Submitted:
16 June 2025
Posted:
17 June 2025
You are already at the latest version
Abstract
Plants have long adapted to the earth’s changing environmental patterns. Yet, 1 with the current rise of abiotic stresses, such as salinity, temperatures, drought, and nutrient 2 depletion occurring at unpredictable rates threaten global agriculture. If this pattern keeps 3 continuing, then long-evolved regulatory mechanisms can become inadequate to keep 4 pace with environmental disturbances. Consequently, to work through these challenges, 5 human-targeted genetic interventions are requisite. In this review, the recent advancements 6 in plant resilience research, from evolutionary mechanisms (polyploidy, epigenetics, gene 7 duplication, etc.) to modern synthetic technologies (CRISPR-Cas, transgene technology, 8 nanotechnology, and artificial intelligence (AI)), are discussed to redefine the boundaries of 9 plant stress tolerance. By integrating these two domain principles, we can understand how 10 the evolutionary mechanisms can help us in designing precision tools to retain or integrate 11 the lost valuable genetic characteristics. Despite these advancements, major hurdles such 12 as limited field trials, specific isoform functional data, and plants’ ability to adopt these 13 resilient traits still remain. With human interventions and technological strategies, we can 14 improve the plant’s resilience. Here we are not replacing natural evolutionary adaptation, 15 but rather we are building a path for better plant adaptation in these environmental crisis 16 situations and laying the road to sustainable food systems.17
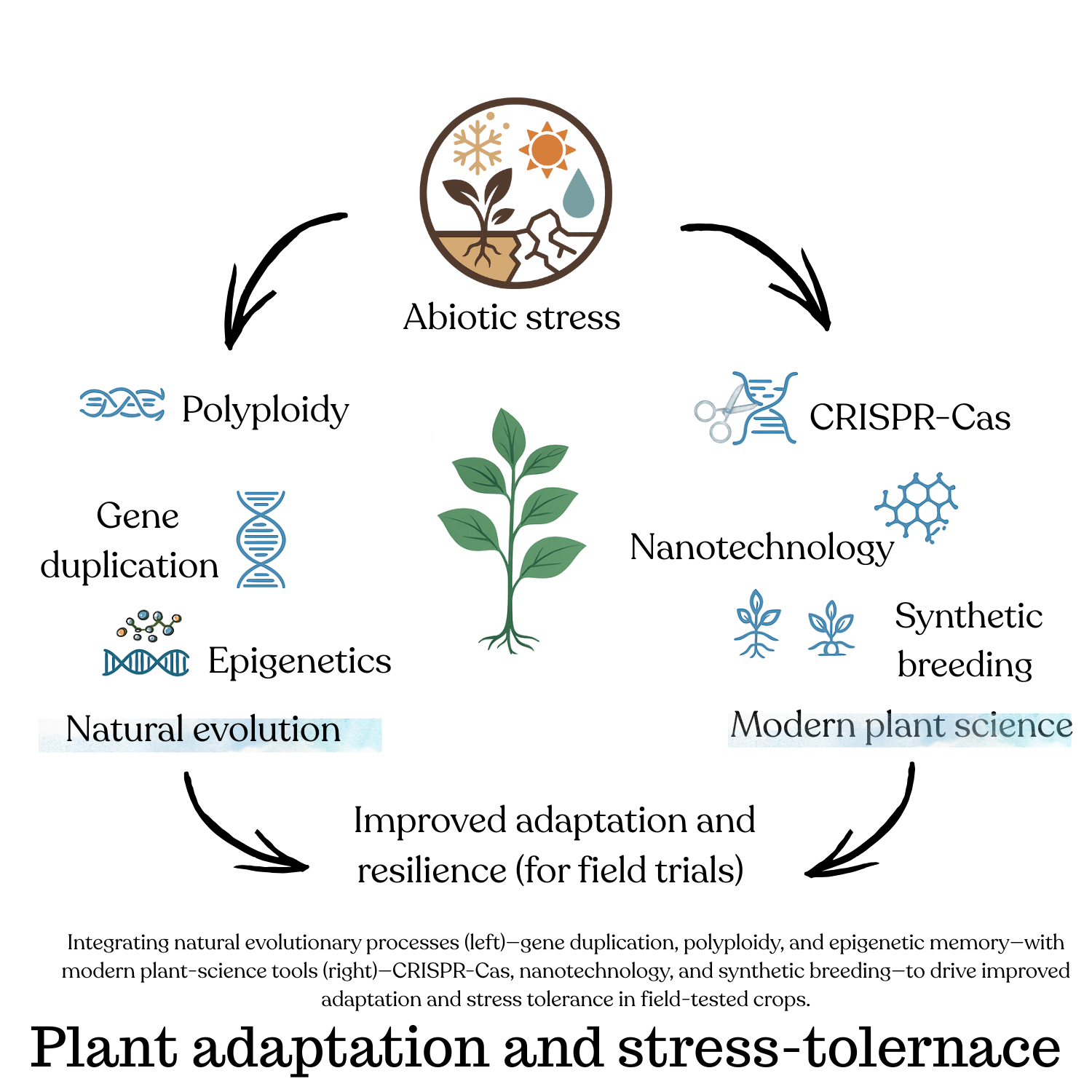
Keywords:
abiotic stress
; CRISPR-Cas
; adaptation
; resilience
; synthetic biology
; precision18
1. Introduction
Through time, plants have endured environmental shifts and adapted themselves, becoming the primary producers in today’s world through many regulatory mechanisms such as gene duplication, polyploidy, and plasticity with the help of evolution. Yet today’s environmental crisis poses a greater challenge to plants to keep up with rapid extreme abiotic stressors such as drought, heat, and salinity just through evolutionary modifications [27]. Natural adaptations are effective, but they are slow-paced to deal with current conditions and pass the adaptive traits across generations.
