
Submitted:
17 June 2025
Posted:
18 June 2025
You are already at the latest version
Abstract
The exponential demand for plant–based meat analogues (PBMAs) can be attributed to the amassing need for sustainable and restorative proteins in the perspective of global food security. However, the development of meat analogues requires suitable physicochemical and techno–functional properties that effectively replicate the appearance, texture, sensorial characteristics, and dietary profile of conventional animal meat. In addition, consumers seek organic, ethical, clean–label, and eco–friendly products that address challenges in the current meat market. This comprehensive review provides updated information about novel protein sources from legumes, oilseeds, cereals, pseudocereals, mushrooms, fungi, and algae species; key ingredients such as binding agents, colorants, flavorants, and chemical constituents to improve the functionality of PBMAs; and technological processing strategies that enhance the microstructural organization of meat substitutes. Moreover, the nutritional composition of phyto–proteins and mycoproteins has been compiled, as understanding the fundamental role of each ingredient in PBMAs is crucial for its formulation. Future research should be directed towards improving hybrid protein–protein interactions, fiber alignment, palate diversification, and thermomechanical techniques that encourage overall health benefits. Strategies for incorporating preservatives to facilitate longer shelf–lives and feasibility for large–scale production should be further explored to drive innovation in the meat analogues industry.
Keywords:
Plant–based Meat Analogues (PBMAs)
; meat substitutes
; GHG emissions
; plant proteins
; mycoproteins
; Texturized Vegetable Proteins (TVPs)
; functional ingredients
; food additives
; nutritional profile
; extrusion technology
1. Introduction
The projected global population is predicted to reach a cumulative 9.7 billion by 2050 according to the United Nations Food and Agriculture Organization [1]. Rapid urbanization and demographic expansion over the past decades have resulted in an increasing demand of approximately 38 million tons for the production of protein–rich meat comprising high nutritional content [12]. However, meat production on an industrial scale level causes major issues such as abundant usage of arable lands, pastures, and water resources; adverse effects faced by aquatic ecosystems, and environmental fluctuations due to greenhouse gases (GHGs) emissions into the atmosphere [3]. While human activities contribute to 51% of the total GHGs, i.e., carbon dioxide (CO2), nitrous oxide (N2O), methane (CH4), and ammonia (NH3), which lead to acidification of ecosystems [4]; livestock production alone is responsible for 1/4th of the world’s emissions [5]. The expected 70% increase in food production in conjunction with the current economic trends for meat intake may lead to the disproportionate availability of feed supplies [6]. Moreover, red and processed meat have been classified as potentially carcinogenic and group–1 carcinogen respectively, by the International Agency for Research on Cancer (IARC) of the World Health Organization (WHO) [7]. Meat consumption has also been associated with detrimental consequences on health such as cardiovascular diseases, type–II diabetes, obesity, colorectal cancers, zoonoses, airborne illnesses, and epidemic outbreaks [8,9]. This raises a necessity for identifying sustainable and affordable alternatives that can fulfil the needs of the growing population without harmful repercussions.
Plant–based meat analogues incorporating meat–like characteristics, have emerged as suitable substitutes that confer benefits in terms of both health and ecological impact, while addressing the limitations of real meat. Proteins obtained from plants are most favored by consumers who are inclined towards minimizing the consumption of animal–derived commodities due to ethical concerns for animal welfare. Meat analogues possess substantial physicochemical characteristics that provide a variety of options for individuals with specific dietary preferences such as vegans, vegetarians, and flexitarians [9,10]. With the steadily growing number of plant–based meat consumers, the global meat substitutes market size is expected to grow tremendously, with a predicted increase of USD 74–263 billion by the year 2030 [11]. Presently, food scientists and industrial experts are exploring techniques that mimic the taste, aroma, texture, and structure of original animal tissue constructs. There are two main types of meat analogues which are being actively studied by the food research community, i.e., lab–grown tissue–engineered meat from cell cultures in contrast to whole animals or organisms, and imitation meat such as plant–based meat or mycoprotein–based meat from ascomycetes or basidiomycetes to improve the juiciness, chewiness, and appearance of analogues [8].
Plant–based proteins primarily derived from legumes, oilseeds, cereals, and pseudocereals have been widely proposed to replace meat as they form a nutritionally balanced diet with sustainable and well–preserved functionality [9]. Similarly, textural and sensorial properties are also important in obtaining realistic meat replications with desired characteristics. The fibrous matrix of spun protein filaments and texturized vegetable proteins (TVPs) constituting plant–based analogues have specific attributes that closely imitate the myofibrillar assembly of meat, thereby contributing to organoleptic structures [12]. Mycoprotein obtained from the mycelia of filamentous fungi has also been used in meat analogues as it is involved in efficient conversion of nutrients and carbohydrate from the growth media with natural meat–like texture, flavors, protein–rich profile, and inherent nutraceutical compounds. The latest industrial advancements utilizing mushroom–based meat analogues also coincide with the United Nation’s Sustainable Development Goals (SDGs) [2]. Additionally, it is known for enhancing digestibility, inducing satiety, synthesizing muscle proteins, and improving lipoprotein profiles due to its rich amino acid constituents and poly–unsaturated fatty acids (PUFAs) [4].
The inclusion of hydrocolloids in meat analogue formulations has been known to improve binding between protein filaments, thus promoting textural properties. Hydrocolloids enhance the fibrous texture of meat analogues, especially when subjected to high moisture extrusion (HME) and high–temperature shear cell (HTSC) processing [13]. Coagulation, texturization, and extrusion of proteins are some of the currently available methods that are used to produce fibrillar oriented structures [14]. Thermomechanical processing (TMP) encompasses these techniques to create textured meat–like arrangements of dry and wet extrudates such as low–moisture (10% – 30%) meat analogues (LMMA) which have a spongy fibrous organization with non–uniform air pockets, and high–moisture (40% – 70%) meat analogues (HMMA) which have well–defined uniform fibrous texture resembling real meat [15].
The main objective of this study is to identify up–to–date advancements in meat analogues utilizing various protein from alternative sources, with a particular focus on their application, combination with texturizing and sensorial agents, incorporation into food substitutes, end–product quality, and implemented technology. This in–depth analysis provides a comprehensive overview of plant–based proteins, oilseeds, cereal and pseudocereal proteins, mushroom–based proteins, algal species, and fungal biomasses; in combination with binding, coloring, chemical, and flavoring agents using extrusion technology, 3D/4D food bioprinting, electrospinning, antisolvent precipitation, and mechanical elongation techniques. The resultant meat analogues were assessed for their physicochemical properties, textural and sensory characteristics, and nutritional amino acid profiles to understand the current established research agenda in shaping consumer acceptance of commercial plant–based meat substitutes.
Figure 1.
Plant–based meat analogues formulation strategies and nutritional composition.
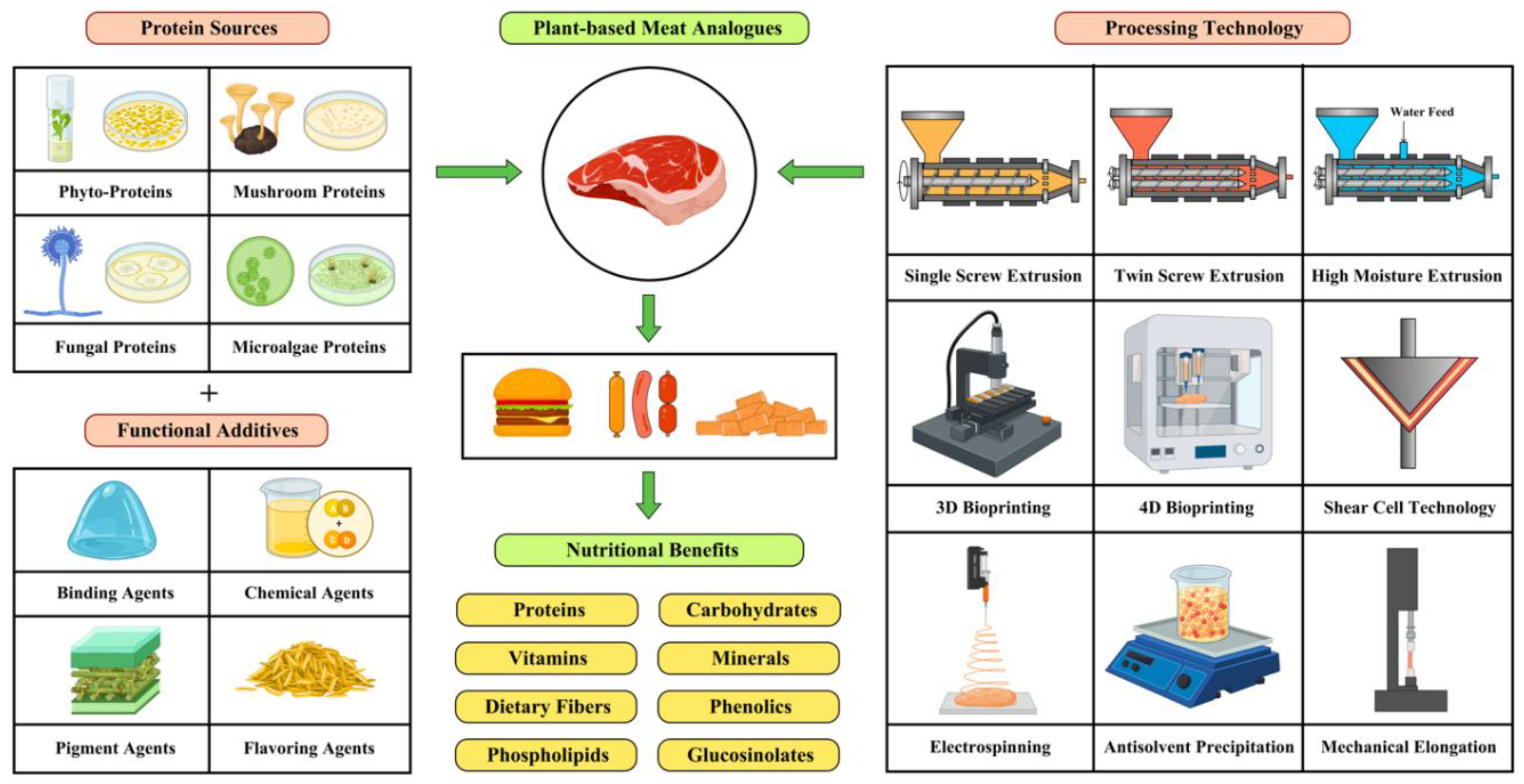
2. Plant Protein Sources
2.1. Proteins from Legumes
2.1.1. Soy Protein
Soy proteins are most commonly used in meat analogues as soy protein isolates (SPI), soy protein concentrate (SPC), defatted soy flour (DSF) [16], and full–fat soy (FFS) forms [17,18]; with the highest protein content (> 90%) and moisture retention found in SPI, making it optimal for HME [19]. The nutritional profile of FFS is constituted by 38–40% protein, 33% carbohydrate, and 18–22% fat with presence of dietary fibers [20,21]. FFS is commonly combined with gluten protein to compensate for its lower protein content compared to other soy forms, contributing towards higher texturization; and imparts a suitable color which improves the appearance of meat analogues [18]. Lin et al. (2025) studied the relationship between 0–12% SPI with four texturized soy proteins (TSPs). TSPs at higher levels revealed an enhanced water binding effect and reduced cooking loss, signifying the importance of texturization in hybridized meat production [22]. Soy proteins also have characteristic functional properties such as gelling, emulsification, lipid–absorbing, and water holding capacities [23]. Furthermore, soy proteins include a good amount of essential amino acids (EAAs) with hydrophilic and hydrophobic properties that facilitate fibrous 3D interlaced matrix formation, along with a Protein Digestibility Corrected Amino Acid Score (PDCAAS) relatively equivalent to that of animal proteins [24]. Soy proteins have high shearing viscosity leading to deformation, alignment, and structural entrapment [25].
2.1.2. Pea Protein
Pea proteins are an alternative source to soy protein with an exceptional EAAs profile, especially lysine and threonine [26]. Pea proteins are listed as non–allergens with a low glycemic index and less gelling capacity than soy proteins [27,28,29]. While pea proteins are largely affordable, has less association to GMO–related issues, and can be cultivated in moderate climate conditions, there are challenges during structurization as heat–induced SPI gels have higher strength when compared to pea protein isolate (PPI) gels [30,31]. A few scientific studies have reported successful PPI structurization through low and high moisture extrusion techniques by the inclusion of wheat gluten (WG) as structural support or hydrocolloids such as cellulose and pectin fibers, which provided viscoelastic strength and anisotropy by matrix elongation [25,32]. Sajib et al. (2023) evaluated the role of isolation temperature and pH levels in PPI–based PBMAs. The study revealed that pH increased from 2.0 to 11.0 resulted in improved yield rates from 37% to 79% at 20°C, with higher lipid absorption, hardness, cohesiveness, chewiness, and gumminess due to protein denaturation and surface hydrophobicity. Similarly, when temperature was augmented from 20°C to 40°C at fixed pH, the solubility, foaming, and gelation capacities were negatively impacted [33]. Pea proteins also play a vital role in preventing leukemia, cancers (breast, lung, colon, pancreas, prostate), cardiovascular diseases, type–II diabetes, colitis, and promoting muscle thickness and gut microbiota, making it a viable alternative to animal meat [34,35].
2.1.3. Mung Bean Protein
Mung bean protein, also known as “green pearl”, is comprised of 25–28% proteins and 1–2% fat content with a nutritional amino acid profile akin to that of soy proteins [36,37]. Furthermore, Mung bean protein isolate (MBPI) has a Digestible Indispensable Amino Acid Score (DIAAS) of 86 which is intermediate to SPIs and PPIs that have a score of 91 and 70 respectively [38,39,40]. Mung bean proteins also improve the texture, gelling, foaming, and emulsification properties of meat analogues as they are made up of 25% albumin and 60% globular proteins [41]. Guo et al. (2024) identified that a 7:3 ratio of MBPI:WG low–moisture extrudates improved protein cross–linking, and a 6:4 ratio of MBPI:WG unfavorably affected the extrudate structures [42]. In another study, Angelis et al. (2023) studied the rheological properties of dry–fractionated MBPIs with 55% protein content via high moisture extrusion cooking (HMEC) with varying moisture levels (40–50%) and temperature (40–170°C). At 70°C, higher complex shear modulus was observed due to starch and MBPI gelation, indicating that higher temperatures have a predominant effect on structural anisotropy, hardness, viscosity, and cohesiveness of the extrudates [43]. Mung bean proteins also have hypoglycemic, immunomodulatory, anti-melanogenic, hepatoprotective, and detoxification properties, offering a nutritionally balanced sustainable source in replacement of animal–based proteins [37,44,45].
2.1.4. Faba Bean Protein
Faba bean protein is a novel alternative protein source which is currently being explored for its protein–rich and diverse properties such as atmospheric nitrogen fixation which is advantageous for the environment [46]. Faba beans provide a favorable source of lysine and can be cultivated in extreme climate conditions, making it easily accessible and sustainable for plant–based meat substitutes [47]. Faba bean proteins have a balanced nutritional composition with 20–41% proteins, 7–9% dietary fibers, 1–2% fat, polyphenols, and other minor compounds [48]. They also possess phenolic acids, flavonoids with antioxidant properties that regulate oxidative stress, inflammation, and prevent degenerative conditions such as Parkinson’s disease [49,50]. Faba beans have exceptional emulsion stabilization and foaming abilities, although to a lesser degree than soy proteins [51]. Fan et al. (2025) investigated the role of HME with 0–6% brewers’ spent grain (BSG) and faba bean proteins, where enhanced fibrous texture was obtained at 3% BSG [52]. Furthermore, Elshamy et al. (2025) studied how dry–fractionated faba bean (FB), yellow pea (YP), and wet–fractionated soybean (SB) protein concentrates influence meat–like texture. Total moisture content (TMC) affected the chewiness, hardness, and cutting strength of PBMAs, with longitudinal fiber orientation at higher temperatures [53]. Thereby, faba bean proteins can be used to produce PBMAs by dry fractionation, wet spinning, and thermomechanical processing [54,55].
2.1.5. Lupin Bean Protein
Lupin proteins are a largely cultivated species of legumes that can be grown in various land conditions with involvement in atmospheric nitrogen fixation and soil phosphate absorption [56]. Lupin protein has one of the highest nutritional amino acid profiles among legumes with 46% total protein content, non–digestible carbohydrates, dietary fibers, mineral bioavailability, and low level of alkaloids [57]. Lupin species such as white, yellow, blue/ narrow–leafed, and pearl lupin have been reported as feasible plant proteins for PBMAs [58,59]. Lupin proteins are also known for their low glycemic index, reducing low–density lipoproteins (LDLs), Trinitrobenzene sulfonic acid (TNBS)–induced colitis inflammation, boosting gastrointestinal function, antioxidant, anti–inflammatory, non–allergenicity, and hypocholesterolemic activities [60,61,62]. Ramos–Diaz et al. (2023) studied different lupin protein blend ratios (30:70, 50:50, and 70:30) using lupin protein isolates (LPI), lupin protein concentrates (LPC), and native lupin flour (LF). The texturized meat analogues (TMAs) with 30% d.m. LF retained more than 50% d.m. protein content, exhibiting fiber formation, and the lesser lupin protein isolate (LPI) content resulted in higher homogeneity [63]. Ayalew et al. (2024) explored the effect of processing temperatures (raw, 130–150°C) and immersion time (raw, 2–6 days). Highest mineral content, protein content, cooking yield, and sensory acceptability were observed at 140°C for a soaking period of 4 days. This study revealed that temperature and soaking conditions have a crucial role in the anti–nutritional attributes of PBMAs [64].
2.1.6. Chickpea Protein
Chickpea protein is the most consumed species of legumes, defined by its characteristic texture, water and oil binding capacity, fat absorbability, emulsion stabilization, solubility, foaming and gelling capacities comparable to that of soy protein isolates (SPI) and whey proteins [65]. The presence of carotenoids in chickpeas is also known for enhancing the color of meat analogues [66]. Chickpea proteins are composed of 12–31% proteins, 58–68% carbohydrates, 4–10% fat, 3–5% dietary fibers, and 10% water [67,68]. Primary processing strategies i.e., germination, dehulling, fermentation, and enzymatic catalyzation have been identified to improve the properties of chickpea proteins together with several extraction methods [69]. These techniques include dry and wet fractionation such as isoelectric precipitation, membrane filtration, air classification, and alkaline/acid/salt extraction to obtain concentrate and isolate forms [70,71,72]. Ma et al. (2023) explored high–pressure homogenization (HPH: 0–150 MPa) at 1–3 cycles on the physicochemical attributes of chickpea proteins, where increased surface hydrophobicity and decreased sulfhydryl content was observed [73]. In another study, Kong et al. (2023) functionalized chickpea protein hydrolysates with trypsin to support muscle and fat cell adhesion, proliferation, and differentiation in cultured meat [74]. Furthermore, Czapalay et al. (2025) investigated how gelling agents can compensate for the absence of adipose tissues, by combining 6% pea starch with 4% chickpea flour and 40% oil at 5–85°C temperature profiles, forming a starch network with adipocyte–like oil pockets [75].

Table 1.
Leguminous Protein Sources for Plant–based Meat Analogues.
| Legume Proteins | Latin Name | Properties | Reference |
|---|---|---|---|
| Soy | Glycine max | Excellent water–holding, emulsification, strong gelling, good texturization. | [76] |
| Pea | Pisum sativum | High emulsifying and water–holding, moderate foaming and gelling stability. | [77] |
| Mung bean | Vigna radiata | Extrudability and texturization, gelling and foaming capacity under medium shear force. | [78] |
| Faba bean | Vicia faba | Good emulsification, foaming, and gelling, improves microstructural properties. | [79] |
| Lupin bean | Lupinus | Good emulsification, foaming, and water–binding, enhances firmness. | [80] |
| Chickpea | Cicer arietinum | Strong water and oil binding, moderate gelation, creates a soft texture. | [81] |
| Kidney bean | Phaseolus vulgaris | Moderate water–holding and gelation; improves texture and elasticity. | [82] |
| Lentil | Lens culinaris | Good water–binding, moderate gelation, emulsification, improves firmness and chewiness. | [83] |
2.2. Proteins from Oilseeds
2.2.1. Hemp Protein
Industrial hemp seeds cultivated locally as a byproduct raw material is nutritionally rich with 20–26% proteins, 25–35% unsaturated fats, >50% linoleic acid, 10–15% dietary fibers, 27–29% carbohydrates, vitamins, and minerals [84,85]. Hemp seeds comprising albumins and globulins, are commonly available in raw, roasted, and oil pressed forms [86]. They serve as a reliable source of nourishment with reduced thyroid hormones, tri–acylglycerol, antioxidant, antihypertensive, hypocholesterolemic, hypoglycemic and neuroprotective effects [87,88]. The essential amino acids (EAAs) in hemp protein are comparable to egg, casein, and soy proteins with an Arg/Lys ratio of 3.78–5.34 [89]. Moreover, processing conditions such as membrane ultrafiltration, alkaline extraction, isoelectric precipitation, and micellization influence the nutritional profile of hemp seeds [90]. Zahari et al. (2023) developed high moisture meat analogues (HMMAs) with hemp seed protein concentrate (HPC) at 60–66% moisture content, 500–900 rpm screw speeds, and 40–150°C temperature profiles. The meat analogue exhibited stronger fibrous structures, texturization, and cutting strength at higher temperatures [91]. Nasrollahzadeh et al. (2022) investigated the role of dry and wet fractionation using five different HPCs. In dry extraction, HPCs maintained their native oligomeric structure with high surface hydrophobicity, solubility, anisotropy, and low gelation. In wet extraction, HPCs had higher bound polyphenols, anisotropy, and viscosity [92]. Thus, hemp seed proteins offer both functionality and viscoelastic properties in plant meat alternatives.
2.2.2. Rapeseed Protein
Rapeseed proteins have tremendous agronomic value owing to the presence of omega–3, omega–6, and unsaturated fatty acids, vitamins, phospholipids, phenolic compounds, tocopherols, sterols, and carotenoids [93]. Rapeseed proteins also have low erucic acid content, overcoming the major limitation of erucic toxicity in hemp proteins which cause myocardial damage and fat body parenchyma [94]. Rapeseed proteins composed of 20% napins (2S albumins) and 60% cruciferins, (12S globulins) are non–allergenic, and have been largely used in bakeries, beverages, flavorings, processed meat products, and dairy industries [95]. Some of the notable bioactivities of rapeseed proteins include Angiotensin–converting enzyme (ACE) inhibition, bile acid binding capacity, immunogenicity, cell growth, cardiovascular regulation, and antithrombotic properties [96,97]. Jia et al. (2021) analyzed the potential of rapeseed protein concentrates (RPC) in meat analogues via shear cell technology. Fibrous textures with 40% d.m. were observed at 140°C and 150°C. Increased WG content in RPCs also improved the color profile, indicating their significance as an optimal protein source after soy protein concentrates (SPC) [98]. In another study, Zhang et al. (2024) investigated the substitution capacity of rapeseed proteins to soy proteins under extrusion cooking technology at 0–50% and 50–0% (w/w) concentrations. 0–20% rapeseed proteins enhanced the hardness and chewiness of extruded analogues. At higher substitution levels, lower expansion, rehydration, and brightness were observed with increased protein stability [99].
2.2.3. Pumpkin Protein
Pumpkin proteins rich in poly-unsaturated fatty acids (PUFAs) have useful nutraceutical compounds with proteins, lipids, essential and non–essential fatty acids. Natural bioactive compounds in pumpkin seeds including flavonoids, carotenoids, and squalene make it a vital component in diverse pharmacological and food sectors [100]. Pumpkin seed proteins have copious amounts of macronutrients and micronutrients which prevent cardiovascular diseases, cancers, deficiency disorders, and protein malnutrition. Pumpkin seeds are also used to treat benign prostate hyperplasia (BHP) and deter the risk of Alzheimer’s and Parkinson’s diseases [101,102]. Pumpkin proteins are composed of 25–37% proteins, 18–25% carbohydrates, 37–45% lipids, and 3–6% dietary fibers [103]. Pumpkin seeds in meat analogues contribute to a savory flavor and chewy texture, as protein materials and oil–based food additives [104]. Choi et al. (2025) prepared high–moisture meat analogues (HMMAs) using SPI and pumpkin protein concentrate (PSC) blends. Increased PSC content resulted in SPI network disruption, low bonding, and alignment, with modified texture and soft gel formation crucial for replicating original meat structures [105]. Furthermore, Kong et al. (2025) prepared gelatin hydrogels using alginate (ALG) combined with PSP for plant–based food 3D–printing applications. Optimal gelation texture, hardness, springiness, and cohesiveness were observed at 2–3% ALG and 2–6% PSP with color resemblance like animal gelatin [106].
Table 2.
