
Submitted:
05 June 2025
Posted:
11 June 2025
You are already at the latest version
Abstract
Climate change poses significant threats to the environment and agricultural productivity. To address these challenges, we aimed to enhance tolerance strategies to salinity stress by applying potassium and calcium to two-year-old potted olive plants grown under saline conditions. To better understand the mitigation of the detrimental effects of salinity stress, we elucidated the interaction between physiological and biochemical responses in olive plants. The results demonstrated that the application of calcium and potassium under salinity stress triggered an adaptive physiological response, notably enhancing photosynthetic efficiency, transpiration rate, and stomatal conductance, especially with the combined treatment of 40 mM KNO₃ and 10 mM CaCl₂. Under salinity conditions, the levels of neoxanthin and violaxanthin decreased, while their increase was strongly associated with higher potassium and calcium concentrations. Under stress conditions, the limited photosynthetic efficiency increase sugar levels that may serve as part of the plant’s adaptive strategy to cope with stress. A negative correlation demonstrated between the photosynthetic rate and osmoprotectant levels. Meanwhile, the positive interaction was depicted among the effective quantum yield of photosystem II (PSII), stomatal conductance, and the photosynthetic rate, underscoring the crucial role of potassium and calcium treatments in maintaining plant physiological and biochemical processes under salt stress.
Keywords:
Climate change
; Calcium
; Olive tree
; Osmoprotectants
; Potassium
; Salt stress
1. Introduction
Climate change is predicted to influence biodiversity, food security, and water purity [1]. Temperature, precipitation, and wind are key climate factors affecting soil formation and subsequent development. Studies have shown that climate change increases salinization, which could affect soil characteristics [2], with over 7% of global land impacted by salinity [3]. Many studies have investigated the negative impacts of salinity on plant metabolic and physiological mechanisms [4], related to the duration of salt exposure, concentrations, and cultivar [5]. Due to water scarcity, Mediterranean areas frequently use saline water for irrigation in olive groves, as olive trees have moderate tolerance to salinity [6]. Currently, Tunisia faces a significant challenge for sustainable agriculture due to critical water shortages, which must be carefully managed to maintain olive production under these difficult conditions, particularly in arid and semi-arid regions. The limited olive trees are often irrigated with brackish water, especially in Tunisia's coastal and arid areas [7]. It is well documented that the controlled application of saline water can enhance olive tree production within specific salinity thresholds [8]. However, the critical situation of climate change worsens water scarcity for irrigation, leading to an increase in saline lands [9].
Based on these challenges, plants exhibit several adaptive metabolic and physiological mechanisms as defense strategies to mitigate the harmful effects of salinity stress. The adaptation strategy was depicted through the tolerance to osmotic stress by an increase in leaf area explained by Na+ exclusion from roots and leaf blades, as well as tissue-specific tolerance to accumulated ions, particularly in response to elevated levels of sodium or, in certain species, chloride (Cl-). These physiological adaptations contribute to coping under saline conditions [8]. Several research studies have demonstrated the importance of nutrients as a supplement to mitigate the effects of salt stress in various crops, including olive trees [10], as well as other crops [11]. Calcium and potassium are essential nutrients that play pivotal roles in plant health and stress mitigation [11]. Calcium is known to strengthen cell walls, regulate ion transport, and improve plant-signaling responses. In contrast, potassium facilitates stomatal regulation and enzyme activation, which are vital for optimal photosynthesis [12].
Olive (Olea europaea L.) is an essential agricultural crop with significant economic and nutritional values, particularly in Mediterranean regions. However, environmental stressors, particularly salt stress, are intensified by climate change. Rising temperatures and shifting precipitation patterns contribute to the salinization of soil and water resources, ultimately reducing agricultural yields and threatening food security. It was previously revealed that the salinity affects the growth of olive trees with an observed significant reduction of the internodes and marked small leaves noted by a thickened mesophyll and cell wall, as well as a decline in blooming leading to a lowering fruit number [13]. Indeed, after a long period of salt stress, the main adverse effects revealed on plants through the abscission of leaves with marked chlorosis and necrosis [14]. The dropping leaves' response to salinity stress was explained as the last defense mechanism against high salt concentrations, marked by a reduction in transpiration rate resulting in an accumulation of toxic ions in the leaves, like Na and Cl [15]. Overall, olive trees are moderately tolerant to salinity, with a marked difference in morphological and pomological traits among cultivars, influencing their adaptability and productivity [16].
Despite the identified strategies to mitigate the salinity stress on different crop productivity, still, the defence and mitigation mechanisms are not well understood, particularly in olive trees. To achieve this objective, we revealed the physiological and biochemical mechanisms in olive plants (cv. Arbequina I18) grown under saline conditions alleviated with two levels of K (10 and 40 mM KNO3) and Ca (10 and 40 mM CaCl2). Understanding alleviation mechanisms through studying biochemical and physiological interactions will develop sustainable agricultural practices to boost olive production in a challenging environment.
2. Materials and Methods
2.1. Plant Material and Experimental Design
The experiment was carried out in a research greenhouse under controlled conditions to reveal the physiological and biochemical mechanisms of the olive plant according to our previous experiment set [10]. Two-year-old own-rooted olive trees were grown in 20 L plastic pots filled with a sand-perlite mixture (1:1). To alleviate the salinity stress, three levels (0, 10, and 40 mM) of supplementary KNO3 and CaCl2 were applied to olive plants irrigated with saline solutions for 23 months. The experiment was designed as a randomized complete block with ten replicates. Each block had two rows of five pots each (see Figure 1 for more details).
2.2. Gas Exchange Measurements and Water Use Efficiency (WUE)
At the end of the experiment, net photosynthetic rates (A), stomatal conductance (Gs), and transpiration rate (E) were recorded for each plant on three to five well-exposed and fully expanded leaves using a portable photosynthesis system (LI-COR Inc. 6200).
Water use efficiency (WUE) was measured as an A/E ratio. These measurements were taken on a sunny day from 8:00 to 10:00 AM, with an average temperature of 28 ± 2 °C and the Photosynthetic Active Radiation between 800-1400 μmol m-2 s-1, ambient relative humidity, and CO2 concentration.
2.3. Modulated Chlorophyll Fluorescence Analyses
Modulated Chl fluorescence measurements were made in attached leaves in the plastic greenhouse with a PAM2100 portable fluorometer (H. Walz, Effeltrich, Germany). F0 was measured by switching on the modulated light at 0.6 kHz; PPFD was less than 0.1 μmol m-2 s-1 at the leaf surface. Fm was measured at 20 kHz with a 1s pulse of 6000 μmol photons m-2 s-1 of white light. The measure was performed according to Genty et al. [17]. Non-photochemical quenching (NPQ) was calculated according to Bilger and Björkman [18]. This experiment was recorded by the end of the experiment and is similar to the photosynthesis measurement with the same fully expanded leaves.
2.4. Photosynthetic Pigment Composition
Leaf disks were taken from the same area of the leaves in which gas exchange and modulated Chl fluorescence were measured [19]. Pigment extracts were thawed on ice, filtered through a 0.45μm filter, and analysed by an isocratic HPLC method.
2.5. Soluble Sugar and Proline
Proline contents were extracted from the fresh tissues of roots, stems, and leaves with 4 mM sulphosalicylic acid and centrifuged at 3000g for 30 min. At the end of the experiment, the proline level was measured according to Bates et al. [20].
Soluble sugar was estimated according to the Robyt and White [21] method. The fresh olive leaves, stems, and roots were boiled for 30 min in 80% methanol at 70 °C. The obtained extract was combined with phenol in the range of 1:1 and mixed with 5 mL of concentrated sulphuric acid. After agitation and cooling, the absorbance was determined using a spectrophotometer at 640 nm. The soluble sugar concentration was determined using glucose solutions to develop a standard curve.
2.6. Statistical Analysis
Data collected during the experiment were subjected to one-way analysis of variance (ANOVA) using the PC software package SPSS (version 20.0; SPSS Inc., Chicago, IL, USA). The mean values were compared using Duncan’s multiple range test (P < 0.05).
