Submitted:
07 June 2025
Posted:
09 June 2025
You are already at the latest version
Abstract
Globally, rice cultivation consumes large amounts of fresh water and urgent improvements in water use efficiency (WUE) are needed to ensure sustainable production, given the increasing water scarcity. While stomatal traits have been a primary focus for enhancing WUE, complex interactions between stomatal and non-stomatal leaf traits remain poorly understood. In this review, we present an analysis of stomatal and non-stomatal leaf traits influencing WUE in rice. The data suggests that optimising stomatal density and size will be insufficient to maximise WUE because non-stomatal traits such as mesophyll conductance, leaf anatomy, and biochemical composition significantly modulate relationships between stomatal conductance and photosynthetic rate. Integrating recent advances in high-throughput phenotyping, multi-omics technologies, and crop modelling, we suggest that combinations of seemingly contradictory traits can enhance WUE without compromising yield potential. We propose a multi-trait breeding framework that leverages both stomatal and non-stomatal adaptations to develop rice varieties with superior WUE and climate resilience. This integrated approach provides a roadmap for accelerating the development of water-efficient rice cultivars, with broad implications for improving WUE in other crops.
Keywords:
climate adaptation
; crop sustainability
; leaf physiology
; rice breeding
; water management
1. Introduction
Water scarcity threatens global food security, mainly because over half the world's population depends on water-intensive crops like rice (Oryza sativa L.) [1,2]. There is substantial evidence that climate change is exacerbating water scarcity, and enhancing the water use efficiency (WUE) of rice crops has become a significant issue for sustaining agricultural productivity [3,4]. Consequently, in addition to testing alternative agronomic methods [5,6,7], researchers worldwide are seeking to breed rice varieties with greater WUE [8,9,10,11,12]. Plant leaves play a pivotal role in water exchange, acting as the primary site for carbon dioxide uptake and water vapour loss through transpiration [4]. Historically, efforts to improve WUE have focused predominantly on stomatal traits, such as stomatal density, size, and conductance. There is, however, mounting evidence that non-stomatal leaf traits, including photosynthetic efficiency, leaf anatomy, and biochemical composition, significantly influence WUE in rice. [13,14,15].
We seek to provide a comprehensive overview of how both stomatal and non-stomatal leaf traits determine WUE in rice. We explore intricate interactions among these traits and their combined impact on water conservation and carbon assimilation. We examine the complex interplay of leaf traits to identify novel strategies for breeding rice varieties with superior WUE without compromising yield potential. Included in our consideration are (1) the varying scales of WUE measurement, from leaf-level gas exchange to whole-plant carbon isotope discrimination, (2) recent advances in the understanding of stomatal behaviour, mesophyll conductance, leaf anatomy, and metabolic adaptations to water stress, (3) emerging technologies, such as high-throughput phenotyping and multi-omic approaches. By integrating these factors, we aim to provide a more holistic understanding of breeding programs for WUE in rice, including potential synergies and trade-offs. Finally, we consider how agronomic practices [16,17] and environmental factors interact with leaf traits to influence WUE under field conditions.
Rice cultivation spans a diverse range of agroecosystems, from traditional continuously flooded paddies to water-limited upland and dryland environments. In flooded systems, WUE traits may contribute to optimising water productivity without imposing stress on the crop, whereas in upland systems, these traits are critical for survival, resilience, and yield stability under drought conditions. The physiological and anatomical mechanisms underlying WUE, such as stomatal behaviour, root architecture, and osmotic adjustment, therefore differ in their functional importance depending on the cultivation system. Recognising this spectrum is essential for developing rice varieties adapted to varying water regimes.
WUE can be broadly defined as the ratio of biomass produced or yield obtained to the amount of water used [9,12]. However, the concept of WUE encompasses multiple scales and definitions depending on the context of measurement. At the leaf level, WUE is typically expressed as the ratio of carbon assimilation to water loss through transpiration [10]. This can be quantified as intrinsic water use efficiency (WUEi), defined as the ratio of net CO2 assimilation rate (An) to stomatal conductance (gs); or alternatively, computed as instantaneous water use efficiency (WUEinst), calculated by dividing An by the transpiration rate (E) [10,11,18]. These leaf-level measurements provide insights into the physiological basis of WUE and are influenced by stomatal and non-stomatal leaf traits. Measurement techniques for leaf-level WUE in rice include gas exchange analysis using infrared gas analysers, which allow for the simultaneous measurement of photosynthesis and transpiration rates [11,19] and carbon isotope discrimination (Δ13C) over the period of leaf development [20,21]. Additionally, chlorophyll fluorescence techniques can provide insights into photosynthetic efficiency, which is closely linked to WUE [22].
At the whole-plant level, WUE can be assessed as the ratio of biomass accumulation to cumulative water use [23,24,25,26]. This approach captures both productive and non-productive water losses. Field-scale measurements often focus on yield-based WUE, calculated as crop yield per unit of water applied or lost through combined evaporation and transpiration [12]. Recent advances in high-throughput phenotyping, including thermal imaging and remote sensing techniques, enable more rapid and precise measurements of WUE-related traits in large populations [27]. These technologies, combined with genomic approaches, are accelerating the identification of genetic determinants of WUE in rice. Understanding the physiological mechanisms underlying WUE variability at the leaf and whole plant level is essential for developing rice varieties with improved drought tolerance and reduced water requirements.
2. Stomatal Leaf Traits and WUE in Rice
Stomatal leaf traits refer to the characteristics and properties of the stomata, including their density, size, distribution, and conductance, which are regulated by controlling gas exchange between leaves and the atmosphere. As highlighted in a recent herbarium study of plant adaptation to climate change [28], there is extensive knowledge of the dozens of genes involved in stomatal development (density, size, distribution).
2.1. Stomatal Density, Size and Arrangement
Recent studies have demonstrated that modifying stomatal density and size through crop engineering offers a promising approach to enhancing WUE and mitigating water scarcity challenges in agricultural crops [4,29]. Rice exhibits unique stomatal structures compared to other plant species, with greater stomatal densities and smaller stomatal sizes. The stomatal complex in rice comprises two dumbbell-shaped guard cells surrounded by subsidiary cells that regulate the stomatal aperture [30,31].
Four distinct models of leaf stomata were described in mutant populations of the purple rice cultivar, Jao Hom Nin, which varied in stomatal density and size [3]. Rice varieties with low stomatal density and smaller stomatal size exhibited improved WUE and biomass production under both field and greenhouse conditions. Similarly, three purple rice mutants were adapted to severe water stress conditions during the reproductive stage of growth, which resulted in low stomatal density and greater WUE under restricted water conditions [32]. Despite these findings, gaps remain regarding the physiological trade-offs of reduced stomatal density, particularly under fluctuating environmental conditions. While smaller, fewer stomata conserve water, they may limit CO₂ uptake under high demand. These dynamics are not yet fully quantified across diverse environments and growth stages.
Genetic manipulation of stomatal traits has shown promise in improving WUE. The high-yielding rice variety IR64 was engineered to reduce stomatal density by enhancing the expression of OsEPF1, a gene involved in rice epidermal patterning [33]. The resulting plants exhibited reduced stomatal conductance and enhanced water conservation. These traits were interpreted as improving drought tolerance while maintaining or improving yields under elevated atmospheric CO2 and high-temperature conditions. A previous study explored the impact of the arrangement of leaf stomata on the physiological responses of rice to differing water availability [34]. Yet, the implications of this spatial arrangement for canopy-level water and carbon fluxes remain underexplored. They proposed that greater stomatal density in the upper leaves (flag leaf and second leaf) with reduced density but larger stomata in the lower leaves (third and fourth leaf) constituted an adaptive strategy (reduced water loss) for genotypes under limited water availability. The study concluded that this stomatal configuration could enhance water productivity in rice under aerobic conditions without yield reduction.
2.2. Stomatal Conductance and Aperture
Leaf stomatal conductance (gs) is a critical physiological parameter influenced by a wide range of environmental and plant factors, including light, temperature, humidity, CO2 concentration, and plant water status [4,35]. The relationship between gs and WUE involves a trade-off between maximising CO2 uptake for photosynthesis and minimising water loss through transpiration. Recent studies have focused on understanding genetic and molecular mechanisms that regulate stomatal conductance and aperture in rice. A previous study revealed that precise control of the stomatal aperture is crucial to maintaining an optimal balance between photosynthesis and water conservation [36]. The study identified genes such as EPF2, OSA1, and SLAC1 involved in stomatal regulation, enabling the enhancement of WUE through genetic manipulation. However, the long-term physiological impacts of manipulating these genes under fluctuating climate conditions remain poorly understood. In particular, the extent to which these molecular interventions affect gas exchange efficiency under combined heat and water stress is underexplored. Comparative evidence from model species like Arabidopsis suggests variability in stomatal responsiveness and downstream signaling efficiency, indicating that gene function may be context-dependent across crops [37].
2.3. Regulation of Transpiration and Carbon Dioxide Uptake
Abscisic acid (ABA) is widely believed to have a central role as a signal for the closure of stomata during water stress [35,38]. Other hormones, such as cytokinins and gibberellins, also contribute to stomatal regulation. Plant circadian rhythms reflect the significance of stomatal conductance, ensuring that stomatal opening and closing are synchronised with environmental light and temperature patterns [38]. This temporal coordination helps optimise WUE and CO2 uptake and contributes to plant growth and survival under varying environmental conditions. Similarly, ion transport and osmotic adjustments within stomatal guard cells facilitate rapid changes in the stomatal aperture, balancing the need for CO2 uptake with water conservation. Fine-tuning these processes enhances plant survival in diverse and changing environments [3,38]. The movement of ions such as potassium (K⁺), chloride (Cl⁻), and calcium (Ca²⁺), along with malate (C₄H₄O₄²⁻), causes water to enter guard cells, leading to stomatal opening for CO₂ uptake during photosynthesis. Conversely, water follows when ions efflux from the guard cells, causing the stomata to close thereby reducing water loss (Figure 1A). The fine-tuning of this mechanism is essential for balancing the demand for CO₂ with the need for water conservation, especially under drought stress [39,40]. Despite this mechanistic clarity, limited studies have explored how these responses are modulated across rice genotypes with varying limited water stress or drought resilience. Moreover, cross-species comparisons of hormonal regulation and ion channel dynamics remain scarce, leaving uncertainties about whether similar mechanisms operate with equal efficiency in monocot cereals versus dicot plants.
In addition to stomatal and cuticular transpiration, grasses such as rice also release water through hydathodes: specialised structures located at leaf margins that facilitate guttation under conditions of high soil moisture and low transpiration demand. Hydathodes, unlike stomata, remain open and serve as low-resistance pathways for liquid exudation, playing a role in water release and nutrient exudation [41]. This process complements transpiration and may contribute to maintaining xylem flow when evaporative demand is low. Furthermore, rice leaves contain well-defined vascular bundles composed of xylem and phloem, which are responsible for the transport of water, nutrients, and photosynthates. These vascular structures are closely associated with companion cells and are integral to leaf physiology, supporting both hydraulic conductance and photosynthate distribution [42].
3. Non-Stomatal Leaf Traits and WUE in Rice
Non-stomatal leaf traits in rice encompass the structural and biochemical characteristics of the leaf which influence photosynthetic efficiency and WUE independent of stomatal behaviour.
3.1. The Role of ΦPSII in Photosynthetic Efficiency
In this review, photosynthetic efficiency is categorised as a non-stomatal leaf trait because it primarily reflects the biochemical and physiological processes within the leaf, including the efficiency of the photosynthetic machinery, such as chloroplasts, enzymes, and photosystems like ΦPSII in converting light energy into chemical energy. Photosynthetic efficiency refers to the ability of plants to convert light energy into chemical energy during photosynthesis [43]. Non-stomatal factors, such as the biochemical capacity of the mesophyll cells, chlorophyll content, and the efficiency of the photosynthetic machinery, play a pivotal role in determining photosynthetic efficiency [4,20,44,45]. These factors can enhance the plant's ability to assimilate CO2 under varying environmental conditions, thereby optimising WUE. By improving photosynthetic efficiency, plants can achieve higher rates of CO₂ assimilation per unit of water transpired. This balance is achieved by optimising stomatal conductance, so that stomata remain partially open to allow CO₂ uptake while reducing excessive water loss. Improved mesophyll conductance, Rubisco efficiency, or enhanced photoprotection mechanisms can also contribute to higher intrinsic WUE under drought stress [46]. This balance between CO2 uptake and water conservation is essential for plant productivity and resilience, making photosynthetic efficiency a key target for crop improvement strategies [47].
3.2. Leaf Anatomy
Mesophyll structure, vein density, and bulliform cells significantly impact CO2 diffusion, light distribution, and water management within leaves [48]. Mesophyll conductance (gm), an important factor influencing CO₂ diffusion, is discussed in more detail in section 3.5. Leaf thickness and cell arrangements influence light distribution and absorption, while chloroplast positioning optimises light harvesting and minimises photodamage. However, these anatomical features often involve trade-offs. For instance, it has been reported that increased vein density in rice was correlated with greater photosynthetic rates but reduced WUE [49], highlighting the complex interplay between structural traits and physiological efficiency.
Aquaporins, a family of integral membrane proteins, play a dual role in regulating both water transport and CO₂ diffusion across leaf tissues, thereby influencing photosynthetic efficiency and WUE in rice. While CO₂ is abundant in the atmosphere, its diffusion into the chloroplasts, where photosynthesis occurs, is limited by mesophyll conductance, which includes movement through intercellular air spaces, cell walls, and membranes. Certain aquaporins, particularly plasma membrane intrinsic proteins (PIPs), facilitate this process by enabling CO₂ to cross cell membranes more efficiently [50]. In parallel, aquaporins influence leaf hydraulic conductance, which refers to the efficiency of water movement from the xylem to the sites of evaporation within the leaf. This conductance affects stomatal behavior and transpiration, thereby indirectly modulating photosynthesis and WUE [48,51]. Environmental factors such as nitrogen availability can regulate aquaporin expression and activity, further linking nutrient status to water and carbon dynamics in leaves [48]. A balanced distribution of aquaporins in mesophyll, bundle sheath, and vascular tissues is considered optimal for improving WUE and enhancing crop resilience under climate stress [52,53].