Most traditional plant breeding methods are constrained by low genetic variability, long generational times and limited trait precision. The key challenges are field validation of laboratory works under real-world conditions, isoform functional data on variants that help stress tolerance, integrating the wild traits without unnecessary conditional limits, and clearing regulatory and safety approvals for these edited crops. Currently around 20-60% of yield is lost on major crops globally depending on region due to abiotic stress [1]. It is crucial to understand that evolution is essential, but with today’s environmental crisis, strategies which can accelerate the yield as well as provide the plant with futuristic pillars to battle the abiotic stresses are needed.
With this regard, engineering and stacking multiple quantitative traits, which can develop resilience in plants against abiotic stress, are vital. This paper mentions natural mechanisms such as epigenetic memory, gene duplication, and alternative splicing, and modern interventions such as transgene technology, nanotechnology, and CRISPR-Cas genome editing. With integrations of these both principal characteristics, tolerant crops to stress can be created and be responsive to future environmental variations.
2. Evolutionary Mechanism for Plant Adaptation
2.1. Gene Duplication and Polyploidy
An alteration in the genome is possible mainly through gene duplication. These duplicated genes can either have new functions or completely different specialised roles to battle the stresses. Gene redundancy impacts plants and helps them survive harmful mutations and environmental shifts [19]. For example, polyploidy in wheat, soybean, and canola contributes to stress resilience by increasing gene redundancy, enhancing biochemical equilibrium, and providing protection against environmental extremities [16].
-
Pros:
- Extra gene copies provide protection against mutations and maintain required functions.
- Polyploidy-derived homologues enhance the stress-coping mechanisms and improve metabolic balance.
-
Cons:
- Entire genome duplications are difficult to induce precisely.
- Polyploid breeding systems require complex crossovers and linkage drags.
-
Unknown:
- How do gene duplicates and homeolog expressions perform under field environmental challenges and their stability in those varied circumstances?
2.2. Epigenetic Memory
DNA methylation and histone modifications can function as the response systems to environmental stimuli. These epigenetic modifications can influence the stress memory and equip the plants to handle chronic or recurring stress [9,24]. For example, drought-induced methylation in rice and tomato has shown promising changes which are perceived over generations [14].
-
Pros
- Stress-induced epigenetic memory is passed on to the next generation and provides transgenerational priming.
- Modifications are reversible, meaning once the stress subsides, plants can reset.
-
Cons
- Methylation due to stress memory markers can revert unpredictably, reducing stability in the long run.
- Current tools lack accuracy and cannot target the epialleles in plants.
-
Unknown
- How long can the epigenetic stress memory markers last across generations and be stable under an uncontrolled field environment?
2.3. Alternative Splicing
DNA methylation and histone modifications can function as the response systems to environmental stimuli. These epigenetic modifications can influence the stress memory and equip the plants to handle chronic or recurring stress [9,24]. For example, drought-induced methylation in rice and tomato has shown promising changes which are perceived over generations [14].
| Pros | ||
| • | Stress-induced epigenetic memory is passed on to the next generation and provides | |
| transgenerational priming. | ||
| • | Modifications are reversible, meaning once the stress subsides, plants can reset. | |
| Cons | ||
| • | Methylation due to stress memory markers can revert unpredictably, reducing stability in the long run. | |
| • | Current tools lack accuracy and cannot target the epialleles in plants. | |
| Unknown | ||
