
Submitted:
09 June 2025
Posted:
13 June 2025
You are already at the latest version
Abstract
Methamphetamine (METH) misuse-induced affective and cognitive dysfunctions cause severe global health and economic burdens. However, the mechanisms underlying METH withdrawal-induced negative emotions and cognitive deficits, as well as the treatment strategies for them, remain elusive. Previous findings suggest that METH use triggers neuroinflammation and neuronal apoptosis, and protein kinase B (AKT), forkhead box protein 1 (FOXO1), and hemeoxygenase-1 (HO-1) are implicated in these processes. In the present study, we aimed to reveal the role and potential mechanisms of luteolin, a flavonoid phytochemical with anti-inflammatory and antioxidative properties, in METH withdrawal-induced negative emotions and cognitive deficits. We found that prolonged METH withdrawal led to an increase in neuronal activity and a decrease in the protein expression of phosphorylated AKT (p-AKT) and HO-1 in the prefrontal cortex (PFC) and caudate putamen (CPu). Luteolin pretreatment partially mitigated these METH withdrawal-induced negative emotions and cognitive deficits, and prevented the abnormal activation of PFC and CPu as well as the downregulation of AKT/HO-1 expression. Notably, we further observed that luteolin inhibited the METH-induced nuclear translocation of FOXO1. Our findings suggest that luteolin may alleviate METH withdrawal-induced affective and cognitive dysfunctions by reducing oxidative injury in the brain through the AKT/FOXO1/HO-1 pathway, highlighting its potential for treating drug addiction-related health issues.
Keywords:
methamphetamine withdrawal
; negative emotions
; cognitive deficits
; luteolin
; protein kinase B (AKT)
; forkhead box protein 1 (FOXO1)
; heme-oxygenase-1 (HO-1)
1. Introduction
Amphetamines are among the most widely misused illicit drugs globally, with over 30 million users according to the World Drug Report 2024. Methamphetamine (METH), a typical amphetamine, is the most misused drug in China [1]. As a potent psychostimulant that can cross the blood brain barrier (BBB) [2], METH exhibits severe neurotoxicity, partly due to oxidative stress injury and neuroinflammation in the brain [3,4,5]. Prolonged METH use can cause addiction and cognitive deficits [6,7]. Moreover, patients often develop psychological disorders during METH withdrawal, particularly anxiety and depression [8,9]. Previous studies have demonstrated that approximately 39% - 64% of METH users exhibite depressive symptoms [10,11]. However, the precise mechanisms underlying METH withdrawal-induced negative emotions and cognitive deficits remain unclear.
Chronic METH use triggers various neural adaptations, including neuronal plasticity and gene expression changes in various brain regions such as the prefrontal cortex (PFC) [12,13] and striatum [14,15]. PFC is associated with cognitive dysfunctions after repeated drug use [16], while the striatum [17] is mainly implicated in the rewarding effects of addictive substances. In the PFC, the inhibitory neurons are activated and excitatory neurons are suppressed to facilitate METH-associated memory retrieval [18]. METH exposure has been shown to increase neuronal activity in the dorsal striatum (also known as the caudate putamen, CPu) [19]. Although mounting evidence has revealed neural circuit and neuronal plasticity changes in multiple brain regions during METH addiction [20,21,22], the activity changes and potential roles of neurons in the CPu and PFC in METH withdrawal-induced negative emotions require further exploration.
Gene expression dynamics are crucial in neural plasticity alterations during substance addiction [23]. Chronic METH use has been shown to decrease the expression of heme-oxygenase-1 (HO-1), a cytoprotective gene protecting cells from METH-induced oxidative stress injury and apoptosis [24,25]. Protein kinase B (AKT) is involved in cell proliferation, differentiation, and survival [26]. The phosphorylation of AKT facilitates the expression of HO-1 [25,27]. Forkhead box protein 1 (FOXO1) is one kind of transcription factor associated with neuroinflammation and apoptosis. Intracerebral hemorrhage has been shown to cause neuroinflammation, neuronal apoptosis, and the activation of FOXO1 [28]. The inhibition of the phosphoinositide 3-kinase (PI3K)/AKT pathway upregulates FOXO1 expression [29]. Thus, we hypothesize that the AKT/FOXO1/HO-1 signaling pathway in the PFC and CPu may play a role in METH withdrawal-induced negative emotions and cognitive deficits.
Recently, food-derived phytochemicals and their derivatives have shown benefit for brain functions [30,31]. Luteolin, a flavonoid, has demonstrated neuroprotective effects in cadmium or ischemic/reperfusion-induced brain injuries [32,33]. It can enhance cell viability and downregulate apoptosis via the PI3K/AKT signaling pathway, thereby ameliorating ischemic stroke [34]. Moreover, luteolin can cross the BBB and protect neurons from apoptosis by inhibiting neuroinflammation and oxidative stress [35,36]. However, the role and molecular mechanisms of luteolin in METH withdrawal-induced affective and cognitive dysfunctions are still unknown.
In this study, we first examined METH withdrawal-induced negative emotions, cognitive deficits, and the AKT/HO-1 signaling expression changes in the PFC and CPu. Neuronal activity alterations in these brain regions after prolonged METH withdrawal were also detected. Then, we investigated the effects of luteolin on METH withdrawal-induced negative emotions and cognitive deficits. The AKT/HO-1 expression and neuronal activity alterations in METH withdrawal after luteolin treatment were further determined. Finally, the dynamics of nuclear translocation of FOXO1 were examined to reveal the potential mechanisms in the protective role of luteolin in METH withdrawal-induced affective and cognitive dysfunctions.
2. Results
2.1. METH Withdrawal Induces Anxiety and Depressive-Like Behaviors, as Well as Cognitive Deficits
We first examined METH withdrawal-induced affective and cognitive function changes (Figure 1A). Our results revealed that the mice did not exhibit anxiety-like behaviors 48 - 72 h after METH withdrawal (on days 9 and 10, Figure 1B and C). However, they showed significant anxiety-like behaviors one-week post-withdrawal. Compared to the Saline (Sal) group, the METH group mice spent less time and showed decreased entries in the open arm of EPM, and spent less time in the central zone of OFT (Figure 1D and E). In NORT, the METH group mice exhibited a decreased recognition index compared to the Sal group in the 24h interval test, but not for the 3h interval (Figure 1F). Additionally, the METH group displayed fewer spontaneous alterations in the Y-maze test, without alterations in the total distance traveled (Figure 1G). Regarding depressive-like behaviors, the METH group mice showed reduced sucrose preference (Figure 1H) and increased immobility time in the FST (Figure 1I), compared to the Sal group. These findings demonstrate that METH withdrawal precipitates negative emotions and cognitive deficits.
2.2. METH Withdrawal Increases the Neuronal Activity in the PFC and CPu
To reveal the METH withdrawal-induced activity changes in neurons in the PFC and CPu, we detected and compared the differences in the c-Fos-positive cell densities in the two brain regions. After the final behavioral test, the mice were immediately sacrificed for tissue collection. The c-Fos-positive cell densities exhibited a significant increase in both the PFC (Figure 2A and B) and CPu (Figure 2C and D) in the METH group compared to the Sal group, demonstrating that prolonged METH withdrawal increased the neuronal activity in these two brain regions.
2.3. METH Withdrawal Downregulates p-AKT and HO-1 Expression in the PFC and CPu
To investigate the potential molecular mechanisms underlying METH withdrawal-induced negative emotions and cognitive deficits, we then measured the AKT and HO-1 protein expression in the PFC and CPu using Western blot analysis. Our results revealed that the p-AKT and HO-1 protein level was significantly decreased in the PFC (Figure 3A and B) and CPu (Figure 3C and D) of METH-exposed mice compared to the Sal-exposed mice, while, there was no difference in the total AKT expression between groups in both regions. These findings suggest that the downregulation of p-AKT and HO-1 expression in both the PFC and CPu may be associated with METH withdrawal-induced negative emotions and cognitive deficits.
2.4. Luteolin Pretreatment Potentially Alleviates METH Withdrawal-Induced Anxiety and Depressive-Like Behaviors, as Well as Cognitive Deficits
To examine the effects of luteolin on METH withdrawal-induced affective and cognitive dysfunctions, we pretreated luteolin 30 min before METH administration and assessed the changes in METH withdrawal-induced anxiety and depressive-like behaviors, as well as cognitive functions (Figure 4A). For anxiety-like behaviors, luteolin pretreatment reversed the METH withdrawal-induced decrease in time spent and number of entries in the open arms of EPM (Figure 4B). There was also a trend of increase in central time spent in the OFT in the luteolin + METH group compared to the vehicle + METH (Veh + METH) group (Figure 4C). Regarding cognitive function, luteolin treatment alleviated the METH withdrawal-induced decrease in recognition index in the 24 h-interval NORT, but not in the 3 h-interval test (Figure 4D). However, no difference in the spontaneous alterations was found among groups in the Y-maze test (Figure 4E). Similarly, for the depression-like behaviors, luteolin treatment prevented the METH withdrawal-induced decrease in sucrose preference in SPT (Figure 4F). Meanwhile, there was a trend of decrease in the immobility time in the luteolin + METH group compared to the Veh + METH group in the FST (Figure 4G). Collectively, these results suggest that luteolin pretreatment partially alleviates METH withdrawal-induced anxiety and depressive like behaviors, as well as cognitive deficits.
2.5. Luteolin Attenuates METH Withdrawal-Induced Abnormal Activation of Neurons in the PFC and CPu
To verify the correlations between the luteolin-induced alleviation of METH withdrawal-associated negative affective/cognitive dysfunctions and neuronal activity changes in the PFC and CPu, we measured the c-Fos expression following luteolin pretreatment in these two brain regions. We found that luteolin treatment inhibited the METH withdrawal-induced activation of neurons in the PFC (Figure 5A and B) and CPu (Figure 5C and D), demonstrating decreased c-Fos-positive cell densities in the luteolin + METH group compared to the Veh + METH group. These results indicate that the protective effects of luteolin on METH withdrawal-induced negative emotions and cognitive deficits may be associated with a downregulation of abnormal neuronal activation in the PFC and CPu.
2.6. Luteolin Prevents METH Withdrawal-Induced Downregulation of p-AKT and HO-1 Expression in the PFC and CPu
To elucidate the potential molecular mechanisms underlying the effects of luteolin on METH withdrawal-induced negative emotions and cognitive deficits, we examined the AKT and HO-1 expression level in the PFC and CPu. Our results demonstrated that luteolin pretreatment reversed the METH withdrawal-induced downregulation of p-AKT and HO-1 expression in the PFC (Figure 6A and B) and CPu (Figure 6C and D). These findings suggest that luteolin may exert a protective effect on METH withdrawal-induced affective and cognitive dysfunctions by potentially activating AKT/HO-1 signaling in the PFC and CPu.