Oilseed Protein Sources for Plant–based Meat Analogues.
| Oilseed Proteins | Latin Name | Properties | Reference |
|---|---|---|---|
| Peanut | Arachis hypogaea | Oil binding and foaming capacity comparable to SPI; higher viscosity and gel formation post–heating. | [107] |
| Hemp | Cannabis sativa | Superior functional attributes such as foaming, gel formation, and WHC. | [108] |
| Rapeseed | Brassica napus | Strong heat–set gel forms by cruciferin under alkaline conditions. | [109] |
| Pumpkin | Cucurbita maxima | High emulsifying capacity and stability, moderate foaming, low gelling, and high WHC. | [110] |
| Flaxseed | Linum usitatissimum | High nutritional value and good techno–functional properties i.e., solubility, foaming, emulsification, gelling, and WHC. | [111] |
| Chia | Salvia hispanica | Excellent water and oil binding properties; rich in plant–based omega–3 fatty acids. | [112] |
| Cottonseed | Gossypium hirsutum | Good emulsification and WHC; contains gossypol, which requires processing for safe consumption. | [113] |
| Sesame | Sesamum indicum | Excellent source of proteins and antioxidants; good emulsification and binding properties. | [114] |
| Safflower | Carthamus tinctorius | Rich in linoleic acid and good water retention aiding in texture formation. | [115] |
| Sunflower | Helianthus annuus | Stable emulsification and foaming with comparable gelling capacity. | [116] |
| Black Cumin | Nigella sativa | Flavor enhancement with its distinctive aroma. Good oil–holding capacity aids in juiciness and texture. | [117] |
2.3. Proteins from Cereal and Pseudocereal
2.3.1. Wheat Protein
Wheat protein has an elastic network structure formed by interchain cross–linking between disulfide bonds of prolamin and glutein residues via partial hydration [24]. Wheat gluten (WG), known for its high viscoelasticity, flexibility, thermal coagulation, and fibrosity, mainly consists of glutenins and gliadins, and acts as a plasticizer in meat analogues [118,119]. Moreover, the nutritional profile of wheat proteins is constituted by 11% proteins, 58% carbohydrates, 2% lipids, and a relatively unbalanced amino acid profile compensated by combining WG with other legume and starch–based proteins [120,121]. WG facilitates protein blends to form independent phases by increasing the content of β–sheets to create dense and compact fibrous structures [122]. It is largely available as binders, fillers, and extenders in vital, texturized, and isolated forms, to improve the overall color stability, juiciness and firmness [123]. While WG is an optimal non–meat ingredient, it is also an allergen which must be taken into consideration for plant–based products. Dai et al. (2024) identified that wheat gluten enzymatic hydrolysates (WGEHs) converted disulfide bonds to free sulfhydryl groups, improving extractability and texturization degrees [124]. Sun et al. (2023) experimented xylose–induced Maillard reactions products (MRPs) at 80–120°C using WGEHs prepared by Flavourzyme [125]. At 120°C, MRPs exhibited formation of intermediates, thermal degradation, protein cross–linking, high umami flavor and low bitterness in plant meat analogues due to the presence of volatile compounds i.e., furans and furanthiols. [126].
2.3.2. Oat Protein
Oat proteins are nutritionally rich plant–derived proteins that have high heat stability due to their globulin content. Oat proteins are made up of 15–20% proteins in whole kernels, carbohydrates, dietary soluble fibers, unsaturated fatty acids, EAAs, vitamins, and minerals [127,128]. Oat proteins contain 1–12% albumins, 50–80% globulins, ~10% glutelins, and 4–15% prolamins [129]. Texturized meat analogues can be produced using oat globulins for rigid and irreversible amyloids, or semi–flexible and reversible amyloid protein nanofibrils [130]. Oat protein isolates and concentrates can be used in meat analogues as they prevent syneresis. They also have a moderate Digestible Indispensable Amino Acid Score (DIAAS) value of 0.57, a higher PDCAAS value than wheat protein, a digestibility rate of 90.3–94.2%, and an efficiency ratio of 2.25–2.38 that is closely equivalent to casein [129]. Oat protein can be used as functional thickeners, emulsifiers, stabilizers, gelling agents, and texture modifiers owing to their foaming, lipid–binding, and WHC [131]. Chemical and enzymatic treatments can improve the functionality of oat proteins which undergo denaturation in liquid and semi–solid products [132]. Brückner–Gühmann et al. (2021) investigated heat–induced gels using oat protein isolate (OPI) with two oat protein hydrolysates at pH levels of 4.5 and 8 under 90°C and 120°C respectively. Trypsin (OPT) and Alcalase (OPA) influenced gelling properties at a 2% degree of hydrolysis with variations in protein aggregation, structure, and lightness [133].
2.3.3. Rice Protein
Rice protein is one of the largest globally–produced cereal crop, composed of 32–78% protein content, including 4–22% albumins, 5–13% globulins, 1–5% prolamins, and 60–80% glutelins [134]. It contains a rich reservoir of lipids, carbohydrates, dietary fibers, vitamins, minerals, and EAAs, depending upon its genotype and growth conditions [135]. Rice protein is renowned for its hypoallergenicity, antidiabetic, anticancer, and antioxidant nature due to its polyphenols, peptides, tocopherols, tocotrienols, γ–oryzanols, and flavonoids [136]. Rice proteins have a high digestibility of 93%, and protein efficiency of 2.02–2.04% [137]. Lee et al. (2022) utilized different substitution ratios of rice protein (RP) to soy protein (SP) ranging from 0–100% (w/w) through low–moisture extrusion cooking (LMEC). SP replacement with RP resulted in increased hardness and density with decreased porosity, expansion ratio, and textural profile. This was due to the short residence time of the dough in the extruder caused by its high flow rate, which led to lower specific mechanical energy (SME) input responsible for protein denaturation and texturization. [138]. Lee et al. (2022) also prepared texturized rice protein (TRP) meat analogues by LMEC using 25–100% and 75–0% RPI–SPI blend ratios, WG, and corn starch. RPI–SPI meat analogues exhibited higher nutritional quality than commercial PBMAs [139]. Similarly, Charlie et al. (2025) developed TVPs using rice bran (R), soy protein (S), and mung bean (M) with 30% moisture content by LMEC at 125°C. MR–TVPs exhibited superior performance in terms of textural integrity, microstructure, density, binding capacity, and rehydration ratio [140].
Table 3.
Cereal Protein Sources for Plant–based Meat Analogues.
| Cereal Proteins | Latin Name | Properties | Reference |
|---|---|---|---|
| Wheat | Triticum aestivum | High in gluten, providing elasticity and binding properties. | [141] |
| Oat | Avena sativa | Good solubility in water, high emulsifying, gelling, and WHC, moderate foaming capacity. | [142] |
| Rice | Oryza sativa | Good solubility in water, high emulsifying and stability, moderate foaming, and low gelling. | [143] |
| Corn (Zein) | Zea mays | Stabilizes oil–in–glycerol emulsion–gels, used as a fat analogue, and stabilizes foam and emulsions. | [144] |
| Barley | Hordeum vulgare | Contains β–glucans, contributing to water retention and gelation. Improves viscosity and mouthfeel. | [145] |
| Millet | Panicum miliaceum | Gluten–free and good water–binding capacity for improved texture. Rich in antioxidants and EAAs. | [146] |
| Sorghum | Sorghum bicolor | Polyphenols support texture and nutrition. Gluten–free good binding capacity for improved juiciness. | [147] |
Table 4.
Pseudocereal Protein Sources for Plant–based Meat Analogues.
| Pseudocereal Proteins | Latin Name | Properties | Reference |
|---|---|---|---|
| Quinoa | Chenopodium quinoa | Higher emulsifying capacity and stability than soy, wheat and pearl millet; low gelling capacity and good water holding capacity. | [148] |
| Amaranth | Amaranthus caudatus | Improves solubility, emulsification, foaming, gelling, and water–holding capabilities subject to pH, temperature, and enzymatic hydrolysis. | [149] |
| Buckwheat | Fagopyrum esculentum | Good water retention and strong emulsification capacity with gluten–free texture enhancement properties. Contributes to a slightly nutty flavor. | [150] |
2.4. Mushroom Species
2.4.1. Agaricus bisporus (Button Mushroom)
Agaricus bisporus (A. bisporus), also known as button or portobello mushroom, is a common edible mycoprotein species from the Agaricaceae family which makes up 35–45% of global mushroom production [151]. A. bisporus has organoleptic and nutraceutical properties with great market demand as it can be cultivated through bio–conversion techniques that convert cellulose into protein–rich biomasses [152]. A. bisporus is low in fat, calories, sodium, and cholesterol, with high levels of proteins, carbohydrates, phenolic compounds, and dietary fibers such as chitin, glucans, ergosterol [153]. Clinical findings have shown that A. bisporus can control weight gain levels, combat cognitive impairment, diabetes, and cancer by enabling natural killer (NK) cells to facilitate immunogenic activity [154]. Polysaccharides in A. bisporus demonstrate antioxidant, anti–inflammatory, anti-obesity, anticancer, immunogenic, and hepatoprotective bioactivities [155]. A. bisporus has notable aroma, taste, and texture influenced by approximately 67–150 volatile compounds and different water–soluble substances that enhance meat–like umami flavor [156]. Fu et al. (2023) partially substituted A. bisporus (AB) mushrooms in chicken breast at 0% (control) to 70% to evaluate its physicochemical and oxidative properties. Increased AB substitution resulted in increased moisture content and PUFAs, with decreased protein content, pH, color saturation, sensory characteristics, and delayed oxidation at 10% AB substitution [157].
2.4.2. Pleurotus ostreatus (Oyster Mushroom)
Pleurotus ostreatus (P. ostreatus), also known as hiratake or oyster mushroom, is a high–quality protein–rich mushroom species of the Pleurotaceae family comprising bioactive nutrients ideal for meat analogues [158]. P. ostreatus is an important nutritional source with 7.3–53.3% proteins, 50–60% carbohydrates, <4% fats, and dietary fibers such as chitin, glucans, cellulose, and hemicelluloses [159]. The vitamins present in P. ostreatus include thiamine, riboflavin, niacin, pyridoxine, retinol, ergocalciferol, and folates [160]. Moreover, P. ostreatus has notable levels of minerals, phenolic compounds, and a balanced EAA profile [161]. The dietary fibers in P. ostreatus have been found to improve physical properties involving texturization, gelation, thickening, emulsification, and stabilization [162]. P. ostreatus also has an exceptional flavor, taste, and chewy texture due to the presence of linoleic acid, palmitic acid, and oleic acid [163]. In addition, the interactions between proteins and polyphenol extracts enhances cross–linkage formation within extrudates [23]. Demircan et al. (2023) studied 3D–printed PBMA formulations using Ganoderma lucidum (GL) Lactarius deliciosus (LD), and Pleurotus ostreatus (PO) mushrooms. The resultant analogues demonstrated shear–thinning, viscoelasticity, and stable structures. P. ostreatus was involved in reducing hardness, stiffness, chewiness, redness, and beany flavors; while increasing the springiness, lightness, and re–printability [164,165].
2.4.3. Lentinus edodes (Shiitake Mushroom)
Lentinus edodes (L. edodes), also known as shiitake mushroom, is the second highest produced mushroom from the Marasmiaceae family, rich in aroma, umami, and phenolic compounds [166]. The fruiting bodies and mycelium extracts possess multifaceted therapeutic constituents such as β–glucans, terpenoids, sterols, flavonoids, and eritadenine responsible for antitumor, anticaries, antimicrobial, antioxidant, hepatoprotective, and immunomodulatory effects [167]. L. edodes is composed of 20–23% proteins, 58–60% carbohydrates, 9–10% dietary fibers, 3–4% lipids, and 4–5% ash content [167]. In particular, the presence of PUFAs moderate blood–cholesterol levels and cell physiological processes [168]. Santhapur et al. (2024) examined shiitake mushroom (SM) and oyster mushroom (OM) with whey protein isolate (WPI). SM had a negligible effect on thermal stability of heat–set WPI compared to OM. WPI exhibited thermal gelation and a higher isoelectric point. Furthermore, SM and OM reduced the hardness, stiffness, and lightness of the WPI gels, while increasing the intensity of its brown color [169]. Choi et al. (2025) investigated L. edodes mycelium (LEM) with different plant protein–based (soy, pea, mung, rice) and polysaccharide–based (agar, carrageenan, gellan gum) hydrocolloids. LEM–protein improved flowability and dispersion; whereas LEM–polysaccharide produced strengthened gel–like structures due to intermolecular β–sheet hydrogen bonding. High gel homogeneity and interconnectivity were observed in LEM–Ca, presenting the significance of L. edodes blends in developing liquid or solid–type PBMAs [170].
2.4.4. Coprinus comatus (Chicken Drumstick Mushroom)
Coprinus comatus (C. comatus), also known as shaggy mane or chicken drumstick mushroom, is a widespread species of the Agaricaceae family. C. comatus has a well–balanced nutritional profile comprising 49.2–76.3% carbohydrates, 11.8–29.5% proteins, 0.6–2.8% dietary fibers, 1.1–5.4% fat, and high levels of EAAs. It has one of the highest PUFAs at 65–66%, mainly composed of linoleic acids, with flavonoids, coumarins, hydroxybenzoic, hydroxycinnamic, and chlorogenic acids [171,172]. C. comatus exhibits anticancer, antidiabetic, antiandrogenic, antimicrobial, antioxidant, anti–inflammatory, acetylcholinesterase inhibitory, and hepatoprotective effects [173,174]. Its major limitation includes allergenic reactions in patients afflicted with dermatitis. They also undergo rapid autolysis and must be consumed at a young stage [175]. Yuan et al. (2022) investigated SPI blends with 15–100% of L. edodes, P. ostreatus, and C. comatus at 35–100% water content via thermos–extrusion. Soy protein isolate–C. comatus (15%) blends with 35% moisture demonstrated texture profiles closely resembling beef. The mushroom meat sausage analogues (MMSA) contained 64 volatile compounds responsible for flavor, identified using gas chromatography-mass spectrometry (GC-MS), thereby indicating the role of fermentation in enhancing aromatic flavor profiles [3]. Ren et al. (2022) analyzed freeze, microwave, infrared, and hot air–drying techniques (FD, MD, IRD, HAD). FD resulted in the highest rehydration ratio and free radical scavenging activity with the lowest color difference in dried C. comatus mushroom samples. MD retained polysaccharides, coumaric acid, and p–hydroxybenzoic acid with higher antioxidant activity. IRD and HAD showed the highest umami concentration, free amino acids, and flavor 5′–nucleotides [176].
Table 5.
Mushroom Proteins for Plant–based Meat Analogues.
| Mushroom Species | Latin Name | Properties | Reference |
|---|---|---|---|
| Button Mushroom | Agaricus bisporus | Enhances juiciness and nutritional profile due to its protein and fiber content, with antioxidant properties. | [177] |
| Oyster Mushroom | Pleurotus ostreatus | Increases protein and fiber content, reduces fat content, softer texture, antioxidant properties. | [178] |
| Shiitake Mushroom | Lentinus edodes | Increases moisture, fiber, nutrition and antioxidant activity, due to methionine and glutamic acid. | [179] |
| Chicken Mushroom | Coprinus comatus | Improves the textural profile and nutritional profile due to its high protein content. | [180] |
2.5. Algae Species
2.5.1. Auxenochlorella protothecoides
Auxenochlorella protothecoides (A. protothecoides), are protein–rich microorganisms of the Chlorellaceae family that can be cultivated in photoautotrophic, mixotrophic, and heterotrophic conditions, with no CO2 emissions and less water consumption [181,182]. They have exceptional antioxidant properties, bioactive compounds, and thickening capacity compared to soy, rice, and whey proteins [183]. Moreover, they are composed of high protein content, chlorophyll and carotenoids, and a high lipid accumulation ability [184]. Sägesser et al. (2024) investigated the structurability of A. protothecoides by HMEC in comparison with soy and pea proteins. A. protothecoides had higher solubility, smaller molecular weight, low hydrophobic sidechains, and pH which affected protein interactions and fibrous network [185]. Caporgno et al. (2020) examined spray–dried A. protothecoides with soy protein concentrate (SPC) to form texturized fibrillar extrudates. 50% of the microalgae biomass was found to considerably influence fiber formation. The increased lipid content led to lubrication, moisture regulation, and improved tenderness while supplementing vitamin B and E [10]. Processing techniques involving denaturation, pH shift, and enzymatic reactions can improve the texture of microalgae–based meat substitutes, making it a promising biomass protein source with ecological advantages and diverse applications in the food industry.
2.5.2. Chlorella vulgaris
Chlorella vulgaris (C. vulgaris), is a freshwater microalgal species belonging to the Chlorellaceae family, composed of 51–58% proteins, 12–17% carbohydrates, 14–22% lipids, vitamins, and minerals [186]. It can be cultivated through phototrophic, mixotrophic, and heterotrophic modes dependent upon external factors such as light intensity and rate of agitation [187,188]. C. vulgaris has numerous food–based applications, encompassing health benefits like detoxification, antioxidation, and immunomodulation [189]. However, its strain type and growth conditions could lead to the risk of contamination due to heavy metal accumulation [190]. These challenges can be tackled by optimizing protected culture systems and advanced processing mechanisms [188]. Gol et al. (2023) combined wet disrupted C. vulgaris (Cv) with PPI by HPH–processing at 10°C to improve gelation. The resultant 14% (w/w) Cv suspensions showed a 10x rise in apparent viscosity, and 9:1 (w/w) PPI–Cv suspensions had 2x storage modulus. Additionally, HMEC PPI–Cv meat analogues retained their hardness, appearance, and anisotropy, with no notable changes post–fractionation and spray drying [191]. Bakhsh et al. (2023) developed PBMAs using Spirulina (SPI), Duck Weed (DW), and Yellow Chlorella (YC). The YC group affected lightness with YC3 having the highest crude fat value, gumminess, and chewiness. Both YC and SPI groups had differing levels of springiness and cohesiveness, but similar DPPH radical scavenging activity [192]. These attributes indicate that C. vulgaris is a novel microalga for plant–based meat products.
2.5.3. Spirulina platensis
Spirulina platensis (S. platensis), is a filamentous cyanobacterium from the Microcoleaceae family recognized for its valuable macro and micronutrients [58,193]. S. platensis can be cultivated in alkaline bodies and has 55–70% proteins, 15–20% carbohydrates, 7% lipids, 30%–35% PUFAs such as gamma–linoleic acid, phenolic acids, polyphenols, and a balanced amino acid profile [194]. It also contains 20% phycobiliproteins such as chlorophyll, phycocyanins, carotenoids, zeaxanthin, and xanthophylls [195]. Furthermore, S. platensis has good protein digestibility and consists of calcium spirulan (Ca–SP), known to inhibit viral diseases such as mumps, measles, cytomegalovirus (CMV), hepatitis C virus (HCV), herpes simplex virus (HSV), human immunodeficiency virus (HIV), and influenza–A (H1N1) [196]. It also has anticancer, antioxidant, anti–inflammatory, neuroprotective, hepatoprotective, hypocholesterolemic, osteogenic, and hormone regulatory effects [197]. Guo et al. (2025) analyzed S. platensis emulsion gels as fat replacers compared to palm oil, oleogel, and soybean oil. S. platensis presented excellent juiciness and heat–tolerance owing to its energy transfer mechanisms and thermally–induced water mobility, with cooking loss and shrinkage equivalent to that of other fats [198]. Afdhaliah et al. (2024) also prepared 30–50% red palm oil (RPO)–Arthrospira platensis protein concentrate (APC)–Carrageenan emulsion gels (EG) as fat substitutes in beef patties. APC effectively stabilized RPO–EG, underscoring its feasibility as a functional ingredient in meat analogues [199].
Table 6.
Algal Proteins for Plant–based Meat Analogues.
| Algae Species | Properties | Reference |
|---|---|---|
| Auxenochlorella protothecoides | High in lipids and carotenoids, potential for enhancing nutritional profile and color. | [200] |
| Chlorella vulgaris | High protein content, vitamins, minerals, and omega–3 fatty acids. Improves nutritional profile, shelf life, and sensory attributes. | [201] |
| Spirulina platensis | High protein content, vitamins, minerals, and rich in EAAs; contributes to improved texture, stability, and color. | [202] |
2.6. Fungi Species
2.6.1. Aspergillus oryzae (Koji Mold)
Aspergillus oryzae (A. oryzae), also known as Koji mold, is an aerobic filamentous fungi belonging to the Aspergillaceae family with both agronomic and medicinal significance, widely used in the production of food processing enzymes [203,204]. Optimal conditions for cultivating A. oryzae include temperature ranges between 32–36°C and pH levels ranging from 2.0–8.0 [205]. While A. oryzae has been commonly employed in fermented products such as soy sauce, soybean paste, rice koji beverage, and black rice vinegar, it has also expanded into PBMAs [206]. Fermented fungus is prepared by introducing A. oryzae to rice, soybean, oat flour, or wheatgrass substrates to produce enzymes responsible for breaking down proteins, fats, and starches into amino acids, fatty acids, and sugars [207]. These substrates also contain nutrients desirable for fungal growth in submerged fermentation (SmF) systems. Yeast and lactic acid bacteria facilitate fermentation to obtain the end–products [208]. Gamarra–Castillo et al. (2022) developed burger patties using A. oryzae, flours, binders, and colorants under moderated conditions of the growth medium. The study revealed that biomass production was maximized at 6 days of incubation with 90 g/L maltodextrin. Additives such as quinoa flour, carboxymethylcellulose, and beet extract resulted in formulations with texture and appearance that closely resembled the control meat [4]. Thus, fungal proteins can be further explored to develop nutritious food in alignment with consumer preferences [209].
2.6.2. Neurospora intermedia
Neurospora intermedia (N. intermedia), is a natural hyphal fungus from the Sordariaceae family with an extensive history in food applications. N. intermedia is commonly utilized in Oncom, Cassava–based beverages, and milk cheese [210]. It can be readily cultivated by submerged and solid–state fermentation (SSF) using straw and molasses carbon substrates to effectively break down complex compounds into protein biomasses [211]. N. intermedia can also be grown on isolated plant byproducts such as brewer’s spent grain, oilseed cakes, pomace, and plant–milk waste [212]. Its processing parameters are largely dependent upon temperature, pH, agitation rate, duration, and inoculum. N. intermedia is normally grown for 3–5 days at 30℃, with pH between 5.5–6.5, inoculum volumes of 1–5%, and agitation rates of 100–200 rpm [213]. N. intermedia has a rapid growth cycle with a high nutritional value and is considered as a safe fungal species as it does not encode mycotoxins when compared to Aspergillus and Penicillium variants [210]. Wang et al. (2024) investigated the role of N. intermedia at 3–7 pH levels and 0–6% soluble starch (SS) to create mycoprotein gel meat (CMGM). At pH 7, CMGM showed good elasticity, recovery, and a scant internal structure; whereas decreased pH resulted in increased water retention and gelling post–steaming. At 6% SS, CMGM exhibited improved hardness and WHC, with reduced porosity and elongation. These findings present the crosslinking ability of mycelial fibers in the gel matrix to produce mycoprotein–based meat substitutes [214].
2.6.3. Fusarium venenatum
Fusarium venenatum (F. venenatum), is a single–celled fungal biomass of the Nectriaceae family composed of a natural fibrous matrix with cellulose, chitin, glucans, 15–45% proteins, dietary fibers, and low fat [215]. Mycoprotein from F. venenatum is produced by continuous fermentation on a glucose substrate at high metabolic rates in an air–lift or solid–state bioreactors [216]. Nutritional supplements typically include vitamins, biotin and choline, and other minerals such as calcium, potassium, magnesium, sulfur, phosphorus, iron, manganese, molybdenum, and copper. In addition, carbon and nitrogen sources, are introduced into the culture medium to fulfil amino acid and sulfate metabolism, lipid and phospholipid biosynthesis, and gluconeogenesis [217,218]. Although F. venenatum has low digestibility, it is well–suited for human consumption as it has low allergenic potential, regulates cholesterol, induces muscle protein anabolism, and controls glucose–insulin levels [219]. Furthermore, F. venenatum contains 1–8% nucleic acid levels which is lower than that of bacteria and yeast, thereby reducing undesirable flavors and providing a meat–like texture to plant–based meat products [220]. Li et al. (2025) investigated the nutritional profile of F. venenatum mycoprotein–based Harbin red sausages, which revealed that increased mycoprotein content and volatile organic compounds subsequently create a rich flavor, promoting consumer acceptance [221].
Table 7.
Fungal Proteins for Plant–based Meat Analogues.
| Fungi Species | Properties | Reference |
|---|---|---|
| Aspergillus oryzae | Imparts a fibrous structure with high protein content and uses various substrates for cultivation. | [222] |
| Neurospora intermedia | Enhances chewability, gelation, and palatability when combined with soluble starch at optimal pH levels. | [223] |
| Fusarium venenatum | Mimics meat–like texture and enhance nutritional profile; widely used in commercial mycoprotein products like Quorn. | [224] |
3. Nutritional Amino Acid Profiling
The nutritional quality of plant-based meat analogues is highly dependent upon their protein content including essential amino acids (EAAs) and non-essential amino acids (NEAAs), lipids, vitamins, minerals, carbohydrates, and dietary fibers, which provide a holistic nutraceutical profile crucial for improving metabolic health. The nutritional profiles of predominant plant-based proteins such as soy, pea, mung bean, faba bean, lupin bean, peanut, and oat proteins are represented in Table 8. Among these plant-based sources, soy protein provides an optimal EAA profile high in lysine, methionine, and iron content, making it suitable for replicating the flavor of conventional meat. Similarly, pea protein contains regulatory branched-chain amino acids (BCAA) such as leucine, isoleucine, and valine, which help facilitate muscle growth and repair. Furthermore, lupin bean protein offers a high methionine content and dietary fibers. High levels of lysine and methionine are important factors in effectively mimicking the sensory properties of meat, as they are responsible for protein synthesis and imparting umami flavors. Among the mycoprotein sources indicated in Table 9, Agaricus bisporus and Pleurotus ostreatus possess balanced EAA profiles with suitable vitamins and minerals. Fungal proteins such as Aspergillus oryzae and Fusarium venenatum also contain high quantities of branched-chain amino acids and histidine, which contribute to a meat-like texture. While Auxenochlorella protothecoides is limited in its lysine and methionine content, it serves as a functional algal protein source primarily due to its rich presence of omega-3 and omega-6 fatty acids.
