Martini, C. , et al. Human oligodendrocyte-like cell differentiation is promoted by TSPO-mediated endogenous steroidogenesis. Biochimica et biophysica acta. Molecular basis of disease 2024, 1870, 167174. [Google Scholar] [CrossRef]
- Dash, P.K.; Mach, S.A.; Moore, A.N. Enhanced neurogenesis in the rodent hippocampus following traumatic brain injury. Journal of neuroscience research 2001, 63, 313–319. [Google Scholar] [CrossRef]
- Richardson, R.M.; Sun, D.; Bullock, M.R. Neurogenesis after traumatic brain injury. Neurosurgery clinics of North America 2007, 18, 169–181. [Google Scholar] [CrossRef]
- Ye, Y.; Zhao, Z.; Xu, H.; Zhang, X.; Su, X.; Yang, Y.; Yu, X.; He, X. Activation of Sphingosine 1-Phosphate Receptor 1 Enhances Hippocampus Neurogenesis in a Rat Model of Traumatic Brain Injury: An Involvement of MEK/Erk Signaling Pathway. Neural plasticity 2016, 2016, 8072156. [Google Scholar] [CrossRef]
- Weston, N.M.; Rolfe, A.T.; Freelin, A.H.; Reeves, T.M.; Sun, D. Traumatic brain injury modifies synaptic plasticity in newly-generated granule cells of the adult hippocampus. Experimental neurology 2021, 336, 113527. [Google Scholar] [CrossRef]
- Wang, X.; Gao, X.; Michalski, S.; Zhao, S.; Chen, J. Traumatic Brain Injury Severity Affects Neurogenesis in Adult Mouse Hippocampus. Journal of neurotrauma 2016, 33, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Bielefeld, P.; Martirosyan, A.; Martin-Suarez, S.; Apresyan, A.; Meerhoff, G.F.; Pestana, F.; Poovathingal, S.; Reijner, N.; Koning, W.; Clement, R.A. , et al. Traumatic brain injury promotes neurogenesis at the cost of astrogliogenesis in the adult hippocampus of male mice. Nature communications 2024, 15, 5222. [Google Scholar] [CrossRef] [PubMed]
- Osier, N.D.; Carlson, S.W.; DeSana, A.; Dixon, C.E. Chronic Histopathological and Behavioral Outcomes of Experimental Traumatic Brain Injury in Adult Male Animals. Journal of neurotrauma 2015, 32, 1861–1882. [Google Scholar] [CrossRef]
- Simon-O'Brien, E.; Gauthier, D.; Riban, V.; Verleye, M. Etifoxine improves sensorimotor deficits and reduces glial activation, neuronal degeneration, and neuroinflammation in a rat model of traumatic brain injury. Journal of neuroinflammation 2016, 13, 203. [Google Scholar] [CrossRef]
- Jazvinscak Jembrek, M.; Radovanovic, V.; Vlainic, J.; Vukovic, L.; Hanzic, N. Neuroprotective effect of zolpidem against glutamate-induced toxicity is mediated via the PI3K/Akt pathway and inhibited by PK11195. Toxicology 2018, 406-407, 58–69. [Google Scholar] [CrossRef]
- Palzur, E.; Edelman, D.; Sakas, R.; Soustiel, J.F. Etifoxine Restores Mitochondrial Oxidative Phosphorylation and Improves Cognitive Recovery Following Traumatic Brain Injury. International journal of molecular sciences 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Chen, J. Moderate traumatic brain injury promotes neural precursor proliferation without increasing neurogenesis in the adult hippocampus. Experimental neurology 2013, 239, 38–48. [Google Scholar] [CrossRef]
- Khalin, I.; Jamari, N.L.; Razak, N.B.; Hasain, Z.B.; Nor, M.A.; Zainudin, M.H.; Omar, A.B.; Alyautdin, R. A mouse model of weight-drop closed head injury: emphasis on cognitive and neurological deficiency. Neural regeneration research 2016, 11, 630–635. [Google Scholar] [CrossRef]
- Kalish, B.T.; Whalen, M.J. Weight Drop Models in Traumatic Brain Injury. Methods in molecular biology 2016, 1462, 193–209. [Google Scholar] [CrossRef]
Figure 1.
Attenuation of neuronal loss in the brains of TBI mice following 2-Cl-MGV-1 treatment compared to vehicle (A) Immunohistochemical staining of NeuN in the brain of TBI mice treated with vehicle (DMSO) for 60 days at 40x magnification (B) Immunohistochemical staining of NeuN in the brain of TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days at 40x magnification. Arrows indicate regions of neuronal injury. Scale bar = 200 μm (C) Immunohistochemical staining of NeuN in the CA3 hippocampal region of TBI mice treated with vehicle (DMSO) for 60 days at 400x magnification (D) Immunohistochemical staining of NeuN in the CA3 hippocampal region of TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days at 400x magnification. Scale bar = 20 μm. (E) Bar graph showing quantification of NeuN staining in the CA3 hippocampal region, expressed as the percentage of DAB signal relative to the number of nuclei. Data is represented as mean ± SEM. ***p< 0.001, unpaired T-Test.