2.7. Luteolin Inhibits METH Withdrawal-Induced Nuclear Translocation of FOXO1
We further investigated the FOXO1 expression dynamics in the PFC (Figure 7A to F) and CPu (Figure 7G to L) following luteolin pretreatment. Immunofluorescence analysis demonstrated that prolonged METH-withdrawal led to an accumulation of FOXO1 in the nucleus of neural cells, suggesting an activation of FOXO1. Luteolin pretreatment could inhibit the nuclear accumulation of FOXO1 in both the PFC (Figure 7A) and CPu (Figure 7G). Next, by using fluorescence co-localization, we found that the fluorescence signals of FOXO1 and the nucleus exhibited strong spatial overlap in the Veh + METH group in both the PFC and CPu (Figure 7B and H). The scatter plot also showed a linear distribution of points along the diagonal in the Veh + METH group in these two brain regions (Figure 7C and I). The above results indicate that METH withdrawal induces co-localization between FOXO1 and the nucleus, suggesting the nuclear translocation of FOXO1. Notably, luteolin pretreatment could attenuate the METH withdrawal-induced nuclear translocation of FOXO1 in both the PFC and CPu. Pearson’s correlation analysis revealed a significant interaction between METH withdrawal and luteolin treatment, with a marked decrease in the R value in the luteolin + METH group compared to the Veh + METH group (Figure 7D and J). Additionally, Western blotting analysis of nuclear, cytoplasmic, and total FOXO1 levels further validated that luteolin treatment mitigated the METH withdrawal-induced increase in nuclear FOXO1 expression (Figure 7E, F, K, and L). Overall, our findings suggest that luteolin pretreatment attenuates the METH withdrawal-induced activation of FOXO1 by inhibiting its nuclear translocation.
3. Discussion
METH withdrawal-induced negative emotions and cognitive deficits cause substantial health and economic burdens to society worldwide [37,38]. However, the underlying mechanisms remain poorly understood, and effective treatments are limited. In the present study, we investigated the role and potential mechanisms of luteolin in METH withdrawal-induced affective and cognitive dysfunctions. Our results demonstrated that METH withdrawal led to anxiety and depressive-like behaviors, as well as cognitive deficits. Neuronal activity was increased and p-AKT/HO-1 expression was decreased in the PFC and CPu after prolonged METH withdrawal. Interestingly, luteolin pretreatment could partially ameliorate the METH withdrawal-induced affective and cognitive dysfunctions, and reverse METH withdrawal-induced abnormal neuron activation and the downregulation of p-AKT/HO-1 signaling. The potentially protective effects of luteolin on METH-induced affective and cognitive dysfunctions may be attributable to the AKT-mediated activation of HO-1 expression and to a decrease in the nuclear translocation of transcription factor FOXO1.
Neuronal activity alterations are critical for neural plasticity in both physiological and pathological conditions, such as aging and drug addiction [39,40]. METH use has been shown to increase neuronal activities, demonstrated as an increase in c-Fos expression, in the orbitofrontal cortex (OFC) and dorsal striatum (CPu) [19,41]. We consistently found that METH withdrawal led to the activation of the brain regions of the PFC and CPu. PFC and CPu have long been implicated in addictive drug-induced rewarding and reinforcing effects [19,42]. However, few studies have investigated their roles in METH withdrawal-induced affective dysfunctions. Interestingly, our findings suggest that neuronal activation in the PFC and CPu may be associated with METH withdrawal-induced negative emotions, offering new insights into their potential roles in drug addiction.
Beyond neuronal activity, gene expression changes also represent long-term neural adaptation after METH exposure [43]. We observed that the expression levels of p-AKT and HO-1 were decreased after prolonged METH withdrawal. AKT-induced activation of HO-1 is known for its antioxidative properties [25,44]. Previous findings have indicated that acute METH exposure increases PI3K/AKT [45] and HO-1 [46] expression, demonstrating compensatory and protective mechanisms to oxidative stress and apoptosis in the brain caused by METH. In accordance with our findings, chronic METH use has been shown to downregulate the AKT and HO-1 expression [25]. Hence, we speculate that the prolonged METH withdrawal-induced downregulation of p-AKT and HO-1 in our study might represent a decompensated response of METH-induced oxidative stress, neuroinflammation, and neuronal apoptosis.
Luteolin is a natural flavonoid compound widely present in medicinal plants [47,48]. Mounting evidence has proven that luteolin has antioxidative and anti-inflammatory properties, alleviating various kinds of diseases such as inflammatory bowel disease [49], cancers [50] and brain injuries [51]. In the context of METH misuse-associated disorders, previous studies have revealed that luteolin prevents METH-induced neurotoxicity [52] and hepatotoxicity [53] by inhibiting apoptosis, autophagy, and inflammation. We found that luteolin pretreatment could partially alleviate METH withdrawal-induced negative emotions and cognitive deficits (episodic memory in NORT, but not spatial memory in Y-maze). Moreover, luteolin prevented prolonged METH withdrawal-induced abnormal activation of the PFC and CPu, as well as the downregulation of AKT/HO-1 signaling. The activation of AKT/HO-1 signaling has been shown to exert anti-apoptotic and antioxidant effects [54,55]. Thus, we suppose that luteolin may relieve affective and cognitive dysfunctions during METH withdrawal by suppressing METH-induced oxidative injuries and neuronal apoptosis in the PFC and CPu through the activation of AKT /HO-1 signaling.
FOXO1 is a pro-apoptotic and pro-inflammation transcription factor. The overexpression of FOXO-1 in macrophages has been shown to increase apoptosis and pro-inflammation cytokine expression, such as that of IL-1beta [56]. The nuclear translocation of FOXO1 accelerates proapoptotic gene expression and causes neuronal death [57]. The inhibition of FOXO1 could attenuate copper-induced apoptosis in neural stem cells [58] and chidamide-triggered pyroptosis in T lymphoblasts [59]. Moreover, the FOXO1 inhibitor suppresses the mevastatin-induced increase in HO-1 expression in cardiac fibroblast and attenuates the anti-inflammatory and antioxidative effects of mevastatin [60]. Unfortunately, few studies have revealed the role of FOXO1 in addictive substance use disorders to date. Our findings demonstrated that METH withdrawal increases but luteolin decreases the nuclear translocation of FOXO1. This may be one of the potential mechanisms underlying the protective effects of luteolin on alleviating METH withdrawal-induced neuronal apoptosis and oxidative injuries, as well as the negative emotions and cognitive deficits.
Our study has several limitations. First, we only investigated the neuronal activity and AKT/FOXO1/HO-1 expression in two representative brain regions (PFC and CPu) in METH withdrawal-induced negative emotions and cognitive deficits. The sub-region functions of these anatomically and functionally heterogeneous brain regions and their projection relationships, interactions of different kinds of neurons, as well as other molecular cascades in other brain regions in the context of METH withdrawal related cognitive and affective disorders still need to be uncovered. Second, we could not verify the specificity of the neuronal activity and AKT/FOXO1/HO-1 signaling changes to the negative emotions and cognitive deficits. Third, although the potential molecular mechanisms underlying the protective effects of luteolin on METH withdrawal-induced negative emotions and cognitive deficits have been preliminary revealed in our study, the detailed and precise molecular cascades and the cellular and synaptic plasticity mechanisms were not fully elucidated. Additionally, we pretreated mice with luteolin before METH in the present study. It would be better to administer luteolin after METH exposure from the view of translational relevance. Therefore, caution should be taken when interpreting the effects of luteolin on alleviating METH withdrawal-induced negative emotions and cognitive deficits. These aspects also provide directions for future research to further explore the in-depth mechanisms and potential invention strategies for METH-associated health issues.
In summary, our results demonstrate that prolonged METH withdrawal increases neuronal activity and suppresses AKT/HO-1 signaling in the PFC and CPu, potentially contributing to negative emotions and cognitive deficits. Luteolin pretreatment may mitigate these effects by normalizing neuronal activity and restoring AKT/HO-1 expression. Furthermore, luteolin reduces the nuclear translocation of FOXO1. These findings suggest that the potentially protective effects of luteolin on METH withdrawal-induced affective and cognitive dysfunctions may stem from attenuating METH-induced oxidative stress and neuronal apoptosis through the AKT/FOXO1/HO-1 pathway. Our results highlight the therapeutic potential of luteolin and its derivatives for treating METH-associated neurological disorders.
4. Materials and Methods
4.1. Animals
A total of 60 male C57BL/6J mice (8 weeks old) were purchased from Vital River Laboratory Animal Technology (Beijing, China). The mice were housed in cages in groups of four with water and standard chow available ad libitum in a controlled environment (temperature of 20℃-24℃, humidity of 40%-60%, and a 12/12h light/dark cycle). All mice were acclimatized to the environment for one week prior to the experiments to avoid stressful effects. Our study protocols adhered to the Guidelines for the Care and Use of Laboratory Animals issued by the National Institutes of Health, USA. Ethical approval for our study was obtained from the Institutional Animal Care and Use Committee of Xi’an Jiaotong University.
4.2. Reagents and Drug Administration Procedure
Methamphetamine hydrochloride powder (China Pharmaceutical and Biological Products, 99.9% purity) was dissolved in 0.9% Sal. Luteolin (Med Chem Express, USA) was dissolved in 0.9% Sal with 5% dimethyl sulfoxide (DMSO, Solarbio, China), 20% polyethylene glycol 300 (PEG300, Solarbio, China), and 2.5% polysorbate 80 (TWEEN 80, Solarbio, China) to promote dissolution. The dose of METH was 3mg/kg and that of luteolin was 20 mg/kg based on previous studies [61,62]. All drugs were freshly prepared before experiments and administered via intraperitoneal (i.p.) injection. To investigate the effects of luteolin on METH withdrawal-induced affective and cognitive dysfunctions, mice were randomly divided into four groups (n = 8 per group) and received one daily injection (i.p.) of Veh + Sal, Luteolin + Sal, Veh + METH, or Luteolin + METH. Luteolin was administrated 30 min before METH injections.
4.3. Behavioral Tests
4.3.1. Anxiety-Like Behaviors
(1) Open field test (OFT)
Mice were placed into the OFT chambers (43 cm length × 43 cm width × 43 cm height) to explore freely for 30 min and the locomotion trajectories were recorded. The total distance traveled, central area distance traveled, and time spent in central area for each mouse were recorded using Smart 3.0 software (Panlab Technology for Bioresearch, Spain).
(2) Elevated plus maze (EPM) test
The EPM apparatus consisted of a central zone (6 cm length× 6 cm width × 6 cm height), two open arms (33 cm length × 6 cm width), and two closed arms (33 cm length × 6 cm width × 6 cm height). Mice were placed into the central zone. Then, the time spent in each arm and the arm entry times were recorded for 5 min using Smart 3.0 software. The percentage of the open arm time was calculated as open arm time / (open arm time + closed arm time).
4.3.2. Cognitive Function Detection
(1) Novel object recognition task (NORT)
The NORT consisted of 3 phases, including the adaptation phase (day 1-2), learning phase (day 3) and test phase (day 3-4). The adaptation phase lasted for 2 days, during which mice were placed in the NORT chambers (30 cm length × 20 cm width × 30 cm height) for 10 min to adapt the apparatus. In the learning phase, two identical objects were put into the chambers 10 cm apart from each other and the side walls. Mice were placed into the chambers to explore for 5 min each time, for 3 times in total with a 15 min interval between each exploration. Three and twenty-four hours after the end of the learning phase, one of the above same objects was replaced with a novel one. Then, mice were placed into the chambers to explore for 5 min and the time spent accessing each object was recorded with Smart3.0 software. The recognition index was calculated as novel object exploring time / (novel object exploring time + familiar object exploring time).