| • | How long can the epigenetic stress memory markers last across generations and be stable under an uncontrolled field environment? | |
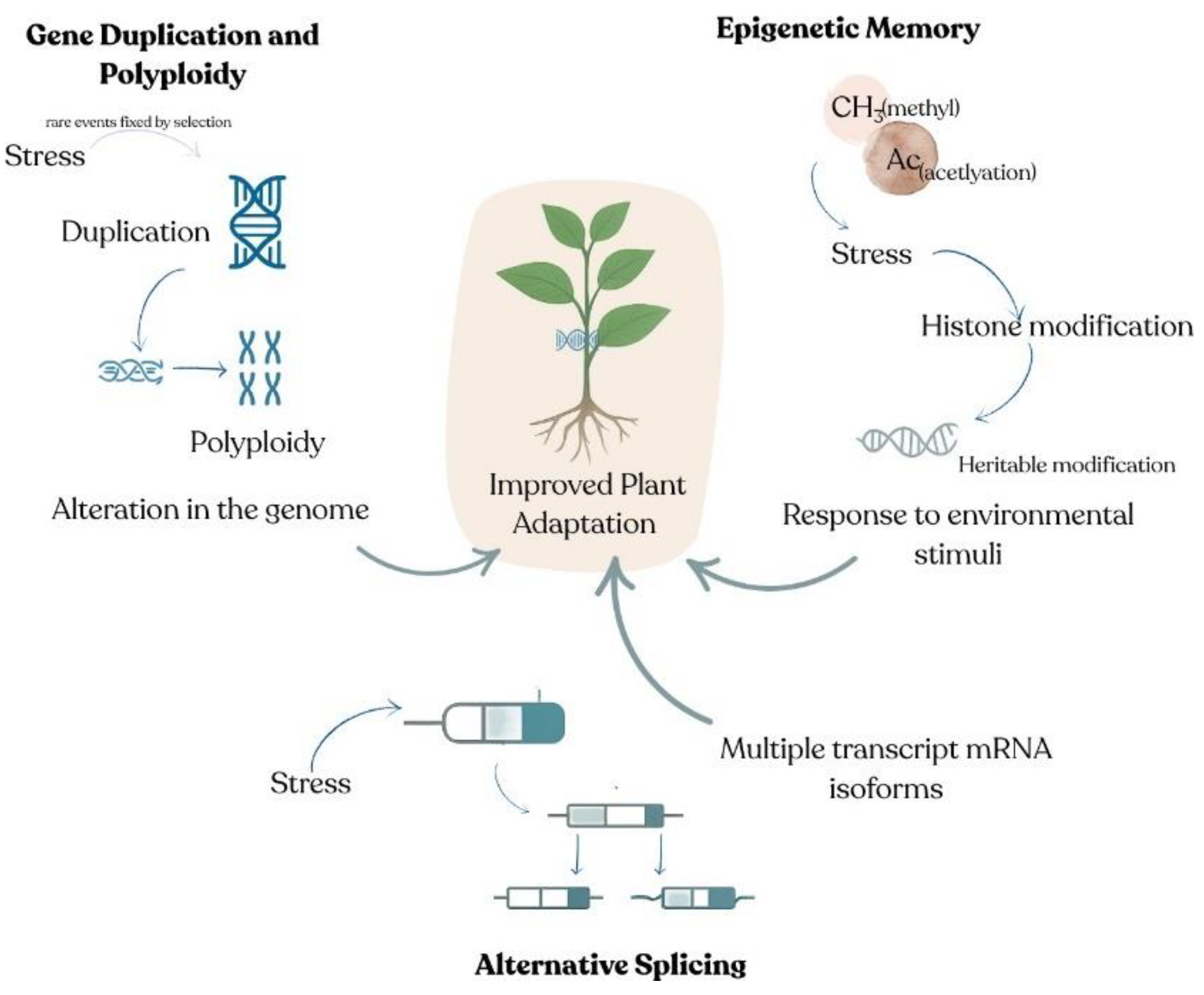
Figure 1.
Schematic overview of key evolutionary mechanisms driving plant stress resilience. (Gene duplication and polyploidy add gene copies that buffer mutations and broaden functionality, while epigenetic memory locks in stress signals via DNA methylation and histone marks. Alternative splicing creates diverse mRNA isoforms to swiftly rewire gene expression—together enhancing plant adaptation to abiotic stress).
Figure 1.
Schematic overview of key evolutionary mechanisms driving plant stress resilience. (Gene duplication and polyploidy add gene copies that buffer mutations and broaden functionality, while epigenetic memory locks in stress signals via DNA methylation and histone marks. Alternative splicing creates diverse mRNA isoforms to swiftly rewire gene expression—together enhancing plant adaptation to abiotic stress).
3. Engineering Strategies for Stress Tolerance in Plants
3.1. CRISPR-Cas Gene Editing
The CRISPR-Cas system advancements have redefined functional genomics and breed- ing methods, allowing researchers to easily edit or modulate the specific regulatory regions. This approach allows effective gene expression modulations without the permanent inte- gration of exogenous sequences into the host [17,27]. For example, editing the OsPYL9 gene with CRISPR-Cas9 in rice, which modulates circadian rhythm, has improved its drought resistance without any negative impact on its yield [25].
Many such tools, like prime or base editors, use CRISPR-Cas9 nickase fusion variants that have enabled the researcher to edit, insert or do point mutations with minimal double-strand breaks [20]. Researchers can edit various stress-adaptive regions with the help of these same principles.
| Pros | ||
| • | Base and prime editors modify single nucleotides with minimal double-strand breaks | |
| and reduce genomic instability. | ||
| • | Transient delivery methods (e.g.,Cas9 RNPs or viral vectors) avoid stable incorporation | |
| of foreign DNA. | ||
| Cons | ||
| • | At low frequencies, screening of off-target mutations is required. | |
| • | Efficient delivery of edits into monocots like wheat and maize is challenging. | |
| Unknown | ||
| • | How do targeted gene edits affect the regulatory mechanism under abiotic stress? | |
3.2. Engineering Gene Networks
With engineered gene circuits responding to environmental stimuli, it is easy to develop promoters that are activated under specific stress conditions. This enables the plants to be more resistant to stress, with high yield and nutritional contents in them [2].
The researchers have implemented synthetic genetic circuits that use Boolean logic gates such as AND, when two or more conditions are met simultaneously and activating gene expressions at that time (e.g., drought AND heat temperatures), and OR when at least one condition is present. These Boolean logic gate configurations are implemented in Arabidopsis to tune the gene expression involved in the root development of the plant and make it respond to specific environmental stimuli; the same stress-responding pathway isobserved in rice [3].
| Pros | ||
| • | Boolean logic gates enable the expression of stress-tolerant genes under specific envi- | |
| ronmental signals. | ||
| • | Modular circuits help in the rapid swapping of sensors and outputs. | |
| Cons | ||
| • | Under non-stress conditions, synthetic circuits might slow the growth due to metabolic load. | |
| • | The data on synthetic circuit function in polyploid crop genomics is limited. | |
| Unknown | ||
| • | How do these circuits function in large-scale field plots under fluctuating environmental stress signals? | |
3.3. Transgene Technology
Transgene technology is a powerful tool in designing new or unique traits for plants to fight against the abiotic stresses and drawbacks of breeding approaches. For example, GmNFYA13 gene overexpression in soybean has shown improved tolerance to drought and salt levels in the soil [18]. With the help of transgene technology, crops can be developed that can withstand the environmental stresses.
