
Submitted:
12 June 2025
Posted:
13 June 2025
You are already at the latest version
Abstract
Military personnel deployed to Iraq and Afghanistan were exposed to emissions from open-air burn pits, where plastics, metals, and medical waste were incinerated. These exposures have been linked to deployment-related respiratory diseases (DRRD) and may also impact neurological health via the lung–brain axis. To investigate molecular mechanisms, adult male rats were exposed to filtered air, naphthalene (a representative volatile organic compound), or a combination of carbon black (surrogate for particulate matter) and naphthalene (CBN) via whole-body inhalation (six hours/day, three consecutive days). Lung, brain, and plasma samples were collected 24 hours after final exposure. Proinflammatory biomarkers were assessed using multiplex electrochemiluminescence and western blotting. Differentially expressed genes (DEGs) were identified by RNA sequencing, and elastic net modeling was used to define exposure-predictive gene signatures. CBN exposure altered inflammatory biomarkers across tissues, with activation of nuclear factor kappa B (NF-κB) signaling. In the lung, gene set enrichment revealed activated pathways related to proliferation and inflammation, while epithelial-mesenchymal transition (EMT) and oxidative phosphorylation were suppressed. In the brain, EMT, inflammation, and senescence pathways were activated, while ribosomal function and oxidative metabolism were downregulated. Elastic net modeling identified a lung gene signature predictive of CBN exposure, including Kcnq3, Tgfbr1, and Tm4sf19. These findings demonstrate that inhalation of a surrogate burn pit mixture induces inflammatory and metabolic gene expression changes in both lung and brain tissues, supporting the utility of this animal model for understanding systemic effects of airborne military toxicants and for identifying potential biomarkers relevant to DRRD and Veteran health.
Keywords:
Military Burn Pit Exposure
; Military Personnel
; Environmental Exposure
; Respiratory Diseases
; Lung Diseases
; Brain Diseases
; Inflammation
; Cytokines
; Gene Expression Profiling
; Oxidative Stress
; Tumor Necrosis Factor-alpha (TNF α)
; Interferon-gamma (IFN γ)
; Air Pollutants
; Nanoparticles
; Polycyclic Aromatic Hydrocarbons
; Volatile Organic Compounds
; Elastic Net
; Gene Set Enrichment Analysis (GSEA)
1. Introduction
Military personnel deployed to combat zones such as Iraq and Afghanistan over the past two decades have been exposed to multiple environmental hazards, including emissions from open-air burn pits used for waste disposal [1,2,3]. Open burn pits were the primary means of waste disposal on military bases in these regions, generating complex mixtures of toxic pollutants that have been linked to respiratory diseases, cancers, and systemic and central inflammation [1,4,5,6,7,8]. The usage of open burn pits, which incinerate a wide range of materials including plastics, metals, medical waste, electronics, chemicals, and petroleum products, has raised significant concerns about potential long-term occupational health effects in Veterans [9]. The Department of Defense estimates that large base sites collectively burned up to 85,000 pounds of waste per day, with the largest burn pit at Joint Base Balad spanning over 25 acres at its peak [10].
Burn pit emissions are characterized by high concentrations of particulate matter (PM), including fine particulate matter with a diameter of 2.5 micrometers or less (PM2.5), which frequently far exceeded U.S. air pollution standards [2,11]. The emissions also contained a complex mixtures of toxic gases, such as polycyclic aromatic hydrocarbons (PAHs) and volatile organic compounds (VOCs), which have been linked to a variety of acute and chronic health issues [10,11,12,13,14]. Exposure to burn pits has been associated with deployment-related respiratory diseases (DRRD) which encompass a spectrum of debilitating pulmonary conditions, including constrictive bronchiolitis and chronic obstructive pulmonary disease [15]. Emerging research also highlights the broader systemic effects of burn pit-derived pollutants, implicating them in neuroinflammation and immune dysregulation through pathways such as the lung-brain axis [4,5,8,16,17,18]. Burn pit toxicants, including PM, VOCs, and PAHs, are linked to neuroinflammation and increased risk of neurodegenerative conditions such as Alzheimer’s disease (AD) and Parkinson’s disease (PD) [8,17,19]. One of the most thoroughly examined hypotheses concerns the adverse health effects of airborne burn pit emissions on the activation of inflammatory responses [9,16,20]. These inflammatory responses are thought to mediate the causal pathways from respiratory exposures to the development of pulmonary, cardiovascular, neurological, and malignant diseases [17,21,22,23]. It is essential to understand these mechanisms to accurately assess the long-term health risks and develop targeted interventions for affected Veterans.
To model military burn pit emissions, we exposed rats to either naphthalene (NA) alone or to a combination of carbon black nanoparticles and naphthalene (CBN). This stepwise design allowed us to isolate the effects of VOCs and PAHs, as represented by naphthalene, as well as their combined effects with PM2.5, represented by carbon black (CB). Our prior studies demonstrated that CB exposures alone can induce both systemic and central inflammation, but that model was limited in its inability to capture the complexity of burn pit toxicants [16]. The addition of naphthalene addressed this gap by incorporating key vapor-phase components common to military burn pit emissions [10,11].
Following the exposures, we conducted protein biomarker profiling, which revealed that the CBN group exhibited the most pronounced inflammatory response across lung, plasma, and brain tissues. Based on this robust protein-level activation of inflammatory pathways, we selected CBN-exposed lung and brain tissues for RNA-seq to explore gene expression changes that underlie this response. We also applied predictive modeling to gene expression in lung, the primary site of contact with airborne burn pit emissions, to identify early molecular indicators of inhalational toxicant exposure. Among all tissues analyzed, the lung exhibited the most robust transcriptional responses, making it an ideal candidate for developing a gene expression signature reflective of direct environmental insults. An elastic net model identified genes predictive of CBN exposure in lung. By delineating key genes and molecular pathways, we identified potential biomarkers of exposure and early indicators of disease progression, supporting our long-term goal of developing diagnostic and therapeutic strategies to help diagnose and treat Veterans affected by military environmental exposures.
2. Results
2.1. Whole-Body Inhalation Exposure of Rats to Sham air, Naphthalene, or Carbon Black with Naphthalene
Rats were exposed via whole-body inhalation to either HEPA-filtered air (sham control group), naphthalene (NA) vapor, or a mixture of carbon black and NA vapor (CBN) for six hours per day over three consecutive days. For the CBN group, the aerosol mass concentration was 7.58 ± 1.55 mg/m³ and the naphthalene vapor concentration was maintained at 10.16 ± 0.01 parts per million (PPM). Aerosol characterization revealed a median particle diameter of 154 nm with a geometric standard deviation of 2.71, confirming that the aerosolized particles were within the respirable range for rodent inhalation studies. These exposure levels were selected to simulate realistic and environmentally relevant conditions based on those reported near military burn pit sites [24]. Naphthalene was chosen for its high volatility, well-characterized toxicological profile, and established role in respiratory tract lesions and carcinogenicity in rodent models [11,13,16,25]. The carbon black concentration represents PM levels that frequently exceed air quality standards in deployed settings. Together, the combined exposure to carbon black and naphthalene was designed to more accurately model the complex airborne pollutant mixtures generated by open-air burn pits during military operations [2,10,11,16].
2.2. Biomarkers of Inflammation
Rats were exposed via whole-body inhalation to either HEPA-filtered air (sham control group), naphthalene (NA) vapor, or a mixture of carbon black and NA vapor (CBN) for six hours per day over three consecutive day In lung tissue, exposure to CBN induced a broad and statistically significant inflammatory response compared to both sham and NA groups (Figure 1). IL-1β levels were markedly elevated in the CBN group (67.68 ± 8.48 pg/mL) relative to NA (54.53 ± 6.04 pg/mL) and sham controls (47.27 ± 8.92 pg/mL). Interestingly, IL-6 levels were significantly decreased in both NA (34.48 ± 13.05 pg/mL) and CBN (40.85 ± 17.37 pg/mL) groups compared to sham animals (103.6 ± 21.72 pg/mL), suggesting a selective suppression of this cytokine. IFN-γ levels increased significantly in both NA (4.96 ± 0.87 pg/mL) and CBN (5.29 ± 0.98 pg/mL) groups compared to sham (2.40 ± 0.53 pg/mL), indicating enhanced immune activation. A similar pattern was observed for IL-4, which rose in NA (0.458 ± 0.169 pg/mL) and CBN (0.514 ± 0.094 pg/mL) groups compared to sham (0.278 ± 0.095 pg/mL).
Anti-inflammatory cytokine IL-10 in lung was also significantly elevated in NA (5.10 ± 0.74 pg/mL) and CBN (5.42 ± 0.99 pg/mL) groups relative to sham (3.19 ± 1.17 pg/mL). KC/GRO (CXCL1), a chemokine involved in neutrophil recruitment, showed the most dramatic increase, reaching 153.2 ± 32.5 pg/mL in the CBN group and 69.96 ± 17.72 pg/mL in the NA group, compared to 42.86 ± 12.13 pg/mL in sham controls. IL-5 levels were also elevated, with values of 18.22 ± 2.23 pg/mL and 14.01 ± 4.06 pg/mL in the CBN and NA groups, respectively, compared to 12.30 ± 1.06 pg/mL in the sham group. IL-13 levels increased significantly in both NA (1.20 pg/mL) and CBN (1.34 pg/mL) exposures relative to sham (0.54 pg/mL). Lastly, TNF-α was significantly elevated in the CBN group (3.51 ± 0.44 pg/mL) and to a lesser extent in the NA group (2.57 ± 0.49 pg/mL), compared to sham (2.38 ± 0.45 pg/mL). Collectively, these data demonstrate that CBN exposure leads to a pronounced and multifaceted inflammatory cytokine response in the lung, with greater effects than NA alone, supporting the utility of the CBN model for studying deployment-related respiratory inflammation.
In plasma, multiple pro-inflammatory cytokines were significantly elevated in the CBN-exposed group compared to sham controls, indicating systemic immune activation following inhalation exposure (Figure 2). IL-4 levels increased significantly in both NA (0.893 ± 0.302 pg/mL) and CBN (0.981 ± 0.232 pg/mL) groups compared to sham (0.043 ± 0.095 pg/mL). IL-6 concentrations were also markedly elevated, with CBN (28.11 ± 9.72 pg/mL) and NA (28.38 ± 9.72 pg/mL) groups both showing significant increases over sham (1.64 ± 9.72 pg/mL). Similarly, IL-10 levels rose substantially in the CBN group (7.25 ± 0.89 pg/mL) and NA group (5.54 ± 0.89 pg/mL) compared to sham (1.75 ± 0.89 pg/mL).
KC/GRO (CXCL1) was strongly induced by CBN exposure in plasma (39.21 ± 4.93 pg/mL) and elevated in NA animals (23.98 ± 4.93 pg/mL), both significantly higher than sham (7.06 ± 4.93 pg/mL;). TNF-α levels also increased in NA (4.76 ± 0.31 pg/mL) and CBN (3.64 ± 0.31 pg/mL) groups compared to sham (1.20 ± 0.31 pg/mL), with NA levels significantly higher than CBN, suggesting a more pronounced acute systemic response in the NA group. IL-1β, IL-5, and IL-13 were not detectable (NT) in plasma from sham animals but were measurable in NA and CBN groups; however, differences among groups did not reach statistical significance. The typical detection range for these cytokines in the MSD V-PLEX Rat Pro-inflammatory Panel is 0.61–2,500 pg/mL. IFN-γ levels showed no significant differences across groups. Collectively, these findings demonstrate a robust systemic inflammatory response to CBN exposures, with patterns of cytokine elevation distinct from those observed in lung tissue.
In brain tissue, several pro-inflammatory cytokines were significantly altered following NA and CBN exposures, indicating a neuroimmune response to inhaled toxicants (Figure 3). IL-1β levels showed no statistically significant differences among groups. However, IL-6 concentrations were significantly reduced in both NA (39.06 ± 4.15 pg/mL) and CBN (35.54 ± 4.15 pg/mL) groups compared to sham (67.05 ± 4.15 pg/mL), suggesting suppression of this cytokine in the brain. Similarly, IL-10 levels were markedly decreased in NA (5.01 ± 0.90 pg/mL) and CBN (5.89 ± 0.90 pg/mL) groups compared to sham (10.06 ± 0.90 pg/mL).
IL-4 levels in brain were also reduced by NA (0.43 ± 0.08 pg/mL) and CBN (0.52 ± 0.08 pg/mL) relative to sham (0.68 ± 0.08 pg/mL). IL-5 concentrations decreased significantly in NA (11.29 ± 1.20 pg/mL) and CBN (12.26 ± 1.20 pg/mL) groups compared to sham (18.69 ± 1.20 pg/mL). IL-13 levels were significantly lower in NA (0.78 ± 0.24 pg/mL) and CBN (0.37 ± 0.24 pg/mL) compared to sham (1.40 ± 0.24 pg/mL). Conversely, IFN-γ was significantly elevated in NA (6.29 ± 0.59 pg/mL) and CBN (6.41 ± 0.59 pg/mL) relative to sham (4.73 ± 0.59 pg/mL), indicating immune activation. KC/GRO (CXCL1) was significantly increased in both NA (2.23 ± 0.20 pg/mL) and CBN (2.14 ± 0.20 pg/mL) compared to sham (1.26 ± 0.20 pg/mL), reflecting enhanced chemokine signaling. TNF-α levels were markedly elevated in the NA group (0.68 ± 0.09 pg/mL) and were even higher in the CBN group (1.27 ± 0.09 pg/mL), both significantly increased compared to sham (0.27 ± 0.09 pg/mL), with CBN also significantly higher than NA.
Together, these results reveal that while NA and CBN exposures suppress several key anti-inflammatory and regulatory cytokines in the brain, they simultaneously enhance pro-inflammatory markers like IFN-γ, CXCL1, and TNF-α. These novel findings suggest a shift toward a neuroinflammatory phenotype. An overview of biomarker response for all tissues is summarized in Table 1.
2.3. Sample Preparation and Next-Generation Sequencing
RNA-seq analysis was conducted using the CBN and sham exposure groups. While NA exposure induced moderate changes in select cytokines, CBN exposure resulted in significant upregulation of a broader array of pro-inflammatory mediators across lung, plasma, and brain compartments. As the goal of transcriptomic profiling was to identify gene expression pathways strongly associated with response to surrogate burn pit exposure, the CBN group was selected to maximize biological signal and relevance to complex toxicant exposures. The sham group served as the reference for baseline expression. This approach enabled a focused and biologically meaningful comparison between high-response and unexposed animals.
RNA-seq analyses of CBN and sham exposed rat lung and brain tissues were used to gain unbiased insights into gene expression associated with the exposures to CBN. The results identified 278 differentially expressed genes (DEGs) in lung with FDR-adjusted p-values below 0.05 (Table S1, Column F). In brain, 19 DEGs were identified with FDR-adjusted p-values below 0.05 (Table S2, Column F). Multidimensional scaling plots visualizing tissue expression dissimilarity between the treatment groups based on the top 200 genes (determined by log2 fold-change) demonstrates the broad effects of CBN exposure compared to sham for lung (Figure 4A) and brain (Figure 4B). The top ten DEGs are labeled in volcano plots of lung (Figure 4C) and brain (Figure 4D), with significant DEGs shown in red (up-regulated in CBN exposed) or blue (down-regulated in CBN exposed). Heat maps demonstrate the 15 most significant DEGs, by FDR, in lung and brain, respectively, (Figure 4E and 4F). Biological relevance and function of the top 15 DEGs are summarized in Table 2.
2.4. Gene Set Enrichment Analysis
Quantitative changes obtained from RNA-seq were analyzed by Gene Set Enrichment Analysis (GSEA) using gene sets from the Molecular Signatures Hallmark, Kyoto Encyclopedia of Genes and Genomes (KEGG), and the SenMayo gene set for senescence-associated genes to identify molecular pathways that were significantly altered by CBN exposure [131,132,133,134]. The SenMayo gene set represents a validated transcriptional signature of cellular senescence derived from multi-tissue expression data and curated to distinguish senescent from non-senescent cells. Its inclusion enabled detection of senescence-related transcriptional programs not captured by conventional pathway databases, making it particularly useful for toxicant models where cellular stress and aging mechanisms may be engaged [134].
In lung, CBN exposure significantly activated a variety of pathways including myelocytomatosis proto-oncogene (Myc) version (v) 1 targets, early region 2 binding factor (E2F) transcription factor targets, cell cycle-related and chemokine signaling (Figure 5A). CBN exposure suppressed specific processes in lung, such as oxidative phosphorylation, Kirsten rat sarcoma viral oncogene homolog (KRAS) signaling down, and EMT (Figure 5A). In the brain, GSEA identified enhanced activity in processes involved with EMT, Wingless/Integrated (Wnt) β-catenin signaling, transforming growth factor (TGF) β signaling, and several inflammatory responses such as cytokine-cytokine receptor interaction signaling (Figure 5B). Processes suppressed in brain with CBN exposure include ribosome, spliceosome, and Myc targets v1.
A significant positive association was determined from the brain RNA-seq data using the SenMayo gene set (NES 1.47, q < 0.03), suggesting that gene expression associated with senescence was elevated due to CBN exposures in the brain (Figure 6). Among the senescence-associated genes upregulated in brain tissue following CBN exposure, three were highlighted for their roles in neuroinflammation, neurodegeneration, and blood-brain barrier (BBB) dysfunction. Interleukin 17 (IL-17), a pro-inflammatory cytokine, is known to contribute to BBB disruption, sustained neuroinflammation and promotes amyloid pathology [135]. AXL receptor tyrosine kinase (Axl) regulates microglial activation and is implicated in neurodegenerative signaling [136]. Endothelial cell-specific molecule 1 (Esm1), a mediator of vascular permeability, suggests BBB compromise [137]. These findings indicate that CBN exposure engages senescence-related transcriptional programs associated with brain vulnerability.
The SenMayo gene set was not significantly enriched in the lung, but several molecular pathways identified by GSEA showed consistent alterations in both lung and brain following CBN exposure. Gene sets significantly activated in both tissues included lysosome [KEGG: ko04142], complement [Hallmark], and endocytosis [KEGG: ko04144] (Figure 5) [138,139,140,141]. These pathways reflect shared biological responses: lysosome activation suggests enhanced clearance of cellular debris and toxic material; complement signaling indicates immune system engagement; and endocytosis reflects increased vesicular trafficking, likely in response to environmental insult. In contrast, oxidative phosphorylation [Hallmark, KEGG: ko00190] was suppressed in both tissues, consistent with impaired mitochondrial energy production and bioenergetic stress [142,143]. Together, these findings suggest a coordinated response to CBN exposure that includes immune activation, cellular adaptation, and mitochondrial dysfunction across lung and brain tissues. In the lung, biological process GO terms significantly enriched among DEGs (FDR p ≤ 0.05) further supported immune involvement, including cellular response to tumor necrosis factor, neutrophil and eosinophil chemotaxis, and chemokine-mediated signaling (Table S3). No significant biological process GO terms were associated with DEGs in the brain.
2.5. NF-κB Activation in Lung and Brain Following CBN Inhalation Exposure
NF-κB is a key transcription factor that regulates innate immune responses, inflammation, and cellular stress pathways. Inhaled environmental toxicants such as particulate matter and VOCs found in burn pit emissions activate NF-κB signaling, leading to nuclear translocation of the p65 subunit and transcription of downstream pro-inflammatory genes, including TNF-α, IL-1β, and KC/GRO (CXCL1). Given its upstream regulatory role, NF-κB serves as a mechanistically informative marker of toxicant-induced immune activation.
To assess NF-κB activation in response to inhalation exposure, we performed western blot analysis of nuclear extracts from lung and brain tissues. CBN exposure significantly increased nuclear NF-κB p65 levels in both tissues compared to sham controls (Figure 7). In the lung, NF-κB p65 abundance was significantly elevated in CBN-exposed animals (Figure 7A–B). This effect was even more pronounced in the brain, (Figure 7C–D).
These results align with the cytokine and RNA-seq analyses showing elevated inflammatory responses across tissues. Notably, GSEA revealed significant alteration of cytokine-mediated responses. Together, these findings confirm that NF-κB activation is a central feature of the lung-brain inflammatory axis following CBN exposure, supporting its utility as a biomarker for inhalation-related immune disruption and dysregulation.