(2) Y-maze test
The Y-maze apparatus consisted of 3 equal arms (30 cm length × 6 cm width × 15 cm height) with an angle of 120°. Mice were placed in the center area and allowed to explore for 5 min. Entering the 3 arms consecutively was defined as a successful alternation. The percentage of alternation was calculated as number of successful alternations / (total arm entries - 2).
4.3.3. Depressive-Like Behaviors
(1) Sucrose preference test (SPT)
The SPT consisted of 2 phases, including the training phase (days 1-3) and test phase (day 5). During the training phase on day 1, mice were put into cages separately for 24 hours with 2 bottles of 1% sucrose water and free food. On days 2-3, one of the 1% sucrose water was replaced with a bottle of normal drinking water, and the positions of sucrose water and drinking water were exchanged at the 24th hour point to avoid position preference. After the training phase, the mice were deprived of water and food for 24 hours. On day 5, the mice were given one bottle of 1% sucrose water and one bottle of drinking water for 24 hours, and the positions of two bottles were exchanged in the middle of the test. The weight loss of the two bottles was recorded. The sucrose preference ratio was calculated as sucrose consumption / (sucrose consumption + normal water consumption).
(2) Forced swim test (FST)
The FST apparatus consisted of several cylindrical transparent tanks (40 cm height × 25 cm diameter). During the FST period, each tank was filled to 30 cm high with water, with a water temperature of 25 - 27 ℃. The total immobile time during the 6 min test was recorded using Smart3.0 software.
4.4. Sample Preparation and Western Blotting
After the behavioral tests, mice were anesthetized with isoflurane and immediately decapitated, and the PFC and CPu regions of the brain were separated on ice. The total protein samples were added with RIPA lysis buffer (Beyotime, China) with 1mM phenylmethylsulfonyl fluoride (PMSF, Beyotime, China) and 1mM phosphatase inhibitor and ultrasonication for extracting the proteins. After centrifuging at 12,000 g for 15 min at 4 ℃, the supernatants were collected and mixed with 5 × loading buffer and then boiled for 5 min. The nuclear and cytoplasmic samples were processed following the instructions provided in the kit (Beyotime, China). The protein concentration was measured using a bicinchoninic acid (BCA) assay (Solarbio, China). Equal amounts of protein (20μg) were loaded onto 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto 0.22 μm polyvinylidene fluoride (PVDF) membranes (Millipore, Darmstadt, Germany). The membranes were blocked with Protein Free Rapid Blocking Buffer (Epizyme, China) at room temperature for 15 min. Next, the PVDF membranes were incubated with primary antibody at 4 ℃ overnight and washed with phosphate-buffered Sal with 0.1% Tween-20 (1 × PBST) three times (10 min per wash) after incubation. After washing, the membranes were incubated with secondary antibody at room temperature for 2 h. Finally, an enhanced chemiluminescence (ECL) kit (Mishushengwu, China) method was used to detect the protein. The expression intensities of the protein bands were analyzed using ImageJ software (National Institute of Health, USA). The primary antibodies used in this study were: p-AKT (Thr 308, 1:1000, Cell Signaling Technology, USA), AKT (1:2000, Cell Signaling Technology, USA), HO-1 (1:2000, Proteintech, China), FOXO1 (1:2000, Proteintech, China), PCNA (1:2000, Proteintech, China), and β-actin (1:5000, Proteintech, China).
4.5. Sample Preparation and Immunofluorescence
Mice were anesthetized with 5% sodium pentobarbital, and then transcardially perfused with 0.9% Sal followed by 4% paraformaldehyde. The brains were collected and post-fixed in 4% paraformaldehyde for 24 h. Then, the brains were removed into 30% sucrose solution dissolved in 0.2M phosphate buffer (PB, pH 7.4) for 48 h of dehydration. The brains were embedded in an optimal-cutting-temperature compound (OCT, Sakura, USA), and cut into 20 μm thick coronal sections on microtome (Leica, Germany). The sections were washed 3 times (5 min per wash) with 1 × PBS and then blocked in 5% goat serum solution (Boster, China) with 0.3% Triton X-100 (Servicebio, China) for 2 h at room temperature. Following this, the sections were incubated with c-Fos (1:400, Cell Signaling Technology, USA), FOXO1 (1:200, Proteintech, China) overnight at 4℃. After washing, the sections were incubated with secondary antibody (1:400, Proteintech, China) for 2 h at room temperature and then washed, and mounted for observation. AntiFade mounting medium (with DAPI) (Servicebio, China) was used to observe the nucleus of the cells. All sections were observed using the fluorescence microscope (Zeiss, Germany). The cFos positive cells were quantified using ImageJ software, with consistent parameter settings applied across all groups. The colocalization analysis and visualization were performed using the colocalization and scatter J tools in ImageJ software.
4.6. Statistical Analysis
GraphPad Prism (version 9.0) was used for all statistical analyses. All data are presented as means ± SEMs. Shapiro-Wilk and Bartlett’s test were used for the normal distribution and homogeneity of variance tests. All the data were normally distributed. Then, data were analyzed using Student’s t-test (two group comparisons) or two-way ANOVA (four group comparisons), accordingly. All post hoc pairwise comparisons were performed using the Bonferroni test. P values < 0.05 were considered statistically significant.
Author Contributions
J.Z. and W.H.: design and conceptualization of the study, revising the manuscript, and obtaining funding. B.G. and R.A.: conducting experiments, analysis and interpretation of the data, drafting the manuscript. M.L., X.W., and J.P.: conducting experiments and revising the manuscript. X.C. and Z.L.: reagents preparation and sample collection. T.L. and X.L.: revising the manuscript. All authors have read and approved the final version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (No. 82101974 and 82371892), the Project of Shaanxi Administration of Traditional Chinese Medicine (NO.2021-ZZ-JC037), the Key Laboratory of Drugs analysis & intelligent-Montoring (No. NNLS-0-202304), and the Natural Science Basic Research Program of Shaanxi (No. 2024JC-YBQN-0894).
Institutional Review Board Statement
The protocols of the present study were approved by the Institutional Animal Care Committee of Xi’an Jiaotong University (No. XJTUAE2023-1268).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data in the present study are available from the corresponding authors upon reasonable request.
Acknowledgments
We would like to thank all the members of our team for their support, and we are grateful to Li Wen (Biomedical Experimental Center of Xi’an Jiaotong University) for their technical support.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| BBB | blood-brain barrier |
| PFC | prefrontal cortex |
| CPu | caudate putamen |
| AKT | protein kinase B |
| FOXO1 | forkhead box protein 1 |
| HO-1 | heme-oxygenase-1 |
| PI3K | phosphoinositide 3-kinase |
| EPM | elevated plus maze |
| OFT | open field test |
| NORT | novel object recognition task |
| SPT | sucrose preference test |
| FST | forced swim test |
References
- Zhang B, Yan X, Li Y, Zhu H, Lu Z, Jia Z: Trends in Methamphetamine Use in the Mainland of China, 2006-2015. Frontiers In Public Health 2022, 10:852837.
- Pang L, Wang Y: Overview of blood-brain barrier dysfunction in methamphetamine abuse. Biomedicine & Pharmacotherapy = Biomedecine & Pharmacotherapie 2023, 161:114478.
- Zhao X, Lu J, Chen X, Gao Z, Zhang C, Chen C, Qiao D, Wang H: Methamphetamine exposure induces neuronal programmed necrosis by activating the receptor-interacting protein kinase 3 -related signalling pathway. Faseb j 2021, 35:e21561. [CrossRef]
- Bellot M, Soria F, López-Arnau R, Gómez-Canela C, Barata C: Daphnia magna an emerging environmental model of neuro and cardiotoxicity of illicit drugs. Environ Pollut 2024, 344:123355. [CrossRef]
- Wang X, Hui R, Li Q, Lu Y, Wang M, Shi Y, Xie B, Cong B, Ma C, Wen D: Gut microbiota-derived trimethylamine N-oxide involved in methamphetamine-induced depression-like behaviors of male mice. Neuropharmacology 2025, 268:110339. [CrossRef]
- Hui R, Xu J, Zhou M, Xie B, Zhou M, Zhang L, Cong B, Ma C, Wen D: Betaine improves METH-induced depressive-like behavior and cognitive impairment by alleviating neuroinflammation via NLRP3 inflammasome inhibition. Prog Neuropsychopharmacol Biol Psychiatry 2024, 135:111093. [CrossRef]
- Wang X, Hu M, Chen J, Lou X, Zhang H, Li M, Cheng J, Ma T, Xiong J, Gao R, et al: Key roles of autophagosome/endosome maturation mediated by Syntaxin17 in methamphetamine-induced neuronal damage in mice. Mol Med 2024, 30:4. [CrossRef]
- Wang X, Hu M, Wu W, Lou X, Gao R, Ma T, Dheen ST, Cheng J, Xiong J, Chen X, Wang J: Indole derivatives ameliorated the methamphetamine-induced depression and anxiety via aryl hydrocarbon receptor along “microbiota-brain” axis. Gut Microbes 2025, 17:2470386. [CrossRef]
- Zhang K, Chen L, Yang J, Liu J, Li J, Liu Y, Li X, Chen L, Hsu C, Zeng J, et al: Gut microbiota-derived short-chain fatty acids ameliorate methamphetamine-induced depression- and anxiety-like behaviors in a Sigmar-1 receptor-dependent manner. Acta Pharm Sin B 2023, 13:4801-4822. [CrossRef]
- Ma J, Sun XJ, Wang RJ, Wang TY, Su MF, Liu MX, Li SX, Han Y, Meng SQ, Wu P, et al: Profile of psychiatric symptoms in methamphetamine users in China: Greater risk of psychiatric symptoms with a longer duration of use. Psychiatry Res 2018, 262:184-192. [CrossRef]