Studies have emphasised the significance of internal leaf anatomy, including aerenchyma and intercellular air spaces, in regulating CO₂ diffusion to the sites of photosynthesis. CO₂ must first dissolve in the thin water layer lining the mesophyll cell walls before diffusing into the symplast and reaching the chloroplasts. Therefore, a greater intercellular surface area, supported by well-developed aerenchyma, enhances both CO₂ diffusion efficiency and evaporative capacity. This anatomical feature contributes to gₘ and plays a critical role in photosynthetic performance and WUE, particularly under stress conditions (Figure 1B) [54,55,56].
Studies have further demonstrated that the midvein-deficient mutant shows higher vein density and photosynthetic efficiency but significantly lower WUE. Although the study did not include water-stress conditions, the mutants are still recommended as promising rice breeding material due to their enhanced photosynthetic efficiency [49]. In a glasshouse experiment with limited water (60% field capacity), it has been observed that high leaf mass area (LMA) mutants exhibited higher WUE and a more significant total number of veins than low-LMA mutants, which had lower WUE. However, high-LMA mutants also showed lower vein density (veins per centimetre) and fewer bulliform cells than their low-LMA counterparts [14].
Abaxial stomatal dominance, where stomata are mainly located on the leaf’s lower surface, helps reduce water loss by limiting stomatal exposure to direct sunlight and wind. This spatial trait can enhance WUE in rice, especially under drought or high vapor pressure conditions. When combined with other anatomical traits like reduced stomatal density and thicker cuticles (discusses in the next section 3.3), abaxial dominance offers a promising target for breeding rice varieties with improved drought resilience and efficient gas exchange [33,57].
3.3. Leaf Cuticle and Epicuticular Wax
Leaf cuticles and epicuticular wax (EW) are critical components in regulating non-stomatal water loss and enhancing WUE in rice. In rice, the β-Ketoacyl-CoA Synthase (KCS) enzyme is essential for the elongation of very long-chain fatty acids, which are major components of cuticular wax [58]. It has been reported that increased expression of the OsCUT1 gene in rice resulted in improved drought tolerance and wax synthesis without reducing yield [59]. On the other hand, the OsGL1-1 gene plays a crucial role in cuticular wax deposition and drought resistance in rice. Mutants with reduced OsGL1-1 expression showed decreased cuticular wax deposition, increased water loss, and enhanced sensitivity to drought compared to wild-type plants [60]. Furthermore, previous research has demonstrated that EW traits contribute to rice resistance against pests, evidenced by EW rice mutants exhibiting reduced resistance to the rice water weevil (Coleoptera: Curculionidae) and the fall armyworm (Lepidoptera: Noctuidae) (Figure 1C) [61].
3.4. Metabolomics Changes in the Leaves
Drought stress triggers complex metabolic responses in plants, regulated by gene activity, which impact vital processes like photosynthesis, respiration, and assimilate translocation [62]. These changes help the plant adjust osmotically, manage reactive oxygen species (ROS), and stabilise cell membranes, crucial for maintaining function during drought.
Drought-tolerant rice cultivars often accumulate greater concentrations of osmoprotectants, including low molecular weight osmolytes such as glycine betaine and proline, as well as organic acids and soluble sugars [62,63]. These compounds help sustain cellular functions and contribute to defence mechanisms under water-limited conditions. Additionally, increased concentrations of antioxidants, phenolic compounds, and flavonoids have been observed in drought-stressed rice leaves, with flavonoids like kaempferol and quercetin playing a significant role in enhancing drought tolerance and modulating WUE [64].
3.5. Mesophyll Conductance (gm) and Intrinsic WUE
gm is a key physiological parameter that governs the rates of CO2 assimilation [44,65]. Von Caemmerer and Evans (2010) suggested that increasing photosynthesis without altering stomatal function can improve WUE, and it also implied that enhancing gs could boost photosynthesis if water is not limited. gm is measured as the rate of CO2 diffusion from intercellular spaces to chloroplasts, which substantially affects WUE. Varieties with greater gm also show improved carbon assimilation per unit of water transpired [15]. Additionally, they mentioned that manipulating gm could increase CO2 availability at Rubisco, the key enzyme in photosynthesis that catalyses the fixation of CO₂ into organic molecules in the Calvin cycle, potentially improving photosynthesis and WUE [66]. Furthermore, Flexas et al. (2013) stated that to simultaneously improve photosynthesis and WUE, genetic manipulation of gm should be done without parallel changes in gs. It was suggested that the optimal trait for enhancing WUE is an increased ratio of gm to gs [67].
For rice, gm plays a significant role in transpiration efficiency (TE) and could affect WUE. Upland rice and O. glaberrima showed a stronger response of TE to the gm/gs ratio under drought conditions, suggesting a link between gm and WUE. Furthermore, it suggested that increasing rice's gm/gs ratio, potentially by modifying leaf anatomy, could enhance both TE and WUE under water-limited conditions [68].
Strategies like genetic manipulation or selective breeding can significantly improve WUE without compromising photosynthetic capacity. The α subunit of a heterotrimeric G protein regulates gm and drought tolerance in rice, suggesting that modifying G protein signalling could enhance WUE in rice cultivars [69].
3.6. Leaf Canopy Architecture
Rice varieties with higher total leaf area and optimised canopy structure exhibited improved WUE under water-limited conditions [34]. Rice mutants with LMA showed increased WUE, increased photosynthetic capacity, and superior carboxylation efficiency compared to low-LMA mutants under both well-watered and water-limited conditions [14,70].
A glasshouse study conducted on the Azucena (tropical japonica) and IR64 (India) rice varieties revealed a correlation between increased leaf width and lower Δ13C. Leaves with higher widths have excellent leaf-boundary layers that resist gaseous diffusion, which would lower the Δ13C and increase WUE [71].
Higher crop density creates dense canopies and increases the thickness of the boundary layer, the layer of still air that surrounds each leaf. This reduces evaporative demand and water loss and potentially improves WUE. However, it may also limit CO₂ diffusion, affecting photosynthesis [72,73].
Erect leaves, which form smaller angles with the stem, allow greater light penetration to lower canopy layers, thereby enhancing overall photosynthetic efficiency and reducing excessive transpiration in the upper canopy. This spatial leaf arrangement has been demonstrated in rice genotypes to improve light interception and optimise water use, contributing to higher WUE [74,75]. Furthermore, studies on spatial leaf arrangement and canopy architecture in rice show that coordinated leaf positioning around the stem improves light distribution throughout the canopy, enhances carbon assimilation, and reduces water loss. This trait is particularly beneficial in high-density planting systems, where maximizing light use efficiency and conserving water are critical for sustaining productivity under variable environmental conditions [76,77].
3.7. Leaf Pubescence and Boundary Layer Resistance
Leaf hairiness (pubescence) has been understudied in rice despite its potential to enhance water conservation and photosynthetic efficiency. Rice varieties show differences in leaf hairiness, from smooth to densely hairy surfaces. These hairs form a boundary layer that slows water vapour diffusion, reducing transpiration and enhancing light reflectance, which can protect the leaf from excessive radiation (Figure 1D) [73,78].
It has been reported that the genetic basis, feasibility of transferring this trait between rice varieties and physiological benefits of leaf hairiness on WUE in rice by introducing a gene from chromosome 6 from hairy-leafed Oryza nivara into the less hairy IR24 variety, creating an introgression line with the hairy-leaf trait (IL-hairy). The IL-hairy plants exhibited warmer leaf surfaces under sunlight due to the increased boundary layer resistance conferred by the leaf hairs and lower transpiration rates under moderate and high light intensities, leading to significantly higher photosynthetic WUE [73].
3.8. Carbon Fixation Efficiency
Carbon fixation efficiency is discussed in this review as a non-stomatal trait because it primarily reflects the internal biochemical capacity of the plant to assimilate carbon, beyond the regulation of gas exchange at the stomatal level. While stomatal conductance indeed influences CO₂ availability in the intercellular spaces, the efficiency with which CO₂ is fixed into carbohydrates, governed by enzymes like Rubisco, chloroplast function, and mesophyll conductance, is largely non-stomatal in nature.
Δ¹³C and carbon isotope ratio (δ¹³C) are widely used indicators of WUE in C₃ plants like rice. Δ¹³C reflects the degree of discrimination against ¹³CO₂ during photosynthesis, which is influenced both by stomatal behaviour and the biochemical processes involved in carbon fixation [20,79]. Higher Δ¹³C values indicate greater discrimination against ¹³C, typically associated with higher stomatal conductance and lower WUE [80]. Conversely, δ¹³C, which is derived from plant tissue and reflects the integrated effects of carbon assimilation over time, is negatively correlated with Δ¹³C [81]. A higher δ¹³C value suggests reduced stomatal conductance and improved carbon fixation efficiency per unit water loss, thus serving as a reliable proxy for assessing WUE in rice, especially under water-limited conditions [82].
4. Integrating Stomatal and Non-Stomatal Traits for WUE Improvement
Improving WUE in rice requires an integrated approach that considers both stomatal and non-stomatal leaf traits. This will enable the development of more effective strategies for sustainable rice production under water-limited conditions.
4.1. Interactions and Trade-Offs Among Leaf Traits
Reducing stomatal density can enhance water conservation [33], but it may also limit CO2 uptake, potentially impacting photosynthetic efficiency. Conversely, increasing gm can improve photosynthetic rates but may lead to greater water loss if not balanced with appropriate stomatal control [45]. Hence, some studies have highlighted the importance of considering multiple traits simultaneously. It has been demonstrated that rice varieties with both optimised stomatal traits and enhanced gm exhibited superior WUE compared to those improved for single traits alone [83]. Further, it has been found that stomatal traits modulated the relationship between leaf nitrogen content and WUE, emphasizing the need for integrated trait selection in breeding programs [84].
4.2. Breeding Strategies for Optimising WUE Through Leaf Traits
Optimising WUE in rice through breeding involves selecting specific leaf traits that improve water management and photosynthetic performance. This multi-trait approach combines traditional breeding with advanced genetic and genomic tools, focusing on stomatal behaviour, leaf morphology, physiological traits, photosynthetic efficiency, and biochemical pathways to develop more water-efficient, productive rice varieties.
Advances in quantitative trait loci (QTL) mapping have allowed the identification of genetic regions linked to desirable leaf traits, such as leaf area, thickness, and orientation. By correlating genetic variations with these traits, QTL mapping helps breeders efficiently integrate these traits into new plant varieties [85,86,87].
Modern genetic technique, marker-assisted selection (MAS), allows for the rapid integration of WUE-related traits by using molecular markers associated with key traits like stomatal conductance, leaf morphology, and Rubisco efficiency. Genomic selection takes this a step further by utilising genome-wide data to predict and select for high WUE traits, streamlining the breeding process and reducing the time needed to develop new varieties. This approach leverages high-throughput genotyping and phenotyping to build predictive models that can accurately identify the best candidates for breeding programs focused on improving WUE [88,89,90].
Recent studies have highlighted the importance of integrating both stomatal and non-stomatal traits to enhance WUE in rice. Stomatal morphology and physiology operate in a coordinated manner, influencing both transpiration efficiency and carbon assimilation [47]. Environmental drivers shape stomatal traits, which in turn interact with broader leaf structural features to modulate WUE at the ecosystem level [91]. Complementing these findings, another study [13] has shown that rice genotypes with low-frequency, large-size stomata exhibited higher intrinsic WUE and more responsive gas exchange under varying vapour pressure deficits. This study also linked epidermal patterning and the expression of key regulatory genes (e.g., ERECTA, TMM, YODA) to stomatal development, reinforcing the need to integrate leaf surface traits with internal physiological and anatomical features. Collectively, these findings support a holistic breeding approach that considers the interplay of multiple leaf traits to develop rice cultivars resilient to climate variability and water-limited environments.
4.3. Agronomic Practices and Environmental Factors Influencing WUE
Anthropogenic water management is an important factor influencing plant performance. Inefficient irrigation methods lead to low WUE and environmental issues. Several alternative rice production methods, such as aerobic rice, direct-seeded rice (DSR), alternate wetting and drying (AWD), and smart irrigation technologies, can improve WUE, though each has specific challenges like high costs or labour intensity [6,92]. Delayed Permanent Water (DPW) is an alternative water management practice extensively used in Australian rice cultivation to increase water productivity for drill-sown rice. In this method, the rice crop is sown and initially managed like conventional drill-sown rice, but the application of permanent water is delayed until at least 50 days after the first flush irrigation [93].
Further, it has been emphasised that to promote water-saving irrigation technology, countries should develop supportive policies, raise farmers' awareness, and offer training and financial assistance tailoring water-saving technologies to specific regions can reduce irrigation water use and improve rice WUE, helping to address global water scarcity challenges [94]. As a country with a well-managed rice industry, the Australian Government's Climate-Smart Agriculture Program provides funding for innovative irrigation projects [95]. Educational campaigns like Brisbane's "Every Drop Counts" raise awareness among farmers about water-saving practices [96]. Organizations such as Irrigation Australia offer training programs and financial subsidies to help farmers adopt new technologies [97].