2.6. Predictive Modeling Using Elastic Net
To identify a gene signature predictive of toxicant exposure, an elastic net regression model was implemented using the R computing environment, [144]. Elastic net was selected for its ability to handle high-dimensional, correlated transcriptomic data by combining L1 and L2 regularization, allowing for sparse but stable gene selection while minimizing overfitting. Compared to lasso, which produced overly sparse models, and ridge regression, which retained too many genes, elastic net offered a balanced solution suited to small-sample, high-feature datasets. Using the 278 differentially expressed genes (FDR < 0.05), the model selected a 97-gene subset predictive of exposure status. Leave-one-out cross-validation (LOO-CV) demonstrated 100% accuracy in distinguishing CBN-exposed from sham lung samples (Figure 8A–B), indicating the model’s strong classification performance and supporting its utility for molecular biomarker discovery.
Among the selected genes, several were particularly informative in characterizing the lung’s transcriptional response to CBN exposure. We highlight ten genes (IgV, Kcnq3, Sirpb3, Tgfbr1, Tm4sf19, St3gal2, Tiam1, Ccl9, Foxn4, and Tm4sf1) that emerged as the top predictors of exposure status (Figure 8B; Table 3). These genes represent key biological functions involved in immune regulation (IgV, Sirpb3, Ccl9, Tgfbr1, St3gal2), ion channel activity and neurogenic inflammation (Kcnq3), fibrosis and epithelial remodeling (Tgfbr1, Tm4sf19, Tiam1), and tissue repair or regenerative capacity (Foxn4, Tm4sf1). Each gene’s biological function and its potential impact on long-term lung health is summarized in Table 3, which highlights their roles in inflammation, oxidative stress, epithelial damage, and impaired regeneration hallmarks of lung injury [20,27,145]. The diversity of pathways represented in this multigene profile underscores the complex molecular landscape triggered by inhaled toxicants like CBN and reflects converging mechanisms that may drive persistent airway pathology.
In contrast to lung tissue, elastic net modeling using brain RNA-seq data did not yield a robust classifier. Only 19 genes in the brain met FDR significance thresholds (as compared to 278 in the lung), and model performance remained near chance (50–60%) despite parameter tuning.
3. Discussion
The findings expand upon our previous model where we demonstrated that whole-body inhalation of nano-sized CB induced systemic inflammatory responses across multiple tissues, including lung, plasma, artery, and brain. Trembley et al. used a single-component particulate exposure and focused on short-term protein-level changes [16]. The current study builds on that foundation by employing a more complex co-exposure to carbon black and naphthalene (CBN) to simulate the multifactorial composition of military burn pit emissions. Importantly, we incorporated transcriptomic profiling in both lung and brain tissues to inform broader and more mechanistically detailed molecular responses.
The data reveal tissue-specific inflammatory patterns across lung, plasma, and brain following CBN exposures. In the lung and plasma, exposure induced robust increases in pro-inflammatory cytokines including IL-4, IL-10, KC/GRO (CXCL1), and TNF-α. Notably, IL-6 was significantly elevated in plasma but suppressed in lung tissue. In contrast, the brain exhibited a divergent cytokine profile characterized by reductions in IL-4, IL-5, IL-6, IL-10, and IL-13, while IFN-γ and KC/GRO were significantly elevated. TNF-α was also markedly increased in the brain, especially in the CBN group. These distinct inflammatory profiles across tissues underscore the presence of a compartmentalized yet interconnected response, supporting the hypothesis of a functional lung–brain axis in the context of complex inhalational exposures. Increased NF-κB nuclear translocation (~2-fold in lung and >3-fold in brain), supports its central role in mediating systemic inflammatory signaling.
Lung transcriptomic analysis showed broad upregulation of genes involved in immune activation, oxidative stress, and epithelial remodeling. Notable genes included Sirpb3, Tgfbr1, and Tm4sf19 (fibrosis and immune regulation), Kcnq3 (sensory modulation), St3gal2 and Tiam1 (cell migration and glycosylation), and Ccl9, Foxn4, Tm4sf1, and IgV gene segments (adaptive immunity and epithelial repair). Elastic net modeling identified a subset of genes as highly predictive of CBN exposure, forming a distinct lung-specific molecular signature of injury. Gene set enrichment analysis (GSEA) corroborated these findings, with enrichment in chemokine signaling, E2F targets, and suppressed oxidative phosphorylation, indicating active immune responses and mitochondrial stress.
In the brain, transcriptomic changes included enrichment of senescence-associated gene sets (SenMayo), Wnt/β-catenin and TGF-β signaling, and suppression of oxidative phosphorylation. Upregulated genes such as Hist1h4m, Zbtb20, Cyp7b1, and FAM107A were associated with epigenetic modulation, neuroinflammation, and lipid metabolism, while downregulated genes like Chchd4 and Pom121L2 implicated impaired mitochondrial and nucleocytoplasmic function. The enrichment of senescence-associated transcripts exclusively in the brain suggests a lower threshold for senescence induction or reduced regenerative capacity compared to the lung. This may contribute to long-term neurological consequences following toxicant exposure.
These findings align with and extend multiple prior models of DRRD. Hathaway et al. demonstrated mitochondrial collapse and oxidative injury in response to carbon black and ozone [145]. Vance et al. showed phase-dependent respiratory suppression from simulated burn pit emissions but did not investigate transcriptional changes [167]. Our model expands these studies by incorporating both volatile chemical and particulate components and revealing molecular evidence for injury in both lung and brain tissues.
Our data also resonate with Gutor et al. (2022), which reported immune-driven airway remodeling in deployment-associated constrictive bronchiolitis, and Gutor et al. (2024), which described a sulfur dioxide (SO₂) exposure model of post-deployment respiratory syndrome (PDRS) [20,168,169]. The latter study revealed pulmonary vascular remodeling and oxidative injury mediated through the ROS–isoLG–SIRT3–SOD2 axis [169]. While Gutor et al. emphasized vascular pathology and pulmonary hypertension, our CBN model demonstrates overlapping pathways, including Noxo1, Tgfbr1, and NF-κB activation, but uniquely captures CNS involvement, highlighting the added value of a multi-tissue toxicology framework. The combined chemical and PM insults of CBN may induce broader systemic effects than SO₂ alone. These models are synthesized in the review by Wang et al. (2023), which calls for harmonized inhalation platforms that more accurately reflect deployment-related exposures [9]. Our study contributes to this effort by integrating chemical and PM toxicants complexity, advanced transcriptomic modeling, and neuroinflammatory endpoints.
Epidemiologic studies have consistently linked burn pit exposures to increased risks of psychiatric conditions, cognitive impairment, and neurodegenerative disease [4,5,8,170,171]. Our observation of elevated TNF-α in both plasma and brain, alongside senescence-linked transcriptional changes, supports the hypothesis that the lung-derived inflammation can disrupt the blood–brain barrier and trigger neuroinflammatory cascades and injury. These immune signals may activate microglia and alter neuronal homeostasis, contributing to the burden of neuropsychiatric disease in exposed Veterans [4,5,8,170,171].
This study has defined biologically coherent alterations in molecular pathways in lung and brain, providing strong evidence for a systemic inflammatory axis following inhalation of burn pit–like emissions. By integrating elastic net modeling, transcriptomic profiling, and cytokine measurement across multiple tissues, our model offers a mechanistic framework for early detection and investigation into DRRDs and neurological diseases following military burn pit exposures. These findings support the expansion of toxicological assessments to include CNS outcomes and provide a foundation for developing clinical diagnostics and therapeutics that address both pulmonary and neuropsychiatric risks in exposed populations.
3.1. Limitations
This study provides valuable insights into the early inflammatory and transcriptional changes induced by surrogate military burn pit exposures. However, several limitations should be acknowledged. First, the exposure protocol used a short-term, subacute model consisting of three 6-hour inhalation sessions over three days. While this design simulates an acute deployment-related exposure, it does not capture the chronic, cumulative nature of military burn pit exposure experienced over many months or years. Longitudinal studies are needed to determine whether these acute molecular signatures persist or evolve with repeated exposures over time.
Second, although our exposure model incorporates two major toxic components found in burn pit emissions, carbon black (as a particulate matter surrogate) and naphthalene (representing VOCs and PAHs), real-world burn pit smoke consists of a far more complex mixture of chemicals, including dioxins, furans, and heavy metals. As such, our CBN mixture does not fully recapitulate the chemical diversity and additive toxicity of actual burn pit emissions. A significant translational challenge remains the inherent complexity and variability of burn pit toxicant mixtures and exposure durations, which may never be fully replicated experimentally. Expanding preclinical models to include diverse exposure mixtures and administration routes (whole-body inhalation, intranasal, oral) will enhance translational relevance and capture real-world exposure variability.
Third, only male rats were used in this study to reduce variability, however sex-specific differences in immune and neuroinflammatory responses are well documented [172]. Inclusion of both sexes in future experiments will be essential for understanding sex-based vulnerability to burn pit-associated diseases.
Fourth, the transcriptomic profiling via RNA-seq was limited to a comparison between CBN-exposed and sham animals, excluding the naphthalene-only (NA) group. While this was a data-driven decision based on cytokine profiling (which showed that CBN induced a more robust and distinct inflammatory signature), it limits the ability to distinguish additive or synergistic effects of particulate and volatile toxicants.
Lastly, while the elastic net modeling demonstrated strong discriminatory power in identifying CBN exposure-related gene signatures, external validation in independent cohorts including human data is necessary before these biomarkers can be translated into diagnostic tools. Similarly, additional studies examining the functional consequences and changes in pulmonary physiology in response to differentially expressed genes will strengthen the biological interpretation of the predictive signatures.
4. Materials and Methods
4.1. Experimental Animals
Male Sprague Dawley rats were obtained from Hilltop Laboratories (Scottdale, PA) and housed in an AAALAC approved facility at West Virginia University (WVU).
4.2. Ethics
All procedures were approved by the WVU Institutional Animal Care and Use Committee (protocol 1602000621) and conformed to the most current National Institutes of Health (NIH) Guidelines for the Care and Use of Laboratory Animals. Housing conditions included 12:12 h light:dark cycle, 20–26 °C, 30–70% relative humidity, acclimatization for 48–72 h prior to any procedure, and ad libitum access to food and water.
4.3. Whole Body Inhalation Exposure
Rats (sham-control group: age 53 ± 1 days; mass 288 ± 3 g; CBN group: age 53 ± 1 days; mass 303 ± 4 g) were exposed to either HEPA filtered air or a mixture of carbon black (target concentration = 6 mg/m3) and naphthalene vapor (target concentration = 10 PPM) for 6 h/day for 3 consecutive days (n = 10/group). A carbon black-only group was not included in this study, as those exposures had been tested in prior experiments [16]. Rats were randomly assigned to either control or experimental groups and were individually housed in cages within a stainless-steel chamber during exposure. The exposure chamber measured 22 x 20 x 20 inches (width x depth x height) with an approximate volume of 144 L. The total airflow through the chamber was approximately 28 LPM during exposures. Bedding material in the bottom of the chamber was soaked with 50 mL of water to provide humidity during exposures. Humidity (30–70%) and temperature (20–26 °C) levels were maintained throughout each exposure period to provide animal comfort.
4.4. Preparation of Naphthalene Vapor and Carbon Black Aerosols
Bulk CB powder (Printex 90®, composed of 99.9% carbon) was obtained as a gift from Evonik (Frankfurt, Germany) and naphthalene crystals (99% purity, Sigma-Aldrich, St. Louise, MO) were acquired. Before each toxicant exposure, thirty grams of naphthalene were placed into a glass beaker with input and output flow ports. The beaker was placed onto a hot plate set to 60 °C and heated for 30 minutes prior to exposure to begin sublimation of the naphthalene into a vapor. After the exposure began, a mass flow controller (MFC) regulated the airflow through the beaker and pushed the naphthalene gas through the system. Fifty grams of CB powder was placed into a high-pressure acoustical generator (HPAG, IEStechno, Morgantown, WV) which aerosolized and de-agglomerated the CB powder. A mass flow controller pushed the aerosolized CB out of the generator and into a Venturi pump (JS-60 M, Vaccon, Medway, MA) to further de-agglomerate the particles. The CB aerosols were then mixed with the naphthalene vapor before being passed into the inhalation exposure chamber (Cube 150, IEStechno, Morgantown, WV). Multiple instruments sampled continuously from the chamber to provide real-time, accurate measurements of the aerosol and gas levels. A light scattering device (PDR-1500, Thermo Environmental Instruments Inc., Franklin, MA) was utilized to determine the aerosol mass concentration and a photo ionization detector (MiniRAE 3000, RAE Systems, Sunnyvale, CA) measured the levels of naphthalene vapor within the chamber. Stable concentration levels were maintained in real-time via software feedback loops. The aerosol mass concentration was adjusted by varying the sound energy in the acoustical generator and the naphthalene vapor level was regulated by varying the airflow through the glass beaker. 37 mm PTFE filters were used for gravimetric measures concurrent with the PDR-1500 measures to obtain a calibration factor. These gravimetric measures were also considered the “gold standard” for the mass concentration measurements, and these levels are reported in the Results section. The MiniRAE 3000 was calibrated daily with isobutylene and a correction factor was applied to get the correct naphthalene levels. The aerosol and vapor leaving the exposure chamber was HEPA and charcoal filtered before entering the house exhaust. The particle count size distribution of the aerosol was measured using a high resolution electrical low-pressure impactor (ELPI, Dekati, Tampere, Finland).
4.5. Tissue Collection and Processing
Animal tissue harvesting was performed 24 h after the final exposure. Euthanasia of the rats was performed via exsanguination under deep anesthesia (5% induction, 2% maintenance with isoflurane gas) followed by organ removal. Plasma aliquots and whole tissues (brain and lung) were snap frozen in liquid nitrogen and stored at −80 °C. Solid tissues were pulverized on dry ice to granular homogenous powder.
4.6. Inflammation Panel
Plasma samples were diluted in assay buffer, and pulverized brain and lung tissues were homogenized in RIPA buffer (Thermo Fisher Scientific, Waltham, MA). Total protein concentrations in tissue lysates were determined using a Direct Detect spectrophotometer (MilliporeSigma, Burlington, MA). Pro-inflammatory cytokines were quantified using the Meso Scale Discovery (MSD) V-PLEX Rat Pro-inflammatory Panel 2 Kit (K15059G; MSD, Rockville, MD, USA), which utilizes a multiplexed immunoassay based on electrochemiluminescence (ECL) detection technology. Concentrations of IFN-γ, IL-1β, IL-4, IL-5, IL-6, IL-10, IL-13, KC/GRO, and TNF-α were measured in brain and lung tissue lysates (50 µg total protein) and diluted plasma samples (25 µL). Data were analyzed using MSD Discovery Workbench 4.0 software [16].
4.7. Western Blot Analysis
Western blotting was used to detect and quantify nuclear fractions isolated using a nuclear extraction reagent (NE-PER; Thermo Scientific, Waltham, MA, USA). Protein concentrations were determined using an infrared spectrometer (Direct Detect; Millipore, Burlington, MA, USA). 25 µg of protein was loaded onto 4-20% precast Tris-Glycine gels (Bio-Rad, Hercules, CA, USA). The protein was transferred using the Trans-Blot Turbo system (Bio-Rad, Hercules, CA, USA). Total protein was stained using LI-COR Revert 700 stain (LI-COR, Lincoln, NE, USA) and imaged using the LI-COR Odyssey FC imager (LI-COR, Lincoln, NE, USA). Blocking was performed using Intercept Blocking Buffer (LI-COR, Lincoln, NE, USA). NF-κB primary antibody was used at 1:1000 dilution (Cell Signaling Technology 6959; Danvers, MA, USA) with incubation performed overnight at 4 °C with agitation. Secondary antibody was used at 1:20,000 dilution for a concentration of 1.0 × 10−1 µg/mL (goat anti-mouse LI-COR 926-32210; LI-COR, Lincoln, NE, USA). Densitometry was performed using Image Studio software (LI-COR, Lincoln, NE, USA).
4.8. Data Analysis
GraphPad Prism 10.2.3 was used for the biomarker and immunoblot data calculations and bar plot visualizations (GraphPad, San Diego, CA, USA). Descriptive statistics are provided for CBN aerosol characterization. MSD data were analyzed using ordinary one-way ANOVA followed by Holm-Šídák’s multiple comparisons test (n = 5–10 per group). The differences between sham-control and CBN exposed groups for western blot data were identified using unpaired t-test or Mann–Whitney tests according to data distribution (n = 8 per group). Significance was established as p-value ≤ 0.05 providing strong evidence against the null hypothesis.
4.9. RNA Sequencing
Tissue and resulting data were processed using a custom pipeline and normalized as previously described [173]. Briefly, the RNA was isolated from lung and brain tissues by the University of Minnesota Genomics Center using a commercial kit (RNeasy Plus Universal kit, Qiagen, Germantown, MD) according to manufacturer’s instructions. RNA was quantified using a RiboGreen assay (Thermo Fisher, Waltham, MA), and the quality was assessed by capillary electrophoresis using a Tapestation (Agilent, Santa Clara, CA). All samples had an RNA integrity number (RIN) ≥ 8. Libraries were prepared using an Illumina TruSeq Stranded Total RNA Library Prep Kit (San Diego, CA), and sequencing was performed using a Novaseq 6000 (Illumina) to generate 150 bp read pairs. Read quality control, trimming, and alignment were performed separately for lung and brain tissues using a custom analysis pipeline (Pipeline for UMII/RI RNASeq Analysis), developed and maintained by the Research Informatics (RI) group at the Minnesota Supercomputing Institute (MSI). Adaptor sequences were trimmed from raw reads using Trimmomatic (v0.33), and the quality of raw and trimmed reads was assessed with FastQC (v0.11.7). Ribosomal RNA (rRNA) contamination, which was low in all samples (≤ 1%), was quantified by aligning a subsample of 10,000 reads pairs from each sample to rRNA sequences from the SILVA database (release 132). Trimmed reads were aligned to the Rattus norvegicus reference genome (mRatBN7.2, Ensembl release 109) using HISAT2 (v2.1.0). Reads with mapping quality < 60 were removed. Raw counts were generated using the featureCounts tool from the Subread package (v1.6.2).
4.10. Differential Gene Expression Analysis
For each tissue, raw counts were imported into R (v4.3.0). Genes shorter than 200 bp or with very low expression (< 1 CPM in at least 50% of samples) were pruned from the count matrix before differential expression testing. Pairwise differential expression testing between CBN-exposed and sham groups was performed separately for brain and lung tissues using a likelihood ratio (LR) test or quasi-likelihood F (QLF)-test, respectively, using EdgeR (v4.0.5). Genes with a false discovery rate (FDR)-adjusted p-value ≤ 0.05 were considered significantly differentially expressed genes (DEGs, Supplemental Tables S1 and S2).
4.11. Gene Set Enrichment Analysis
Expressed genes from brain or lung tissues were ranked by multiplying -log10 (p-value) by the sign of the fold-change from differential expression testing, where higher ranked genes have higher expression in CBN-exposed than sham tissues and lower ranked genes have lower expression in CBN-exposed than sham tissues. Gene set enrichment analysis (GSEA) using the GSEA function from the clusterProfiler R package (v4.10.0) was performed using the pre-ranked genes and combined gene sets that included rat Hallmark gene sets (MSigDB), the SenMayo gene set with human gene symbols mapped to rat symbols (MSigDB), and KEGG pathways. Hallmark and KEGG pathways were not included in the GSEA if they were not applicable to brain or lung tissues. For example, the Alzheimer disease KEGG pathway was removed for lung, and the Hallmark bile acid metabolism pathway was removed for both tissues. For all GSEA, only gene sets with at least 10 genes and at most 500 genes were analyzed. Overrepresentation analysis with all biological process gene ontology (GO) terms was performed separately for both tissues using the topGO R package (v. 2.58.0) with the default weight01 algorithm. Gene sets and GO terms with q-value ≤ 0.05 were considered significantly enriched/overrepresented. GSEA summary plots were created using the ggplot2 R package (v 3.5.2), heatmaps were produced using the pheatmap R package (v 1.0.12), and individual pathway enrichment plots were produced using the R package enrichplot (v 1.24.2).
4.12. Predictive Modeling Using Elastic Net
To build a predictive model of CBN exposure we extended our analysis to include computational methods to identify predictors that remain associated with exposure after accounting for correlations with the expression of other genes. To achieve this, a penalized regression model, which adds a penalty for models with more predictors, was applied to the lung tissue expression data for 278 differentially expressed genes (FDR p-value ≤ 0.05) [174]. Penalized regression helps select a subset of the most predictive genes, in this case genes predictive of CBN exposure, by reducing the influence of less relevant ones [175]. Different methods have been developed to exclude or down-weight the importance of genes to produce a robust, predictive signature that depends on a smaller subset of genes. We tested several methods, including lasso, elastic net, and ridge regression, with lasso being the most conservative and ridge regression the least conservative [176]. Elastic net, which selected 97 genes, was chosen as a balanced option given the small sample size. An extended explanation of this model is included in the Supplemental Methods.
