
Submitted:
10 July 2024
Posted:
12 July 2024
You are already at the latest version
Abstract
The phenotypic manifestation of lead toxicity includes cognitive, learning, and memory decline which is associated with neuronal injuries and death and been a major challenge in public health especially in developing countries. Thus, we explore the prophylactic potentials of luteolin in prefrontal Pb intoxication. Thirty-six (36) male Wistar Rats were used for this research and di-vided into six (6) groups of six animals each and were treated orally as follows: Group 1-Normal saline (0.5ml daily for 14 days); Group 2- PbCl3 (50mg/kg of PbCl3 daily for 14 days); Group 3- luteolin (100mg/kg of LUT daily for 14 days); Group 4- luteolin and PbCl3 (50mg/kg PbCl3 and 100mg/kg LUT daily for 14 days); Group 5- PbCl3 then luteolin (50mg/kg of PbCl3 daily for 14 days followed by 100mg/kg of LUT daily for 14 days); Group 6- luteolin then PbCl3 (100mg/kg of LUT daily for 14days followed by 50mg/kg of PbCl3 daily for 14 days). We measured cerebral oxidative redox parameters, G6PDH, IL-6, acetylcholinesterase levels, and total protein profiles using ELISA and biochemical assay kits. We also examined cerebral morphology and the Nissl profile with H&E and toluidine blue staining techniques, as well as astrocytic morphology and synaptic integrity with anti-GFAP and anti-synaptophysin antibodies. As a result of Pb perturba-tion, luteolin treatment inhibits MDA, IL-6, and AChE expression while increasing antioxidant enzyme activities (SOD, GPx) and G6PDH. Furthermore, luteolin has significant prophylactic po-tential against Pb-induced neuronal injuries and modulates astrocyte actions, preserving synap-tic integrity and thus improving cognition, learning, and memory. Taken together, our findings indicate that luteolin may be a promising candidate for managing and treating Pb neural intoxi-cation.
Keywords:
Neuroanatomy
; lead toxicity
; prophylactic potentials of luteolin
; neurodegeneration
1. Introduction
Lead poisoning, primarily affecting children, is a critical global public health concern, contributing to an estimated 600,000 new cases of mental disorders annually, especially in developing countries [1,2]. Lead (Pb), a heavy metal pervasive in nature, has emerged as a prominent environmental toxin due to its widespread use in industrial and household applications [3]. Exposure to lead during pregnancy, infancy, or early childhood poses a substantial risk, impairing the development of the nervous system. This can lead to cognitive dysfunction, significant reductions in memory and learning abilities, as well as motor and behavioral deficits [4,5].
The mechanisms underpinning lead toxicity are multifaceted and include oxidative stress, alterations in membrane biophysics, disruptions in cellular signaling, and interference with neuro-transmission. Lead triggers the generation of reactive oxygen species (ROS), deactivates biological enzymes by binding to their sulfhydryl groups, and displaces essential ions like calcium and, to a lesser extent, zinc. These events, in turn, trigger calmodulin-dependent processes and inflict damage primarily upon the prefrontal cortex, hippocampus, and cerebellum. These detrimental effects are associated with a range of neurological disorders, starting with fundamental conditions such as mental retardation and neuronal damage, progressing to more complex impairments including nerve dysfunction and behavioral deficits. Additionally, they can resemble conditions seen in more advanced neurodegenerative diseases like Alzheimer's, Parkinson's, and schizophrenia [6,7]. Mitochondrial activity produces ATP, which ensures the proper operation of all cellular activities. This ATP generation process frequently results in the production of free radicals, which are extremely harmful to the body; thus, there is a biological system known as the antioxidative system that is responsible for the removal of free radicals produced during ATP production.
Furthermore, lead hinders neurotransmitter release, causing disruptions in vital systems like GABAergic, dopaminergic, and cholinergic pathways, as well as NMDA-ion channel inhibition, particularly during the neonatal period [8]. Lead has been shown to activate capillary protein kinase C. Protein kinase C (PKC) is a family of enzymes involved in various cellular processes, including signal transduction and the regulation of cell growth and differentiation. When lead activates PKC, it can trigger a cascade of cellular events, potentially leading to harmful effects on cell function and signaling. It also inhibits the activity of the Na/K-ATPase enzyme. Na/K-ATPase is responsible for maintaining the balance of sodium and potassium ions in and out of cells, which is crucial for proper cell function, including nerve cell function. Inhibition of Na/K-ATPase by lead disrupts this ion balance, potentially affecting the normal functioning of cells, including neurons. These actions culminate in the formation of a permeability transition pore, ultimately leading to programmed cell death processes and subsequent mitochondrial self-destruction [9,10]. In summary, lead toxicity triggers apoptosis, excitotoxicity (affecting neurotransmitter storage, release, and modification), and alters key components such as neurotransmitter receptors, mitochondria, second messengers, cerebrovascular endothelial cells, astroglia, and oligodendroglia.
In light of this grave public health issue, there has been growing interest in exploring alternative approaches for mitigating the adverse effects of lead toxicity. Medicinal plants and their bioactive compounds have garnered attention as potential remedies for various diseases [11,12]. Flavones, a class of non-essential nutrients, have enjoyed historical recognition across diverse cultures for their nutraceutical benefits [12]. Luteolin, a natural flavonoid abundant in numerous plants, stands out for its diverse therapeutic properties, including anticancer, antioxidant, wound healing, anti-diabetic, anti-bacterial, anti-inflammatory, and anti-proliferative effects [12,13]. Its unique structural composition, characterized by C6-C3-C6 arrangement, hydroxyl moieties, and 2-3 double bonds, underlies its biochemical and biological activities [14]. Numerous studies have demonstrated luteolin's ability to inhibit lipid peroxidation, downregulate inducible nitric oxide synthase (iNOS), restore and enhance antioxidant enzymes, and modulate inflammatory mediators in animal models of various diseases [14,15,16,17].
In this study, we delve into the structural and morphological alterations as well as molecular changes occurring in the prefrontal cortex due to lead toxicity in Wistar rats. Additionally, we investigated the potential prophylactic effects of luteolin, shedding light on its role in safeguarding against the neurological consequences of lead exposure. This research aims to contribute valuable insights into the therapeutic potential of luteolin in mitigating the detrimental impact of lead toxicity on the central nervous system, offering promise for the development of novel strategies to combat this pressing public health challenge.
2. Materials and Methods
2.1. Animal Procurement and Care
Male Wistar Rats (160-180g of about 6-7 weeks old) were housed in the animal house of the University of Ilorin’s Faculty of Basic Medical Sciences, College of Health Sciences. All animals had free access to drinking water and a pelletized diet and were given 14 days to acclimate prior to the start of the experiment. The protocols followed in this study adhere strictly to the guidelines on animal handling of the institutional Animal Care and Use Committee (IACUC) approved by Ethical Review Committee of College of Health Science, University of Ilorin, Nigeria (NIH Publications No. 8023, revised 1978).
2.2. Reagents Purchase and Preparation of Solutions
Luteolin (MERCK, Sigma-Aldrich, Germany) was prepared in normal saline (100mg/5ml). Crystalline salt of lead chloride PbCl3 (MERCK, Sigma-Aldrich, Germany) was dissolved in normal saline (50mg/5ml). Both solutions were prepared freshly daily and kept at 4ºC before administration.
2.3. Experimental Designs
Thirty-six (36) male Wistar Rats were used for this research and divided into six (6) groups of six animals each and were treated orally as follows: Group 1-Normal saline (0.5ml daily for 14 days); Group 2- PbCl2 (50mg/kg of PbCl2 daily for 14 days); Group 3- luteolin (100mg/kg of LUT daily for 14 days); Group 4- luteolin and PbCl2 (50mg/kg PbCl2 and 100mg/kg LUT daily for 14 days); Group 5- PbCl2 then luteolin (50mg/kg of PbCl2 daily for 14 days followed by 100mg/kg of LUT daily for 14 days); Group 6- luteolin then PbCl2 (100mg/kg of LUT daily for 14days followed by 50mg/kg of PbCl2 daily for 14 days). We assess mitochondrion function by comparing the levels of antioxidative enzymes and malondialdehyde, a byproduct of lipid peroxidation, across experimental groups.
2.4. Tissue Processing
Following the last administration, rats were given 20mg/kgBw of ketamine intra-peritoneally to euthanize them before undergoing transcardial perfusion with a flush of 50 mL of 0.4 M PBS (pH 7.4) followed by 500 mL of 4% paraformaldehyde (PFA). The brain tissue was then excised, rinsed in 0.25 M sucrose three times for 5 minutes each, and post-fixed in 4% PFA for 24 hours before being stored in 30% sucrose at 4 oC until further processing. Rats used in enzymatic studies were sacrificed via cervical dislocation, excised, rinsed in 0.25 M sucrose three times for five minutes each, and placed in 30 percent sucrose at 4 ° C. The cerebral sections were obtained and routinely processed to produce paraffin wax-embedded blocks for histology and antigen retrieval immunohistochemistry. Histological demonstration of cerebral cytoarchitecture and Nissl profiling were performed in paraffin wax-embedded sections stained in hematoxylin and eosin (H&E) and toluidine blue.