| Pros | ||
| • | Under laboratory and greenhouse conditions, transgenes provide a high level of | |
| expression of specific resilience genes. | ||
| • | With well-characterized promoters, the trait stability expressions are possible. | |
| Cons | 150 | |
| • | Approvals from regulatory boards and the public vary depending on regions. | |
| • | Over generations, transgenic silencing occurs and leads to reduced trait stability. | |
| Unknown | ||
| • | How do introduced transgenes and native genes interact with each other under stress conditions in field settings? | |
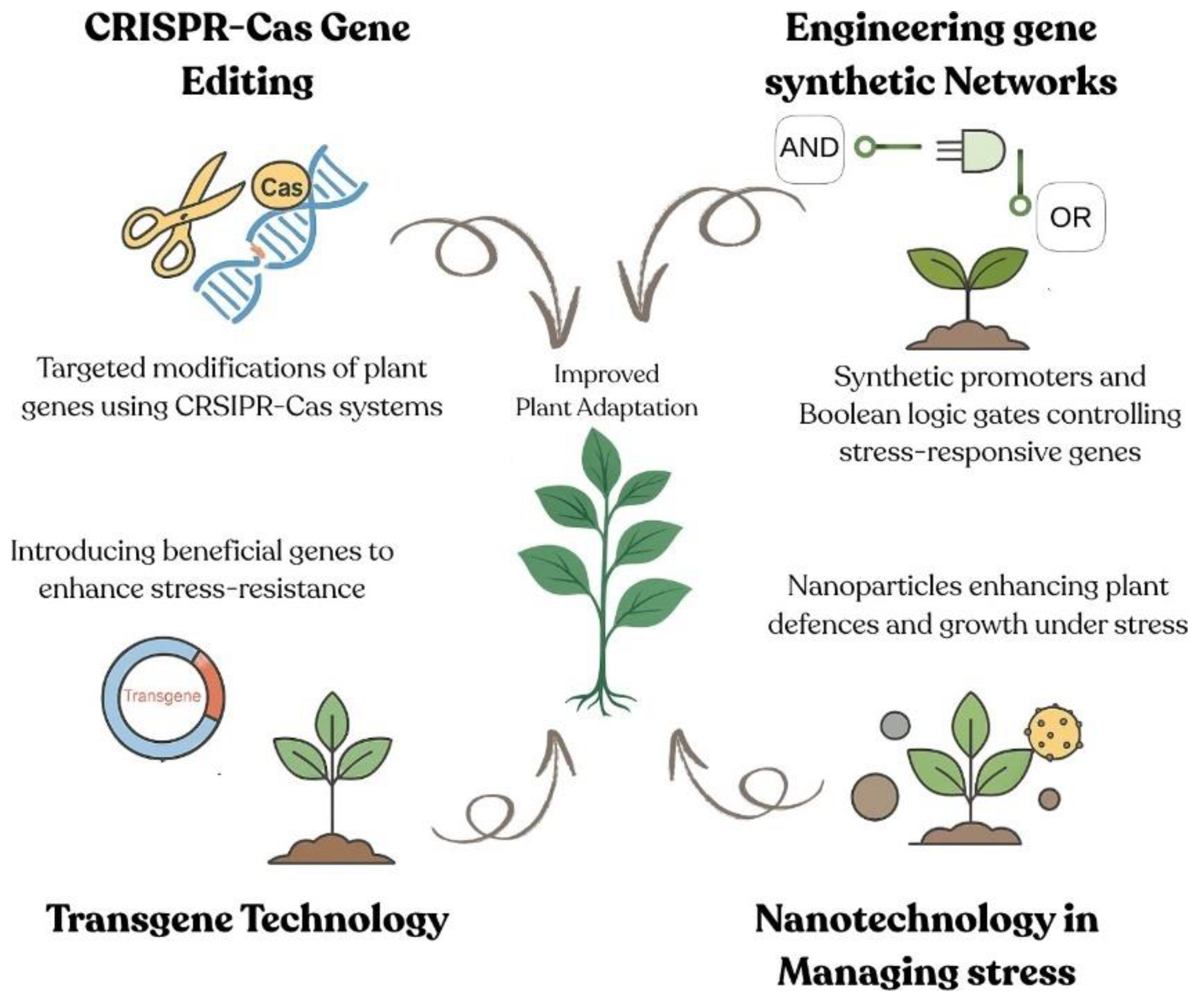
Figure 2.
Integrating synthetic biology tools to boost plant stress tolerance. (CRISPR-Cas gene editing enables precise targeting of endogenous loci; engineered gene circuits with synthetic promot- ers and Boolean logic gates allow condition-specific activation of stress-responsive genes; transgene technology introduces novel or enhanced traits for resilience; and nanotechnology delivers nutrients or protective agents directly to plant cells—all converging to improve adaptation under abiotic stress).
Figure 2.
Integrating synthetic biology tools to boost plant stress tolerance. (CRISPR-Cas gene editing enables precise targeting of endogenous loci; engineered gene circuits with synthetic promot- ers and Boolean logic gates allow condition-specific activation of stress-responsive genes; transgene technology introduces novel or enhanced traits for resilience; and nanotechnology delivers nutrients or protective agents directly to plant cells—all converging to improve adaptation under abiotic stress).
3.4. Nanotechnology in Managing Stress
Many studies have listed the importance of selected nanoparticles’ role in photosyn-thesis, oxidative stress, and delivering the required nutrients into plant cells, though species and dosage vary widely [6]. Many studies have emphasised the importance of nanoparti- cles’ role in mitigating the abiotic stress. For example, cerium oxide (CeO2) nanoparticles enhance the functions of antioxidant enzymes and reduce the oxidative damage in apple plants under drought conditions and improve plant growth in water deficit conditions [23].
-
Pros
- Nanoparticles deliver nutrients or signalling molecules directly into plant cells with ease and hence enhance the functions of specific targeted activity.
- Engineered design allows the controlled release of particles under specific abiotic stress conditions.
-
Cons
- The specific size and dose of nanoparticles suitable for soil types and crops are yet to be identified.
- Toxicity to nontarget organisms and soil microbes is the biggest concern.