4.13. Availability of Data and Materials
Most data generated or analyzed during this study are included in this published article [and its supplementary information files]. Raw fastq files and processed data files containing raw counts and trimmed mean of M-values (TMM) normalized counts per million (CPM) and log2 CPM values (produced by EdgeR) for all samples are available through the Gene Expression Omnibus (GEO) accession number GSE281993. Other data is available from the corresponding author on reasonable request.
5. Conclusions
This study highlights key biological mechanisms linking inhalational toxicants from burn pit exposures to systemic inflammation and multi-organ dysfunction. By characterizing the potential molecular and immune signatures in both lung and brain tissues, our findings contribute to efforts to develop toxicological risk assessments, enable earlier diagnosis, and support targeted care for Veterans. These insights align with broader public health goals to mitigate the long-term effects of air pollution and occupational exposures [27,145,177].
Incorporating digital biomarkers, non-invasive imaging technologies, and integrated diagnostics represents a long-term translational goal aimed at enhancing our ability to detect, monitor, and understand burn pit-related health effects. These advanced approaches are not yet widely implemented but hold promise as scalable, non-invasive tools to assess physiological and inflammatory responses in affected populations [2,18,178]. Embedding such innovations within a broader community of care model uniting basic scientists, environmental toxicologists, clinicians, and the Veteran communities we serve ensures that the scientific discovery is rapidly translated into meaningful, accessible healthcare solutions [2,178,179]. This model reinforces the value of interdisciplinary collaboration to address the complexity of environmental toxicant exposures [2,18,178,179].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Lung_EdgeR_Likelihood_Ratio_Test_Results; Table S2: Brain_EdgeR_Likelihood_Ratio_Test_Results; Table S3: Lung_topGO_significantly_enriched_BP_GO_Terms; Supplementary Methods: Statistical model for selecting a predictive gene signature of CBN exposure.
Author Contributions
JHT, TAB, and BJS wrote the original draft. BJS and HLW contributed to the conceptualization of the predictive modeling. JHT and TAB contributed to the overall conceptualization, planning, sample processing, and manuscript editing. HLW performed the predictive modeling and associated manuscript writing. AH conducted the raw sequence data, differential gene expression, and GSEA analysis, and contributed to the transcriptomic data interpretation, figure preparation and editing of the manuscript. TG and TRN contributed to the conceptualization, whole-body exposure experiments, tissue acquisition, manuscript review and editing. CHW, JPN, and PB provided scientific advice and assisted with manuscript review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the following sources: U.S. Department of Veterans Affairs, Veterans Health Administration funding through Biomedical Laboratory Research and Development Service (I01BX004146, TAB); Clinical Science Research and Development Service (I01CX002701, JHT); Military Exposures Research Program Field Planning Award (TAB, JHT); Rehabilitation Research and Development Service (A4836-C, HLW, RECOVER; U.S. National Institutes of Health WV-CTSI U54 GM104942 (TRN); University of Minnesota Genomic Center (UMGC) Pilot Grant (TB and JHT).
Institutional Review Board Statement
All animal procedures were approved by the West Virginia University (WVU) Institutional Animal Care and Use Committee (protocol number 1602000621, approval March 1, 2018 and renewed on March 1, 2024). Animal procedures were conducted in accordance with the most recent National Institutes of Health (NIH) Guidelines for the Care and Use of Laboratory Animals. No human subjects work was performed or included.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
We would like to acknowledge the Burn Pits 360 Veteran Nonprofit Advocacy Group and their Scientific Advisory Committee, led by Dr. Julie Tomaska and co-founder Rosie Lopez Torres, for their valuable guidance, including providing critical insights on the first-hand experiences and impacts of burn pit exposures on U.S. Veterans. The authors acknowledge the University of Minnesota Genomics Center (UMGC) and the University of Minnesota Supercomputing Institute (MSI) for providing resources that contributed to the research results reported in this paper. We also thank Drs. Todd Knutson and Sarah Munro at MSI for their expert guidance in RNA-seq analysis. Additional thanks to Simon So for his technical assistance.
Conflicts of Interest
All authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results. Dr. Tammy Butterick discloses her role as Co-Chair of the Scientific Advisory Board for the Burn Pits 360 Veteran Nonprofit Advocacy Group. This position is unpaid, voluntary, and does not constitute a financial conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AD | Alzheimer’s disease |
| BP GO | Biological process gene ontology |
| CB | Carbon black powder |
| CBN | Carbon black nanoparticles and naphthalene vapor |
| ConB | Constrictive bronchiolitis |
| DEGs | Differentially expressed genes |
| DRRD | Deployment-Related Respiratory Diseases |
| ELP | Electrical low-pressure impactor |
| EMT | Epithelial to mesenchymal transition |
| FDR | False discovery rate |
| GEO | Gene Expression Omnibus |
| GSEA | Gene Set Enrichment Analysis |
| HPAG | High-pressure acoustical generator |
| LOO | Leave-one-out |
| LR | Likelihood ratio |
| MFC | Mass flow controller |
| MSD | Meso Scale Discovery |
| MSI | Minnesota Supercomputing Institute |
| NIH | National Institutes of Health |
| PD | Parkinson’s disease |
| PM | Particulate matter |
| PM2.5 | Particulate matter < 2.5 microns |
| QLF | Quasi-likelihood F |
| RI | Research Informatics |
| RIN | RNA integrity number |
| RNA-seq | RNA sequencing |
| rRNA | Ribosomal RNA |
| VOCs | Volatile organic compounds |
References
- Hirzel, K.L.; Balmer, J. Airborne Hazards and Open Burn Pit Exposures. Workplace Health Saf 2023, 71, 352. [CrossRef]
- Trembley, J.H.; Barach, P.; Tomaska, J.M.; Poole, J.T.; Ginex, P.K.; Miller, R.F.; Lindheimer, J.B.; Szema, A.M.; Gandy, K.; Siddharthan, T.; et al. Current understanding of the impact of United States military airborne hazards and burn pit exposures on respiratory health. Part Fibre Toxicol 2024, 21, 43. [CrossRef]
- National Academies, S.; Engineering; Health; Medicine, D.; Board on Population, H.; Public Health, P.; Committee on the Respiratory Health Effects of Airborne Hazards Exposures in the Southwest Asia Theater of Military, O. Respiratory Health Effects of Airborne Hazards Exposures in the Southwest Asia Theater of Military Operations 2020. [CrossRef]
- Hoisington, A.J.; Stearns-Yoder, K.A.; Kovacs, E.J.; Postolache, T.T.; Brenner, L.A. Airborne Exposure to Pollutants and Mental Health: A Review with Implications for United States Veterans. Curr Environ Health Rep 2024, 11, 168-183. [CrossRef]
- Penuelas, V.L.; Lo, D.D. Burn pit exposure in military personnel and the potential resulting lung and neurological pathologies. Frontiers in Environmental Health 2024, 3. [CrossRef]
- Perveen, M.M.; Mayo-Malasky, H.E.; Lee-Wong, M.F.; Tomaska, J.M.; Forsyth, E.; Gravely, A.; Klein, M.A.; Trembley, J.H.; Butterick, T.A.; Promisloff, R.A.; et al. Gross Hematuria and Lower Urinary Tract Symptoms Associated With Military Burn Pits Exposures in US Veterans Deployed to Iraq and Afghanistan. J Occup Environ Med 2023, 65, 740-744. [CrossRef]
- Savitz, D.A.; Woskie, S.R.; Bello, A.; Gaither, R.; Gasper, J.; Jiang, L.; Rennix, C.; Wellenius, G.A.; Trivedi, A.N. Deployment to Military Bases With Open Burn Pits and Respiratory and Cardiovascular Disease. JAMA Netw Open 2024, 7, e247629. [CrossRef]
- Brooks, A.W.; Sandri, B.J.; Nixon, J.P.; Nurkiewicz, T.R.; Barach, P.; Trembley, J.H.; Butterick, T.A. Neuroinflammation and Brain Health Risks in Veterans Exposed to Burn Pit Toxins. Int J Mol Sci 2024, 25, 9759. [CrossRef]
- Wang, X.; Doherty, T.A.; James, C. Military burn pit exposure and airway disease: Implications for our Veteran population. Ann Allergy Asthma Immunol 2023, 131, 720-725. [CrossRef]
- Masiol, M.; Mallon, C.T.; Haines, K.M., Jr.; Utell, M.J.; Hopke, P.K. Airborne Dioxins, Furans, and Polycyclic Aromatic Hydrocarbons Exposure to Military Personnel in Iraq. J Occup Environ Med 2016, 58, S22-30. [CrossRef]
- Woodall, B.D.; Yamamoto, D.P.; Gullett, B.K.; Touati, A. Emissions from small-scale burns of simulated deployed U.S. military waste. Environ Sci Technol 2012, 46, 10997-11003. [CrossRef]
- Aurell, J.; Gullett, B.K.; Yamamoto, D. Emissions from open burning of simulated military waste from forward operating bases. Environ Sci Technol 2012, 46, 11004-11012. [CrossRef]
- Blasch, K.W.; Kolivosky, J.E.; Heller, J.M. Environmental Air Sampling Near Burn Pit and Incinerator Operations at Bagram Airfield, Afghanistan. J Occup Environ Med 2016, 58, S38-43. [CrossRef]
- Engelbrecht, J.P.; McDonald, E.V.; Gillies, J.A.; Jayanty, R.K.; Casuccio, G.; Gertler, A.W. Characterizing mineral dusts and other aerosols from the Middle East--Part 2: grab samples and re-suspensions. Inhal Toxicol 2009, 21, 327-336. [CrossRef]
- Falvo, M.J.; Sotolongo, A.M.; Osterholzer, J.J.; Robertson, M.W.; Kazerooni, E.A.; Amorosa, J.K.; Garshick, E.; Jones, K.D.; Galvin, J.R.; Kreiss, K.; et al. Consensus Statements on Deployment-Related Respiratory Disease, Inclusive of Constrictive Bronchiolitis: A Modified Delphi Study. Chest 2023, 163, 599-609. [CrossRef]
- Trembley, J.H.; So, S.W.; Nixon, J.P.; Bowdridge, E.C.; Garner, K.L.; Griffith, J.; Engles, K.J.; Batchelor, T.P.; Goldsmith, W.T.; Tomaska, J.M.; et al. Whole-body inhalation of nano-sized carbon black: a surrogate model of military burn pit exposure. BMC Res Notes 2022, 15, 275. [CrossRef]
- Block, M.L.; Calderon-Garciduenas, L. Air pollution: mechanisms of neuroinflammation and CNS disease. Trends Neurosci 2009, 32, 506-516. [CrossRef]
- Hoisington, L.; Lowry, C.A.; McDonald, L.T.; Krefft, S.D.; Rose, C.S.; Kovacs, E.J.; Brenner, L.A. First Annual PACT Act Research Symposium on Veterans Health: A Colorado PACT Act Collaboration (CoPAC) Initiative. Mil Med 2024, 189, 80-84. [CrossRef]
- Heusinkveld, H.J.; Wahle, T.; Campbell, A.; Westerink, R.H.S.; Tran, L.; Johnston, H.; Stone, V.; Cassee, F.R.; Schins, R.P.F. Neurodegenerative and neurological disorders by small inhaled particles. Neurotoxicology 2016, 56, 94-106. [CrossRef]
- Gutor, S.S.; Richmond, B.W.; Du, R.H.; Wu, P.; Lee, J.W.; Ware, L.B.; Shaver, C.M.; Novitskiy, S.V.; Johnson, J.E.; Newman, J.H.; et al. Characterization of Immunopathology and Small Airway Remodeling in Constrictive Bronchiolitis. Am J Respir Crit Care Med 2022, 206, 260-270. [CrossRef]
- Trageser, K.J.; Sebastian-Valverde, M.; Naughton, S.X.; Pasinetti, G.M. The Innate Immune System and Inflammatory Priming: Potential Mechanistic Factors in Mood Disorders and Gulf War Illness. Front Psychiatry 2020, 11, 704. [CrossRef]
- Garshick, E.; Abraham, J.H.; Baird, C.P.; Ciminera, P.; Downey, G.P.; Falvo, M.J.; Hart, J.E.; Jackson, D.A.; Jerrett, M.; Kuschner, W.; et al. Respiratory Health after Military Service in Southwest Asia and Afghanistan. An Official American Thoracic Society Workshop Report. Ann Am Thorac Soc 2019, 16, e1-e16. [CrossRef]
- Nurkiewicz, T.R.; Porter, D.W.; Hubbs, A.F.; Stone, S.; Chen, B.T.; Frazer, D.G.; Boegehold, M.A.; Castranova, V. Pulmonary nanoparticle exposure disrupts systemic microvascular nitric oxide signaling. Toxicol Sci 2009, 110, 191-203. [CrossRef]
- Command, U.S.A.P.H. Screening Health Risk Assessments, Joint Base Balad, Iraq, 11 May–19 June 2009; U.S. Army Center for Health Promotion and Preventive Medicine: Aberdeen Proving Ground, MD, July 2010.
- Buckpitt, A.; Boland, B.; Isbell, M.; Morin, D.; Shultz, M.; Baldwin, R.; Chan, K.; Karlsson, A.; Lin, C.; Taff, A.; et al. Naphthalene-induced respiratory tract toxicity: metabolic mechanisms of toxicity. Drug Metab Rev 2002, 34, 791-820. [CrossRef]
- Ganter, M.T.; Roux, J.; Miyazawa, B.; Howard, M.; Frank, J.A.; Su, G.; Sheppard, D.; Violette, S.M.; Weinreb, P.H.; Horan, G.S.; et al. Interleukin-1β Causes Acute Lung Injury via αvβ5 and αvβ6 Integrin–Dependent Mechanisms. Circulation Research 2008, 102, 804-812, doi:doi:10.1161/CIRCRESAHA.107.161067.
- Hill, W.; Lim, E.L.; Weeden, C.E.; Lee, C.; Augustine, M.; Chen, K.; Kuan, F.C.; Marongiu, F.; Evans, E.J., Jr.; Moore, D.A.; et al. Lung adenocarcinoma promotion by air pollutants. Nature 2023, 616, 159-167. [CrossRef]
- Piao, C.H.; Fan, Y.; Nguyen, T.V.; Song, C.H.; Kim, H.T.; Chai, O.H. PM2.5 exposure regulates Th1/Th2/Th17 cytokine production through NF-κB signaling in combined allergic rhinitis and asthma syndrome. Int Immunopharmacol 2023, 119, 110254. [CrossRef]
- Li, Y.; Liu, Y.; Liang, Z.; Jia, S.; Liu, Y.; Li, J.; Liu, J.; Shi, J.; Yuan, P.; Zhu, J.; et al. Plasma IL4 Levels Linked to Pulmonary Hypertension Severity and Outcome. Clin Exp Pharmacol Physiol 2025, 52, e70040. [CrossRef]
- Wang, H.; Tumes, D.J.; Hercus, T.R.; Yip, K.H.; Aloe, C.; Vlahos, R.; Lopez, A.F.; Wilson, N.; Owczarek, C.M.; Bozinovski, S. Blocking the human common beta subunit of the GM-CSF, IL-5 and IL-3 receptors markedly reduces hyperinflammation in ARDS models. Cell Death Dis 2022, 13, 137. [CrossRef]
- Liva, S.M.; de Vellis, J. IL-5 induces proliferation and activation of microglia via an unknown receptor. Neurochem Res 2001, 26, 629-637. [CrossRef]
- Voiriot, G.; Razazi, K.; Amsellem, V.; Tran Van Nhieu, J.; Abid, S.; Adnot, S.; Mekontso Dessap, A.; Maitre, B. Interleukin-6 displays lung anti-inflammatory properties and exerts protective hemodynamic effects in a double-hit murine acute lung injury. Respiratory Research 2017, 18, 64. [CrossRef]
- Kopf, M.; Baumann, H.; Freer, G.; Freudenberg, M.; Lamers, M.; Kishimoto, T.; Zinkernagel, R.; Bluethmann, H.; Köhler, G. Impaired immune and acute-phase responses in interleukin-6-deficient mice. Nature 1994, 368, 339-342. [CrossRef]
- Duque Ede, A.; Munhoz, C.D. The Pro-inflammatory Effects of Glucocorticoids in the Brain. Front Endocrinol (Lausanne) 2016, 7, 78. [CrossRef]
- Frieler, R.A.; Meng, H.; Duan, S.Z.; Berger, S.; Schütz, G.; He, Y.; Xi, G.; Wang, M.M.; Mortensen, R.M. Myeloid-specific deletion of the mineralocorticoid receptor reduces infarct volume and alters inflammation during cerebral ischemia. Stroke 2011, 42, 179-185. [CrossRef]
- Ware, L.B.; Matthay, M.A. The acute respiratory distress syndrome. N Engl J Med 2000, 342, 1334-1349. [CrossRef]
- Liu, X.; Qian, X.; Xing, J.; Wang, J.; Sun, Y.; Wang, Q.g.; Li, H. Particulate Matter Triggers Depressive-Like Response Associated With Modulation of Inflammatory Cytokine Homeostasis and Brain-Derived Neurotrophic Factor Signaling Pathway in Mice. Toxicological Sciences 2018, 164, 278-288. [CrossRef]
- Brombacher, T.M.; Nono, J.K.; De Gouveia, K.S.; Makena, N.; Darby, M.; Womersley, J.; Tamgue, O.; Brombacher, F. IL-13-Mediated Regulation of Learning and Memory. J Immunol 2017, 198, 2681-2688. [CrossRef]
- Hershey, G.K.K. IL-13 receptors and signaling pathways: An evolving web. Journal of Allergy and Clinical Immunology 2003, 111, 677-690. [CrossRef]
- Allouche, J.; Cremoni, M.; Brglez, V.; Graça, D.; Benzaken, S.; Zorzi, K.; Fernandez, C.; Esnault, V.; Levraut, M.; Oppo, S.; et al. Air pollution exposure induces a decrease in type II interferon response: A paired cohort study. EBioMedicine 2022, 85, 104291. [CrossRef]
- Li, Y.; Lin, B.; Hao, D.; Du, Z.; Wang, Q.; Song, Z.; Li, X.; Li, K.; Wang, J.; Zhang, Q.; et al. Short-term PM(2.5) exposure induces transient lung injury and repair. J Hazard Mater 2023, 459, 132227. [CrossRef]
- Kayalar, Ö.; Rajabi, H.; Konyalilar, N.; Mortazavi, D.; Aksoy, G.T.; Wang, J.; Bayram, H. Impact of particulate air pollution on airway injury and epithelial plasticity; underlying mechanisms. Front Immunol 2024, 15, 1324552. [CrossRef]
- Erickson, M.A.; Jude, J.; Zhao, H.; Rhea, E.M.; Salameh, T.S.; Jester, W.; Pu, S.; Harrowitz, J.; Nguyen, N.; Banks, W.A.; et al. Serum amyloid A: an ozone-induced circulating factor with potentially important functions in the lung-brain axis. Faseb j 2017, 31, 3950-3965. [CrossRef]