3.1. Nutritional Profile of Plant Protein Sources
Table 8.
Nutritional Value of Major Plant Proteins.
| Total Protein Content (g/100g) |
Chicken Breast [225] |
Soy [226] |
Pea [227] |
Mung bean [228] |
Faba bean [229] |
Lupin bean [230] |
Peanut [231] |
Oat [232] |
| Isoleucine (Ile) | 4.64 | 2.0 – 4.2 | 2.3 – 4.5 | 3.9 | 3.9 | 3.5 | 0.8 – 1.5 | 3.8 – 4.1 |
| Histidine (His) | 3.04 | 1.0 – 2.3 | 1.6 – 2.5 | 2.8 | 2.4 | 2.7 | 0.6 – 1.1 | 2.1 – 2.4 |
| Leucine (Leu) | 8.27 | 3.0 – 6.7 | 5.7 – 6.4 | 7.4 | 7.4 | 6.8 | 1.6 – 2.9 | 6.9 – 7.6 |
| Methionine (Met) | 2.84 | 0.5 – 1.1 | 0.3 – 1.1 | 1.3 | 0.8 | 2.8 | 0.3 – 0.5 | 2.2 – 3.3 |
| Lysine (Lys) | 7.55 | 2.7 – 5.3 | 4.7 – 5.7 | 6.2 | 7.0 | 4.5 | 0.9 – 1.5 | 3.5 – 4.1 |
| Phenylalanine (Phe) | 4.14 | 2.1 – 4.5 | 3.7 – 5.5 | 9.0 | 4.1 | 4.9 | 1.3 – 2.2 | 5.0 – 5.5 |
| Tryptophan (Trp) | 1.04 | 0.5 – 1.1 | 0.7 – 1 | 0.6 | 0.8 | 0.6 | 0.2 – 0.4 | 0.8 – 0.9 |
| Threonine (Thr) | 4.51 | 1.7 – 3.1 | 2.5 – 3.9 | 2.0 | 3.4 | 2.9 | 0.8 – 1.1 | 3.1 – 3.4 |
| Valine (Val) | 5.07 | 2.0 –4.0 | 2.7 – 5.0 | 4.6 | 4.3 | 3.2 | 1.0 – 1.8 | 5.2 – 5.8 |
| Alanine (Ala) | 5.33 | 1.9 – 3.5 | 3.2 – 4.3 | 3.6 | 4.1 | 2.9 | 0.9 – 1.7 | 4.4 – 4.9 |
| Arginine (Arg) | 6.44 | 3.1 – 6.6 | 6.6 | 6.4 | 9.4 | 10.0 | 3.0 – 5.0 | 6.1 – 7.1 |
| Aspartic acid (Asp)/Asparagine (Asn) | 9.06 | 10.2 – 12.0 | 8.9 – 11.5 | 8.5 | 10.7 | 9.7 | 3.0 – 4.9 | 7.6 – 8.7 |
| Cysteine (Cys) | 1.01 | 0.6 – 1.0 | 0.2 – 1.0 | 1.3 | 1.3 | 2.8 | 0.3 – 0.6 | 1.9 – 2.5 |
| Glutamic acid/Glutamine (Glu) | 13.52 | 7.8 –17.5 | 12.9 –13.2 | 12.5 | 16.5 | 22 | 5.2 – 8.3 | 20.9 – 27.3 |
| Glycine (Gly) | 4.0 | 1.8 – 3.6 | 2.8 – 4.1 | 3.2 | 4.3 | 3.7 | 1.5 | 4.6 – 5.3 |
| Proline (Pro) | 3.4 | 2.3 – 4.9 | 3.1 – 4.5 | 3.0 | 3.9 | 3.2 | 1.1 – 1.9 | 5.5 – 6.9 |
| Serine (Ser) | 3.99 | 2.3 – 4.5 | 3.6 – 5.3 | 3.8 | 4.6 | 4.3 | 1.2 – 1.7 | 3.8 – 5.6 |
| Tyrosine (Tyr) | 3.54 | 1.5 – 3.2 | 2.6 – 3.8 | 9.0 | 2.7 | 4.9 | 1.0 – 1.6 | 2.7 – 3.5 |
| Total Fat Content (g/100g) | Chicken Breast | Soy |
Pea |
Mung bean | Faba bean | Lupin bean |
Peanut |
Oat |
| Saturated fat (g) | 1.0 – 1.2 | 2.88 | 0.071 | 0.348 | 0.066 | 1.16 | 6.28 | 1.11 |
| Monounsaturated fat (g) | 0.8 – 1.2 | 4.4 | 0.035 | 0.161 | 0.079 | 3.94 | 24.4 | 1.98 |
| Polyunsaturated fat (g) | 0.8 | 11.3 | 0.187 | 0.384 | 0.164 | 2.44 | 15.6 | 2.3 |
| Essential fatty acids–Omega–3 (mg) | 32 – 107 | 1330 | 35 | 27 | 12 | 446 | 3.0 | 100 |
| EFA– Omega–6 (mg) | 559 | 9920 | 152 | 357 | 152 | 2000 | 15600 | 2200 |
| Vitamin Content | Chicken Breast | Soy |
Pea |
Mung bean | Faba bean | Lupin bean |
Peanut |
Oat |
| Vitamin A (RAE, UI) (μg, UI) | 6, 21 | 1, 22 | 38, 765 | 6, 114 | 1, 15 | – | – | – |
| Vitamin B1 (Thiamin) (mg) | 0.07 | 0.874 | 0.266 | 0.233 | 0.097 | 0.64 | 0.64 | 0.46 |
| Vitamin B2 (Riboflavin) (mg) | 0.114 | 0.87 | 0.132 | 0.621 | 0.089 | 0.22 | 0.135 | 0.155 |
| Vitamin B3 (Niacin) (mg) | 9.45 –13.7 | 1.62 | 2.09 | 2.25 | 0.711 | 2.19 | 12.1 | 1.125 |
| Vitamin B5 (Pantothenic acid) (mg) | 0.965 | 0.793 | 0.104 | 1.91 | 0.157 | 0.75 | 1.77 | – |
| Vitamin B6 (mg) | 0.6 –1.0 | 0.377 | 0.169 | 0.382 | 0.072 | 0.357 | 0.348 | 0.1 |
| Vitamin B9 (Folate) (μg) | 4.0 | 375 | 65 | 625 | 104 | 355 | 246 | 56 |
| Vitamin C (Total Ascorbic acid) (mg) | – | 6.0 | 40 | 4.8 | 0.3 | 4.8 | – | – |
| Vitamin D | 0.1 –2.5 | – | – | – | – | – | – | – |
| Vitamin E (alpha–toco–pherol) (mg) | 0.27 | 0.85 | 0.13 | 0.51 | 0.02 | – | 8.33 | 0.42 |
| Vitamin K (Phylloquinone) (μg) | 0.3 | 47 | 24.8 | 9.0 | 2.9 | – | – | 2.0 |
| Mineral Content | Chicken Breast | Soy |
Pea |
Mung bean | Faba bean | Lupin bean |
Peanut |
Oat |
| Calcium (Ca) (mg) | 6.0 –18.0 | 277 | 25 | 132 | 36 | 176 | 92 | 52 |
| Iron (Fe) (mg) | 1.04 –1.07 | 15.7 | 1.47 | 6.74 | 1.5 | 4.36 | 4.58 | 4.25 |
| Magnesium (Mg) (mg) | 29 | 280 | 33 | 189 | 43 | 198 | 168 | 138 |
| Phosphorous (P) (mg) | 228 | 704 | 108 | 367 | 125 | 440 | 376 | 410 |
| Potassium (K) (mg) | 343 –460 | 1880 | 244 | 1250 | 268 | 1010 | 705 | 362 |
| Sodium (Na) (mg) | 74 | 2.0 | 5.0 | 15 | 5.0 | 15 | 18 | 6.0 |
| Zinc (Zn) (mg) | 1.0 –1.6 | 4.89 | 1.24 | 2.68 | 1.01 | 4.75 | 3.27 | 3.64 |
| Copper (Cu) (mg) | 0.049 | 1.66 | 0.176 | 0.941 | 0.259 | 1.02 | 1.14 | 0.391 |
| Manganese (Mn) (mg) | 0.017 | 2.52 | 0.41 | 1.04 | 0.421 | 2.38 | 1.93 | – |
| Selenium (Se) (μg) | 28.4 | 17.8 | 1.8 | 8.2 | 2.6 | 8.2 | 7.3 | 28.9 |
| Nutritional Content | Chicken Breast | Soy |
Pea |
Mung bean | Faba bean | Lupin bean |
Peanut |
Oat |
| Ash (g/100g) | 1.06 | 4.87 | 0.87 | 3.32 | 0.81 | 3.28 | 2.33 | – |
| Carbohydrates (g/100g) | – | 30.2 | 14.4 | 62.6 | 19.6 | 40.4 | 16.1 | 67.7 |
| Total Dietary Fibers (g/100g) | – | 9.3 | 5.7 | 16.3 | 5.4 | 18.9 | 8.5 | 10.1 |
| Total Sugars (Glucose, Fructose, Lactose, Maltose, Galactose) (g/100g) | – | 7.33 | 5.67 | 6.6 | 1.82 | – | 4.72 | 0.99 |
| Calories (kcal) | 165 | 446 | 81 | 347 | 460 | 371 | 567 | 379 |
3.2. Nutritional Profile of Mushroom/Algae/Fungi Species
Table 9.
Nutritional Value of Major Mushroom/Algae/Fungi Species.
| Total Protein Content (g/100g) |
Chicken Breast [225] |
A. bisporus [233] |
P. ostreatus [234] |
L. edodes [168] |
A. oryzae . [209] |
A. protothecoides [235] |
P. limosum [236] |
F. venenatum [237] |
| Isoleucine (Ile) | 4.64 | 1.37 | 1.02 | 0.62 | 13.2 | 0.31 | NS | 1.51 |
| Histidine (His) | 3.04 | 0.17 | 0.27 | 0.84 | 0.75 | 0.09 – 0.96 | NS | 7.22 |
| Leucine (Leu) | 8.27 | 1.20 | 1.23 | 1.29 | 2.5 | NS | NS | 1.90 |
| Methionine (Met) | 2.84 | 0.22 | 0.26 | 0.42 | 0.59 | Low | NS | 4.21 |
| Lysine (Lys) | 7.55 | 1.09 | 1.18 | 1.91 | 1.83 | 0.184 – 0.224 | NS | 5.81 |
| Phenylalanine (Phe) | 4.14 | 0.19 | 0.25 | 0.83 | 4.7 | Low | NS | 3.01 |
| Tryptophan (Trp) | 1.04 | 0.3 | 0.4 | 0.28 | 0.15 | 0.36 | NS | NS |
| Threonine (Thr) | 4.51 | 0.36 | 0.38 | 1.01 | 0.3 | 0.23 | NS | 3.31 |
| Valine (Val) | 5.07 | 0.68 | 0.42 | 1.05 | 1.27 | NS | NS | 6.05 |
| Alanine (Ala) | 5.33 | 0.77 | 0.62 | 1.17 | 2.52 | NS | NS | 2.41 |
| Arginine (Arg) | 6.44 | 0.74 | 0.23 | 2.45 | 1.72 | NS | NS | 7.12 |
| Aspartic acid (Asp)/Asparagine (Asn) | 9.06 | 1.92 | 0.87 | 1.73 | 1.69 | NS | NS | NS |
| Cysteine (Cys) | 1.01 | 0.14 | 0.18 | 0.08 | 0.5 –1.0 | NS | NS | 2.11 |
| Glutamic acid/Glutamine (Glu) | 13.52 | 1.06 | 0.98 | 4.93 | 5.7 | 0.56 – 6.88 | NS | NS |
| Glycine (Gly) | 4.0 | 0.6 | 0.47 | 0.89 | 0.57 | NS | NS | 3.50 |
| Proline (Pro) | 3.4 | 0.69 | 0.68 | 0.82 | 2.5 | NS | NS | NS |
| Serine (Ser) | 3.99 | 0.62 | 0.26 | 1.04 | 6.19 | NS | NS | NS |
| Tyrosine (Tyr) | 3.54 | 0.28 | 0.65 | 0.54 | 0.72 | Low | NS | NS |
| Total Fat Content | Chicken Breast | A. bisporus | P. ostreatus | L. edodes | A. oryzae | A. protothecoides | P. limosum | F. venenatum |
| Total Fat Content (g) | 3.5 – 3.6 | 0.34 – 0.41 | 0.41 | 0.49 – 3.0 | 3 – 5 | 21 | 2.9 | 2.9 |
| Saturated fat (g) | 1.0 – 1.2 | 0.09 | 0.06 | ~0.23 | ~1.2 | NS | 0.7 | 0.7 |
| Monounsaturated fat (g) | 0.8 – 1.2 | 0.21 – 0.23 | 0.03 | ~0.3 | NS | NS | 0.5 | 0.5 |
| Polyunsaturated fat (g) | 0.8 | 0.07 – 0.1 | 0.12 | ~0.2– 0.24 | NS | High | 1.4 | 1.8 |
| Essential fatty acids–Omega–3 (mg) | 0.032– 0.107 | 0.1 | Low | Low | Low | High | NS | 6.9 |
| EFA– Omega–6 (mg) | 0.55 – 3.5 | 53 – 68 | 3.1 – 5.4 | 1.04 | 0.22 | 27.6 | 3.0 | 3.0 |
| Vitamin Content | Chicken Breast | A. bisporus | P. ostreatus | L. edodes | A. oryzae | A. protothecoides | P. limosum | F. venenatum |
| Vitamin A (RAE, UI) (mg, UI) | 0.006, 21 | 0 | 0.23 – 21, 2.93 | 0.01 | NS | NS | NS | 0 |
| Vitamin B1 (Thiamin) (mg) | 0.07 | 1.05 | NS | 0.05 | 0.2 | 0.1 | NS | 0.07 |
| Vitamin B2 (Riboflavin) (mg) | 0.114 | 0.42 | NS | 0.15 | 0.3 | 0.5 | NS | 0.114 |
| Vitamin B3 (Niacin) (mg) | 9.45 – 13.7 | 4.55 | NS | 0.99 | 3.6 | 2.0 | NS | ~13.7 |
| Vitamin B5 (Pantothenic acid) (mg) | 0.965 | 1.75 | NS | 0.5 | 1.5 | 0.5 | NS | 1.58 |
| Vitamin B6 (mg) | 0.6 – 1.0 | 0.082 | NS | 0.1 | 0.1 | 0.2 | NS | 0.921 |
| Vitamin B9 (Folate) (μg) | 4.0 | NS | NS | 21.51 | 500 | 100 | NS | NS |
| Vitamin C (Total Ascorbic acid) (mg) | – | NS | 16.46 | 2.1 | NS | 1.0 | NS | 0 |
| Vitamin D (μg) | 0.1 – 2.5 | 0.2 | 29 | NS | NS | NS | NS | 0.025 |
| Vitamin E (alpha–toco–pherol) (mg) | 0.27 | NS | 21.50 | NS | NS | 0.5 | NS | NS |
| Vitamin K (Phylloquinone) (μg) | 0.3 | NS | NS | NS | NS | NS | NS | NS |
| Mineral Content | Chicken Breast | A. bisporus | P. ostreatus | L. edodes | A. oryzae | A. protothecoides | P. limosum | F. venenatum |
| Calcium (Ca) (mg) | 6.0 –18 | 0.047 | 342 – 410 | 18 | 2.0 | 20 | NS | 5 |
| Iron (Fe) (mg) | 1.04 – 1.07 | 0.013 | 17 – 21 | 0.9 | 10 | 5.0 | NS | 0.45 |
| Magnesium (Mg) (mg) | 29 | 115 | 7.0 | 40.7 | NS | NS | NS | NS |
| Phosphorous (P) (mg) | 228 | 860 | 695 – 1060 | 778 | NS | NS | NS | NS |
| Potassium (K) (mg) | 343 – 460 | 4015 | 2080 – 2280 | 356 | 230 | 300 | NS | 255 |
| Sodium (Na) (mg) | 74 | 3.0 | 193 | NS | NS | NS | NS | NS |
| Zinc (Zn) (mg) | 1.0 – 1.6 | 0.013 | 12.96 | 1.0 | 1.5 | 1.0 | NS | 0.8 |
| Copper (Cu) (mg) | 0.049 | 52 – 350 | 91 – 116 | 14.8 | NS | NS | NS | NS |
| Manganese (Mn) (mg) | 0.017 | 4.8 | 16 – 23 | 2.0 | NS | NS | NS | NS |
| Selenium (Se) (μg) | 28.4 | NS | NS | 46.1 | NS | NS | NS | NS |
| Nutritional Content | Chicken Breast | A. bisporus | P. ostreatus | L. edodes | A. oryzae | A. protothecoides | P. limosum | F. venenatum |
| Ash (g/100g) | 1.06 | 1.35 | 8.22 | 6.0 | 4.0 | 4.0 | 4.0 | 2.0 |
| Carbohydrates (g/100g) | – | 3.3 | 43.42 | 64 | 50 | 20 | 50 | 12 |
| Total Dietary Fibers (g/100g) | – | 11.01 | 21 – 47 | 33.6 | 14 | NS | 30 | 6.0 |
| Total Sugars (Glucose, Fructose, Lactose, Maltose, Galactose) (g/100g) | – | 14.08 | 1.11 | 4.40 | ~2.0 | 3.5 | NS | 1.0 |
| Calories (kcal) | 165 | 22 | 33 | 34 | 300 | 300 | 300 | 93 |
[Note: NS → Not Specified].
4. Formulatory Composition
4.1. Binding Agents
4.1.1. Carboxymethyl Cellulose (CMC)
Carboxymethylcellulose (CMC), a cellulose derivative, is widely used in plant–based meat analogues since it enhances texture, moisture retention, and structural stability. CMC acts as a hydrocolloid, possessing remarkable water–holding capacity and viscosity, forming a 3D network structure that improves rheological properties and overall mouthfeel of meat analogues. Earlier studies suggest that CMC may help lower food products’ glycemic index. Through the formation of a matrix around starch granules, CMC retards enzymatic hydrolysis, digestion, and nutrient absorption [238]. Additionally, CMC has been shown to be successful in lowering the absorption of fat in fried foods. 1% CMC–coated French fries lowered fat absorption to 65.1%, compared to 14.1% fat in uncoated samples [239]. CMC is compatible with other edible gums–kinds like guar gum or xanthan gum. It is essential for reproducing the fibrous meandering structure in meat analogues since it reduces interfacial tension between the water and oil phases, which is perfect for emulsification [240]. In varying pH conditions and processing conditions, CMC’s robustness in industrial applications. While its cost is higher than alternatives like starch and pectin, CMC remains a preferred choice due to its superior ability to deliver desired texture, binding, and moisture retention in plant–based meat analogues.
4.1.2. Methylcellulose (MC)
Methylcellulose is an important binder and texturizing agent. It is frequently utilized because it may decrease oil leakage and purge loss, increase emulsion stability, and improve textural qualities. It’s particularly effective in water and fat binding, helping to retain moisture and fat in the products. MC is the most commonly used ingredient in PBMA burgers because of its binding capacity and unique reversible thermal gelation [241]. It helps to bind texturized vegetable proteins (TVP) and other ingredients together in comminated and emulsion–type PBMA products, such as burgers and sausages. The binding results in a consistent gel network, which improves the product’s mouthfeel and texture. Despite its effectiveness, methylcellulose has an “E number” designation, a European Union (EU) labelling system used to indicate the presence of food additives which may not appeal to consumers who perceive additives as inorganic constituents [242]. However, it remains a key ingredient in many commercial plant–based meat products due to its functionality. Alternatives such as alginate, hydroxypropyl methylcellulose, and other hydrocolloids are considered but they cannot match methylcellulose’s unique properties.
4.1.3. Xanthan Gum (X)
Plant–based meat substitutes majorly utilize xanthan gum, an extracellular polysaccharide fermented from Xanthomonas species, because of its gelling, thickening, stabilizing, and emulsifying qualities [239]. It enhances texture and fibrous structure by encouraging the development of cohesive and organized networks in protein matrices, particularly in formulations that contain wheat gluten and pea protein isolate [243]. Additionally, xanthan gum interacts with pectin and low acyl gellan gum, to improve the mechanical qualities and mimic a meat–like texture. Since xanthan gum has a higher yield strength than protein–based binding agents, it may be used to create intermediate fibres and improve structural compactness in soy protein isolate (SPI) blends [244]. Xanthan gum also serves to enhance both 3D printing performance along with gel stability in plant–based formulations, rendering syneresis impossible, delaying staling, and capable of maintaining shape at freeze–thaw cycles [245]. Although the utilization of hydrocolloids such as xanthan gum has raised controversy upon several occasions due to their nutritional effects and safety, no significant evidence is available to date to support complaints or concerns about health hazards (EFSA Panel on Food Additives and Nutrient Sources added to Food, 2018).
4.1.4. Carrageenan (CA)
Red seaweeds like Chondrus crispus and Eucheuma sp. are the source of Carrageenan, a polysaccharide used in the food industry for its thickening, stabilizing, and emulsifying qualities [246]. The range of applications includes the stabilization of emulsions in dairy and meat products, as well as an improvement in the texture and lipid oxidation reduction of formulations such as sausages, ham, and PBMAs [247]. Additionally, carrageenan is essential for enhancing emulsion stability and water–holding capacity, both of which are critical for yielding the appropriate texture in meat substitutes. The textural properties in mushroom–based sausage analogues that include carrageenan improve when compared with soy protein concentrate and casein, including reduced purge loss and gumminess. Earlier studies showed that its gelling and thickening ability improves product stability and consistency as well as cooking loss reduction [248]. Carrageenan is also utilized to make edible films and coatings for fresh meals. Beyond food, it has other health benefits like regulating cholesterol levels and aiding in wound healing [249]. Kappa–carrageenan (k–carrageenan) is one of the most widely used forms and is particularly known for its strong gelling properties, which are commonly used in meat analogues for structural integrity.
4.1.5. Guar Gum (GuG)
A multipurpose polysaccharide derived from Cyamopsis tetragonolobus seeds, Guar gum is well-known for its ability to bind, thicken, and emulsify [250]. Its high water-holding capacity (WHC) and viscosity–modifying characteristics make it a significant component in plant–based meat preparations. It increases water retention and enhance 3D printing performance in soy protein isolate gels while inhibiting syneresis during freeze–thaw cycles [251]. In food–based applications, guar gum reduces the absorption of fats, for instance, French fries coated with 1% guar gum that saw a decrease of 60.2% fat content in relation to the control samples [252]. Methods to retain the moisture content and textural properties in meat analogues have been researched extensively to ensure improved juiciness and mouthfeel. Studies have revealed that addition of hydrocolloids such as guar gum in plant-based meat formulations enhances the water holding capacity, subsequently simulating the fibrous structure of real meat [14]. Despite occasional debates over the nutritional impact of edible gums, including guar gum, no substantial evidence has been found to suggest any health risks, further solidifying its role as a safe and effective binding agent (EFSA Panel on Food Additives and Nutrient Sources added to Food, 2018). Furthermore, guar gum can effectively interact with other hydrocolloids and proteins, allowing for the creation of a harmonious texture that is crucial for product acceptance. This property is essential for companies seeking to appeal to a broad range of consumers.
4.1.6. Gellan Gum (GeG)
Gellan gum, a microbial hydrocolloid derived from Sphingomonas sp., is widely renowned for its remarkable gelling properties [253]. Plant-based meat substitutes and various products, including jams, jellies, crackers, chicken nuggets, yogurt, and frozen desserts, can benefit from their ability to produce strong, stable gels that withstand heat and pH changes [240]. Consequently, it elevates the cohesion and mouthfeel of the finished product. When appropriately balanced salt concentrations are employed, the network of gellan in a protein matrix forms a cohesive structure [244]. Gellan gum has two forms–high acyl and low acyl, each exhibiting different properties and performance in plant-based meat formulations. High Acyl Gellan Gum form is a more flexible gel because it is softer and more elastic than acyl groups. Therefore, it is useful for applications in which a tender texture is desirable, such as frozen desserts and yogurts. On the other hand, low acyl gellan gum forms firm, brittle gels since they lack acyl groups. Its structural rigidity and fibrousness are desirable in meat analogs and textured products. When combined, these forms can balance textural properties to mimic the fibrous structure of traditional meat. Salt concentrations further enhance plant- based meat’s functionality, mechanical, and textural characteristics [254].
4.1.7. Other Hydrocolloid–Based Binders in PBMAs
Pectin is a polysaccharide predominantly found in the cell walls of fruits and is widely recognized for its gelling properties. Although primarily used in jams and jellies, pectin’s ability to form gels makes it a candidate for PBMAs. Pectin has proven to enhance the structural integrity of meat analogues, contributing to a more cohesive protein matrix when combined with other hydrocolloids. Low methylated pectin with calcium chloride (CaCl2) in blends containing PPI has been found to facilitate gelation, resulting in thin fibril formations within the protein matrix, thereby improving the overall texture [244]. Pectin’s unique egg–box model mechanism allows it to stabilize gels through electrostatic interactions with cations, which is crucial for achieving the desired consistency in meat alternatives [254]. Furthermore, pectin can lower the glycemic index, making it an advantageous ingredient in PBMA formulations [238].