Nutrient management such as applying balanced fertilisers, particularly nitrogen, in appropriate amounts can enhance plant growth and photosynthetic efficiency, leading to better WUE [9]. Sowing techniques, such as direct seeding and proper transplanting density, can optimise plant establishment and water utilisation. Direct seeding generally achieves higher WUE than traditional transplanting, though wet seeding by broadcast, a common method, results in lower WUE compared to traditional transplanting [98]. Moreover, uncontrolled weeds compete with rice for water and nutrients. It has been found that while weed control does not affect total water consumption, it influences WUE through its impact on yield [99]. Higher temperatures can raise transpiration rates, but practices like mulching help regulate plant temperature and reduce water loss. Non-flooded plastic film mulching and wheat straw mulching are effective options for improving WUE in rice cultivation [7]. Therefore, optimising WUE in rice production requires multiple approaches, integrating breeding for better leaf traits, effective agronomic practices, and consideration of environmental factors to achieve sustainable rice production in water-limited conditions.

Table 1.
Comprehensive list of rice genes involved in (A) stomatal and (B) non-stomatal leaf traits affecting water use efficiency. The table includes genes spanning key functional categories including stomatal development and regulation, cuticular wax biosynthesis, leaf morphology, stress response pathways, and metabolic adaptations. Locus IDs correspond to the Rice Annotation Project and were verified via the SNP-Seek database from the International Rice Research Institute [100]. Directional trends in the WUE Association column indicate: ↑ = positive association with increased WUE (gene overexpression or enhanced function improves water use efficiency); ↓ = negative association with WUE (gene overexpression or enhanced function reduces water use efficiency); ± = variable or context-dependent association (effects depend on environmental conditions, genetic background, or developmental stage).
Table 1.
Comprehensive list of rice genes involved in (A) stomatal and (B) non-stomatal leaf traits affecting water use efficiency. The table includes genes spanning key functional categories including stomatal development and regulation, cuticular wax biosynthesis, leaf morphology, stress response pathways, and metabolic adaptations. Locus IDs correspond to the Rice Annotation Project and were verified via the SNP-Seek database from the International Rice Research Institute [100]. Directional trends in the WUE Association column indicate: ↑ = positive association with increased WUE (gene overexpression or enhanced function improves water use efficiency); ↓ = negative association with WUE (gene overexpression or enhanced function reduces water use efficiency); ± = variable or context-dependent association (effects depend on environmental conditions, genetic background, or developmental stage).
| A. Stomatal leaf Traits | |||
|---|---|---|---|
| Gene | Locus ID | Role | WUE Association |
| OsEP3 | LOC_Os02g15950 | Regulates stomatal guard cell development [101]. ERECT PANICLE3 |
↑ WUE Optimises stomatal development for better water regulation |
| OsKAT1 | LOC_Os01g55200 | Regulates stomatal opening and closing by facilitating potassium ion flux, which directly affects stomatal movement and plant water regulation [102,103] K+ TRANSPORTER 1 |
↑ WUE improves stomatal control |
| OsSDD1 | LOC_Os03g0143100 | Mutations lead to higher stomatal density [13] STOMATAL DENSITY AND DISTRIBUTION |
↓ WUE increases water loss via excess stomata |
| OsTMM | LOC_Os01g02060 | Mutations can lead to irregular stomatal spacing, affecting gas exchange efficiency [13]. TOO MANY MOUTHS |
↓ WUE disrupts stomatal patterning |
| OsEPF1 | LOC_Os04g0637300 | Overexpress reduce stomatal density [36] EPIDERMAL PATTERNING FACTOR1 |
↑ WUE reduces transpiration |
| OsSPCH1 | LOC_Os06g33450 | Essential for the initiation of stomatal lineage, termination of meristemoid fate and the transition to guard mother cell (GMC) identity. Overexpression expression leads to higher stomatal density [33,104] SPEECHLESS1 |
↓ WUE Overexpression results in increased water loss |
| OsSPCH2 | LOC_Os02g15760 | alterations in stomatal size and density in stress conditions. Overexpression increases density [13,105] SPEECHLESS2 |
↓ WUE Overexpression results in increased water loss |
| OsMUTE | LOC_Os05g51820 | Essential for the initiation of stomatal lineage, termination of meristemoid fate and the transition to GMC identity. Involves in the differentiation of stomatal precursor cells. It plays a critical role in determining stomatal size by regulating the development of guard cells [104] | ↑ WUE ensures proper stomatal function |
| OsFAMA | LOC_Os05g50900 | Essential for the initiation of stomatal lineage, termination of meristemoid fate and the transition to GMC identity [104] | ↑ WUE Maintains correct stomatal identity for optimal function |
| OsFLP | LOC_Os07g43420 | Regulates the orientation of GMC symmetrical division [104] FOUR LIPS |
↑ WUE Regulates stomatal spacing for efficient gas exchange |
| B. Non-stomatal leaf traits | |||
| Gene | Locus ID | Role | WUE Association |
| OSH43 | LOC_Os03g56110 LOC_Os03g57560 |
Overexpression results in broader leaves, increased tiller number, and more flowers [106]. ORYZA SATIVA HOMEOBOX43 | ↑ WUE No direct evidence. |
| OsABCG9 | LOC_Os04g44610 | Mutations can lead to reduced cuticular wax content, resulting in increased sensitivity to drought and other environmental stresses[107] | ↓ WUE Mutation increases drought sensitivity due to water loss |
| GL1 | LOC_Os05g02730 | controlling wax synthesis in rice leaves [108] GLOSSY1 |
↑ WUE Improves cuticle barrier to reduce water loss |
| OSGL1-1 | LOC_Os09g25850 LOC_Os09g25850 |
Increase leaf cuticular wax deposition and enhance drought tolerance [60] GLOSSY1-1 |
↑ WUE Enhances drought resistance |
| OsGL1-2 | LOC_Os02g08230 | Overexpression increases cuticular wax production and improves drought tolerance [108] GLOSSY1-2 |
↑ WUE Overexpression reduces water loss |
| OsWR1 | LOC_Os02g10760 | Overexpression increases the expression of genes involved in wax synthesis [109] WAX SYNTHESIS REGULATORY GENE1 | ↑ WUE Overexpression reduces water loss |
| OsCutA1 | LOC_Os10g23204 | Overexpression increases cuticular wax production and improves drought tolerance, a promising gene for engineering rice plants with enhanced drought tolerance [59] | ↑ WUE Overexpression reduces water loss |
| YGL1 | LOC_Os05g28200 | Overexpression results in darker green leaves, increased chlorophyll content, and increased photosynthetic activity. This leads to improved growth and increased yields [110]YELLOW-GREEN LEAF1 | ↑ WUE Overexpression improves carbon assimilation efficiency |
| OsHB2 | LOC_Os10g33960 | Overexpression of the OsHB2 gene results in longer roots, broader leaves, and increased tolerance to drought and salinity stress [111] HOMOEOBOX1 | ↑ WUE Overexpression improves water capture and leaf area for photosynthesis |
| RCN1 | LOC_Os11g05470 | Inhibiting flowering transition and delaying heading under drought. Many rachis branches in the panicle and high yield [112] ROOTS CURL IN NAPHTHYLPHTHALAMIC ACID1 |
± WUE Delays maturity but may improve yield under drought |
| OsCKX2 | LOC_Os01g10110 | Produce larger leaves, more flowers, and higher yields [113] CYTOKININ OXIDASE/DEHYDROGENASE2 |
↑ WUE Improves resource allocation for higher yield |
| Gn1a | LOC_Os01g10110 | Improves grain yield, resulting in more flowers and larger grains [114] GRAIN NUMBER 1a identical to CKX2 |
↑ WUE Increases yield potential under stress |
| NAL3 | LOC_Os12g01120 | Overexpression results in broader leaves, reduced tiller number, increased lateral root development, and larger grains [115]NARROW LEAF3 | ↑ WUE Overexpression enhances soil water access and light interception |
| NRL1 | LOC_Os12g37190 | Overexpression results in wider leaves, increased plant height, larger vascular bundles, stronger stems, and higher yields [106,116] NARROW AND ROLLED LEAF1 |
↑ WUE Overexpression improves plant robustness and water transport |
| NAL1 | LOC_Os04g52479 | Overexpression results in broader leaves, reduced tiller number, increased lateral root development, and larger grains [117] NARROW LEAF1 |
↑ WUE Improves growth under water-limited conditions |
| NAL7 | LOC_Os03g06654 | Increased expression could have wider leaves, taller plants, and higher yields [106,117]. NARROW LEAF7 |
↑ WUE Enhances light interception and productivity |
| OsSPL14 | LOC_Os08g39890 | Regulates tiller number and panicle branching, which can also lead to increased grain yield [118]. SQUAMOSA PROMOTOR BINDING PROTEIN-LIKE14 |
↑ WUE Enhances yield per unit water use |
| OsSPL9 | LOC_Os05g33810 | Increases grain number per panicle and yield, and regulates drought tolerance, which can improve rice performance in dry environments [119]. SQUAMOSA PROMOTOR BINDING PROTEIN-LIKE9 |
↑ WUE Enhances productivity under drought stress |
| WSL1 | LOC_Os06g39750 | Involved in the biosynthesis of cuticular wax in rice leaves [120] WHITE STRIPED LEAF1 |
↑ WUE Reinforces leaf cuticle to reduce evaporation |
| OsMyb6 | LOC_Os06g10350 | Overexpression in rice improves drought and salinity tolerance [121] MYB-TKF6 |
↑ WUE Overexpression enhances stress response and water retention |
| OsPIP1;1 | LOC_Os02g44630 | facilitate osmotic water transport across membranes [122] PLASMAMEMBRANE INTRINSIC PROTEIN1;1 |
± WUE Can improve water uptake or increase loss depending on conditions |
| OsPIP2;1 | LOC_Os07g26690 | contributes to water transport and is highly expressed in roots and leaves [122] PLASMAMEMBRANE INTRINSIC PROTEIN1;1 |
± WUE Context-dependent water transport regulation |
| OsNAC6 | LOC_Os01g66120 | Regulates stress responses, including drought tolerance, by modulating root architecture and other physiological traits. Overexpressing transgenic plants displayed an accelerated leaf senescence phenotype at the grain-filling stage [123] NAM, ATAF1/2, and CUC2-FAMILY6 |
± WUE Accelerates senescence; balance needed for net benefit |
| OsDREB2B | LOC_Os05g27930 | Involves in water- and heat-shock stress responses and tolerance [124] DEHYDRATION-RESPONSE ELEMENT-BINDING PROTEIN2 |
↑ WUE Boosts adaptive response under stress |
| OsbZIP23 | LOC_Os02g52780 | Overexpression shows improved tolerance to drought and high-salinity stresses and sensitivity to ABA [125] BASIC-ZIPPER23 |
↑ WUE Enhances tolerance through ABA sensitivity |
| CFL1 | LOC_Os02g31140 | Reduced expression resulted in the reinforcement of cuticle structure [126] CURLY FLAG LEAF1 |
↑ WUE Reduced expression improves barrier to water loss |
| OsCHR4 | LOC_Os07g31450 LOC_Os07g32430 |
regulates leaf morphogenesis and cuticle wax formation [127] CHROMATIN REMODELLING FACTOR4 |
↑ WUE Enhances wax formation for water conservation |
| OsABA8OX3 | LOC_Os09g28390 | Control ABA level and drought stress resistance in rice [128] ABA 8´HYDROXYLASE3 |
↑ WUE Enhances wax formation for water conservation |
| OsAPX7 | LOC_Os04g35520 | involved in signalling transduction pathways related to drought stress response [129] ASCORBATE PEROXIDASE7 |
↑ WUE Activates antioxidant pathways under stress |
| OsTPS1 | LOC_Os01g23530 LOC_Os05g44210 LOC_Os05g44310 LOC_Os05g44300 LOC_Os08g34580 |
enhance the abiotic stress tolerance by increasing the amount of trehalose and proline [130] TREHALOSE-6-PHOSPHATE SYNTHASE1 |
↑ WUE Accumulates osmoprotectants for drought resilience |
4.4. WUE Optimization Strategies Between Traditional and Dryland Rice Farming Systems
Rice cultivation spans a diverse range of agroecosystems, from continuously flooded lowland paddies to water-limited upland or dryland fields. These contrasting environments impose distinct physiological constraints and selection pressures on rice plants, resulting in divergent strategies for optimising WUE. Figure 2 compares key physiological, anatomical, and agronomic traits of rice grown in traditional paddy systems versus dryland/upland systems.
In traditional irrigated paddy systems, the presence of standing water creates a saturated microenvironment with high humidity and low evaporative demand. This decouples plant water status from atmospheric vapor pressure deficit (VPD), reducing transpirational losses [131]. Consequently, irrigated rice typically develops shallow root systems and extensive aerenchyma tissue to facilitate internal oxygen transport under anaerobic conditions. Stomatal conductance in these systems tends to be relatively high and less responsive to VPD fluctuations. Additionally, non-stomatal adaptations such as epicuticular wax deposition and leaf rolling are minimal, as the consistently moist conditions reduce the need for such drought-avoidance traits [6,132,133,134].
In contrast, dryland or upland rice is cultivated under aerobic conditions where water availability is often intermittent and limiting. These environments favor more conservative water use strategies. Upland rice varieties typically exhibit deeper and more highly branched root systems to access subsoil moisture. They also demonstrate tighter stomatal regulation, with greater sensitivity to both soil water potential and atmospheric VPD. Non-stomatal adaptations are more pronounced, including increased epicuticular wax accumulation, enhanced leaf rolling during drought stress, and the production of osmoprotectants such as proline and soluble sugars that help maintain cellular integrity under dehydration [6].
Management practices to enhance WUE also differ significantly between the two systems. In irrigated paddy fields, techniques like AWD have been adopted to reduce water inputs while maintaining yield stability. This method intermittently drains fields to encourage deeper rooting and reduce methane emissions. In dryland systems, practices such as mulching, deficit irrigation, and optimised planting densities are employed to reduce surface evaporation, improve soil moisture retention, and enhance root-soil contact [135,136]. These contrasting strategies underscore the importance of context-specific breeding and agronomic interventions. While some core physiological principles of WUE, such as efficient stomatal control and root system architecture, are relevant across systems, their functional significance and genetic regulation vary with environmental and management conditions.