Figure 1.
Attenuation of neuronal loss in the brains of TBI mice following 2-Cl-MGV-1 treatment compared to vehicle (A) Immunohistochemical staining of NeuN in the brain of TBI mice treated with vehicle (DMSO) for 60 days at 40x magnification (B) Immunohistochemical staining of NeuN in the brain of TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days at 40x magnification. Arrows indicate regions of neuronal injury. Scale bar = 200 μm (C) Immunohistochemical staining of NeuN in the CA3 hippocampal region of TBI mice treated with vehicle (DMSO) for 60 days at 400x magnification (D) Immunohistochemical staining of NeuN in the CA3 hippocampal region of TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days at 400x magnification. Scale bar = 20 μm. (E) Bar graph showing quantification of NeuN staining in the CA3 hippocampal region, expressed as the percentage of DAB signal relative to the number of nuclei. Data is represented as mean ± SEM. ***p< 0.001, unpaired T-Test.
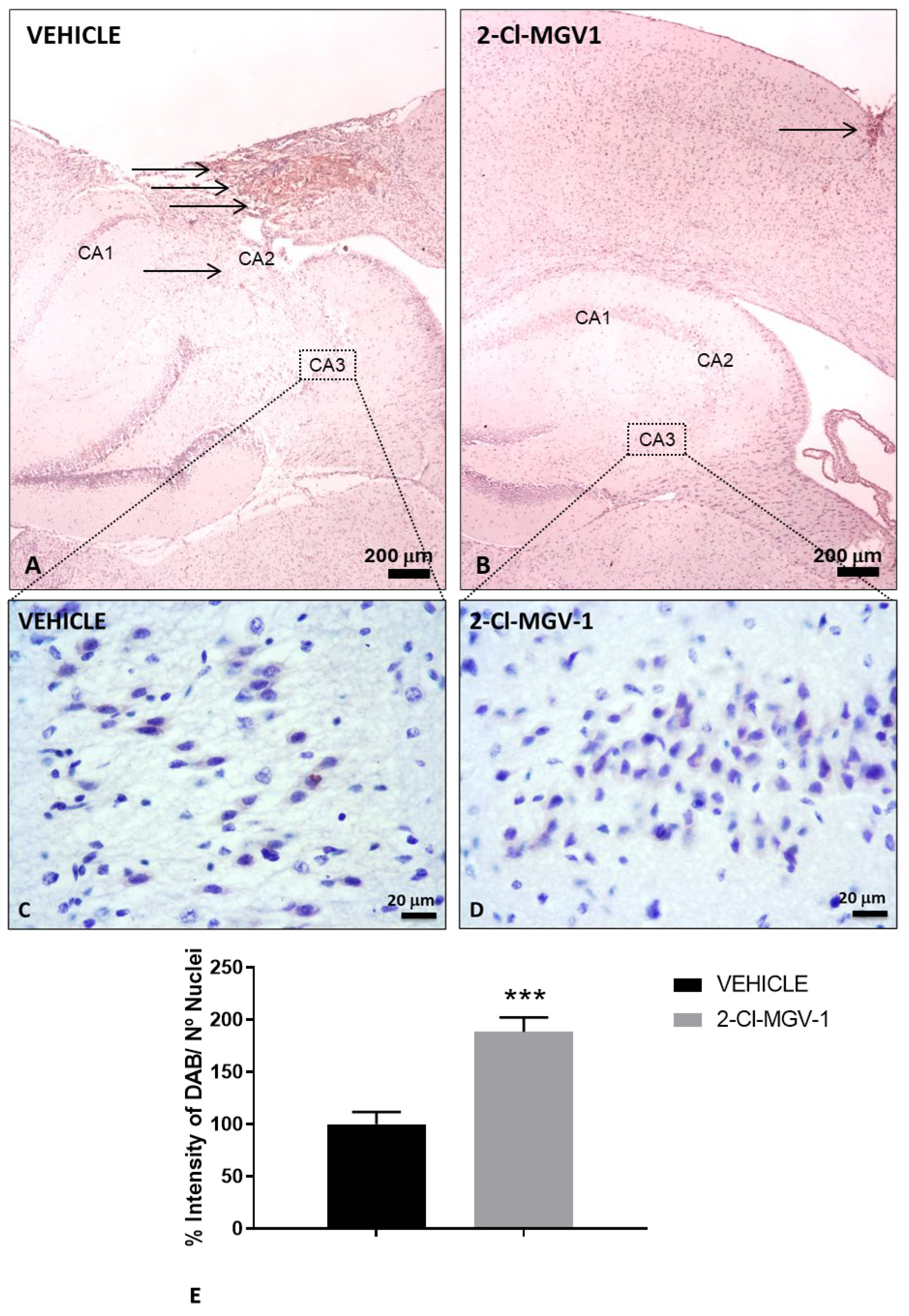
Figure 2.