2.5. Immunohistochemistry
Serial sections of the cerebrum (15 m) (The brain) were cut from paraffin blocks, and protein cross-linkages were removed in the sections by activating the antigens with 0.1 percent trypsin for 20 minutes at room temperature. Endogenous peroxidase was inhibited with hydrogen peroxide, and nonspecific protein reactions were reduced with 5% bovine serum albumin (BSA). Each slide received 500 μL of diluted primary antibody and was incubated overnight at 4 ° C. Anti-GFAP and anti-Synaptophysin primary antibodies (Cell Signaling Technologies) were diluted at 1:100 in blocking buffer (10% calf serum, 1% BSA, and 0.1 percent Triton X-100 in 0.1 M PBS). After that, the secondary biotinylated antibody was desalted and diluted in PBS (pH 8.0) before being applied to tissue sections. The secondary antibody was incubated for 30 minutes at room temperature in the humidity chamber. The immunogenic reaction was initiated with 3, 3 ′DAB and intensified with a methenamine silver kit (as directed by the manufacturer). The sections were counterstained with hematoxylin and then treated with 1 percent acid alcohol to reduce the intensity of the counterstain. Histology and immunohistochemistry images were captured with an Olympus binocular research microscope (Olympus, New Jersey, USA) linked to an Amscope Camera (5.0 MP).
2.6. Preparation of Tissue Sample for Biochemical Analyses
Cerebellum tissue homogenate was prepared with ice-cold 0.25 M sucrose (Sigma) within automated homogenizer at 4 ° C. The tissue homogenate was centrifuged for 10 min (IEC: CENTRA-GP8R, DJB Labcare, UK) in a microfuge at 12,000 rpm to obtain the supernatant. The supernatants were aspirated into plain labeled glass cuvette placed in ice. SOD, MDA and GPx, G6PDH, TP, AChE, and IL-6 activities were assayed according to the manufacturer’s instruction in the assay kit pack (Abcam).
2.7. Data Analysis
All data obtained from quantitative studies were analyzed using GraphPad Prism® software (Version 5.1) and tested for analysis of variance (ANOVA) with Tukey’s multiple comparisons test. Significance was set at p<0.05.
3. Results
3.1. Luteolin Inhibits Lead-Induced Lipid Peroxidation and Enhances Antioxidative Enzymes
Lead poisoning caused a notable drop in GPx (Figure 1a) and SOD (Figure 1b) levels while elevating MDA (Figure 1c) levels in the brain compared to the control groups (CON), signifying increased oxidative stress and impaired mitochondrial function (p<0.01, p<0.05, p<0.01, respectively). Similarly, when compared to the groups treated with luteolin (LUT), the differences were statistically significant (GPx p<0.005, SOD p<0.005, and MDA p<0.005, respectively). This indicates that lead exposure induced oxidative stress and disrupted normal mitochondrial activity in the brain. Luteolin treatment improves antioxidative enzyme activity and inhibits lipid peroxidation, as evidenced by significant improvements in cerebral GPx, SOD, and a drastic reduction in MDA formation in groups treated with luteolin before, concurrently, and after lead intoxication.
3.2. Luteolin Improves Glucose 6-Phosphate Dehydrogenase Following Lead Intoxication
To investigate glucose metabolism in the pathogenic and intervention groups, we measured the level of G6PDH in the cerebrum (Figure 1d). We discovered that lead intoxication disrupted glucose bioenergetics, as evidenced by a significant decrease in G6PDH expression and activity compared to the CON (p0.005) and LUT (p0.005) groups. The glucose bioenergetics in the groups treated with luteolin concurrently, before and after lead intoxication, were normalized because G6PDH levels were comparable to those in the control groups.
3.3. Luteolin Prevented Pro-Inflammatory Cytokine Activation Following Lead Intoxication
In both normal and pathological conditions, cytokines play important roles in modulating various cellular responses of neurons. Cytokines actively participate in and support long-term plasticity, learning, and memory processes in physiological conditions. In contrast, when cytokines levels are elevated during inflammation, the effects of cytokines are mostly detrimental to memory mechanisms, associated behaviors, and homeostatic plasticity. We measured cerebral IL-6 expression and examined its modulatory patterns during lead intoxication and luteolin interventional treatments. Lead treatment inhibits synaptic plasticity, implying impaired memory and associated behaviors, as evidenced by increased expression of cerebral IL-6 after lead intoxication when compared to the CON and LUT groups (p<0.005). In lead toxicity, luteolin treatment normalized IL-6 activity, improving synaptic plasticity and memory condition in the cerebrum, as IL-6 expression was reduced in the LUTPb, Pb + LUT, and LUT + Pb groups, though this was only significant in the LUT + Pb group (p<0.05).
3.4. Lead Intoxication Increased Acetylcholinesterase (AChE) Activity in the Cerebrum: Treatment with Luteolin Normalizes It
We measured AChE activity in the cerebral homogenates of lead-intoxicated rats and found that AChE was significantly higher after lead toxicity compared to the control groups (CON (p<0.005), LUT (p<0.005)), indicating poor signal conductivity and transmission. However, luteolin treatment before, during, and after lead intoxication reduced and normalized AChE activity to levels comparable to the control groups.
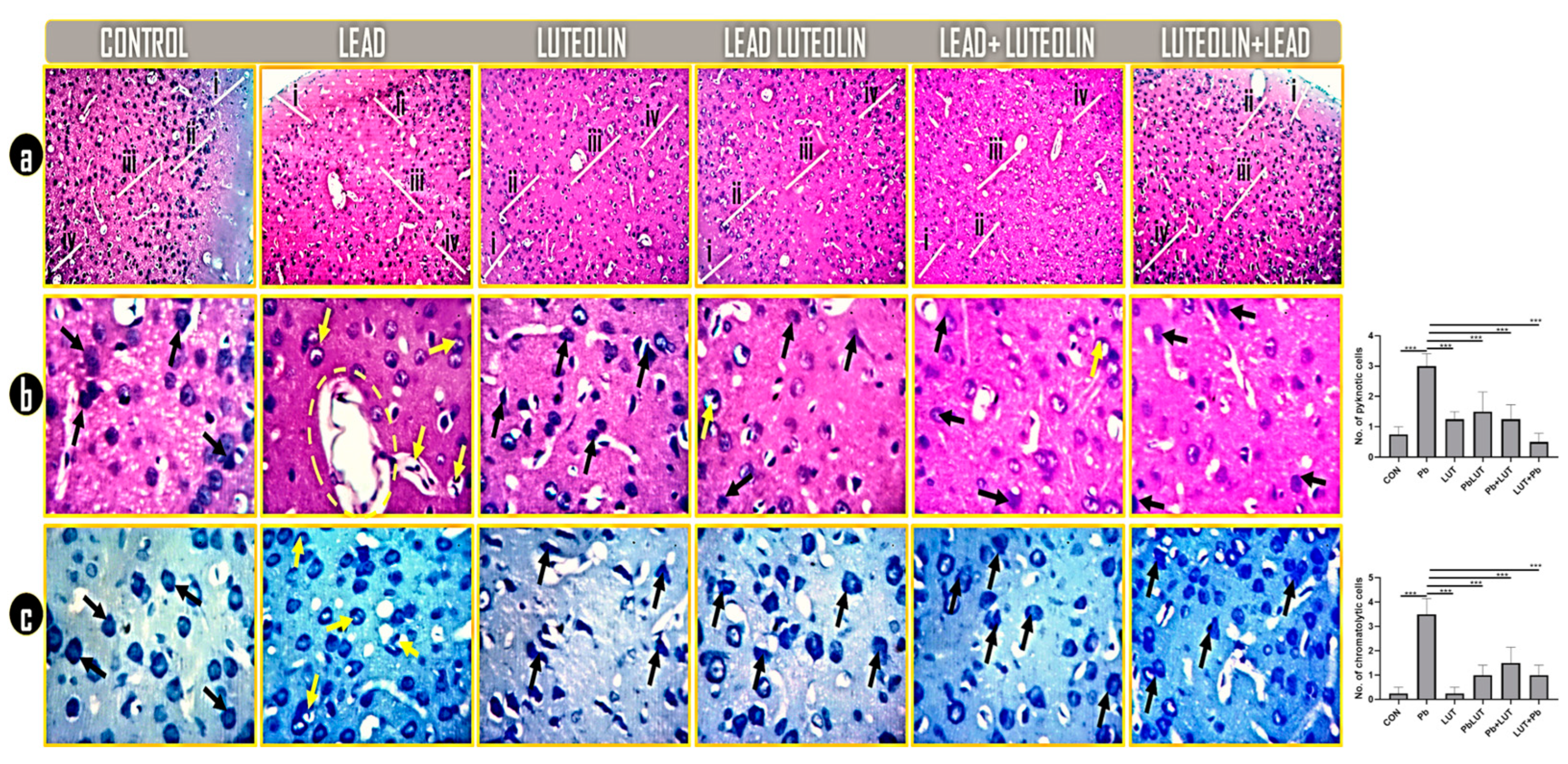
3.5. Luteolin Normalized Histological and Nissl Profiling Changes Caused by Lead Toxicity
Histological alterations and Nissl profiling was investigated with H&E and toluidine blue. histology of the control groups (control and luteolin) shows regular neuronal population, normal cytoarchitecture and distinguished prefrontal layers with regular pyramidal neurons with normal cell body and processes (black arrows). Rats intoxicated with lead shows neurons with degenerative features (pyknosis and necrosis) scattered within the neuropil with distorted cytoarchitecture and fragmented PFC layers. Cytoplasmic inclusions, nuclear materials and neuronal membrane are greatly compromised in this group (yellow arrows and circle). Pyramidal neurons in PFC of rats treated with luteolin concomitantly with (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication was markedly similar to that of the control and are deeply stained (black arrows) with no or few degenerated neurons (yellow arrows) as in case of (PbLUT) and (Pb+LUT) groups (Figure 2a&b). Chromatogenic Nissl substances and regular neuronal density were seen in PFC of the control groups. However, lead intoxication resulted to highly chromatolytic Nissl granules with pyknotic, necrotic and fragmented pyramidal cells having axons that are degenerated (yellow arrows). The chromatogenic nature and cellular density of neurons in the groups treated with luteolin concomitant (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication was marked resemblance (Figure 2c).