- Zhang J, Xie Y, Su H, Tao J, Sun Y, Li L, Liang H, He R, Han B, Lu Y, et al: Prevalence and correlates of depressive symptoms during early methamphetamine withdrawal in Han Chinese population. Drug Alcohol Depend 2014, 142:191-196. [CrossRef]
- Pratelli M, Hakimi AM, Thaker A, Jang H, Li HQ, Godavarthi SK, Lim BK, Spitzer NC: Drug-induced change in transmitter identity is a shared mechanism generating cognitive deficits. Nat Commun 2024, 15:8260. [CrossRef]
- Shilling PD, Kuczenski R, Segal DS, Barrett TB, Kelsoe JR: Differential regulation of immediate-early gene expression in the prefrontal cortex of rats with a high vs low behavioral response to methamphetamine. Neuropsychopharmacology 2006, 31:2359-2367. [CrossRef]
- Li J, Zhu L, Su H, Liu D, Yan Z, Ni T, Wei H, Goh ELK, Chen T: Regulation of miR-128 in the nucleus accumbens affects methamphetamine-induced behavioral sensitization by modulating proteins involved in neuroplasticity. Addict Biol 2021, 26:e12881. [CrossRef]
- Vilca SJ, Margetts AV, Höglund L, Fleites I, Bystrom LL, Pollock TA, Bourgain-Guglielmetti F, Wahlestedt C, Tuesta LM: Microglia contribute to methamphetamine reinforcement and reflect persistent transcriptional and morphological adaptations to the drug. Brain Behav Immun 2024, 120:339-351. [CrossRef]
- Wang Y, Liu J, Yue S, Chen L, Singh A, Yu T, Calipari ES, Wang ZJ: Prefrontal cortex excitatory neurons show distinct response to heroin-associated cue and social stimulus after prolonged heroin abstinence in mice. Neuropsychopharmacology 2025. [CrossRef]
- Ding J, Huang J, Tang X, Shen L, Hu S, He J, Liu T, Yu Z, Liu Y, Wang Q, et al: Low and high dose methamphetamine differentially regulate synaptic structural plasticity in cortex and hippocampus. Front Cell Neurosci 2022, 16:1003617. [CrossRef]
- Hu YB, Deng X, Liu L, Cao CC, Su YW, Gao ZJ, Cheng X, Kong D, Li Q, Shi YW, et al: Distinct roles of excitatory and inhibitory neurons in the medial prefrontal cortex in the expression and reconsolidation of methamphetamine-associated memory in male mice. Neuropsychopharmacology 2024, 49:1827-1838. [CrossRef]
- Liu M, Mu S, Han W, Tan X, Liu E, Hang Z, Zhu S, Yue Q, Sun J: Dopamine D1 receptor in orbitofrontal cortex to dorsal striatum pathway modulates methamphetamine addiction. Biochem Biophys Res Commun 2023, 671:96-104. [CrossRef]
- Barbano MF, Qi J, Chen E, Mohammad U, Espinoza O, Candido M, Wang H, Liu B, Hahn S, Vautier F, Morales M: VTA glutamatergic projections to the nucleus accumbens suppress psychostimulant-seeking behavior. Neuropsychopharmacology 2024, 49:1905-1915. [CrossRef]
- Wang M, Xu L, Zhao D, Wang W, Xu L, Cao Y, Meng F, Liu J, Li C, Jiang S: The glutamatergic projections from the PVT to mPFC govern methamphetamine-induced conditional place preference behaviors in mice. J Affect Disord 2025, 371:289-304. [CrossRef]
- Chen T, Su H, Li R, Jiang H, Li X, Wu Q, Tan H, Zhang J, Zhong N, Du J, et al: The exploration of optimized protocol for repetitive transcranial magnetic stimulation in the treatment of methamphetamine use disorder: A randomized sham-controlled study. EBioMedicine 2020, 60:103027. [CrossRef]
- Mukherjee D, Gonzales BJ, Ashwal-Fluss R, Turm H, Groysman M, Citri A: Egr2 induction in spiny projection neurons of the ventrolateral striatum contributes to cocaine place preference in mice. Elife 2021, 10. [CrossRef]
- Wei T, Li JD, Wang YJ, Zhao W, Duan F, Wang Y, Xia LL, Jiang ZB, Song X, Zhu YQ, et al: p-Nrf2/HO-1 Pathway Involved in Methamphetamine-induced Executive Dysfunction through Endoplasmic Reticulum Stress and Apoptosis in the Dorsal Striatum. Neurotox Res 2023, 41:446-458.
- Meng X, Zhang C, Guo Y, Han Y, Wang C, Chu H, Kong L, Ma H: TBHQ Attenuates Neurotoxicity Induced by Methamphetamine in the VTA through the Nrf2/HO-1 and PI3K/AKT Signaling Pathways. Oxid Med Cell Longev 2020, 2020:8787156.
- Yang W, Li Y, Tang Y, Tao Z, Yu M, Sun C, Ye Y, Xu B, Zhao X, Zhang Y, Lu X: Mesenchymal stem cells overexpressing neuropeptide S promote the recovery of rats with spinal cord injury by activating the PI3K/AKT/GSK3β signaling pathway. Stem Cell Res Ther 2025, 16:100.
- Wang L, Chen Y, Sternberg P, Cai J: Essential roles of the PI3 kinase/Akt pathway in regulating Nrf2-dependent antioxidant functions in the RPE. Invest Ophthalmol Vis Sci 2008, 49:1671-1678. [CrossRef]
- Deng S, Jin P, Sherchan P, Liu S, Cui Y, Huang L, Zhang JH, Gong Y, Tang J: Recombinant CCL17-dependent CCR4 activation alleviates neuroinflammation and neuronal apoptosis through the PI3K/AKT/Foxo1 signaling pathway after ICH in mice. J Neuroinflammation 2021, 18:62.
- Nageswaran V, Carreras A, Reinshagen L, Beck KR, Steinfeldt J, Henricsson M, Ramezani Rad P, Peters L, Strässler ET, Lim J, et al: Gut Microbial Metabolite Imidazole Propionate Impairs Endothelial Cell Function and Promotes the Development of Atherosclerosis. Arterioscler Thromb Vasc Biol 2025. [CrossRef]
- Koppula S, Wankhede NL, Sammeta SS, Shende PV, Pawar RS, Chimthanawala N, Umare MD, Taksande BG, Upaganlawar AB, Umekar MJ, et al: Modulation of cholesterol metabolism with Phytoremedies in Alzheimer’s disease: A comprehensive review. Ageing Res Rev 2024, 99:102389.
- Singh AA, Katiyar S, Song M: Phytochemicals Targeting BDNF Signaling for Treating Neurological Disorders. Brain Sci 2025, 15.
- Ma HY, Wang J, Wang J, Guo Z, Qin XY, Lan R, Hu Y: Luteolin attenuates cadmium neurotoxicity by suppressing glial inflammation and supporting neuronal survival. Int Immunopharmacol 2025, 152:114406. [CrossRef]
- Li H, Li JX, Zeng YD, Zheng CX, Dai SS, Yi J, Song XD, Liu T, Liu WH: Luteolin ameliorates ischemic/reperfusion injury by inhibiting ferroptosis. Metab Brain Dis 2025, 40:159.
- Luo S, Li H, Mo Z, Lei J, Zhu L, Huang Y, Fu R, Li C, Huang Y, Liu K, et al: Connectivity map identifies luteolin as a treatment option of ischemic stroke by inhibiting MMP9 and activation of the PI3K/Akt signaling pathway. Exp Mol Med 2019, 51:1-11.
- Wang Z, Zeng M, Wang Z, Qin F, Chen J, He Z: Dietary Luteolin: A Narrative Review Focusing on Its Pharmacokinetic Properties and Effects on Glycolipid Metabolism. Journal of Agricultural and Food Chemistry 2021, 69:1441-1454. [CrossRef]
- Kempuraj D, Thangavel R, Kempuraj DD, Ahmed ME, Selvakumar GP, Raikwar SP, Zaheer SA, Iyer SS, Govindarajan R, Chandrasekaran PN, Zaheer A: Neuroprotective effects of flavone luteolin in neuroinflammation and neurotrauma. BioFactors (Oxford, England) 2021, 47:190-197. [CrossRef]
- Jiang P, Sun J, Zhou X, Lu L, Li L, Xu J, Huang X, Li J, Gong Q: Dynamics of intrinsic whole-brain functional connectivity in abstinent males with methamphetamine use disorder. Drug Alcohol Depend Rep 2022, 3:100065. [CrossRef]
- Moulis L, Le SM, Hai VV, Huong DT, Minh KP, Oanh KTH, Rapoud D, Quillet C, Thi TTN, Vallo R, et al: Gender, homelessness, hospitalization and methamphetamine use fuel depression among people who inject drugs: implications for innovative prevention and care strategies. Front Psychiatry 2023, 14:1233844.
- Gooijers J, Pauwels L, Hehl M, Seer C, Cuypers K, Swinnen SP: Aging, brain plasticity, and motor learning. Ageing Res Rev 2024, 102:102569.
- Wang Y, Guo R, Chen B, Rahman T, Cai L, Li Y, Dong Y, Tseng GC, Fang J, Seney ML, Huang YH: Cocaine-induced neural adaptations in the lateral hypothalamic melanin-concentrating hormone neurons and the role in regulating rapid eye movement sleep after withdrawal. Mol Psychiatry 2021, 26:3152-3168. [CrossRef]
- Tanaka T, Ago Y, Umehara C, Imoto E, Hasebe S, Hashimoto H, Takuma K, Matsuda T: Role of Prefrontal Serotonergic and Dopaminergic Systems in Encounter-Induced Hyperactivity in Methamphetamine-Sensitized Mice. Int J Neuropsychopharmacol 2017, 20:410-421. [CrossRef]
- Li X, Xu X, Feng Q, Zhou N, He Y, Liu Y, Tai H, Kim HY, Fan Y, Guan X: Glutamatergic pathways from medial prefrontal cortex to paraventricular nucleus of thalamus contribute to the methamphetamine-induced conditioned place preference without affecting wakefulness. Theranostics 2025, 15:1822-1841. [CrossRef]
- Oladapo A, Deshetty UM, Callen S, Buch S, Periyasamy P: Single-Cell RNA-Seq Uncovers Robust Glial Cell Transcriptional Changes in Methamphetamine-Administered Mice. Int J Mol Sci 2025, 26. [CrossRef]
- Yin Y, Mu F, Zhang L, Zhao J, Gong R, Yin Y, Zheng L, Du Y, Jin F, Wang J: Wedelolactone activates the PI3K/AKT/NRF2 and SLC7A11/GPX4 signalling pathways to alleviate oxidative stress and ferroptosis and improve sepsis-induced liver injury. J Ethnopharmacol 2025, 344:119557.
- Shen S, Zhang M, Ma M, Rasam S, Poulsen D, Qu J: Potential Neuroprotective Mechanisms of Methamphetamine Treatment in Traumatic Brain Injury Defined by Large-Scale IonStar-Based Quantitative Proteomics. Int J Mol Sci 2021, 22.
- Yu H, Peng Y, Dong W, Shen B, Yang G, Nie Q, Tian Y, Qin L, Song C, Chen B, et al: Nrf2 attenuates methamphetamine-induced myocardial injury by regulating oxidative stress and apoptosis in mice. Hum Exp Toxicol 2023, 42:9603271231219488. [CrossRef]
- Yuan J, Zhang K, Yang L, Cheng X, Chen J, Guo X, Cao H, Zhang C, Xing C, Hu G, Zhuang Y: Luteolin attenuates LPS-induced damage in IPEC-J2 cells by enhancing mitophagy via AMPK signaling pathway activation. Front Nutr 2025, 12:1552890. [CrossRef]
- Zhan XZ, Bo YW, Zhang Y, Zhang HD, Shang ZH, Yu H, Chen XL, Kong XT, Zhao WZ, Teimonen T, et al: Luteolin inhibits diffuse large B-cell lymphoma cell growth through the JAK2/STAT3 signaling pathway. Front Pharmacol 2025, 16:1545779.