5. Future Perspectives and Research Directions
The exploration of WUE in rice is entering an exciting new era, driven by technological advances and interdisciplinary approaches. This last section examines emerging technologies that are revolutionising our ability to study leaf traits and WUE, from high-resolution imaging techniques to multi-omics approaches (Table 2). It also addresses the key challenges facing researchers and breeders in developing water-efficient rice cultivars and discusses the broader implications of this work for sustainable agriculture and climate resilience. By highlighting these future directions, we aim to provide a roadmap for researchers and emphasise the critical importance of continued innovation in this field.
5.1. Emerging Technologies and Approaches for Studying Leaf Traits and WUE
Recent technological advancements have revolutionised our ability to study leaf traits and WUE in rice at unprecedented levels of detail and scale. These innovations span a wide range of disciplines, from advanced imaging techniques to molecular biology approaches, offering researchers powerful new tools to unravel the complexities of plant-water interactions. Confocal microscopy offers high-resolution, 3D images of leaf tissues, focusing on cellular structures and stomatal dynamics [137]. Its advantage lies in cellular-level visualisation, but it is limited to small sample sizes and controlled environments. Thermal imaging with infrared thermography measures canopy temperature, indicating plant water status and transpiration rates. This method is rapid and non-invasive, yet sensitive to environmental fluctuations such as wind and ambient temperature [138,139]. X-ray micro-computed tomography (Micro-CT) and magnetic resonance imaging (MRI) provide non-destructive 3D imaging of leaf anatomy and water distribution. These offer exceptional anatomical insights but require costly equipment and may not be feasible for field-scale studies [140,141]. Chlorophyll fluorescence imaging, such as pulse amplitude modulation (PAM) fluorometry, assesses photosynthetic efficiency. While informative, its interpretation requires specialised knowledge, and results can vary depending on leaf age and environmental stress [142,143]. Field-based platforms use sensors and imaging systems on tractors [144], while greenhouses and growth chambers utilise automated systems to monitor conditions [145]. Unmanned aerial vehicles (UAVs) equipped with multispectral, hyperspectral, and thermal cameras capture data on plant health and water status from above [146]. UAVs enable rapid coverage of large areas but depend on weather and require data processing expertise [147]. Hyperspectral imaging captures detailed spectral information to assess physiological and biochemical traits [148]. Light detection and ranging (LIDAR) creates high-resolution 3D maps of plant structures, helping to evaluate leaf area, canopy architecture, and how these factors influence light interception, transpiration, and overall water use [149]. Both tools provide deep insights into spatial traits, though data complexity and processing time remain significant challenges.
In addition to the stable carbon isotopes in Δ13C, oxygen and hydrogen isotopes (δ18O) and δ2H) offer insights into water sources and usage dynamics [150]. Near-infrared spectroscopy (NIRS) can also analyse the chemical composition of plant tissues, which correlates with water content and WUE [151]. Raman spectroscopy analyses molecular compositions and structures to study biochemical changes in leaves [152]. These spectroscopic methods are fast and non-destructive but may require calibration against destructive sampling methods [153].
Genomic approaches like CRISPR-Cas9 and gene editing, along with transcriptomics, proteomics, and metabolomics, are powerful tools for studying WUE. CRISPR-Cas9 enables precise gene modifications to improve WUE, while transcriptomics and proteomics analyse gene expression and protein profiles to identify key WUE regulators [154]. Metabolomics examines the complete set of plant metabolites to understand metabolic pathways related to water use and stress responses [155]. These molecular approaches allow for targeted breeding and trait discovery, though they are resource-intensive and require validation under field conditions.
Big data and machine learning are transforming the study of WUE by using predictive modelling to combine environmental, genomic, and phenotypic data for optimising breeding strategies. Machine learning algorithms analyse large datasets to identify patterns affecting WUE, such as soil moisture and climate conditions [156]. Satellite imagery and geographic information systems (GIS) provide large-scale vegetation, soil, and climate data to monitor and manage WUE across regions. Smart irrigation systems utilise soil moisture sensors and weather forecasts to optimise water use and reduce waste [157]. While these digital tools offer scalability and precision, their effectiveness depends on data quality, infrastructure, and model accuracy.
Compared to traditional methods, such as manual trait measurements, gas exchange analysis using portable photosynthesis systems, or basic soil moisture probes, these emerging technologies offer greater speed, scale, precision, and data integration. However, they also demand higher technical expertise, infrastructure, and investment. A balanced application that combines traditional and emerging methods may provide the most comprehensive insight into rice physiology and WUE improvement [158].
5.2. Challenges in Developing Water-Efficient Rice Cultivars
Despite significant advances in our understanding of WUE in rice, the development of water-efficient cultivars faces several complex challenges. These obstacles span genetic, physiological, and agronomic domains, necessitating a multidisciplinary approach to breeding and crop improvement. Key challenges include the trade-off between improving WUE and potential reductions in biomass or yield. Genetic complexity is another challenge, as WUE is influenced by many genes and their interactions with the environment [71,168]. Additionally, the diversity of rice-growing ecosystems, from flooded paddies to rainfed fields, necessitates a multifaceted approach to breeding [169]. Therefore, variations in soil type, fertility, and water-holding capacity further complicate efforts to develop effective cultivars.
High-throughput phenotyping for WUE can be challenging due to the technical difficulty and cost of advanced technologies, which may not be accessible to all breeding programs, especially in developing countries. Additionally, traditional rice farming practices and the historical significance of rice cultivation may lead to resistance among traditional farmers to adopt new cultivars due to concerns about their performance or preference for established methods.
Implications for sustainable agriculture and climate resilience. The development of water-efficient rice cultivars has far-reaching implications that extend beyond immediate agricultural productivity, touching on global issues of food security, environmental sustainability, and climate change adaptation. By addressing these critical challenges, advances in rice WUE have the potential to transform agricultural practices and contribute significantly to sustainable development goals. Developing and using water-efficient rice cultivars can reduce irrigation needs, conserving water resources and supporting agriculture in water-scarce areas. This approach lowers energy requirements for water distribution, promotes sustainable farming, and ensures stable yields despite variable water availability, enhancing food security and farmer incomes, especially in drought-prone regions. By improving crop resilience to water stress, these cultivars help farmers adapt to climate change, minimize crop failure risks, and reduce economic losses. Additionally, efficient water use can decrease methane emissions from flooded fields and benefit local ecosystems by lessening the depletion of natural water bodies [1,170,171].
A comprehensive understanding of the interplay between stomatal and non-stomatal traits is vital for enhancing WUE in rice under climate variability. Stomatal traits directly influence transpiration and gas exchange, while non-stomatal traits; such as leaf morphology, cuticular wax load, and root architecture; contribute to water retention and drought adaptation. Together, these traits modulate the plant’s physiological response to biotic and abiotic stresses, offering complementary pathways to improve WUE without compromising yield potential. Integrating both trait types in breeding strategies supports the development of cultivars that are better adapted to diverse and water-limited environments, aligning with the broader goals of sustainable agriculture and climate resilience [13,91].
5.3. Interaction with Root Traits
Another important consideration that is often overlooked is that WUE in rice involves a complex interaction between leaf and root traits. Deeper rooting in rice genotypes increases WUE under drought conditions by reducing the internal concentration of CO2 (Ci) within the leaf, highlighting variability in root traits and gas exchange parameters among genotypes. Extensive root systems help plants access more water, but efficient stomatal control is vital to prevent excessive water loss. Roots signal leaves to adjust stomatal conductance based on soil water availability. Efficient WUE enhances root growth by providing more photosynthates, further improving water acquisition [11]. It was found that gs and transpiration rates decrease during soil drying before leaf water potential declines, indicating the importance of root signaling [172]. Hence, the interaction of leaf and root traits in enhancing WUE is also an underexplored area of study that warrants further investigation.
6. Towards Enhanced WUE in Rice
The concept map in Figure 3 encapsulates the complex nature of WUE in rice, highlighting the intricate relationships between stomatal and non-stomatal traits. This visual representation underscores the complexity of WUE as a trait and the need for integrated approaches in future research and breeding efforts. Stomatal traits such as density, size, and conductance interact closely with non-stomatal factors like leaf anatomy, photosynthetic efficiency, and metabolic adaptations. The integration and optimisation branch of the map emphasises the importance of considering trade-offs between these traits and leveraging emerging technologies for crop improvement. This holistic view guides future research directions, suggesting that advancements in WUE will likely come from a synergistic approach that combines traditional breeding with cutting-edge technologies like CRISPR gene editing and high-throughput phenotyping. The map also highlights the critical role of environmental interactions, reminding researchers of the need to develop rice varieties that maintain high WUE across diverse and changing climatic conditions.
Moving forward, the challenge lies in translating this complex understanding into practical breeding strategies and agronomic practices. Interdisciplinary collaboration will be key, bringing together expertise in plant physiology, genetics, agronomy, and data science. By addressing the interconnected aspects of WUE illustrated in this concept map, researchers can develop more resilient and water-efficient rice varieties, contributing significantly to sustainable agriculture in the face of global water scarcity and climate change.
7. Conclusion
There is an intricate relationship between stomatal and non-stomatal leaf traits and both collectively have an impact on WUE in rice. Understanding the complex interactions among these traits is essential for developing rice varieties with improved WUE, as well as for implementing agronomic practices that optimise water management in rice cultivation. Further investigation will reveal specific beneficial varieties for special agronomic practices. By focusing on both stomatal and non-stomatal leaf traits, researchers can develop climate-resilient rice varieties that ensure food security for future generations. Insights from this review can guide breeding programs to develop new crop varieties with enhanced WUE, ensuring sustainable production in water-scarce regions. Precision agriculture practices can be optimised based on specific leaf traits linked to efficient water use. The review offers a foundation for future research aimed at elucidating the mechanisms of WUE, facilitating the discovery of novel genetic targets and pathways.
Funding
This work was supported by funding from AgriFutures Australia Project No. PRP-013062 and PRO-016914.
Data Availability Statement
Data sharing is not applicable to this article as it is a review of existing literature. No new data were created or analysed.
Acknowledgments
We would like to express our gratitude to AgriFutures Australia for providing the stipend and funding this research project. We also thank Swinburne University of Technology for the Tuition Fee Scholarship. The three figures used in this publication are grahically illustrated by Samantha and Elisha Butardo.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| WUE | Water Use Efficiency |
| gs | stomatal conductance |
| Δ13C | carbon isotope discrimination |
| gm | mesophyll conductance |
References
- FAO. Water Scarcity – One of the greatest challenges of our time. 2019; Available from: https://www.fao.org/newsroom/story/Water-Scarcity-One-of-the-greatest-challenges-of-our-time/en.
- WHO. 1 in 3 people globally do not have access to safe drinking water – UNICEF, WHO. 2019 27/12/2022]; Available from: https://www.who.int/news/item/18-06-2019-1-in-3-people-globally-do-not-have-access-to-safe-drinking-water-unicef-who.
- Pitaloka, M.K., et al., Induced Genetic Variations in Stomatal Density and Size of Rice Strongly Affects Water Use Efficiency and Responses to Drought Stresses. Front Plant Sci, 2022. 13: p. 801706. [CrossRef]
- Bertolino, L.T., R.S. Caine, and J.E. Gray, Impact of Stomatal Density and Morphology on Water-Use Efficiency in a Changing World. Front Plant Sci, 2019. 10: p. 225. [CrossRef]
- Chapagain, T., A. Riseman, and E. Yamaji, Achieving more with less water: Alternate wet and dry irrigation (AWDI) as an alternative to the conventional water management practices in rice farming. Journal of Agricultural Science, 2011. 3(3): p. 3. [CrossRef]
- Mallareddy, M., et al., Maximizing water use efficiency in rice farming: A comprehensive review of innovative irrigation management technologies. Water, 2023. 15(10): p. 1802. [CrossRef]
- Zhang, Z., et al., Yield, grain quality and water use efficiency of rice under non-flooded mulching cultivation. Field Crops Research, 2008. 108(1): p. 71-81. [CrossRef]
- Borrell, A., A. Garside, and S. Fukai, Improving efficiency of water use for irrigated rice in a semi-arid tropical environment. Field Crops Research, 1997. 52(3): p. 231-248. [CrossRef]
- Fukai, S. and J. Mitchell, Factors determining water use efficiency in aerobic rice. Crop and Environment, 2022. 1(1): p. 24-40. [CrossRef]
- Gago, J., et al., Opportunities for improving leaf water use efficiency under climate change conditions. Plant Science, 2014. 226: p. 108-119. [CrossRef]
- Gobu, R., et al., Unlocking the Nexus between Leaf-Level Water Use Efficiency and Root Traits Together with Gas Exchange Measurements in Rice (Oryza sativa L.). Plants, 2022. 11(9): p. 1270. [CrossRef]
- Hatfield, J.L. and C. Dold, Water-Use Efficiency: Advances and Challenges in a Changing Climate. Front Plant Sci, 2019. 10: p. 103. [CrossRef]
- Ramachandra, A., et al., Decoding stomatal characteristics regulating water use efficiency at leaf and plant scales in rice genotypes. Planta, 2024. 260(3): p. 56. [CrossRef]
- Reddy, S.H., et al., Leaf mass area determines water use efficiency through its influence on carbon gain in rice mutants. Physiologia Plantarum, 2020. 169(2): p. 194-213. [CrossRef]
- Xiong, D. and J. Flexas, From one side to two sides: the effects of stomatal distribution on photosynthesis. New Phytol, 2020. 228(6): p. 1754-1766. [CrossRef]
- Akram, H., et al., Impact of water deficit stress on various physiological and agronomic traits of three basmati rice (Oryza sativa L.) cultivars. J Anim Plant Sci, 2013. 23(5): p. 1415-1423.