3. Results
3.1. Gas, Exchange, and Water Use Efficiency
Salinity stress influenced the physiological key mechanisms of olive plants, especially the photosynthesis rate, stomatal conductance, and transpiration, with an average reduction of 70%, 36.7%, and 44.5%, respectively, compared to the control. Thus, the water use efficiency was marked by an average decrease of 44.3% under both salinity doses (Figure 2). Overall, KNO3 or CaCl2 enhanced the physiological mechanisms of olive plants under saline treatments. It was noted that the best alleviation was induced by 40 mM KNO3 and 10 mM CaCl2, which was marked by an enhancement of photosynthetic rate and stomatal conductance by an average increase of 73.4% and 31%, respectively, under 200 mM NaCl. The same applied doses of potassium and calcium induced a medium effective impact for improving transpiration rates and Water Use Efficiency by a respective increase of 50.25 and 45.65% under 200 mM NaCl (Figure 2).
3.2. Impact on Chlorophyll Fluorescence
The control of olive leaves had the maximal photochemical efficiency of PSII measured as Fv/Fm values of 0.8. Salinity stress significantly decreased this ratio with registered values of 0.76 and 0.73 at 100 and 200 mM NaCl, respectively (Figure 3 A). The application of potassium and calcium alleviated the salinity stress, in which the Fv/Fm value was markedly approached to the control value through the application of 40 mM K under a salt concentration of 100 mM (Figure 3A). The effective quantum yield of PSII (ΦPSII) was influenced by salinity with a significant decrease of 44% and 62% at 100 mM and 200 mM NaCl, respectively (Figure 3 B). This effect was related to a substantial reduction in the efficiency of excitation energy capture by open PSII reaction centers, which was depicted by an average decrease of 40.5% for Φexc and photochemical quenching (qP) under salinity doses. In contrast, plants cultivated in the presence of K and Ca showed a slight decrease in the amount used and captured within the electron transfer chain (Fig. 3C, D) compared to the control. Contrary to all the mentioned fluorescence parameters, salinity stress significantly enhanced the non-photochemical quenching (NPQ) by an average increase of 67% under salt stress (Figure 3E).
3.3. Soluble Sugar and Proline Accumulation
A significant increase in proline and soluble sugar contents was recorded in the different parts of the olive tree (Figure 4). It was noted that olive leaves accumulate the highest level of soluble sugars compared with the other tissues under salinity stress and compared to the control (Figure 4A). This level was significantly accumulated in old leaves by 87.37 μmol.g-1 at 200 mM NaCl rather than at 100 mM NaCl with a 59.65 μmol.g-1 level. The lowest accumulation of soluble sugar was recorded in stems by values that did not exceed 10.28 μmol.g-1 at 200 mM compared to the control (5.64 μmol.g-1). However, this increase in the accumulation levels of soluble sugar was decreased through the potassium and calcium applications in all plant parts compared to plants grown under salt stress. The highest marked enhancement of soluble sugars was recorded in young and old leaves, an increase of 7.8% at the lowest dose of potassium (10 K) and calcium (10 Ca) under 100 mM NaCl (Figure 4A).
The proline content was markedly influenced, especially in old leaves, with an increase of 4.4 times under salt stress compared to the control. While the lowest level was recorded in stems with an accumulation of 0.67 μmol.g-1, under salt stress compared to the control (0.32 μmol.g-1) (Figure 4 B). Besides that, the applied potassium and calcium decreased the proline accumulation in all plant organs under salt stress. This decrease was markedly noted in young leaves and thin stems by an average reduction of 30.3% at 40 mM KNO3 under salinity doses. In the medium roots, the decrease was markedly depicted at 40 mM CaCl2 in plants grown at 100 mM NaCl, while at 10 mM CaCl2, the proline level increased in all plant parts. At 200 mM NaCl, the proline content was significantly reduced in thin and medium roots by an average reduction of 38.6% at 10 mM CaCl2 (Figure 4 B).
3.4. Photosynthetic Pigments
The obtained results from this experiment showed that 100 and 200 mM NaCl salinity reduced photosynthesis pigments, including chlorophyll a (chla), chlorophyll b (chlb), total chlorophyll (chla+b), and total carotenoid (car) in comparison to the control (Table 1). Potassium and calcium-treated plants showed higher photosynthetic pigment contents compared to plants grown under saline stress. The highest levels were obtained at 40 mM KNO3 and 40 mM CaCl2 at 100 mM NaCl. Besides that, salinity stress decreased the neoxanthin (N) and violaxanthin (V) pigments, while an increase of anteraxanthin (A), lutein(L), carotene (C), and zeaxanthin (Z) was observed only under 200 mM NaCl (Table 1). Potassium and calcium supplies improved these pigments, which are related to treatments. The total pool of xanthophyll cycle pigments (VAZ, sum of violaxanthin + antheraxanthin and zeaxanthin) increased under a high salinity dose. On the other hand, potassium and calcium induced a marked decrease under both salt treatments, but to different extents among treatments. In addition, the salinity increased the lutein and chlorophyll ratio and carotene and chlorophyll ratio by 27.75 and 32.86%, respectively, at salinity doses. Salinity increased the (V+A+Z)/Chl ratio by 15.45 and 28.46% at 100 mM and 200 mM NaCl, respectively, but it was decreased by potassium and calcium applications (Table 1).
3.5. Physiological and Biochemical Interactions
Overall, this study highlights the importance of the positive interaction between the effective quantum yield of PSII (ΦPSII), stomatal conductance (Gs), and photosynthetic rate (A), to adapt plants for maintaining their physiological and biochemical processes (Figure 5). This positive correlation between photosynthetic rate and Gs can indicate how effectively plants can regulate their stomatal openings to optimize carbon dioxide uptake and water conservation under salinity stress (Figure 5 a, b). This study has shown that higher Gs correlate with increased ΦPSII, as stomatal opening ensures a continuous supply of CO2 for photosynthesis. A clear negative correlation was found between photosynthetic rate and osmoprotectant molecules such as proline and sugar (Figure 5 c, d, e, f). Understanding this relationship led researchers to develop strategies to improve plant tolerance to salinity stress and enhance crop productivity.
4. Discussion
Salinity stress adversely affects plant productivity by reducing growth and yield, primarily due to decreased carbon fixation from stomatal closure, restricted photosynthesis, influencing osmotic adaptation, and nutritional imbalances. A similar observed adverse effect of salinity stress on the physiological mechanisms of plants has been observed with those reported by several previous research works on olive trees [4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22]. According to our previous results [10], the application of potassium (as KNO3) and Calcium (as CaCl2) is considered an effective supplement to alleviate the negative effects of salinity in olive plants, improving growth parameters and ensuring membrane stability. Additionally, the present study aimed to investigate the role of physiological and biochemical interactions in mitigating salt stress in olive plants. Thus, our results are similar to Chartzoulakis et al. [23], which revealed that the potassium at 100 mM increased photosynthesis in olive leaves. Cha-um et al. [24] showed that the low level of potassium maintained the stabilization of photosynthetic pigments in rice, with better growth under 200 mM NaCl. In addition, potassium is known for its role in osmotic adjustment of the guard cell, controlling the stomatal movements resulting in CO2 assimilation in photosynthesis [23]. Jafari et al. [25] showed that a high level of Ca prompted photosynthesis at 80 mM NaCl dramatically from 17.5 to 27.4 μmol CO2.m-2.s-1 in sorghum. It is well known that potassium is an important supplement for plant growth and is crucial to maintain a balance of nutrients to prevent adverse effects on plants under salinity stress [26]. Similar to the photosynthetic rate, the stomatal conductance was reduced under salinity stress compared to the control [7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27]. The combination of salinity with the addition of both calcium and potassium supplements to the plants increased stomatal conductance. These results are in agreement with the initial hypothesis that extra calcium may ameliorate the effect of salinity on stomatal conductance, which has been revealed in rice [28], guava [29], cucumber [30], and endive [31]. It was noted that the reduction of transpiration by salinity was related to the reduced stomatal conductance under salinity stress. However, supplementary foliar nutrient application increased the transpiration rate. The highest alleviation effect was induced at 40 mM KNO3 and 40 mM CaCl2, under 100 mM NaCl. It was concluded in previous results that olive trees under salinity stress are subject to important physiological changes, such as a marked decrease in the rate of photosynthetic assimilation (A), stomatal conductance (gs), and transpiration (E) [32,33,34]. Therefore, water use efficiency (WUE) was significantly impacted by salinity stress but showed moderate improvement with the application of 40 mM KNO3 and 10 mM CaCl2. Trabelsi et al. [35], who reported a reduction of WUE in both young and old olive leaves of plants grown under salinity stress, highlighted this effect. Some studies have shown that K plays an important role in growth and water use efficiency in olives [36], and could be useful in ameliorating the biotic stress effect in olive plants [37].