Figure 2.
a-c: Prefrontal cortex metabolic, acetylcholinesterase and inflammatory activities determined by glucose-6-phosphate dehydrogenase (G6PDH), acetylcholinesterase (AChE) and interleukin -6 (IL-6). G6PDH expression in the cortical lysate was significantly reduced following lead intoxication in comparison to other treatment groups (fig 2a). G6PDH expression in the cortex was normalized following luteolin treatment with (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication. Lead intoxication resulted to cortical inflammation as evidenced by increase of IL-6 expression in rat cerebral lysate when comparison to other experimental groups. Expectantly, treatment with luteolin downregulates IL-6 expression with (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication thereby inhibiting cortical inflammation (fig 2b). Acetylcholinesterase activities were significantly higher in the lead-treated group, as was cerebellar AChE upregulation, when compared to other treatment groups. AChE activities were normalized by treating rats with (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication, as evidenced by a decrease in the expression of cerebral AChE in these groups (fig 2c). (*=p<0.05, **=p<0.01, and ***=p<0.005).
Figure 2.
a-c: Prefrontal cortex metabolic, acetylcholinesterase and inflammatory activities determined by glucose-6-phosphate dehydrogenase (G6PDH), acetylcholinesterase (AChE) and interleukin -6 (IL-6). G6PDH expression in the cortical lysate was significantly reduced following lead intoxication in comparison to other treatment groups (fig 2a). G6PDH expression in the cortex was normalized following luteolin treatment with (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication. Lead intoxication resulted to cortical inflammation as evidenced by increase of IL-6 expression in rat cerebral lysate when comparison to other experimental groups. Expectantly, treatment with luteolin downregulates IL-6 expression with (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication thereby inhibiting cortical inflammation (fig 2b). Acetylcholinesterase activities were significantly higher in the lead-treated group, as was cerebellar AChE upregulation, when compared to other treatment groups. AChE activities were normalized by treating rats with (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication, as evidenced by a decrease in the expression of cerebral AChE in these groups (fig 2c). (*=p<0.05, **=p<0.01, and ***=p<0.005).
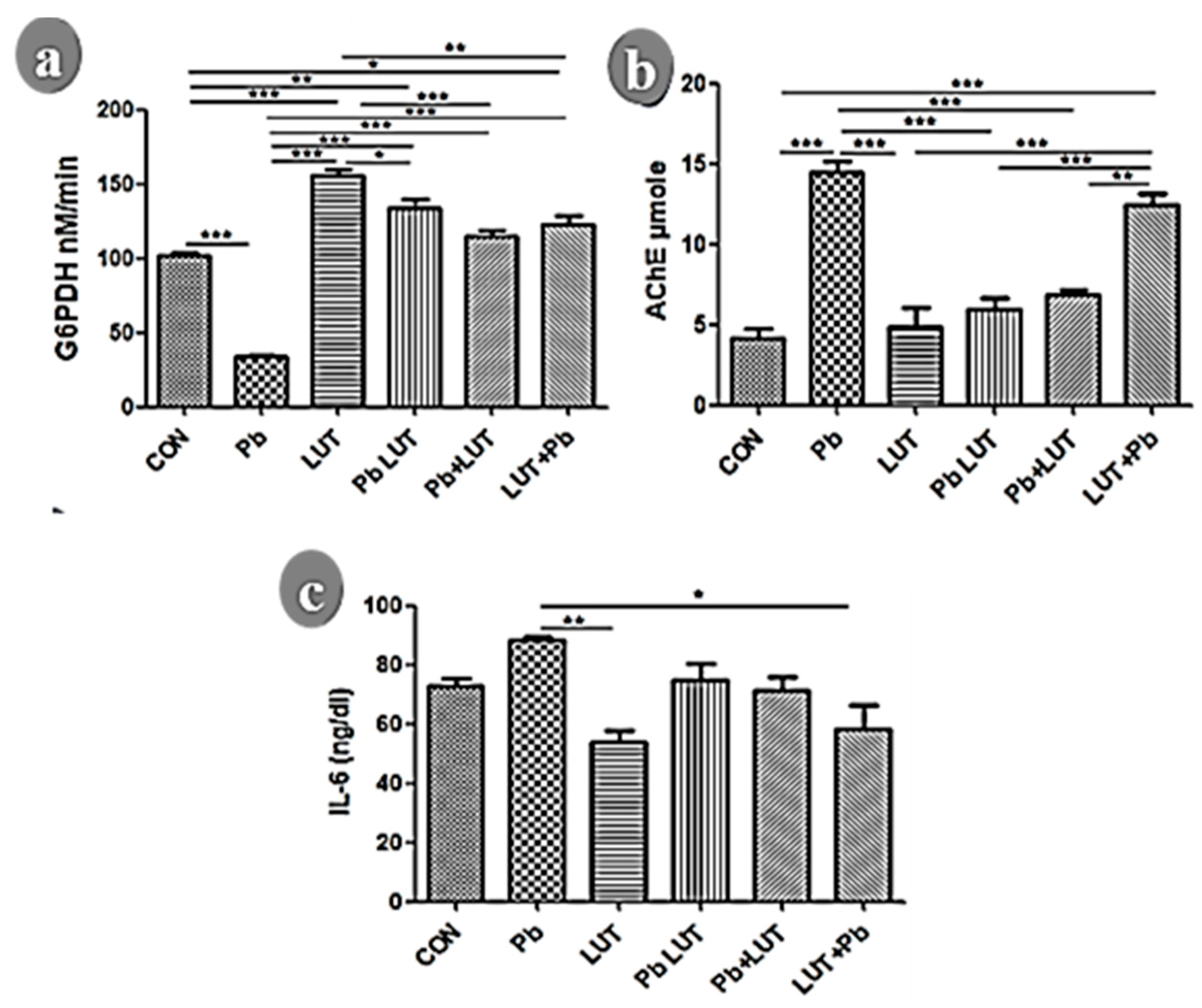
Figure 3.
a-c: Representative photomicrographs revealing the general histomorphology and characteristic Nissl profile of the PFC of treated rats as demonstrated by Hematoxylin and Eosin stain (a and b) and Toluidine blue stain (c). Rats in the control groups (control and luteolin) shows regular neuronal population, normal cytoarchitecture and distinguished prefrontal layers with regular pyramidal neurons with normal cell body and processes (black arrows). Rats intoxicated with lead shows neurons with degenerative features (pyknosis and necrosis) scattered within the neuropil with distorted cytoarchitecture and fragmented PFC layers. Cytoplasmic inclusions, nuclear materials and neuronal membrane are greatly compromised in this group (yellow arrows and circle). Pyramidal neurons in PFC of rats treated with luteolin concomitantly with (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication was markedly similar to that of the control and are deeply stained (black arrows) with no or few degenerated neurons (yellow arrows) as in case of (PbLUT) and (Pb+LUT) groups (fig 2a&b). Chromatogenic Nissl substances and regular neuronal density were seen in PFC of the control groups. However, lead intoxication resulted to highly chromatolytic Nissl granules with pyknotic, necrotic and fragmented pyramidal cells having axons that are degenerated (yellow arrows). The chromatogenic nature and cellular density of neurons in the groups treated with luteolin concomitant (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication was marked resemblance (fig 2c) (40x: 400x).