- Yang S, Duan H, Yan Z, Xue C, Niu T, Cheng W, Zhang Y, Zhao X, Hu J, Zhang L: Luteolin Alleviates Ulcerative Colitis in Mice by Modulating Gut Microbiota and Plasma Metabolism. Nutrients 2025, 17. [CrossRef]
- Mahwish, Imran M, Naeem H, Hussain M, Alsagaby SA, Al Abdulmonem W, Mujtaba A, Abdelgawad MA, Ghoneim MM, El-Ghorab AH, et al: Antioxidative and Anticancer Potential of Luteolin: A Comprehensive Approach Against Wide Range of Human Malignancies. Food Sci Nutr 2025, 13:e4682. [CrossRef]
- Kempuraj D, Thangavel R, Kempuraj DD, Ahmed ME, Selvakumar GP, Raikwar SP, Zaheer SA, Iyer SS, Govindarajan R, Chandrasekaran PN, Zaheer A: Neuroprotective effects of flavone luteolin in neuroinflammation and neurotrauma. Biofactors 2021, 47:190-197.
- Tan XH, Zhang KK, Xu JT, Qu D, Chen LJ, Li JH, Wang Q, Wang HJ, Xie XL: Luteolin alleviates methamphetamine-induced neurotoxicity by suppressing PI3K/Akt pathway-modulated apoptosis and autophagy in rats. Food Chem Toxicol 2020, 137:111179.
- Zhang KK, Wang H, Qu D, Chen LJ, Wang LB, Li JH, Liu JL, Xu LL, Yoshida JS, Xu JT, et al: Luteolin Alleviates Methamphetamine-Induced Hepatotoxicity by Suppressing the p53 Pathway-Mediated Apoptosis, Autophagy, and Inflammation in Rats. Front Pharmacol 2021, 12:641917.
- Wang TJ, Hou WC, Hsiao BY, Lo TH, Chen YT, Yang CH, Shih YT, Liu JC: 2-Hydroxyl hispolon reverses high glucose-induced endothelial progenitor cell dysfunction through the PI3K/Akt/eNOS and AMPK/HO-1 pathways. Br J Pharmacol 2025.
- Guo M, Fu W, Zhang X, Li T, Ma W, Wang H, Wang X, Feng S, Sun H, Zhang Z, et al: Total flavonoids extracted from the leaves of Murraya paniculata (L.) Jack prevents acetaminophen-induced liver injury by activating Keap1/Nrf2 and PI3K/AKT/mTOR signaling pathway. J Ethnopharmacol 2025, 344:119562. [CrossRef]
- Gupta P, Srivastav S, Saha S, Das PK, Ukil A: Leishmania donovani inhibits macrophage apoptosis and pro-inflammatory response through AKT-mediated regulation of β-catenin and FOXO-1. Cell Death Differ 2016, 23:1815-1826. [CrossRef]
- Zhou J, Li H, Li X, Zhang G, Niu Y, Yuan Z, Herrup K, Zhang YW, Bu G, Xu H, Zhang J: The roles of Cdk5-mediated subcellular localization of FOXO1 in neuronal death. J Neurosci 2015, 35:2624-2635.
- Zhang Y, Wang M, Tang L, Yang W, Zhang J: FoxO1 silencing in Atp7b(-/-) neural stem cells attenuates high copper-induced apoptosis via regulation of autophagy. J Neurochem 2024, 168:2762-2774.
- Li X, Liu B, Huang D, Ma N, Xia J, Zhao X, Duan Y, Li F, Lin S, Tang S, et al: Chidamide triggers pyroptosis in T-cell lymphoblastic lymphoma/leukemia via the FOXO1/GSDME axis. Chin Med J (Engl) 2024.
- Lee IT, Yang CC, Lin YJ, Wu WB, Lin WN, Lee CW, Tseng HC, Tsai FJ, Hsiao LD, Yang CM: Mevastatin-Induced HO-1 Expression in Cardiac Fibroblasts: A Strategy to Combat Cardiovascular Inflammation and Fibrosis. Environ Toxicol 2025, 40:264-280. [CrossRef]
- Ren Q, Ma M, Yang C, Zhang JC, Yao W, Hashimoto K: BDNF-TrkB signaling in the nucleus accumbens shell of mice has key role in methamphetamine withdrawal symptoms. Transl Psychiatry 2015, 5:e666.
- Kou JJ, Shi JZ, He YY, Hao JJ, Zhang HY, Luo DM, Song JK, Yan Y, Xie XM, Du GH, Pang XB: Luteolin alleviates cognitive impairment in Alzheimer’s disease mouse model via inhibiting endoplasmic reticulum stress-dependent neuroinflammation. Acta Pharmacol Sin 2022, 43:840-849. [CrossRef]
Figure 1.
METH withdrawal-induced negative emotions and cognitive deficits. (A) Experimental procedure. (B-E) Anxiety-like behaviors. (B and D) Time spent and number of entries in the open arm (%) in the EPM test. (C and E) Time spent in the central zone (%) and total distance traveled in the OFT. (F-G) Cognitive function (episodic and spatial memories) test. (F) Time spent accessing the new object (%) in the NORT. (G) Spontaneous alternations (%) in the Y-maze test. (H-I) Depressive-like behaviors. (H) Sucrose consumption (%) in the sucrose preference test. (I) Immobility time in the FST. Data are expressed as the mean ± SEM; n = 8/group. * P < 0.05, ** P < 0.01, *** P < 0.001, compared to the Sal group. Sal, saline; METH, methamphetamine; EPM, elevated plus maze; OFT, open field test; NORT, novel object recognition test; SPT, sucrose preference test; FST, forced swim test.
Figure 1.
METH withdrawal-induced negative emotions and cognitive deficits. (A) Experimental procedure. (B-E) Anxiety-like behaviors. (B and D) Time spent and number of entries in the open arm (%) in the EPM test. (C and E) Time spent in the central zone (%) and total distance traveled in the OFT. (F-G) Cognitive function (episodic and spatial memories) test. (F) Time spent accessing the new object (%) in the NORT. (G) Spontaneous alternations (%) in the Y-maze test. (H-I) Depressive-like behaviors. (H) Sucrose consumption (%) in the sucrose preference test. (I) Immobility time in the FST. Data are expressed as the mean ± SEM; n = 8/group. * P < 0.05, ** P < 0.01, *** P < 0.001, compared to the Sal group. Sal, saline; METH, methamphetamine; EPM, elevated plus maze; OFT, open field test; NORT, novel object recognition test; SPT, sucrose preference test; FST, forced swim test.
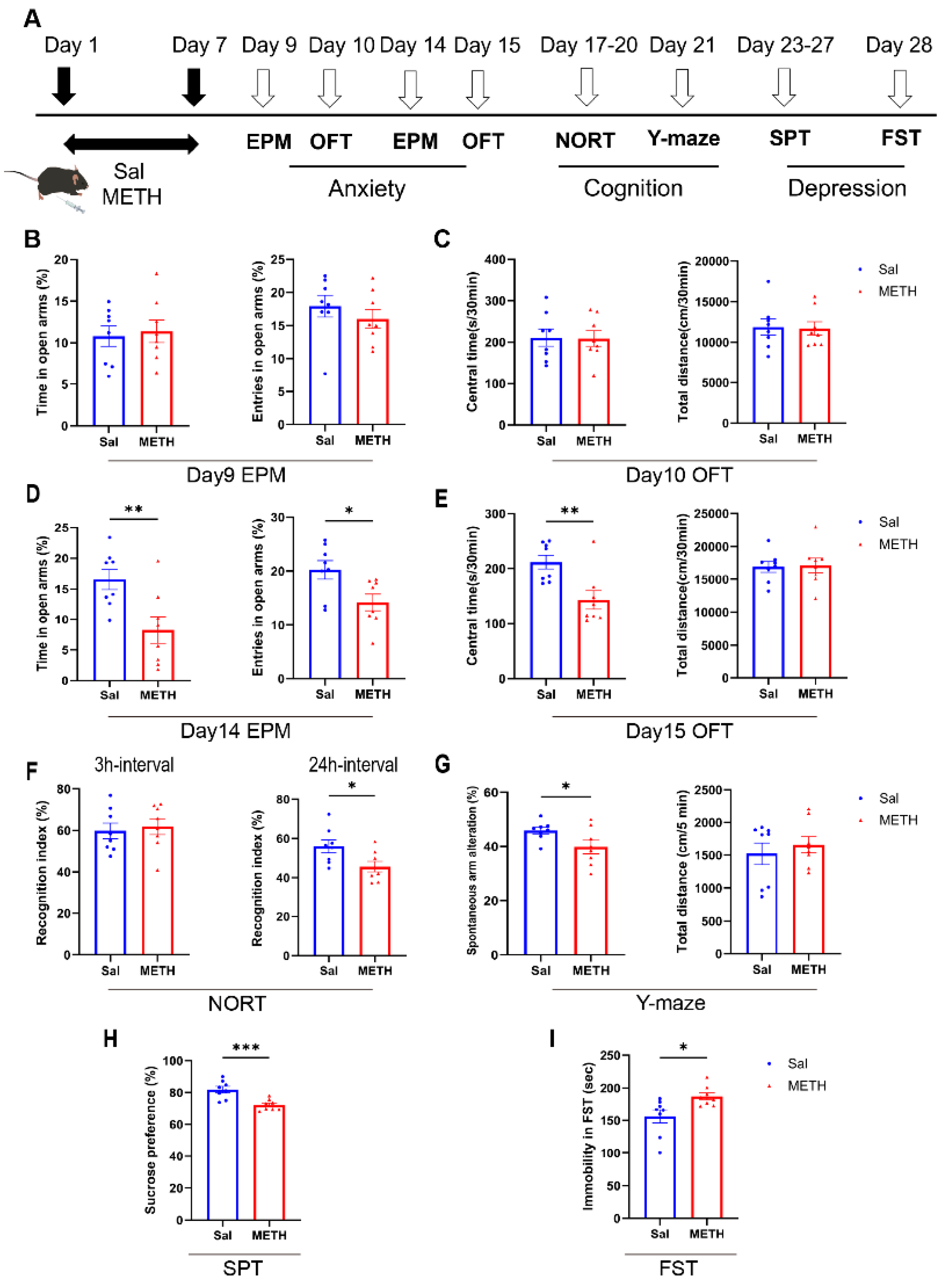
Figure 2.
METH withdrawal activated c-Fos expression in the PFC and CPu. (A-B) Representative images of c-Fos staining and quantitative analysis of the density of c-Fos positive cells in the PFC. (C-D) Representative images of c-Fos staining and quantitative analysis of the density of c-Fos positive cells in the CPu. (A and C). Scale bars represent 200 μm under low magnification and 50 μm under high magnification. Data are presented as the mean ± SEM; n = 3/group, 2 slices per mouse. **** P < 0.0001, compared to the Sal group. Sal, saline; METH, methamphetamine; PFC, prefrontal cortex; CPu, caudate putamen.