- McCarrick, S.; Malmborg, V.; Gren, L.; Danielsen, P.H.; Tunér, M.; Palmberg, L.; Broberg, K.; Pagels, J.; Vogel, U.; Gliga, A.R. Pulmonary exposure to renewable diesel exhaust particles alters protein expression and toxicity profiles in bronchoalveolar lavage fluid and plasma of mice. Arch Toxicol 2025, 99, 797-814. [CrossRef]
- Ghosh, A.; Payton, A.; Gallant, S.C.; Rogers, K.L., Jr.; Mascenik, T.; Hickman, E.; Love, C.A.; Schichlein, K.D.; Smyth, T.R.; Kim, Y.H.; et al. Burn Pit Smoke Condensate-Mediated Toxicity in Human Airway Epithelial Cells. Chem Res Toxicol 2024, 37, 791-803. [CrossRef]
- Kim, Y.H.; Warren, S.H.; Kooter, I.; Williams, W.C.; George, I.J.; Vance, S.A.; Hays, M.D.; Higuchi, M.A.; Gavett, S.H.; DeMarini, D.M.; et al. Chemistry, lung toxicity and mutagenicity of burn pit smoke-related particulate matter. Part Fibre Toxicol 2021, 18, 45. [CrossRef]
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: uncovering the molecular mechanisms. Nat Rev Neurosci 2007, 8, 57-69. [CrossRef]
- Marín-Palma, D.; Fernandez, G.J.; Ruiz-Saenz, J.; Taborda, N.A.; Rugeles, M.T.; Hernandez, J.C. Particulate matter impairs immune system function by up-regulating inflammatory pathways and decreasing pathogen response gene expression. Scientific Reports 2023, 13, 12773. [CrossRef]
- Shukla, A.; Timblin, C.; BeruBe, K.; Gordon, T.; McKinney, W.; Driscoll, K.; Vacek, P.; Mossman, B.T. Inhaled particulate matter causes expression of nuclear factor (NF)-kappaB-related genes and oxidant-dependent NF-kappaB activation in vitro. Am J Respir Cell Mol Biol 2000, 23, 182-187. [CrossRef]
- Debarba, L.K.; Jayarathne, H.S.M.; Stilgenbauer, L.; dos Santos, A.L.T.; Koshko, L.; Scofield, S.; Sullivan, R.; Mandal, A.; Klueh, U.; Sadagurski, M. Microglia Mediate Metabolic Dysfunction From Common Air Pollutants Through NF-κB Signaling. Diabetes 2024, 73, 2065-2077. [CrossRef]
- van Beek, E.M.; Cochrane, F.; Barclay, A.N.; van den Berg, T.K. Signal Regulatory Proteins in the Immune System. The Journal of Immunology 2005, 175, 7781-7787. [CrossRef]
- Roselli, M.; Fernando, R.I.; Guadagni, F.; Spila, A.; Alessandroni, J.; Palmirotta, R.; Costarelli, L.; Litzinger, M.; Hamilton, D.; Huang, B.; et al. Brachyury, a driver of the epithelial-mesenchymal transition, is overexpressed in human lung tumors: an opportunity for novel interventions against lung cancer. Clin Cancer Res 2012, 18, 3868-3879. [CrossRef]
- Hu, Y.; Lu, Y.; Xing, F.; Hsu, W. FGFR1/MAPK-directed brachyury activation drives PD-L1-mediated immune evasion to promote lung cancer progression. Cancer Lett 2022, 547, 215867. [CrossRef]
- Park, K.H.; Choi, Y.J.; Min, W.K.; Lee, S.H.; Kim, J.; Jeong, S.H.; Lee, J.H.; Choi, B.M.; Kim, S. Particulate matter induces arrhythmia-like cardiotoxicity in zebrafish embryos by altering the expression levels of cardiac development- and ion channel-related genes. Ecotoxicol Environ Saf 2023, 263, 115201. [CrossRef]
- Liang, C.; Ding, R.; Sun, Q.; Liu, S.; Sun, Z.; Duan, J. An Overview of Adverse Outcome Pathway Links between PM2.5 Exposure and Cardiac Developmental Toxicity. Environment & Health 2024, 2, 105-113. [CrossRef]
- Sinkevicius, K.W.; Kriegel, C.; Bellaria, K.J.; Lee, J.; Lau, A.N.; Leeman, K.T.; Zhou, P.; Beede, A.M.; Fillmore, C.M.; Caswell, D.; et al. Neurotrophin receptor TrkB promotes lung adenocarcinoma metastasis. Proc Natl Acad Sci U S A 2014, 111, 10299-10304. [CrossRef]
- Zhang, S.-y.; Hui, L.-p.; Li, C.-y.; Gao, J.; Cui, Z.-s.; Qiu, X.-s. More expression of BDNF associates with lung squamous cell carcinoma and is critical to the proliferation and invasion of lung cancer cells. BMC Cancer 2016, 16, 171. [CrossRef]
- Wu, D.-m.; Liu, T.; Deng, S.-h.; Han, R.; Xu, Y. SLC39A4 expression is associated with enhanced cell migration, cisplatin resistance, and poor survival in non-small cell lung cancer. Scientific Reports 2017, 7, 7211. [CrossRef]
- Jiang, Y.; Zhan, H.; Zhang, Y.; Yang, J.; Liu, M.; Xu, C.; Fan, X.; Zhang, J.; Zhou, Z.; Shi, X.; et al. ZIP4 promotes non-small cell lung cancer metastasis by activating snail-N-cadherin signaling axis. Cancer Lett 2021, 521, 71-81. [CrossRef]
- Hu, Y.; Hu, Q.; Ansari, M.; Riemondy, K.; Pineda, R.; Sembrat, J.; Leme, A.S.; Ngo, K.; Morgenthaler, O.; Ha, K.; et al. Airway-derived emphysema-specific alveolar type II cells exhibit impaired regenerative potential in COPD. Eur Respir J 2024, 64. [CrossRef]
- Wang, J.; Zhang, X.; Long, M.; Yuan, M.; Yin, J.; Luo, W.; Wang, S.; Cai, Y.; Jiang, W.; Chao, J. Macrophage-derived GPNMB trapped by fibrotic extracellular matrix promotes pulmonary fibrosis. Commun Biol 2023, 6, 136. [CrossRef]
- Taya, M.; Hammes, S.R. Glycoprotein Non-Metastatic Melanoma Protein B (GPNMB) and Cancer: A Novel Potential Therapeutic Target. Steroids 2018, 133, 102-107. [CrossRef]
- Payapilly, A.; Guilbert, R.; Descamps, T.; White, G.; Magee, P.; Zhou, C.; Kerr, A.; Simpson, K.L.; Blackhall, F.; Dive, C.; et al. TIAM1-RAC1 promote small-cell lung cancer cell survival through antagonizing Nur77-induced BCL2 conformational change. Cell Rep 2021, 37, 109979. [CrossRef]
- Carrera Silva, E.A.; Chan, P.Y.; Joannas, L.; Errasti, A.E.; Gagliani, N.; Bosurgi, L.; Jabbour, M.; Perry, A.; Smith-Chakmakova, F.; Mucida, D.; et al. T cell-derived protein S engages TAM receptor signaling in dendritic cells to control the magnitude of the immune response. Immunity 2013, 39, 160-170. [CrossRef]
- Shin, E.; Kwon, Y.; Jung, E.; Kim, Y.J.; Kim, C.; Hong, S.; Kim, J. TM4SF19 controls GABP-dependent YAP transcription in head and neck cancer under oxidative stress conditions. Proc Natl Acad Sci U S A 2024, 121, e2314346121. [CrossRef]
- Fu, F.; Yang, X.; Zheng, M.; Zhao, Q.; Zhang, K.; Li, Z.; Zhang, H.; Zhang, S. Role of Transmembrane 4 L Six Family 1 in the Development and Progression of Cancer. Front Mol Biosci 2020, 7, 202. [CrossRef]
- Rose, C.E., Jr.; Lannigan, J.A.; Kim, P.; Lee, J.J.; Fu, S.M.; Sung, S.S. Murine lung eosinophil activation and chemokine production in allergic airway inflammation. Cell Mol Immunol 2010, 7, 361-374. [CrossRef]
- Russo, R.C.; Ryffel, B. The Chemokine System as a Key Regulator of Pulmonary Fibrosis: Converging Pathways in Human Idiopathic Pulmonary Fibrosis (IPF) and the Bleomycin-Induced Lung Fibrosis Model in Mice. Cells 2024, 13. [CrossRef]
- Russo, R.C.; Ryffel, B. The Chemokine System as a Key Regulator of Pulmonary Fibrosis: Converging Pathways in Human Idiopathic Pulmonary Fibrosis (IPF) and the Bleomycin-Induced Lung Fibrosis Model in Mice. Cells 2024, 13, 2058.
- Brass, D.M.; Yang, I.V.; Kennedy, M.P.; Whitehead, G.S.; Rutledge, H.; Burch, L.H.; Schwartz, D.A. Fibroproliferation in LPS-induced airway remodeling and bleomycin-induced fibrosis share common patterns of gene expression. Immunogenetics 2008, 60, 353-369. [CrossRef]
- Azfar, M.; van Veen, S.; Houdou, M.; Hamouda, N.N.; Eggermont, J.; Vangheluwe, P. P5B-ATPases in the mammalian polyamine transport system and their role in disease. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 2022, 1869, 119354. [CrossRef]
- Zhou, Y.; Xu, B.; Zhou, Y.; Liu, J.; Zheng, X.; Liu, Y.; Deng, H.; Liu, M.; Ren, X.; Xia, J.; et al. Identification of Key Genes With Differential Correlations in Lung Adenocarcinoma. Front Cell Dev Biol 2021, 9, 675438. [CrossRef]
- Carnesecchi, S.; Deffert, C.; Pagano, A.; Garrido-Urbani, S.; Métrailler-Ruchonnet, I.; Schäppi, M.; Donati, Y.; Matthay, M.A.; Krause, K.H.; Barazzone Argiroffo, C. NADPH oxidase-1 plays a crucial role in hyperoxia-induced acute lung injury in mice. Am J Respir Crit Care Med 2009, 180, 972-981. [CrossRef]
- Seimetz, M.; Sommer, N.; Bednorz, M.; Pak, O.; Veith, C.; Hadzic, S.; Gredic, M.; Parajuli, N.; Kojonazarov, B.; Kraut, S.; et al. NADPH oxidase subunit NOXO1 is a target for emphysema treatment in COPD. Nat Metab 2020, 2, 532-546. [CrossRef]
- Zhou, C.; Gao, Y.; Ding, P.; Wu, T.; Ji, G. The role of CXCL family members in different diseases. Cell Death Discovery 2023, 9, 212. [CrossRef]
- Liang, M.; Jiang, Z.; Huang, Q.; Liu, L.; Xue, Y.; Zhu, X.; Yu, Y.; Wan, W.; Yang, H.; Zou, H. Clinical Association of Chemokine (C-X-C motif) Ligand 1 (CXCL1) with Interstitial Pneumonia with Autoimmune Features (IPAF). Scientific Reports 2016, 6, 38949. [CrossRef]
- Mionnet, C.; Buatois, V.; Kanda, A.; Milcent, V.; Fleury, S.; Lair, D.; Langelot, M.; Lacoeuille, Y.; Hessel, E.; Coffman, R.; et al. CX3CR1 is required for airway inflammation by promoting T helper cell survival and maintenance in inflamed lung. Nature Medicine 2010, 16, 1305-1312. [CrossRef]
- Bain, C.C.; MacDonald, A.S. The impact of the lung environment on macrophage development, activation and function: diversity in the face of adversity. Mucosal Immunology 2022, 15, 223-234. [CrossRef]
- Pilette, C.; Ouadrhiri, Y.; Godding, V.; Vaerman, J.P.; Sibille, Y. Lung mucosal immunity: immunoglobulin-A revisited. Eur Respir J 2001, 18, 571-588. [CrossRef]
- Chae, J.; Choi, J.; Chung, J. Polymeric immunoglobulin receptor (pIgR) in cancer. J Cancer Res Clin Oncol 2023, 149, 17683-17690. [CrossRef]
- Kobayashi, K.; Suzukawa, M.; Watanabe, K.; Arakawa, S.; Igarashi, S.; Asari, I.; Hebisawa, A.; Matsui, H.; Nagai, H.; Nagase, T.; et al. Secretory IgA accumulated in the airspaces of idiopathic pulmonary fibrosis and promoted VEGF, TGF-β and IL-8 production by A549 cells. Clin Exp Immunol 2020, 199, 326-336. [CrossRef]
- Guardado, S.; Ojeda-Juárez, D.; Kaul, M.; Nordgren, T.M. Comprehensive review of lipocalin 2-mediated effects in lung inflammation. Am J Physiol Lung Cell Mol Physiol 2021, 321, L726-l733. [CrossRef]
- Tanahashi, H.; Iwamoto, H.; Yamaguchi, K.; Sakamoto, S.; Horimasu, Y.; Masuda, T.; Nakashima, T.; Ohshimo, S.; Fujitaka, K.; Hamada, H.; et al. Lipocalin-2 as a prognostic marker in patients with acute exacerbation of idiopathic pulmonary fibrosis. Respiratory Research 2024, 25, 195. [CrossRef]
- Treekitkarnmongkol, W.; Hassane, M.; Sinjab, A.; Chang, K.; Hara, K.; Rahal, Z.; Zhang, J.; Lu, W.; Sivakumar, S.; McDowell, T.L.; et al. Augmented Lipocalin-2 Is Associated with Chronic Obstructive Pulmonary Disease and Counteracts Lung Adenocarcinoma Development. American Journal of Respiratory and Critical Care Medicine 2021, 203, 90-101. [CrossRef]
- Cavallaro, D.; Guerrieri, M.; Cattelan, S.; Fabbri, G.; Croce, S.; Armati, M.; Bennett, D.; Fossi, A.; Voltolini, L.; Luzzi, L.; et al. Markers of Bronchiolitis Obliterans Syndrome after Lung Transplant: Between Old Knowledge and Future Perspective. Biomedicines 2022, 10, 3277.
- Vitorakis, N.; Piperi, C. Insights into the Role of Histone Methylation in Brain Aging and Potential Therapeutic Interventions. Int J Mol Sci 2023, 24. [CrossRef]
- Taylor, B.C.; Young, N.L. Histone H4 proteoforms and post-translational modifications in the Mus musculus brain with quantitative comparison of ages and brain regions. Anal Bioanal Chem 2023, 415, 1627-1639. [CrossRef]
- Calderón-Garcidueñas, L.; Herrera-Soto, A.; Jury, N.; Maher, B.A.; González-Maciel, A.; Reynoso-Robles, R.; Ruiz-Rudolph, P.; van Zundert, B.; Varela-Nallar, L. Reduced repressive epigenetic marks, increased DNA damage and Alzheimer's disease hallmarks in the brain of humans and mice exposed to particulate urban air pollution. Environ Res 2020, 183, 109226. [CrossRef]
- Jakovcevski, M.; Akbarian, S. Epigenetic mechanisms in neurological disease. Nature Medicine 2012, 18, 1194-1204. [CrossRef]
- Edamatsu, M.; Miyano, R.; Fujikawa, A.; Fujii, F.; Hori, T.; Sakaba, T.; Oohashi, T. Hapln4/Bral2 is a selective regulator for formation and transmission of GABAergic synapses between Purkinje and deep cerebellar nuclei neurons. J Neurochem 2018, 147, 748-763. [CrossRef]
- Pintér, P.; Alpár, A. The Role of Extracellular Matrix in Human Neurodegenerative Diseases. International Journal of Molecular Sciences 2022, 23, 11085.
- Back, S.A.; Tuohy, T.M.F.; Chen, H.; Wallingford, N.; Craig, A.; Struve, J.; Luo, N.L.; Banine, F.; Liu, Y.; Chang, A.; et al. Hyaluronan accumulates in demyelinated lesions and inhibits oligodendrocyte progenitor maturation. Nature Medicine 2005, 11, 966-972. [CrossRef]
- Doeppner, T.R.; Herz, J.; Bähr, M.; Tonchev, A.B.; Stoykova, A. Zbtb20 Regulates Developmental Neurogenesis in the Olfactory Bulb and Gliogenesis After Adult Brain Injury. Mol Neurobiol 2019, 56, 567-582. [CrossRef]
- Zhao, Y.; Gai, C.; Yu, S.; Song, Y.; Gu, B.; Luo, Q.; Wang, X.; Hu, Q.; Liu, W.; Liu, D.; et al. Liposomes-Loaded miR-9-5p Alleviated Hypoxia-Ischemia-Induced Mitochondrial Oxidative Stress by Targeting ZBTB20 to Inhibiting Nrf2/Keap1 Interaction in Neonatal Mice. Antioxid Redox Signal 2025, 42, 512-528. [CrossRef]
- Solleiro-Villavicencio, H.; Rivas-Arancibia, S. Effect of Chronic Oxidative Stress on Neuroinflammatory Response Mediated by CD4+T Cells in Neurodegenerative Diseases. Frontiers in Cellular Neuroscience 2018, Volume 12 - 2018. [CrossRef]
- Wang, Y.; Cai, M.; Lou, Y.; Zhang, S.; Liu, X. ZBTB20-AS1 promoted Alzheimer's disease progression through ZBTB20/GSK-3β/Tau pathway. Biochemical and Biophysical Research Communications 2023, 640, 88-96. [CrossRef]
- Shih, J.C.; Chen, K.; Ridd, M.J. Monoamine oxidase: from genes to behavior. Annu Rev Neurosci 1999, 22, 197-217. [CrossRef]
- Singh, C.; Bortolato, M.; Bali, N.; Godar, S.C.; Scott, A.L.; Chen, K.; Thompson, R.F.; Shih, J.C. Cognitive abnormalities and hippocampal alterations in monoamine oxidase A and B knockout mice. Proc Natl Acad Sci U S A 2013, 110, 12816-12821. [CrossRef]
- Beucher, L.; Gabillard-Lefort, C.; Baris, O.R.; Mialet-Perez, J. Monoamine oxidases: A missing link between mitochondria and inflammation in chronic diseases ? Redox Biology 2024, 77, 103393. [CrossRef]
- Zou, X.; Gao, S.; Li, J.; Li, C.; Wu, C.; Cao, X.; Xia, S.; Shao, P.; Bao, X.; Yang, H.; et al. A monoamine oxidase B inhibitor ethyl ferulate suppresses microglia-mediated neuroinflammation and alleviates ischemic brain injury. Front Pharmacol 2022, 13, 1004215. [CrossRef]
- Hughes, T.M.; Rosano, C.; Evans, R.W.; Kuller, L.H. Brain cholesterol metabolism, oxysterols, and dementia. J Alzheimers Dis 2013, 33, 891-911. [CrossRef]
- Zhang, X.; Lv, C.; An, Y.; Liu, Q.; Rong, H.; Tao, L.; Wang, Y.; Wang, Y.; Xiao, R. Increased Levels of 27-Hydroxycholesterol Induced by Dietary Cholesterol in Brain Contribute to Learning and Memory Impairment in Rats. Mol Nutr Food Res 2018, 62. [CrossRef]
- Dunk, M.M.; Rapp, S.R.; Hayden, K.M.; Espeland, M.A.; Casanova, R.; Manson, J.E.; Shadyab, A.H.; Wild, R.; Driscoll, I. Plasma oxysterols are associated with serum lipids and dementia risk in older women. Alzheimers Dement 2024, 20, 3696-3704. [CrossRef]
- Björkhem, I.; Leoni, V.; Meaney, S. Genetic connections between neurological disorders and cholesterol metabolism. Journal of Lipid Research 2010, 51, 2489-2503. [CrossRef]
- Rat Genome Database, R. ENSRNOG00000064885: Predicted Ensembl rat gene. Available online: https://rgd.mcw.edu/ (accessed on 04/25/2025).
- Martin, I.; Kim, J.W.; Lee, B.D.; Kang, H.C.; Xu, J.C.; Jia, H.; Stankowski, J.; Kim, M.S.; Zhong, J.; Kumar, M.; et al. Ribosomal protein s15 phosphorylation mediates LRRK2 neurodegeneration in Parkinson's disease. Cell 2014, 157, 472-485. [CrossRef]
- Asano, Y.; Kishida, S.; Mu, P.; Sakamoto, K.; Murohara, T.; Kadomatsu, K. DRR1 is expressed in the developing nervous system and downregulated during neuroblastoma carcinogenesis. Biochemical and Biophysical Research Communications 2010, 394, 829-835. [CrossRef]
- van Olst, L.; Simonton, B.; Edwards, A.J.; Forsyth, A.V.; Boles, J.; Jamshidi, P.; Watson, T.; Shepard, N.; Krainc, T.; Argue, B.M.R.; et al. Microglial mechanisms drive amyloid-β clearance in immunized patients with Alzheimer’s disease. Nature Medicine 2025. [CrossRef]
- Xue, C.; Chu, Q.; Shi, Q.; Zeng, Y.; Lu, J.; Li, L. Wnt signaling pathways in biology and disease: mechanisms and therapeutic advances. Signal Transduction and Targeted Therapy 2025, 10, 106. [CrossRef]
- Halleskog, C.; Mulder, J.; Dahlström, J.; Mackie, K.; Hortobágyi, T.; Tanila, H.; Kumar Puli, L.; Färber, K.; Harkany, T.; Schulte, G. WNT signaling in activated microglia is proinflammatory. Glia 2011, 59, 119-131. [CrossRef]
- Ensembl. Gene: ENSRNOG00000069713 – Rattus norvegicus. Available online: https://useast.ensembl.org/Rattus_norvegicus/Gene/Splice?db=core;g=ENSRNOG00000069713;r=X:15395785-15398610;t=ENSRNOT00000101970 (accessed on 04/25/2025).