Figure 3.
a-c: Representative photomicrographs revealing the general histomorphology and characteristic Nissl profile of the PFC of treated rats as demonstrated by Hematoxylin and Eosin stain (a and b) and Toluidine blue stain (c). Rats in the control groups (control and luteolin) shows regular neuronal population, normal cytoarchitecture and distinguished prefrontal layers with regular pyramidal neurons with normal cell body and processes (black arrows). Rats intoxicated with lead shows neurons with degenerative features (pyknosis and necrosis) scattered within the neuropil with distorted cytoarchitecture and fragmented PFC layers. Cytoplasmic inclusions, nuclear materials and neuronal membrane are greatly compromised in this group (yellow arrows and circle). Pyramidal neurons in PFC of rats treated with luteolin concomitantly with (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication was markedly similar to that of the control and are deeply stained (black arrows) with no or few degenerated neurons (yellow arrows) as in case of (PbLUT) and (Pb+LUT) groups (fig 2a&b). Chromatogenic Nissl substances and regular neuronal density were seen in PFC of the control groups. However, lead intoxication resulted to highly chromatolytic Nissl granules with pyknotic, necrotic and fragmented pyramidal cells having axons that are degenerated (yellow arrows). The chromatogenic nature and cellular density of neurons in the groups treated with luteolin concomitant (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication was marked resemblance (fig 2c) (40x: 400x).
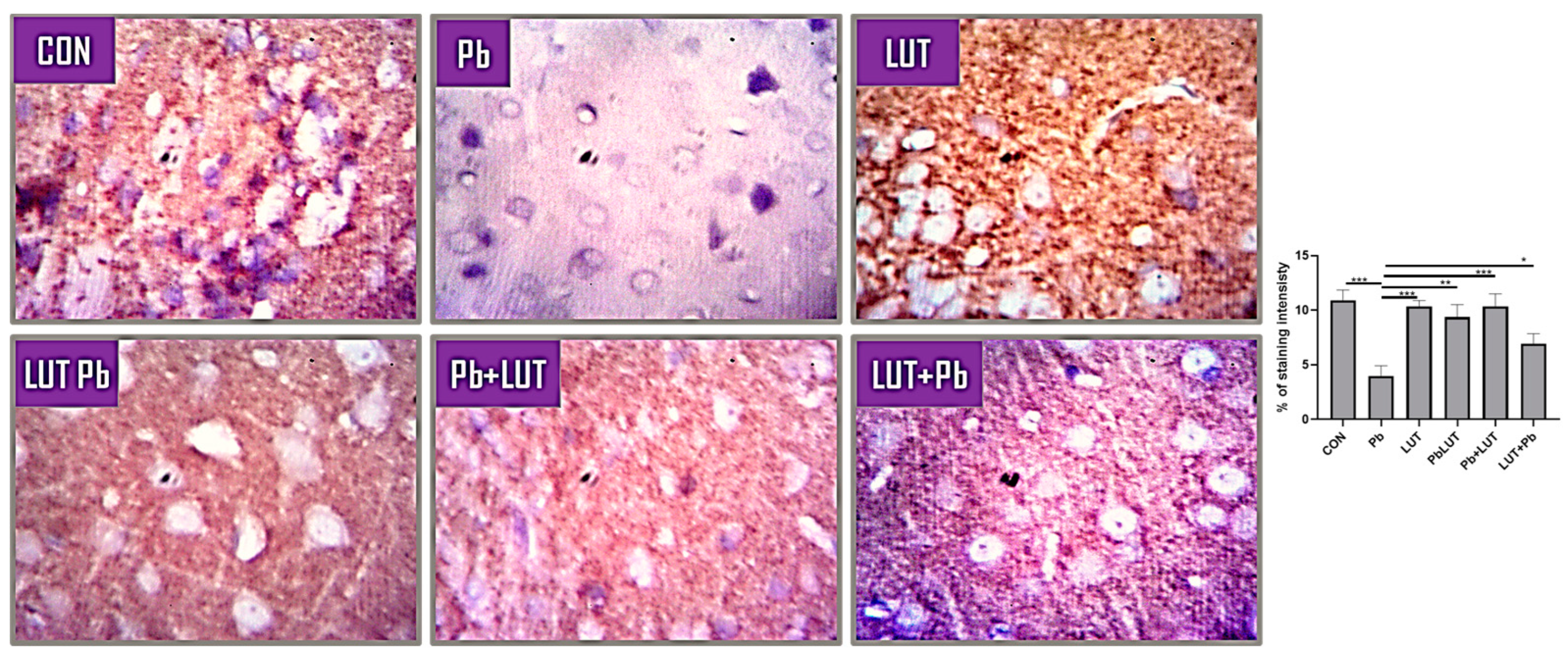
3.6. Lead-Induced Astrogliosis and Luteolin Therapeutic Effects
Astrocytes are key players for neuronal survival, and perturbation of astrocytic functions can be a major cause of neurodegeneration. The process of glial scar formation exemplifies the interaction between the neuroprotective and neurotoxic effects of reactive gliosis. Thus, we investigated astrocytic density, morphology and distribution using GFAP immunostaining. The morphology of astrocytes was altered by lead treatment, resulting in an increase in density with reactive astroglia and the presence of hypertrophic cells within the cerebral, indicating astrogliosis. LUT treatment improves astrocyte density and morphology in rats in the LUT+Pb, Pb+LUT, and PbLUT groups slightly more than in the control groups (Figure 4).
3.7. Luteolin Improved Long Term Plasticity and Cognition in Lead-Induced Synaptic Alteration
Lead intoxication resulted to immuno-negativity to anti-rats synaptophysin which is an indication of loss of synaptic functioning and communication. Treatment using luteolin during, before and after lead intoxication was effective in normalizing the synaptic integrity as there was immune reactivity to synaptophysin in this group that is comparable to that of the control groups (Figure 5).
4. Discussion
Physiological equilibrium is maintained in the mitochondrial metabolism and ATP production by enzymes (superoxide dismutase, catalase, peroxidase) or low molecular weight oxidants (chelating agents, glutathione, NADPH), whereas oxidative stress occurs when there is an imbalance in reactive oxygen (ROS) production and scavenging rate. Uncontrolled production, accumulation of ROS, and decreased antioxidant capacity are major etiologies of several diseases, usually resulting in damaged cell lipids, proteins, nucleic acid, and mitochondrial functions, ultimately leading to apoptotic and necrotic cell death. [18,19,20] Because of their high unsaturated lipid content, brain tissues are particularly vulnerable to free radical attacks, particularly in the axonal layer of the hippocampus and cortical region. [21]
Lead has been reported as a potent prooxidant, known to increase lipid peroxides and inhibit antioxidant enzyme activity via calcium replacement, as well as neurotransmitter production inhibition and disruption of their activities in a different region of the brain, resulting in oxidative damage and excitotoxicity, culminating in neurodegeneration. [6,9] These toxic effects of Pb account for the marked depletion in SOD and GPx levels and an increase in MDA expression in cerebral lysate in our current findings. Lead intoxication causes free radical damage via two distinct, albeit related, pathways: (1) the generation of reactive oxygen species (ROS), and (2) the direct depletion of antioxidant reserves, which it achieves by inhibiting mitochondrial function, resulting in a massive influx of Ca2+ into the mitochondria, thereby improving mitochondrial electron transport, which favors the generation of reactive oxygen species (ROS), and inactivating antioxidant enzymes by binding to their sulfhydryl complex and displacing zinc (Zn) in the enzyme-coenzyme complexes rendering them ineffective as an antioxidant. [22,23,24] Concurrent, pre-treatment, and post-treatment with luteolin normalized the physiological equilibrium in the cerebral cellular redox, as there was a substantial increase in SOD, GPx, and a drastic reduction in MDA expression in these groups. Various studies shown that luteolin inhibits lipid peroxidation, downregulates inducible nitric oxide synthase (iNOS), restores and improves anti-oxidant enzymes. [14,15,16] The use of flavonoids and other polyphenolic compounds derived from plants is thought to be a promising strategy for preventing neurodegenerative diseases like Alzheimer’s and Parkinson’s. [20] The underlying mechanism involving luteolin actions against lead induced cerebral oxidative damages could be due to its richness as a flavonoid which makes it a potent antioxidant.