Brown seaweed is the natural alginate source, a polymer sold as sodium alginate [255,256]. Because of its free hydroxyl and carboxyl groups, they interact with water effectively and can be used as a gelling agent, thickener, stabilizer, and emulsifier. It serves as a source of dietary fiber that may help reduce cholesterol and blood glucose levels. Notable sources include Laminaria sp. and Sargassum sp. Alginate improves product stability throughout freeze-thaw cycles and provides exceptional adhesion. Additionally, it improves the texture by binding vegetable proteins and reducing oil absorption [257]. Overall, while alginate may be less commonly utilized than other binders in PBMAs, its unique properties present promising opportunities for innovation.
Agar, a polysaccharide derived from red seaweeds such as Gelidium spp. and Gracilaria spp., is recognized for its stabilizing, thickening, and gelling properties [258]. In the context of PBMAs, agar can enhance texture and provide desirable elasticity, making it an effective binder that contributes to meat alternatives’ overall mouthfeel and structure. Its capacity to gel enables better cohesiveness between plant proteins. Additionally, agar’s high melting point ensures that the structural integrity of PBMAs is maintained during cooking. Given its functionality and cost-effectiveness due to the abundant availability of seaweeds, agar presents a viable way to improve the quality of plant-based meat substitutes.
Gum Arabic, sometimes called gum acacia, is an exudate from Acacia trees characterized by branched polysaccharides, including β–D–galactopyranosyl units [259]. In PBMAs, gum Arabic enhances emulsification and foaming properties, particularly in sausage–type meat analogues, where it helps maintain stable emulsions. It improves binding efficiency, maintains suspension, and contributes to bulk volume and cloudiness in formulations. Its functionality at alkaline pH further supports its use in various meat alternatives. Gum Arabic enhances texture and improves mouthfeel, making it a valuable ingredient in plant-based meat products.
Konjac mannan gum, derived from the tubers of Amorphophallus konjac, is a polysaccharide characterized by high molecular weight and water solubility [260]. It is an excellent gelling and emulsifying agent, making it particularly useful as a binder. In plant-based formulations, konjac mannan gum helps reduce moisture loss during cooking and frying, improving meat alternatives’ overall texture and sensory attributes. For instance, vegan meat sausages formulated with konjac mannan gum have been shown to closely resemble traditional meat sausages in terms of hardness, slicing ability, and water–holding capacity [261]. Additionally, its ability to enhance non-digestible fiber content makes konjac mannan gum suitable for low–calorie food applications.
Locust bean gum, derived from the seeds of the Ceratonia siliqua tree, is a high–molecular–weight polysaccharide recognized as a natural food additive [280]. It can prevent aggregation, stabilize emulsions, and improve texture by reducing syneresis and stabilizing freeze-thawing in starch–based formulations [262]. Its composition—primarily D–mannopyranose units with side chains of D–galactopyranose—allows it to function effectively as a binder while supporting gut health and reducing fat absorption [282]. Overall, locust bean gum is a valuable ingredient for enhancing the stability and sensory attributes of PBMAs.
Figure 2.
(A) Plant-based meat analogues with different concentrations of hydrocolloids (Control: plant-based patty without polysaccharide; Car–1: 1% κ–carrageenan, Car–2: 2% κ–carrageenan; LBG–1: 1% locust bean gum, LBG–2: 2% locust bean gum; AG–1: 1% Arabic gum, AG–2: 2% Arabic gum; GeG–1: 1% gellan gum, GeG–2: 2% gellan gum; GuG–1: 1% guar gum, GuG–2: 2% guar gum; XG–1: 1% xanthan gum, XG–2: 2% xanthan gum; BP: Beef patty); (B) Sensory properties of plant-based meat analogues; (C) Water–holding capacity (WHC) and cooking yield of plant-based meat analogues. Reproduced with permission [263]. Copyright Multidisciplinary Digital Publishing Institute (MDPI), 2023.
Figure 2.
(A) Plant-based meat analogues with different concentrations of hydrocolloids (Control: plant-based patty without polysaccharide; Car–1: 1% κ–carrageenan, Car–2: 2% κ–carrageenan; LBG–1: 1% locust bean gum, LBG–2: 2% locust bean gum; AG–1: 1% Arabic gum, AG–2: 2% Arabic gum; GeG–1: 1% gellan gum, GeG–2: 2% gellan gum; GuG–1: 1% guar gum, GuG–2: 2% guar gum; XG–1: 1% xanthan gum, XG–2: 2% xanthan gum; BP: Beef patty); (B) Sensory properties of plant-based meat analogues; (C) Water–holding capacity (WHC) and cooking yield of plant-based meat analogues. Reproduced with permission [263]. Copyright Multidisciplinary Digital Publishing Institute (MDPI), 2023.
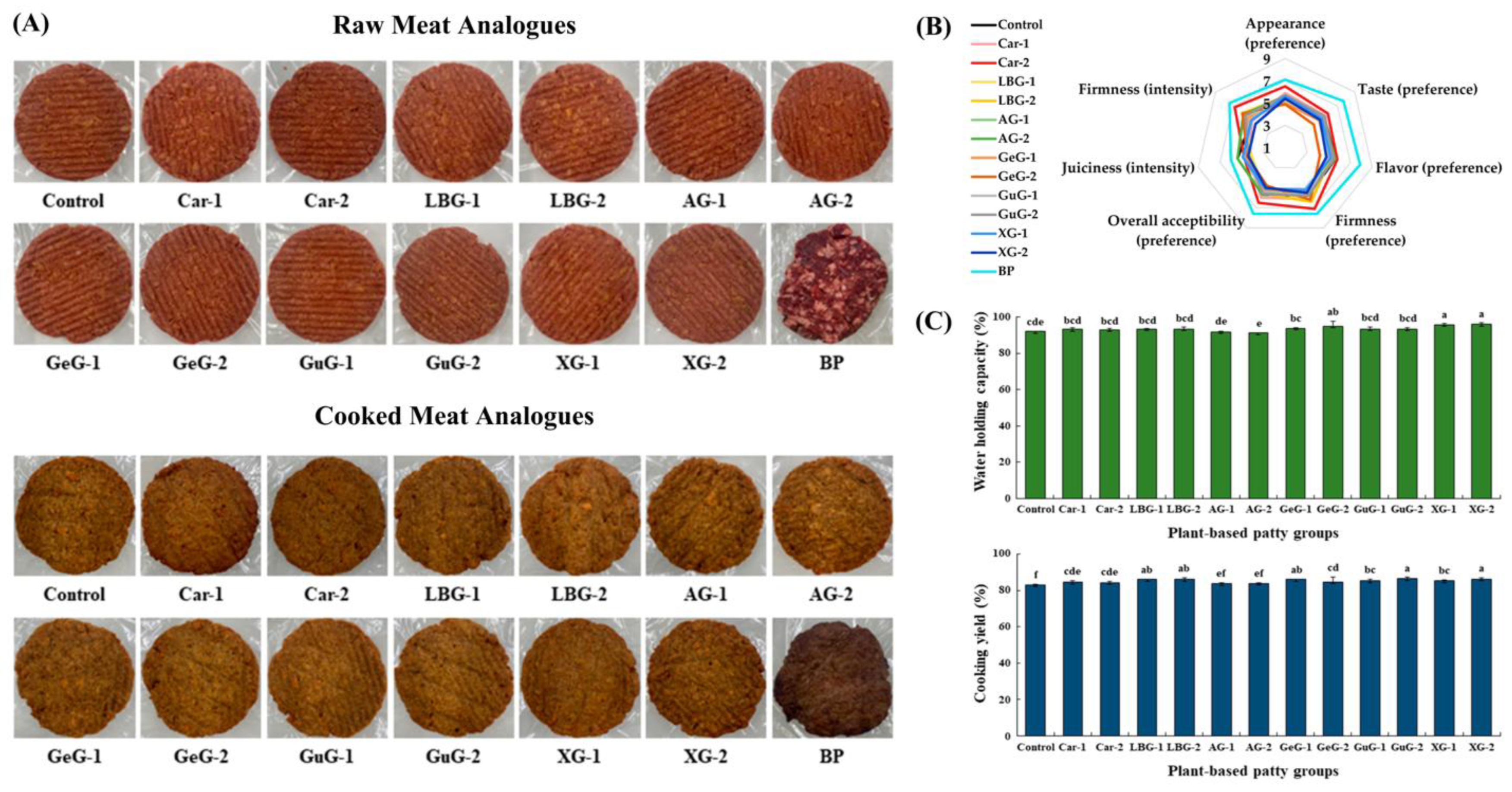
Table 10.
Types of Hydrocolloids in Meat Analogues.
| Hydrocolloids (Food additive code) | Latin Name | Properties | Reference |
|---|---|---|---|
| Carrageenan (E407) |
Chondrus crispus | Acts as a binder, stabilizer, and moisture retainer.
|
[264] |
| Xanthan gum (E415) |
Xanthomonas campestris | Exhibits pseudoplasticity. Maintains stability over a wide temperature (10–80°C) and pH (5–10) range. | [265] |
| Guar gum (E412) |
Cyamopsis tetragonoloba | Functions as a thickener, texture enhancer and fat substitute. Maintains stability in a wide pH range (1.0–10.5). | [266] |
| Konjac gum (E425) | Amorphophallus konjac | Exhibits texture control and forms thermo–reversible and irreversible gels. Stable at 85°C in the presence of mild alkali (pH 9–10). | [267] |
| Low acyl Gellan gum/ Gelzan (E418) | Sphingomonas elodea | Forms strong, brittle gels that are heat– and pH–stable, providing a fibrous structure. | [268] |
| Locust Bean Gum (E410) | Ceratonia siliqua | Functions as a thickener, emulsifier, and stabilizer. Has a neutral flavor and prebiotic properties that improve hydration. | [269] |
| Gum Arabic/Acacia (E414) | Senegalia senegal | Highly water-soluble, low viscosity, and optimal at pH 3.5. Structure–forming transitions shift to higher pH as protein content increases. | [270] |
| Flaxseed gum | Linum usitatissimum. | Improves mouthfeel, moisture retention, and uniform texture while preventing ingredient separation. | [271] |
Table 11.
Types of Starches and Enzymes in Meat Analogues.
| Starches | Latin Name | Properties | Reference |
|---|---|---|---|
| Corn starch | Zea mays | Enhances viscosity and elasticity. Exhibits WAC and gelatinization properties. Reduces off–flavors. | [272] |
| Modified starch | Amylum modificatum | Enhances viscosity, elasticity, resilience, chewiness, and ductility. Reduces off–flavors and off–colors. | [273] |
| Potato starch | Solanum tuberosum | Increases fibrous content through thermo–irreversible gelatinization and freeze-thawing processes. | [274] |
| Wheat starch | Triticum vulgare | Exhibits thermal stability, retrogradation properties, and swelling index. It develops fibrousness due to glutenins and gliadins ratio. | [275] |
| Maltodextrin | – | Functions as a thickener, binding agent, and mouthfeel enhancer. Provides a creamy, spreadable texture similar to hydrogenated fat. | [276] |
| Transglutaminase | – | Induces crosslinks between protein molecules, improving binding properties and cutting strength. Enhances the overall texture. | [277] |
Table 12.
Types of Crosslinking Gelling Agents in Meat Analogues.
| Cross–linking Gelling Agents | Latin Name | Properties | Reference |
|---|---|---|---|
| Calcium ions (Ca2+, lactate, acetate) + Sodium alginate/ alginin | – | Functions as an adhesive and forms a cold–set gel in the presence of divalent cations, improving texture and moisture retention. | [278] |
| Agar | Gracilaria verrucosa | Serves as a plant–based alternative to gelatin. Provides palatability and does not require high sugar concentrations to form a gel. | [279] |
| Pectin | Saccharomyces cerevisiae | High– or low–methyl–esterified pectin forms gels when combined with sugar and acid. | [280] |
4.2. Coloring Agents
Natural colorants in plant–based meat analogues have attracted significant attention because of growing concern over synthetic colorants’ safety and health risks. Numerous natural colors, including betalains, carotenoids, anthocyanins, chlorophyll, and heme proteins, have been investigated for potential use in plant–based meat substitutes [281]. However, each pigment poses shortcomings concerning stability, pH sensitivity, thermal degradation, and oxidation, [282]. Leghemoglobin (LegH), one of the most used pigments, has shown great promise since it resembles myoglobin, the primary protein that gives animal meat its red hue. LegH is produced via microbial fermentation, wherein genetically modified yeast or bacteria are used for the production of a heme-containing protein with properties/functions similar to those of animal-derived myoglobin. Despite the success achieved in color matching, concerns about consumer acceptance of genetically modified organisms remain an ongoing challenge [281].
Chlorophyll–derived pigments have also been investigated as natural colorants. Chlorophyll is utilized to synthesize Fe–pheophytin through ion exchange methods, resulting in a more stable pigment [283]. However, chlorophyll and its derivatives are very sensitive to pH and oxidation, requiring stabilization techniques, including encapsulation and emulsification. Anthocyanin extraction from sources like black beans has been investigated for adding red to plant–based meat substitutes. Chromatographic examination of the isolated anthocyanins revealed that, on a temporal level, absorption peaks scanned at 277, 515, and 546 nm identified delphinidin–3–mono glucoside (D3G) at 3.1 and petunidin–3–mono glucoside (P3G) at 5.1 [283]. Such pigments provide natural alternatives to synthetic red dyes but are very pH–pH-sensitive, changing from red in acidic to blue or green in alkaline conditions [281]. Carotenoids like lycopene, beta–carotene, and astaxanthin are another class of natural pigments used in PBMAs. Depending on concentration and processing conditions, these pigments are sourced from tomatoes, carrots, and microalgae–based colors within a range of red to orange. Studies have also shown that the meshing of carotenoids with other pigments, like anthocyanins or betalains, can be more dynamic and stable. However, due to high lipophilicity, delivery systems are necessary to stabilize and enhance their bioavailability [282].
Betalains, which are produced from plants like red beet, have shown the ability to replicate the color of fresh meat. When red beet (0.4–1.5 mg/g) and cacao pigments (1.1–1.3 mg/g) were combined, the color produced matched that of well-cooked beef [284]. The ideal concentrations were determined using response surface methodology (RSM). However, betalains are very sensitive to light, temperature, and pH, necessitating co–pigmentation and microencapsulation. Microbial pigments from Monascus, Fusarium venenatum, and Neurospora intermedia have been studied for their ability to impart natural red colors with a pH-dependent color change in plant-based meat analogues [285]. However, certain microbial pigments produced by Monascus, can also create hazardous byproducts, requiring purification measures. Overall, the development of natural pigments for plant–based meat analogues must employ multidisciplinary approaches wherein pigment stability, safety, and consumer acceptability are considered.
Table 13.
Types of Coloring Agents in Meat Analogues.
| Coloring Agents | Latin Name | Shade | Pigments | Reference |
|---|---|---|---|---|
| Annatto extract/ Achiote | Bixa orellana | Yellow–Orange to Red | Carotenoids – Bixin and Norbixin | [286] |
| Caramel | Calamellus | Golden Brown | Heated sugars – Caramelization | [287] |
| Malt extract | Hordeum vulgare | Brown | Heated grains – Maillard reaction | [288] |
| Beet extract | Beta vulgaris | Red–Purple | Betalains – Betanin | [289] |
| Elderberry extract | Sambucus nigra | Purple–Red | Anthocyanins | [290] |
| Lycopene | Lycopersicon esculentum | Red | Carotenoids | [291] |
| Paprika | Capsicum annuum | Yellow–Orange to Red | Carotenoids – Capsanthin and Capsorubin | [292] |
| Turmeric |
Curcuma longa | Right Yellow | Curcuminoids | [293] |
| Spirulina extract | Arthrospira platensis | Blue–Green | Phycocyanins | [294] |
| Chlorophyllin | Chlorophylle | Green | Chlorophyll | [295] |
| Pomegranate concentrate | Punica granatum | Red–Purple | Anthocyanins and Ellagitannins | [296] |
4.3. Chemical Agents
The preparation of plant–based meat analogues involves the use of several chemical agents to regulate texture, stability, color, and other properties concerning product quality. These compounds primarily mimic the sensory and functional attributes of meat. Understanding their roles can allow manufacturers to advance the development of PBMAs to illustrate the characteristics and properties of meat adequately. Certain chemical agents are used to fortify the taste and control oxidation. One example is monosodium glutamate (MSG), which largely replicates the meaty flavor of animal protein from traditional meat [297]. Sodium erythorbate and ascorbic acid act as antioxidants that lower lipid oxidation and stabilize plant–based proteins, therefore minimizing undesirable off-flavors [298]. Dealing with oxidation is an ongoing challenge among plant–based formulations; naturally, highly unsaturated lipids can degrade quickly because of rancidity. Antioxidants eliminate this constraint and facilitate stability and freshness for extended periods. On the other hand, Choline chloride acts as an emulsifier, allowing fat and moisture to distribute uniformly within plant–based compositions. The emulsifying materials are highly useful in preventing different phases from separation and influencing the end product’s mouthfeel [299].
The texture of plant–based meat analogues is pivotal for mimicking the mouthfeel of conventional meat. Sodium metabisulfite and potassium metabisulfite are employed to maintain protein interactions and to inhibit discoloration [3]. Sodium chloride and calcium chloride are used to improve water retention and gelling properties, producing a distinctive chewier structure. Salt ions interact with plant proteins, enhancing their gelling ability and bonding power [300]. Glycerin is popularly included to stimulate moisture retention, thus allowing for a juicier bite. It acts as a humectant, preventing the product from becoming overly dry while maintaining textural quality throughout storage and cooking. Hydrocolloids such as methylcellulose and carrageenan are frequently used to give elasticity and create structural cohesion, strengthening the meat–like consistency [301]. With balance among these texture enhancing agents, manufacturers can achieve more realistic meat–like textures when fortifying plant-based products. Choline chloride offers further stabilizing support to these formulations by impeding phase separation whilst enhancing moisture retention [302].
Nonane, methanol, and acetonitrile aid lipid extraction and flavor compound analysis to identify ideal processing conditions. Hydrochloric acid and sodium hydroxide act as pH regulators during protein isolation, improving protein solubility and structural properties. Altering the pH is important to optimize protein functionality, whereby properties such as WHC and gel formation are affected [303]. Other popular natural colorants are purple potato powder, which provides a high brightness level, and beetroot extract, a rich source of betanin that imitates the red shades of raw meat. In addition, some analyses use bromophenol blue to monitor changes in pH. These additives ensure that product quality and visual presentation remain consistent, which is vital to how the products are received in the marketplace. Overall, they provide very critical considerations that formulate appropriate manufacturing parameters, satisfactory sensory qualities, functional stability, and consumer satisfaction with the finished product. Continuous advancements in ingredient technology and processing techniques are expected to refine the use of these chemical agents, further improving the sensory and nutritional attributes of plant-based meats.
Table 14.
Types of Chemical Agents in Meat Analogues.
| Cellulose | Chemical Formula | Properties | Reference |
|---|---|---|---|
| Carboxymethyl cellulose (CMC) | (C6H10O5)n | Odorless, white, or yellowish powder. Preserves structure and stops ingredient separation. | [304] |
| Methylcellulose (MC) (E461) | C20H38O11 | Known for its binding capacity and unique reversible thermal gelation. Functions as a stabilizer and emulsifier. | [241] |
| Hydroxypropyl methylcellulose (HPMC) (E464) | C56H108O30 | Provides binding, gelling, texture improvement, and stabilization. Functions as an emulsifier with thermal gelation properties. | [305] |
4.4. Flavoring Agents
Flavor and sensory aspects are central to consumer acceptance of plant–protein–based meat substitutes. This quest to imitate meat comes with the challenge of replicating taste profiles via Maillard reaction, lipid oxidation, and umami compounds. Thus, various odor and umami enhancers have been integrated into multiple formulations, including yeast extracts, nucleotides, and lipid-based fermentation products [302]. In meat analogues, hydrocolloids like methylcellulose, konjac gum, and carrageenan can enhance binding and water retention, improving juiciness and cohesiveness [306]. The addition of monosodium glutamate (MSG) and mushroom extracts increases the richness of flavors in plant-based alternatives. Aging processes often intensify this flavor by breaking down proteins into umami-rich amino acids. Yeast extracts are an inexpensive flavor enhancer that owes their umami character to nucleotides like Inosine monophosphate (IMP) and guanosine monophosphate (GMP). Research indicates that when combined with amino acids and peptides from hydrolysed proteins, yeast extract enhances the meat–like tastes of plant-based alternatives. Soy sauce and Aspergillus oryzae–fermented products impart savory notes and mask undesirable bean flavors from a legume–based proteins. The effects of fermented plant extracts derived primarily from wheat and barley have also been identified, as they curtail bitterness and improve palatability [297].
Many lipid-based solutions–including vegetable oils infused with heme–like molecules and volatile compounds give meat analogues their characteristic aroma and juiciness. Heme proteins like soy–derived leghemoglobin closely mimic the iron-rich animal muscle protein, myoglobin [307]. Other lipid sources such as coconut oil and algal lipids contribute to mouthfeel and act as delivery systems for lipid-soluble volatile compounds produced during TMP. The meaty fragrance production mainly depends on lipid oxidation pathways, where PUFAs serve as precursors to aldehydes and ketones [300]. Heat-induced reduction of sugars and amino acids by the Maillard reaction produces a new class of volatile chemicals, such as pyrazines, thiols, and aldehydes that resemble the distinctive grilled and roasted flavors of cooked meat. It has been demonstrated that adding cysteine and ribose to plant–based analogues increases the synthesis of volatile chemicals. [308]. Furthermore, sulfur-containing compounds such as thiols derived from methionine contribute significantly to the umami notes of meat alternatives [118]. Modern biotechnology has brought about additional fermentation-derived aroma compounds, particularly short–chain fatty acids, esters, and ketones with fatty, cheesy, and buttery notes, via microbial strains such as Bacillus and Lactobacillus.
Modifying plant proteins through enzymatic action has also been used to minimize off–flavor associated with grassy and beany notes, which are usually specific to protein sources like soy and pea. Palatability is improved by the enzymatic breakdown of proteins into smaller peptides and free amino acids [309]. Different innovative culinary approaches are being explored, including precision fermentation and metabolic engineering of yeast strains to generate customized flavor compounds like those found in meat [107]. Despite advancements, several challenges remain in achieving a balanced meat-like flavor. The interactions between plant proteins and added flavor enhancers are complicated and lead to inconsistent tastes among different formulations. Variation in the flavors of meat analogues is largely influenced by ingredient sources, processing techniques, and storage conditions, all of which contribute to consumer taste perception [310]. With consumers shifting towards clean-label foods, more integrated studies should focus on plant-derived umami compounds from seaweeds, tomato concentrates, and miso fermentation in place of artificial additives [76].
Table 15.
Types of Flavoring Powders and Pulps in Meat Analogues.
| Powders and Pulps | Latin Name | Properties | Reference |
|---|---|---|---|
| Purple potato powder | Ipomoea batatas | Provides natural color, enhances texture, and is rich in antioxidants. | [311] |
| Konjac powder | Amorphophallus konjac | Functions as a gelling agent, imparts a chewy texture, and is low in calories. | [312] |
| Meat flavor powder |
– | Adds a savory, umami–rich flavor. | [313] |
| Paprika powder |
Capsicum annuum | Contributes color with a mild, slightly sweet flavor. | [314] |
| Cumin/Jeera powder |
Cuminum cyminum | Imparts an earthy, warm, and slightly nutty flavor. | [315] |
| Citric acid powder | Acidum citricum | Provides a tangy flavor and helps extend shelf life. | [316] |
| Ascorbic acid powder | Acidum ascorbicum | A source of vitamin C, prevents oxidation, and maintains freshness. | [317] |
| Mustard powder | Brassica juncea | Delivers a sharp, tangy, and slightly spicy flavor. | [318] |
| Onion powder |
Allium cepa | Adds a rich, savory base flavor. | [313] |
| Garlic powder | Allium sativum | Enhances depth and richness of flavor. | [314] |
| Panela/Jaggery powder | Saccharum officinarum | Provides a subtle sweetness that balances flavors. | [319] |
| Tomato powder | Solanum lycopersicum | Enhances natural color and imparts a rich, tangy flavor. | [320] |
| Pepper powder | Piper nigrum | Adds depth with a bold, spicy kick. | [321] |
| Flaxseed powder | Linum usitatissimum | High in omega–3 fatty acids and dietary fiber, contributing to texture and nutrition. | [322] |
| Ground jackfruit pulp | Artocarpus heterophyllus | Provides a fibrous texture and is a good source of dietary fiber. | [323] |
Table 16.
Types of Flavoring Agents in Meat Analogues.
| Flavoring Agents | Properties | Reference |
|---|---|---|
| Monosodium glutamate/ Ajinomoto | A sodium salt of glutamic acid that enhances savory, umami flavors. | [324] |
| Yeast extracts powder | Provides umami flavor and helps mask bitter or earthy off–notes. | [301] |
| Soy leghemoglobin | A heme protein derived from genetically modified yeast; it enhances the meaty taste in plant–based products. | [325] |
| Beet and lemon juice | Adds natural red color and brightness to foods. | [310] |
| Cooked onion and carrot juice concentrates | Enhances savory depth, while natural carrot pigments improve color. | [326] |
| Salt | Enhances overall flavor by balancing and intensifying taste perception. | [327] |
Table 17.