-
Unknown
- What happens to nanoparticles over time in an agricultural ecosystem, and how do they interact with nutrient cycles and microbes or other organisms and their potential effects on them? This is an example of a quote.
4. Integration of Evolutionary Mechanism and Engineered Strategies
Evolutionary mechanisms, such as gene duplication, polyploidy, and epigenetic modifications, provide adaptive mechanisms for developing plant resilience, but these mechanisms occur at slow rates and are unpredictable with current fast-changing environmental dynamics.
Engineered tools (CRISPR-Cas, transgenics, synthetic gene circuits, and nanoparticles) can provide the required speed, precision, and flexibility, but these are less sophisticated [27] when compared to natural adaptations which are better adapted to environment and can be integrated into existing cells and respond to even subtle changes in environments. With the integration of insights from evolutionary adaptations with modern technologies, we can develop crops with resilient genes or transcription factors to abiotic stresses. For example, epigenetic mapping of plants can identify stress-prone regions, like drought, and can be used to guide the synthetic gene circuit promoters to regulate the gene expression and study their function in detail and understand their role in developing resilient crops in the future. The integration of these methods is not to mimic the expression blindly but to understand the logic behind it and translate it with precision and develop tools that build resilience to posed environmental changes while respecting ecological diversity while abiding by rules of nature, limiting the impacts on non-target organisms, and local gene pool.
5. Gaps in Current Research
With many technological advancements, critical gaps still persist:
5.1. Limited Field Validations
Many studies have revealed the potential of engineered traits for stress tolerance. But actual practice in the complexity of environmental conditions remains unexplored. Field trials in practical settings are required to validate the traits’ efficiency, adaptability, and stability across generations.
5.2. Limited Understanding of Isoform-Specific Functions
Multiple isoforms can be generated using alternative splicing (AS), but we still need to study further about their functions and regulatory mechanisms. A full understanding of these forms can help in understanding the genetic intervention that enables stress tolerance in plants.
5.3. Underutilisation of Wild Relatives
The undomesticated relatives of crops have various adaptive traits that have developed over many years of survival mechanisms. While continuous breeding in particular regions has a positive impact on yield, the overall genetic diversity of plant species is narrowed to a bottleneck. With the help of modern technologies like gene editing, speed breeding, and genomic selection, researchers are required to break the traditional barriers in an ethical way to develop sustainable agriculture systems.
5.4. Ineffective Interaction Across Disciplines
Many researchers work independently to arrive at a solution. These fragmented approaches treat functional genomics, phenomics, epigenomics, and artificial intelligence (AI) as independent domains, and vital connections for crop improvement will be lost. We can work in the form of a cross-disciplinary team integrating functional genomics, phenomics, epigenomics, and artificial intelligence (AI) to build productive, resilient, and sustainable crops.
5.5. Regulatory Acceptances
The genome editing faces uncertainties from regulatory boards and the public, particularly concerning ethical and environmental safety. For example, transgenic GMOs face > 30 percent public rejection rates in the EU, whereas gene-edited crops using SDN-1 are considered non-GMO in the US and Argentina, leading to permissive regulation [13].
6. Future of Resilient Crop Design
We propose a few strategies to overcome limitations in designing future-proof crops that withstand current dynamic environmental shifts as follows:
6.1. Isoform-Specific Functional Genomics
- Most gene editing strategies include knockout or overexpression of target genes. However, with assistance from alternative splicing, multiple mRNA isoforms can be produced from a single gene, which can serve as an adaptive mechanism under stress [7]. These isoforms can have different functions and be used to design crops with improved efficacy instead of whole-genome modification; this approach minimises off- target modifications and enhances stress adaptation while preserving native regulation. For example, in rice, OsDREB2B gene isoforms can confer heat tolerance independent of the DREB pathway [8].– Further field trials needed to be conducted on rice and wheat under controlled drought conditions to identify specific tissue expressions and the resilience correlation of stress-responsive isoforms, such as DREB2B or SR proteins.
- Future work: which stress-responsive isoforms can yield stability under abiotic stresses needs to be studied in field conditions to understand their survival rates over two or more seasons.
6.2. Integration of Wild Germplasm with the Help of Genomic-Assisted Breeding
-
Wild relatives of any plant varieties have a vast range of gene pools, which have been developed over the years with the help of natural selection under express environ- mental conditions. Their integration into modern-day commercial or non-commercial crops can enhance resilience with the desired speed and specificity needed for improv- ing crop traits. These integrations can be achieved by the following methods:
- -
- AI haplotype mapping uses a diverse variety of genome library data to mine specific stress-resilient alleles. For example, use of AI-augmented haplotype map- ping to identify superior drought-responsive haplotypes in > 1000 rice accessions to increase yield in water-deficit field trial conditions with > 80 percent predictive accuracy [22].
- -
- With rapid-generation advancement (speed breeding) along with genomic selection (GS) allele stacking, genes can be transferred from one species to another and improve their allele efficiency. For example, use of both speed breeding and GS cycles for early detection of drought-tolerant alleles via GS panels in wheat [10].
- -
- Through CRISPR-Cas, we can edit gene lines to recreate important or beneficial wild alleles by bypassing linkage completely instead of just crossing. For example, with base editors of CRISPR-Cas9 we can reconstruct beneficial domesticated alleles via precise nucleotide substitutions, minimising double-strand breaks and linkage drag, while preserving stress-resistant wild traits [15].
- -
- For example, in wild barley, genes of salt tolerance can be mapped and edited into modern varieties without traditional breeding approaches and improve their salinity [26].
- Future work: the targeted editing of transcripts such as m6A needed further understanding of how to accelerate the recovery from abiotic stresses.