Some studies have shown that salt stress inhibits PSII photochemistry [38]. The decline of ФPSII by salinity might cause excess light energy, which would increase the excitation pressure on PSII, raising the probability of reactive oxygen species (ROS) generation and photo-inhibition of PSII [39]. The increase in non-photochemical quenching (NPQ) may indicate a reduced demand for the products of electron transport, which are typically utilized for assimilation, leading to the dissipation of excess light energy [40]. Potassium and calcium application enhanced the operating yield of PSII and chlorophyll, with the maximum impact observed at 40 mM KNO3 and 40 mM CaCl2 in plants exposed to salt stress. Chlorophyll concentration can be used as a sensitive indicator of the cellular metabolic state; therefore, its decrease indicates toxicity in tissues due to the accumulation of ions. This result agreed with Kaya et al. [41], who found that the chlorophyll concentration was reduced in strawberry plants under 35 mM NaCl compared to the control, while the potassium at 3 mM (K2SO4) increased the chlorophyll concentration. Besides that, the photosynthetic pigment was influenced by salinity stress and was alleviated by potassium and calcium, which is in agreement with the obtained results of Melgar et al. [22].
The present study showed that the increase in proline content was related to NaCl concentrations. Proline could protect the photosynthetic activity of salt-stressed olive trees by regulating hydration and osmotic adjustment, thus promoting growth even under stressful conditions [42,43,44]. Ben Ahmed et al. [45] noted similar results in olive cv. Chemlali. Poury et al. [46] outlined that proline amid the tolerance mechanisms developed by plants against environmental stress. Overall, the supplied KNO3 and CaCl2 decreased proline content, which was more effective at 40 mM at 100 mM NaCl. The calcium application induced the ameliorated adverse effect of salinity on proline content [47]. According to Wang et al. [26], the application of potassium significantly reduced the uptake of Na+ and enhanced K+ levels in plants under salinity conditions, leading to an osmotic regulation with proline accumulation. Potassium maintains ionic and osmotic balance in plant cells and reduces the need for proline accumulation as an osmoprotectant. In addition, potassium can also enhance the activity of enzymes involved in proline metabolism, promoting the efficient utilization of proline in plants under salt-stress conditions. The proline accumulation in plants was reviewed by Jin et al. [48] and Zhu [49] through the involvement of proline key enzyme P5CS in this upregulation when plants were supplemented with potassium under saline stress. Potassium can influence stress-signalling pathways, including those involving calcium, which is integral for activating stress-responsive genes.
Similarly to proline content, salinity increased soluble sugar content in all plant parts. This accumulation may represent a key vital mechanism for olive trees under salt constraint by allowing them to prevent tissue dehydration and protect their cells from oxidative damage [44]. However, potassium supply has been shown to mitigate the negative effects of salt stress on soluble sugar accumulation in plants. Furthermore, the potassium and calcium application under salt stress decreased the soluble sugars in olive tissues. Farooq et al. [50] demonstrated that potassium supplementation under salt stress significantly increased soluble sugar content in olive leaves. Zhao et al. [51] highlighted that calcium supplementation increased sugar accumulation by stabilizing cell membranes and promoting the activity of transport proteins involved in sugar movement within the plant. Meanwhile, calcium stabilizes cell structures and acts as a signaling molecule that upregulates the gene responsible for sugar metabolism.
The recorded positive correlation indicates that higher stomatal conductance (Gs) is associated with increased ΦPSII, which may reduce photorespiration and minimize ROS formation, protecting PSII and sustaining ΦPSII [52]. While Stomatal conductance is proportional to CO2 concentration and air humidity [53], Salinity stress may lead to decreased stomatal conductance as well as chlorophyll content, which affects olive trees' ability to perform photosynthesis efficiently [54]. However, potassium and calcium supplementation enhance the interaction between stomatal conductance (Gs) and ΦPSII [55]. The detrimental effects of salt on photosynthesis, stomatal conductance, and transpiration rate may be considerably mitigated by adding K and Ca to irrigation solutions [56]. This suggests that the effect of potassium and calcium alleviates salt stress in olive plants and can maintain a higher photosynthetic rate under salinity stress, possibly due to their ability to regulate stomatal conductance more effectively. However, the clear negative correlation between photosynthetic rate and osmoprotectant molecules could be explained by the stomatal closure, reduction of CO2 uptake, or damage to the photosystem II (PSII) [57]. Stress conditions can limit photosynthetic efficiency, causing a rise in sugar levels could be part of a plant's stress adaptation strategy, where accumulated sugars help mitigate damage by serving as osmolytes, stabilizing cellular structures, or acting as signals that trigger protective pathways [58]. Overall, the potassium and calcium significantly enhanced the osmoprotectant molecule levels in olive tissues, which are related to salinity tolerance. Moreover, they protect physiological processes against harmful inorganic compounds [59]. In addition, soluble sugar and proline are considered efficient means of preserving the potential of a passive osmotic process in the cytoplasm and in maintaining ribosomes and proteins against the harmful impacts of Na+ ions.
5. Conclusions
Salt stress negatively influences olive trees by affecting physiological and biochemical efficiency. The high salinity levels influence the osmotic stress, resulting in reduced stomatal conductance and impaired gas exchange, ultimately affecting the tree’s ability to photosynthesize. Thus, highlighting the effective strategies to enhance salt tolerance in olive trees is critical for maintaining their productivity in increasingly saline conditions. The obtained results confirmed mainly the impact of potassium and calcium supplies in reducing the detrimental effects of salinity in olive trees.
Author Contributions
Conceptualization, A.L. and H.B.; methodology, A.L., H.B; software, A.K., F.M; validation, A.L. and H.B. and A.LA.; formal analysis, A.LA., F.M., A.K.; investigation, A.LA., A.K., H.B.; data curation, H.B and A.L.; writing—original draft preparation, A.LA., A.K., H.B.; writing—review and editing, A.LA., H.B., A.L, A.K; supervision, A.L.; project administration, H.B.; funding acquisition, A.K. All authors have read and agreed to the published version of the manuscript
Funding
This research was funded by a grant from Tunisian LPOI (LR16 IO 03-SERS-Tunisia) and African Plant Nutrition Institute (APNI) ‘Knowledge on 4R nutrient management to support olive production and establishment of sufficient nutrient ranges for intensive olive plantations in Tunisia
Data Availability Statement
Data contained in the present work are original.