Mitochondrial ATP production via oxidative phosphorylation from glucose metabolism is critical for proper brain function and is the underlying concept of cellular homeostasis. Almost all neurodegenerative pathologies are caused by a disruption in either central or peripheral glucose bioenergetics, which affects almost every level of cellular or metabolic cascades. Although both G6PDH and LDH are involved in energy production, they play very different roles. G6PDH generates NADPH, which is used as a reducing agent in many metabolic pathways. For example, it is used to reduce glutathione, an important antioxidant. In contrast, LDH catalyzes the interconversion of pyruvate and lactate, a reaction that is involved in energy production through glycolysis. G6PDH and LDH work together to ensure that the cell has the necessary cofactors and intermediates for energy production. This may be affected by oxidative modification, and decreased activity may contribute to the changes in glucose metabolism seen in some nervous disorders. [25] Thus we checked the level of cerebral G6PDH to check the preventive, protective and therapeutic potentials of luteolin in lead induced perturbed glucose bioenergetics pathway. We discovered that lead intoxication disrupted glucose bioenergetics, as evidenced by a significant decrease in G6PDH expression and activity compared to the control groups. Experiments have shown that lead activates protein kinase C in capillary cells while inhibiting Na+/K+-ATPase, interfering with energy metabolism which support our findings. [9] G6PD is a pentose shunt housekeeping enzyme that catalyzes the oxidation of glucose-6-phosphate to 6-phosphogluconolactone, which reduces NADP+ to NADPH. NADPH is a pivotal molecule that protects cells from oxidative-induced damage by modulating GSH activities. Under normal conditions, disulfide bridges (-S-S-) formed by oxidation of sulfhydryl (SH) groups are reduced by GSH to SH-groups in the presence of glutathione peroxidase, which is then oxidized to GSSG and restored to GSH. Because G6PD maintains the level of NADPH, its disruption has been linked to a variety of diseases, including G6PDH-deficient RBCs, liver diseases, gastrointestinal diseases, and neurodegenerative diseases. [25,26] The aforementioned mechanisms might be the underlying mechanism by which lead induces oxidative stress and lipid peroxidation and subsequent glucose metabolic disruption and ATP production in this study. The glucose bioenergetics in the groups treated with luteolin concurrently, before and after lead intoxication, were normalized because G6PDH levels were comparable to those in the control groups. Luteolin has been shown in both in vivo and in vitro studies to have a potent effect on the mechanistic pathways involved in the inhibition of ATP production during cellular toxicity. [20,27] The aforementioned studies support Luteolin’s ability to restore and improve glucose bioenergetics following lead intoxication, as reported in this study. The underlying mechanisms by which luteolin inhibits protein kinase C thereby normalizing Na+/K+-ATPase and modulate G6PDH activity and thus NADPH.
Cytokines are bioactive proteins that immune cells produce in response to pathogens or injury and are abundantly expressed by activated glial and neuronal cells in various brain regions, playing pivotal roles in a variety of normal and pathological processes such as sleep regulation, neuronal development, blood-brain barrier alteration, neurotransmitter metabolism modulation, and synaptic plasticity. [28,29] Due to their involvement in the inflammatory process in patients’ brains with different neurodegenerative diseases they have become vital in understanding the pathogenesis as well as finding potent therapy for these diseases. We investigated Luteolin’s modulatory role in Pb induced cerebral inflammatory process by measuring the level of IL-6. When compared to the control groups, lead treatment increased the expression of cerebral IL-6, indicating that inflammation is one of its pathogenesis mechanisms. IL-6 is a potent pro-inflammatory cytokine that stimulates prostaglandin synthesis, neutrophil influx, and T and B lymphocyte activation and proliferation. Several experiments demonstrated that Pb exacerbated the occurrence of inflammation by elevating various types of pro-inflammatory cytokines, [28] which supports the upregulation of cerebral IL-6 following lead intoxication in this study. Lead underlying mechanisms may involve modulating the MAPK and nuclear factor kappa-B (NF-B) pathways, which involve activator protein-1 (AP-1) and IKK activity, respectively, initiating the upregulation of transcription factors that produce pro-inflammatory cytokines, as well as activating the PI3K-AKT pathway, which further exacerbates the inflammatory process during lead intoxication. [28] Luteolin treatment normalized cerebrum inflammatory response during lead intoxication as shown by marked decrease of cerebrum IL-6 comparable to the control groups. The results of this study collaborate previous reports. The underlying mechanisms of luteolin’s anti-inflammatory actions are its ability to suppress AP-1 and NF-kappa B activation, IkB degradation, and COX-2 expression as a result of its antioxidant and anti-inflammatory properties due to its flavone content. [30,31,32]
Cerebral AChE levels were measured in Pb neurotoxicity and the modulatory effects of luteolin was investigated because the enzyme is involved in cognition, learning and memory. [33] Cholinergic neurons, which are positive markers for memory evolution and development, as well as cholinergic-related disorders that release acetylcholine, a key factor in neuro-cognition, have been linked to a variety of neurodegenerative diseases. [34,35] Previous research has shown that Pb cholinotoxic effects cause a functional change in cholinergic and glutamatergic neurotransmission, resulting in low IQ and impaired cognition without binding to free thiol groups, [35,36] which adds to the current study’s findings. Luteolin treatment, on the other hand, reduced and normalized AChE activity to levels comparable to the control groups. AChE is a key enzyme in the CNS, and inhibiting it raises neural acetylcholine levels, which is one of the therapies for symptomatic relief of mild to moderate Alzheimer’s disease and an important aspect for Alzheimer’s drug development. There have been several reports of it being inhibited after being treated with luteolin. [37,38] Increased ROS and PKC activity have been linked to disruption of AChE synthesis, [39] highlighting the potential underlying mechanisms of luteolin in this study.
Histological investigation shows that Pb intoxication resulted to pyramidal neurons with degenerative features (pyknosis and necrosis) scattered within the neuropil with distorted cytoarchitecture, high cellular density and fragmented PFC layers. Luteolin’s ability to inhibit Pb cytotoxicity may be a key component of its therapeutic potentials. Furthermore, Nissl integrity in pyramidal neurons with the prefrontal cortex of Pb intoxicated rats show marked chromatolytic changes in comparison to the control group, depicting intercellular protein synthesis inhibition and perturbed endoplasmic activity. Nissl granules are responsible for the synthesis of receptor proteins and polypeptides and can change due to pathological conditions, resulting in their disintegration, a condition known as chromatolysis, which can be triggered by axotomy, ischemia, cell toxicity, and exhaustion, or virus infections, ultimately leading to cell death and neurodegeneration. [40] Present finding collaborates with previous reports that showed the toxic effect of Pb on Nissl bodies in neurons. [28,41] Pb disrupt DNA and proteins activities resulting to ER stress and protein macromolecules insult which may be responsible for the chromatolytic changes and cellular decline observed in this current study.
Astrocytes are a critical component of the blood-brain barrier (BBB) that transports neurotrophic factors to neurons. It is important in modulating neuronal immune response, potential excitotoxicity, and synaptic plasticity, all of which improve cognition, learning, and memory. [42,43,44,45] Pb has been demonstrated to be toxic to oligodendrocytes and astrocytes. Pb is thought to disrupt the main structural components of the BBB by primarily injuring astrocytes, followed by endothelial microvasculature damage, [44] thereby activating cascades involved in inflammation, excitotoxicity, synaptic degradation, which reduces cognition, learning, and memory, and neuronal apoptosis ultimately. Following lead intoxication, we observe morphology changes in astrocytes, including an increase in density with reactive astroglia and the presence of hypertrophic cells within the cerebral, indicating astrogliosis. In this study, the toxic effect of Pb on astrocytes is considered to be responsible for synaptic degradation and loss of signal transduction, resulting in a decline in cognition, learning, and memory. The ability of luteolin to prevent astrogliosis and normalize astrocyte morphology is an important aspect of its ability to stop synaptic degradation and preserve and improve cerebral synapses, which reflects improved cognition, learning, and memory. An interaction between increased tissue ROS and astrogliosis has been described. Furthermore, IL-6 and glutamate have been shown to modulate astrocyte activities that rely on high energy consumption, highlighting a possible four-fold mechanism of luteolin’s actions in this study.
Contribution to Knowledge
The findings presented in this study hold significant importance for the scientific community and broader society by shedding light on several critical aspects related to lead toxicity and the potential therapeutic role of luteolin:
Understanding Lead Toxicity Mechanisms: this research provides a comprehensive overview of the mechanisms underlying lead toxicity, emphasizing its role as a prooxidant that disrupts cellular equilibrium. It highlights how lead-induced oxidative stress, lipid peroxidation, and disruption of antioxidant enzymes contribute to neurodegenerative processes. This understanding is fundamental for addressing lead poisoning-related health issues.
The study also underscores the potential of luteolin, a natural flavonoid found in plants, as a therapeutic agent against lead-induced neurotoxicity. Luteolin's ability to restore antioxidant enzyme levels, mitigate lipid peroxidation, and reduce inflammation demonstrates its promise as a neuroprotective agent.