6.3. Development of Smart Phenotypic Field Platforms
-
Real-world applications are more complex and fluctuating compared to laboratory conditions. To address this issue, the implementation of smart field testing is required, with the combinational use of nanotechnology, sensors, and artificial intelligence (AI) not just for labs but for regional institutions globally, which can transform field trials into more adaptive and effective centres for testing:
- -
- In-plant and soil nanosensors can enable continuous real-time tracking of ROS, ABA, ion fluxes, etc. For example, carbon-dot-embedded graphene nanosheets de- tect ROS fluctuation [5]; dual-functional sensors based on charged gold nanorods (AuNRs) are used to detect ABA levels [28]; and the use of electrochemical nanosensors along with metal-decorated carbon nanotube electrodes detects real-time soil nitrate (NO3-) fluxes limited to parts per billion (ppb) range [12].
- -
- Drone imaging and AI models can detect stress signatures with the help of canopy temperatures and spectral shifts. For example, thermal imaging of stom- atal closure in wheat fields [4]; the combination of thermal, multispectral, and hyperspectral sensors and drones with VNIR can detect the temperature shifts with <1 m resolution, and AI helps in predicting the water stress with 85 percent accuracy and vegetation index abnormalities [21].
- -
- We can equip plants with engineered environmental sensing synthetic switches combined with sensors and regulators to respond to any external signals (e.g., controlled stress-oscillation circuits to detect ROS and ABA levels [11]).
- Future work: how the delivery system of stress-protective genes with tissue-specific promoters can help in improving resilience without any permanent transformative changes.
6.4. Stakeholder Communication and Engagement
- Any research is never autonomous of its intended target group. A transparent com- munication is needed between all farmers, policymakers, regulators, and the public in understanding the benefits and risks of bringing technologies together for crop improvement. Collaborative networks needed to be established with experts from genomics, data science, agronomy, and social sciences, to develop integrated, innovative and regulatory frameworks for increased acceptance towards crop varieties.
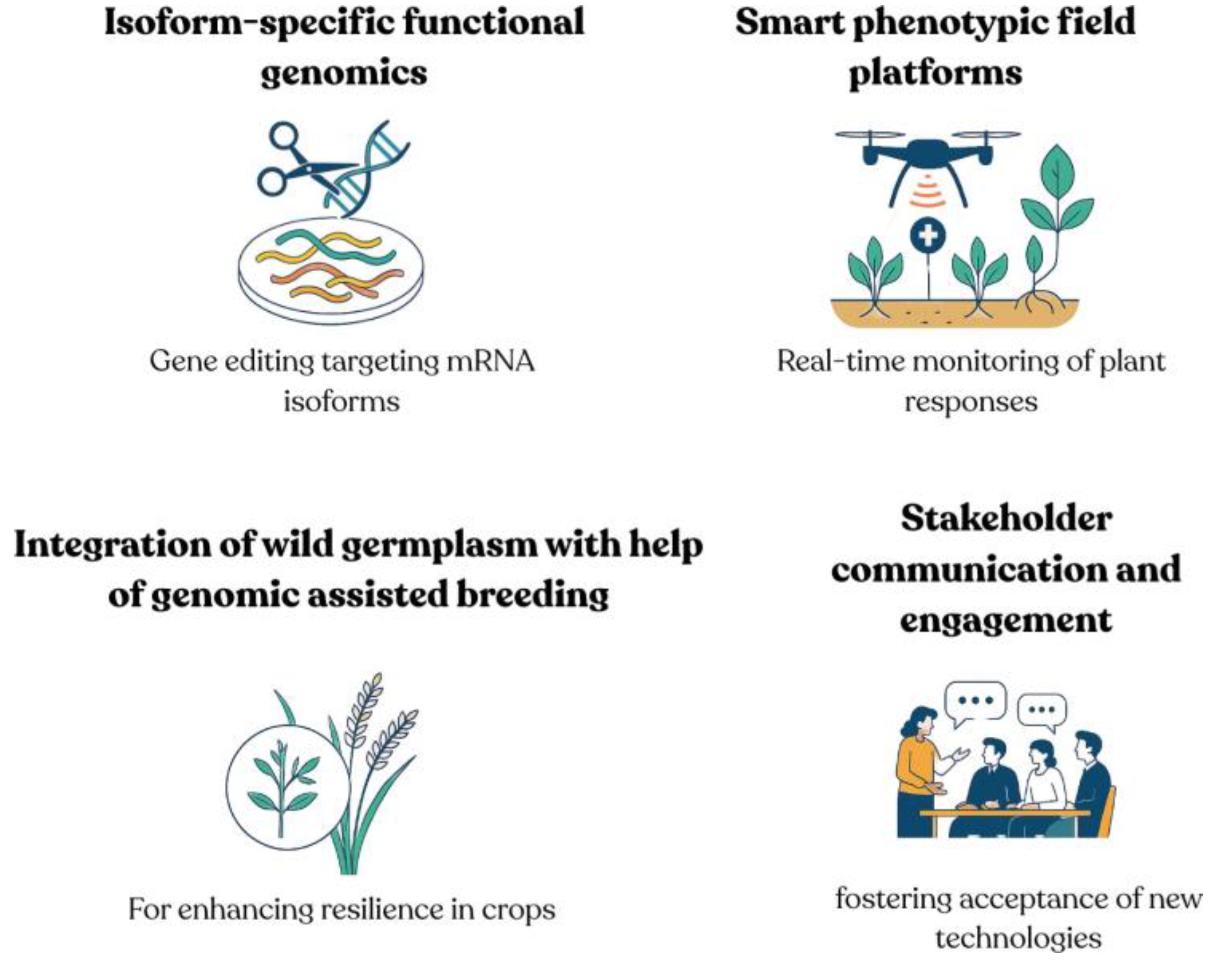
Figure 3.