Types of Sweetening Agents in Meat Analogues.
| Sweeteners | Properties | Reference |
|---|---|---|
| Dextrose | 20% less sweet than sucrose, contributes to the Maillard reaction and caramelization when combined with cysteine. | [328] |
| Glucose | Actively participates in the Maillard reaction, enhancing browning and flavor development. | [329] |
| Sucrose | Serves as a bulking agent and preservative. Hydrolyzed into glucose and fructose during processing. | [330] |
| Fructose | Undergoes the Maillard reaction, though its effect is less pronounced compared to glucose and dextrose. | [331] |
| Sugar alcohols (erythritol, sorbitol) | Lower glycemic impact, contribute to a smooth, creamy texture, help retain moisture, and prevent drying. | [332] |
| Brown Sugar | Imparts a rich caramel flavor due to its molasses content. | [333] |
Table 18.
Emulsification Agents in Meat Analogues.
| Emulsifying Agents | Latin Name | Properties | Reference |
|---|---|---|---|
| Corn oil | Maydis oleum raffinatum | Rich in PUFAs, primarily linoleic acid (58–62%). Contains high levels of phytosterols (8,300–25,500 ppm) and tocopherols (1,130–1,830 ppm). | [334] |
| Soy oil | Soiae oleum raffinatum | Contains PUFAs like linoleic acid (48–58%) and isoflavones. | [335] |
| Peanut/ Groundnut oil |
Arachis hypogaea | High in monounsaturated fats, primarily oleic acid (45–72%). Mild flavor with a high smoke point. Used in oleogel production for fat stabilization. | [336] |
| Rapeseed oil | Brassica campestris | Rich in unsaturated fats, particularly oleic acid. Contributes to smooth texture and stability. | [337] |
| Canola oil | Brassica napus | Contains 7% saturated fat, monounsaturated fat, and ALA omega–3 fatty acids. It has a mild flavor, and a high smoke point. | [338] |
| Sunflower oil |
Helianthus annuus | High in PUFAs, mainly linoleic acid (55–75%). High smoke point and naturally rich in vitamin E. | [339] |
| Safflower oil | Carthamus tinctorius | Contains high levels of PUFAs, mainly linoleic acid. Enhances texture and moisture retention. | [340] |
| Palm oil |
Elaeis guineensis | Semi–solid at room temperature, contributing to moisture retention and succulence. | [341] |
| Red palm oil | Elaeis guineensis (or) Elaeis oleifera | Contains saturated and monounsaturated fats, with high palmitic acid and carotenoids. Enhances mouthfeel, juiciness, and richness in formulations. | [342] |
| Coconut oil | Cocos Nucifera | 92% saturated fat, making it a stable fat source. Used in 3D–printable fat analogues for meat substitutes when combined with glucomannan. | [343] |
| Orange oil | Citrus sinensis | Acts as a masking agent to reduce the bean odor of soy–based products. Exhibits strong antioxidant activity (DPPH radical scavenging activity). | [344] |
5. Existing Technology
5.1. Single Screw Extrusion
Plant–based meat analogues (PBMAs) are majorly produced using single–screw extrusion due to their simplicity and cheap operating costs. A single revolving screw is used to apply pressure to a combination of plant proteins, water, and other ingredients. Heat and shear from this process cause the proteins to become denatured and fibrous. It is capable of producing a range of textures, including meat–like ones. Nevertheless, in comparison to more sophisticated techniques such as high–moisture extrusion (HME) or twin–screw extrusion, the fibrous structures produced are generally less defined, leading to a product that lacks the chewiness and structural integrity of actual meat. Despite this limitation, single–screw extrusion remains a feasible option for producing plant–based meat alternatives in large quantities, especially for products such as textured vegetable protein (TVP). This extrusion process is also energy efficient and runs at comparatively low expenses, making it ideal for bulk production [345]. Furthermore, improvements in extrusion parameters like screw speed, moisture content, and temperature can improve the texture and protein quality of the finished product [346]. Further, it has also been proven to enhance digestibility in plant proteins by denaturing, unfolding, and cross–linking them during the process. While twin–screw extrusion is more suitable for producing a meat-like structure compared to single–screw extrusion, its economic feasibility and scalability make it a widely accessible technology for developing PBMAs [347].
5.2. Twin Screw Extrusion
Twin–screw extrusion (TSE) is a more sophisticated and intricate process than single–screw extrusion, providing better control over the protein structure. It involves the use of two intermeshed screws, generating greater shear stress and enabling easier manipulation of plant protein networks, which aids in forming layered and fibrous structures. The use of twin–screw extrusion has been shown to be especially useful in producing high–moisture PBMAs that closely mimic the meat–like mouthfeel and texture of natural meat. With the fine control of processing conditions like screw speed, temperature, and pressure, TSE can produce fibrous, elastic, and meat–like products [348]. Specifically, TSE enables protein molecules to align into layered configurations that mimic muscle fibers in meats from animals [308]. The technique can also combine different protein sources, such as soy, wheat gluten, and pea protein, to maximize the final product’s mouthfeel and structural quality [349]. Current research aims at maximizing the extrusion process, such as screw design and thermal versus mechanical energy input balance, to enhance product quality and manufacturing productivity. Through optimization of these parameters, TSE has the potential to meet the increasing demand for PBMAs.
5.3. High Moisture Extrusion
High moisture extrusion (HME) is among the most effective techniques for manufacturing PBMAs that mimic the fibrous structure of animal meat [309]. HME is a process where a protein blend is extruded at high pressure and temperature, typically with a moisture level above 70%. The synergy of shear forces, heat, and pressure produces a fibrous structure that is similar to the texture of whole–muscle meats such as chicken and beef [350]. One of the significant benefits of HME is that it can enhance the mouthfeel of PBMAs. By adjusting temperature, screw rate, and moisture, the texture may be modified [351]. To encourage protein alignment and limit expansion, HME uses a twin–screw extruder and cold die at above 50% moisture content, resulting in a final product that resembles structured meat [352]. The capability of synthesizing analogues from proteins such as soy, hemp, pea, lentils, and faba beans has made HME a widely accepted process in the manufacture of PBMAs [353]. Despite its merits, HME is energy-demanding and needs sophisticated machinery, which may raise production costs and make scaling difficult [297]. However, HME is still among the most promising technologies for PBMA production, with high productivity, nutrient retention, and minimal waste production [308]. Although soy, wheat, and peanut proteins are frequently used in HME, research on other protein sources, including hemp and lupin, has increased the technology’s applicability.
5.4.3. D–Bioprinting
3D–bioprinting technology has been a groundbreaking tool in developing PBMAs. Recent research by Keerthana et al. (2020), illustrated the capability of 3D–printing to manufacture mushroom–based meat analogues. [152]. 3D–printing or additive manufacturing is used to produce customized texture, shape, and nutritional composition, where food bioinks are added in layers to produce meat analogues with the texture of muscle fibers [354]. However, printing mushroom products is difficult owing to its fibrous network, which can clog the printing nozzle. Extrusion, binder jetting, and inkjet printing are some 3D printing techniques used in the food processing industry. Foods that are semi–solid, such meat purees and doughs, work well for extrusion–based printing. Inkjet printing is utilized in low–viscosity fluids; whereas binder jetting accommodates powder forms such as sugar or cocoa powders. Of the various methods, the extrusion technique stands out as the most widely researched for food printing, where nozzle diameter, material deposition rate, and print speed influence the printability and structure of the final product [355]. The other two main aspects of 3D food printing include its capacity to provide high degrees of customization, such that food texture, shape, and nutritional composition can be tailored. Studies have also investigated the application of food hydrocolloids to enhance printability [356,357]. While there are limitations in mirroring the intricate fibrous texture of animal meat, advancements in protein sources continue to enhance the quality of meat analogues [358,359].
5.5.4. D–Bioprinting
Building upon this, 4D food printing adds the aspect of time and environmental stimuli so that printed foods can transform in terms of shape, color, taste, or nutritional content. This new technology uses food–grade shear–thinning inks, which can flow when subjected to external force but become stable once the force is released. This property allows for the development of food structures that are capable of dynamically changing during printing or post-processing. One of the capabilities of 4D printing is the conversion of shiitake mushrooms and potato purees, with ergosterol being converted to vitamin D2 through UV irradiation [360]. This concomitant conversion illustrates how 4D printing can produce sophisticated shapes in food and improve nutritional qualities. Synergistic integration of 4D printing with other processing steps can add much value to food products [361]. This capacity to alter food structure and nutritional value creates new opportunities for personalized nutrition and functional foods [360]. 4D–bioprinting technology has excellent potential for developing novel, flexible food items that can satisfy various customer demands and tastes [152].
5.6. Shear Cell Technology
Shear Cell Technology (SCT) is used to create plant-based meat substitutes that closely mimic the structure of animal meat [14]. It consists of rotating blades or cylinders that subject plant protein blends to high shear forces, forming elongated, fibrous strands [362]. SCT can control the network alignment and protein density with high precision as it increases the cohesiveness and elasticity of the protein [307]. SCT also consumes less energy than conventional extrusion processes, making it more economical for small-scale production [363]. The resultant products have a very high level of anisotropy and can mimic fibrous characteristics effectively [364]. However, SCT also faces limitations in scaling up and ensuring consistency in the finished product. Control of shear force and temperature is needed to achieve the desired texture, which can be challenging in large–scale production. Denaturation of the proteins due to excessive shear stress can potentially affect the quality of PBMAs. Nonetheless, SCT remains a promising technology for producing PBMAs, with ongoing studies focused on improving processing efficiency.
Figure 3.
(A) Schematic diagram of the high–moisture extrusion process and microscopic images of Pea Protein Isolate (PPI):Soybean Meal (SM) meat analogues at varying concentration ratios. Reproduced with permission [364]. Copyright Multidisciplinary Digital Publishing Institute (MDPI), 2024. (B) Cryo–images of high–moisture extruded meat analogues in dead–stop trials at TMaterial = 95°C and 125°C, with extrudate morphology at four different material temperatures. (C, D) Velocity, shear stresses, tensile stresses, and temperature profiles along the die length. Reproduced with permission [365]. Copyright Multidisciplinary Digital Publishing Institute (MDPI), 2021.
Figure 3.
(A) Schematic diagram of the high–moisture extrusion process and microscopic images of Pea Protein Isolate (PPI):Soybean Meal (SM) meat analogues at varying concentration ratios. Reproduced with permission [364]. Copyright Multidisciplinary Digital Publishing Institute (MDPI), 2024. (B) Cryo–images of high–moisture extruded meat analogues in dead–stop trials at TMaterial = 95°C and 125°C, with extrudate morphology at four different material temperatures. (C, D) Velocity, shear stresses, tensile stresses, and temperature profiles along the die length. Reproduced with permission [365]. Copyright Multidisciplinary Digital Publishing Institute (MDPI), 2021.

5.7. Electrospinning
Electrospinning is a possible technique for creating micro–fibrous structures in PBMAs, which uses an electric field to draw a molten substance or polymer solution into thin strands. Plant proteins such as pea and soy may be electrospun to produce PBMAs, which are a network of fibres that resemble the fibrous structure of animal muscle tissue [350]. The fibers can be harvested onto a substrate to create a scaffold, which can be processed further into a meat–like form. Electrospinning is especially beneficial for producing textured products with a high surface area and increased protein alignment, which replicates the muscle fiber structure. This process enables great control over the diameter, orientation, and density of the fibers, which directly influence the end texture and mouthfeel of the PBMA [307]. Nevertheless, it poses problems of scaling up to industrial applications since it is still labor–intensive and necessitates optimizing solution viscosity, voltage, and the collection system [366]. Despite complications, electrospinning is promising for fabricating plant-based meat products with complex fiber structures mimicking the mouthfeel and texture of actual meat. Future studies will probably be directed toward optimizing the electrospinning process to enhance yield and scalability for commercial production.
5.8. Antisolvent Precipitation
Antisolvent precipitation is a technique employed to create plant protein networks through the precipitation of proteins from a solvent system, commonly using ethanol or acetic acid as solvents [14]. In antisolvent precipitation, a concentrated protein solution is combined with water or another antisolvent, precipitating the proteins and creating a solid network structure. This technique allows for thick, cohesive protein matrices with meat–like textures and is typically used with proteins like zein, soy, or wheat gluten. The networked precipitates can be additionally treated and processed to enhance texture and stability, which makes them favorable for PBMAs. Antisolvent precipitation provides an effective and straightforward approach to producing protein networks with high structural integrity. By regulating precipitation conditions, including temperature, solvent, and protein concentration, it is possible to adjust the final product’s texture [258]. One disadvantage of this approach is the possibility of denaturation or loss of protein functionality, particularly when employing severe solvents such as ethanol or acetic acid. Moreover, it might be necessary to add steps to eliminate impurities of solvents and test the product’s safety for eating. In spite of these complexities, antisolvent precipitation remains a valuable process for creating PBMAs that have better texture and protein integrity [367].
5.9. Mechanical Elongation
Mechanical elongation is a technique used to stretch plant protein matrices to replicate meat’s fibrous structure. Protein mixes are often subjected to a mechanical force throughout the procedure, which causes the proteins to lengthen into long fibres resembling the muscle fibers in animal flesh [14]. The elongation process can be done using various techniques, including stretching doughy protein mixtures or employing specific machines, stretching and twisting the protein network. Mechanical elongation is usually blended with other processing methods, including high–moisture extrusion, to improve the final product’s texture and extensibility [370]. The final product is more integrative and contains better chewability, bringing the texture closer to animal muscle tissue. One of the strengths of mechanical stretching is that it can regulate the extent of fiber alignment and protein matrix density, which greatly influences the final texture of the PBMA. Yet, some of the challenges are to optimize the elongation conditions, i.e., the applied force, the protein blend employed, and the moisture level, to get the required texture without over–denaturing the proteins. Mechanical elongation is also energy–consuming and may require special equipment. Despite all these difficulties, this method has successfully produced high–quality PBMAs with improved fibre structures that resemble whole–muscle meat[368].
Figure 4.
(A) 3D–printed meat analogues with different infill patterns and ratios. (B) Viscosity curves of composite inks: (a) textured soybean protein (TSP) with varying hydrocolloids, and (b) drawing soy protein (DSP) with varying hydrocolloids. Reproduced with permission [368]. Copyright Multidisciplinary Digital Publishing Institute (MDPI), 2021. (C, D) SEM images and texture profile analysis of fibrous material produced from zein (ZP) by mechanical elongation (ME), antisolvent precipitation (AS), and electrospinning (ES) with SPI control. Reproduced with permission [14]. Copyright Elsevier, 2020.
Figure 4.
(A) 3D–printed meat analogues with different infill patterns and ratios. (B) Viscosity curves of composite inks: (a) textured soybean protein (TSP) with varying hydrocolloids, and (b) drawing soy protein (DSP) with varying hydrocolloids. Reproduced with permission [368]. Copyright Multidisciplinary Digital Publishing Institute (MDPI), 2021. (C, D) SEM images and texture profile analysis of fibrous material produced from zein (ZP) by mechanical elongation (ME), antisolvent precipitation (AS), and electrospinning (ES) with SPI control. Reproduced with permission [14]. Copyright Elsevier, 2020.
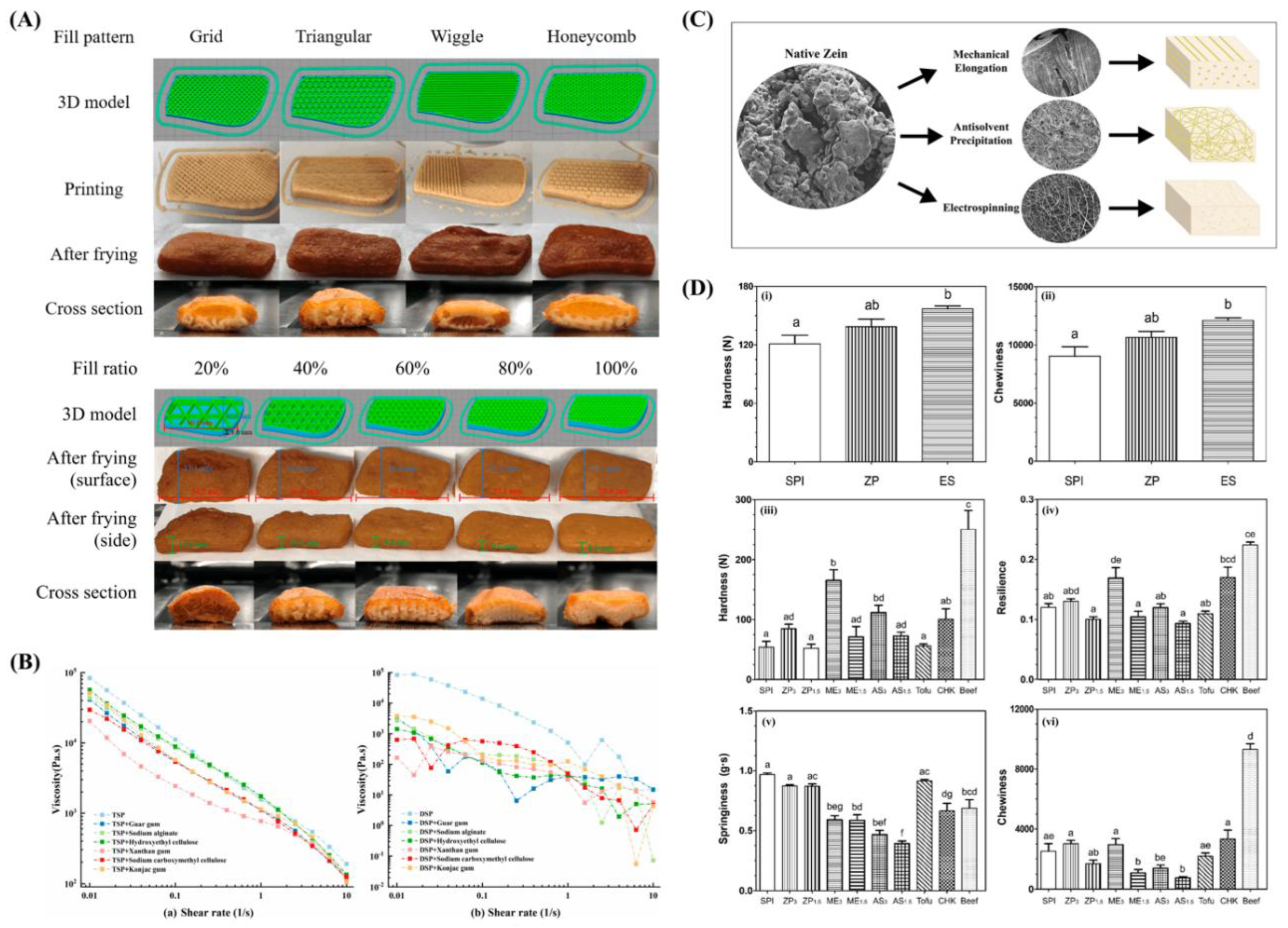
6. Conclusion
The PBMA industry is constantly expanding as more people prefer meat alternatives due to rising demand, ecological concerns, and changing preferences. From early soy–based originals to evolved multi-ingredient analogues present today, the history of PBMAs demonstrates perseverance in gaining sensory and nutrient equivalency to conventional meat with no concomitant environmental repercussions. The development of meat analogues is a precise balancing process, requiring proportionate blends of plant–derived proteins, polysaccharides, and lipids. This synergy of ingredients created through plant biochemistry, allows for the production of products that highly mimic the mouthfeel associated with meat consumption. Technological advancements also have a significant role in controlling both material composition and architecture. These technologies, coupled with novel flavoring and coloring agents, enhance the overall taste and texture. The alternative meat sector is expected to continue its growth in the near future. Nonetheless, several challenges still remain, including the presence of off–note flavor profiles and micronutrient deficiencies. These limitations can be addressed through further exploration into volatile and non–volatile compounds to improve savoriness, minimize additives, and use fortified nutrient–dense plant resources per clean–label trends. Meeting consumer needs in the context of taste, texture, acceptability, and affordability are also critical to the success of the meat analogues industry. Essential strategies for overcoming food neophobia include encouraging familiarity with PBMAs through effective marketing, raising awareness, and emphasizing the role of sustainability in conscious food production and consumption. PBMAs have the potential to advance the global food scene, where increased investments in R&D, curated formulations, and processing technologies contribute to the development of ethical consumer-oriented food systems.
Author Contributions
The submitted original manuscript is not part of any previously published article and is not under consideration elsewhere. All the authors of this manuscript have significantly contributed to its preparation as follows: Asfa Sultana Sher (Conceptualization, Data Collection, Data Interpretation, Manuscript Drafting), Vidya Sri Padmanaban (Conceptualization, Data Collection, Data Interpretation, Manuscript Drafting), Dr. Swathi Sudhakar (Conceptualization, Supervision, Critical Revisions, Manuscript Review), Dr. Lakshminath Kundanati (Conceptualization, Supervision, Critical Revisions, Manuscript Review).
Funding
No funding was received to assist with the preparation of this manuscript.
Data Availability Statement
Data sharing is not applicable to this article as no new data was generated during this study.
Acknowledgments
Not applicable.
Conflict of Interest
All authors have no conflicts of interest.
References
- FAO (2018) The future of food and agriculture - Alternative pathways to 2050. 224.
- Singh U, Tiwari P, Kelkar S, et al. (2023) Edible mushrooms: A sustainable novel ingredient for meat analogs. eFood 4:e122. [CrossRef]
- Yuan X, Jiang W, Zhang D, et al. (2022) Textural, sensory and volatile compounds analyses in formulations of sausages analogue elaborated with edible mushrooms and soy protein isolate as meat substitute. Foods 11:52. [CrossRef]
- Gamarra-Castillo O, Echeverry-Montaña N, Marbello-Santrich A, et al. (2022) Meat Substitute Development from Fungal Protein (Aspergillus oryzae). Foods 11:2940. [CrossRef]
- Ritchie H (2019) Food production is responsible for one-quarter of the world’s greenhouse gas emissions. Our World in Data.
- Ederer P, Baltenweck I, Blignaut JN, et al. (2023) Affordability of meat for global consumers and the need to sustain investment capacity for livestock farmers. Anim Front 13:45. [CrossRef]
- 2015; IARC (2015) Red Meat and Processed Meat IARC Monographs on the Evaluation of Carcinogenic Risks to Humans Volume 114.