Figure 2.
METH withdrawal activated c-Fos expression in the PFC and CPu. (A-B) Representative images of c-Fos staining and quantitative analysis of the density of c-Fos positive cells in the PFC. (C-D) Representative images of c-Fos staining and quantitative analysis of the density of c-Fos positive cells in the CPu. (A and C). Scale bars represent 200 μm under low magnification and 50 μm under high magnification. Data are presented as the mean ± SEM; n = 3/group, 2 slices per mouse. **** P < 0.0001, compared to the Sal group. Sal, saline; METH, methamphetamine; PFC, prefrontal cortex; CPu, caudate putamen.
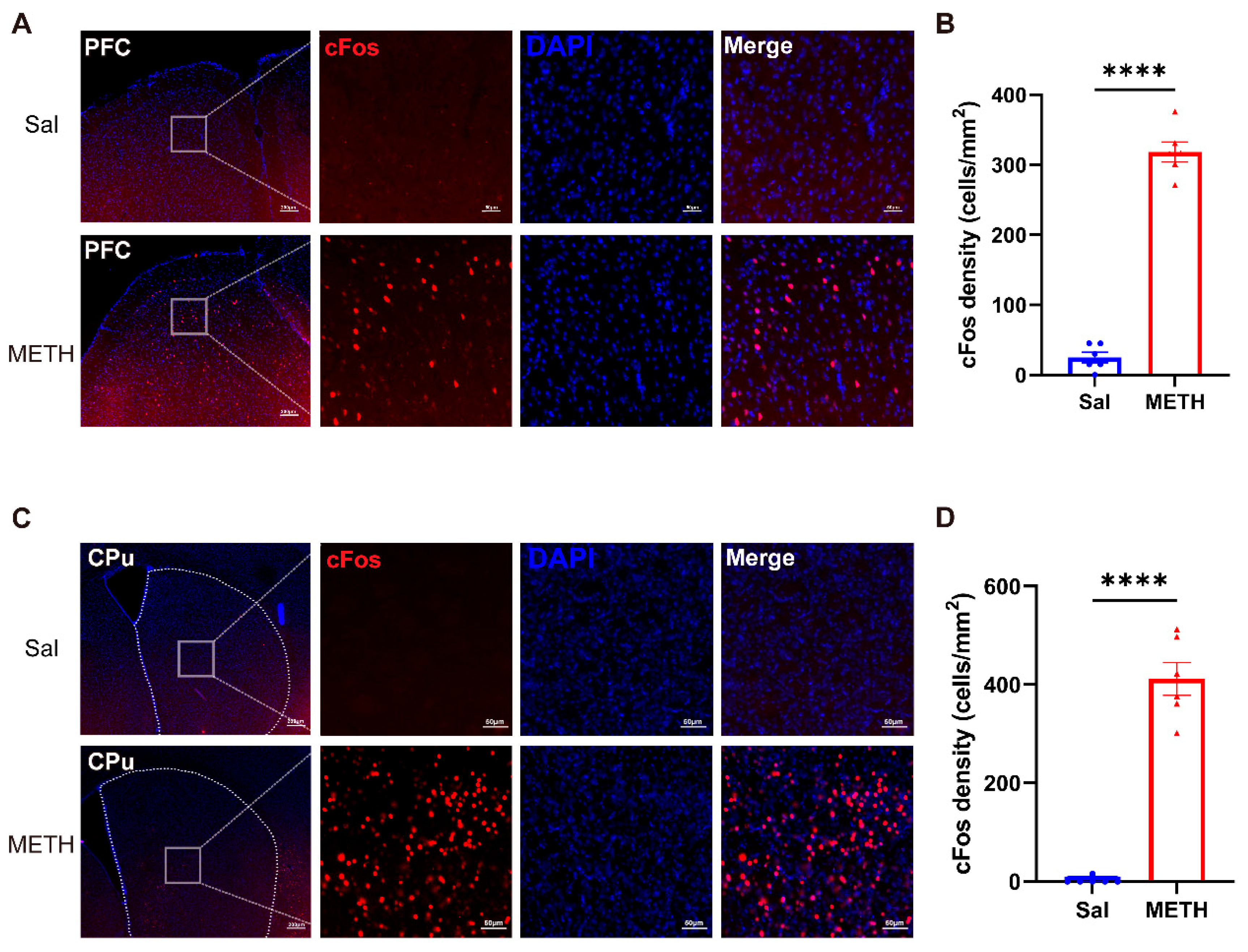
Figure 3.
METH withdrawal downregulated p-AKT and HO-1 expression in the PFC and CPu. (A-B) Protein levels of p-AKT(Thr308), total-AKT (t-AKT), HO-1, and β-actin in the PFC. (C-D) Protein levels of p-AKT(Thr308), t-AKT, HO-1, and β-actin in the CPu. Data are presented as the mean ± SEM; n = 6/group, ** P < 0.01, *** P < 0.001, **** P < 0.0001, compared to the Sal group. Sal, saline; METH, methamphetamine; PFC, prefrontal cortex; CPu, caudate putamen.
Figure 3.
METH withdrawal downregulated p-AKT and HO-1 expression in the PFC and CPu. (A-B) Protein levels of p-AKT(Thr308), total-AKT (t-AKT), HO-1, and β-actin in the PFC. (C-D) Protein levels of p-AKT(Thr308), t-AKT, HO-1, and β-actin in the CPu. Data are presented as the mean ± SEM; n = 6/group, ** P < 0.01, *** P < 0.001, **** P < 0.0001, compared to the Sal group. Sal, saline; METH, methamphetamine; PFC, prefrontal cortex; CPu, caudate putamen.
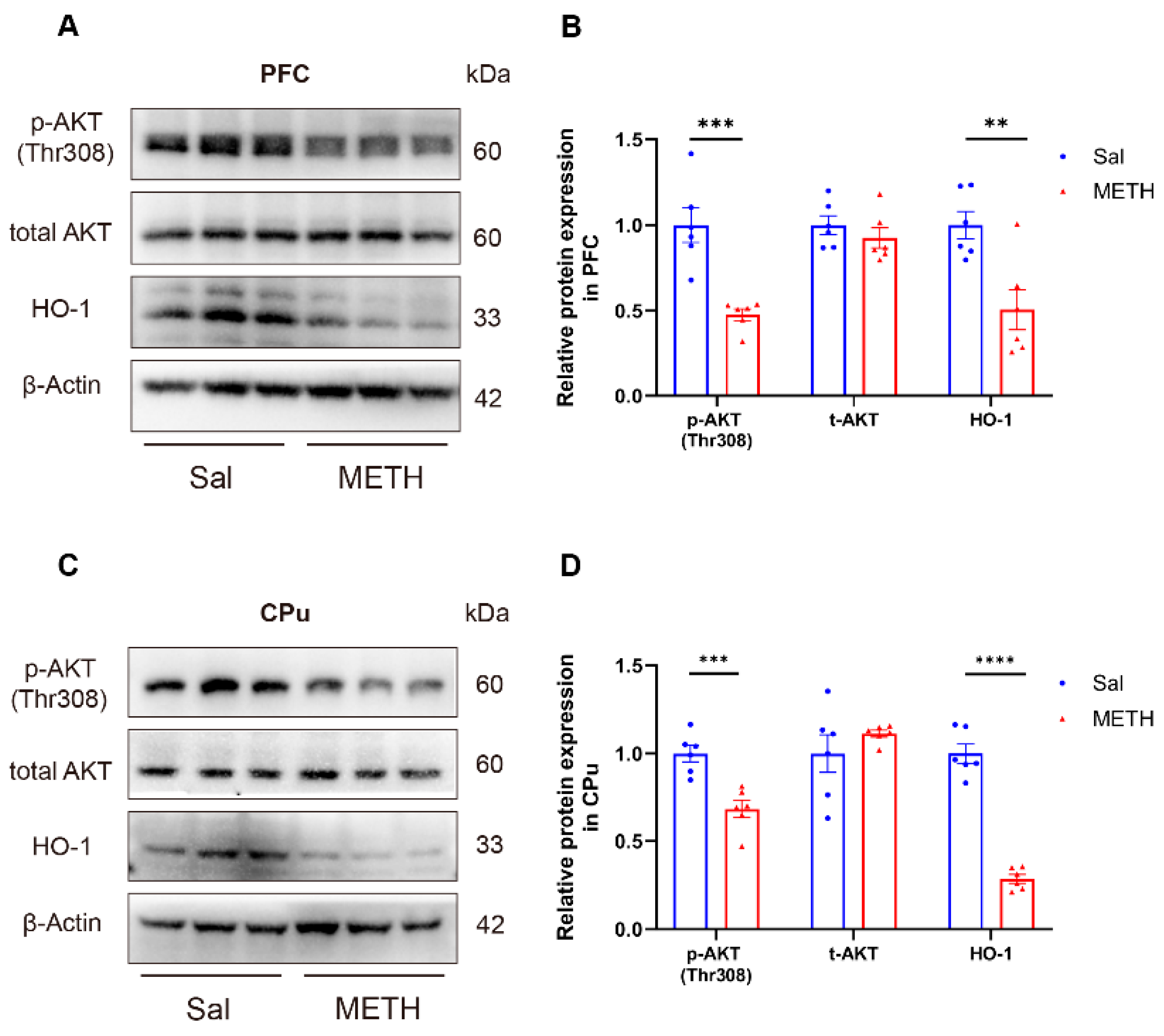
Figure 4.
Luteolin pretreatment alleviated METH withdrawal-induced negative emotions and cognitive deficits. (A) Experimental procedure. (B-C) Anxiety-like behaviors. (B) Time spent and number of entries in the open arm (%) in the EPM test. (C) Time spent in the central zone (%) and total distance traveled in the OFT. (D-E) Cognition function test. (D) Time spent accessing the new object (%) in the NORT. (E) Spontaneous alternations (%) in the Y-maze test. (F-G) Depressive-like behaviors. (F) Sucrose consumption (%) in the sucrose preference test. (G) Immobility time in the FST. Data are expressed as the mean ± SEM; n = 8/group. * P < 0.05, ** P < 0.01, **** P < 0.0001, compared to the Sal group; ## P < 0.01 compared to the Veh + METH group. Sal, saline; Veh, vehicle; Lut, luteolin; METH, methamphetamine; EPM, elevated plus maze; OFT, open field test; NORT, novel object recognition test; SPT, sucrose preference test; FST, forced swim test.
Figure 4.
Luteolin pretreatment alleviated METH withdrawal-induced negative emotions and cognitive deficits. (A) Experimental procedure. (B-C) Anxiety-like behaviors. (B) Time spent and number of entries in the open arm (%) in the EPM test. (C) Time spent in the central zone (%) and total distance traveled in the OFT. (D-E) Cognition function test. (D) Time spent accessing the new object (%) in the NORT. (E) Spontaneous alternations (%) in the Y-maze test. (F-G) Depressive-like behaviors. (F) Sucrose consumption (%) in the sucrose preference test. (G) Immobility time in the FST. Data are expressed as the mean ± SEM; n = 8/group. * P < 0.05, ** P < 0.01, **** P < 0.0001, compared to the Sal group; ## P < 0.01 compared to the Veh + METH group. Sal, saline; Veh, vehicle; Lut, luteolin; METH, methamphetamine; EPM, elevated plus maze; OFT, open field test; NORT, novel object recognition test; SPT, sucrose preference test; FST, forced swim test.