Attenuation of neuroinflammation in TBI mice following 2-Cl-MGV-1 treatment compared to vehicle. (A, C) Immunohistochemical staining of TSPO in the brain cortex of TBI mice treated with vehicle (DMSO) for 60 days. (B, D) Immunohistochemical staining of TSPO in the brain cortex of TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days. Images were captured at 100x or 400x magnification. Scale bars = 100 μm or 20 μm, respectively. (E) Bar graph showing quantification of TSPO staining in the cortical region near the injury site, expressed as the percentage of DAB signal relative to the number of nuclei. Data are presented as mean ± SEM. ***p < 0.001, unpaired t-test.
Figure 2.
Attenuation of neuroinflammation in TBI mice following 2-Cl-MGV-1 treatment compared to vehicle. (A, C) Immunohistochemical staining of TSPO in the brain cortex of TBI mice treated with vehicle (DMSO) for 60 days. (B, D) Immunohistochemical staining of TSPO in the brain cortex of TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days. Images were captured at 100x or 400x magnification. Scale bars = 100 μm or 20 μm, respectively. (E) Bar graph showing quantification of TSPO staining in the cortical region near the injury site, expressed as the percentage of DAB signal relative to the number of nuclei. Data are presented as mean ± SEM. ***p < 0.001, unpaired t-test.
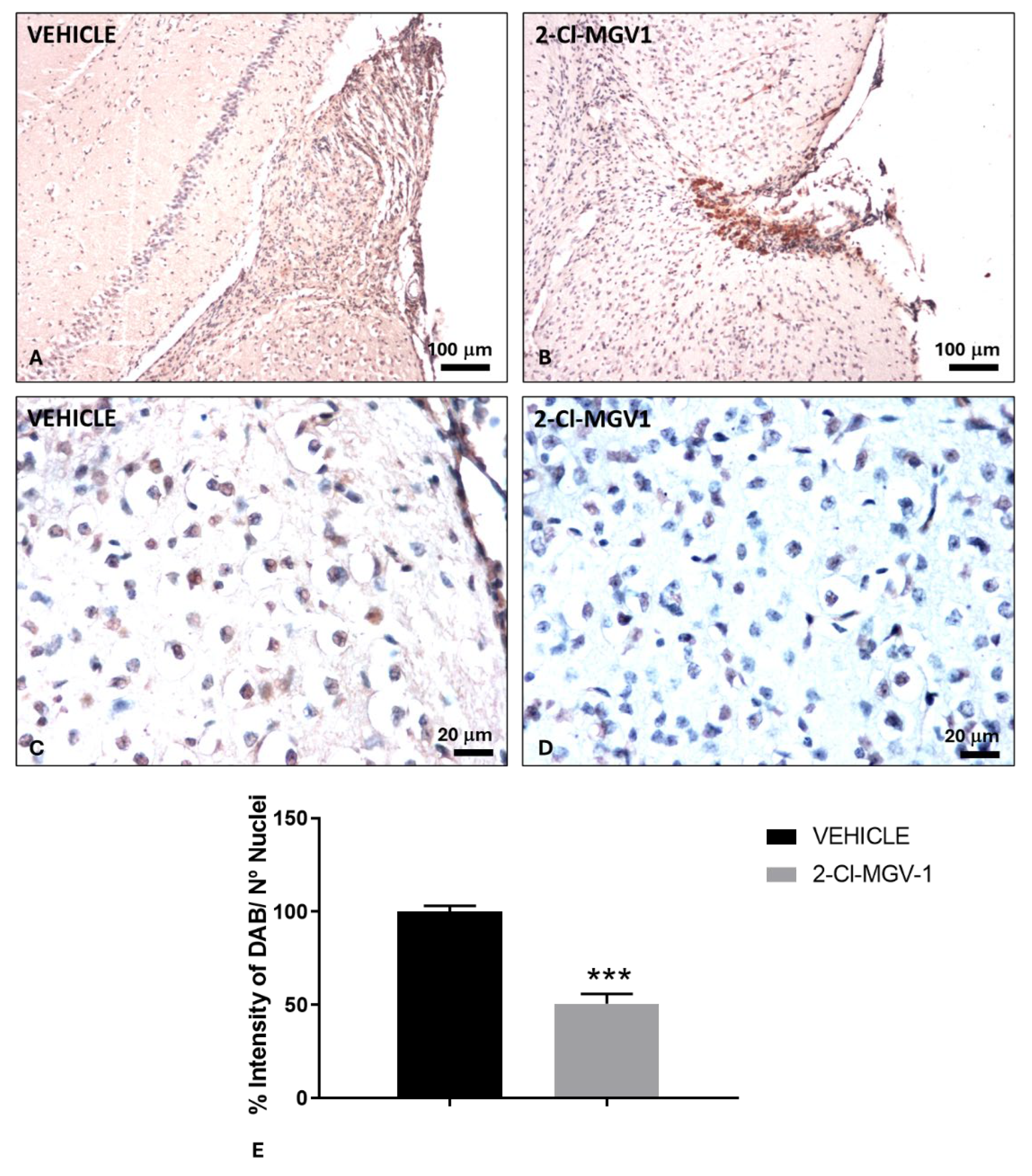
Figure 3.