- Qureshi, I.A.; Mehler, M.F. Emerging roles of non-coding RNAs in brain evolution, development, plasticity and disease. Nature Reviews Neuroscience 2012, 13, 528-541. [CrossRef]
- Zeng, H.-X.; Qin, S.-J.; Andersson, J.; Li, S.-P.; Zeng, Q.-G.; Li, J.-H.; Wu, Q.-Z.; Meng, W.-J.; Oudin, A.; Kanninen, K.M.; et al. The emerging roles of particulate matter-changed non-coding RNAs in the pathogenesis of Alzheimer's disease: A comprehensive in silico analysis and review. Environmental Pollution 2025, 366, 125440. [CrossRef]
- Hirano, A.; Hsu, P.K.; Zhang, L.; Xing, L.; McMahon, T.; Yamazaki, M.; Ptáček, L.J.; Fu, Y.H. DEC2 modulates orexin expression and regulates sleep. Proc Natl Acad Sci U S A 2018, 115, 3434-3439. [CrossRef]
- Lane, J.M.; Qian, J.; Mignot, E.; Redline, S.; Scheer, F.; Saxena, R. Genetics of circadian rhythms and sleep in human health and disease. Nat Rev Genet 2023, 24, 4-20. [CrossRef]
- van den Ameele, J.; Krautz, R.; Cheetham, S.W.; Donovan, A.P.A.; Llorà-Batlle, O.; Yakob, R.; Brand, A.H. Reduced chromatin accessibility correlates with resistance to Notch activation. Nature Communications 2022, 13, 2210. [CrossRef]
- Chuikov, S.; Levi, B.P.; Smith, M.L.; Morrison, S.J. Prdm16 promotes stem cell maintenance in multiple tissues, partly by regulating oxidative stress. Nat Cell Biol 2010, 12, 999-1006. [CrossRef]
- Schröder, S.; Fuchs, U.; Gisa, V.; Pena, T.; Krüger, D.M.; Hempel, N.; Burkhardt, S.; Salinas, G.; Schütz, A.L.; Delalle, I.; et al. PRDM16-DT is a novel lncRNA that regulates astrocyte function in Alzheimer's disease. Acta Neuropathol 2024, 148, 32. [CrossRef]
- Xu, X.; Liu, C.; Xu, Z.; Tzan, K.; Zhong, M.; Wang, A.; Lippmann, M.; Chen, L.C.; Rajagopalan, S.; Sun, Q. Long-term exposure to ambient fine particulate pollution induces insulin resistance and mitochondrial alteration in adipose tissue. Toxicol Sci 2011, 124, 88-98. [CrossRef]
- Ensembl. Senp5-ps1 (Small ubiquitin-like modifier (SUMO) protease 5 pseudogene) [ENSRNOG00000068362.1]. Available online: https://useast.ensembl.org/Rattus_norvegicus/Gene/Summary?db=core;g=ENSRNOG00000068362;r=2:95501188-95504509;t=ENSRNOT00000094190 (accessed on 27 April).
- Pink, R.C.; Wicks, K.; Caley, D.P.; Punch, E.K.; Jacobs, L.; Carter, D.R. Pseudogenes: pseudo-functional or key regulators in health and disease? Rna 2011, 17, 792-798. [CrossRef]
- Henley, J.M.; Craig, T.J.; Wilkinson, K.A. Neuronal SUMOylation: mechanisms, physiology, and roles in neuronal dysfunction. Physiol Rev 2014, 94, 1249-1285. [CrossRef]
- NCBI, G. POM121 transmembrane nucleoporin-like 2 (POM121L2), Rattus norvegicus (Rat). Gene ID: 94026. Available online: https://www.ncbi.nlm.nih.gov/gene/94026 (accessed on 26 April 2025).
- Coyne, A.N.; Rothstein, J.D. Nuclear pore complexes — a doorway to neural injury in neurodegeneration. Nature Reviews Neurology 2022, 18, 348-362. [CrossRef]
- Coyne, A.N.; Rothstein, J.D. The ESCRT-III protein VPS4, but not CHMP4B or CHMP2B, is pathologically increased in familial and sporadic ALS neuronal nuclei. Acta Neuropathol Commun 2021, 9, 127. [CrossRef]
- Komiyama, K.; Iijima, K.; Kawabata-Iwakawa, R.; Fujihara, K.; Kakizaki, T.; Yanagawa, Y.; Yoshimoto, Y.; Miyata, S. Glioma facilitates the epileptic and tumor-suppressive gene expressions in the surrounding region. Sci Rep 2022, 12, 6805. [CrossRef]
- Meyer, K.; Buettner, S.; Ghezzi, D.; Zeviani, M.; Bano, D.; Nicotera, P. Loss of apoptosis-inducing factor critically affects MIA40 function. Cell Death & Disease 2015, 6, e1814-e1814. [CrossRef]
- Modjtahedi, N.; Tokatlidis, K.; Dessen, P.; Kroemer, G. Mitochondrial Proteins Containing Coiled-Coil-Helix-Coiled-Coil-Helix (CHCH) Domains in Health and Disease. Trends Biochem Sci 2016, 41, 245-260. [CrossRef]
- Guerreiro, R.; Gibbons, E.; Tábuas-Pereira, M.; Kun-Rodrigues, C.; Santo, G.C.; Bras, J. Genetic architecture of common non-Alzheimer's disease dementias. Neurobiol Dis 2020, 142, 104946. [CrossRef]
- Bogorodskiy, A.; Okhrimenko, I.; Burkatovskii, D.; Jakobs, P.; Maslov, I.; Gordeliy, V.; Dencher, N.A.; Gensch, T.; Voos, W.; Altschmied, J.; et al. Role of Mitochondrial Protein Import in Age-Related Neurodegenerative and Cardiovascular Diseases. Cells 2021, 10. [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proceedings of the National Academy of Sciences 2005, 102, 15545-15550, doi:doi:10.1073/pnas.0506580102.
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Research 2022, 51, D587-D592. [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Research 2000, 28, 27-30. [CrossRef]
- Saul, D.; Kosinsky, R.L.; Atkinson, E.J.; Doolittle, M.L.; Zhang, X.; LeBrasseur, N.K.; Pignolo, R.J.; Robbins, P.D.; Niedernhofer, L.J.; Ikeno, Y.; et al. A new gene set identifies senescent cells and predicts senescence-associated pathways across tissues. Nature Communications 2022, 13, 4827. [CrossRef]
- Cao, M.; Liu, J.; Zhang, X.; Wang, Y.; Hou, Y.; Song, Q.; Cui, Y.; Zhao, Y.; Wang, P. IL-17A promotes the progression of Alzheimer’s disease in APP/PS1 mice. Immunity & Ageing 2023, 20, 74. [CrossRef]
- Huang, Y.; Happonen, K.E.; Burrola, P.G.; O'Connor, C.; Hah, N.; Huang, L.; Nimmerjahn, A.; Lemke, G. Microglia use TAM receptors to detect and engulf amyloid β plaques. Nat Immunol 2021, 22, 586-594. [CrossRef]
- Pfau, S.J.; Langen, U.H.; Fisher, T.M.; Prakash, I.; Nagpurwala, F.; Lozoya, R.A.; Lee, W.-C.A.; Wu, Z.; Gu, C. Characteristics of blood–brain barrier heterogeneity between brain regions revealed by profiling vascular and perivascular cells. Nature Neuroscience 2024, 27, 1892-1903. [CrossRef]
- Choi, A.M.K.; Ryter, S.W.; Levine, B. Autophagy in human health and disease. The New England Journal of Medicine 2013, 368, 651-662. [CrossRef]
- Nixon, R.A. The role of autophagy in neurodegenerative disease. Nature Medicine 2013, 19, 983-997. [CrossRef]
- Park, M.; Pei, R. Endocytosis and its roles in lung diseases. International Journal of Molecular Sciences 2020, 21, 6021. [CrossRef]
- Saheki, Y.; De Camilli, P. Synaptic vesicle endocytosis. Cold Spring Harbor Perspectives in Biology 2012, 4, a005645. [CrossRef]
- Cloonan, S.M.; Choi, A.M.K. Mitochondria in lung disease. Journal of Clinical Investigation 2016, 126, 809-820. [CrossRef]
- Lin, M.-T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787-795. [CrossRef]
- R Core Team. R: A language and environment for statistical computing. Available online: https://www.R-project.org/ (accessed on.
- Hathaway, Q.A.; Majumder, N.; Goldsmith, W.T.; Kunovac, A.; Pinti, M.V.; Harkema, J.R.; Castranova, V.; Hollander, J.M.; Hussain, S. Transcriptomics of single dose and repeated carbon black and ozone inhalation co-exposure highlight progressive pulmonary mitochondrial dysfunction. Part Fibre Toxicol 2021, 18, 44. [CrossRef]
- Gao, E.; Shao, W.; Zhang, C.; Yu, M.; Dai, H.; Fan, T.; Zhu, Z.; Xu, W.; Huang, J.; Zhang, Y.; et al. Lung Epithelial Cells Can Produce Antibodies Participating In Adaptive Humoral Immune Responses. bioRxiv 2021, 2021.2005.2013.443498. [CrossRef]
- Kato, A.; Hulse, K.E.; Tan, B.K.; Schleimer, R.P. B-lymphocyte lineage cells and the respiratory system. Journal of Allergy and Clinical Immunology 2013, 131, 933-957. [CrossRef]
- Sun, H.; Lin, A.H.; Ru, F.; Patil, M.J.; Meeker, S.; Lee, L.Y.; Undem, B.J. KCNQ/M-channels regulate mouse vagal bronchopulmonary C-fiber excitability and cough sensitivity. JCI Insight 2019, 4. [CrossRef]
- Hayashi, A.; Ohnishi, H.; Okazawa, H.; Nakazawa, S.; Ikeda, H.; Motegi, S.; Aoki, N.; Kimura, S.; Mikuni, M.; Matozaki, T. Positive regulation of phagocytosis by SIRPbeta and its signaling mechanism in macrophages. J Biol Chem 2004, 279, 29450-29460. [CrossRef]
- National Center for Biotechnology, I. SIRPB3P signal regulatory protein beta 3, pseudogene [Homo sapiens (human)]. NCBI Gene 2025.
- Shen, Q.; Zhao, L.; Pan, L.; Li, D.; Chen, G.; Chen, Z.; Jiang, Z. Soluble SIRP-Alpha Promotes Murine Acute Lung Injury Through Suppressing Macrophage Phagocytosis. Front Immunol 2022, 13, 865579. [CrossRef]
- Deng, Z.; Fan, T.; Xiao, C.; Tian, H.; Zheng, Y.; Li, C.; He, J. TGF-β signaling in health, disease and therapeutics. Signal Transduction and Targeted Therapy 2024, 9, 61. [CrossRef]
- Wu, M.; Wang, Z.; Shi, X.; Zan, D.; Chen, H.; Yang, S.; Ding, F.; Yang, L.; Tan, P.; Ma, R.Z.; et al. TGFβ1-RCN3-TGFBR1 loop facilitates pulmonary fibrosis by orchestrating fibroblast activation. Respiratory Research 2023, 24, 222. [CrossRef]
- Lee, T.-H.; Yeh, C.-F.; Lee, Y.-T.; Shih, Y.-C.; Chen, Y.-T.; Hung, C.-T.; You, M.-Y.; Wu, P.-C.; Shentu, T.-P.; Huang, R.-T.; et al. Fibroblast-enriched endoplasmic reticulum protein TXNDC5 promotes pulmonary fibrosis by augmenting TGFβ signaling through TGFBR1 stabilization. Nature Communications 2020, 11, 4254. [CrossRef]
- Aschner, Y.; Downey, G.P. Transforming Growth Factor-β: Master Regulator of the Respiratory System in Health and Disease. Am J Respir Cell Mol Biol 2016, 54, 647-655. [CrossRef]
- Choi, C.; Jeong, Y.L.; Park, K.-M.; Kim, M.; Kim, S.; Jo, H.; Lee, S.; Kim, H.; Choi, G.; Choi, Y.H.; et al. TM4SF19-mediated control of lysosomal activity in macrophages contributes to obesity-induced inflammation and metabolic dysfunction. Nature Communications 2024, 15, 2779. [CrossRef]
- Chung, C.-T.; Yeh, K.-C.; Lee, C.-H.; Chen, Y.-Y.; Ho, P.-J.; Chang, K.-Y.; Chen, C.-H.; Lai, Y.-K.; Chen, C.-T. Molecular profiling of afatinib-resistant non-small cell lung cancer cells in vivo derived from mice. Pharmacological Research 2020, 161, 105183. [CrossRef]
- Zhao, X.; Yue, Y.; Wang, X.; Zhang, Q. Bioinformatics analysis of PLA2G7 as an immune-related biomarker in COPD by promoting expansion and suppressive functions of MDSCs. Int Immunopharmacol 2023, 120, 110399. [CrossRef]
- Karhadkar, T.R.; Pilling, D.; Cox, N.; Gomer, R.H. Sialidase inhibitors attenuate pulmonary fibrosis in a mouse model. Scientific Reports 2017, 7, 15069. [CrossRef]
- Krick, S.; Helton, E.S.; Easter, M.; Bollenbecker, S.; Denson, R.; Zaharias, R.; Cochran, P.; Vang, S.; Harris, E.; Wells, J.M.; et al. ST6GAL1 and α2-6 Sialylation Regulates IL-6 Expression and Secretion in Chronic Obstructive Pulmonary Disease. Front Immunol 2021, 12, 693149. [CrossRef]
- Fulkerson, P.C.; Zimmermann, N.; Hassman, L.M.; Finkelman, F.D.; Rothenberg, M.E. Pulmonary Chemokine Expression Is Coordinately Regulated by STAT1, STAT6, and IFN-γ. The Journal of Immunology 2004, 173, 7565-7574. [CrossRef]
- Yan, H.H.; Jiang, J.; Pang, Y.; Achyut, B.R.; Lizardo, M.; Liang, X.; Hunter, K.; Khanna, C.; Hollander, C.; Yang, L. CCL9 Induced by TGFβ Signaling in Myeloid Cells Enhances Tumor Cell Survival in the Premetastatic Organ. Cancer Res 2015, 75, 5283-5298. [CrossRef]
- Yao, Y.; Zhang, T.; Qi, L.; Liu, R.; Liu, G.; Li, J.; Sun, C. Identification of Four Genes as Prognosis Signatures in Lung Adenocarcinoma Microenvironment. Pharmgenomics Pers Med 2021, 14, 15-26. [CrossRef]
- Li, S.; Xiang, M. Foxn4 influences alveologenesis during lung development. Dev Dyn 2011, 240, 1512-1517. [CrossRef]
- Zhang, X.; Huang, X.; Zhang, X.; Lai, L.; Zhu, B.; Lin, P.; Kang, Z.; Yin, D.; Tian, D.; Chen, Z.; et al. The miR-941/FOXN4/TGF-β feedback loop induces N2 polarization of neutrophils and enhances tumor progression of lung adenocarcinoma. Frontiers in Immunology 2025, Volume 16 - 2025. [CrossRef]
- Aros, C.J.; Pantoja, C.J.; Gomperts, B.N. Wnt signaling in lung development, regeneration, and disease progression. Communications Biology 2021, 4, 601. [CrossRef]
- Vance, S.A.; Kim, Y.H.; George, I.J.; Dye, J.A.; Williams, W.C.; Schladweiler, M.J.; Gilmour, M.I.; Jaspers, I.; Gavett, S.H. Contributions of particulate and gas phases of simulated burn pit smoke exposures to impairment of respiratory function. Inhal Toxicol 2023, 35, 129-138. [CrossRef]
- Gutor, S.S.; Miller, R.F.; Blackwell, T.S.; Polosukhin, V.V. Environmental and occupational bronchiolitis obliterans: new reality. EBioMedicine 2023, 95, 104760. [CrossRef]
- Gutor, S.S.; Salinas, R.I.; Nichols, D.S.; Bazzano, J.M.R.; Han, W.; Gokey, J.J.; Vasiukov, G.; West, J.D.; Newcomb, D.C.; Dikalova, A.E.; et al. Repetitive sulfur dioxide exposure in mice models post-deployment respiratory syndrome. Am J Physiol Lung Cell Mol Physiol 2024, 326, L539-L550. [CrossRef]
- Sibener, L.; Zaganjor, I.; Snyder, H.M.; Bain, L.J.; Egge, R.; Carrillo, M.C. Alzheimer's Disease prevalence, costs, and prevention for military personnel and veterans. Alzheimers Dement 2014, 10, S105-110. [CrossRef]
- Tanner, C.M.; Goldman, S.M.; Ross, G.W.; Grate, S.J. The disease intersection of susceptibility and exposure: chemical exposures and neurodegenerative disease risk. Alzheimers Dement 2014, 10, S213-225. [CrossRef]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nature Reviews Immunology 2016, 16, 626-638. [CrossRef]
- So, S.W.; Nixon, J.P.; Bernlohr, D.A.; Butterick, T.A. RNAseq Analysis of FABP4 Knockout Mouse Hippocampal Transcriptome Suggests a Role for WNT/beta-Catenin in Preventing Obesity-Induced Cognitive Impairment. Int J Mol Sci 2023, 24. [CrossRef]
- Breheny P, H.J. Coordinate descent algorithms for nonconvex penalized regression, with applications to biological feature selection. Annals of Applied Statistics 2011, 5, 232-253.
- Fan, J.; Li, R. Variable selection via nonconcave penalized likelihood and its oracle properties. Journal of the American Statistical Association 2001, 96, 1348-1360.
- Friedman, J.; Hastie, T.; Tibshirani, R. Regularization Paths for Generalized Linear Models via Coordinate Descent. J Stat Softw 2010, 33, 1-22.
- Chen, F.; Zhang, W.; Mfarrej, M.F.B.; Saleem, M.H.; Khan, K.A.; Ma, J.; Raposo, A.; Han, H. Breathing in danger: Understanding the multifaceted impact of air pollution on health impacts. Ecotoxicology and Environmental Safety 2024, 280, 116532. [CrossRef]
- Trembley, J.H.; Barach, P.; Tomáška, J.M.; Poole, J.T.; Ginex, P.K.; Miller, R.F.; Sandri, B.J.; Szema, A.M.; Gandy, K.; Siddharthan, T.; et al. Veterans Affairs Military Toxic Exposure Research Conference: Veteran-centric Approach and Community of Practice. Mil Med 2024. [CrossRef]
- Ginex, P.K.; Barach, P.; Boffetta, P.; Poole, J.T.; Trembley, J.H.; Tomấška, J.; Klein, M.A.; Butterick, T.A. Exposure-Informed Care Following Toxic Environmental Exposures: A Lifestyle Medicine Approach. Am J Lifestyle Med 2025, 15598276251327106. [CrossRef]
- Tay, J.K.; Narasimhan, B.; Hastie, T. Elastic Net Regularization Paths for All Generalized Linear Models. Journal of Statistical Software 2023, 106, 1-31.
- Tibshirani, R. Regression shrinkage and selection via the lasso. Journal of the Royal Statistical Society Series B: Statistical Methodology 1996, 58, 267-288.
- Breheny, P.; Huang, J. Coordinate descent algorithms for nonconvex penalized regression, with applications to biological feature selection. Annals of Applied Statistics 2011, 5, 232-253.
- Zou, H.; Hastie, T. Regression shrinkage and selection via the elastic net, with applications to microarrays. Journal of the Royal Statistical Society Series B 2003, 67, 301-320.
- Cheng, H.; Garrick, D.J.; Fernando, R.L. Efficient strategies for leave-one-out cross validation for genomic best linear unbiased prediction. Journal of Animal Science and Biotechnology 2017, 8, 1-5.
Figure 1.
Proinflammatory protein profiling in lung. Lung tissues were collected 24 hours post-exposure and analyzed using a multiplex electrochemiluminescence assay (MSD V-PLEX Rat Pro-inflammatory Panel 2). Concentrations of IL 1β, IL 6, IFN γ, IL 4, IL 10, KC/GRO (CXCL1), IL 5, IL 13, and TNF α were quantified. Data were analyzed using ordinary one-way ANOVA followed by Holm-Šídák’s multiple comparisons test. Statistical significance is indicated as follows: *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. Error bars represent mean ± SEM.
Figure 1.
Proinflammatory protein profiling in lung. Lung tissues were collected 24 hours post-exposure and analyzed using a multiplex electrochemiluminescence assay (MSD V-PLEX Rat Pro-inflammatory Panel 2). Concentrations of IL 1β, IL 6, IFN γ, IL 4, IL 10, KC/GRO (CXCL1), IL 5, IL 13, and TNF α were quantified. Data were analyzed using ordinary one-way ANOVA followed by Holm-Šídák’s multiple comparisons test. Statistical significance is indicated as follows: *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. Error bars represent mean ± SEM.