- Yadav, N., et al., Physiological response and agronomic performance of drought tolerance mutants of Aus rice cultivar Nagina 22 (Oryza sativa L). Field Crops Research, 2023. 290: p. 108760. [CrossRef]
- Condon, A.G., et al., Improving intrinsic water-use efficiency and crop yield. Crop science, 2002. 42(1): p. 122-131.
- Suleiman, S.O., et al., Grain yield and leaf gas exchange in upland NERICA rice under repeated cycles of water deficit at reproductive growth stage. Agricultural Water Management, 2022. 264: p. 107507. [CrossRef]
- Centritto, M., et al., Leaf gas exchange, carbon isotope discrimination, and grain yield in contrasting rice genotypes subjected to water deficits during the reproductive stage. J Exp Bot, 2009. 60(8): p. 2325-39. [CrossRef]
- Wang, Z., et al., Relationships between stable isotope natural abundances (δ13C and δ15N) and water use efficiency in rice under alternate wetting and drying irrigation in soils with high clay contents. Frontiers in Plant Science, 2022. 13: p. 1077152. [CrossRef]
- Wang, X., et al., Leaf rolling precedes stomatal closure in rice (Oryza sativa) under drought conditions. J Exp Bot, 2023. 74(21): p. 6650-6661. [CrossRef]
- Blankenagel, S., et al., Generating Plants with Improved Water Use Efficiency. Agronomy, 2018. 8(9): p. 194. [CrossRef]
- Bramley, H., N.C. Turner, and K.H.M. Siddique, Water Use Efficiency, in Genomics and Breeding for Climate-Resilient Crops: Vol. 2 Target Traits, C. Kole, Editor. 2013, Springer Berlin Heidelberg: Berlin, Heidelberg. p. 225-268.
- Brendel, O., The relationship between plant growth and water consumption: a history from the classical four elements to modern stable isotopes. Annals of Forest Science, 2021. 78(2): p. 47. [CrossRef]
- Petrík, P., et al., Leaf physiological and morphological constraints of water-use efficiency in C3 plants. AoB PLANTS, 2023. 15(4). [CrossRef]
- Pandya, P., et al., THERMAL IMAGING AND ITS APPLICATION IN IRRIGATION WATER MANAGEMENT. AgriTech Today, 2023: p. 40.
- Lang, P.L., et al., Century-long timelines of herbarium genomes predict plant stomatal response to climate change. Nature Ecology & Evolution, 2024: p. 1-13. [CrossRef]
- Mohammed, U., et al., IRice plants overexpressing OsEPF1 show reduced stomatal density and increased root cortical aerenchyma formation. Scientific reports, 2019. 9(1): p. 5584.
- Agurla S, et al., Mechanism of Stomatal Closure in Plants Exposed to Drought and Cold Stress. Adv Exp Med Biol., 2018. 1081: p. 215-232.
- Laza, M.R.C., et al., Quantitative trait loci for stomatal density and size in lowland rice. Euphytica, 2010. 172: p. 149-158. [CrossRef]
- Phunthong, C., et al., Rice mutants, selected under severe drought stress, show reduced stomatal density and improved water use efficiency under restricted water conditions. Frontiers in Plant Science, 2024. 15: p. 1307653. [CrossRef]
- Caine, R.S., et al., Rice with reduced stomatal density conserves water and has improved drought tolerance under future climate conditions. New Phytologist, 2019. 221(1): p. 371-384. [CrossRef]
- Ramachandra, A., B. PRABHU, and M. Sheshshayee, Relevance of Stomatal Traits in Determining the Water Use and Water Use Efficiency in Rice Genotypes Adapted to Different Cultivation Systems. Mysore Journal of Agricultural Sciences, 2023. 57(1).
- Malini, M., et al., Abscisic-Acid-Modulated Stomatal Conductance Governs High-Temperature Stress Tolerance in Rice Accessions. Agriculture, 2023. 13(3): p. 545. [CrossRef]
- Wang, Y., et al., Stomata conductance as a goalkeeper for increased photosynthetic efficiency. Current Opinion in Plant Biology, 2022. 70: p. 102310.
- Villalobos-López, M.A., et al., Biotechnological advances to improve abiotic stress tolerance in crops. International Journal of Molecular Sciences, 2022. 23(19): p. 12053. [CrossRef]
- Yari Kamrani, Y., et al., Regulatory role of circadian clocks on ABA production and signaling, stomatal responses, and water-use efficiency under water-deficit conditions. Cells, 2022. 11(7): p. 1154. [CrossRef]
- Hsiao, T.C., et al., Influence of osmotic adjustment on leaf rolling and tissue death in rice (Oryza sativa L.). Plant Physiology, 1984. 75(2): p. 338-341. [CrossRef]
- Wang, X., et al., Rice Responses to Abiotic Stress: Key Proteins and Molecular Mechanisms. International Journal of Molecular Sciences, 2025. 26(3): p. 896. [CrossRef]
- Singh, S., Guttation: Quantification, Microbiology and Implications for Phytopathology, in Progress in Botany: Vol. 75, U. Lüttge, W. Beyschlag, and J. Cushman, Editors. 2014, Springer Berlin Heidelberg: Berlin, Heidelberg. p. 187-214.
- Aloni, R., Leaf Development and Vascular Differentiation, in Vascular Differentiation and Plant Hormones, R. Aloni, Editor. 2021, Springer International Publishing: Cham. p. 141-162.
- Li, R., et al., Research progress in improving photosynthetic efficiency. International Journal of Molecular Sciences, 2023. 24(11): p. 9286. [CrossRef]
- Flexas, J., et al., Mesophyll diffusion conductance to CO2: an unappreciated central player in photosynthesis. Plant Sci, 2012. 193-194: p. 70-84. [CrossRef]
- Flexas, J., et al., Mesophyll conductance to CO2 and Rubisco as targets for improving intrinsic water use efficiency in C3 plants. Plant, Cell & Environment, 2016. 39(5): p. 965-982. [CrossRef]
- Ye, Z.-P., et al., Quantifying light response of leaf-scale water-use efficiency and its interrelationships with photosynthesis and stomatal conductance in C3 and C4 species. Frontiers in Plant Science, 2020. 11: p. 374. [CrossRef]
- Haworth, M., et al., Integrating stomatal physiology and morphology: evolution of stomatal control and development of future crops. Oecologia, 2021. 197(4): p. 867-883. [CrossRef]
- Terashima, I., et al., Leaf Functional Anatomy in Relation to Photosynthesis. Plant Physiology, 2010. 155(1): p. 108-116. [CrossRef]
- Kang, G., et al., Leaf microstructure and photosynthetic characteristics of a rice midvein-deficient mutant dl-14. Biologia plantarum, 2022. 66: p. 172-177. [CrossRef]
- Chen, J., et al., Aquaporins and CO2 diffusion across biological membrane. Frontiers in Physiology, 2023. 14: p. 1205290. [CrossRef]
- Ding, L., et al., Aquaporin Expression and Water Transport Pathways inside Leaves Are Affected by Nitrogen Supply through Transpiration in Rice Plants. International Journal of Molecular Sciences, 2018. 19(1): p. 256. [CrossRef]
- Raza, Q., et al., Genomic diversity of aquaporins across genus Oryza provides a rich genetic resource for development of climate resilient rice cultivars. BMC Plant Biology, 2023. 23(1): p. 172. [CrossRef]
- Nguyen, M.X., S. Moon, and K.-H. Jung, Genome-wide expression analysis of rice aquaporin genes and development of a functional gene network mediated by aquaporin expression in roots. Planta, 2013. 238(4): p. 669-681. [CrossRef]
- Flexas, J., et al., CO2 Diffusion Inside Photosynthetic Organs, in The Leaf: A Platform for Performing Photosynthesis, W.W. Adams Iii and I. Terashima, Editors. 2018, Springer International Publishing: Cham. p. 163-208.
- Oguchi, R., et al., Leaf Anatomy and Function, in The Leaf: A Platform for Performing Photosynthesis, W.W. Adams Iii and I. Terashima, Editors. 2018, Springer International Publishing: Cham. p. 97-139.
- Takahashi, H. and M. Nakazono, Cavity Tissue for the Internal Aeration in Plants, in Responses of Plants to Soil Flooding, J.-I. Sakagami and M. Nakazono, Editors. 2024, Springer Nature Singapore: Singapore. p. 105-117.
- Muir, C.D., Making pore choices: repeated regime shifts in stomatal ratio. Proceedings of the Royal Society B: Biological Sciences, 2015. 282(1813): p. 20151498. [CrossRef]
- Wang, X., et al., A β-Ketoacyl-CoA Synthase Is Involved in Rice Leaf Cuticular Wax Synthesis and Requires a CER2-LIKE Protein as a Cofactor. Plant Physiology, 2016. 173(2): p. 944-955. [CrossRef]
- Gao, X., et al., A β-ketoacyl-CoA Synthase OsCUT1 Confers Increased Drought Tolerance in Rice. Rice Science, 2022. 29(4): p. 353-362. [CrossRef]
- Qin, B.-X., et al., Rice OsGL1-1 is involved in leaf cuticular wax and cuticle membrane. Molecular Plant, 2011. 4(6): p. 985-995. [CrossRef]
- Bernaola, L., et al., Epicuticular Wax Rice Mutants Show Reduced Resistance to Rice Water Weevil (Coleoptera: Curculionidae) and Fall Armyworm (Lepidoptera: Noctuidae). Environmental Entomology, 2021. 50(4): p. 948-957. [CrossRef]
- Pradhan, A.K., et al., Chapter 26 - Comparative Metabolomics Approach Towards Understanding Chemical Variation in Rice Under Abiotic Stress, in Advances in Rice Research for Abiotic Stress Tolerance, M. Hasanuzzaman, et al., Editors. 2019, Woodhead Publishing. p. 537-550.
- Hassanein, A., et al., Differential metabolic responses associated with drought tolerance in egyptian rice. J. Appl. Biol. Biotechnol, 2021. 9(4): p. 37-46.
- Jan, R., et al., Drought and UV radiation stress tolerance in rice is improved by overaccumulation of non-enzymatic antioxidant flavonoids. Antioxidants, 2022. 11(5): p. 917. [CrossRef]
- Pons, T.L., et al., Estimating mesophyll conductance to CO2: methodology, potential errors, and recommendations. Journal of Experimental Botany, 2009. 60(8): p. 2217-2234. [CrossRef]
- von Caemmerer, S. and J.R. Evans, Enhancing C3 photosynthesis. Plant Physiology, 2010. 154(2): p. 589-592.
- Flexas, J., et al., Diffusional conductances to CO 2 as a target for increasing photosynthesis and photosynthetic water-use efficiency. Photosynthesis research, 2013. 117: p. 45-59. [CrossRef]
- Ouyang, W., et al., Stomatal conductance, mesophyll conductance, and transpiration efficiency in relation to leaf anatomy in rice and wheat genotypes under drought. Journal of Experimental Botany, 2017. 68(18): p. 5191-5205. [CrossRef]
- Zait, Y., Á. Ferrero-Serrano, and S.M. Assmann, The α subunit of the heterotrimeric G protein regulates mesophyll CO2 conductance and drought tolerance in rice. New Phytologist, 2021. 232(6): p. 2324-2338. [CrossRef]
- Ye, M., et al., High leaf mass per area Oryza genotypes invest more leaf mass to cell wall and show a low mesophyll conductance. AoB PLANTS, 2020. 12(4). [CrossRef]
- This, D., et al., Genetic analysis of water use efficiency in rice (Oryza sativa L.) at the leaf level. Rice Science, 2010. 3(1): p. 72-86. [CrossRef]
- Baloch, A., et al., Optimum plant density for high yield in rice (Oryza sativa L.). Asian J. Plant Sci, 2002. 1(1): p. 25-27. [CrossRef]
- Hamaoka, N., et al., A hairy-leaf gene, BLANKET LEAF, of wild Oryza nivara increases photosynthetic water use efficiency in rice. Rice (N Y), 2017. 10(1): p. 20. [CrossRef]
- Zhang, Y., et al., Modeling the leaf angle dynamics in rice plant. PLoS One, 2017. 12(2): p. e0171890. [CrossRef]
- Luo, X., et al., Phytohormones signaling and crosstalk regulating leaf angle in rice. Plant cell reports, 2016. 35: p. 2423-2433. [CrossRef]
- Chang, T.-G., et al., A three-dimensional canopy photosynthesis model in rice with a complete description of the canopy architecture, leaf physiology, and mechanical properties. Journal of Experimental Botany, 2019. 70(9): p. 2479-2490. [CrossRef]
- Burgess, A.J., et al., Exploring relationships between canopy architecture, light distribution, and photosynthesis in contrasting rice genotypes using 3D canopy reconstruction. Frontiers in plant science, 2017. 8: p. 734. [CrossRef]
- T, S. and D. Vijayalakshmi, Physiological and biotechnological approach to improve water use efficiency in rice (Oryza sativa L.): Review. Journal of Pharmacognosy and Phytochemistry, 2020. 9(5): p. 1044-1048. [CrossRef]
- Farquhar, G.D., J.R. Ehleringer, and K.T. Hubick, Carbon isotope discrimination and photosynthesis. Annual review of plant physiology and plant molecular biology, 1989. 40(1): p. 503-537.