Neurogenesis markers in the SVZ of TBI mice following 2-Cl-MGV-1 treatment compared to vehicle. Immunohistochemical staining of TSPO (A-C), Nestin (D-F), NeuroD1 (G-I) and β-Tubulin III (J-L) in the SVZ of TBI mice. (A, D, G, J) Brain sections from TBI mice treated with vehicle (DMSO) for 60 days. (B, E, H, K) Brain sections from TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days. Bar charts showing quantification of TSPO (C), Nestin (F), NeuroD1 (I) and β-Tubulin III (L) levels, expressed as the percentage of DAB signal relative to the number of nuclei. Images were captured at 400x magnification. Scale bar = 20 μm. Data is represented as mean ± SEM. ***p< 0.001, unpaired T-Test.
Figure 3.
Neurogenesis markers in the SVZ of TBI mice following 2-Cl-MGV-1 treatment compared to vehicle. Immunohistochemical staining of TSPO (A-C), Nestin (D-F), NeuroD1 (G-I) and β-Tubulin III (J-L) in the SVZ of TBI mice. (A, D, G, J) Brain sections from TBI mice treated with vehicle (DMSO) for 60 days. (B, E, H, K) Brain sections from TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days. Bar charts showing quantification of TSPO (C), Nestin (F), NeuroD1 (I) and β-Tubulin III (L) levels, expressed as the percentage of DAB signal relative to the number of nuclei. Images were captured at 400x magnification. Scale bar = 20 μm. Data is represented as mean ± SEM. ***p< 0.001, unpaired T-Test.
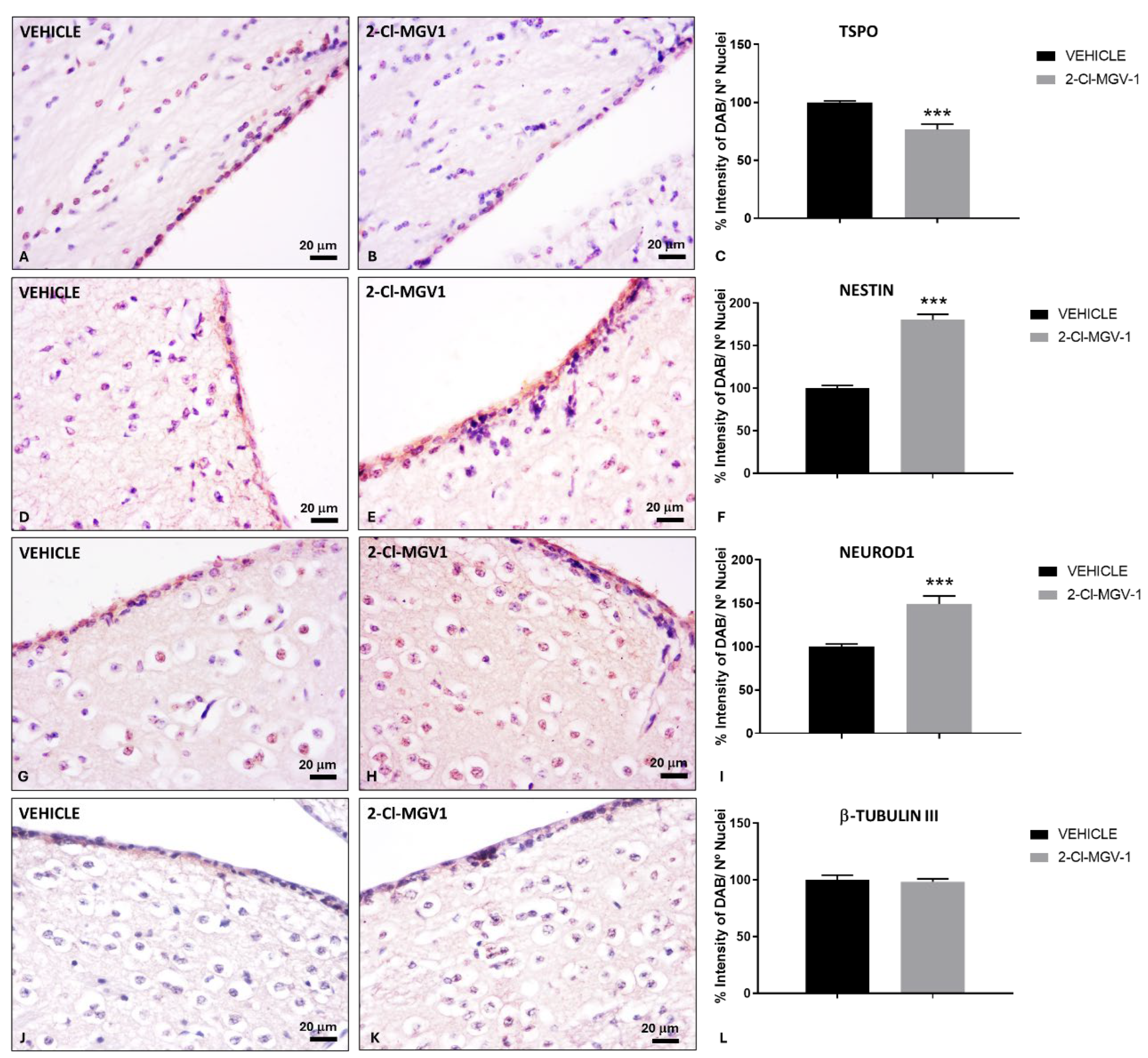
Figure 4.