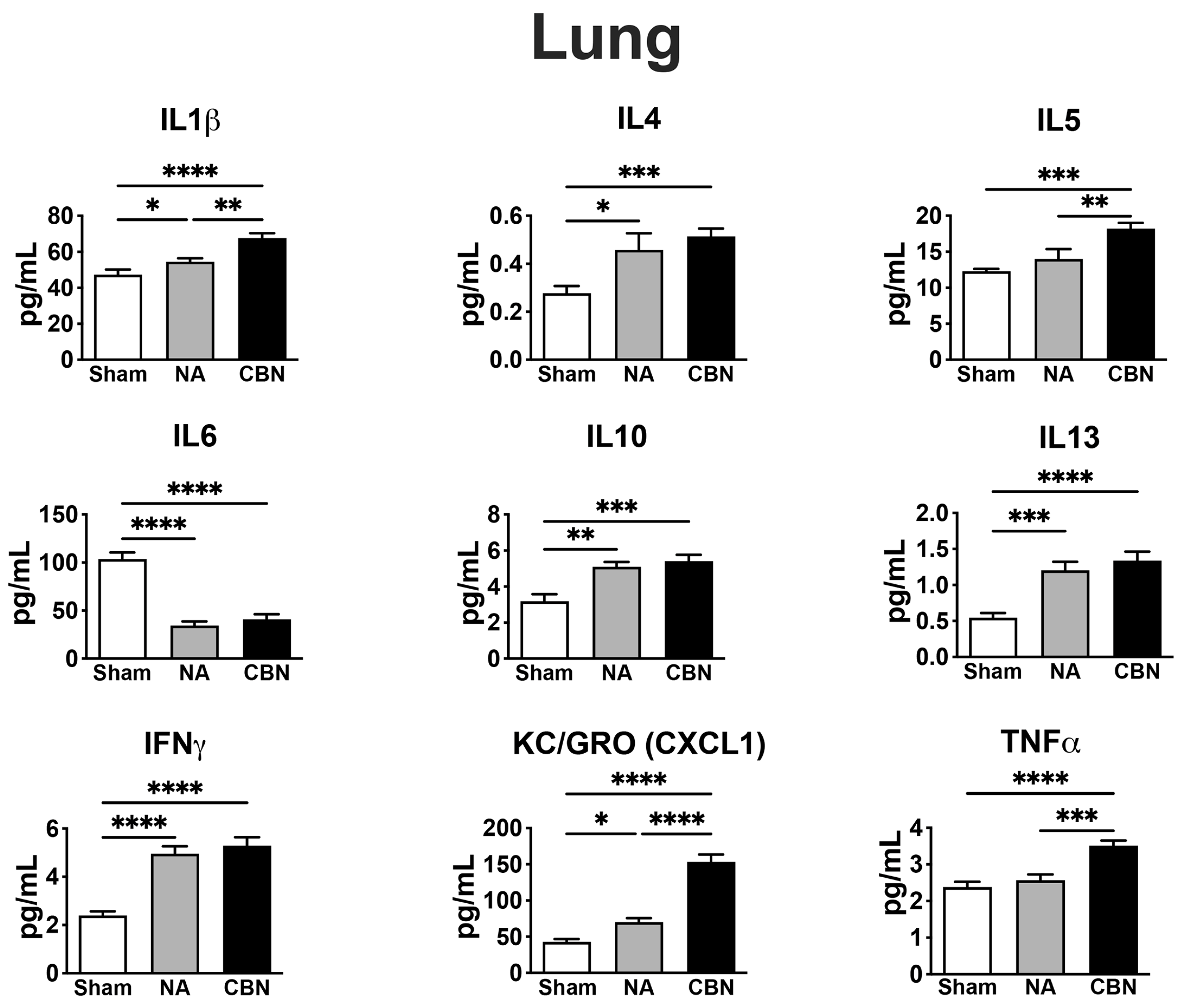
Figure 2.
Proinflammatory protein profiling in plasma. Plasma was collected 24 hours post-exposure and analyzed using a multiplex electrochemiluminescence assay (MSD V-PLEX Rat Pro-inflammatory Panel 2). Statistical significance was determined by ordinary one-way ANOVA with Holm-Šídák’s post hoc test and is denoted as follows: *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. Error bars represent mean ± SEM. nt, not detected. ns, not significant.
Figure 2.
Proinflammatory protein profiling in plasma. Plasma was collected 24 hours post-exposure and analyzed using a multiplex electrochemiluminescence assay (MSD V-PLEX Rat Pro-inflammatory Panel 2). Statistical significance was determined by ordinary one-way ANOVA with Holm-Šídák’s post hoc test and is denoted as follows: *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. Error bars represent mean ± SEM. nt, not detected. ns, not significant.
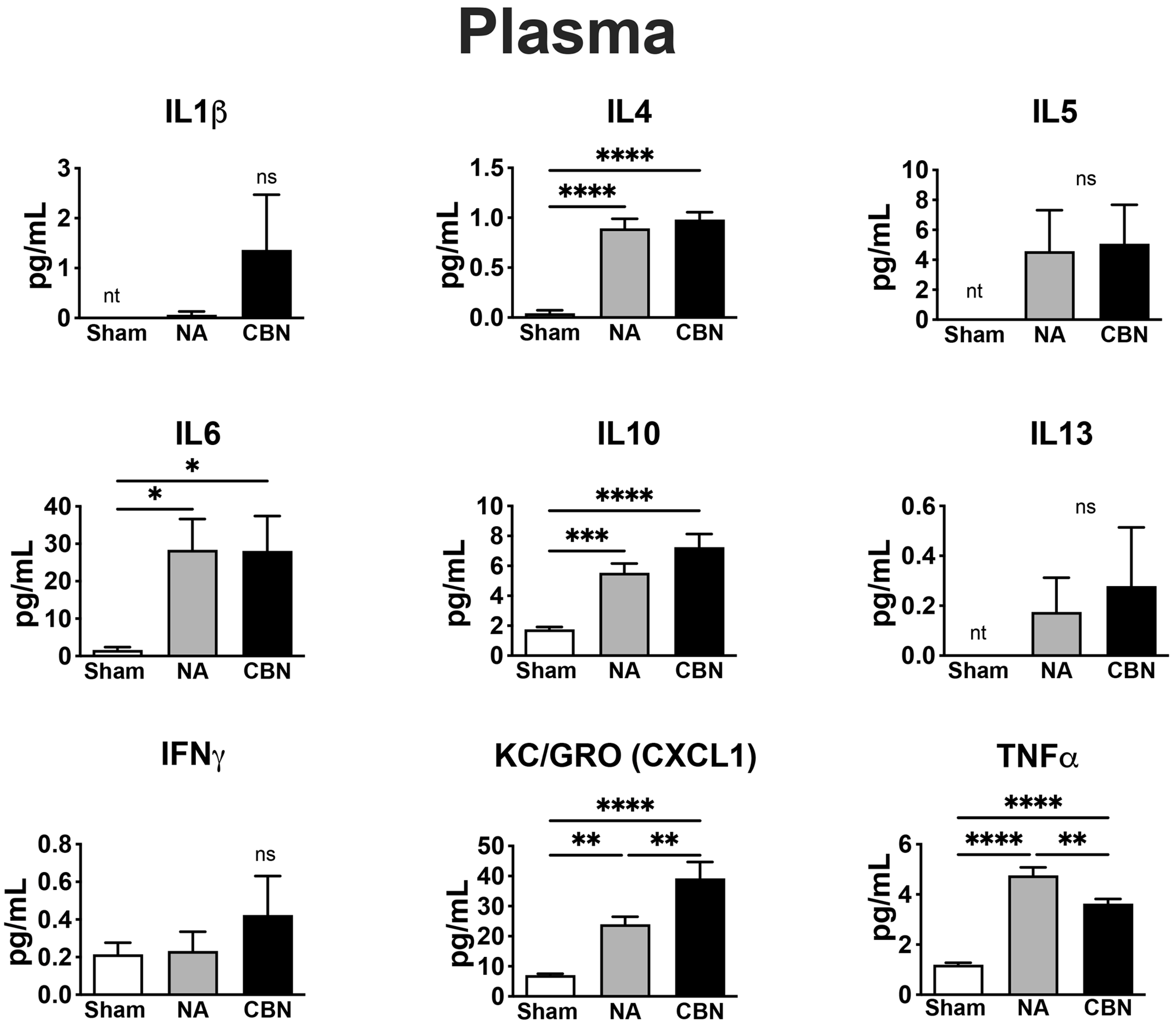
Figure 3.
Proinflammatory protein profiling in brain. Brain tissues were harvested 24 hours post-exposure and analyzed using a multiplex electrochemiluminescence assay (MSD V-PLEX Rat Pro-inflammatory Panel 2). Statistical significance was determined by ordinary one-way ANOVA with Holm-Šídák’s post hoc test and is denoted as follows: *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. Error bars represent mean ± SEM. ns, not significant.
Figure 3.
Proinflammatory protein profiling in brain. Brain tissues were harvested 24 hours post-exposure and analyzed using a multiplex electrochemiluminescence assay (MSD V-PLEX Rat Pro-inflammatory Panel 2). Statistical significance was determined by ordinary one-way ANOVA with Holm-Šídák’s post hoc test and is denoted as follows: *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. Error bars represent mean ± SEM. ns, not significant.
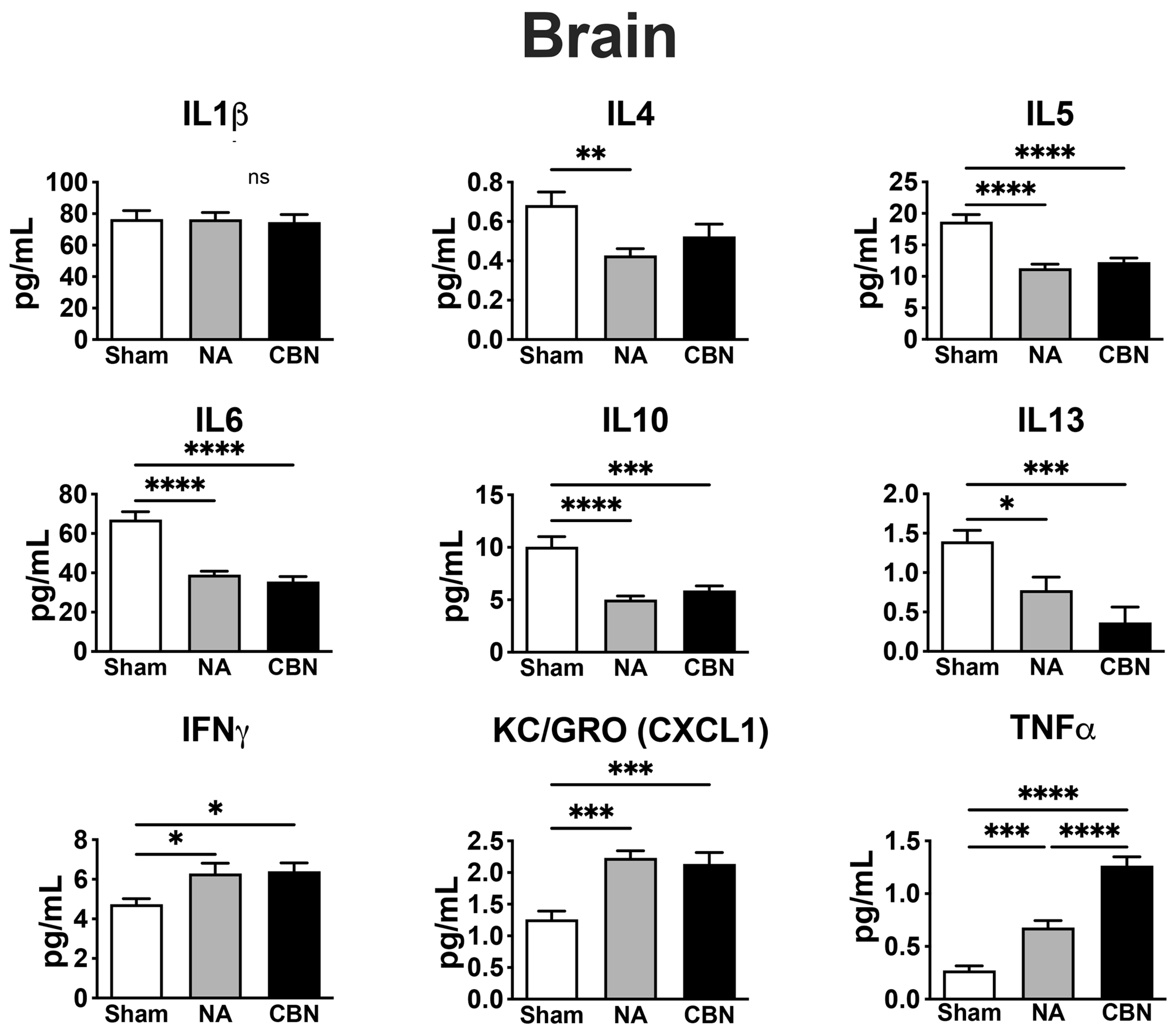
Figure 4.
Summary of RNA sequencing differential expression analysis. Multidimensional scaling plots showing differences between rats within each tissue type based on expression of 200 genes with the greatest log2 fold-change difference between groups treated with sham air (control) and CBN for lung (A) or brain (B). Volcano plots summarizing differential expression results. Significantly differentially expressed (DE) genes (FDR p-value < 0.05) are shown in red (up-regulated in CBN exposed) or blue (down-regulated in CBN exposed), and the top ten DE genes are labeled for lung (C) or brain (D). Heatmap showing scaled log2 counts per million (z-scores) for the top 15 DE genes in lung (E) or brain (F).
Figure 4.
Summary of RNA sequencing differential expression analysis. Multidimensional scaling plots showing differences between rats within each tissue type based on expression of 200 genes with the greatest log2 fold-change difference between groups treated with sham air (control) and CBN for lung (A) or brain (B). Volcano plots summarizing differential expression results. Significantly differentially expressed (DE) genes (FDR p-value < 0.05) are shown in red (up-regulated in CBN exposed) or blue (down-regulated in CBN exposed), and the top ten DE genes are labeled for lung (C) or brain (D). Heatmap showing scaled log2 counts per million (z-scores) for the top 15 DE genes in lung (E) or brain (F).
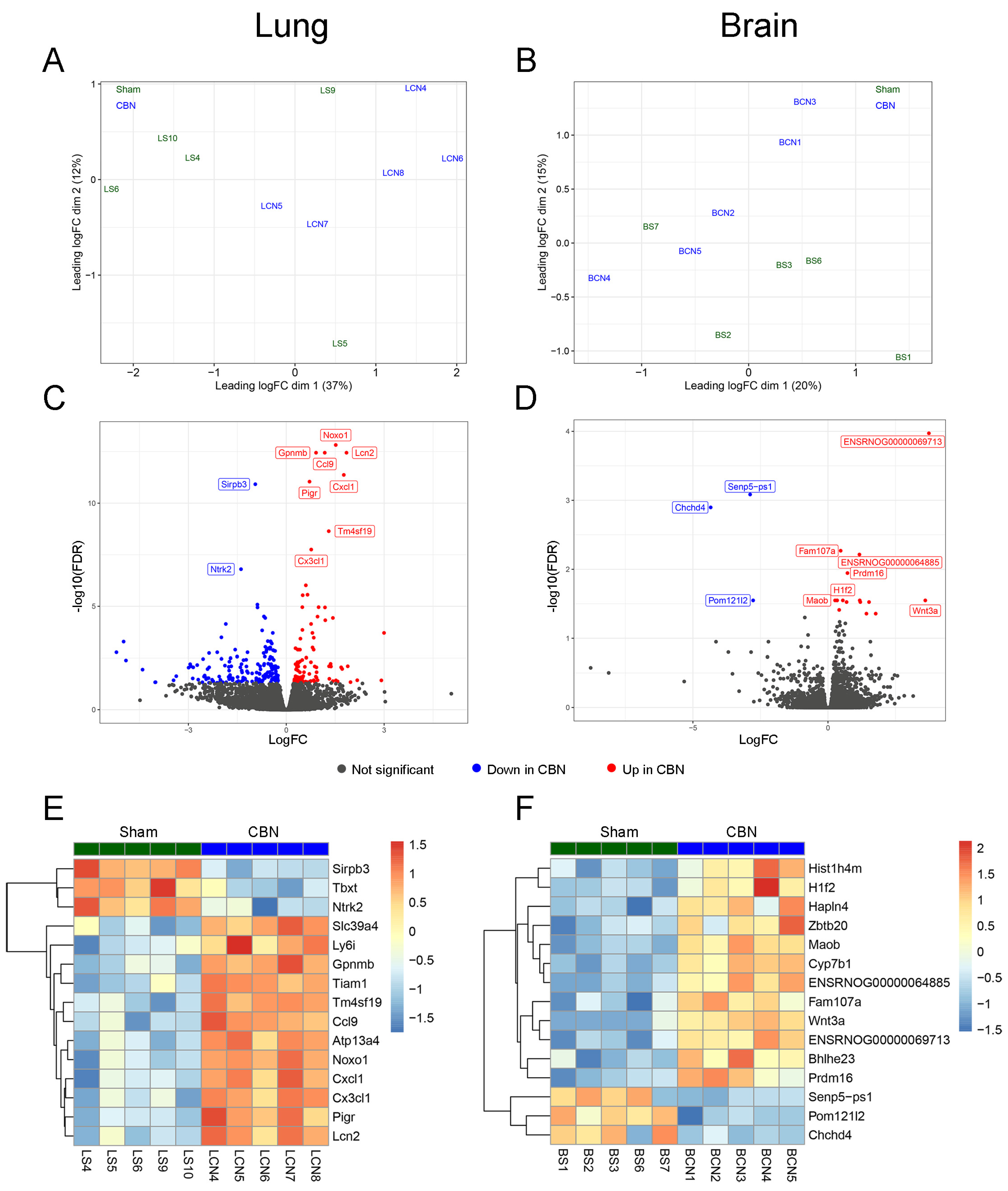
Figure 5.
Summary of gene set enrichment analysis (GSEA) results. GSEA was performed separately in brain and lung tissues using Hallmark, KEGG, and the Saul Sen Mayo (genes upregulated in senescent cells) gene sets. Normalized enrichment scores are shown for all gene sets tested in lung (A) and brain (B) with significantly enriched gene sets colored in red, activated in carbon black naphthalene (CBN)-treated, or blue, suppressed in CBN-treated samples).
Figure 5.
Summary of gene set enrichment analysis (GSEA) results. GSEA was performed separately in brain and lung tissues using Hallmark, KEGG, and the Saul Sen Mayo (genes upregulated in senescent cells) gene sets. Normalized enrichment scores are shown for all gene sets tested in lung (A) and brain (B) with significantly enriched gene sets colored in red, activated in carbon black naphthalene (CBN)-treated, or blue, suppressed in CBN-treated samples).