It helped to study the disruption of glucose bioenergetics and ATP production in the context of lead toxicity, highlighting the crucial role of G6PDH in cellular energy metabolism. The normalization of G6PDH levels by luteolin suggests its potential in preserving proper brain function.
The investigation into AChE levels reveals the potential cognitive benefits of luteolin. Since cognitive decline is a hallmark of neurodegenerative diseases, luteolin's ability to normalize AChE activity has implications for conditions such as Alzheimer's disease.
The study provides valuable histological and cellular insights into the impact of lead on neuronal integrity, endoplasmic reticulum function, and astrocyte morphology. Understanding these cellular changes can help identify potential targets for therapeutic interventions.
Our study elucidates the role of pro-inflammatory cytokines, particularly IL-6, in lead-induced neuroinflammation. Luteolin's ability to modulate cytokine levels highlights its anti-inflammatory properties and potential in mitigating neuroinflammatory responses.
The study contributes to the growing body of evidence supporting the use of natural compounds like luteolin in the prevention and treatment of neurodegenerative diseases. This aligns with the broader scientific interest in exploring alternative, plant-derived therapies for various health conditions. Lastly, this research not only advances our understanding of the intricate mechanisms involved in lead toxicity but also offers a promising avenue for the development of interventions to mitigate its detrimental effects on the nervous system. The potential therapeutic role of luteolin as demonstrated in this study opens up exciting possibilities for future research and the development of novel treatments for neurodegenerative diseases and lead poisoning.
In conclusion, the findings of this study support and elicit the notion that lead (Pb) causes neurotoxicity and cerebral degradation through oxidative stress, disruption of AChE, IL-6, and G6PDH activity, as evidenced by excessive free radical production, depletion of the endogenous antioxidant system and G6PDH activity, and upregulation of AChE and IL-6, all of which result in astrogliosis, synaptic loss, and subsequent neuronal death. Intriguingly, luteolin exhibits a desirable and potent ability in mitigating the neurotoxic effects of Pb and is thus recommended as a potential candidate for Pb poisoning treatments.
Author Contributions
Conceptualization, Leviticus Oghenevurinrin Arietarhire; Data curation, Aisha Iyabo Ayuba, Olushola Oladapo Oluyomi and Emmanuel Oliseh Ikechuku; Formal analysis, Keji- Salmat Temilola Keji-Taofik and Joseph Igbo Enya; Funding acquisition, Leviticus Oghenevurinrin Arietarhire, Keji- Salmat Temilola Keji-Taofik and Emmanuel Oliseh Ikechuku; Investigation, Aisha Iyabo Ayuba and Evi Ndudi Imonikosaye; Methodology, Leviticus Oghenevurinrin Arietarhire; Project administration, Leviticus Oghenevurinrin Arietarhire, Aisha Iyabo Ayuba and Evi Ndudi Imonikosaye; Resources, Keji- Salmat Temilola Keji-Taofik, Joseph Igbo Enya, Olushola Oladapo Oluyomi and Olanrewaju John Afees; Software, Joseph Igbo Enya; Supervision, Leviticus Oghenevurinrin Arietarhire and Olanrewaju John Afees; Validation, Aisha Iyabo Ayuba and Ukwubile Ileje Inelo; Visualization, Joseph Igbo Enya and Emmanuel Oliseh Ikechuku; Writing – original draft, Leviticus Oghenevurinrin Arietarhire; Writing – review & editing, Keji- Salmat Temilola Keji-Taofik, Joseph Igbo Enya, Olushola Oladapo Oluyomi, Ukwubile Ileje Inelo and Olugbogi Ezekiel.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Naranjo, V.I.; Hendricks, M.; Jones, K.S. Lead toxicity in children: An unremitting public health problem. Pediatric Neurology 2020, 113, 51–55. [Google Scholar] [CrossRef]
- Miles-Novelo, A.; Anderson, C.A. Climate Change and Human Behavior: Impacts of a Rapidly Changing Climate on Human Aggression and Violence. Elements in Applied Social Psychology 2022. [Google Scholar]
- Nejad, A.R.; Hatamian, M.; Kafi, M.; Souri, M.K.; Shahbazi, K. Interaction of lead and nitrate on growth characteristics of ornamental judas tree (Cercis siliquastrum L.). Open Agriculture 2018, 3, 670–677. [Google Scholar] [CrossRef]
- Olness, K. Effects on brain development leading to cognitive impairment: A worldwide epidemic. Journal of Developmental & Behavioral Pediatrics 2003, 24, 120–130. [Google Scholar]
- Masindi, V.; Muedi, K.L. Environmental contamination by heavy metals. Heavy metals 2018, 10, 115–132. [Google Scholar]
- Khan, D.A.; Qayyum, S.; Saleem, S.; Khan, F.A. Lead-induced oxidative stress adversely affects health of the occupational workers. Toxicology and Industrial Health 2008, 24, 611–618. [Google Scholar] [CrossRef]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS sources in physiological and pathological conditions. Oxidative medicine and cellular longevity 2016, 2016. [Google Scholar] [CrossRef]
- Sanders, T.; Liu, Y.; Buchner, V.; Tchounwou, P.B. Neurotoxic effects and biomarkers of lead exposure: A review. Reviews on environmental health 2009, 24, 15–46. [Google Scholar] [CrossRef]
- Mason, L.H.; Harp, J.P.; Han, D.Y. Pb neurotoxicity: Neuropsychological effects of lead toxicity. BioMed research international 2014, 2014. [Google Scholar] [CrossRef]
- Shabani, S. A mechanistic view on the neurotoxic effects of air pollution on central nervous system: Risk for autism and neurodegenerative diseases. Environmental Science and Pollution Research 2021, 28, 6349–6373. [Google Scholar] [CrossRef]
- Donno, D.; Boggia, R.; Zunin, P.; Cerutti, A.K.; Guido, M.; Mellano, M.G.; Beccaro, G.L.; et al. Phytochemical fingerprint and chemometrics for natural food preparation pattern recognition: An innovative technique in food supplement quality control. Journal of food science and technology 2016, 53, 1071–1083. [Google Scholar] [CrossRef] [PubMed]
- Lodhi, S.; Vadnere, G.P.; Sharma, V.K.; Usman, M.R. Marrubium vulgare L.: A review on phytochemical and pharmacological aspects. J. Intercult. Ethnopharmacol 2017, 6, 429–452. [Google Scholar] [CrossRef]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.M. Luteolin, a flavonoid with potential for cancer prevention and therapy. Current cancer drug targets 2008, 8, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Pandurangan, A.K.; Esa, N.M. Luteolin, a bioflavonoid inhibits colorectal cancer through modulation of multiple signaling pathways: A review. Asian Pacific Journal of Cancer Prevention 2014, 15, 5501–5508. [Google Scholar] [CrossRef] [PubMed]
- Manju, V.; Nalini, N. Protective role of luteolin in 1, 2-dimethylhydrazine induced experimental colon carcinogenesis. Cell Biochemistry and Function: Cellular biochemistry and its modulation by active agents or disease 2007, 25, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Ashokkumar, P.; Sudhandiran, G. Protective role of luteolin on the status of lipid peroxidation and antioxidant defense against azoxymethane-induced experimental colon carcinogenesis. Biomedicine & Pharmacotherapy 2008, 62, 590–597. [Google Scholar]
- Damani, J.J.; De Souza, M.J.; VanEvery, H.L.; Strock, N.C.; Rogers, C.J. The Role of Prunes in Modulating Inflammatory Pathways to Improve Bone Health in Postmenopausal Women. Advances in Nutrition 2022. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Sayre, L.M.; Perry, G.; Smith, M.A. Oxidative stress and neurotoxicity. Chemical research in toxicology 2008, 21, 172–188. [Google Scholar] [CrossRef]
- Wszelaki, N.; Melzig, M.F. (2013). Additive protective effects of luteolin and pyruvate against 6-hydroxydopamine and 3-hydroxykynurenine induced neurotoxicity in SH-SY5Y cells.