- Mazumder MAR, Sukchot S, Phonphimai P, et al. (2023) Mushroom–Legume-Based Minced Meat: Physico-Chemical and Sensory Properties. Foods 12:. [CrossRef]
- Zahari I, Ferawati F, Helstad A, et al. (2020) Development of High-Moisture Meat Analogues with Hemp and Soy Protein Using Extrusion Cooking. Foods 2020, Vol 9, Page 772 9:772. [CrossRef]
- Caporgno MP, Böcker L, Müssner C, et al. (2020) Extruded meat analogues based on yellow, heterotrophically cultivated Auxenochlorella protothecoides microalgae. Innovative Food Science & Emerging Technologies 59:102275. [CrossRef]
- Caputo V, Sun J, Staples AJ, Taylor H (2024) Market outlook for meat alternatives: Challenges, opportunities, and new developments. Trends Food Sci Technol 148:104474. [CrossRef]
- Małecki J, Muszy S, Sołowiej BG, et al. (2021) Proteins in Food Systems—Bionanomaterials, Conventional and Unconventional Sources, Functional Properties, and Development Opportunities. Polymers (Basel) 13:2506. [CrossRef]
- Taghian Dinani S, Broekema NL, Boom R, van der Goot AJ (2023) Investigation potential of hydrocolloids in meat analogue preparation. Food Hydrocoll 135:108199. [CrossRef]
- Mattice KD, Marangoni AG (2020) Comparing methods to produce fibrous material from zein. Food Research International 128:108804. [CrossRef]
- Ahmad MI, Farooq S, Ali U, et al. (2024) Molecular phenomena associated with the formation of fibrous structure of plant-based meat analogues. Trends Food Sci Technol 153:104743. [CrossRef]
- Gulkirpik E, Donnelly A, Nowakunda K, et al. (2023) Evaluation of a low-resource soy protein production method and its products. Front Nutr 10:1067621. [CrossRef]
- Choi HW, Hahn J, Kim HS, Choi YJ (2025) Microstructural and textural characteristics of blend gels and high-moisture meat analogs of soy protein isolate and faba bean protein concentrate. Food Chem 467:142184. [CrossRef]
- Jeon YH, Gu BJ, Ryu GH (2023) Investigating the Potential of Full-Fat Soy as an Alternative Ingredient in the Manufacture of Low- and High-Moisture Meat Analogs. Foods 2023, Vol 12, Page 1011 12:1011. [CrossRef]
- Zhang T, Dou W, Zhang X, et al. (2021) The development history and recent updates on soy protein-based meat alternatives. Trends Food Sci Technol 109:702–710. [CrossRef]
- Toomer OT, Oviedo-Rondón EO, Ali M, et al. (2024) Full-Fat Soybean Meals as an Alternative Poultry Feed Ingredient-Feed Processing Methods and Utilization-Review and Perspective. Animals (Basel) 14:. [CrossRef]
- Kumari S, Dambale AS, Samantara R, et al. (2025) Introduction, History, Geographical Distribution, Importance, and Uses of Soybean (Glycine max L.). Soybean Production Technology 1–17. [CrossRef]
- Lin W, Barbut S (2025) Effects of 0–12% soy proteins (four texturized and one isolate) on a lean hybrid meat system: cooking loss, texture, dynamic rheology, microstructure, and T2 NMR. Applied Food Research 5:100747. [CrossRef]
- Cho SY, Ryu GH (2021) Effects of Oyster Mushroom Addition on Physicochemical Properties of Full Fat Soy-based Meat Analog by Extrusion Process. Food Engineering Progress 25:85–94. [CrossRef]
- Hafizur Rahman Bhuiyan M, Yeasmen N, Ngadi M (2024) Restructuring plant-derived composites towards the production of meat-analog based coated fried food. Food Chem 443:138482. [CrossRef]
- Schreuders FKG, Dekkers BL, Bodnár I, et al. (2019) Comparing structuring potential of pea and soy protein with gluten for meat analogue preparation. J Food Eng 261:32–39. [CrossRef]
- Shanthakumar P, Klepacka J, Bains A, et al. (2022) The Current Situation of Pea Protein and Its Application in the Food Industry. Molecules 27:5354. [CrossRef]
- Rogers H, Dora M, Tsolakis N, Kumar M (2024) Plant-based Food Supply Chains: Recognising Market Opportunities and Industry Challenges of Pea Protein. Applied Food Research 4:100440. [CrossRef]
- Zhang L, Apea-Bah FB, Chen X, et al. (2023) The physicochemical and structural properties and in vitro digestibility of pea starch isolated from flour ground by milling and air classification. Food Chem 419:136086. [CrossRef]
- Zhang S, Huang W, Roopesh MS, Chen L (2022) Pre-treatment by combining atmospheric cold plasma and pH-shifting to prepare pea protein concentrate powders with improved gelling properties. Food Research International 154:111028. [CrossRef]
- Sha L, Koosis AO, Wang Q, et al. (2021) Interfacial dilatational and emulsifying properties of ultrasound-treated pea protein. Food Chem 350:129271. [CrossRef]
- Ge J, Sun C, Chang Y, et al. (2023) Understanding the differences in heat-induced gel properties of twelve legume proteins: A comparative study. Food Research International 163:112134. [CrossRef]
- Li Y, Qi X, Rong L, et al. (2024) Effect of gellan gum on the rheology, gelling, and structural properties of thermally induced pea protein isolate gel. Food Hydrocoll 147:109379. [CrossRef]
- Sajib M, Forghani B, Kumar Vate N, Abdollahi M (2023) Combined effects of isolation temperature and pH on functionality and beany flavor of pea protein isolates for meat analogue applications. Food Chem 412:135585. [CrossRef]
- Johansson M, Xanthakis E, Langton M, et al. (2021) Mixed legume systems of pea protein and unrefined lentil fraction: Textural properties and microstructure. LWT 144:111212. [CrossRef]
- Wang J, Kadyan S, Ukhanov V, et al. (2022) Recent advances in the health benefits of pea protein (Pisum sativum): bioactive peptides and the interaction with the gut microbiome. Curr Opin Food Sci 48:100944. [CrossRef]
- Sidiq M, Muzzaffar S, Masoodi FA, Irfan S (2024) Impact of ultrasonication on the physicochemical, structural, thermal and functional properties of Mung bean protein. Measurement: Food 16:100205. [CrossRef]
- Brishti FH, Chay SY, Muhammad K, et al. (2021) Texturized mung bean protein as a sustainable food source: Effects of extrusion on its physical, textural and protein quality. Innovative Food Science & Emerging Technologies 67:102591. [CrossRef]
- Han F, Moughan PJ, Li J, Pang S (2020) Digestible Indispensable Amino Acid Scores (DIAAS) of Six Cooked Chinese Pulses. Nutrients 12:3831. [CrossRef]
- Hammer L, Moretti D, Bétrix CA, et al. (2024) In vitro DIAAS of Swiss soybean cultivars using the INFOGEST model: Increase in protein quality from soybean to soymilk and tofu. Food Research International 178:113947. [CrossRef]
- Herreman L, Nommensen P, Pennings B, Laus MC (2020) Comprehensive overview of the quality of plant- And animal-sourced proteins based on the digestible indispensable amino acid score. Food Sci Nutr 8:5379–5391. [CrossRef]
- Siddiquy M, Ghamry M, Golshany H, et al. (2024) Structural and functional characterization of mung bean protein-peach gum conjugate through the Maillard reaction as a novel encapsulation agent. Prog Org Coat 188:108201. [CrossRef]
- Guo R, Sun B, Zhu Y, et al. (2024) Low-moisture extruded mung bean protein isolate and wheat gluten: Structure, techno-functional characteristics and establishment of rehydration kinetics of the products. LWT 210:116844. [CrossRef]
- De Angelis D, Opaluwa C, Pasqualone A, et al. (2023) Rheological properties of dry-fractionated mung bean protein and structural, textural, and rheological evaluation of meat analogues produced by high-moisture extrusion cooking. Curr Res Food Sci 7:100552. [CrossRef]
- Kartikeyan A, Vasudevan V, Peter AJ, et al. (2022) Effect of incubation period on the glycosylated protein content in germinated and ungerminated seeds of mung bean (Vigna radiata (L.) Wilczek). Int J Biol Macromol 217:633–651. [CrossRef]
- Hou D, Zhao Q, Yousaf L, et al. (2020) Consumption of mung bean (Vigna radiata L.) attenuates obesity, ameliorates lipid metabolic disorders and modifies the gut microbiota composition in mice fed a high-fat diet. J Funct Foods 64:103687. [CrossRef]
- Augustin MA, Cole MB (2022) Towards a sustainable food system by design using faba bean protein as an example. Trends Food Sci Technol 125:1–11. [CrossRef]
- Tuccillo F, Kantanen K, Wang Y, et al. (2022) The flavor of faba bean ingredients and extrudates: Chemical and sensory properties. Food Research International 162:112036. [CrossRef]
- Saldanha do Carmo C, Knutsen SH, Malizia G, et al. (2021) Meat analogues from a faba bean concentrate can be generated by high moisture extrusion. Future Foods 3:100014. [CrossRef]
- De Silva D, Liu Y, Smith MA, et al. (2024) Distribution of Vicine, Convicine and Levodopa in Faba Bean Plant Tissues Determined by UltraHigh Performance Liquid Chromatography-Electrospray Ionization Mass Spectrometry. Journal of Chromatography Open 5:100127. [CrossRef]
- Choi YM, Shin MJ, Lee S, et al. (2024) Anti-nutrient factors, nutritional components, and antioxidant activities of faba beans (Vicia faba L.) as affected by genotype, seed traits, and their interactions. Food Chem X 23:101780. [CrossRef]
- Liu C, Pei R, Heinonen M (2022) Faba bean protein: A promising plant-based emulsifier for improving physical and oxidative stabilities of oil-in-water emulsions. Food Chem 369:130879. [CrossRef]
- Fan Y, Annamalai PK, Bhandari B, Prakash S (2025) Characteristics of faba bean protein-based high-moisture meat analogues incorporating brewers’ spent grain through extrusion. Innovative Food Science & Emerging Technologies 100:103919. [CrossRef]
- Elshamy AF, Rosentrater KA, Ghnimi S, et al. (2025) High-moisture meat analogs produced from dry-fractionated Faba bean, yellow pea, and functional soy proteins: Effects of mixture design and extrusion parameters on texture properties. Innovative Food Science & Emerging Technologies 100:103927. [CrossRef]
- Kamani MH, Liu J, Fitzsimons SM, et al. (2024) Determining the influence of fava bean pre-processing on extractability and functional quality of protein isolates. Food Chem X 21:101200. [CrossRef]
- Kantanen K, Chamlagain B, Succi V, et al. (2024) Fermentation of faba bean flour as a pre-treatment in high-moisture extrusion for simultaneous fortification of vitamin B12 and reduction of raffinose oligosaccharides in meat analogues. Future Foods 10:100518. [CrossRef]
- Shrestha S, Hag L van t., Haritos VS, Dhital S (2021) Lupin proteins: Structure, isolation and application. Trends Food Sci Technol 116:928–939. [CrossRef]
- Kebede YS, Teferra TF (2023) Isoelectric point isolation and characterization of proteins from lupine cultivars as influenced by chemical and thermal treatments. Heliyon 9:e14027. [CrossRef]
- Palanisamy M, Töpfl S, Berger RG, Hertel C (2019) Physico-chemical and nutritional properties of meat analogues based on Spirulina/lupin protein mixtures. European Food Research and Technology 245:1889–1898. [CrossRef]
- Abreu B, Lima J, Rocha A (2023) Consumer Perception and Acceptability of Lupin-Derived Products: A Systematic Review. Foods 12:1241. [CrossRef]
- Mota J, Casimiro S, Fernandes J, et al. (2022) Lupin Protein Concentrate as a Novel Functional Food Additive That Can Reduce Colitis-Induced Inflammation and Oxidative Stress. Nutrients 14:2102. [CrossRef]
- Kamran F, Phillips M, Harman DG, Reddy N (2023) Antioxidant activities of lupin (Lupinus angustifolius) protein hydrolysates and their potential for nutraceutical and functional foods. Food Chemistry Advances 2:100297. [CrossRef]
- Yaver E, Bilgiçli N (2021) Ultrasound-treated lupin (Lupinus albus L.) flour: Protein- and fiber-rich ingredient to improve physical and textural quality of bread with a reduced glycemic index. LWT 148:. [CrossRef]
- Ramos-Diaz JM, Oksanen S, Kantanen K, et al. (2023) Characterization of texturized meat analogues containing native lupin flour and lupin protein concentrate/isolate. Heliyon 9:. [CrossRef]
- Ayalew DB, Abera BD, Adiss YL (2024) Effect of roasting temperature and soaking time on the nutritional, antinutrional and sensory properties of protein-based meat analog from lupine. Heliyon 10:e33122. [CrossRef]
- Kazemzadeh Pournaki S, Biswas A, Hall C (2024) Effects of storage conditions on chemistry and technological properties of different cultivars of Chickpea. J Agric Food Res 16:101066. [CrossRef]
- Sehar S, Rabail R, Munir S, et al. (2023) An insight into anticancer perspectives of chickpea bioactive compounds. Food Chemistry Advances 3:100453. [CrossRef]
- Madurapperumage A, Tang L, Thavarajah P, et al. (2021) Chickpea (Cicer arietinum L.) as a Source of Essential Fatty Acids – A Biofortification Approach. Front Plant Sci 12:734980. [CrossRef]
- Begum N, Khan QU, Liu LG, et al. (2023) Nutritional composition, health benefits and bio-active compounds of chickpea (Cicer arietinum L.). Front Nutr 10:1218468. [CrossRef]
- Xing Q, Dekker S, Kyriakopoulou K, et al. (2020) Enhanced nutritional value of chickpea protein concentrate by dry separation and solid state fermentation. Innovative Food Science & Emerging Technologies 59:102269. [CrossRef]
- Wetterauw K, Spitzer M, Nafingah R, et al. (2024) Dry fractionation of chickpea flour: Impact of de-oiling and flow aids. Powder Technol 446:120180. [CrossRef]
- Liu R, Flanagan BM, Ratanpaul V, Gidley MJ (2025) Valorising legume protein extraction side-streams: Isolation and characterisation of fibre-rich and starch-rich co-products from wet fractionation of five legumes. Food Hydrocoll 164:111191. [CrossRef]
- Yang JS, Dias FFG, Pham TTK, et al. (2024) A sequential fractionation approach to understanding the physicochemical and functional properties of aqueous and enzyme-assisted aqueous extracted black bean proteins. Food Hydrocoll 146:109250. [CrossRef]
- Ma Y, Zhang J, He J, et al. (2023) Effects of high-pressure homogenization on the physicochemical, foaming, and emulsifying properties of chickpea protein. Food Research International 170:112986. [CrossRef]
- Kong Y, Toh NP, Wu Y, Huang D (2023) Trypsin-treated chickpea protein hydrolysate enhances the cytoaffinity of microbeads for cultured meat application. Food Research International 173:113299. [CrossRef]
- Czapalay ES, Dobson S, Marangoni AG (2025) Legume Starch and Flour-Based Emulsion Gels as Adipose Tissue Mimetics in Plant-Based Meat Products. Future Foods 100578. [CrossRef]
- Lee EJ, Hong GP (2023) Effect of the double heating cycle on the thermal gelling properties of vicilin fractions from soy, mung bean, red bean and their mixture with soy glycinin. Food Hydrocoll 137:108370. [CrossRef]
- Kornet R, Yang J, Venema P, et al. (2022) Optimizing pea protein fractionation to yield protein fractions with a high foaming and emulsifying capacity. Food Hydrocoll 126:107456. [CrossRef]
- Seetapan N, Raksa P, Limparyoon N, et al. (2023) High moisture extrusion of meat analogues using mung bean (Vigna radiata L.) protein and flour blends: investigations on morphology, texture and rheology. Int J Food Sci Technol 58:1922–1930. [CrossRef]
- Boukid F, Castellari M (2022) How can processing technologies boost the application of faba bean (Vicia faba L.) proteins in food production? eFood 3:e18. [CrossRef]
- Arzami AN, de Carvalho DM, Vilaplana F, et al. (2022) Narrow-leafed lupin (Lupinus angustifolius L.): Characterization of emulsification and fibre properties. Future Foods 6:100192. [CrossRef]
- Patil ND, Bains A, Sridhar K, et al. (2024) Extraction, Modification, Biofunctionality, and Food Applications of Chickpea (Cicer arietinum) Protein: An Up-to-Date Review. Foods 13:1398. [CrossRef]
- Wani IA, Andrabi SN, Sogi DS, Hassan I (2020) Comparative study of physicochemical and functional properties of flours from kidney bean (Phaseolus vulgaris L.) and green gram (Vigna radiata L.) cultivars grown in Indian temperate climate. Legume Science 2:e11. [CrossRef]
- Carlin J; Der T;, Wanasundara JPD;, et al. (2023) Generating Multi-Functional Pulse Ingredients for Processed Meat Products—Scientific Evaluation of Infrared-Treated Lentils. Foods 12:1722. [CrossRef]
- Tănase Apetroaei V, Pricop EM, Istrati DI, Vizireanu C (2024) Hemp Seeds (Cannabis sativa L.) as a Valuable Source of Natural Ingredients for Functional Foods—A Review. Molecules 2024, Vol 29, Page 2097 29:2097. [CrossRef]
- Banskota AH, Jones A, Hui JPM, Stefanova R (2022) Triacylglycerols and Other Lipids Profiling of Hemp By-Products. Molecules 27:2339. [CrossRef]
- Chen H, Xu B, Wang Y, et al. (2023) Emerging natural hemp seed proteins and their functions for nutraceutical applications. Food Science and Human Wellness 12:929–941. [CrossRef]
- Santos-Sánchez G, Álvarez-López AI, Ponce-España E, et al. (2022) Hempseed (Cannabis sativa) protein hydrolysates: A valuable source of bioactive peptides with pleiotropic health-promoting effects. Trends Food Sci Technol 127:303–318. [CrossRef]
- Karabulut G, Kahraman O, Pandalaneni K, et al. (2023) A comprehensive review on hempseed protein: Production, functional and nutritional properties, novel modification methods, applications, and limitations. Int J Biol Macromol 253:127240. [CrossRef]
- Cabral EM, Zhu X, Garcia-Vaquero M, et al. (2023) Recovery of Protein from Industrial Hemp Waste (Cannabis sativa, L.) Using High-Pressure Processing and Ultrasound Technologies. Foods 12:2883. [CrossRef]
- Eckhardt L, Bu F, Franczyk A, et al. (2024) Hemp (Cannabis sativa L.) protein: Impact of extraction method and cultivar on structure, function, and nutritional quality. Curr Res Food Sci 8:100746. [CrossRef]
- Zahari I, Purhagen JK, Rayner M, et al. (2023) Extrusion of high-moisture meat analogues from hempseed protein concentrate and oat fibre residue. J Food Eng 354:111567. [CrossRef]
- Nasrollahzadeh F, Roman L, Swaraj VJS, et al. (2022) Hemp (Cannabis sativa L.) protein concentrates from wet and dry industrial fractionation: Molecular properties, nutritional composition, and anisotropic structuring. Food Hydrocoll 131:107755. [CrossRef]
- Tileuberdi N, Turgumbayeva A, Yeskaliyeva B, et al. (2022) Extraction, Isolation of Bioactive Compounds and Therapeutic Potential of Rapeseed (Brassica napus L.). Molecules 27:8824. [CrossRef]
- Peeters K, Tenorio AT (2022) Comparing Analytical Methods for Erucic Acid Determination in Rapeseed Protein Products. Foods 11:815. [CrossRef]
- Stolte N, Vettel J, Möllers C (2022) Genetic variation for seed storage protein composition in rapeseed (Brassica napus) and development of near-infrared reflectance spectroscopy calibration equations. Plant Breeding 141:408–417. [CrossRef]
- Duan X, Dong Y, Zhang M, et al. (2023) Identification and molecular interactions of novel ACE inhibitory peptides from rapeseed protein. Food Chem 422:136085. [CrossRef]
- Shen J, Liu Y, Wang X, et al. (2023) A Comprehensive Review of Health-Benefiting Components in Rapeseed Oil. Nutrients 15:999. [CrossRef]
- Jia W, Curubeto N, Rodríguez-Alonso E, et al. (2021) Rapeseed protein concentrate as a potential ingredient for meat analogues. Innovative Food Science & Emerging Technologies 72:102758. [CrossRef]
- Zhang Y, Shao F, Wan X, et al. (2024) Effects of rapeseed protein addition on soybean protein-based textured protein produced by low-moisture extrusion: Changes in physicochemical attributes, structural properties and barrel flow behaviors. Food Hydrocoll 149:109631. [CrossRef]
- Singh A, Kumar V (2024) Pumpkin seeds as nutraceutical and functional food ingredient for future: A review. Grain & Oil Science and Technology 7:12–29. [CrossRef]
- Aziz A, Noreen S, Khalid W, et al. (2023) Pumpkin and Pumpkin Byproducts: Phytochemical Constitutes, Food Application and Health Benefits. ACS Omega 8:23346. [CrossRef]
- Zerafatjou N, Amirzargar M, Biglarkhani M, et al. (2021) Pumpkin seed oil (Cucurbita pepo) versus tamsulosin for benign prostatic hyperplasia symptom relief: a single-blind randomized clinical trial. BMC Urol 21:. [CrossRef]
- Singh A, Kumar V (2022) Nutritional, phytochemical, and antimicrobial attributes of seeds and kernels of different pumpkin cultivars. Food Front 3:182–193. [CrossRef]
- Mehvish Habib, Sakshi Singh, Alok Sagar N, et al. (2025) Physicochemical and functional characterization of pumpkin seed protein isolate. Sustainable Food Technology. [CrossRef]
- Choi HW, Hahn J, Kim HS, Choi YJ (2025) Thermorheological properties and structural characteristics of soy and pumpkin seed protein blends for high-moisture meat analogs. Food Chem 464:141768. [CrossRef]
- Kong Y, Lin S, Chen S, et al. (2025) Pumpkin seed protein-based hydrogel as gelatin mimics and edible inks in 3D-Printed food. Food Hydrocoll 162:111014. [CrossRef]
- Kumar P, Sharma N, Ahmed MA, et al. (2022) Technological interventions in improving the functionality of proteins during processing of meat analogs. Front Nutr 9:1044024. [CrossRef]
- El-Sohaimy SA, Androsova NV, Toshev AD, El Enshasy HA (2022) Nutritional Quality, Chemical, and Functional Characteristics of Hemp (Cannabis sativa ssp. sativa) Protein Isolate. Plants 2022, Vol 11, Page 2825 11:2825. [CrossRef]
- Silventoinen P, Kortekangas A, Nordlund E, Sozer N (2022) Impact of Phytase Treatment and Calcium Addition on Gelation of a Protein-Enriched Rapeseed Fraction. Food Bioproc Tech 15:1422–1435. [CrossRef]
- Gadekar YP, Jairath G, Soni A, et al. (2025) Utilisation of pumpkin seed (Cucurbita maxima) as a meat matrix preservative: Influence on colour and lipid stabilities. Meat Sci 223:. [CrossRef]
- Kowalczewski L, Ró MB, Zá Nska ˙, et al. (2022) Flaxseed Bioactive Compounds: Chemical Composition, Functional Properties, Food Applications and Health Benefits-Related Gut Microbes. Foods 11:3307. [CrossRef]
- Motyka S, Skała E, Ekiert H, Szopa A (2023) Health-promoting approaches of the use of chia seeds. J Funct Foods 103:105480. [CrossRef]
- Huang X, Hu Y, Li Z, et al. (2025) Dephenolization Methods, Quality Characteristics, Applications, and Advancements of Dephenolized Cottonseed Protein: Review. Foods 2025, Vol 14, Page 628 14:628. [CrossRef]
- Wei P, Zhao F, Wang Z, et al. (2022) Sesame (Sesamum indicum L.): A Comprehensive Review of Nutritional Value, Phytochemical Composition, Health Benefits, Development of Food, and Industrial Applications. Nutrients 14:4079. [CrossRef]
- Song C, Lin Y, Hong P, et al. (2022) Low-Content Pre-Emulsified Safflower Seed Oil Enhances the Quality and Flavor of the Nemipterus Virgatus Surimi Gel. Gels 8:106. [CrossRef]
- Tantawy AA, Ali M, Kurbonova M, et al. (2025) Functional properties of sunflower protein concentrates extracted using different anti-greening agents- low-fat whipping cream preparation. LWT 218:117456. [CrossRef]
- Albakry Z, Karrar E, Mohamed Ahmed IA, et al. (2022) Nutritional Composition and Volatile Compounds of Black Cumin (Nigella sativa L.) Seed, Fatty Acid Composition and Tocopherols, Polyphenols, and Antioxidant Activity of Its Essential Oil. Horticulturae 8:575. [CrossRef]
- Wang J, Sun X, Xu X, et al. (2022) Wheat Flour-Based Edible Films: Effect of Gluten on the Rheological Properties, Structure, and Film Characteristics. Int J Mol Sci 23:11668. [CrossRef]
- Zhang X, Zhao Y, Zhang T, et al. (2023) Potential of hydrolyzed wheat protein in soy-based meat analogues: Rheological, textural and functional properties. Food Chem X 20:100921. [CrossRef]
- Wieser H, Koehler P, Scherf KA (2020) The Two Faces of Wheat. Front Nutr 7:517313. [CrossRef]
- Siddiqi RA, Singh TP, Rani M, et al. (2020) Diversity in Grain, Flour, Amino Acid Composition, Protein Profiling, and Proportion of Total Flour Proteins of Different Wheat Cultivars of North India. Front Nutr 7:141. [CrossRef]
- Zhang R, Yang Y, Liu Q, et al. (2023) Effect of Wheat Gluten and Peanut Protein Ratio on the Moisture Distribution and Textural Quality of High-Moisture Extruded Meat Analogs from an Extruder Response Perspective. Foods 2023, Vol 12, Page 1696 12:1696. [CrossRef]
- Sun Y, Dong M, Bai J, et al. (2024) Preparation and properties of high-soluble wheat gluten protein-based meat analogues. J Sci Food Agric 104:42–50. [CrossRef]
- Dai H, An H (2024) Study on the structural characteristics of wheat gluten enzymatic hydrolysates and their effect on the texturization degree of high-moisture plant-protein extrudates. J Cereal Sci 119:103974. [CrossRef]
- Flavourzyme® | Novozymes. https://www.novozymes.com/en/products/animal-protein/flavourzyme. Accessed 2 May 2025.