Acknowledgments
We appreciate all the people who have collaborated on this project.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Pacifici, M.; Foden, W.B.; Visconti, P.; Watson, J.E.M.; Butchart, S.H.M.; Kovacs, K.M.; Scheffers, B.R.; Hole, D.G.; Martin, T.G.; Akçakaya, H.R.; Corlett, R.T.; Huntley, B.; Bickford, D.; Carr, J.A.; Hoffmann, A.A.; Midgley, G.F.; Kelly, P.P.; Pearson, R.G.; Williams, S.E.; Willis, S.G.; Young, B.; Rondinini, C. Assessing species vulnerability to climate change. Nat. Clim. Chang. 2015, 5, 215–225. [Google Scholar] [CrossRef]
- Okur, B.; Orçen, N. Soil salinization and climate change. In Climate change and soil interactions; Prasad, M.N.V., Pietrzykowski, M., Eds.; Elsevier: Amsterdam, 2020; pp. 331–350. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Azimi, M.; Khoshzaman, T.; Taheri, M.; Dadras, A.R. Evaluation of salinity tolerance of three olive (Olea europaea L.) cultivars. J. Cent. Eur. Agric. 2021, 22, 571–581. [Google Scholar] [CrossRef]
- Vigo, C.; Therios, I.N.; Bosabalidis, M. Plant growth, nutrient concentration and leaf anatomy of olive plants irrigated with diluted seawater. J. Plant Nutr. 2005, 28, 1001–1021. [Google Scholar] [CrossRef]
- Kapulnik, Y.; Tsror, L.; Zipori, I.; Hazanovsky, M.; Wininger, S.; Dag, A. Effect of AMF application on growth, productivity and susceptibility to Verticillium Wilt of olives grown under desert conditions. Symbiosis 2010, 52, 103–111. [Google Scholar] [CrossRef]
- Boussadia, O.; Zgallai, H.; Mzid, N.; Zaabar, R.; Braham, M.; Doupis, G.; Koubouris, G. Physiological responses of two olive cultivars to salt stress. Plants 2023, 12, 1926. [Google Scholar] [CrossRef] [PubMed]
- Vergine, M.; Palm, E.R.; Salzano, A.M.; Negro, C.; Nissim, W.G.; Sabbatini, L.; Balestrini, R.; Pinto, M.C.; Dipierro, N.; Gohari, G.; Fotopoulos, V.; Mancuso, S.; Luvisi, A.; Bellis, L.; Scaloni, A.; Vita, F. Water and nutrient availability modulate the salinity stress response in Olea europaea cv. Arbequina. Plant Stress 2024, 14, 100648. [Google Scholar] [CrossRef]
- Hassan, I.F.; Maybelle, G.; Bedour, A.L.; Proietti, P.; Regni, L. Salinity stress effects on three different olive cultivars and the possibility of their cultivation in reclaimed lands. Plant Arch. 2020, 20, 2378–2382. [Google Scholar]
- Larbi, A.; Kchaou, H.; Gaaliche, B.; Gargouri, K.; Boulal, H.; Morales, F. Supplementary potassium and calcium improves salt tolerance in olive plants. Sci. Hortic. 2020, 260, 108912. [Google Scholar] [CrossRef]
- Taha, R.S.; Seleiman, M.F.; Alotaibi, M.; Alhammad, B.A.; Rady, M.M.; Mahdi, A.H.A. Exogenous potassium treatments elevate salt tolerance and performances of Glycine max L. by boosting antioxidant defense system under actual saline field conditions. Agronomy 2020, 10, 1741. [Google Scholar] [CrossRef]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Gucci, R.; Lombardini, L.; Tattini, M. Analysis of leaf water relations in leaves of two olive (Olea europaea) cultivars differing in tolerance to salinity. Tree Physiol. 1997, 17, 13–21. [Google Scholar] [CrossRef]
- Rugini, E.; Baldoni, L.; Muleo, R.; Sebastiani, L. The Olive Tree Genome; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Loupassaki, M.H.; Chartzoulakis, K.S.; Digalaki, N.B.; Androulakis, I.I. Effects of salt stress on concentration of nitrogen, phosphorus, potassium, calcium, magnesium, and sodium in leaves, shoots, and roots of six olive cultivars. J. Plant Nutr. 2002, 25, 2457–2482. [Google Scholar] [CrossRef]
- Tattini, M.; Melgar, J.C.; Traversi, M.L. Responses of Olea europaea to high salinity: a brief-ecophysiological review. Adv. Hort. Sci. 2008, 22, 159–173. [Google Scholar]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef]
- Kchaou, H.; Larbi, A.; Chaieb, M.; Sagardoy, R.; Msallem, M.; Morales, F. genotypic differentiation in the stomatal response to salinity and contrasting photosynthetic and photoprotection responses in five olive (Olea europaea L.) Cultivars. Sci. Hortic. 2013, 160, 129–138. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Robyt, J.F.; White, B.J. Biochemical Techniques: Theory and Practice; Brooks/Cole: Monterey, CA, USA, 1987; p. 217. [Google Scholar]
- Melgar, J.C.; Mohamed, Y.; Ben Yahmed, J.; Abadia, A.; Abadia, J.; Gomez, J.A. Long-term responses of olive trees to salinity. Agric. Water Manag. 2009, 96, 1105–1113. [Google Scholar] [CrossRef]
- Chartzoulakis, K.; Loupassaki, M.; Bertaki, M.; Androulakis, I. Response of two olive cultivars to salt stress and potassium supplement. J. Plant Nutr. 2006, 29, 2063–2078. [Google Scholar] [CrossRef]
- Cha-um, S.; Yooyongwech, S.; Supaibulwatana, K.; Kirdmanee, C. Water relations, pigment stabilization, photosynthetic abilities and growth improvement in salt stressed rice plants treated with exogenous potassium nitrate application. Int. J. Plant Prod. 2010, 4, 187–198. [Google Scholar]
- Jafari, M.H.S.; Abbasi, A.R.; Mousavi, S.F. Interactive effects of NaCl induced salinity, calcium, and potassium on physio-morphological traits of Sorghum (Sorghum bicolor L.). Pak. J. Bot. 2009, 41, 3053–3063. [Google Scholar]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef]
- Tattini, M.; Traversi, M.L.; Pinelli, P.; Leonardi, C.; Noferini, M.; Faccio, A.; Fini, A. Contrasting response mechanisms to root-zone salinity in three co-occurring Mediterranean woody evergreens: A physiological and biochemical study. Funct. Plant Biol. 2009, 36, 551–563. [Google Scholar] [CrossRef]
- Sultana, N.; Ikeda, T.; Kashem, M.A. Effect of foliar spray of nutrient solutions on photosynthesis, dry matter accumulation and yield in seawater-stressed rice. Environ. Exp. Bot. 2001, 46, 129–140. [Google Scholar] [CrossRef]
- Ebert, G.; Eichert, T.; Fernández, V.; Müller, B.; Römheld, V. Ameliorating effects of Ca(NO₃)₂ on growth, mineral uptake and photosynthesis of NaCl-stressed guava seedlings (Psidium guajava L.). Sci. Hortic. 2002, 93, 125–135. [Google Scholar] [CrossRef]
- Kaya, C.; Higgs, D. Ameliorative effects of potassium phosphate on salt stressed pepper and cucumber. J. Plant Nutr. 2002, 26, 807–820. [Google Scholar] [CrossRef]