Next-generation strategies for resilient crop development. ((Top left) Isoform-specific functional genomics uses targeted gene editing to manipulate stress-responsive mRNA variants. (Top right) Smart phenotyping platforms combine real-time nanosensors and drone imaging to monitor plant stress responses in the field. (Bottom left) Integration of wild germplasm via genomic- assisted breeding leverages natural diversity for enhanced resilience. (Bottom right) Stakeholder communication and engagement foster public acceptance and informed decision-making around new agricultural technologies).
Figure 3.
Next-generation strategies for resilient crop development. ((Top left) Isoform-specific functional genomics uses targeted gene editing to manipulate stress-responsive mRNA variants. (Top right) Smart phenotyping platforms combine real-time nanosensors and drone imaging to monitor plant stress responses in the field. (Bottom left) Integration of wild germplasm via genomic- assisted breeding leverages natural diversity for enhanced resilience. (Bottom right) Stakeholder communication and engagement foster public acceptance and informed decision-making around new agricultural technologies).
7. Conclusion
The plant resilience story is evolving rapidly every day. In this review, we highlighted how evolutionary mechanisms such as gene duplication, epigenetics, etc., with modern tools such as CRISPR-Cas, synthetic biology, etc., can bring out the powerful approach to tackle environmental challenges and make crops resilient. Instead of just replacing nature, we can translate evolutionary insights into engineered solutions and pave the path for future agriculture, where crops will be stress tolerant, adaptive, responsive and resilient, ready to handle not only present environmental conditions but also future ones. We recommend the use of multi-season field trials of alternative splicing to understand more about agronomic benefits of diverse stress-responsive traits and their stability across generations and different environmental conditions and creating interdisciplinary teams for sensor-augmented phenotyping to interpret nanosensors, drones and AI platforms usage in field trial settings. With integration expertise, the next generation of edited crops can move from laboratories to agricultural lands with reliability and prowess.
Author Contributions
All work (conceptualization; methodology; investigation; original draft preparation; review and editing; visualization) was carried out by V.S.S.P. The author has read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
During the preparation of this manuscript, the author used Canva \- (www.canva.com) for figure creation, QuillBot (www.quillbot.com) for language editing, and OpenAI to assist with reference formatting; the author has reviewed and approved all content. The author also thanks the anonymous reviewers for their constructive feedback and acknowledges the foundational work of all the researchers cited in this manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic Strategies for Improving Crop Yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef]
- Belcher, M.S.; Vuu, K.M.; Zhou, A.; Mansoori, N.; Agosto Ramos, A.; Thompson, M.G.; Scheller, H.V.; Loqué, D.; Shih, P.M. Design of Orthogonal Regulatory Systems for Modulating Gene Expression in Plants. Nat. Chem. Biol. 2020, 16, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Brophy, J.A.N.; Magallon, K.J.; Duan, L.; Zhong, V.; Ramachandran, P.; Kniazev, K.; Dinneny, J.R. Synthetic Genetic Circuits as a Means of Reprogramming Plant Roots. Science 2022, 377, 747–751. [Google Scholar] [CrossRef]
- Costa, J.M.; Grant, O.M.; Chaves, M.M. Thermography to Explore Plant–Environment Interactions. J. Exp. Bot. 2013, 64, 3937–3949. [Google Scholar] [CrossRef] [PubMed]
- Demirer, G.S.; Zhang, H.; Matos, J.L.; Goh, N.S.; Cunningham, F.J.; Sung, Y.; Chang, R.; Aditham, A.J.; Chio, L.; Cho, M.J.; Staskawicz, B.; Landry, M.P. High Aspect Ratio Nanomaterials Enable Delivery of Functional Genetic Material without DNAIntegration in Mature Plants. Nat. Nanotechnol. 2019, 14, 456–464. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Saad, A.M.; Soliman, S.M.; Salem, H.M.; Desoky, E.M.; Babalghith, A.O.; El-Tahan, A.M.; Ibrahim, O.M.; Ebrahim, A.A.M.; Abd El-Mageed, T.A.; Elrys, A.S.; Elbadawi, A.A.; El-Tarabily, K.A.; AbuQamar, S.F. Role of Nanoparticles inEnhancing Crop Tolerance to Abiotic Stress: A Comprehensive Review. Front. Plant Sci. 2022, 13, 946717. [Google Scholar] [CrossRef]
- Filichkin, S.; Priest, H.D.; Megraw, M.; Mockler, T.C. Alternative Splicing in Plants: Directing Traffic at the Crossroads ofAdaptation and Environmental Stress. Curr. Opin. Plant Biol. 2015, 24, 125–135. [Google Scholar]