Increased accumulation of neuronal precursor cells in the injured cortex of TBI mice following 2-Cl-MGV-1 treatment compared to vehicle (A) Representative immunohistochemical staining of NeuroD1 in the brain cortex of TBI mice treated with vehicle (DMSO) for 60 days. (B) Representative immunohistochemical staining of NeuroD1 in the brain cortex of TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days. (C) Bar graph showing quantification of NeuroD1 levels, expressed as the percentage of DAB intensity relative to the number of nuclei. Microscopy images were captured at 400x magnification. Scale bar = 20 μm. Data are represented as mean ± SEM. **p < 0.01, unpaired t-test.
Figure 4.
Increased accumulation of neuronal precursor cells in the injured cortex of TBI mice following 2-Cl-MGV-1 treatment compared to vehicle (A) Representative immunohistochemical staining of NeuroD1 in the brain cortex of TBI mice treated with vehicle (DMSO) for 60 days. (B) Representative immunohistochemical staining of NeuroD1 in the brain cortex of TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days. (C) Bar graph showing quantification of NeuroD1 levels, expressed as the percentage of DAB intensity relative to the number of nuclei. Microscopy images were captured at 400x magnification. Scale bar = 20 μm. Data are represented as mean ± SEM. **p < 0.01, unpaired t-test.
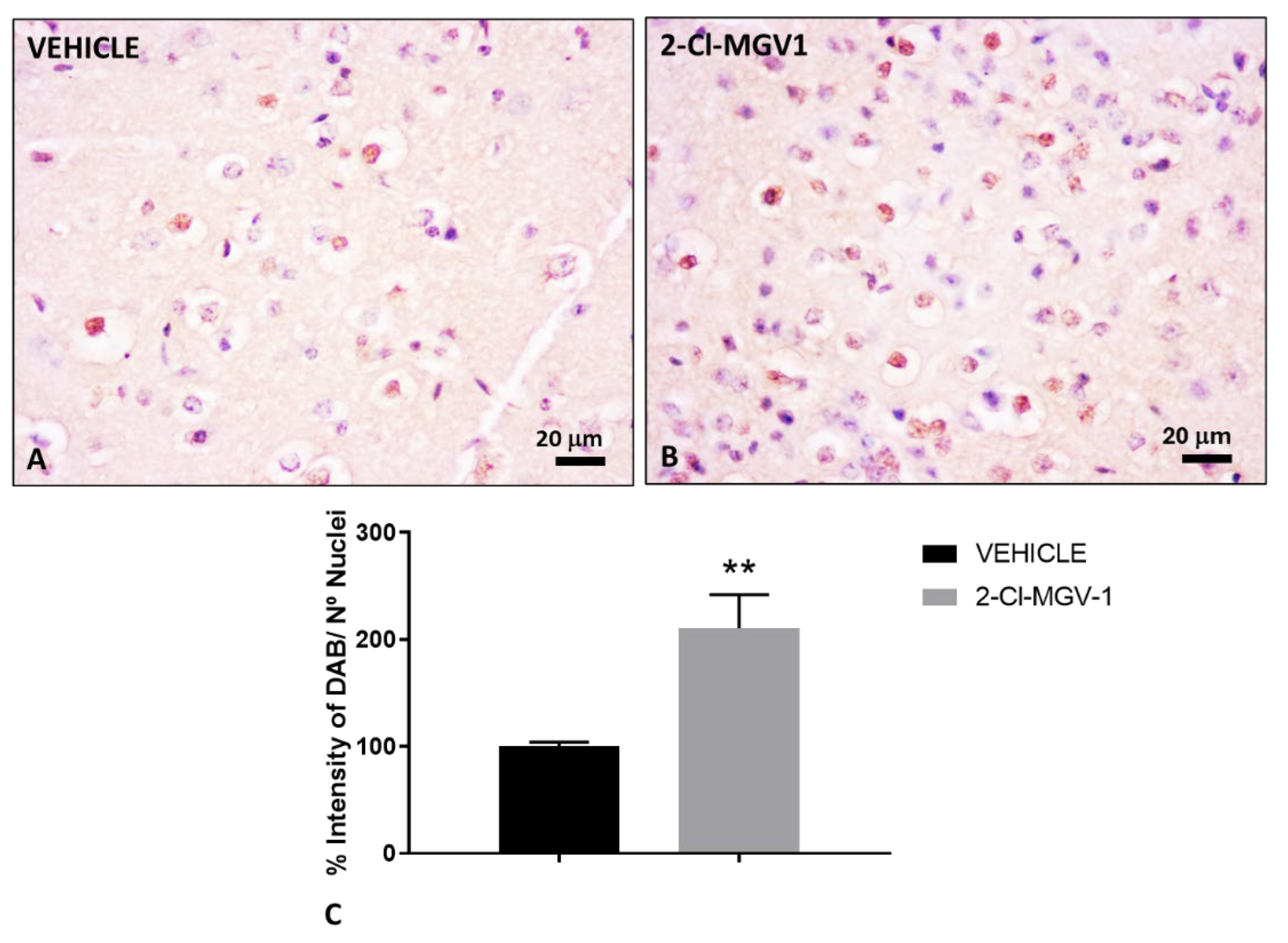
Figure 5.