- Tzortzakis, N.G. Potassium and calcium enrichment alleviate salinity-induced stress in hydroponically grown endives. Hort. Sci. 2010, 37, 155–162. [Google Scholar] [CrossRef]
- Roussos, P.A.; Koubouris, G.C.; Tsantili, E. Intra- and inter-cultivar impacts of salinity stress on leaf photosynthetic performance, carbohydrates and nutrient content of nine indigenous Greek olive cultivars. Acta Physiol. Plant. 2017, 39, 136. [Google Scholar] [CrossRef]
- Buono, D.; Muzzalupo, I.; Perri, E.; Rinaldi, R.; Rinaldi, A.; Caira, S.; Ruggiero, A.; Balestrieri, M.L. Effects of Megafol on the olive cultivar ‘Arbequina’ grown under severe saline stress in terms of physiological traits, oxidative stress, antioxidant defenses, and cytosolic Ca²⁺. Front. Plant Sci. 2021, 11, 603576. [Google Scholar]
- Skodra, C.; Pateraki, I.; Madesis, P.; Kalaitzis, P. Unraveling salt-responsive tissue-specific metabolic pathways in olive tree. Physiol. Plant. 2021, 173, 13565. [Google Scholar] [CrossRef]
- Trabelsi, L.; Smaoui, A.; Ksouri, R.; Triki, M.A.; Oueslati, S. The effect of drought and saline water on the nutritional behaviour of the olive tree (Olea europaea L.) in an arid climate. S. Afr. J. Bot. 2024, 165, 126–135. [Google Scholar] [CrossRef]
- Restrepo-Diaz, H.; Benlloch, M.; Navarro, C.; Fernandez, J.E. Potassium fertilization of rainfed olive orchards. Sci. Hortic. 2008, 116, 399–403. [Google Scholar] [CrossRef]
- Abdolzadeh, A.; Karimi, E.; Sadeghipour, H.R. Increasing salt tolerance in olive, Olea europaea L. plants by supplemental potassium nutrition involves changes in ion accumulation and anatomical attributes. Int. J. Plant Prod. 2009, 3, 49–60. [Google Scholar]
- El Khouni, A.; El Aouni, M.H.; Ghnaya, T.; Abdelly, C.; Gouia, H. Structural and functional integrity of Sulla carnosa photosynthetic apparatus under iron deficiency conditions. Plant Biol. 2018, 20, 415–425. [Google Scholar] [CrossRef]
- Netondo, G.W.; Onyango, J.C.; Beck, E. Sorghum and salinity II. Gas exchange and chlorophyll fluorescence of sorghum under salt stress. Crop Sci. 2004, 44, 806–811. [Google Scholar]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Kaya, C.; Kirnak, H.; Higgs, D.; Saltali, K. Supplementary calcium enhances plant growth and fruit yield in strawberry cultivars grown at high (NaCl) salinity. Sci. Hortic. 2002, 93, 65–74. [Google Scholar] [CrossRef]
- Boualem, S.; Boutaleb, S.; Boukhalfa-Deraoui, N. Effect of salinity on the physiological behavior of the olive tree (variety Sigoise). J. Fundam. Appl. Sci. 2019, 11, 525–538. [Google Scholar]
- Ayaz, M.; Hakki, E.E.; Ercisli, S.; Cakir, O.; Tanyolac, B. Three Turkish olive cultivars display contrasting salt stress-coping mechanisms under high salinity. Trees 2021, 35, 1283–1298. [Google Scholar] [CrossRef]
- El Yamani, M.; Cordovilla, M.P. Tolerance mechanisms of olive tree (Olea europaea) under saline conditions. Plants 2024, 13, 2094. [Google Scholar] [CrossRef] [PubMed]
- Ben Ahmed, C.; Ben Rouina, B.; Sensoy, S.; Boukhris, M.; Abdallah, F.B. Changes in water relations, photosynthetic activity and proline accumulation in one-year-old olive trees (Olea europaea L. cv. Chemlali) in response to NaCl salinity. Acta Physiol. Plant. 2008, 30, 553–560. [Google Scholar] [CrossRef]
- Poury, N.; Kholdebarin, B.; Shariati, M. Effects of salinity and proline on growth and physiological characteristics of three olive cultivars. Gesunde Pflanzen 2023, 75, 1169–1111. [Google Scholar] [CrossRef]
- Mozafari, M.R.; Johnson, C.; Hatziantoniou, S.; Demetzos, C. Nanoliposomes and their applications in food nanotechnology. J. Liposome Res. 2008, 18, 309–327. [Google Scholar] [CrossRef]
- Jin, Z.; Wang, Z.; Ma, Q.; Sun, L.; Liu, Z.; Zhai, S.; Xu, J.; Cui, X.; Jiang, D.; Cao, W. Potassium supply improves salt tolerance in wheat through better K/Na ratio and enhanced antioxidative metabolism. Plant Sci. 2015, 230, 103–111. [Google Scholar]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Nawaz, A.; Chaudhary, H.J.; Rehman, A.; Nadeem, F.; Ali, Q.; Zahoor, R. Morphological, physiological and biochemical aspects of zinc seed priming-induced drought tolerance in faba bean. Sci. Hortic. 2021, 281, 109894. [Google Scholar] [CrossRef]
- Zhao, Y.; Ma, Q.; Jin, Z.; Wang, Z.; Wang, J.; Jia, H.; Chen, W.; Deng, Q.; Ding, Y. Physiological, proteomic and metabolomic analysis provide insights into Bacillus sp. mediated salt tolerance in wheat. Plant Cell Rep. 2022, 41, 95–118. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Keeping a positive carbon balance under adverse conditions: Responses of photosynthesis and respiration to water stress. Physiol. Plant. 2004, 122, 545–552. [Google Scholar] [CrossRef]
- Villalobos, F.J.; López-Bernal, Á.; García-Tejera, O.; Testi, L. Is olive crop modelling ready to assess the impacts of global change? Front. Plant Sci. 2023, 14, 1249793. [Google Scholar] [CrossRef]
- Karaca, C.; Aslan, G.E.; Kurunc, A.; Baştug, R.; Navarro, A.; Buyuktas, D. Effect of irrigation water salinity on physiological parameters and yield of tomato plants across phenological stages. Earth Sci. Hum. Constr. 2024, 4, 177–186. [Google Scholar] [CrossRef]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Ma, Y.; Yuan, Z.; Wei, Z.; Yan, F.; Liu, X.; Li, X.; Hou, J.; Hao, Z.; Liu, F. Stomatal and non-stomatal regulations of photosynthesis in response to salinity, and K and Ca fertigation in cotton (Gossypium hirsutum L. cv.). Environ. Exp. Bot. 2025, 230, 106092. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in c3 plants: stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Guerfel, M.; Baccouri, O.; Boujnah, D.; Chaïbi, W.; Zarrouk, M. impacts of water stress on gas exchange, water relations, chlorophyll content and leaf structure in the two main tunisian olive (Olea europaea L.) cultivars. Sci. Hortic. 2009, 119, 257–263. [Google Scholar] [CrossRef]
- Rady, M.M.; Taha, R.S.; Semida, W.M.; Alharby, H.F. Modulation of salt stress effects on Vicia faba L. plants grown on a reclaimed-saline soil by salicylic acid application. Rom. Agric. Res. 2017, 34, 175–185. [Google Scholar]
Figure 1.
Experimental design.