- Sun A, Chen L, Wu W, et al. (2023) The potential meat flavoring generated from Maillard reaction products of wheat gluten protein hydrolysates-xylose: Impacts of different thermal treatment temperatures on flavor. Food Research International 165:112512. [CrossRef]
- Wouters AGB, Nicolai T (2024) Self-assembly of oat proteins into various colloidal states as function of the NaCl concentration and pH. Food Hydrocoll 149:109603. [CrossRef]
- Rafique H, Dong R, Wang X, et al. (2022) Dietary-Nutraceutical Properties of Oat Protein and Peptides. Front Nutr 9:950400. [CrossRef]
- Kumar L, Sehrawat R, Kong Y (2021) Oat proteins: A perspective on functional properties. LWT 152:112307. [CrossRef]
- Zhou J, Li T, Peydayesh M, et al. (2022) Oat Plant Amyloids for Sustainable Functional Materials. Advanced Science 9:2104445. [CrossRef]
- Zhao J, Ni D, Bhandari B, et al. (2024) Impacts of oat flour fortification on the rheological, microstructural, digestibility, and sensory characteristics of low-fat, high-protein almond-based gels. Food Hydrocoll 151:109849. [CrossRef]
- Sargautis D, Kince T (2023) Effect of Enzymatic Pre-Treatment on Oat Flakes Protein Recovery and Properties. Foods 12:965. [CrossRef]
- Brückner-Gühmann M, Kratzsch A, Sozer N, Drusch S (2021) Oat protein as plant-derived gelling agent: Properties and potential of modification. Future Foods 4:100053. [CrossRef]
- Senarathna S, Mel R, Malalgoda M (2024) Utilization of cereal-based protein ingredients in food applications. J Cereal Sci 116:103867. [CrossRef]
- Wang D, Li T, Zhang X, et al. (2025) Modification of rice protein and its components: Enhanced fibrils formation and improved foaming properties. Food Hydrocoll 158:110575. [CrossRef]
- Rivero Meza SL, Cañizares L, Dannenberg B, et al. (2024) Sustainable rice bran protein: Composition, extraction, quality properties and applications. Trends Food Sci Technol 145:104355. [CrossRef]
- Li S, Jiang Z, Wang F, et al. (2020) Characterization of rice glutelin fibrils and their effect on in vitro rice starch digestibility. Food Hydrocoll 106:105918. [CrossRef]
- Lee JS, Choi I, Han J (2022) Construction of rice protein-based meat analogues by extruding process: Effect of substitution of soy protein with rice protein on dynamic energy, appearance, physicochemical, and textural properties of meat analogues. Food Research International 161:111840. [CrossRef]
- Lee JS, Oh H, Choi I, et al. (2022) Physico-chemical characteristics of rice protein-based novel textured vegetable proteins as meat analogues produced by low-moisture extrusion cooking technology. LWT 157:113056. [CrossRef]
- Charlie EA, Angrainy H, Kantrong H (2025) Exploring the use of rice bran and mung bean as soy substitutes in low-moisture extruded plant-based meat. Innovative Food Science & Emerging Technologies 100:103916. [CrossRef]
- Qu M, Jiang P, Zhu Y, et al. (2024) Effects of glutenin/gliadin ratio and calcium ion on the structure and gelatinity of wheat gluten protein under heat induction. Food Biosci 58:103704. [CrossRef]
- Boukid F (2021) Oat proteins as emerging ingredients for food formulation: where we stand? European Food Research and Technology 247:535–544. [CrossRef]
- Wu W, Li F, Wu X (2021) Effects of rice bran rancidity on oxidation, structural characteristics and interfacial properties of rice bran globulin. Food Hydrocoll 110:106123. [CrossRef]
- Li XL, Meng R, Xu BC, et al. (2022) Function emulsion gels prepared with carrageenan and zein/carboxymethyl dextrin stabilized emulsion as a new fat replacer in sausages. Food Chem 389:. [CrossRef]
- Dangi N, Yadav BS, Yadav RB (2020) Barley β-glucan concentrate and its acid hydrolysate for the modification of dough making and rheological properties of water chestnut flour. Int J Biol Macromol 164:253–264. [CrossRef]
- Abdollahzadeh A, Vazifedoost M, Didar Z, et al. (2024) Comparison of the effect of hydroxyl propyl methyl cellulose, pectin, and concentrated raisin juice on gluten-free bread based on rice and foxtail millet flour. Food Sci Nutr 12:439–449. [CrossRef]
- Bulgaru V, Netreba N, Ghendov-Mosanu A (2025) Pre-Treatment of Vegetable Raw Materials (Sorghum Oryzoidum) for Use in Meat Analog Manufacture. Applied Sciences 2025, Vol 15, Page 349 15:349. [CrossRef]
- Park J-H, Lee Y-J, Lim J-G, et al. (2021) Effect of Quinoa (Chenopodium quinoa Willd.) Starch and Seeds on the Physicochemical and Textural and Sensory Properties of Chicken Meatballs during Frozen Storage. Foods 2021, Vol 10, Page 1601 10:1601. [CrossRef]
- Zhu F (2023) Amaranth proteins and peptides: Biological properties and food uses. Food Research International 164:112405. [CrossRef]
- Sofi SA, Ahmed N, Farooq A, et al. (2022) Nutritional and bioactive characteristics of buckwheat, and its potential for developing gluten-free products: An updated overview. Food Sci Nutr 11:2256. [CrossRef]
- Bartkiene E, Zarovaite P, Starkute V, et al. (2023) Changes in Lacto-Fermented Agaricus bisporus (White and Brown Varieties) Mushroom Characteristics, including Biogenic Amine and Volatile Compound Formation. Foods 12:2441. [CrossRef]
- Keerthana K, Anukiruthika T, Moses JA, Anandharamakrishnan C (2020) Development of fiber-enriched 3D printed snacks from alternative foods: A study on button mushroom. J Food Eng 287:110116. [CrossRef]
- He M, Condict L, Richardson SJ, et al. (2024) Molecular characterization of interactions between lectin - a protein from common edible mushroom (Agaricus bisporus) - and dietary carbohydrates. Food Hydrocoll 146:109253. [CrossRef]
- Bell V, Silva CRPG, Guina J, Fernandes TH (2022) Mushrooms as future generation healthy foods. Front Nutr 9:1050099. [CrossRef]
- Kim H;, Jeon Y-E;, Kim S-M;, et al. (2023) Agaricus bisporus Extract Exerts an Anti-Obesity Effect in High-Fat Diet-Induced Obese C57BL/6N Mice by Inhibiting Pancreatic Lipase-Mediated Fat Absorption. Nutrients 15:4225. [CrossRef]
- Du X, Sissons J, Shanks M, Plotto A (2021) Aroma and flavor profile of raw and roasted Agaricus bisporus mushrooms using a panel trained with aroma chemicals. LWT 138:110596. [CrossRef]
- Fu Q, Yang J, Lv L, et al. (2023) Effects of replacing chicken breast meat with Agaricus bisporus mushrooms on the qualities of emulsion-type sausages. LWT 184:114983. [CrossRef]
- Törős G, El-Ramady H, Prokisch J (2022) Edible Mushroom of Pleurotus spp.:A Case Study of Oyster Mushroom (Pleurotus ostreatus L.). Environment, Biodiversity and Soil Security 6:51–59. [CrossRef]
- Ghafoor A, Niazi AR (2024) Pleurotus spp: an ultimate solution to the emerging calamities of the world. N Z J Bot. [CrossRef]
- Fulgoni VL, Agarwal S (2021) Nutritional impact of adding a serving of mushrooms on usual intakes and nutrient adequacy using National Health and Nutrition Examination Survey 2011–2016 data. Food Sci Nutr 9:1504. [CrossRef]
- Bouranis L, Aguchem RN, Chibuogwu CC, et al. (2021) Nutrient and Antinutrient Composition of Pleurotus ostreatus Grown on Different Substrates. Biology and Life Sciences Forum 2022, Vol 11, Page 69 11:69. [CrossRef]
- Mazumder MAR, Sujintonniti N, Chaum P, et al. (2023) Developments of Plant-Based Emulsion-Type Sausage by Using Grey Oyster Mushrooms and Chickpeas. Foods 2023, Vol 12, Page 1564 12:1564. [CrossRef]
- Vargas-Sánchez RD;, Rodríguez-Carpena M;, Fernández-López JG;, et al. (2022) Pleurotus Genus as a Potential Ingredient for Meat Products. Foods 11:779. [CrossRef]
- Demircan E, Aydar EF, Mertdinc (Mertdinç) Z, et al. (2023) 3D printable vegan plant-based meat analogue: Fortification with three different mushrooms, investigation of printability, and characterization. Food Research International 173:113259. [CrossRef]
- Zhang Z, Zang M, Chen J, et al. (2024) Effect of the mycelium of oyster mushrooms on the physical and flavor properties of a plant-based beef analogue. LWT 198:116029. [CrossRef]
- Hajdú P, Abdalla Z, El-Ramady H, Prokisch J (2022) Edible Mushroom of Lentinula spp.: A Case Study of Shiitake (Lentinula edodes L.) Cultivation. Environment, Biodiversity and Soil Security 6:41–49. [CrossRef]
- Kaya M, Cam M (2022) Eritadenine: Pressurized liquid extraction from Lentinula edodes and thermal degradation kinetics. Sustain Chem Pharm 29:100809. [CrossRef]
- Yu CX, Zhang YR, Ren YF, et al. (2023) Composition and contents of fatty acids and amino acids in the mycelia of Lentinula edodes. Food Sci Nutr 11:4038–4046. [CrossRef]
- Santhapur R, Jayakumar D, McClements DJ (2024) Development and Characterization of Hybrid Meat Analogs from Whey Protein-Mushroom Composite Hydrogels. Gels 2024, Vol 10, Page 446 10:446. [CrossRef]
- Choi H, Gwon HG, Kim D, et al. (2025) Structural characterization and rheological analysis of Lentinus edodes mycelium (mycoprotein)–hydrocolloid composites for food formulation. Future Foods 11:100568. [CrossRef]
- Dimopoulou M, Kolonas A, Mourtakos S, et al. (2022) Nutritional Composition and Biological Properties of Sixteen Edible Mushroom Species. Applied Sciences 2022, Vol 12, Page 8074 12:8074. [CrossRef]
- Stilinović N, Čapo I, Vukmirović S, et al. (2020) Chemical composition, nutritional profile and in vivo antioxidant properties of the cultivated mushroom Coprinus comatus. R Soc Open Sci 7:. [CrossRef]
- Ratnaningtyas NI, Hernayanti, Ekowati N, et al. (2022) Antioxidant Activities and Properties of Coprinus comatus Mushroom Both Mycelium and Fruiting Body Extracts In Streptozotocin-Induced Hyperglycemic Rats Model. Biosaintifika 14:9–21. [CrossRef]
- Zawadzka A, Kobus-Cisowska J, Szwajgier D, et al. (2022) Dual functional cholinesterase inhibitors and complexing of aluminum ions of five species of fungi family depended of drying conditions and extraction process - In vitro study. LWT 154:112712. [CrossRef]
- Yang H, Zheng Z, Zhou H, et al. (2022) Proteomics Reveals the Mechanism Underlying the Autolysis of Postharvest Coprinus comatus Fruiting Bodies. J Agric Food Chem 70:1346–1357. [CrossRef]
- Ren S, Zheng E, Zhao T, et al. (2022) Evaluation of physicochemical properties, equivalent umami concentration and antioxidant activity of Coprinus comatus prepared by different drying methods. LWT 162:113479. [CrossRef]
- Hollweg G, Trindade PCO, dos Santos BA, et al. (2024) Development of Plant-Based Burgers with Partial Replacement of Texturized Soy Protein by Agaricus bisporus: Effects on Physicochemical and Sensory Properties. Foods 13:. [CrossRef]
- Tokarczyk G, Felisiak K, Adamska I, et al. (2023) Effect of Oyster Mushroom Addition on Improving the Sensory Properties, Nutritional Value and Increasing the Antioxidant Potential of Carp Meat Burgers. Molecules 28:6975. [CrossRef]
- Rangel-Vargas E, Rodriguez JA, Domínguez R, et al. (2021) Edible Mushrooms as a Natural Source of Food Ingredient/Additive Replacer. Foods 10:2687. [CrossRef]
- Qu H, Dong Y, Liu C, et al. (2023) Effects of Coprinus comatus (chicken drumstick mushroom) on the quality characteristics of low nitrite-backfat Cantonese sausages. LWT 187:115339. [CrossRef]
- Liu K, Li J, Xing C, et al. (2023) Characterization of Auxenochlorella protothecoides acyltransferases and potential of their protein interactions to promote the enrichment of oleic acid. Biotechnology for Biofuels and Bioproducts 16:1–16. [CrossRef]
- Korozi E, Tsagou V, Kefalogianni I, et al. (2022) Continuous Culture of Auxenochlorella protothecoides on Biodiesel Derived Glycerol under Mixotrophic and Heterotrophic Conditions: Growth Parameters and Biochemical Composition. Microorganisms 10:541. [CrossRef]
- Mavrommatis A, Tsiplakou E, Zerva A, et al. (2023) Microalgae as a Sustainable Source of Antioxidants in Animal Nutrition, Health and Livestock Development. Antioxidants 2023, Vol 12, Page 1882 12:1882. [CrossRef]
- Maltsev Y, Maltseva K, Kulikovskiy M, Maltseva S (2021) Influence of Light Conditions on Microalgae Growth and Content of Lipids, Carotenoids, and Fatty Acid Composition. Biology 2021, Vol 10, Page 1060 10:1060. [CrossRef]
- Sägesser C, Kallfelz JM, Boulos S, et al. (2024) Structurability of microalgae, soy and pea protein for extruded high-moisture meat analogues. Food Hydrocoll 156:110290. [CrossRef]
- Mtaki K, Kyewalyanga MS, Mtolera MSP (2021) Supplementing wastewater with NPK fertilizer as a cheap source of nutrients in cultivating live food (Chlorella vulgaris). Ann Microbiol 71:1–13. [CrossRef]
- Abdur Razzak S, Bahar K, Islam KMO, et al. (2024) Microalgae cultivation in photobioreactors: sustainable solutions for a greener future. Green Chemical Engineering 5:418–439. [CrossRef]
- De Gol C, Snel S, Rodriguez Y, Beyrer M (2023) Gelling capacity of cell-disrupted Chlorella vulgaris and its texture effect in extruded meat substitutes. Food Structure 37:100332. [CrossRef]
- Farag MR, Alagawany M, Mahdy EAA, et al. (2023) Benefits of Chlorella vulgaris against Cadmium Chloride-Induced Hepatic and Renal Toxicities via Restoring the Cellular Redox Homeostasis and Modulating Nrf2 and NF-KB Pathways in Male Rats. Biomedicines 11:2414. [CrossRef]
- Expósito N, Carafa R, Kumar V, et al. (2021) Performance of chlorella vulgaris exposed to heavy metal mixtures: Linking measured endpoints and mechanisms. Int J Environ Res Public Health 18:1–19. [CrossRef]
- De Gol C, Snel S, Rodriguez Y, Beyrer M (2023) Gelling capacity of cell-disrupted Chlorella vulgaris and its texture effect in extruded meat substitutes. Food Structure 37:100332. [CrossRef]
- Bakhsh A, Park J, Baritugo KA, et al. (2023) A holistic approach toward development of plant-based meat alternatives through incorporation of novel microalgae-based ingredients. Front Nutr 10:1110613. [CrossRef]
- Gentscheva G, Nikolova K, Panayotova V, et al. (2023) Application of Arthrospira platensis for Medicinal Purposes and the Food Industry: A Review of the Literature. Life 13:845. [CrossRef]
- Balestri F, Podgórska-Kryszczuk I (2024) Spirulina—An Invaluable Source of Macro- and Micronutrients with Broad Biological Activity and Application Potential. Molecules 29:5387. [CrossRef]
- Mohammed IA, Ruengjitchatchawalya M, Paithoonrangsarid K (2023) Cultivation manipulating zeaxanthin-carotenoid production in Arthrospira (Spirulina) platensis under light and temperature stress. Algal Res 76:103315. [CrossRef]
- Singh U, Gandhi HA, Nikita, et al. (2023) Cyanometabolites: molecules with immense antiviral potential. Arch Microbiol 205:164. [CrossRef]
- Uzlaşır T, Selli S, Kelebek | Hasim (2024) Spirulina platensis and Phaeodactylum tricornutum as sustainable sources of bioactive compounds: Health implications and applications in the food industry. Future Postharvest and Food 1:34–46. [CrossRef]
- Guo J, Huang Y, Gu X, Meng Z (2025) Spirulina platensis protein-based emulsion gel as fat substitute in meat analogs: Evaluation performance across post-processing. Food Chem 463:141414. [CrossRef]
- Afdhaliah N, Ningrum A, Setiowati AD (2024) Red Palm Oil Gelled Emulsion Stabilized by Spirulina (Arthrospira platensis) Protein and Carrageenan as Fat Replacer in Beef Patty. Food Bioproc Tech 1–15. [CrossRef]
- Qu Y, Chen X, Ma B, et al. (2022) Extracellular Metabolites of Heterotrophic Auxenochlorella protothecoides: A New Source of Bio-Stimulants for Higher Plants. Mar Drugs 20:569. [CrossRef]
- Abdel-Moatamed BR, El-Fakhrany AEMA, Elneairy NAA, et al. (2024) The Impact of Chlorella vulgaris Fortification on the Nutritional Composition and Quality Characteristics of Beef Burgers. Foods 13:1945. [CrossRef]
- Sahil S, Bodh S, Verma P (2024) Spirulina platensis: A comprehensive review of its nutritional value, antioxidant activity and functional food potential. J Cell Biotechnol 1–14. [CrossRef]
- Yang H, Song C, Liu C, Wang P (2024) Synthetic Biology Tools for Engineering Aspergillus oryzae. Journal of Fungi 10:34. [CrossRef]
- Sun Z, Wu Y, Long S, et al. (2024) Aspergillus oryzae as a Cell Factory: Research and Applications in Industrial Production. Journal of Fungi 10:248. [CrossRef]
- Seidler Y, Rimbach G, Lüersen K, et al. (2024) The postbiotic potential of Aspergillus oryzae – a narrative review. Front Microbiol 15:1452725. [CrossRef]
- Yamashita H (2021) Koji starter and koji world in Japan. Journal of Fungi 7:. [CrossRef]
- Murai T, Annor GA (2025) Improving the Nutritional Profile of Intermediate Wheatgrass by Solid-State Fermentation with Aspergillus oryzae Strains. Foods 2025, Vol 14, Page 395 14:395. [CrossRef]
- Brandão M, Marques DJ, Sousa S, et al. (2025) Lactic Acid Bacteria and Yeast Fermentation to Improve the Nutritional Value of Ulva rigida. Marine Drugs 2025, Vol 23, Page 106 23:106. [CrossRef]
- Rousta N, Hellwig C, Wainaina S, et al. (2021) Filamentous Fungus Aspergillus oryzae for Food: From Submerged Cultivation to Fungal Burgers and Their Sensory Evaluation—A Pilot Study. Foods 2021, Vol 10, Page 2774 10:2774. [CrossRef]
- Maini Rekdal V, Villalobos-Escobedo JM, Rodriguez-Valeron N, et al. (2024) Neurospora intermedia from a traditional fermented food enables waste-to-food conversion. Nat Microbiol 9:2666. [CrossRef]
- Amara AA, El-Baky NA (2023) Fungi as a Source of Edible Proteins and Animal Feed. Journal of Fungi 2023, Vol 9, Page 73 9:73. [CrossRef]
- Rekdal VM, Villalobos-Escobedo JM, Rodriguez-Valeron N, et al. (2024) Multi-omics analysis of a traditional fermented food reveals a byproduct-associated subpopulation of Neurospora intermedia for waste-to-food upcycling. bioRxiv 3:2024.07.24.604980. [CrossRef]
- Toghiani J, Fallah N, Nasernejad B, et al. (2025) Production of protein-rich fungal biomass from pistachio dehulling waste using edible Neurospora intermedia. Sci Rep 15:5873. [CrossRef]
- Wang B, Lu H, Lou H, et al. (2024) Synthesis and characterization of Neurospora intermedia-based composite mycoprotein gel meat: Insight into the effect of pH and soluble starch on water-holding capacity and texture properties. Food Hydrocoll 155:110190. [CrossRef]
- Li K, Qiao K, Xiong J, et al. (2023) Nutritional Values and Bio-Functional Properties of Fungal Proteins: Applications in Foods as a Sustainable Source. Foods 12:4388. [CrossRef]
- Zhu J, Chen W, Chen Y, et al. (2025) CFD optimization of an air lift fermenter for Fusarium venenatum fermentation. Bioresour Technol Rep 29:102024. [CrossRef]
- Majumder R, Miatur S, Saha A, Hossain S (2024) Mycoprotein: production and nutritional aspects: a review. Sustainable Food Technology 2:81–91. [CrossRef]
- Tong S, Chen W, Hong R, et al. (2024) Efficient Mycoprotein Production with Low CO2 Emissions through Metabolic Engineering and Fermentation Optimization of Fusarium venenatum. J Agric Food Chem 72:604–612. [CrossRef]
- Akinsemolu AA, Onyeaka HN (2025) Mycoproteins as sustainable food sources: current applications and future prospects. Discover Applied Sciences 7:1–11. [CrossRef]
- D’Almeida AP, de Albuquerque TL (2025) Is It Possible to Produce Meat Without Animals? The Potential of Microorganisms as Protein Sources. Fermentation 2025, Vol 11, Page 24 11:24. [CrossRef]
- Li XL, Qi XN, Deng JC, et al. (2025) Characterization of Fusarium venenatum Mycoprotein-Based Harbin Red Sausages. Foods 14:556. [CrossRef]
- Devanthi PVP, Pratama F, Pramanda IT, et al. (2024) Exploring the Potential of Aspergillus oryzae for Sustainable Mycoprotein Production Using Okara and Soy Whey as Cost-Effective Substrates. Journal of Fungi 10:. [CrossRef]
- Elhalis H, See XY, Osen R, et al. (2023) The potentials and challenges of using fermentation to improve the sensory quality of plant-based meat analogs. Front Microbiol 14:. [CrossRef]
- Jeon H, Shin S, Winarto J, et al. (2024) Genetic approach to discover a valuable gene for enhanced nutritional value in the edible filamentous fungus Fusarium venenatum. Food Front 5:2556–2565. [CrossRef]
- Phumsombat P, Trisakwattana K, Ittithanaput N, et al. (2024) Synbiotic and protein-enriched low-fat Sao Hai rice ice cream. Quality Assurance and Safety of Crops and Foods 16:14–27. [CrossRef]
- Soybeans, mature seeds, raw - USDA FoodData Central Food Details. https://fdc.nal.usda.gov/food-details/174270/nutrients. Accessed 9 Mar 2025.
- Peas, green, raw - USDA FoodData Central Food Details. https://fdc.nal.usda.gov/food-details/170419/nutrients. Accessed 9 Mar 2025.
- Mung beans, mature seeds, raw - USDA FoodData Central Food Details. https://fdc.nal.usda.gov/food-details/174256/nutrients. Accessed 9 Mar 2025.
- Broadbeans (fava beans), mature seeds, cooked, boiled, without salt - USDA FoodData Central Food Details. https://fdc.nal.usda.gov/food-details/173753/nutrients. Accessed 9 Mar 2025.
- Lupins, mature seeds, raw - USDA FoodData Central Food Details. https://fdc.nal.usda.gov/food-details/172423/nutrients. Accessed 9 Mar 2025.
- Peanuts, all types, raw - USDA FoodData Central Food Details. https://fdc.nal.usda.gov/food-details/172430/nutrients. Accessed 9 Mar 2025.
- Oats, raw - USDA FoodData Central Food Details. https://fdc.nal.usda.gov/food-details/1101825/nutrients. Accessed 9 Mar 2025.