- Condon, A.G., et al., Breeding for high water-use efficiency. J Exp Bot, 2004. 55(407): p. 2447-60. [CrossRef]
- Farquhar, G. and R. Richards, Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Functional Plant Biology, 1984. 11(6): p. 539-552. [CrossRef]
- Impa, S.M., et al., Carbon Isotope Discrimination Accurately Reflects Variability in WUE Measured at a Whole Plant Level in Rice. Crop Science, 2005. 45(6): p. 2517-2522. [CrossRef]
- Qu, M., et al., Rapid stomatal response to fluctuating light: an under-explored mechanism to improve drought tolerance in rice. Functional Plant Biology, 2016. 43(8): p. 727-738. [CrossRef]
- Wu, A., et al., Quantifying impacts of enhancing photosynthesis on crop yield. Nature plants, 2019. 5(4): p. 380-388. [CrossRef]
- Raju, B.R., et al., Discovery of QTLs for water mining and water use efficiency traits in rice under water-limited condition through association mapping. Molecular Breeding, 2016. 36(3): p. 35. [CrossRef]
- Vinarao, R., et al., QTL Validation and Development of SNP-Based High Throughput Molecular Markers Targeting a Genomic Region Conferring Narrow Root Cone Angle in Aerobic Rice Production Systems. Plants (Basel), 2021. 10(10): p. 2099. [CrossRef]
- Yue, B., et al., QTL Analysis for Flag Leaf Characteristics and Their Relationships with Yield and Yield Traits in Rice. Acta Genetica Sinica, 2006. 33(9): p. 824-832. [CrossRef]
- Hoang, G.T., et al., Genome-wide association mapping of leaf mass traits in a Vietnamese rice landrace panel. PLoS One, 2019. 14(7): p. e0219274. [CrossRef]
- Phetluan, W., et al., Candidate genes affecting stomatal density in rice (Oryza sativa L.) identified by genome-wide association. Plant Science, 2023. 330: p. 111624. [CrossRef]
- Yang, W., et al., Genome-wide association study of rice (Oryza sativa L.) leaf traits with a high-throughput leaf scorer. Journal of experimental botany, 2015. 66(18): p. 5605-5615.
- Liu, C., et al., Relationships of stomatal morphology to the environment across plant communities. Nature Communications, 2023. 14(1): p. 6629. [CrossRef]
- Zhang, Y., et al., Water use efficiency and physiological response of rice cultivars under alternate wetting and drying conditions. ScientificWorldJournal, 2012. 2012: p. 287907. [CrossRef]
- NSW-DPI, DPI Primefact, B. Dunn, Editor. 2023.
- Haonan, Q., et al., Current status of global rice water use efficiency and water-saving irrigation technology recommendations. Journal of Agronomy and Crop Science, 2023. 209(5): p. 734-746. [CrossRef]
- Farmonaut. Revolutionizing Australian rice farming: Sustainable water-saving technologies for enhanced productivity. 2024; Available from: https://farmonaut.com/australia/revolutionizing-australian-rice-farming-sustainable-water-saving-technologies-for-enhanced-productivity/.
- 2, S.I.f.P.P., Smart irrigation control in rice growing systems: Application in bankless channel irrigation. 2021.
- Authority., M.D.B. New technology helping rice growers use less water. 2025; Available from: https://www.mdba.gov.au/news-and-events/newsroom/new-technology-helping-rice-growers-use-less-water.
- Thakur, A., et al., Evaluation of planting methods in irrigated rice. Archives of Agronomy and Soil Science, 2004. 50(6): p. 631-640. [CrossRef]
- Sangavi, S. and S. Porpavai, Impact of irrigation scheduling and weed management on water use efficiency and yield of direct dry seeded rice. Madras Agricultural Journal, 2018. 105(march (1-3)): p. 1. [CrossRef]
- IRRI. SNP-Seek database.; Available from: https://snp-seek.irri.org/.
- Yu, H., et al., Decreased photosynthesis in the erect panicle 3 (ep3) mutant of rice is associated with reduced stomatal conductance and attenuated guard cell development. Journal of Experimental Botany, 2015. 66(5): p. 1543-1552. [CrossRef]
- Hwang, H., et al., Unique features of two potassium channels, OsKAT2 and OsKAT3, expressed in rice guard cells. PLoS One, 2013. 8(8): p. e72541. [CrossRef]
- Moon, S.-J., et al., A dominant negative OsKAT2 mutant delays light-induced stomatal opening and improves drought tolerance without yield penalty in rice. Frontiers in Plant Science, 2017. 8: p. 772. [CrossRef]
- Wu, Z., et al., Multiple transcriptional factors control stomata development in rice. New Phytologist, 2019. 223(1): p. 220-232. [CrossRef]
- Chen, H., et al., Genetic bases of the stomata-related traits revealed by a genome-wide association analysis in rice (Oryza sativa L.). Frontiers in genetics, 2020. 11: p. 611. [CrossRef]
- Yang, W., et al., Genome-wide association study of rice (Oryza sativa L.) leaf traits with a high-throughput leaf scorer. J Exp Bot, 2015. 66(18): p. 5605-15.
- Nguyen, V.N., et al., OsABCG9 is an important ABC transporter of cuticular wax deposition in rice. Frontiers in Plant Science, 2018. 9: p. 960. [CrossRef]
- Islam, M.A., et al., Characterization of Glossy1-homologous genes in rice involved in leaf wax accumulation and drought resistance. Plant Mol Biol, 2009. 70(4): p. 443-56. [CrossRef]
- Wang, Y., et al., An ethylene response factor OsWR1 responsive to drought stress transcriptionally activates wax synthesis related genes and increases wax production in rice. Plant Mol Biol, 2012. 78(3): p. 275-88. [CrossRef]
- Sheng, Z., et al., Yellow-Leaf 1 encodes a magnesium-protoporphyrin IX monomethyl ester cyclase, involved in chlorophyll biosynthesis in rice (Oryza sativa L.). PLoS One, 2017. 12(5): p. e0177989. [CrossRef]
- Li, C. and B. Zhang, MicroRNAs in control of plant development. Journal of cellular physiology, 2016. 231(2): p. 303-313. [CrossRef]
- Wang, Y., et al., RICE CENTRORADIALIS 1, a TFL1-like Gene, Responses to Drought Stress and Regulates Rice Flowering Transition. Rice (N Y), 2020. 13(1): p. 70. [CrossRef]
- Li, M., et al., Mutations in the F-box gene LARGER PANICLE improve the panicle architecture and enhance the grain yield in rice. Plant biotechnology journal, 2011. 9(9): p. 1002-1013. [CrossRef]
- Sakamoto, T., Phytohormones and rice crop yield: strategies and opportunities for genetic improvement. Transgenic Res, 2006. 15(4): p. 399-404. [CrossRef]
- Ishiwata, A., et al., Two WUSCHEL-related homeobox genes, narrow leaf2 and narrow leaf3, control leaf width in rice. Plant Cell Physiol, 2013. 54(5): p. 779-92. [CrossRef]
- Hu, J., et al., Identification and characterization of NARROW AND ROLLED LEAF 1, a novel gene regulating leaf morphology and plant architecture in rice. Plant Molecular Biology, 2010. 73(3): p. 283-292. [CrossRef]
- Sonah, H., et al., Molecular mapping of quantitative trait loci for flag leaf length and other agronomic traits in rice (Oryza sativa). Cereal Research Communications, 2012. 40(3): p. 362-372. [CrossRef]
- Miura, K., et al., OsSPL14 promotes panicle branching and higher grain productivity in rice. Nat Genet, 2010. 42(6): p. 545-9. [CrossRef]
- Hu, L., et al., OsSPL9 Regulates Grain Number and Grain Yield in Rice. Front Plant Sci, 2021. 12: p. 682018. [CrossRef]
- Yu, D., et al., Wax Crystal-Sparse Leaf1 encodes a β–ketoacyl CoA synthase involved in biosynthesis of cuticular waxes on rice leaf. Planta, 2008. 228(4): p. 675-685. [CrossRef]
- Tang, Y., et al., Overexpression of a MYB Family Gene, OsMYB6, Increases Drought and Salinity Stress Tolerance in Transgenic Rice. Front Plant Sci, 2019. 10: p. 168. [CrossRef]
- Sakurai, J., et al., Identification of 33 rice aquaporin genes and analysis of their expression and function. Plant and Cell Physiology, 2005. 46(9): p. 1568-1577. [CrossRef]
- Zhou, Y., et al., Identification and functional characterization of a rice NAC gene involved in the regulation of leaf senescence. BMC Plant Biology, 2013. 13: p. 1-13. [CrossRef]
- Matsukura, S., et al., Comprehensive analysis of rice DREB2-type genes that encode transcription factors involved in the expression of abiotic stress-responsive genes. Molecular Genetics and Genomics, 2010. 283: p. 185-196. [CrossRef]
- Xiang, Y., et al., Characterization of OsbZIP23 as a key player of the basic leucine zipper transcription factor family for conferring abscisic acid sensitivity and salinity and drought tolerance in rice. Plant physiology, 2008. 148(4): p. 1938-1952. [CrossRef]
- Wu, R., et al., CFL1, a WW Domain Protein, Regulates Cuticle Development by Modulating the Function of HDG1, a Class IV Homeodomain Transcription Factor, in Rice and Arabidopsis The Plant Cell, 2011. 23(9): p. 3392-3411. [CrossRef]
- Guo, T., et al., Mutations in the rice OsCHR4 gene, encoding a CHD3 family chromatin remodeler, induce narrow and rolled leaves with increased cuticular wax. International Journal of Molecular Sciences, 2019. 20(10): p. 2567. [CrossRef]
- Cai, S., et al., A key ABA catabolic gene, OsABA8ox3, is involved in drought stress resistance in rice. PLoS One, 2015. 10(2): p. e0116646. [CrossRef]
- Jardim-Messeder, D., et al., Stromal Ascorbate Peroxidase (OsAPX7) Modulates Drought Stress Tolerance in Rice (Oryza sativa). Antioxidants, 2023. 12(2): p. 387. [CrossRef]
- Li, H.-W., et al., Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta, 2011. 234(5): p. 1007-1018. [CrossRef]
- Cabangon, R.J., et al., Effect of irrigation method and N-fertilizer management on rice yield, water productivity and nutrient-use efficiencies in typical lowland rice conditions in China. Paddy and Water Environment, 2004. 2(4): p. 195-206. [CrossRef]
- He, Q., et al., Released control of vapor pressure deficit on rainfed rice evapotranspiration responses to extreme droughts in the subtropical zone. Plant and Soil, 2024: p. 1-19. [CrossRef]
- Haque, M.M., D.J. Mackill, and K.T. Ingram, Inheritance of Leaf Epicuticular Wax Content in Rice. Crop Science, 1992. 32(4): p. 865-868. [CrossRef]
- Wang, C., et al., Comparison of Agronomic and Physiological Characteristics for Rice Varieties Differing in Water Use Efficiency under Alternate Wetting and Drying Irrigation. Agronomy, 2024. 14(9): p. 1986. [CrossRef]
- Gao, H., et al., Biodegradable film mulching reduces the climate cost of saving water without yield penalty in dryland rice production. Resources, Conservation and Recycling, 2023. 197: p. 107071. [CrossRef]
- Malumpong, C., et al., Alternate Wetting and Drying (AWD) in broadcast rice (Oryza sativa L.) management to maintain yield, conserve water, and reduce gas emissions in Thailand. Agricultural Research, 2021. 10: p. 116-130. [CrossRef]
- Omasa, K., et al., 3D Confocal laser scanning microscopy for the analysis of chlorophyll fluorescence parameters of chloroplasts in intact leaf tissues. Plant Cell Physiol, 2009. 50(1): p. 90-105. [CrossRef]
- Luan, Y., et al., Improving the performance in crop water deficit diagnosis with canopy temperature spatial distribution information measured by thermal imaging. Agricultural Water Management, 2021. 246: p. 106699. [CrossRef]
- Jones, H.G., et al., Thermal infrared imaging of crop canopies for the remote diagnosis and quantification of plant responses to water stress in the field. Functional Plant Biology, 2009. 36(11): p. 978-989. [CrossRef]
- Mathers, A.W., et al., Investigating the microstructure of plant leaves in 3D with lab-based X-ray computed tomography. Plant Methods, 2018. 14: p. 99. [CrossRef]
- Van As, H., Intact plant MRI for the study of cell water relations, membrane permeability, cell-to-cell and long distance water transport. Journal of Experimental Botany, 2007. 58(4): p. 743-756.
- Gotarkar, D., et al., High-Throughput Analysis of Non-Photochemical Quenching in Crops Using Pulse Amplitude Modulated Chlorophyll Fluorometry. J Vis Exp, 2022(185): p. e63485.