Treatment with 2-Cl-MGV-1 reduces the accumulation of neuronal precursor cells in the olfactory bulb of TBI mice compared to vehicle (A, C) Representative images of NeuroD1 immunohistochemical staining in the olfactory bulb of TBI mice treated with vehicle (DMSO) for 60 days. (B, D) Representative images of NeuroD1 immunohistochemical staining in the olfactory bulb of TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days. (E) Bar graph showing quantification of NeuroD1 levels, expressed as the percentage of DAB intensity relative to the number of nuclei in the mitral cell layer (ML) and granule cell layer (GCL) of the olfactory bulb. Microscopy photos were captured at 400x magnification. Scale bar = 20 μm. Data is represented as mean ± SEM. ***p< 0.001, unpaired T-Test. GL, Glomerular layer. Arrows indicate the presence of neuronal precursor cells or neuroblasts.
Figure 5.
Treatment with 2-Cl-MGV-1 reduces the accumulation of neuronal precursor cells in the olfactory bulb of TBI mice compared to vehicle (A, C) Representative images of NeuroD1 immunohistochemical staining in the olfactory bulb of TBI mice treated with vehicle (DMSO) for 60 days. (B, D) Representative images of NeuroD1 immunohistochemical staining in the olfactory bulb of TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days. (E) Bar graph showing quantification of NeuroD1 levels, expressed as the percentage of DAB intensity relative to the number of nuclei in the mitral cell layer (ML) and granule cell layer (GCL) of the olfactory bulb. Microscopy photos were captured at 400x magnification. Scale bar = 20 μm. Data is represented as mean ± SEM. ***p< 0.001, unpaired T-Test. GL, Glomerular layer. Arrows indicate the presence of neuronal precursor cells or neuroblasts.