- Olajide, O.J.; Fatoye, J.O.; Idowu, O.F.; Ilekoya, D.; Gbadamosi, I.T.; Gbadamosi, M.T.; Asogwa, N.T. Reversal of behavioral decline and neuropathology by a complex vitamin supplement involves modulation of key neurochemical stressors. Environmental Toxicology and Pharmacology 2018, 62, 120–131. [Google Scholar] [CrossRef]
- Lidsky, T.I.; Schneider, J.S. Lead neurotoxicity in children: Basic mechanisms and clinical correlates. Brain 2003, 126, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Patrick, L. Lead Toxicity, a review of the literature. Part I: Exposure, Evaluation, and treatment. Alternative medicine review 2006, 11. [Google Scholar]
- Flora, G.; Gupta, D.; Tiwari, A. Toxicity of lead: A review with recent updates. Interdisciplinary toxicology 2012, 5, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Zhang, D.; Chen, J.; Qin, Y.Y.; Sheng, R.; Feng, X.; Qin, Z.H.; et al. G6PD plays a neuroprotective role in brain ischemia through promoting pentose phosphate pathway. Free Radical Biology and Medicine 2017, 112, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, P.R.; Bates, K.A.; Porter, T.; Teimouri, E.; Perry, G.; Steele, J.W.; Verdile, G.; et al. Latrepirdine: Molecular mechanisms underlying potential therapeutic roles in Alzheimer’s and other neurodegenerative diseases. Translational psychiatry 2013, 3, e332. [Google Scholar] [CrossRef] [PubMed]
- Qiusheng, Z.; Yuntao, Z.; Rongliang, Z.; Dean, G.; Changling, L. Effects of verbascoside and luteolin on oxidative damage in brain of heroin treated mice. Die Pharmazie-An International Journal of Pharmaceutical Sciences 2005, 60, 539–543. [Google Scholar]
- Metryka, E.; Chibowska, K.; Gutowska, I.; Falkowska, A.; Kupnicka, P.; Barczak, K.; Baranowska-Bosiacka, I.; et al. Lead (Pb) exposure enhances expression of factors associated with inflammation. International journal of molecular sciences 2018, 19, 1813. [Google Scholar] [CrossRef] [PubMed]
- Bourgognon, J.M.; Cavanagh, J. The role of cytokines in modulating learning and memory and brain plasticity. Brain and Neuroscience Advances 2020, 4, 2398212820979802. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Li, Q.Z. Prediction of apoptosis protein subcellular location using improved hybrid approach and pseudo-amino acid composition. Journal of Theoretical Biology 2007, 248, 377–381. [Google Scholar] [CrossRef]
- Ziyan, L.; Yongmei, Z.; Nan, Z.; Ning, T.; Baolin, L. Evaluation of the anti-inflammatory activity of luteolin in experimental animal models. Planta medica 2007, 73, 221–226. [Google Scholar] [CrossRef]
- Nabavi, S.F.; Braidy, N.; Gortzi, O.; Sobarzo-Sanchez, E.; Daglia, M.; Skalicka-Woźniak, K.; Nabavi, S.M. Luteolin as an anti-inflammatory and neuroprotective agent: A brief review. Brain research bulletin 2015, 119, 1–11. [Google Scholar] [CrossRef]
- Amjad, S.; Umesalma, S. (2015). Journal of Molecular Biomarkers & Diagnosis.
- Taïr, K.; Kharoubi, O.; Taïr, O.A.; Hellal, N.; Benyettou, I.; Aoues, A. Aluminium-induced acute neurotoxicity in rats: Treatment with aqueous extract of Arthrophytum (Hammada scoparia). Journal of acute disease 2016, 5, 470–482. [Google Scholar] [CrossRef]
- Nwobi, N.L.; Nwobi, J.C.; Akinosun, M.O.; Atulomah, N.O.; Nwazuoke, I.A.; Anetor, J.I. Impaired Antioxidant-Defence Status in Nigerian Children with Elevated Blood Lead Levels: A Possible Predisposing Factor to Chronic Diseases. Journal of Krishna Institute of Medical Sciences (JKIMSU) 2020, 9. [Google Scholar]
- Ademuyiwa OU, R.N.; Ugbaja, R.N.; Rotimi, S.O.; Abam, E.; Okediran, B.S.; Dosumu, O.A.; Onunkwor, B.O. Erythrocyte acetylcholinesterase activity as a surrogate indicator of lead-induced neurotoxicity in occupational lead exposure in Abeokuta, Nigeria. Environmental Toxicology and Pharmacology 2007, 24, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Li, M.; Tian, Y.; Liu, J.; Shang, J. Luteolin inhibits ROS-activated MAPK pathway in myocardial ischemia/reperfusion injury. Life Sciences 2015, 122, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Musa, A.; Shady, N.H.; Ahmed, S.R.; Alnusaire, T.S.; Sayed, A.M.; Alowaiesh, B.F.; Abdelmohsen, U.R.; et al. Antiulcer potential of olea europea l. Cv. arbequina leaf extract supported by metabolic profiling and molecular docking. Antioxidants 2021, 10, 644. [Google Scholar] [CrossRef] [PubMed]
- Monturiol-Gross, L.; Flores-Díaz, M.; Pineda-Padilla, M.J.; Castro-Castro, A.C.; Alape-Giron, A. Clostridium perfringens phospholipase C induced ROS production and cytotoxicity require PKC, MEK1 and NFκB activation. PLoS ONE 2014, 9, e86475. [Google Scholar] [CrossRef] [PubMed]
- Vinters, H.V.; Kleinschmidt-DeMasters, B.K. (2018). General pathology of the central nervous system. In Greenfield’s Neuropathology-Two Volume Set (pp. 25-82). CRC Press.
- Hussein, M.T.; Nemah, M.N. (2015, October). Modeling and control of quadrotor systems. In 2015 3rd RSI International Conference on Robotics and Mechatronics (ICROM) (pp. 725-730). IEEE.
- Sattler, R.; Rothstein, J.D. Regulation and dysregulation of glutamate transporters. Neurotransmitter Transporters 2006, 277–303. [Google Scholar]
- Schousboe, A.; Waagepetersen, H.S. Role of astrocytes in glutamate homeostasis: Implications for excitotoxicity. Neurotoxicity research 2005, 8, 221–225. [Google Scholar] [CrossRef]
- Sanders, T.; Liu, Y.; Buchner, V.; Tchounwou, P.B. Neurotoxic effects and biomarkers of lead exposure: A review. Reviews on environmental health 2009, 24, 15–46. [Google Scholar] [CrossRef]
- Halassa, M.M.; Haydon, P.G. Integrated brain circuits: Astrocytic networks modulate neuronal activity and behavior. Annual review of physiology 2010, 72, 335–355. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
a-c: Prefrontal cortex oxidative redox evaluated by superoxide dismutase (SOD), gluthione peroxidase (GPx), and Malondialdehyde (MDA). The cortical homogenate of rats intoxicated with lead disrupt the activity of the antioxidative system as evidence by significant downregulation of GPx (fig 1a) and SOD (fig 1b) expression when compared across other experimental groups. Treatments with luteolin with (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication normalized the antioxidative system activity in the cortical homogenate as the level of SOD and GPx expression was significantly increase which similar that of the control groups (CON & LUT). Lead intake (Pb) promotes lipid peroxidation in rat cortical lysates as shown by significant increase of MDA level in relation to other experimental groups (fig 1c). Luteolin inhibits lipid peroxidation among groups treated with, before, and after lead intake as the level of cortical MDA was significant decreased similarly to that of the control groups. (*=p<0.05, **=p<0.01, and ***=p<0.005).
Figure 1.
a-c: Prefrontal cortex oxidative redox evaluated by superoxide dismutase (SOD), gluthione peroxidase (GPx), and Malondialdehyde (MDA). The cortical homogenate of rats intoxicated with lead disrupt the activity of the antioxidative system as evidence by significant downregulation of GPx (fig 1a) and SOD (fig 1b) expression when compared across other experimental groups. Treatments with luteolin with (PbLUT), before (LUT+Pb), and after (Pb+LUT) lead intoxication normalized the antioxidative system activity in the cortical homogenate as the level of SOD and GPx expression was significantly increase which similar that of the control groups (CON & LUT). Lead intake (Pb) promotes lipid peroxidation in rat cortical lysates as shown by significant increase of MDA level in relation to other experimental groups (fig 1c). Luteolin inhibits lipid peroxidation among groups treated with, before, and after lead intake as the level of cortical MDA was significant decreased similarly to that of the control groups. (*=p<0.05, **=p<0.01, and ***=p<0.005).