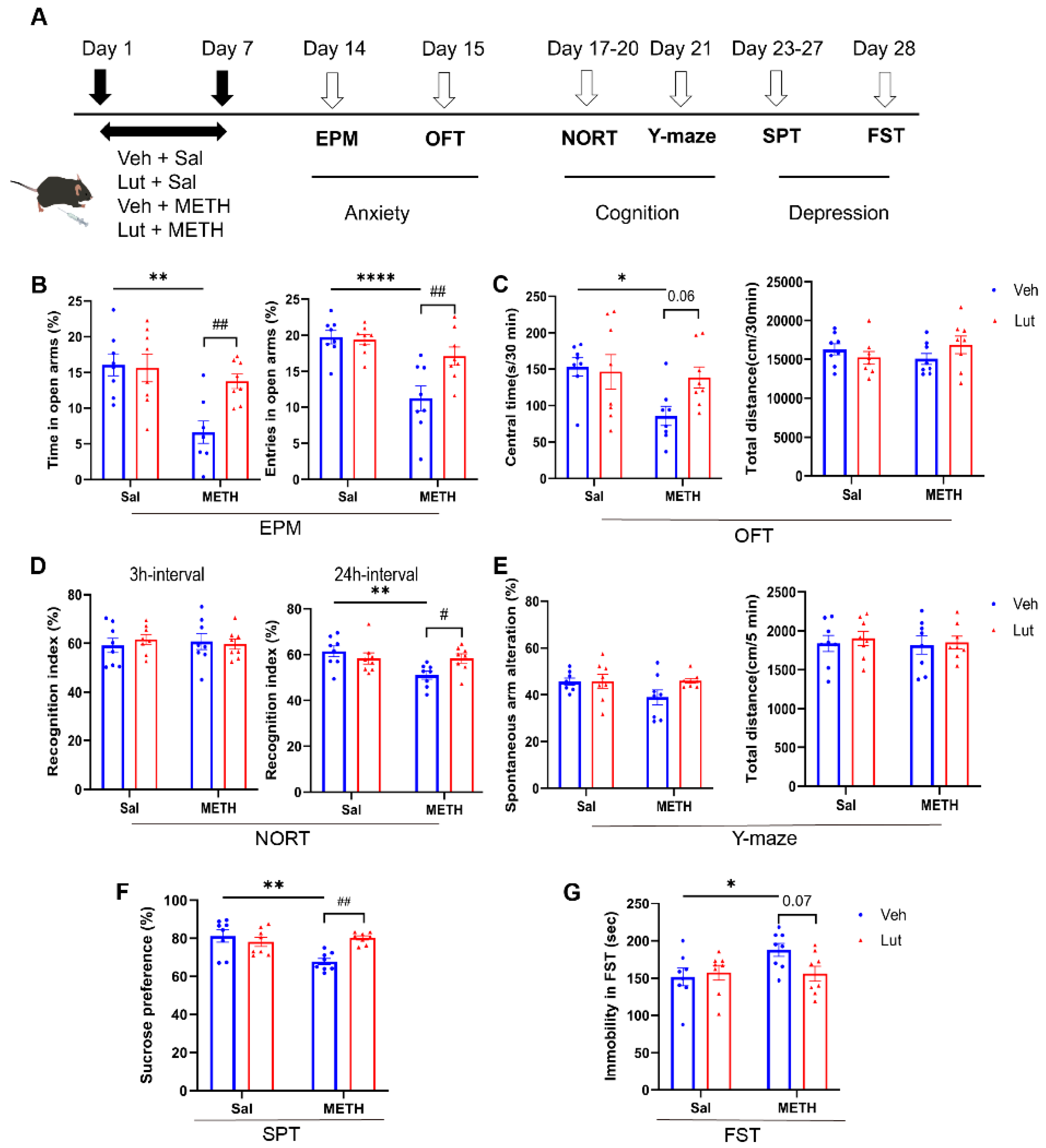
Figure 5.
Luteolin pretreatment downregulated METH withdrawal-induced activation of c-Fos expression in the PFC and CPu. (A-B) Representative images of c-Fos staining and quantitative analysis of the density of c-Fos positive cells in the PFC. (C-D) Representative images of c-Fos staining and quantitative analysis of the density of c-Fos positive cells in the CPu. (A and C) Scale bars represent 200 μm under low magnification and 50 μm under high magnification. Data are presented as the mean ± SEM; n = 3/group, 2 slices per mouse. **** P < 0.0001, compared to the Veh + Sal group; #### P <0.0001 compared to the Lut + METH group. Sal, saline; METH, methamphetamine; Veh, vehicle; Lut, luteolin; PFC, prefrontal cortex; CPu, caudate putamen.
Figure 5.
Luteolin pretreatment downregulated METH withdrawal-induced activation of c-Fos expression in the PFC and CPu. (A-B) Representative images of c-Fos staining and quantitative analysis of the density of c-Fos positive cells in the PFC. (C-D) Representative images of c-Fos staining and quantitative analysis of the density of c-Fos positive cells in the CPu. (A and C) Scale bars represent 200 μm under low magnification and 50 μm under high magnification. Data are presented as the mean ± SEM; n = 3/group, 2 slices per mouse. **** P < 0.0001, compared to the Veh + Sal group; #### P <0.0001 compared to the Lut + METH group. Sal, saline; METH, methamphetamine; Veh, vehicle; Lut, luteolin; PFC, prefrontal cortex; CPu, caudate putamen.
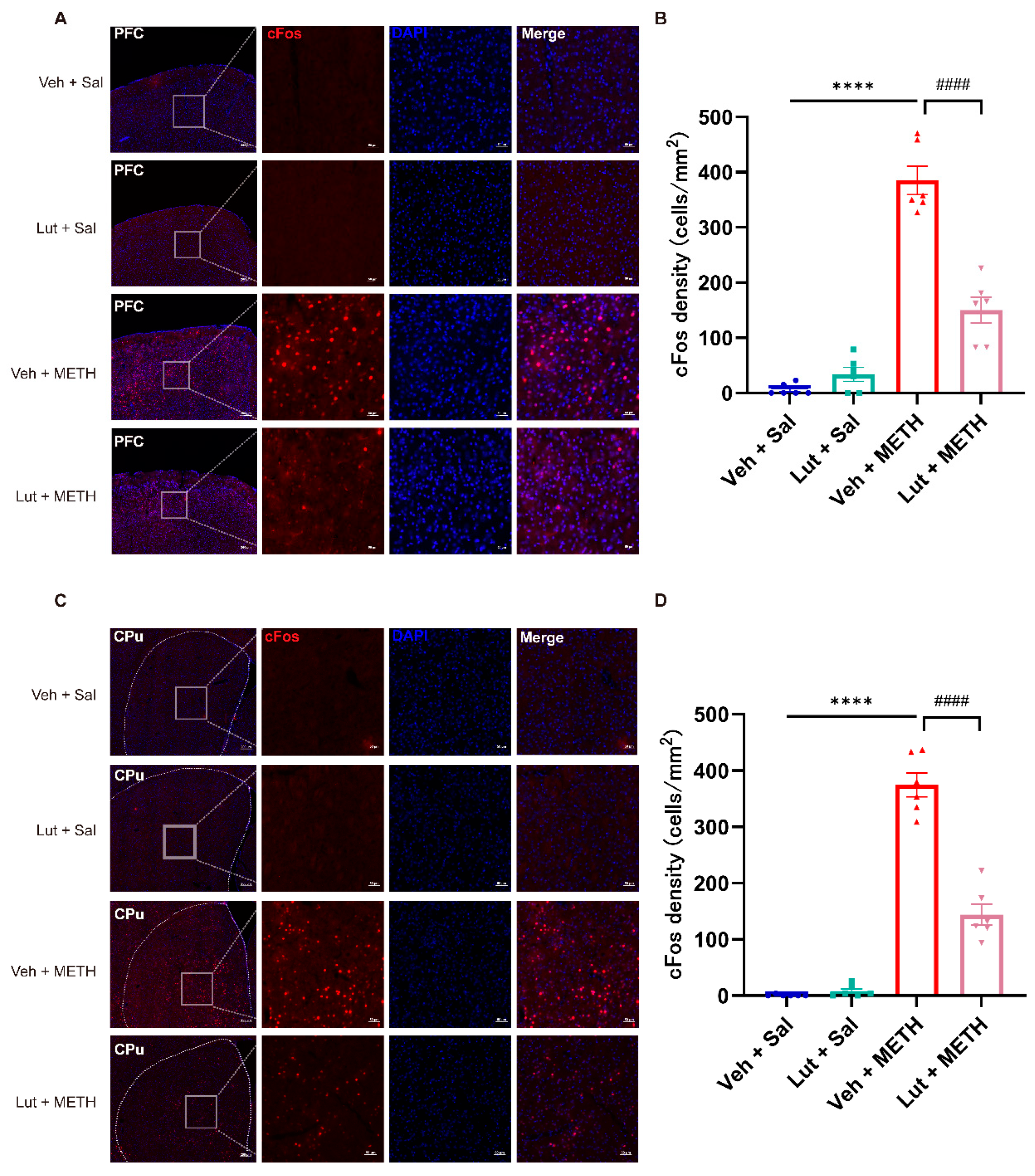
Figure 6.
Luteolin pretreatment reversed METH withdrawal-induced downregulation of p-AKT and HO-1 in the PFC and CPu. (A-B) Protein levels of p-AKT(Thr308), total-AKT (t-AKT), HO-1, and β-actin in the PFC. (C-D) Protein levels of p-AKT(Thr308), t-AKT, HO-1, and β-actin in the CPu. Data are presented as the mean ± SEM; n = 6/group, * P < 0.05, ** P < 0.01, **** P < 0.0001, compared to the Veh + Sal group; # P < 0.05, #### P < 0.0001, compared to the Lut + METH group. Sal, saline; METH, methamphetamine; Veh, vehicle; Lut, luteolin; PFC, prefrontal cortex; CPu, caudate putamen.
Figure 6.
Luteolin pretreatment reversed METH withdrawal-induced downregulation of p-AKT and HO-1 in the PFC and CPu. (A-B) Protein levels of p-AKT(Thr308), total-AKT (t-AKT), HO-1, and β-actin in the PFC. (C-D) Protein levels of p-AKT(Thr308), t-AKT, HO-1, and β-actin in the CPu. Data are presented as the mean ± SEM; n = 6/group, * P < 0.05, ** P < 0.01, **** P < 0.0001, compared to the Veh + Sal group; # P < 0.05, #### P < 0.0001, compared to the Lut + METH group. Sal, saline; METH, methamphetamine; Veh, vehicle; Lut, luteolin; PFC, prefrontal cortex; CPu, caudate putamen.