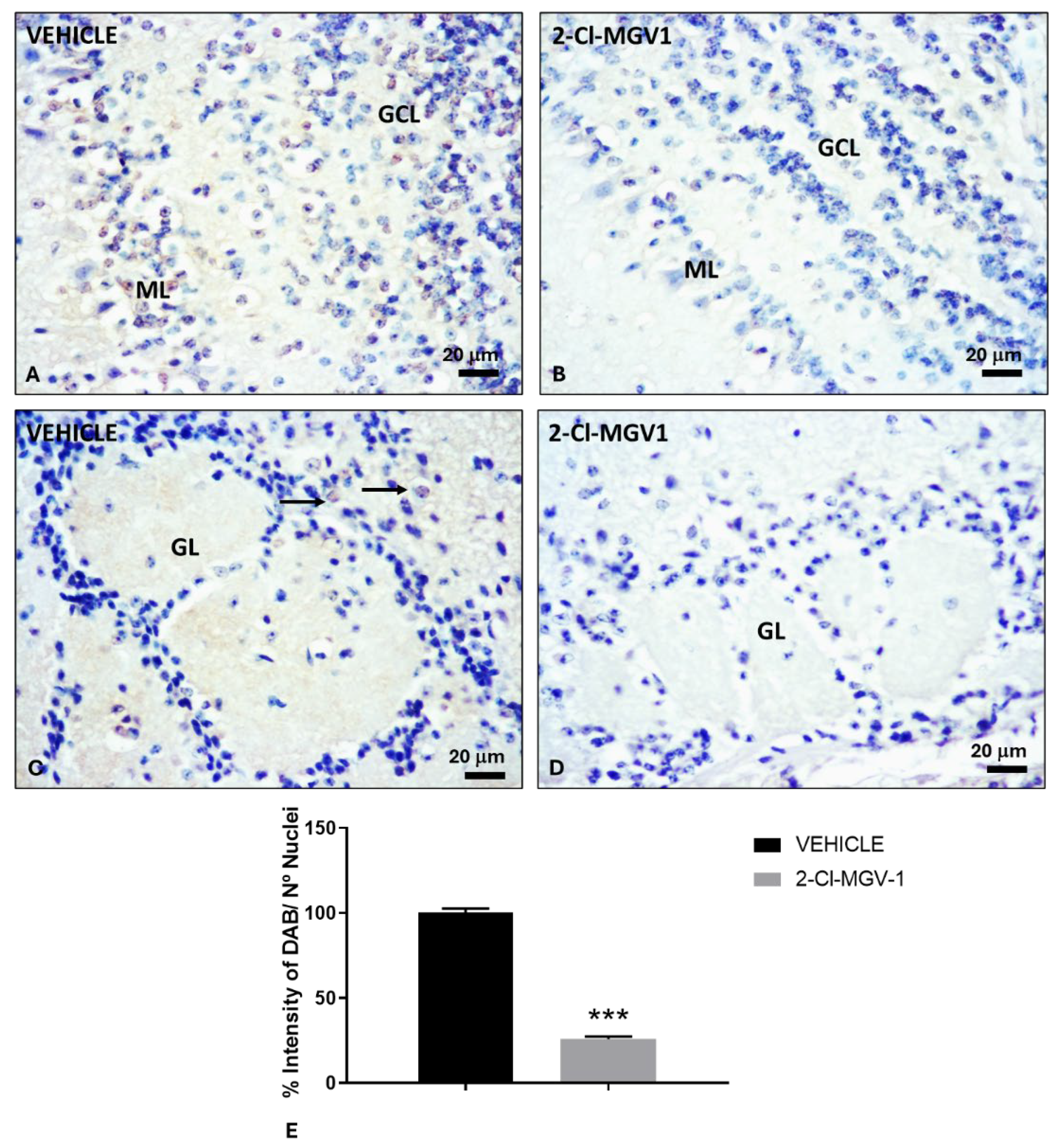
Figure 6.
Reduction of neurogenesis markers in the hippocampal dentate gyrus of TBI mice treated with 2-Cl-MGV-1 compared to vehicle. Immunohistochemical staining of TSPO (A-C), Nestin (D-F), NeuroD1 (G-I) and β-Tubulin III (J-L) in the hippocampal dentate gyrus. (A, D, G, J) Brain sections from TBI mice treated with vehicle (DMSO) for 60 days. (B, E, H, K) Brain sections from TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days. Bar graphs show quantification of TSPO (C), Nestin (F), NeuroD1 (I) and β-Tubulin III (L) levels, expressed as the percentage of DAB intensity relative to the number of nuclei. Microscopy photos were captured at 400x magnification. Scale bar =20 μm. Data is represented as mean ± SEM. ***p< 0.001, unpaired T-Test.
Figure 6.
Reduction of neurogenesis markers in the hippocampal dentate gyrus of TBI mice treated with 2-Cl-MGV-1 compared to vehicle. Immunohistochemical staining of TSPO (A-C), Nestin (D-F), NeuroD1 (G-I) and β-Tubulin III (J-L) in the hippocampal dentate gyrus. (A, D, G, J) Brain sections from TBI mice treated with vehicle (DMSO) for 60 days. (B, E, H, K) Brain sections from TBI mice treated with 7.5 mg/kg of 2-Cl-MGV-1 for 60 days. Bar graphs show quantification of TSPO (C), Nestin (F), NeuroD1 (I) and β-Tubulin III (L) levels, expressed as the percentage of DAB intensity relative to the number of nuclei. Microscopy photos were captured at 400x magnification. Scale bar =20 μm. Data is represented as mean ± SEM. ***p< 0.001, unpaired T-Test.