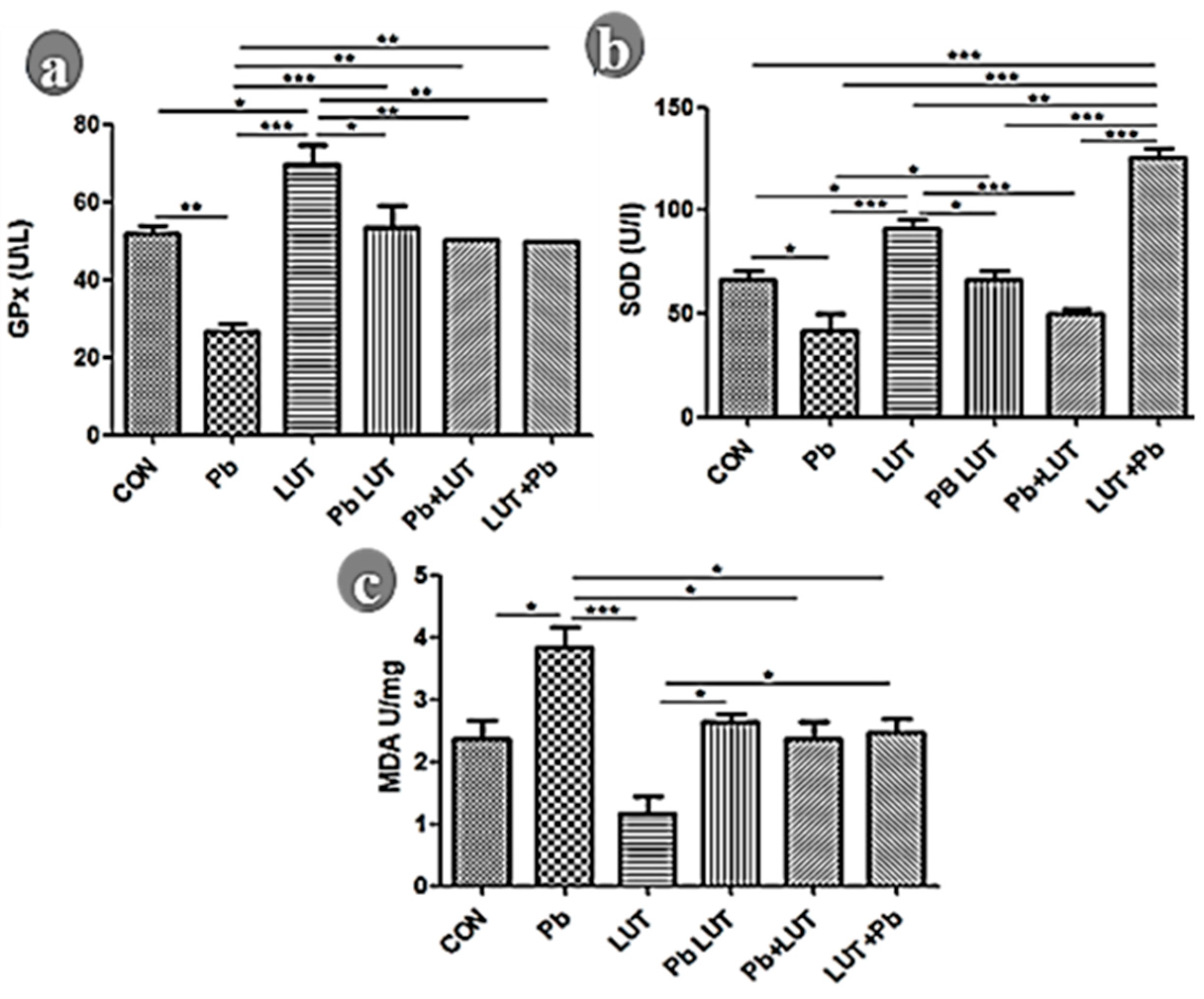
Figure 4.
GFAP anti-rat antibody demonstrates astrocyte immunoreactivity across experimental groups. Normal astrocyte density and morphology are seen in the cerebral cortex of rats in the control and LUT groups (black arrows), with sparse astrocytes around neurons and between layers and regular processes. The morphology of astrocytes was altered by lead treatment (red arrows), resulting in an increase in density with reactive astroglia and the presence of hypertrophic cells within the cerebral, indicating astrogliosis. LUT treatment improves astrocyte density and morphology in rats in the LUT+Pb, Pb+LUT, and PbLUT groups slightly more than in the control groups (400x).
Figure 4.
GFAP anti-rat antibody demonstrates astrocyte immunoreactivity across experimental groups. Normal astrocyte density and morphology are seen in the cerebral cortex of rats in the control and LUT groups (black arrows), with sparse astrocytes around neurons and between layers and regular processes. The morphology of astrocytes was altered by lead treatment (red arrows), resulting in an increase in density with reactive astroglia and the presence of hypertrophic cells within the cerebral, indicating astrogliosis. LUT treatment improves astrocyte density and morphology in rats in the LUT+Pb, Pb+LUT, and PbLUT groups slightly more than in the control groups (400x).
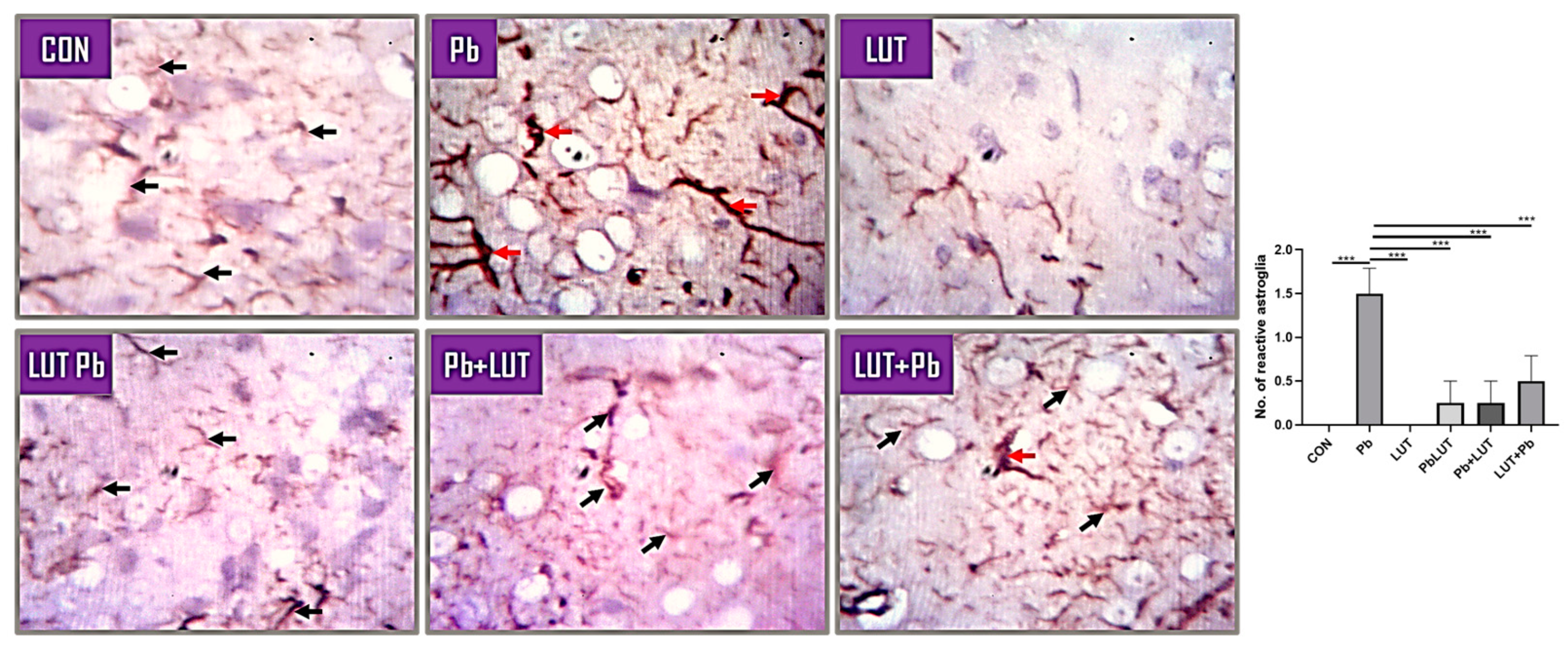
Figure 5.
Immunohistochemistry of rats’ cerebral cortex from various experimental groups demonstrating synaptic density and integrity as measured by synaptophysin. The immunostaining for anti-synaptophysin was used to compare the synaptic density and integrity of neurons in the cerebral cortex of rats across treatment groups. The cerebral of rats in the control and LUT groups was positive for synaptophysin immunostaining, indicating effective communication between neurons. Because there was no reactivity to synaptophysin immunostaining, the cerebral cortex of rats in the pathology group (Lead) had compromised synaptic vesicles, density, and integrity. Post-treatment (Pb+LUT), pre-treatment (LUT+Pb), and concurrent treatment with LUT shows reactivity to synaptophysin immunostaining that is comparable to the control groups (400x).
Figure 5.
Immunohistochemistry of rats’ cerebral cortex from various experimental groups demonstrating synaptic density and integrity as measured by synaptophysin. The immunostaining for anti-synaptophysin was used to compare the synaptic density and integrity of neurons in the cerebral cortex of rats across treatment groups. The cerebral of rats in the control and LUT groups was positive for synaptophysin immunostaining, indicating effective communication between neurons. Because there was no reactivity to synaptophysin immunostaining, the cerebral cortex of rats in the pathology group (Lead) had compromised synaptic vesicles, density, and integrity. Post-treatment (Pb+LUT), pre-treatment (LUT+Pb), and concurrent treatment with LUT shows reactivity to synaptophysin immunostaining that is comparable to the control groups (400x).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