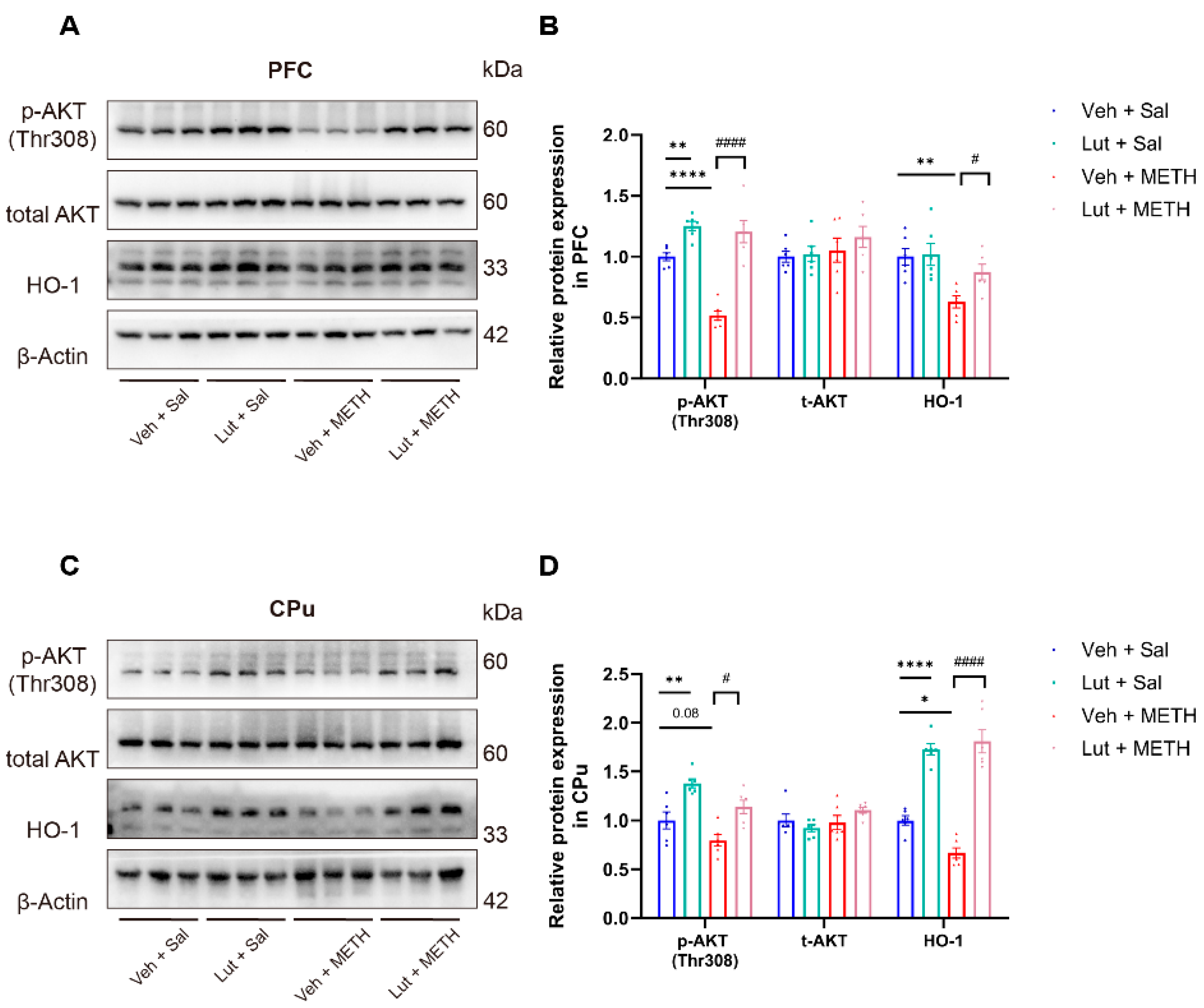
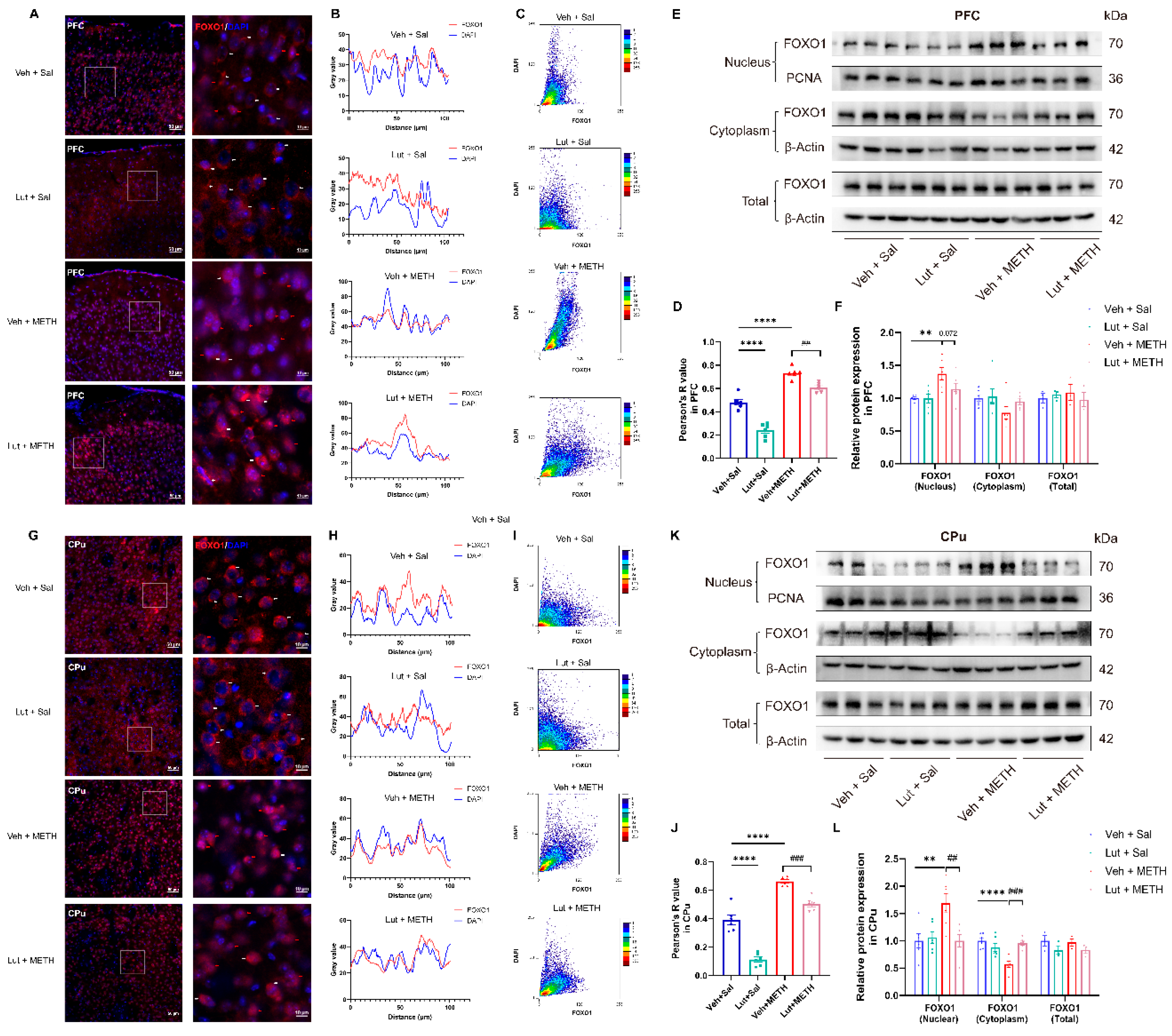
Figure 7.
Luteolin pretreatment prevented METH withdrawal-induced activation of FOXO1 in the PFC and CPu. (A) FOXO1 immunofluorescence in the PFC. (B) Co-localization curve plot of FOXO1 and DAPI in the PFC. (C) Scatter plot showing the fluorescence co-localization of FOXO1 and nucleus in the PFC. (D). Pearson’s correlation for the colocalization of FOXO1 and DAPI in the PFC. (E) Representative immunoblot images of FOXO1 expression in the PFC. (F) Comparisons of relative protein expression level of FOXO1 among groups in the PFC. (G) FOXO1 immunofluorescence in the CPu. (H) Co-localization curve plot of FOXO1 and DAPI in the CPu. (I) Scatter plot showing the fluorescence co-localization of FOXO1 and nucleus in the CPu. (J). Pearson’s correlation for the colocalization of FOXO1 and DAPI in the CPu. (K) Representative immunoblot images of FOXO1 expression in the CPu. (L) Comparisons of relative protein expression level of FOXO1 among groups in the CPu. (A and G) Scale bars represent 50 μm under low magnification and 10 μm under high magnification, n = 3/group, 2 slices per mouse. Red arrows represent nuclear translocation and white arrows represent cytoplasm localization of FOXO1. (F and L) Data are presented as the mean ± SEM, n = 6/group for nucleus/cytoplasm FOXO1; n = 3/group for total FOXO1. ** P < 0.01, **** P < 0.0001 compared to the Veh + Sal group; ## P < 0.01, ### P < 0.001 compared to the Lut + METH group. Sal, saline; METH, methamphetamine; Veh, vehicle; Lut, luteolin; PFC, prefrontal cortex; CPu, caudate putamen.
Figure 7.
Luteolin pretreatment prevented METH withdrawal-induced activation of FOXO1 in the PFC and CPu. (A) FOXO1 immunofluorescence in the PFC. (B) Co-localization curve plot of FOXO1 and DAPI in the PFC. (C) Scatter plot showing the fluorescence co-localization of FOXO1 and nucleus in the PFC. (D). Pearson’s correlation for the colocalization of FOXO1 and DAPI in the PFC. (E) Representative immunoblot images of FOXO1 expression in the PFC. (F) Comparisons of relative protein expression level of FOXO1 among groups in the PFC. (G) FOXO1 immunofluorescence in the CPu. (H) Co-localization curve plot of FOXO1 and DAPI in the CPu. (I) Scatter plot showing the fluorescence co-localization of FOXO1 and nucleus in the CPu. (J). Pearson’s correlation for the colocalization of FOXO1 and DAPI in the CPu. (K) Representative immunoblot images of FOXO1 expression in the CPu. (L) Comparisons of relative protein expression level of FOXO1 among groups in the CPu. (A and G) Scale bars represent 50 μm under low magnification and 10 μm under high magnification, n = 3/group, 2 slices per mouse. Red arrows represent nuclear translocation and white arrows represent cytoplasm localization of FOXO1. (F and L) Data are presented as the mean ± SEM, n = 6/group for nucleus/cytoplasm FOXO1; n = 3/group for total FOXO1. ** P < 0.01, **** P < 0.0001 compared to the Veh + Sal group; ## P < 0.01, ### P < 0.001 compared to the Lut + METH group. Sal, saline; METH, methamphetamine; Veh, vehicle; Lut, luteolin; PFC, prefrontal cortex; CPu, caudate putamen.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