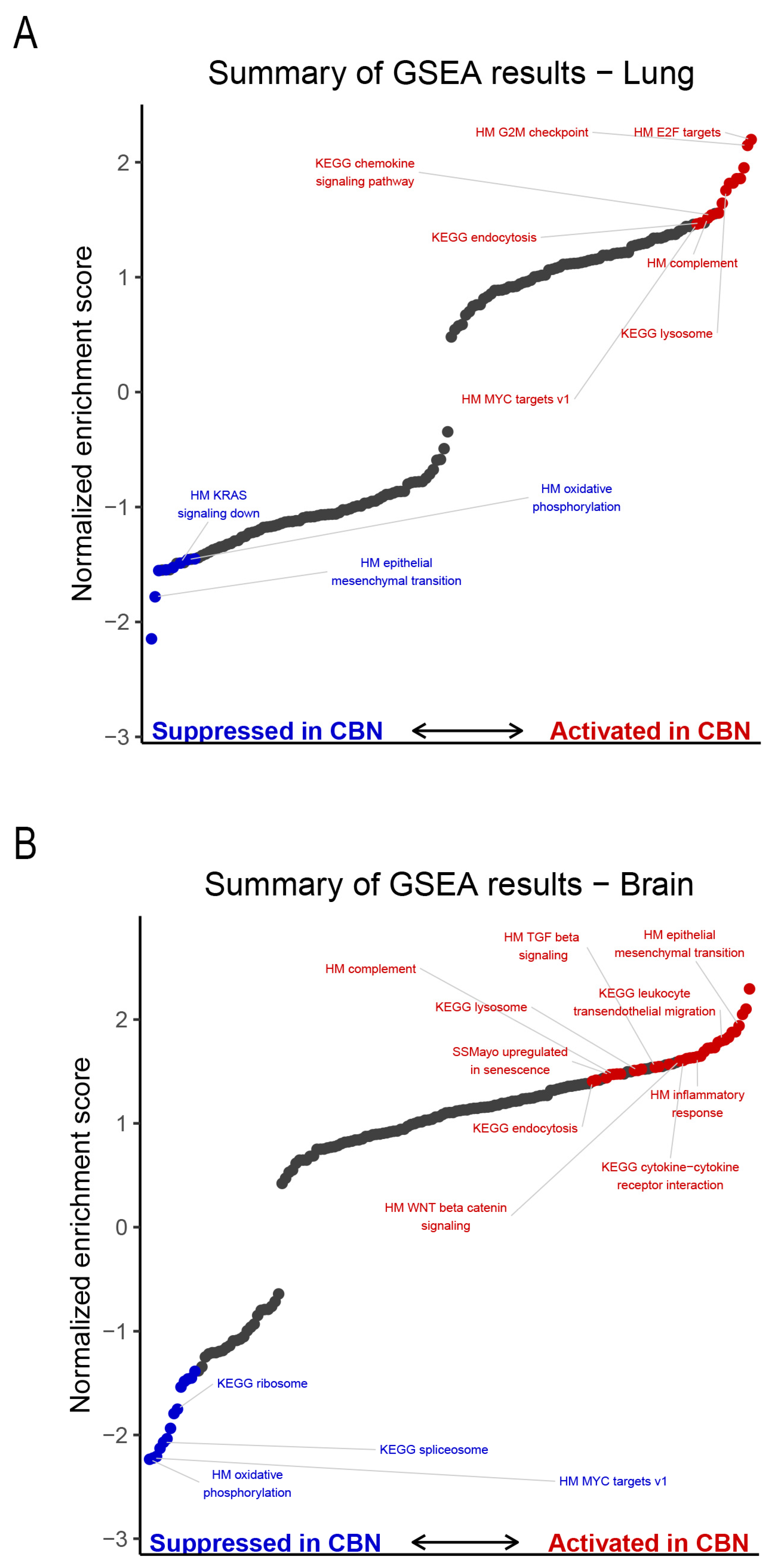
Figure 6.
The Saul Sen Mayo gene set (genes upregulated in senescent cells) was significantly activated in brains of CBN-treated rats. (A) Enrichment plot showing the running enrichment score for genes in the Saul Sen Mayo gene set using a ranked list of all expressed genes based on results of differential expression analysis between CBN-treated and sham control brain tissue. (B) Heatmap showing log₂ fold-changes (log₂FC) of each sample over the mean of the comparison group for leading-edge genes (genes that contribute to the enrichment signal) from the Saul Sen Mayo gene set. Genes are ordered in the heatmap by hierarchical clustering.
Figure 6.
The Saul Sen Mayo gene set (genes upregulated in senescent cells) was significantly activated in brains of CBN-treated rats. (A) Enrichment plot showing the running enrichment score for genes in the Saul Sen Mayo gene set using a ranked list of all expressed genes based on results of differential expression analysis between CBN-treated and sham control brain tissue. (B) Heatmap showing log₂ fold-changes (log₂FC) of each sample over the mean of the comparison group for leading-edge genes (genes that contribute to the enrichment signal) from the Saul Sen Mayo gene set. Genes are ordered in the heatmap by hierarchical clustering.
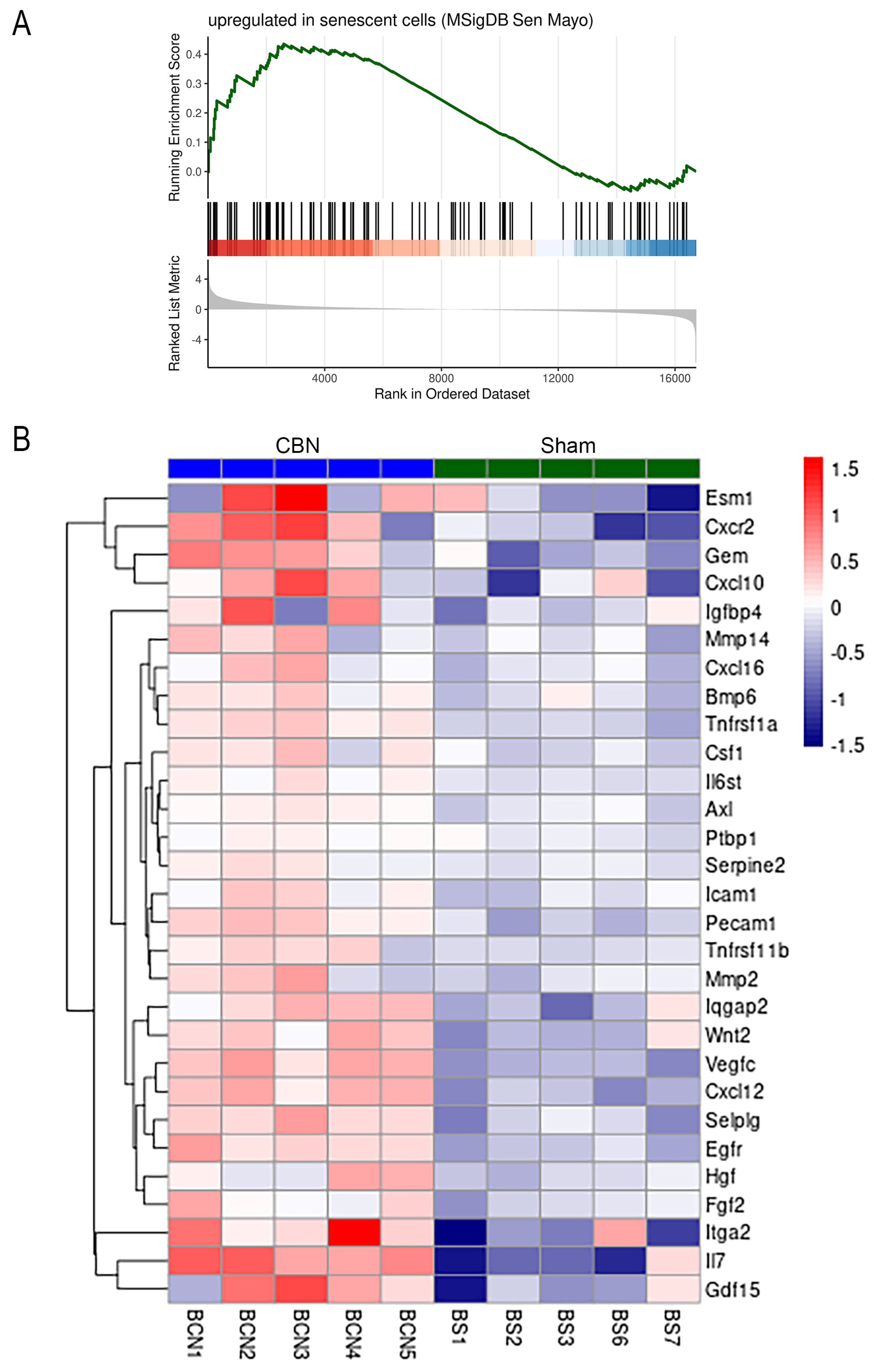
Figure 7.
Nuclear NF κB p65 protein levels are elevated in lung and brain following CBN exposure. Western blot and densitometry analyses of NF κB p65 in nuclear extracts from rat lung (A-B) and brain (C-D) tissues following exposure CBN or sham control. (A) Western blot of NF κB p65 (65 kDa) in lung nuclear extracts from sham and CBN-exposed rats. Total protein stain is shown as a loading control. (B) Quantification of NF κB p65 nuclear abundance in lung shows a significant increase in the CBN-exposed group compared to sham (***p < 0.001). (C) Western blot of NF κB p65 in brain nuclear extracts with total protein loading control. (D) Densitometry analysis reveals a robust increase in brain nuclear NF κB p65 levels following CBN exposure (****p < 0.0001). Densitometry values were normalized to total protein per sample. Data are presented as mean ± SEM.
Figure 7.
Nuclear NF κB p65 protein levels are elevated in lung and brain following CBN exposure. Western blot and densitometry analyses of NF κB p65 in nuclear extracts from rat lung (A-B) and brain (C-D) tissues following exposure CBN or sham control. (A) Western blot of NF κB p65 (65 kDa) in lung nuclear extracts from sham and CBN-exposed rats. Total protein stain is shown as a loading control. (B) Quantification of NF κB p65 nuclear abundance in lung shows a significant increase in the CBN-exposed group compared to sham (***p < 0.001). (C) Western blot of NF κB p65 in brain nuclear extracts with total protein loading control. (D) Densitometry analysis reveals a robust increase in brain nuclear NF κB p65 levels following CBN exposure (****p < 0.0001). Densitometry values were normalized to total protein per sample. Data are presented as mean ± SEM.
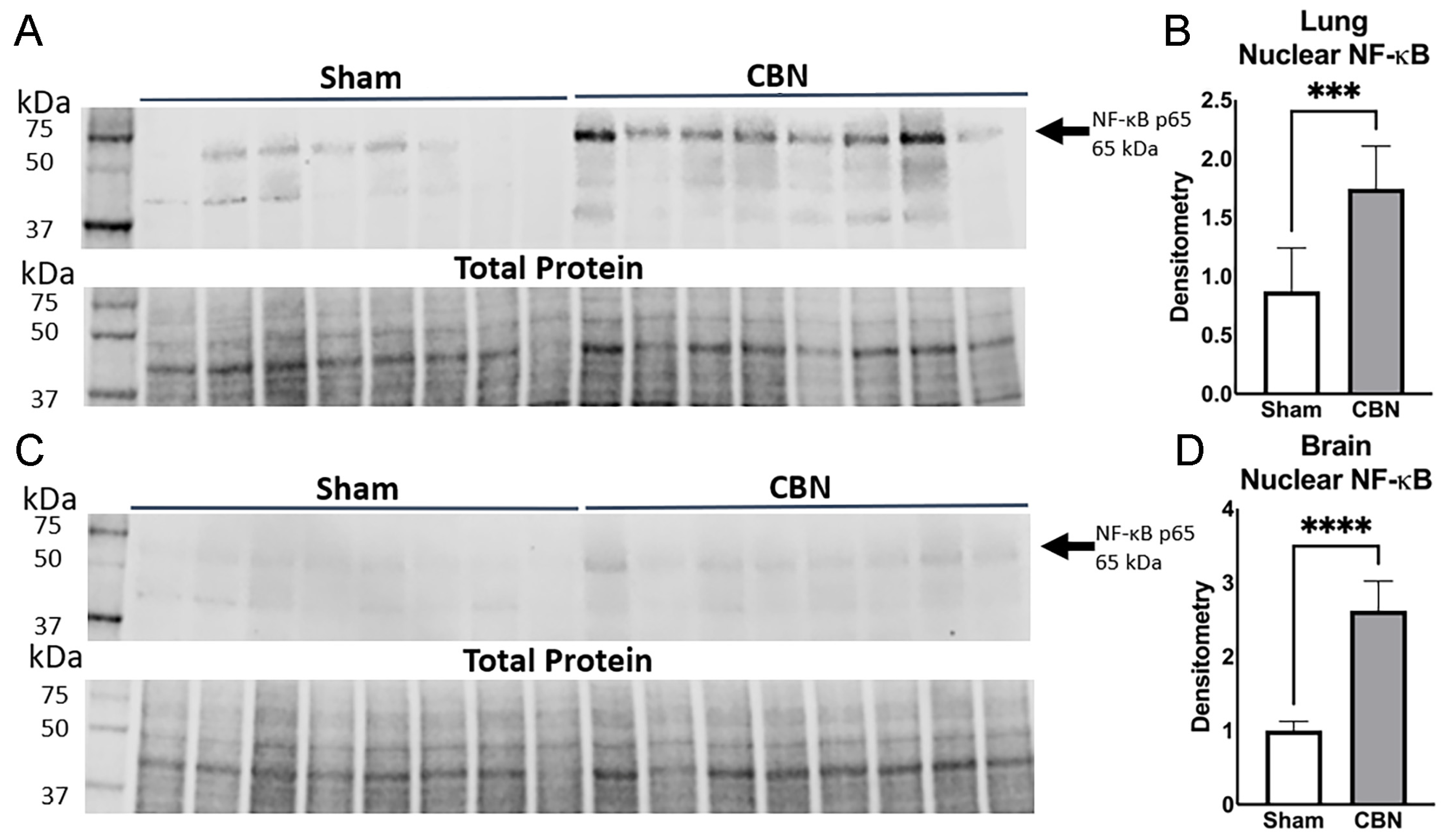
Figure 8.
Elastic net regression model predicts CBN exposure based on gene expression profiles in lung. (A) Binary classification plot showing the estimated probability that each biological replicate belongs to the CBN exposure group versus the sham control, based on elastic net modeling using 278 differentially expressed genes (DEG) between CBN and sham control groups. (B) Added-variable plots illustrate the relationship between gene predictability (x-axis) and standardized expression difference (y-axis) for the final 97 genes retained in the elastic net model. Each point represents a gene, colored by its relative contribution to the model. The top 10 predictive genes are labeled.
Figure 8.
Elastic net regression model predicts CBN exposure based on gene expression profiles in lung. (A) Binary classification plot showing the estimated probability that each biological replicate belongs to the CBN exposure group versus the sham control, based on elastic net modeling using 278 differentially expressed genes (DEG) between CBN and sham control groups. (B) Added-variable plots illustrate the relationship between gene predictability (x-axis) and standardized expression difference (y-axis) for the final 97 genes retained in the elastic net model. Each point represents a gene, colored by its relative contribution to the model. The top 10 predictive genes are labeled.
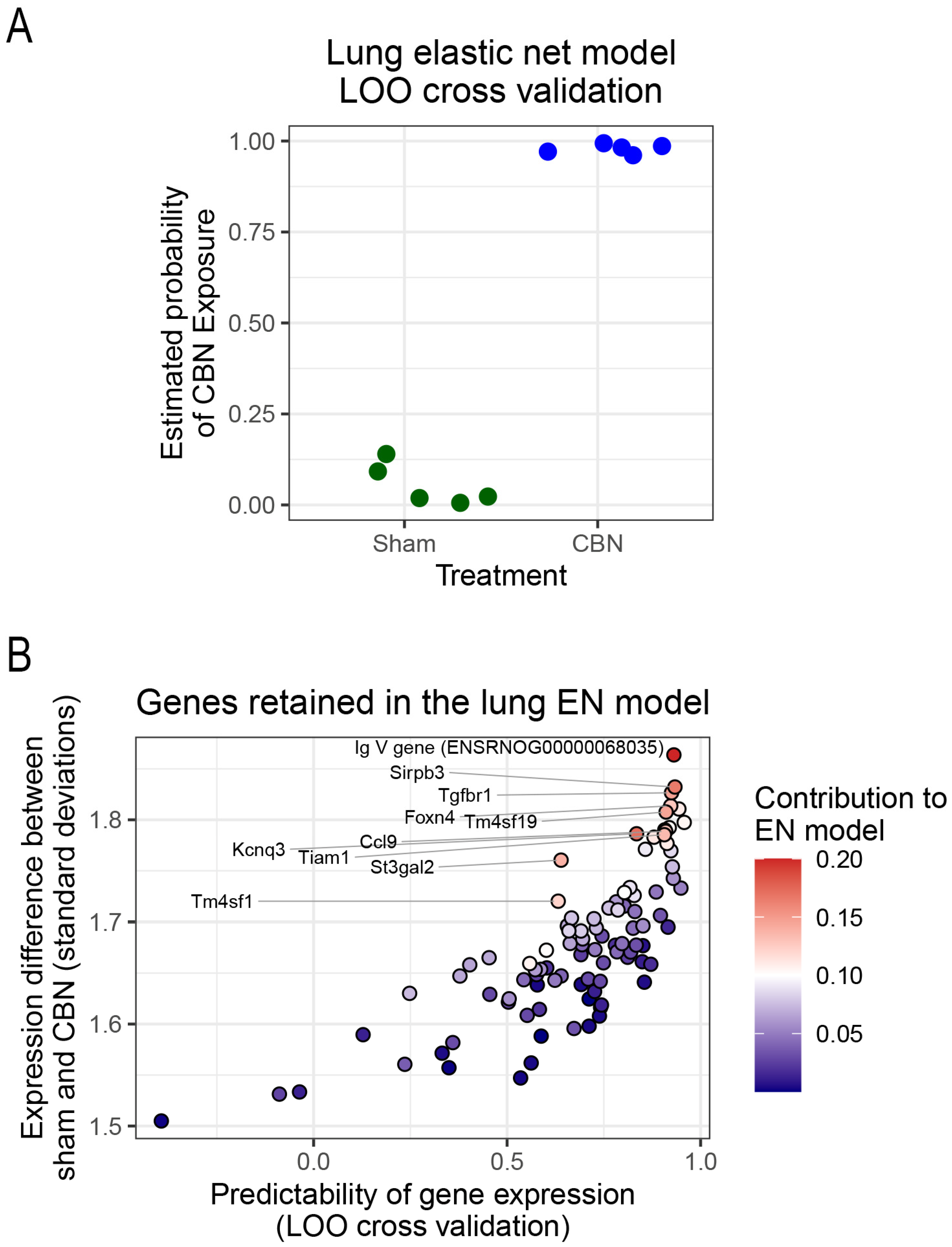

Table 1.
Protein Biomarkers Altered by Inhalation Exposure and Biological Relevance.
| Biomarker | Tissue | NA | CBN | Biological Role / Relevance |
|---|---|---|---|---|
| IL1β | Lung | ↑ | ↑ | IL1β is a proinflammatory cytokine involved in acute lung inflammation with key roles in local and systemic inflammation and is commonly elevated following particulate matter exposure [16,26,27]. |
| IL4 | Lung Plasma Brain |
↑ ↑ ↓ |
↑ ↑ ↓ |
IL4 is a Th2 cytokine that plays a role in allergic airway inflammation, pulmonary hypertension and in regulating neuroimmune signaling [16,28,29]. |
| IL5 | Lung Brain |
↑ ↓ |
↑ ↓ |
IL5 promotes eosinophil activity; upregulated in lung but suppressed in brain, suggesting compartmentalized immune activation [16,30,31]. |
| IL6 | Lung Brain Plasma |
↓ ↓ ↑ |
↓ ↓ ↑ |
IL6 is a pleiotropic cytokine; suppressed in lung and brain, but elevated in plasma during systemic inflammation from inhaled toxicants[32,33,34,35]. IL6 deficiency in acute lung injury is associated with acute respiratory distress syndrome (ARDS) [32,36] |
| IL10 | Lung Plasma Brain |
↑ ↑ ↓ |
↑ ↑ ↓ |
IL10 is an anti-inflammatory cytokine; increased in plasma and lung but decreased in brain, suggesting differential regulation under toxic stress [16,37]. |
| IL13 | Lung Brain |
↑ ↓ |
↑ ↓ |
IL13 mediates mucus production and drives fibrosis and remodeling in lung and has context-dependent pro- or anti-inflammatory effects in the brain [28,38,39]. Decreased IL13 in the CNS is linked to increased neurodegeneration, highlighting how toxicant exposures differentially regulate lung and brain immune responses [39]. |
| IFNγ | Lung Brain |
↑ ↑ |
↑ ↑ |
IFNγ, a Th1 cytokine, plays a dual role in lung and brain toxicity. In lung, it promotes macrophage activation and inflammation, contributing to tissue injury after toxicant exposure and supporting neuroprotective immune regulation [9,16,40]. In brain, IFN-γ both exacerbates neuroinflammation and supports neuroprotective immune regulation [9,16,40]. |
| KC/GRO (CXCL1) | Lung Plasma Brain |
↑ ↑ ↑ |
↑ ↑ ↑ |
KC/GRO (CXCL1) recruits neutrophils, is expressed by macrophages, and is markedly elevated in lung, plasma, and brain after exposures, a hallmark of lung and brain injury [8,41,42,43,44,45,46]. |
| TNFα | Lung Plasma Brain |
↑ | ↑↑ | TNFα is a key proinflammatory mediator that drives lung injury through macrophage activation and neutrophil recruitment and promotes microglial activation and neurodegeneration in the brain following toxic exposures [8,9,16,47,48]. |
| NF-κB (Western Blot) | Lung Brain |
N/A N/A |
↑ ↑ |
NF-κB is a transcription factor activated by oxidative and inflammatory stress, driving lung and brain injury [16,49]. PM exposures upregulate NF-κB in lung, promoting inflammation and epithelial damage [49]. In brain, NF-κB activation contributes to neuroinflammation, metabolic dysfunction, and neurodegeneration [47,50]. |
Table 2.