- Krishnamoorthi R, Srinivash M, Mahalingam PU, Malaikozhundan B (2022) Dietary nutrients in edible mushroom, Agaricus bisporus and their radical scavenging, antibacterial, and antifungal effects. Process Biochemistry 121:10–17. [CrossRef]
- Effiong ME, Umeokwochi CP, Afolabi IS, Chinedu SN (2023) Assessing the nutritional quality of Pleurotus ostreatus (oyster mushroom). Front Nutr 10:1279208. [CrossRef]
- Canelli G, Tarnutzer C, Carpine R, et al. (2020) Biochemical and Nutritional Evaluation of Chlorella and Auxenochlorella Biomasses Relevant for Food Application. Front Nutr 7:565996. [CrossRef]
- Zhang C, Wu X, Chen J, Zhou J (2024) Novel fungal alternative proteins from Penicillium limosum for enhancing structural and functional properties of plant-based meat analogues. Food Chem 444:138627. [CrossRef]
- Pobiega K, S˛ Ekul J, Pakulska A, et al. (2024) Fungal Proteins: Sources, Production and Purification Methods, Industrial Applications, and Future Perspectives. Applied Sciences 2024, Vol 14, Page 6259 14:6259. [CrossRef]
- Alshammari NA, Taylor MA, Stevenson R, et al. (2021) Effect of Intake of Food Hydrocolloids of Bacterial Origin on the Glycemic Response in Humans: Systematic Review and Narrative Synthesis. Nutrients 13:2407. [CrossRef]
- Vila-Clarà G, Vila-Martí A, Vergés-Canet L, Torres-Moreno M (2024) Exploring the Role and Functionality of Ingredients in Plant-Based Meat Analogue Burgers: A Comprehensive Review. Foods 13:1258. [CrossRef]
- Pirsa S, Hafezi K (2023) Hydrocolloids: Structure, preparation method, and application in food industry. Food Chem 399:133967. [CrossRef]
- Bakhsh A, Lee S-J, Lee E-Y, et al. (2021) A Novel Approach for Tuning the Physicochemical, Textural, and Sensory Characteristics of Plant-Based Meat Analogs with Different Levels of Methylcellulose Concentration. Foods 10:560. [CrossRef]
- Battacchi D, Verkerk R, Pellegrini N, et al. (2020) The state of the art of food ingredients’ naturalness evaluation: A review of proposed approaches and their relation with consumer trends. Trends Food Sci Technol 106:434–444. [CrossRef]
- Taghian Dinani S, Zhang Y, Vardhanabhuti B, Jan van der Goot A (2023) Enhancing textural properties in plant-based meat alternatives: The impact of hydrocolloids and salts on soy protein-based products. Curr Res Food Sci 7:. [CrossRef]
- Taghian Dinani S, Broekema NL, Boom R, van der Goot AJ (2023) Investigation potential of hydrocolloids in meat analogue preparation. Food Hydrocoll 135:108199. [CrossRef]
- Tan H, Nie S (2021) Functional hydrocolloids, gut microbiota and health: picking food additives for personalized nutrition. FEMS Microbiol Rev 45:. [CrossRef]
- Aga MB, Dar AH, Nayik GA, et al. (2021) Recent insights into carrageenan-based bio-nanocomposite polymers in food applications: A review. Int J Biol Macromol 192:197–209. [CrossRef]
- Liu F, Hou P, Zhang H, et al. (2021) Food-grade carrageenans and their implications in health and disease. Compr Rev Food Sci Food Saf 20:3918–3936. [CrossRef]
- Sha L, Xiong YL (2020) Plant protein-based alternatives of reconstructed meat: Science, technology, and challenges. Trends Food Sci Technol 102:51–61. [CrossRef]
- Cheng C, Chen S, Su J, et al. (2022) Recent advances in carrageenan-based films for food packaging applications. Front Nutr 9:. [CrossRef]
- Tahmouzi S, Meftahizadeh H, Eyshi S, et al. (2023) Application of guar (Cyamopsis tetragonoloba L.) gum in food technologies: A review of properties and mechanisms of action. Food Sci Nutr 11:4869–4897. [CrossRef]
- Yu J, Li D, Wang L, Wang Y (2023) Improving freeze-thaw stability and 3D printing performance of soy protein isolate emulsion gel inks by guar & xanthan gums. Food Hydrocoll 136:108293. [CrossRef]
- Salehi F (2020) Effect of coatings made by new hydrocolloids on the oil uptake during deep-fat frying: A review. J Food Process Preserv 44:. [CrossRef]
- Eghbaljoo H, Sani IK, Sani MA, et al. (2022) Advances in plant gum polysaccharides; Sources, techno-functional properties, and applications in the food industry - A review. Int J Biol Macromol 222:2327–2340. [CrossRef]
- Einhorn-Stoll U, Archut A, Eichhorn M, Kastner H (2021) Pectin - Plant protein systems and their application. Food Hydrocoll 118:106783. [CrossRef]
- Abka-khajouei R, Tounsi L, Shahabi N, et al. (2022) Structures, Properties and Applications of Alginates. Mar Drugs 20:364. [CrossRef]
- Gheorghita Puscaselu R, Lobiuc A, Dimian M, Covasa M (2020) Alginate: From Food Industry to Biomedical Applications and Management of Metabolic Disorders. Polymers (Basel) 12:2417. [CrossRef]
- Kyriakopoulou K, Keppler JK, van der Goot AJ (2021) Functionality of Ingredients and Additives in Plant-Based Meat Analogues. Foods 2021, Vol 10, Page 600 10:600. [CrossRef]
- Bisht B, Lohani UC, Kumar V, et al. (2022) Edible hydrocolloids as sustainable substitute for non-biodegradable materials. Crit Rev Food Sci Nutr 62:693–725. [CrossRef]
- Prasad N, Thombare N, Sharma SC, Kumar S (2022) Gum arabic – A versatile natural gum: A review on production, processing, properties and applications. Ind Crops Prod 187:115304. [CrossRef]
- Khezerlou A, Zolfaghari H, Banihashemi SA, et al. (2021) Plant gums as the functional compounds for edible films and coatings in the food industry: A review. Polym Adv Technol 32:2306–2326. [CrossRef]
- Inguglia ES, Song Z, Kerry JP, et al. (2023) Addressing Clean Label Trends in Commercial Meat Processing: Strategies, Challenges and Insights from Consumer Perspectives. Foods 12:2062. [CrossRef]
- Brassesco ME, Brandão TRS, Silva CLM, Pintado M (2021) Carob bean (Ceratonia siliqua L.): A new perspective for functional food. Trends Food Sci Technol 114:310–322. [CrossRef]
- Han JH, Keum DH, Hong SJ, et al. (2023) Comparative Evaluation of Polysaccharide Binders on the Quality Characteristics of Plant-Based Patties. Foods 2023, Vol 12, Page 3731 12:3731. [CrossRef]
- Udo T, Mummaleti G, Mohan A, et al. (2023) Current and emerging applications of carrageenan in the food industry. Food Research International 173:113369. [CrossRef]
- Chaturvedi S, Kulshrestha S, Bhardwaj K, Jangir R (2021) A Review on Properties and Applications of Xanthan Gum. Microbial Polymers: Applications and Ecological Perspectives 87–107. [CrossRef]
- Tahmouzi S, Meftahizadeh H, Eyshi S, et al. (2023) Application of guar (Cyamopsis tetragonoloba L.) gum in food technologies: A review of properties and mechanisms of action. Food Sci Nutr 11:4869–4897. [CrossRef]
- Gao Y, Luo C, Zhang J, et al. (2022) Konjac glucomannan improves the gel properties of low salt myofibrillar protein through modifying protein conformation. Food Chem 393:133400. [CrossRef]
- Dinani ST, Broekema NL, Boom RM, van der Goot AJ (2023) When and how should low acyl gellan gum be added to the protein blends to improve meat analogue texture? Journal of Food Measurement and Characterization 17:6609–6619. [CrossRef]
- Nasrallah K, Khaled S, El Khatib S, Krayem M (2024) Nutritional, biochemical and health properties of Locust beans and its applications in the food industry: a review. J Food Sci Technol 61:621–630. [CrossRef]
- Williams PA, Phillips GO (2021) Gum arabic. In: Handbook of Hydrocolloids. Elsevier, pp 627–652.
- Lorenc F, Jarošová M, Bedrníček J, et al. (2022) Structural Characterization and Functional Properties of Flaxseed Hydrocolloids and Their Application. Foods 11:2304. [CrossRef]
- Xia Y, Qian J, Zhao Y, et al. (2023) Effects of food components and processing parameters on plant-based meat texture formation and evaluation methods. J Texture Stud 54:394–409. [CrossRef]
- Dobson S, Laredo T, Marangoni AG (2022) Particle filled protein-starch composites as the basis for plant-based meat analogues. Curr Res Food Sci 5:892–903. [CrossRef]
- Bühler JM, Schlangen M, Möller AC, et al. (2022) Starch in Plant-Based Meat Replacers: A New Approach to Using Endogenous Starch from Cereals and Legumes. Starch - Stärke 74:. [CrossRef]
- Rashwan AK, Younis HA, Abdelshafy AM, et al. (2024) Plant starch extraction, modification, and green applications: a review. Environ Chem Lett 22:2483–2530. [CrossRef]
- Choi HW, Kim J-H, Ham SH, et al. (2025) Effect of heating time and drying method on the functional properties of soy protein isolate–maltodextrin conjugates for plant-based meringue cookies. Future Foods 11:100587. [CrossRef]
- Yu X, Li T, Yue M, et al. (2025) Impact of transglutaminase on structural and rheological properties of pea protein-cornmeal-wheat gluten blends for meat analogue production. J Food Eng 390:112412. [CrossRef]
- Taghian Dinani S, Zhang Y, Vardhanabhuti B, Jan van der Goot A (2023) Enhancing textural properties in plant-based meat alternatives: The impact of hydrocolloids and salts on soy protein-based products. Curr Res Food Sci 7:100571. [CrossRef]
- Öztürk-Kerimoğlu B (2021) A promising strategy for designing reduced-fat model meat emulsions by utilization of pea protein-agar agar gel complex. Food Structure 29:100205. [CrossRef]
- Zhang J, Liu P, Wu A, et al. (2024) Towards understanding pectin-protein interaction and the role of pectin in plant-based meat analogs constructing. LWT 202:116325. [CrossRef]
- Wu H, Sakai K, Zhang J, McClements DJ (2024) Plant-based meat analogs: color challenges and coloring agents. Food, Nutrition and Health 1:4. [CrossRef]
- Mariano E, Lee DY, Yun SH, et al. (2024) The Color-Developing Methods for Cultivated Meat and Meat Analogues: A Mini-Review. Food Sci Anim Resour 44:356–371. [CrossRef]
- Bakhsh A, Cho C, Baritugo KA, et al. (2023) Production and Analytical Aspects of Natural Pigments to Enhance Alternative Meat Product Color. Foods 12:1281. [CrossRef]
- Ryu KK, Kang YK, Jeong EW, et al. (2023) Applications of various natural pigments to a plant-based meat analog. LWT 174:114431. [CrossRef]
- Elhalis H, See XY, Osen R, et al. (2023) The potentials and challenges of using fermentation to improve the sensory quality of plant-based meat analogs. Front Microbiol 14:. [CrossRef]
- Nieto G, Martínez-Zamora L, Peñalver R, et al. (2023) Applications of Plant Bioactive Compounds as Replacers of Synthetic Additives in the Food Industry. Foods 13:47. [CrossRef]
- Shi X, Wang Z, Fang Z (2024) Effects of Incorporating Caramel, Carrot, and Tomato Powder on the Quality Characteristics of Soy Protein-Based Meat Patties. Foods 13:2224. [CrossRef]
- Bugoni M, Takiya CS, Grigoletto NathaliaTS, et al. (2022) Dry malt extract from barley partially replacing ground corn in diets of dairy cows: Nutrient digestibility, ruminal fermentation, and milk composition. J Dairy Sci 105:5714–5722. [CrossRef]
- Fernández-López J, Ponce-Martínez AJ, Rodríguez-Párraga J, et al. (2023) Beetroot juices as colorant in plant-based minced meat analogues: Color, betalain composition and antioxidant activity as affected by juice type. Food Biosci 56:103156. [CrossRef]
- Rocchetti G, Becchi PP, Lucini L, et al. (2022) Elderberry (Sambucus nigra L.) Encapsulated Extracts as Meat Extenders against Lipid and Protein Oxidation during the Shelf-Life of Beef Burgers. Antioxidants 11:2130. [CrossRef]
- Wen W, Chen X, Huang Z, et al. (2022) Dietary lycopene supplementation improves meat quality, antioxidant capacity and skeletal muscle fiber type transformation in finishing pigs. Animal Nutrition 8:256–264. [CrossRef]
- Ryu KK, Kang YK, Jeong EW, et al. (2023) Applications of various natural pigments to a plant-based meat analog. LWT 174:114431. [CrossRef]
- Augustyńska-Prejsnar A, Topczewska J, Ormian M, et al. (2022) The Effect of the Addition Turmeric on Selected Quality Characteristics of Duck Burgers Stored under Refrigeration. Applied Sciences 12:805. [CrossRef]
- Bakhsh A, Park J, Baritugo KA, et al. (2023) A holistic approach toward development of plant-based meat alternatives through incorporation of novel microalgae-based ingredients. Front Nutr 10:. [CrossRef]
- Martins T, Barros AN, Rosa E, Antunes L (2023) Enhancing Health Benefits through Chlorophylls and Chlorophyll-Rich Agro-Food: A Comprehensive Review. Molecules 28:5344. [CrossRef]
- El-Anany A, F.M. Ali R, Almujaydil MS, et al. (2024) Innovate plant-based burger patties using defatted sesame cake flour, chickpea flour, coffee silver skin and pomegranate juice as natural colorant: effects on nutritional and acceptability aspect. Nutr Food Sci 54:934–950. [CrossRef]
- Sun C, Ge J, He J, et al. (2021) Processing, Quality, Safety, and Acceptance of Meat Analogue Products. Engineering 7:674–678. [CrossRef]
- Guo Z, Teng F, Huang Z, et al. (2020) Effects of material characteristics on the structural characteristics and flavor substances retention of meat analogs. Food Hydrocoll 105:105752. [CrossRef]
- Zhao Y, Zhao X, Sun P, et al. (2023) Effects of adding other protein products on textural properties of soy protein concentrate-based meat analogs. J Texture Stud 54:410–419. [CrossRef]
- Chen Q, Chen Z, Zhang J, et al. (2023) Application of lipids and their potential replacers in plant-based meat analogs. Trends Food Sci Technol 138:645–654. [CrossRef]
- Jiang W, Yang X, Li L (2024) Flavor of extruded meat analogs: A review on composition, influencing factors, and analytical techniques. Curr Res Food Sci 8:100747. [CrossRef]
- Zhang C, Hua Y, Li X, et al. (2020) Key volatile off-flavor compounds in peas (Pisum sativum L.) and their relations with the endogenous precursors and enzymes using soybean (Glycine max) as a reference. Food Chem 333:127469. [CrossRef]
- Li R, Guo Y, Dong A, Yang X (2023) Protein-based emulsion gels as materials for delivery of bioactive substances: Formation, structures, applications and challenges. Food Hydrocoll 144:108921. [CrossRef]
- Cheng W, Zhu Y, Jiang G, et al. (2023) Sustainable cellulose and its derivatives for promising biomedical applications. Prog Mater Sci 138:101152. [CrossRef]
- Lu Z, Lee P-R, Yang H (2023) Using HPMC to improve sensory properties of vegan omelet analogue: Effect of HPMC on water retention, oil adsorption, and thermal gelation. Food Hydrocoll 144:108938. [CrossRef]
- Bao H, Wang Y, Huang Y, et al. (2025) The Beneficial Role of Polysaccharide Hydrocolloids in Meat Products: A Review. Gels 11:55. [CrossRef]
- Godschalk-Broers L, Sala G, Scholten E (2022) Meat Analogues: Relating Structure to Texture and Sensory Perception. Foods 11:2227. [CrossRef]
- Choton S, Gupta N, Bandral JD, et al. (2020) Extrusion technology and its application in food processing: A review. Pharma Innov 9:162–168. [CrossRef]
- Jang J, Lee D-W (2024) Advancements in plant based meat analogs enhancing sensory and nutritional attributes. NPJ Sci Food 8:50. [CrossRef]
- Fernández-López J, Ponce-Martínez AJ, Rodríguez-Párraga J, et al. (2023) Beetroot juices as colorant in plant-based minced meat analogues: Color, betalain composition and antioxidant activity as affected by juice type. Food Biosci 56:103156. [CrossRef]
- Jin S-K, Shin T-S, Yim D-G (2020) Effects of partial substitution of nitrites with purple-fleshed sweet potato powder on physicochemical characteristics of sausages. J Anim Sci Technol 62:702–712. [CrossRef]
- Huang L, Ren Y, Li H, et al. (2022) Create Fat Substitute From Soybean Protein Isolate/Konjac Glucomannan: The Impact of the Protein and Polysaccharide Concentrations Formulations. Front Nutr 9:. [CrossRef]
- (2020) The Flavor of Plant-Based Meat Analogues. Cereal Foods World 65:. [CrossRef]
- Kale P, Mishra A, Annapure US (2022) Development of vegan meat flavour: A review on sources and techniques. Future Foods 5:100149. [CrossRef]
- Liu X, Chen W, Sun M, et al. (2025) The Effect of Cumin on the Formation of β-Carboline Heterocyclic Amines in Smoked Meat and Simulated Systems. Foods 14:299. [CrossRef]
- Gu Z, Liu S, Duan Z, et al. (2021) Effect of citric acid on physicochemical properties and protein structure of low-salt restructured tilapia (<scp> Oreochromis mossambicus </scp>) meat products. J Sci Food Agric 101:1636–1645. [CrossRef]
- Wu H, Sakai K, Zhang J, McClements DJ (2024) Plant-based meat analogs: color challenges and coloring agents. Food, Nutrition and Health 1:4. [CrossRef]
- Grygier A (2023) Mustard Seeds as a Bioactive Component of Food. Food Reviews International 39:4088–4101. [CrossRef]
- Kaur R, Prasad K (2024) Development and characterization of chickpea based ready to use replacement beverage mix. Journal of Food Measurement and Characterization 18:3595–3618. [CrossRef]
- Lyu X, Ying D, Zhang P, Fang Z (2024) Effect of Whole Tomato Powder or Tomato Peel Powder Incorporation on the Color, Nutritional, and Textural Properties of Extruded High Moisture Meat Analogues. Food Bioproc Tech 17:231–244. [CrossRef]
- Kołodziejczak K, Onopiuk A, Szpicer A, Poltorak A (2021) Meat Analogues in the Perspective of Recent Scientific Research: A Review. Foods 11:105. [CrossRef]
- Baig MA, Ajayi FF, Hamdi M, et al. (2025) Recent Research Advances in Meat Analogues: A Comprehensive Review on Production, Protein Sources, Quality Attributes, Analytical Techniques Used, and Consumer Perception. Food Reviews International 41:236–267. [CrossRef]
- Achinna P, Malleboina P, Joshi AA, et al. (2024) Advancing plant-based meat analogs: Composite blend of pulse protein reinforcing structure with fibrous mushroom, jackfruit seed powder and carboxymethyl cellulose. Food Science and Technology International. [CrossRef]
- Kumar S, Kumar V, Sharma R, et al. (2021) Plant Proteins as Healthy, Sustainable and Integrative Meat Alternates. In: Veganism - a Fashion Trend or Food as a Medicine. IntechOpen.
- Reyes TF, Agrawal P, Chan T, et al. (2023) The Safety of Soy Leghemoglobin Protein Preparation Derived from Pichia pastoris Expressing a Soy Leghemoglobin Gene from Glycine max: In Vitro and In Vivo Studies. J Toxicol 2023:7398724. [CrossRef]
- Vega EN, Ciudad-Mulero M, Fernández-Ruiz V, et al. (2023) Natural Sources of Food Colorants as Potential Substitutes for Artificial Additives. Foods 12:4102. [CrossRef]
- Ryu J, Rosenfeld SE, McClements DJ (2024) Creation of plant-based meat analogs: Effects of calcium salt type on structure and texture of potato protein-alginate composite gels. Food Hydrocoll 156:110312. [CrossRef]
- Samborska K, Boostani S, Geranpour M, et al. (2021) Green biopolymers from by-products as wall materials for spray drying microencapsulation of phytochemicals. Trends Food Sci Technol 108:297–325. [CrossRef]
- Fu S, Ma Y, Wang Y, et al. (2023) Contents and Correlations of Nε-(carboxymethyl)lysine, Nε-(carboxyethyl)lysine, Acrylamide and Nutrients in Plant-Based Meat Analogs. Foods 12:1967. [CrossRef]
- Huang M, Mehany T, Xie W, et al. (2022) Use of food carbohydrates towards the innovation of plant-based meat analogs. Trends Food Sci Technol 129:155–163. [CrossRef]
- Peñaranda I, Garrido MD (2024) Viability of fructooligosaccharides as substitutes for methylcellulose reduction in plant-based burgers. Food Hydrocoll 154:110104. [CrossRef]
- Baydin T, Dille MJ, Aarstad OA, et al. (2023) The impact of sugar alcohols and sucrose on the physical properties, long-term storage stability, and processability of fish gelatin gels. J Food Eng 341:111334. [CrossRef]
- Liu J, Wan P, Xie C, Chen D-W (2021) Key aroma-active compounds in brown sugar and their influence on sweetness. Food Chem 345:128826. [CrossRef]
- Safaei P, Bayat G, Mohajer A (2024) Comparison of fish oil supplements and corn oil effects on serum lipid profile: a systematic review and meta-analysis of randomized controlled trials. Syst Rev 13:54. [CrossRef]
- Rizzo G, Baroni L (2018) Soy, Soy Foods and Their Role in Vegetarian Diets. Nutrients 10:43. [CrossRef]
- Arya SS, Salve AR, Chauhan S (2016) Peanuts as functional food: a review. J Food Sci Technol 53:31–41. [CrossRef]
- Shen J, Liu Y, Wang X, et al. (2023) A Comprehensive Review of Health-Benefiting Components in Rapeseed Oil. Nutrients 15:999. [CrossRef]
- Rahim MA, Ayub H, Sehrish A, et al. (2023) Essential Components from Plant Source Oils: A Review on Extraction, Detection, Identification, and Quantification. Molecules 28:6881. [CrossRef]
- Kołodziejczak K, Onopiuk A, Szpicer A, Poltorak A (2023) The Effect of Type of Vegetable Fat and Addition of Antioxidant Components on the Physicochemical Properties of a Pea-Based Meat Analogue. Foods 13:71. [CrossRef]
- N AA, Sontakke M (2023) Plant-based emulsifiers: Sources, extraction, properties and applications. Pharma Innov 12:08–16. [CrossRef]
- Ahmad Hairi AN, Taufik AM, Zainal Abidin SAS (2023) Product innovation: palm oil fat in plant-based meat. In: Innovation of Food Products in Halal Supply Chain Worldwide. Elsevier, pp 57–66.
- Loganathan R, Subramaniam KM, Radhakrishnan AK, et al. (2017) Health-promoting effects of red palm oil: evidence from animal and human studies. Nutr Rev 75:98–113. [CrossRef]
- Sanders C, Dobson S, Marangoni AG (2024) Effect of saturated and unsaturated fat on the physical properties of plant-based cheese. Curr Res Food Sci 9:100832. [CrossRef]
- Cho Y, Bae J, Lee J, Choi M-J (2023) Storage Stability of Meat Analogs Supplemented with Vegetable Oils. Foods 12:3586. [CrossRef]
- Vatansever S, Hall C (2024) Plant proteins for dry extruded products. In: Functionality of Plant Proteins. Elsevier, pp 339–372.
- Ribeiro G, Piñero M-Y, Parle F, et al. (2024) Optimizing Screw Speed and Barrel Temperature for Textural and Nutritional Improvement of Soy-Based High-Moisture Extrudates. Foods 13:1748. [CrossRef]
- He J, Evans NM, Liu H, Shao S (2020) A review of research on plant-based meat alternatives: Driving forces, history, manufacturing, and consumer attitudes. Compr Rev Food Sci Food Saf 19:2639–2656. [CrossRef]
- Mazumder MdAR, Panpipat W, Chaijan M, et al. (2023) Role of plant protein on the quality and structure of meat analogs: A new perspective for vegetarian foods. Future Foods 8:100280. [CrossRef]
- Arora S, Kataria P, Nautiyal M, et al. (2023) Comprehensive Review on the Role of Plant Protein As a Possible Meat Analogue: Framing the Future of Meat. ACS Omega 8:23305–23319. [CrossRef]
- Schmid E, Farahnaky A, Adhikari B, Torley PJ (2022) High moisture extrusion cooking of meat analogs: A review of mechanisms of protein texturization. Compr Rev Food Sci Food Saf 21:4573–4609. [CrossRef]
- Choi HW, Hahn J, Kim H-S, Choi YJ (2025) The influence of cooling die temperature gradients on the texture of high-moisture meat analogs. Food Chem 468:142403. [CrossRef]
- Maung TT, Gu BY, Kim MH, Ryu GH (2020) Fermentation of texturized vegetable proteins extruded at different moisture contents: effect on physicochemical, structural, and microbial properties. Food Sci Biotechnol 29:897–907. [CrossRef]
- Köllmann N, Schreuders FKG, Zhang L, van der Goot AJ (2023) On the importance of cooling in structuring processes for meat analogues. J Food Eng 350:111490. [CrossRef]
- Zhao L, Zhang M, Chitrakar B, Adhikari B (2021) Recent advances in functional 3D printing of foods: a review of functions of ingredients and internal structures. Crit Rev Food Sci Nutr 61:3489–3503. [CrossRef]
- Piyush, Kumar R, Kumar R (2020) 3D printing of food materials: A state of art review and future applications. Mater Today Proc 33:1463–1467. [CrossRef]
- Ko HJ, Wen Y, Choi JH, et al. (2021) Meat analog production through artificial muscle fiber insertion using coaxial nozzle-assisted three-dimensional food printing. Food Hydrocoll 120:106898. [CrossRef]
- Wen Y, Kim HW, Park HJ (2022) Effects of transglutaminase and cooking method on the physicochemical characteristics of 3D-printable meat analogs. Innovative Food Science & Emerging Technologies 81:103114. [CrossRef]
- Chao C, Park HJ, Kim HW (2024) Effect of l-cysteine on functional properties and fibrous structure formation of 3D-printed meat analogs from plant-based proteins. Food Chem 439:137972. [CrossRef]
- Akharume FU, Aluko RE, Adedeji AA (2021) Modification of plant proteins for improved functionality: A review. Compr Rev Food Sci Food Saf 20:198–224. [CrossRef]
- Chen J, Zhang M, Mujumdar AS, Phuhongsunge P (2022) 4D printing induced by microwave and ultrasound for mushroom mixtures: Efficient conversion of ergosterol into vitamin D2. Food Chem 387:132840. [CrossRef]
- Yang W, Tu A, Ma Y, et al. (2021) Chitosan and Whey Protein Bio-Inks for 3D and 4D Printing Applications with Particular Focus on Food Industry. Molecules 27:173. [CrossRef]
- Du Q, Tu M, Liu J, et al. (2023) Plant-based meat analogs and fat substitutes, structuring technology and protein digestion: A review. Food Research International 170:112959. [CrossRef]
- Cornet SH V., Snel SJE, Schreuders FKG, et al. (2022) Thermo-mechanical processing of plant proteins using shear cell and high-moisture extrusion cooking. Crit Rev Food Sci Nutr 62:3264–3280. [CrossRef]
- Wang Z, Tian Y, Lou F, Guo Z (2024) Effect of Pea Protein Isolate–Soybean Meal Ratio on Fiber Structure and Texture Properties of High-Moisture Meat Analogs. Foods 13:3818. [CrossRef]
- Wittek P, Ellwanger F, Karbstein HP, Emin MA (2021) Morphology Development and Flow Characteristics during High Moisture Extrusion of a Plant-Based Meat Analogue. Foods 2021, Vol 10, Page 1753 10:1753. [CrossRef]
- Chiang JH, Tay W, Ong DSM, et al. (2021) Physicochemical, textural and structural characteristics of wheat gluten-soy protein composited meat analogues prepared with the mechanical elongation method. Food Structure 28:100183. [CrossRef]
- Zahari I, Östbring K, Purhagen JK, Rayner M (2022) Plant-Based Meat Analogues from Alternative Protein: A Systematic Literature Review. Foods 11:2870. [CrossRef]
- Chen Y, Zhang M, Bhandari B, et al. (2021) 3D Printing of Steak-like Foods Based on Textured Soybean Protein. Foods 2021, Vol 10, Page 2011 10:2011. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