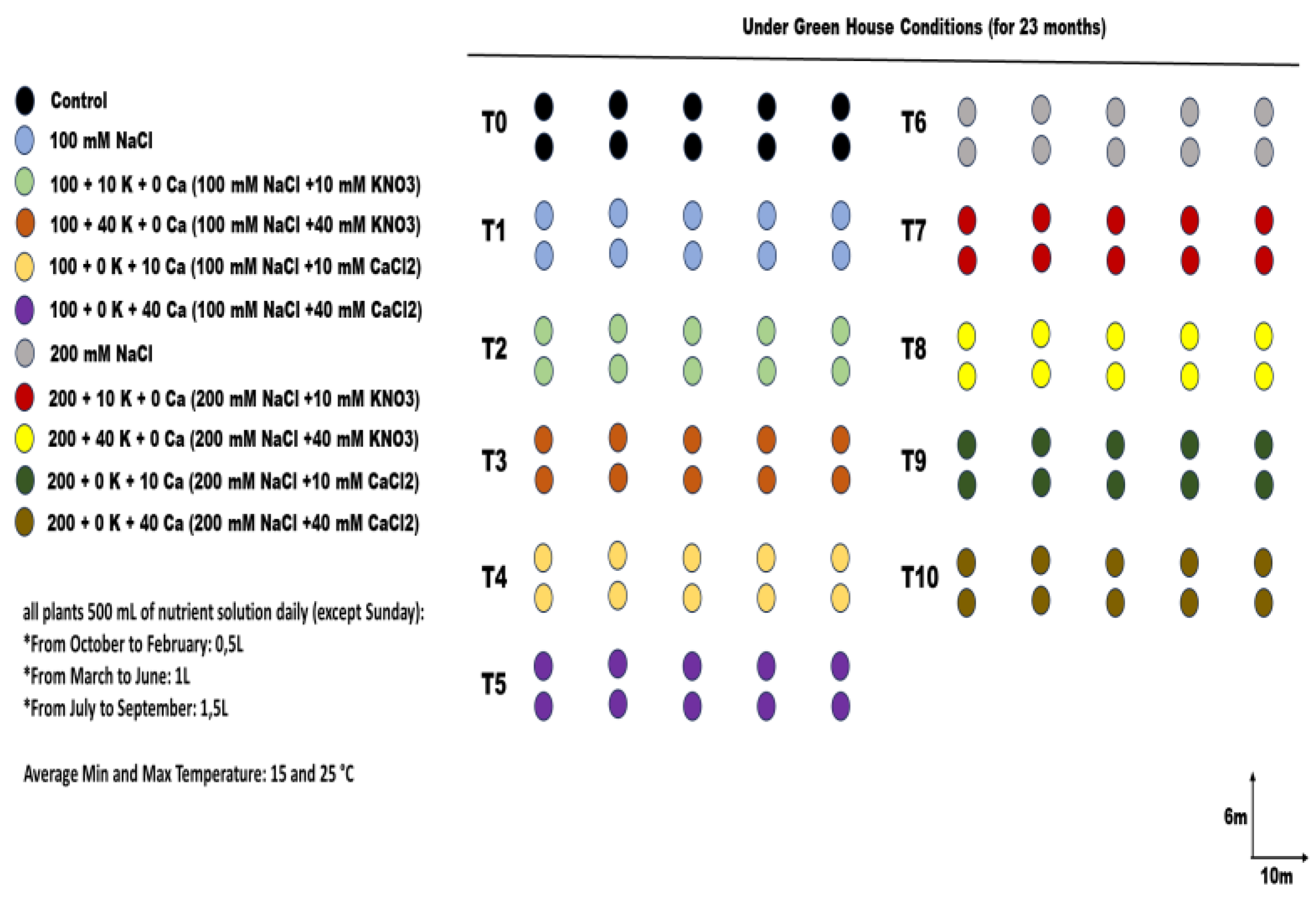
Figure 2.
Stomatal conductance (gs)(a), photosynthetic rate (A) (b), transpiration (E) (c), water use efficiency (A/E) (d), in ArbequinaI18 olive plants under NaCl (100 and 200 mM), supplemented with potassium (10 and 40 mM KNO3) and calcium (10 and 40 mM CaCl2). The bars on each column show standard error. Values (N = 5 ± S.E.). Different letters in columns indicate significant differences among treatments at P < 0.05.
Figure 2.
Stomatal conductance (gs)(a), photosynthetic rate (A) (b), transpiration (E) (c), water use efficiency (A/E) (d), in ArbequinaI18 olive plants under NaCl (100 and 200 mM), supplemented with potassium (10 and 40 mM KNO3) and calcium (10 and 40 mM CaCl2). The bars on each column show standard error. Values (N = 5 ± S.E.). Different letters in columns indicate significant differences among treatments at P < 0.05.
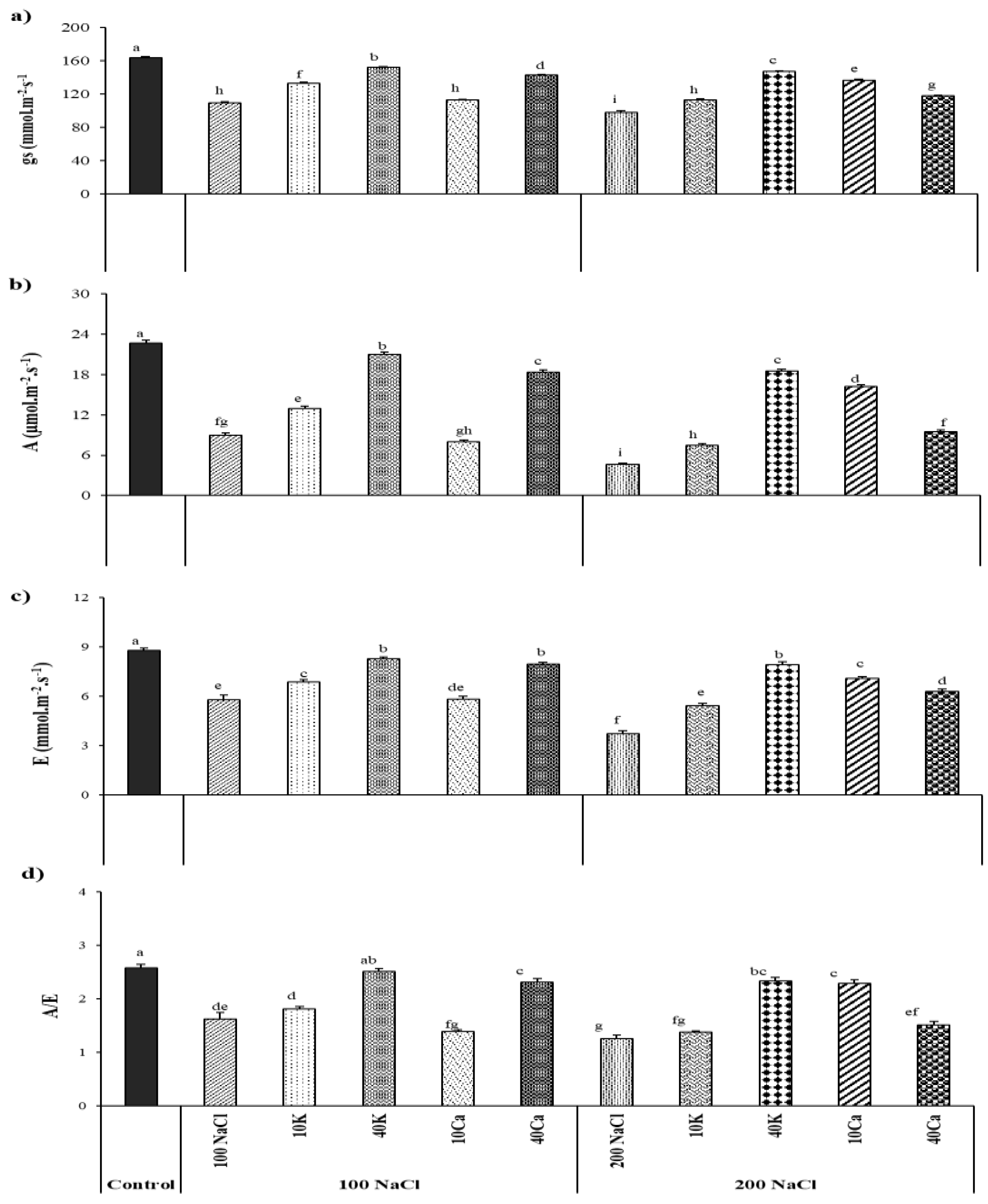
Figure 3.
Fv/Fm, ΦPSII, Φexc, QP. NPQ of ArbequinaI18 cultivar under NaCl (100 and 200 mM) supplemented with potassium (10 and 40 mM KNO3) and calcium (10 and 40 mM CaCl2). The bars on each column show standard error. Values (N = 5 ± S.E.). Different letters in columns indicate significant differences among treatments at P < 0.05.
Figure 3.
Fv/Fm, ΦPSII, Φexc, QP. NPQ of ArbequinaI18 cultivar under NaCl (100 and 200 mM) supplemented with potassium (10 and 40 mM KNO3) and calcium (10 and 40 mM CaCl2). The bars on each column show standard error. Values (N = 5 ± S.E.). Different letters in columns indicate significant differences among treatments at P < 0.05.