Top 15 DEGs in Lung and Brain: Relevance to Toxicant-Induced Injury.
| LUNG | ||
|
Gene Symbol Decreased or Increased |
Full Gene Name | Biological Function and Disease Relevance |
| Sirpb3 | Signal-regulatory protein beta-3 | Sirpb3 is a non-coding pseudogene in the signal regulatory protein (SIRP) family, which includes receptors expressed on myeloid cells. Although it does not encode a protein, Sirpb3 may influence immune signaling through RNA-based mechanisms, similar to SIRPB1, which promotes myeloid activation and inflammation[51]. Persistent activation of SIRP-related pathways can amplify myeloid signaling, driving chronic inflammation, impaired epithelial repair, tissue damage, and fibrosis. In contrast, reduced expression may impair phagocytic clearance and antigen resolution, increasing susceptibility to infection and promoting maladaptive lung remodeling. [51]. |
| Tbxt | T-box transcription factor T; alias, brachyury | Fundamentally important for lung development, reactivation in adult tissue is associated with lung injury responses, fibrosis via TGF-β, Wnt signaling, and cancer, influencing tissue remodeling and repair outcomes [52,53]. Downregulation of Tbxt in the lung may impair epithelial regeneration and repair of injured airways, leading to persistent epithelial damage. It also disrupts necessary matrix remodeling, which can result small airway injury and contribute to chronic inflammation and may increase the risk of diseases like bronchiolitis obliterans and early-stage COPD [54,55]. |
| Ntrk2 | Neurotrophic receptor tyrosine kinase 2 | Encodes for neurotrophic receptor tyrosine kinase 2 (TrkB). Reduced expression of Ntrk2 (TrkB) may impair neurotrophic signaling and epithelial repair, potentially hindering recovery from lung injury. In contrast, increased Ntrk2 expression is associated with aggressive tumor behavior, enhanced metastasis, and poor survival in lung adenocarcinoma [56,57]. |
| Slc39a4 | Solute carrier family 39 (zinc transporter), member 4; alias, Zrt like protein-4 (Zip4) | Encodes for zinc transporter-4 (Zip-4) that is overexpressed in non-small cell lung cancer, where it promotes metastasis by activating epithelial-mesenchymal transition (EMT)-related pathways [58]. ZIP4 promotes NSCLC progression and metastasis by upregulating the snail-N-cadherin signaling axis, thereby facilitating EMT and enhancing cell migration and invasion[59]. |
| Ly6i | Lymphocyte antigen 6 complex, locus I | Surface marker selectively expressed in a dysfunctional subpopulation of alveolar type II cells in emphysematous lungs. Ly6i+ cells exhibit impaired regenerative capacity, increased senescence, and inflammation, contributing to progressive tissue damage in COPD [60]. Its presence may lead to long-term respiratory decline and increased risk of chronic lung failure [60]. |
| Gpnmb | Glycoprotein non-metastatic melanoma protein B | Transmembrane protein expressed in injured lung epithelial and immune cells, where it regulates cell adhesion, immune response, and tissue repair. In lung injury and cancer, it promotes tumor growth and invasion through membrane signaling and its shed ectodomain, activating integrins and MMPs. Up-regulation of Gpnmb in lung leads to its accumulation in fibrotic ECM. Enhances fibroblast proliferation, migration, and fibrotic protein production, contributing to progressive tissue remodeling [61,62]. |
| Tiam1 | T-cell lymphoma invasion and metastasis 1 | Regulates cell migration and cytoskeletal remodeling through Rac1 signaling. Tiam1 may also influence fibroblast activation and endothelial barrier function during lung injury[63,64]. Long-term dysregulation of Tiam1 affects lung health in a disease-dependent manner. Overexpression promotes tumor invasion and metastasis in both small cell and non-small cell lung cancer (NSCLC), while reduced expression may impair tissue repair and endothelial barrier integrity [63,64]. |
| Tm4sf19 | Transmembrane 4 L six family member 19; alias, transmembrane 4L | Transmembrane protein expressed in lung epithelial cells, where it participates in signal transduction, transcriptional regulation, and response to oxidative stress. It influences GABP-mediated YAP signaling pathways in maintaining epithelial integrity [65]. May also regulate immune cell interactions, contributing to immune surveillance and inflammation in the lung tissue[66]. Altered Tm4sf19 expression is linked to NSCLC tumor progression and may also contribute to inflammation or epithelial dysfunction in COPD and lung fibrosis. Its effects depend on cellular stress and immune signaling, potentially protecting or worsening disease based on context [65,66]. |
| Ccl9 | Chemokine (C-C motif) ligand 9; alias, macrophage inflammatory protein-1 gamma (MIP-1γ) | Chemokine expressed in lung tissue that recruits monocytes, macrophages, and dendritic cells via the CCR1 receptor[67] to support innate immune responses during lung injury or infection [68]. Long-term dysregulation of Ccl9 in the lung contributes to chronic inflammation and immune-driven tissue remodeling. Its persistent upregulation is associated with fibrosis and may worsen outcomes in diseases like asthma, COPD, and pulmonary fibrosis. Ccl9 may also amplify lung responses to environmental exposures, promoting progressive damage over time [69,70]. |
| Atp13a4 | ATPase 13A4 | P-type ATPase that transports polyamines and cations using ATP hydrolysis; helps maintain ion balance and may support epithelial integrity, stress responses, and signaling [71]. Its regulation of calcium and polyamines suggests a role in oxidative stress and tissue remodeling [72]. Atp13a4 amplification has been linked to lung cancer prognosis [72]. Dysregulation of polyamine or ion transport via ATP13A4 could also contribute to epithelial dysfunction [72]. |
| Noxo1 | NADPH oxidase organizer 1 | Regulates ROS generation in lungs; elevated in response to air pollution and oxidative injury [73]. Oxidative stress regulator implicated in fibrosis and progressive lung disease such as COPD [74] |
| Cxcl1 | C-X-C motif chemokine ligand 1; alias, KC/GRO | Chemokine that recruits neutrophils to inflamed lung tissue; key in early lung immune response to toxins[75,76]. |
| Cx3cl1 | C-X3-C motif chemokine ligand 1; alias, fractalkine | Chemokine expressed by lung epithelial, endothelial, and smooth muscle cells in both membrane-bound and soluble forms. It regulates monocyte and macrophage recruitment through the receptor Cx3cr1, supporting immune cell adhesion, cytokine release, and tissue remodeling [77,78]. In chronic lung diseases like COPD, asthma, and IPF, dysregulated CX3CL1 expression drives persistent macrophage-mediated inflammation and fibrosis, contributing to progressive tissue damage [77,78]. Elevated protein expression in lung tissue demonstrated in preclincal burn pit models [16]. |
| Pigr | Polymeric immunoglobulin receptor | Immunoglobulin receptor (PIGR) transports IgA and IgM across lung epithelial cells to generate secretory IgA at the mucosal surface, where it neutralizes inhaled pathogens and toxins. PIGR expression is regulated by inflammatory cytokines, enabling immune defense and limiting immune response [79,80,81]. Loss of PIGR regulation may increase susceptibility to infection and chronic respiratory illness. Overexpression in the lung often reflects epithelial stress or immune activation, as observed in COPD, asthma, IPF, and lung cancer. While initially protective, sustained PIGR elevation can drive epithelial remodeling, disrupt immune tolerance, and promote chronic inflammation, contributing to airway hyperreactivity and fibrosis [80,81] |
| Lcn2 | Lipocalin-2 | Iron-binding glycoprotein produced by lung epithelial and immune cells, playing key roles in inflammation, innate immunity, and iron regulation. While it helps protect against bacterial infection by sequestering iron, excessive LCN2 expression contributes to chronic inflammation, oxidative stress, and lung tissue damage [82,83]. Upregulated in both COPD and constrictive bronchiolitis, contributing to inflammation, immune dysregulation, and airway structural damage. Elevation can lead to iron imbalance and oxidative stress, chronic tissue injury and impaired repair. Can promote airway remodeling, fibrosis, and lung function decline, increasing the risk of lung cancer [82,84,85]. |
| BRAIN | ||
| Gene Symbol (Accession) | Full Gene Name | Biological Function and Disease Relevance |
| Hist1h4m | Histone cluster 1 H4 family member | Histone protein linked to epigenetic modulation under stress; altered in neurotoxic exposure models [86,87,88]. H4-mediated epigenetic alteration may contribute to memory deficits and aging-related brain dysfunction [86,87,88]. |
| H1f2 | H1 histone family member 2 | Histone protein involved in chromatin structure; dysregulation linked to neurodegenerative changes [86,88,89]. Altered expression found in neuroinflammatory and cognitive impairment models [86,88,89]. |
| Hapln4 | Hyaluronan and proteoglycan link protein 4; alias, brain link protein 2 (Bral2) | Encodes an extracellular matrix protein that links hyaluronic acid to proteoglycans, supporting synaptic architecture and stability [90]. Enriched at GABAergic synapses, Hapln4 plays critical roles in nervous system development, synaptic maturation, and maintaining extracellular homeostasis [90]. Elevated Hapln4 expression can impair synaptic transmission, restrict neuroplasticity, and promote neuroinflammation following injury or toxicant exposure, increasing vulnerability to cognitive dysfunction and neurodegenerative diseases [91,92]. |
| Zbtb20 | Zinc finger and BTB domain containing 20 | Transcription factor essential for brain development, regulating neuronal differentiation, synaptic plasticity, and memory formation [93]. It also modulates oxidative stress and inflammatory responses, positively regulating pro-inflammatory cytokines during neuroinflammation and injury [94]. Linked to Alzheimer’s pathology and cognitive decline after toxic exposures [95,96]. |
| Maob | Monoamine oxidase B | Encodes monoamine oxidase B, a mitochondrial enzyme that degrades neurotransmitters like dopamine and serotonin, regulating neurotransmitter balance and producing hydrogen peroxide[97,98,99]. Its activity, enriched in astrocytes and neurons, links it to oxidative stress in the brain [100]. Dysregulated MAOB activity leads to elevated oxidative stress, mitochondrial dysfunction, and neuronal damage [100]. Upregulation of Maob has been implicated in neurodegenerative diseases such as PD and AD, contributing to cognitive decline, neuroinflammation, and progressive dopaminergic neuron loss [100]. |
| Cyp7b1 | Cytochrome P450 family 7 subfamily B member 1; alias, oxysterol 7-alpha-hydroxylase brain | Encodes a cytochrome P450 enzyme that plays a crucial role in the metabolism of neurosteroids and cholesterol-derived molecules within the brain[101]. It helps regulate the synthesis of specific oxysterols and neuroactive steroids, impacting neuronal protection, signaling, and inflammation. Dysregulation of Cyp7b1 can increase vulnerability to neurodegeneration, inflammation, and lipid imbalances seen in conditions like Alzheimer’s disease [101,102,103,104]. |
|
ENSRNOG00000064885 (only Ensembl ID available) |
Predicted Ensembl rat gene | Predicted to encode a ribosomal protein that maintains ribosome integrity, facilitates translation, and participates in ribonucleoprotein complexes essential for protein synthesis [105]. Ribosomal dysfunction may impair neuronal protein synthesis and plasticity, increasing susceptibility to oxidative stress, inflammation, and neurodegeneration after toxicant exposure [106]. |
| Fam107a | Family with sequence similarity 107 member A; alias, Down-Regulated in Renal Cell Carcinoma 1 (Drr1) | Stress-inducible, actin-binding protein that regulates cytoskeletal dynamics, supporting synaptic function, neural cell survival, and adaptation to stress [107]. Fam107a also modulates microglial activation, linking neural stress responses to neuroinflammation [108]. Sustained upregulation supports neural protection and reduced inflammation, but chronic dysregulation may cause maladaptive inflammation or synaptic dysfunction. Balanced Fam107a expression is crucial for long-term brain health [108]. |
| Wnt3a | Wingless/Intergrated (Wnt) family member3A |
Protein coding gene for Wnt3A glycoprotein involved in Wnt signaling. Plays a vital role in brain development and function by promoting neurogenesis, supporting neuronal differentiation and synaptic plasticity, protecting against oxidative stress, and maintaining neural stem cell niches [109]. Wnt3a acts as a double-edged sword in the adult brain, supporting repair, neurogenesis, and immune regulation under balanced activation, but potentially exacerbating neuroinflammation and degeneration when overactivated, particularly in chronic disease or toxic exposures [110]. |
|
ENSRNOG00000069713 (only Ensembl ID available) |
Predicted Ensembl rat gene | Predicted to function as a long noncoding RNA (lncRNA), Unknown experimentally, but typical lncRNAs regulate gene expression, chromatin remodeling, RNA stability, and may affect ribosome-related processes indirectly [111]. Dysregulation of lncRNAs may impact gene expression to impair memory, plasticity, and promote neurodegeneration after toxicant exposures [112,113]. |
| Bhlhe23 | Basic helix-loop-helix family member e23; alias Dec2 | Transcription factor crucial for neuronal differentiation and retinal development and may also influence circadian rhythm regulation in the brain. Its activity helps shape the development and specialization of certain neuron types [114,115]. Upregulation of Bhlhe23 may increase orexin neuron activity, reducing sleep duration and extending wakefulness. This can lead to impaired cognition, mood disturbances, and increased risk of neuroinflammation and neurodegeneration over time [114]. |
| Prdm16 | PR/SET domain 16 | Transcriptional regulator involved in neurogenesis and the maintenance of neural stem cells [116]. It promotes neuronal differentiation and protects against oxidative stress, supporting normal brain development and function[117]. Dysregulation of Prdm16 in the brain may compromise neurogenesis, increase oxidative stress susceptibility, and negatively affect cognition and long-term brain health, especially under stress or environmental challenge [17,118,119]. |
| Senp5-ps1 | Small ubiquitin-like modifier (SUMO) protease 5 pseudogene | Pseudogene variant of Senp5; non-functional transcript, may regulate their corresponding genes by acting as molecular decoys for microRNAs or generating noncoding RNAs that influence gene expression [120]. Pseudogenes of SUMO proteases may influence neurodegeneration and neuroinflammation by modulating gene expression linked to neuronal survival and cellular stress responses [121]. Changes in their activity could alter susceptibility to neurodegenerative and neuroinflammatory diseases [122]. |
| Pom121L2 | POM121 transmembrane nucleoporin-like 2 | Predicted to encode a nucleoporin-like protein that binds nuclear localization sequences and acts as a structural constituent of the nuclear pore complex. It likely facilitates RNA export and protein import between the nucleus and cytoplasm [123]. Disruption of nuclear pore complex proteins impairs nucleocytoplasmic transport, leading to blood-brain barrier dysfunction, oxidative stress, and chronic neuroinflammation, which collectively increase the risk of neurodegeneration. [124,125]. In a glioma model, Pom121L2 was found to be downregulated in the peritumoral brain region characterized by neuroinflammation and epileptogenic activity [126]. |
| Chchd4 | Coiled-coil-helix-coiled-coil-helix domain containing 4; alias, Mitochondrial intermembrane space import and assembly protein 40 (Mia40) | Essential for mitochondrial protein import; mitochondrial respiration and redox regulation[127], lipid metabolism[128], downregulation suggests energy metabolism deficits in brain [129]. Linked to impaired neuronal energy supply and mitochondrial dysfunction in brain disease[130] |
Table 3.
Lung Elastic Net Predictive Gene Panel – Top 10 Contributing Genes.
| Gene Symbol |
Full Gene Name | Biological Function and Disease Relevance |
|---|---|---|
| IgV gene | Immunoglobulin variable region | Predicted transcript, IgV gene expression in lung injury reflects the activation of the adaptive immune system, particularly B cell recruitment and antibody production [146,147]. While activation of IgV may be protective, sustained IgV upregulation is associated with chronic inflammation, tissue remodeling, and fibrosis, as observed in lung diseases such as COPD and interstitial lung disease (ILD) [146,147]. |
| Kcnq3 | Potassium voltage-gated channel subfamily Q member 3 | Potassium channel subunit that regulates membrane potential and sensory neuron excitability in the lung, influencing neuropeptide release, inflammation, and lung injury severity[148].Kcnq3 dysregulation can lead to chronic sensory neuron hyperexcitability, promoting airway hypersensitivity, inflammation, and persistent cough. Long-term, this may contribute to airway remodeling and worsening of chronic lung diseases such as asthma or COPD [148]. |
| Sirpb3 | Signal-regulatory protein beta-3 | Sirpb3 is a pseudogene in the signal regulatory protein (SIRP) family, which includes transmembrane receptors involved in myeloid cell signaling [149]. Though non-coding, Sirpb3 may regulate innate immune responses through RNA-based mechanisms [150]. Based on its similarity to Sirpb1 which promotes myeloid activation and inflammation Sirpb3 may influence immune signaling and contribute to lung inflammatory states [149,150,151]. |
| Tgfrb1 | Transforming growth factor-beta receptor type 1 | Receptor for TGF-β signaling involved in immune regulation and fibrosis; controls lung cell differentiation and matrix production, activation promoting inflammation [152,153].Persistent Tgfbr1 activation promotes chronic inflammation, subepithelial fibrosis, smooth muscle thickening, and airway remodeling; impairs epithelial repair and regeneration, leading to fixed airflow obstruction associated with ConB and progressive disease in IPF, COPD, and asthma [154,155]. |
| Tm4sf19 | Transmembrane 4 L six family member 19 | Tetraspanin-like membrane protein that alters lysosomal degradation in macrophages, oxidative stress, and intracellular lipid content and cellular adhesion and immune signaling lysosomal activity in macrophages [156,157]. Dysregulation of Tm4sf19 may impair macrophage lysosomal function and promote oxidative stress, contributing to chronic inflammation, defective lipid clearance, and persistent immune activation processes that underlie airway remodeling and progression of chronic lung diseases such as COPD [156,158]. |
| St3Gal2 | ST3 beta-galactoside alpha-2,3-sialyltransferase 2 | Functions as a sialyltransferase enzyme that catalyzes the addition of sialic acid to glycoproteins and glycolipids, specifically in an α2,3 linkage to galactose residues [159]. This sialylation regulates protein stability, cell signaling, and immune cell interactions. Elevated ST3GAL2 in fibrotic lungs coincides with reduced sialylation, disrupting glycoprotein function and promoting fibrosis [160]. |
| Tiam1 | T-cell lymphoma invasion and metastasis-inducing protein 1 | Cell migration, cytoskeletal remodeling, Rac1 signaling, [64], promotes small cell lung cancer[63]. In the lung, elevated Tiam1 expression has been linked to enhanced invasiveness and metastasis in small cell lung cancer (SCLC). Its dysregulation may also contribute to abnormal epithelial remodeling in chronic lung disease [63]. |
| Ccl9 | Chemokine (C-C motif) ligand 9 | Functions in the lung as a chemokine primarily produced by myeloid cells such as macrophages and dendritic cells. It promotes the recruitment of immune cells through its receptor CCR1, contributing to inflammatory responses and shaping the immune microenvironment [161,162] . It promotes recruitment of dendritic cells and other leukocytes, amplifying inflammatory responses in diseases like asthma [161]. In cancer, especially lung metastasis models, CCL9 induced by TGF-β signaling in myeloid cells enhances tumor cell survival and creates a pro-metastatic lung microenvironment by recruiting immunosuppressive cells [162]. |
| Foxn4 | Forkhead box N4 | Transcription factor expressed in proximal airway epithelial cells during lung development; its loss impairs alveologenesis, leading to enlarged alveoli, thin walls, and poor septation[163,164]. Foxn4 is critical for normal lung development, with its loss leading to impaired alveolar structure and maturation. In lung adenocarcinoma, Foxn4 acts as a tumor suppressor by inhibiting TGF-β signaling and epithelial-mesenchymal transition. Its downregulation promotes tumor progression and inflammation via neutrophil polarization [165]. |
| Tm4sf1 | Transmembrane 4 L six family member 1 |
Tetraspanin-like membrane protein that serves as a marker of Wnt-responsive alveolar epithelial progenitor (AEP) cells, contributing to alveolar repair and regeneration after lung injury [66,166]. Supports lung repair by marking Wnt-responsive alveolar progenitor cells involved in regenerating alveolar epithelium after injury [166]. However, in lung cancer particularly NSCLC its overexpression promotes tumor growth, invasion, and epithelial-mesenchymal transition via PI3K/AKT and JAK/STAT signaling. This dual role makes TM4SF1 both a regenerative marker and a potential oncogenic driver [66]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