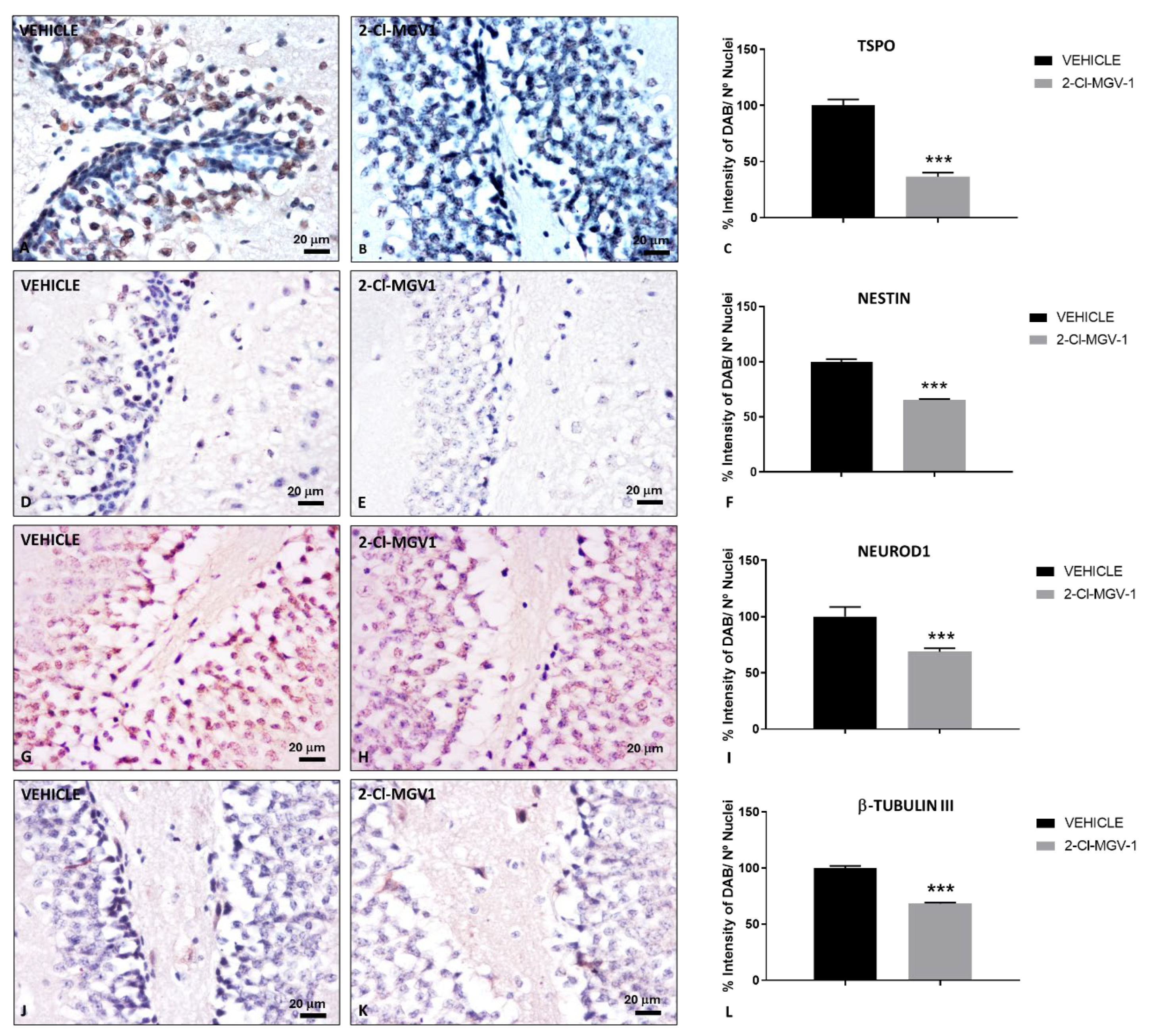

Table 1.
Primary antibodies for immunohistochemistry.
| Antibody | Antigen | Catalog Nº | Company |
|---|---|---|---|
| Anti-TSPO | The 18-kDa translocator protein | PA5-75544 | Invitrogen |
| Anti-Nestin | Neuroectodermal stem cell protein | N5413 | Sigma-Aldrich |
| Anti-NeuroD1 | Neurogenic differentiation factor 1 | #ABE991 | Millipore |
| Anti-TUBB3 | β-Tubulin III | T2200 | Sigma-Aldrich |
| Anti-NeuN | Neuronal nuclear antigen | AB4301175 | Sigma-Aldrich |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