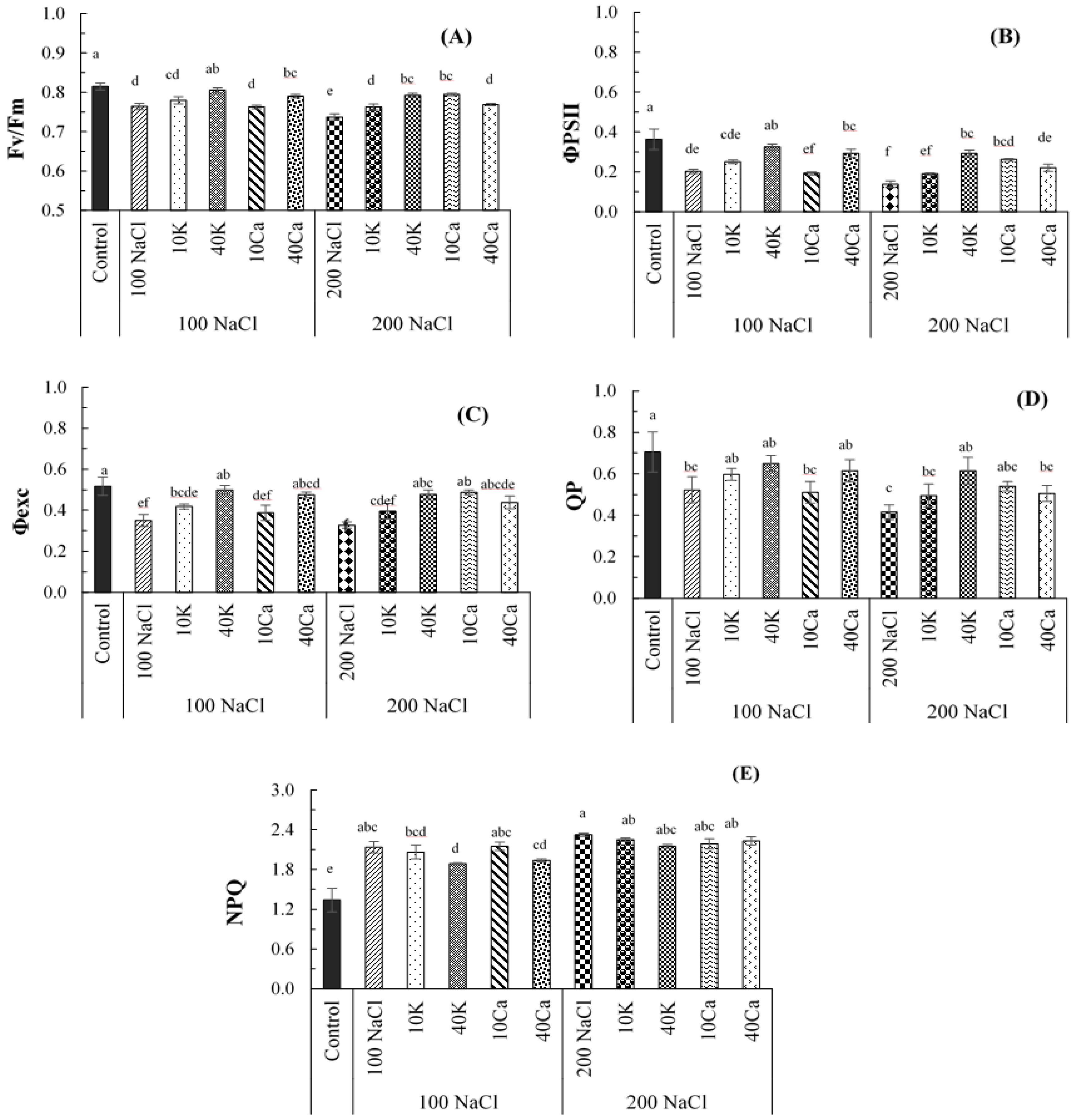
Figure 4.
Proline contents of Arbequina I18 cultivars under NaCl (100 and 200 mM), supplemented with potassium (10 and 40 mM KNO3) and calcium (10 and 40 mM CaCl2). The bars on each column show standard error. Values (N = 5 ± S.E.). Different letters in columns indicate significant differences among treatments at P < 0.05.
Figure 4.
Proline contents of Arbequina I18 cultivars under NaCl (100 and 200 mM), supplemented with potassium (10 and 40 mM KNO3) and calcium (10 and 40 mM CaCl2). The bars on each column show standard error. Values (N = 5 ± S.E.). Different letters in columns indicate significant differences among treatments at P < 0.05.
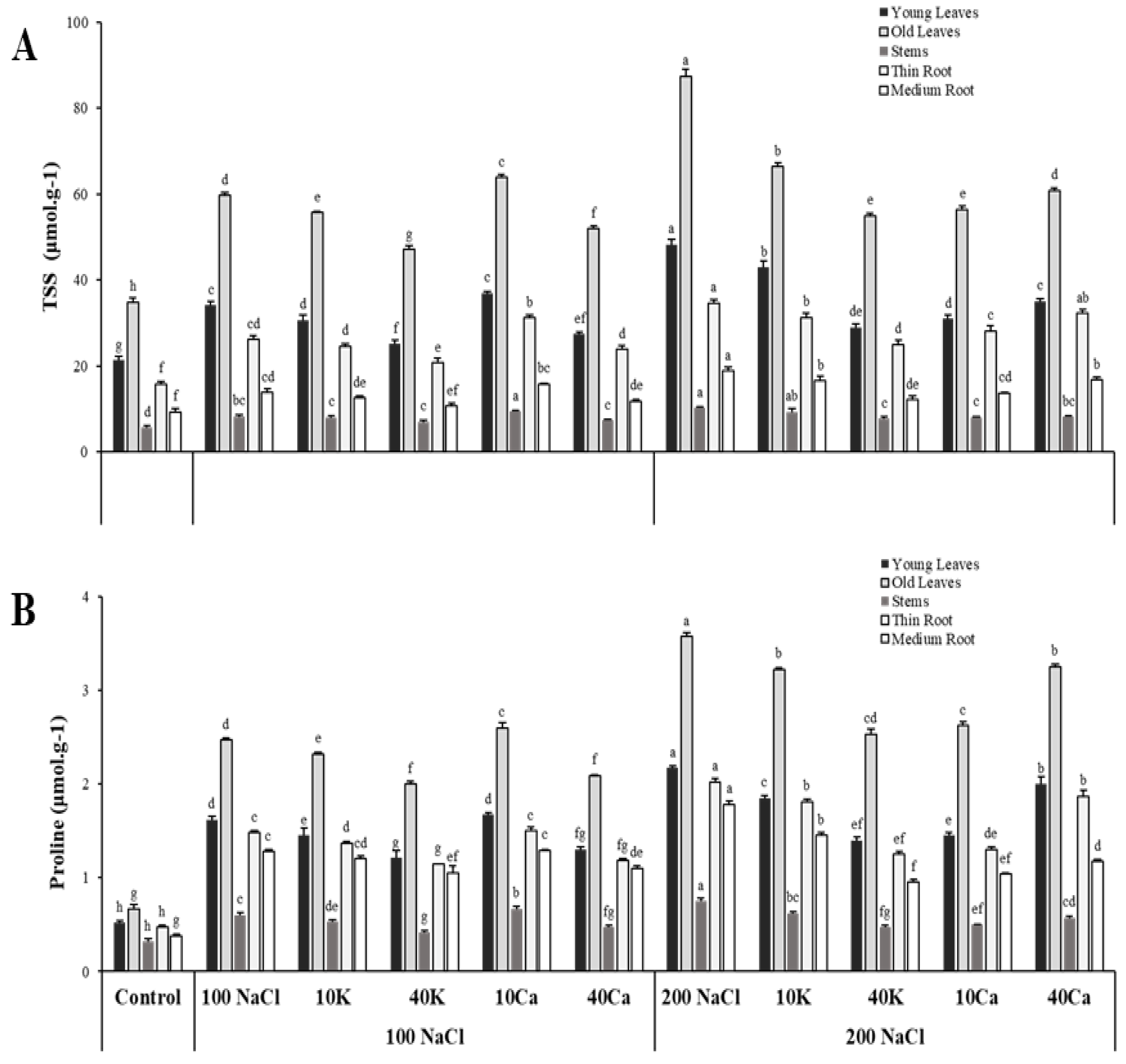
Figure 5.