- Ganie, S.A.; Reddy, A.S.N. Stress-Induced Changes in Alternative Splicing Landscape in Rice: Functional Significance of SpliceIsoforms in Stress Tolerance. Biology 2021, 10, 309. [Google Scholar] [CrossRef]
- Gallusci, P.; Dai, Z.; Génard, M.; Gauffretau, A.; Leblanc-Fournier, N.; Richard-Molard, C.; Vile, D.; Brunel-Muguet, S. Epigenetics for Plant Improvement: Current Knowledge and Modeling Avenues. Trends Plant Sci. 2017, 22, 610–623. [Google Scholar] [CrossRef]
- Jighly, A.; Lin, Z.; Pembleton, L.W.; Cogan, N.O.I.; Spangenberg, G.C.; Hayes, B.J.; Daetwyler, H.D. Boosting Genetic Gain inAllogamous Crops via Speed Breeding and Genomic Selection. Front. Plant Sci. 2019, 10, 1364. [Google Scholar] [CrossRef]
- Khan, A.; Lister, R. Synthetic Gene Circuits in Plants: Recent Advances and Challenges. Quant. Plant Biol. 2025, 6, e6. [Google Scholar] [CrossRef] [PubMed]
- Kundu, M.; Krishnan, P.; Chobhe, K.A.; Manjaiah, K.M.; Pant, R.P.; Chawla, G. Fabrication of Electrochemical Nanosensor forDetection of Nitrate Content in Soil Extract. Journal of Soil Science and Plant Nutrition 2022, 22, 2777–2792. [Google Scholar] [CrossRef]
- Lassoued, R.; Macall, D.; Smyth, S.; Phillips, P.; Hesseln, H. Risk and Safety Considerations of Genome Edited Crops: ExpertOpinion. Curr. Res. Biotechnol. 2019, 1, 100011. [Google Scholar] [CrossRef]
- Li, J.; Yang, D.L.; Huang, H.; Zhang, G.; He, L.; Pang, J.; Lozano-Durán, R.; Lang, Z.; Zhu, J.K. Epigenetic Memory MarksDetermine Epiallele Stability at Loci Targeted by De Novo DNA Methylation. Nat. Plants 2020, 6, 661–674. [Google Scholar] [CrossRef]
- Li, T.; Yang, X.; Yu, Y.; Si, X.; Zhai, X.; Zhang, H.; Dong, W.; Gao, C.; Xu, C. Domestication of Wild Tomato Is Accelerated byGenome Editing. Nat. Biotechnol. 2018, 36, 890–895. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, L.; Wei, X.; Datta, T.; Wei, F.; Xie, Z. Polyploidization: A Biological Force That Enhances Stress Resistance. Int. J.Mol. Sci. 2024, 25, 1957. [Google Scholar] [CrossRef]
- Lin, Q.; Zong, Y.; Xue, C.; Wang, S.; Jin, S.; Zhu, Z.; Wang, Y.; Anzalone, A.V.; Raguram, A.; Doman, J.L.; Liu, D.R.; Gao, C. PrimeGenome Editing in Rice and Wheat. Nat. Biotechnol. 2020, 38, 582–585. [Google Scholar] [CrossRef]
- Ma, X.J.; Fu, J.D.; Tang, Y.M.; Yu, T.F.; Yin, Z.G.; Chen, J.; Zhou, Y.B.; Chen, M.; Xu, Z.S.; Ma, Y.Z. GmNFYA13 Improves Salt and Drought Tolerance in Transgenic Soybean Plants. Front. Plant Sci. 2020, 11, 587244. [Google Scholar] [CrossRef] [PubMed]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.H. Evolution of Gene Duplication in Plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef]
- Rodríguez-Leal, D.; Lemmon, Z.H.; Man, J.; Bartlett, M.E.; Lippman, Z.B. Engineering Quantitative Trait Variation for CropImprovement by Genome Editing. Cell 2017, 171, 470–480.e8. [Google Scholar] [CrossRef]
- Sharma, H.; Sidhu, H.; Bhowmik, A. Remote Sensing Using Unmanned Aerial Vehicles for Water Stress Detection: A ReviewFocusing on Specialty Crops. Drones 2025, 9, 241. [Google Scholar] [CrossRef]
- ]ref-journal Singh, P.; Sundaram, K.T.; Vinukonda, V.P.; Venkateshwarlu, C.; Paul, P.J.; Pahi, B.; Gurjar, A.; Singh, U.M.; Kalia,S. ; Kumar, A.; Singh, V.K.; Sinha, P. Superior Haplotypes of Key Drought-Responsive Genes Reveal Opportunities for theDevelopment of Climate-Resilient Rice Varieties. Commun. Biol. 2024, 7, 89. [Google Scholar]
- Soleymani, S.; Piri, S.; Aazami, M.A.; Salehi, B. Cerium Oxide Nanoparticles Alleviate Drought Stress in Apple Seedlings byRegulating Ion Homeostasis, Antioxidant Defense, Gene Expression, and Phytohormone Balance. Sci. Rep. 2025, 15, 11805. [Google Scholar] [CrossRef] [PubMed]
- Thiebaut, F.; Hemerly, A.S.; Ferreira, P.C.G. A Role for Epigenetic Regulation in the Adaptation and Stress Responses ofNon-Model Plants. Front. Plant Sci. 2019, 10, 246. [Google Scholar] [CrossRef]
- Usman, B.; Nawaz, G.; Zhao, N.; Liao, S.; Liu, Y.; Li, R. Precise Editing of the OsPYL9 Gene by RNA-Guided Cas9 NucleaseConfers Enhanced Drought Tolerance and Grain Yield in Rice (Oryza sativa L.) by Regulating Circadian Rhythm and AbioticStress Responsive Proteins. Int. J. Mol. Sci. 2020, 21, 7854. [Google Scholar] [CrossRef]
- Varshney, R.K.; Singh, V.K.; Kumar, A.; Powell, W.; Sorrells, M.E. Can Genomics Deliver Climate-Change Ready Crops? Curr.Opin. Plant Biol. 2018, 45, 205–211. [Google Scholar]
- Zafar, S.A.; Zaidi, S.S.; Gaba, Y.; Singla-Pareek, S.L.; Dhankher, O.P.; Li, X.; Mansoor, S.; Pareek, A. Engineering Abiotic StressTolerance via CRISPR/Cas-Mediated Genome Editing. J. Exp. Bot. 2020, 71, 470–479. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, W.; Zhang, H.; Wang, S.; Li, X.; Naqvi, S.M.Z.A.; Hu, J. Dual-Functional SERRS and Fluorescent Aptamer Sensor for Abscisic Acid Detection via Charged Gold Nanorods. Front. Chem. 2022, 10, 965761. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