- Baker, N.R., Chlorophyll fluorescence: a probe of photosynthesis in vivo. Annu. Rev. Plant Biol., 2008. 59(1): p. 89-113. [CrossRef]
- Thorp, K.R., et al., High-throughput phenotyping of crop water use efficiency via multispectral drone imagery and a daily soil water balance model. Remote Sensing, 2018. 10(11): p. 1682. [CrossRef]
- Langstroff, A., et al., Opportunities and limits of controlled-environment plant phenotyping for climate response traits. Theor Appl Genet, 2022. 135(1): p. 1-16. [CrossRef]
- Fullana-Pericàs, M., et al., High-throughput phenotyping of a large tomato collection under water deficit: Combining UAVs’ remote sensing with conventional leaf-level physiologic and agronomic measurements. Agricultural Water Management, 2022. 260: p. 107283. [CrossRef]
- Berni, J.A., et al., Thermal and narrowband multispectral remote sensing for vegetation monitoring from an unmanned aerial vehicle. IEEE Transactions on geoscience and Remote Sensing, 2009. 47(3): p. 722-738. [CrossRef]
- Asaari, M.S.M., et al., Non-destructive analysis of plant physiological traits using hyperspectral imaging: A case study on drought stress. Computers and Electronics in Agriculture, 2022. 195: p. 106806. [CrossRef]
- Omasa, K., F. Hosoi, and A. Konishi, 3D lidar imaging for detecting and understanding plant responses and canopy structure. J Exp Bot, 2007. 58(4): p. 881-98. [CrossRef]
- Zhang, M., et al., Effects of Water Isotope Composition on Stable Isotope Distribution and Fractionation of Rice and Plant Tissues. Journal of Agricultural and Food Chemistry, 2024. 72(16): p. 8955-8962. [CrossRef]
- Wang, M., et al., Evaluation of water-use efficiency in foxtail millet (Setaria italica) using visible-near infrared and thermal spectral sensing techniques. Talanta, 2016. 152: p. 531-9. [CrossRef]
- Pérez-Bueno, M.L., M. Pineda, and M. Barón, Phenotyping plant responses to biotic stress by chlorophyll fluorescence imaging. Frontiers in plant science, 2019. 10: p. 477268. [CrossRef]
- Schulz, H. and M. Baranska, Identification and quantification of valuable plant substances by IR and Raman spectroscopy. Vibrational Spectroscopy, 2007. 43(1): p. 13-25. [CrossRef]
- Rathnasamy, S.A., et al., Altering Stomatal Density for Manipulating Transpiration and Photosynthetic Traits in Rice through CRISPR/Cas9 Mutagenesis. Current Issues in Molecular Biology, 2023. 45(5): p. 3801-3814. [CrossRef]
- Kosová, K., et al., Drought stress response in common wheat, durum wheat, and barley: transcriptomics, proteomics, metabolomics, physiology, and breeding for an enhanced drought tolerance. Drought stress tolerance in plants, vol 2: molecular and genetic perspectives, 2016: p. 277-314.
- Sharma, N., et al., Data-driven approaches to improve water-use efficiency and drought resistance in crop plants. Plant Sci, 2023. 336: p. 111852. [CrossRef]
- Mo, X., et al., Prediction of crop yield, water consumption and water use efficiency with a SVAT-crop growth model using remotely sensed data on the North China Plain. Ecological Modelling, 2005. 183(2-3): p. 301-322. [CrossRef]
- Araus, J.L. and J.E. Cairns, Field high-throughput phenotyping: the new crop breeding frontier. Trends in plant science, 2014. 19(1): p. 52-61. [CrossRef]
- Huang, L., et al., Comparative transcriptome sequencing of tolerant rice introgression line and its parents in response to drought stress. BMC genomics, 2014. 15: p. 1-16. [CrossRef]
- Lenka, S.K., et al., Comparative analysis of drought-responsive transcriptome in Indica rice genotypes with contrasting drought tolerance. Plant biotechnology journal, 2011. 9(3): p. 315-327.
- Chung, P.J., et al., Transcriptome profiling of drought responsive noncoding RNAs and their target genes in rice. BMC Genomics, 2016. 17(1): p. 563. [CrossRef]
- Wu, X., et al., Advances in crop proteomics: PTMs of proteins under abiotic stress. Proteomics, 2016. 16(5): p. 847-865. [CrossRef]
- Lawas, L.M.F., et al., Metabolic responses of rice cultivars with different tolerance to combined drought and heat stress under field conditions. GigaScience, 2019. 8(5). [CrossRef]
- Liang, Y., et al., Transcriptome profiling of two rice genotypes under mild field drought stress during grain-filling stage. AoB PLANTS, 2021. 13(4). [CrossRef]
- Zargar, S.M., et al., Physiological and multi-omics approaches for explaining drought stress tolerance and supporting sustainable production of rice. Frontiers in Plant Science, 2022. 12: p. 803603. [CrossRef]
- Roy, N., P. Debnath, and H.S. Gaur, Adoption of Multi-omics Approaches to Address Drought Stress Tolerance in Rice and Mitigation Strategies for Sustainable Production. Molecular Biotechnology, 2025: p. 1-13. [CrossRef]
- Cao, J., et al., Integrating multi-source data for rice yield prediction across China using machine learning and deep learning approaches. Agricultural and Forest Meteorology, 2021. 297: p. 108275. [CrossRef]
- Xu, H., et al., Detection of epistatic and gene-environment interactions underlying three quality traits in rice using high-throughput genome-wide data. BioMed Research International, 2015. 2015(1): p. 135782.
- Bouman, B.A., et al., Rice: feeding the billions. 2007.
- Singh, N., D. Patel, and G. Khalekar, Methanogenesis and methane emission in rice/paddy fields. Sustainable Agriculture Reviews 33: Climate Impact on Agriculture, 2018: p. 135-170.
- IRRI. Rice to zero hunger. 2019 2023.10.05]; Available from: https://www.irri.org/world-food-day-2019-rice-zero-hunger.
- Siopongco, J.D., et al., Stomatal responses in rainfed lowland rice to partial soil drying; evidence for root signals. Plant Production Science, 2008. 11(1): p. 28-41. [CrossRef]
Figure 1.
Anatomical structures and physiological mechanisms regulating water use efficiency in rice leaves. (A) Ion transport mechanisms in guard and subsidiary cells during stomatal dynamics. The left panel illustrates stomatal opening, initiated by activation of plasma membrane H⁺-ATPases, which drive H⁺ efflux and establish a membrane potential that facilitates the influx of K⁺, Cl⁻, and water (H₂O), resulting in increased guard cell turgor and pore opening. Malate²⁻ is indicated within the guard cell as a key osmotic anion. Right panel shows stomatal closing with efflux of ions, leading to guard cell shrinkage and reduced aperture. (B) Carbon dioxide diffusion pathway from atmosphere to chloroplast stroma, showing CO₂ movement through the boundary layer, plasma membrane, cell wall, intercellular air space, and chloroplast envelope. CO₂-permeable aquaporins in the plasma membrane facilitate efficient CO₂ transport, enhancing mesophyll conductance for photosynthesis. (C) Cross-sectional view of leaf cuticle architecture showing the multilayer barrier structure: outer epicuticular wax crystals, cuticle proper, cuticular layer, and underlying epidermal cell. This waxy barrier regulates non-stomatal water loss and provides protection against environmental stress. (D) Comparison of leaf surface boundary layer effects on temperature gradients and water vapour (H₂O) diffusion. Upper panel demonstrates how leaf pubescence (trichomes) creates a thicker boundary layer, increasing the temperature gradient and slowing water vapour diffusion, thereby improving water conservation and thermal regulation. Lower panel shows smooth leaf surface with thinner boundary layer and steeper temperature gradient from leaf surface to ambient air.
Figure 1.
Anatomical structures and physiological mechanisms regulating water use efficiency in rice leaves. (A) Ion transport mechanisms in guard and subsidiary cells during stomatal dynamics. The left panel illustrates stomatal opening, initiated by activation of plasma membrane H⁺-ATPases, which drive H⁺ efflux and establish a membrane potential that facilitates the influx of K⁺, Cl⁻, and water (H₂O), resulting in increased guard cell turgor and pore opening. Malate²⁻ is indicated within the guard cell as a key osmotic anion. Right panel shows stomatal closing with efflux of ions, leading to guard cell shrinkage and reduced aperture. (B) Carbon dioxide diffusion pathway from atmosphere to chloroplast stroma, showing CO₂ movement through the boundary layer, plasma membrane, cell wall, intercellular air space, and chloroplast envelope. CO₂-permeable aquaporins in the plasma membrane facilitate efficient CO₂ transport, enhancing mesophyll conductance for photosynthesis. (C) Cross-sectional view of leaf cuticle architecture showing the multilayer barrier structure: outer epicuticular wax crystals, cuticle proper, cuticular layer, and underlying epidermal cell. This waxy barrier regulates non-stomatal water loss and provides protection against environmental stress. (D) Comparison of leaf surface boundary layer effects on temperature gradients and water vapour (H₂O) diffusion. Upper panel demonstrates how leaf pubescence (trichomes) creates a thicker boundary layer, increasing the temperature gradient and slowing water vapour diffusion, thereby improving water conservation and thermal regulation. Lower panel shows smooth leaf surface with thinner boundary layer and steeper temperature gradient from leaf surface to ambient air.
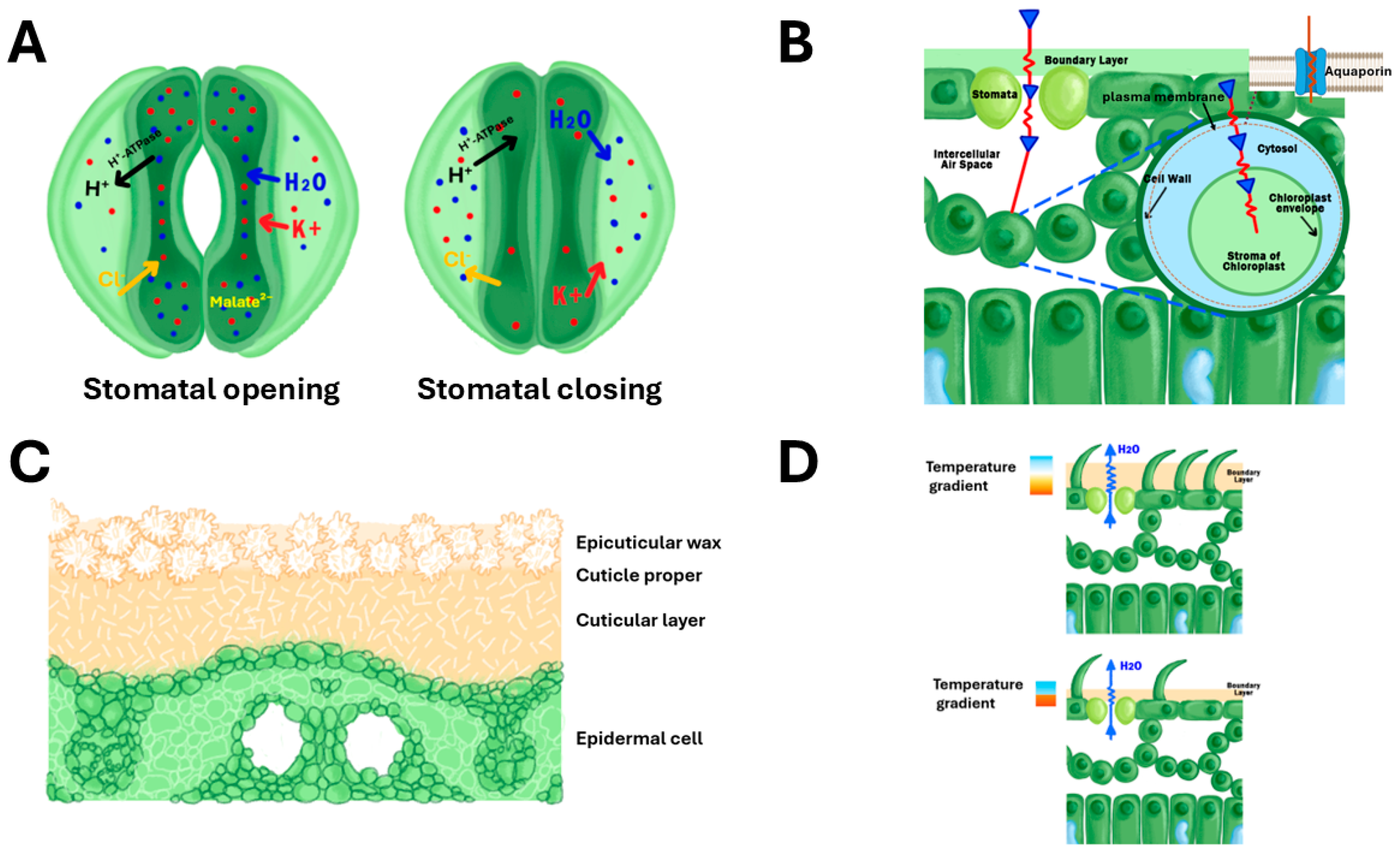
Figure 2.
Comparative analysis of physiological adaptations and management strategies in two distinct rice production systems. The diagram contrasts traditional irrigated paddy systems (left) operating under anaerobic, flooded conditions with dryland/upland systems (right) under aerobic, water-limited conditions. Traditional paddy system features shallow root architecture with aerenchyma tissue for oxygen transport under waterlogged conditions, higher stomatal conductance with reduced sensitivity to vapour pressure deficit (VPD), minimal epicuticular wax deposition, and water depth management through alternate wetting and drying practices. Dryland/upland system exhibits deeper, more extensive root systems for enhanced soil moisture capture, responsive stomatal regulation with higher sensitivity to soil moisture deficits, increased epicuticular wax accumulation and leaf rolling for water conservation, enhanced osmoprotectant production, and water-conserving management practices including mulching and deficit irrigation strategies. Water droplets illustrate different soil moisture distribution patterns characteristic of each system.
Figure 2.
Comparative analysis of physiological adaptations and management strategies in two distinct rice production systems. The diagram contrasts traditional irrigated paddy systems (left) operating under anaerobic, flooded conditions with dryland/upland systems (right) under aerobic, water-limited conditions. Traditional paddy system features shallow root architecture with aerenchyma tissue for oxygen transport under waterlogged conditions, higher stomatal conductance with reduced sensitivity to vapour pressure deficit (VPD), minimal epicuticular wax deposition, and water depth management through alternate wetting and drying practices. Dryland/upland system exhibits deeper, more extensive root systems for enhanced soil moisture capture, responsive stomatal regulation with higher sensitivity to soil moisture deficits, increased epicuticular wax accumulation and leaf rolling for water conservation, enhanced osmoprotectant production, and water-conserving management practices including mulching and deficit irrigation strategies. Water droplets illustrate different soil moisture distribution patterns characteristic of each system.