Interactions with photosynthetic rate (A), stomatal conductance (gs), and osmoprotectant (SS: soluble sugar; Pr: proline); OL: Old leaves; YL: Young leaves; (a): A/gs; (b): A/ ФPSII; (c): A/SS YL; (d): A/SS OL; (e): A/Pr YL; (f): A/Pr OL.
Figure 5.
Interactions with photosynthetic rate (A), stomatal conductance (gs), and osmoprotectant (SS: soluble sugar; Pr: proline); OL: Old leaves; YL: Young leaves; (a): A/gs; (b): A/ ФPSII; (c): A/SS YL; (d): A/SS OL; (e): A/Pr YL; (f): A/Pr OL.
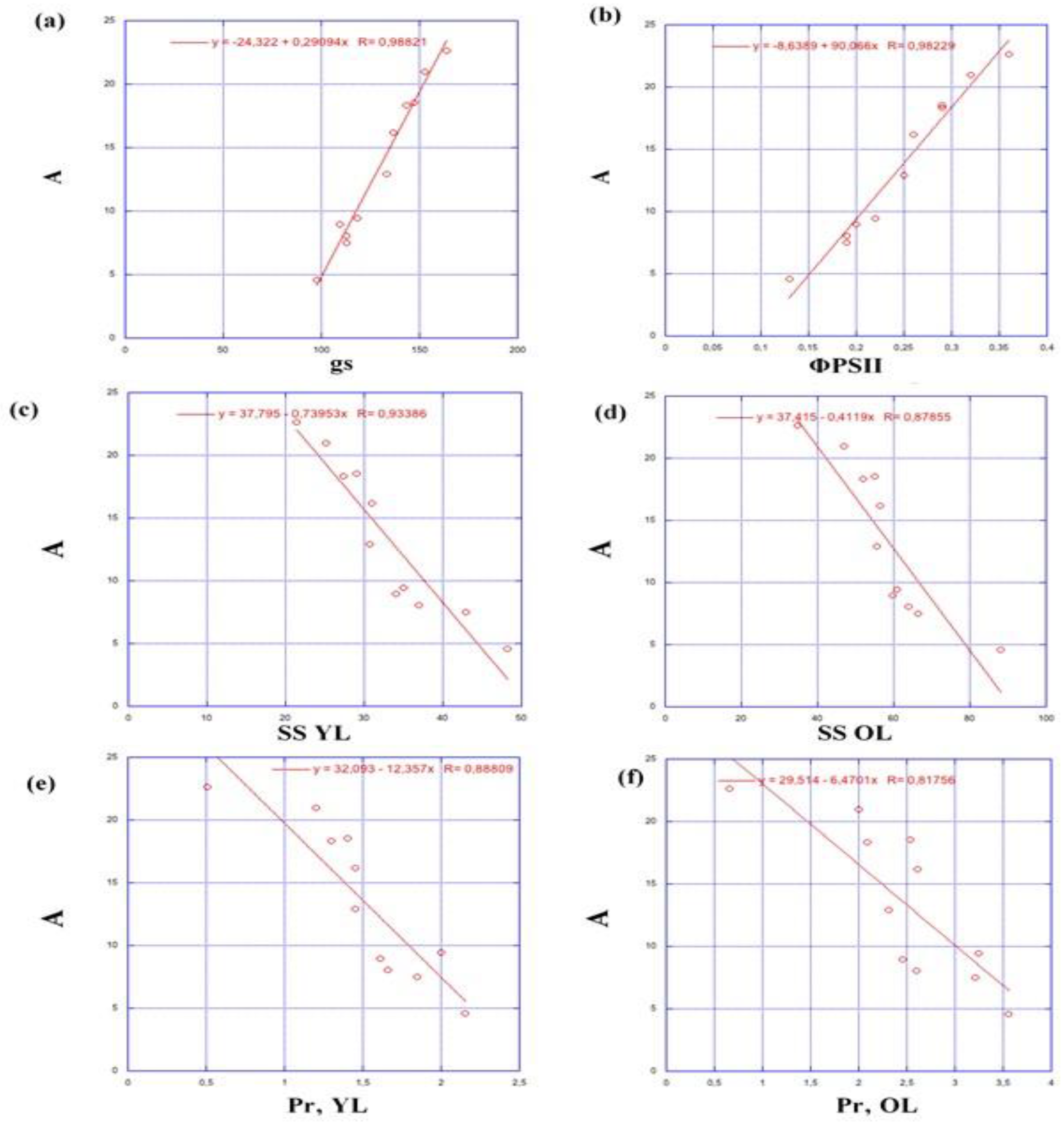

Table 1.
Total Chl (Chl a+b), Chl a/b, β-carotene, lutein, neoxanthin, and violaxanthin + antheraxanthin + zeaxanthin (VAZ) (mmol pigment m-2). Lutein/Chl and V+A+Z/Chl (mol pigment mol-1 Chl).
Table 1.
Total Chl (Chl a+b), Chl a/b, β-carotene, lutein, neoxanthin, and violaxanthin + antheraxanthin + zeaxanthin (VAZ) (mmol pigment m-2). Lutein/Chl and V+A+Z/Chl (mol pigment mol-1 Chl).
| Treatment | Total Chl | Chl a/b | β-Carotene | Lutein | Neoxanthin | VAZ | (A+Z)/(V+A+Z) | Lutein/Chl | V+A+Z/Chl |
|---|---|---|---|---|---|---|---|---|---|
| Control | 655±10bc | 2.54±0.67a | 54±90a | 91±17a | 31±50abc | 63±16a | 0.17±0.04cd | 0.14±0.01abc | 0.09±0.01ab |
| 100 | 556±44c | 3.18±0.39a | 61±19abc | 98±18a | 26±00abcd | 60±12a | 0.28±0.08abc | 0.18±0.01abc | 0.11±0.02ab |
| 100-10K | 588±73c | 2.87±0.35a | 61±29abc | 109±33a | 27±90abcd | 61±21a | 0.26±0.09bcd | 0.18±0.04abc | 0.10±0.02ab |
| 100-40K | 652±74bc | 2.66±0.82a | 57±50ab | 93±13a | 29±00abcd | 60±90a | 0.24±0.08bcd | 0.14±0.04abc | 0.09±0.02ab |
| 100-10Ca | 894±23ab | 3.05±0.14a | 73±38ab | 98±34a | 21±30cd | 47±21a | 0.11±0.09d | 0.12±0.07bc | 0.06±0.03b |
| 100-40Ca | 627±16c | 2.74±0.71a | 49±11ab | 94±50a | 35±13ab | 60±13a | 0.26±0.08bcd | 0.15±0.05abc | 0.10±0.01ab |
| 200 | 552±98c | 3.32±0.62a | 70±11c | 116±23a | 18±90d | 68±15a | 0.42±0.07a | 0.21±0.07a | 0.13±0.04a |
| 200-10K | 620±87c | 3.10±0.19a | 67±90bc | 105±50a | 31±30abc | 79±59a | 0.25±0.13bcd | 0.17±0.03abc | 0.12±0.07a |
| 200-40K | 961±24a | 3.05±0.13a | 47±16abc | 98±70a | 38±80a | 57±10a | 0.23±0.04bcd | 0.11±0.03bc | 0.06±0.02b |
| 200-10Ca | 1031±11a | 3.08±0.02a | 57±11abc | 103±60a | 35±40ab | 60±10a | 0.25±0.05bcd | 0.10±0.01c | 0.06±0.00b |
| 200-40Ca | 587±10c | 3.22±0.07a | 69±17ab | 105±11a | 23±40bcd | 68±17a | 0.36±0.08ab | 0.18±0.04ab | 0.11±0.01ab |
Values (N = 5± S.E.). Means followed by the same letter are not significantly different at P < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