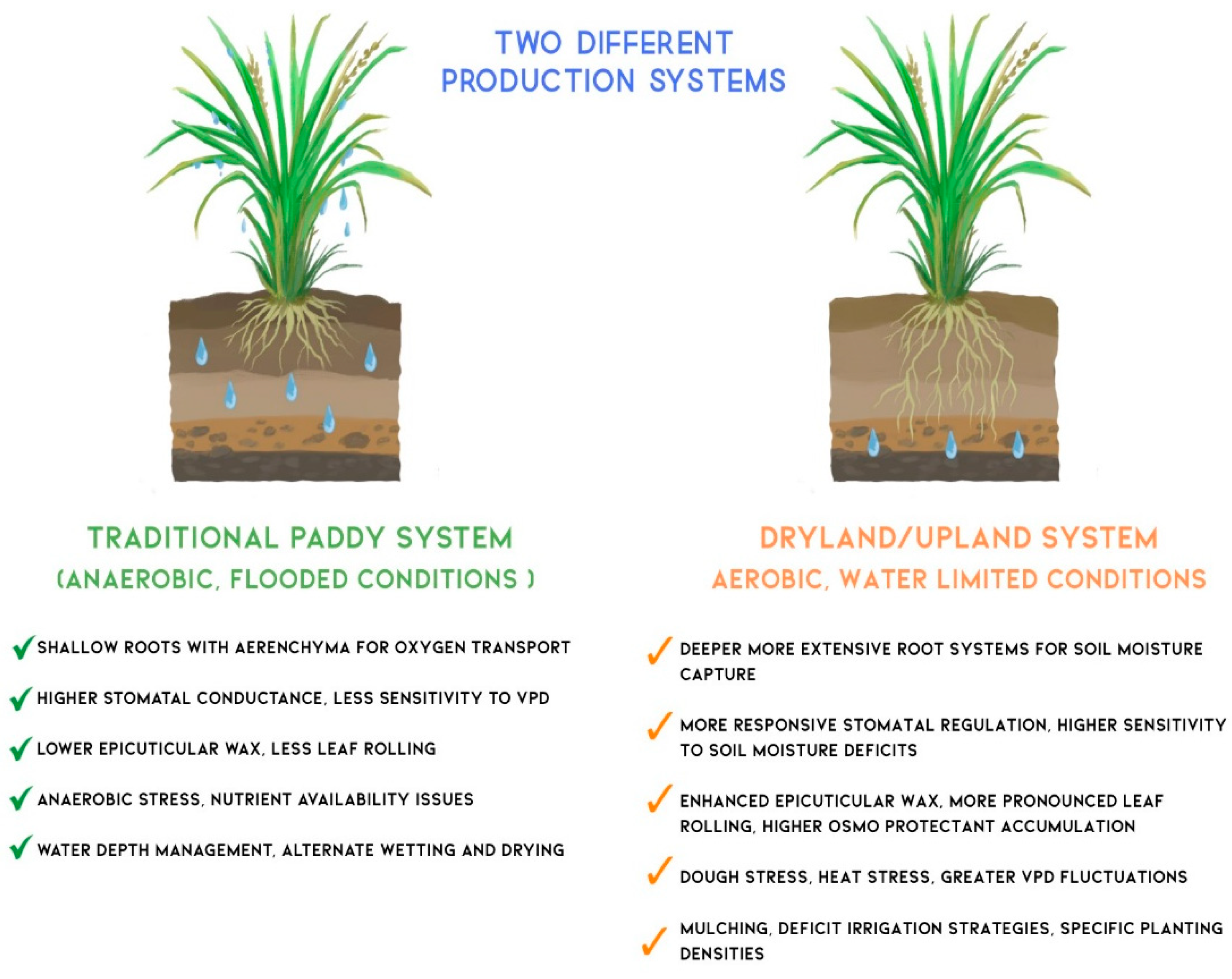
Figure 3.
Conceptual framework illustrating the integrated approach to water use efficiency (WUE) improvement in rice through stomatal and non-stomatal leaf traits. The diagram demonstrates how stomatal leaf traits (left side: stomatal conductance, density and size, ABA signaling pathways, and aperture control mechanisms) and non-stomatal leaf traits (right side: photosynthetic efficiency, leaf cuticle and epicuticular wax deposition, mesophyll conductance, metabolic adaptations, and leaf anatomy) both contribute to overall WUE in rice. The bidirectional arrow between trait categories indicates their interactive and complementary nature in determining water use efficiency. The central integration and optimisation component emphasizes the requirement for holistic approaches incorporating emerging technologies (multi-omics, high-throughput phenotyping), environmental interactions (genotype × environment effects), trade-offs between traits (balancing water conservation with carbon assimilation), and strategic breeding approaches (marker-assisted selection, genomic selection) to develop water-efficient rice cultivars. The rice plants represent the two major cultivation systems (traditional paddy and dryland/upland) discussed throughout the review, highlighting that WUE optimisation strategies must be adapted to specific production environments. This framework provides a roadmap for researchers and breeders to systematically approach WUE improvement through coordinated manipulation of multiple physiological and anatomical traits.
Figure 3.
Conceptual framework illustrating the integrated approach to water use efficiency (WUE) improvement in rice through stomatal and non-stomatal leaf traits. The diagram demonstrates how stomatal leaf traits (left side: stomatal conductance, density and size, ABA signaling pathways, and aperture control mechanisms) and non-stomatal leaf traits (right side: photosynthetic efficiency, leaf cuticle and epicuticular wax deposition, mesophyll conductance, metabolic adaptations, and leaf anatomy) both contribute to overall WUE in rice. The bidirectional arrow between trait categories indicates their interactive and complementary nature in determining water use efficiency. The central integration and optimisation component emphasizes the requirement for holistic approaches incorporating emerging technologies (multi-omics, high-throughput phenotyping), environmental interactions (genotype × environment effects), trade-offs between traits (balancing water conservation with carbon assimilation), and strategic breeding approaches (marker-assisted selection, genomic selection) to develop water-efficient rice cultivars. The rice plants represent the two major cultivation systems (traditional paddy and dryland/upland) discussed throughout the review, highlighting that WUE optimisation strategies must be adapted to specific production environments. This framework provides a roadmap for researchers and breeders to systematically approach WUE improvement through coordinated manipulation of multiple physiological and anatomical traits.
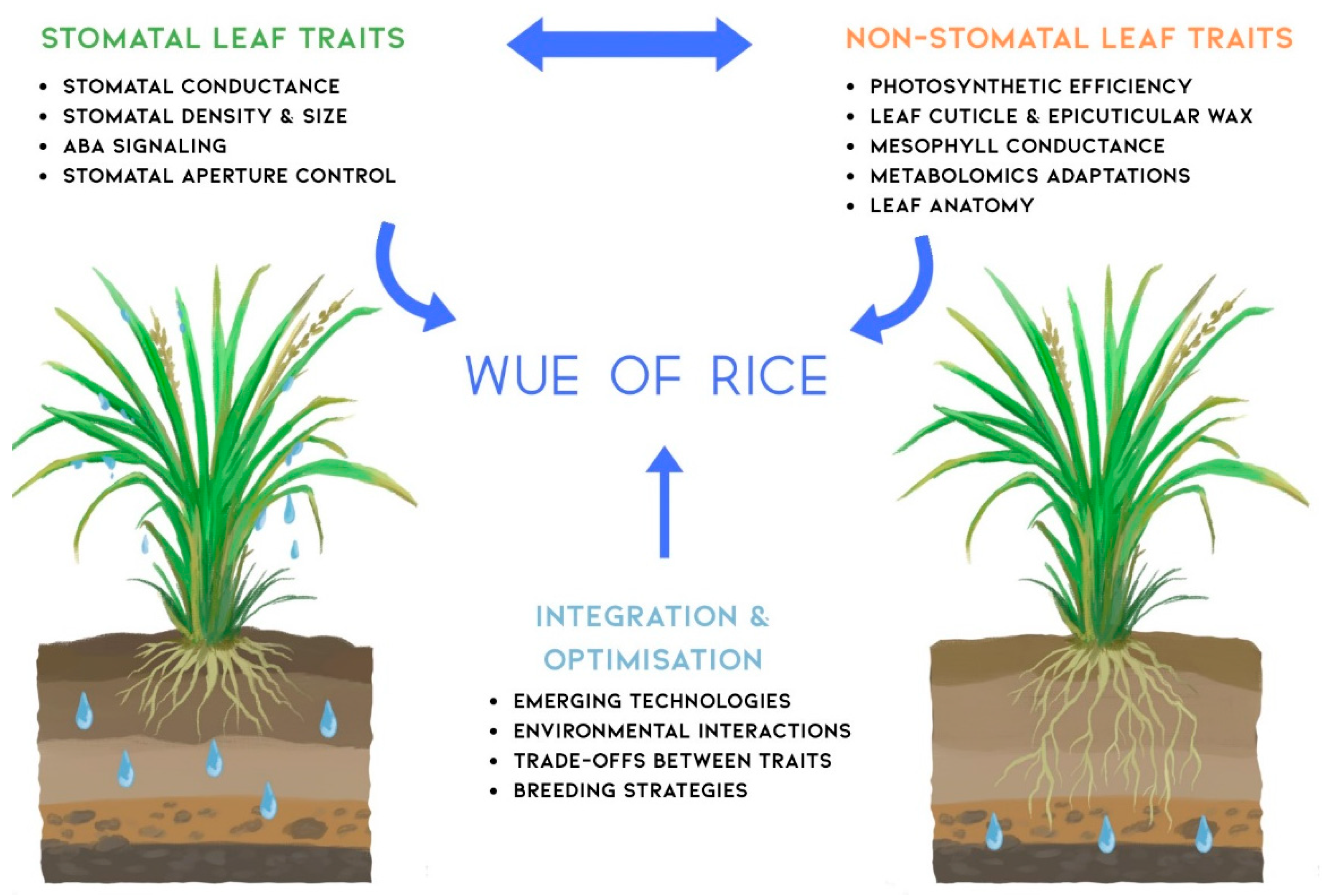
Table 2.
Multi-omics approaches for studying water use efficiency in rice. The table summarises key technologies, their specific applications to WUE research, notable research findings, and representative references across different omics platforms. Genomics approaches focus on gene identification and editing; transcriptomics on expression profiling; proteomics on protein-level changes; metabolomics on biochemical responses; phenomics on high-throughput trait measurement; and integrative approaches on systems-level understanding of WUE mechanisms.
Table 2.
Multi-omics approaches for studying water use efficiency in rice. The table summarises key technologies, their specific applications to WUE research, notable research findings, and representative references across different omics platforms. Genomics approaches focus on gene identification and editing; transcriptomics on expression profiling; proteomics on protein-level changes; metabolomics on biochemical responses; phenomics on high-throughput trait measurement; and integrative approaches on systems-level understanding of WUE mechanisms.
| Omics Level | Key Technologies |
Applications to WUE |
Notable Findings |
References |
|---|---|---|---|---|
| Genomics | GWAS, QTL mapping, CRISPR-Cas9, Transgenic overexpression | Identification of genomic regions associated with WUE; Targeted gene editing of stomatal regulators | Transgenic overexpression and CRISPR/Cas9-mediated editing of the OsEPF1 gene in rice have been shown to significantly reduce stomatal density, leading to improved drought tolerance and altered photosynthetic performance. These findings highlight OsEPF1 as a key regulator of WUE. | [33,89,154] |
| Transcriptomics | RNA-Seq, microarray analysis, RT-qPCR | Profiling gene expression networks under drought; Comparative transcriptome profiling of drought-tolerant and sensitive rice genotypes; Identification of drought-responsive noncoding RNAs and their regulatory targets | Transcriptome analysis revealed hundreds of drought-responsive genes, including OsDREB2A and OsLEA3, which are key regulators in ABA-mediated drought response pathways, 66 miRNAs and 98 lncRNAs were differentially expressed under drought; miR171f-5p targeted Os03g0828701-00, suggesting a role in drought adaptation. | [159,160,161] |
| Proteomics | LC-MS/MS, iTRAQ, 2D-electrophoresis | Identification of drought-responsive proteins; Analysis of PTMs during water deficit | Over 2000 proteins were detected in rice leaves under drought; 42 showed significant changes. Key drought-responsive proteins included actin depolymerizing factor, S-like ribonuclease, and chloroplastic dehydroascorbate reductase. PTMs such as phosphorylation, ubiquitination, and glycosylation modulate protein function under drought, contributing to stress tolerance. | [161,162] |
| Metabolomics | GC-MS, LC-MS, NMR spectroscopy | Profiling of osmoprotectants and secondary metabolites under water stress, identifying antioxidant compounds that enhance WUE. | Flag leaves exhibited cultivar-specific increases in proline, sucrose, and malate under combined drought and heat stress. Overaccumulation of flavonoids, such as kaempferol and quercetin, enhances drought and UV tolerance by reducing oxidative damage, overexpressing Flavanone 3-Hydroxylase showed higher kaempferol and quercetin levels, lower ROS and salicylic acid, and upregulated expression of DHN and UVR8 genes. | [64,163,164] |
| Phenomics | Thermal imaging, hyperspectral analysis, chlorophyll fluorescence, LiDAR | High-throughput field screening; Non-invasive measurement of physiological traits | Thermal imaging improves detection of crop water deficit by capturing spatial canopy temperature variations, enabling non-invasive and real-time assessment of plant responses to drought stress under field conditions, 3D LiDAR enables precise canopy structure analysis linked to stomatal function and transpiration. | [138,139,148,149] |
| Integrative Multi-Omics | Network analysis, systems biology, machine learning | Integration of genomic, transcriptomic, proteomic, and metabolomic data; Predictive modeling of WUE traits | Multi-omics integration revealed gene, protein, and metabolite interactions enhancing drought tolerance, highlighting the potential of big-omics data to breed drought-resilient rice with improved WUE under climate change. Multi-omics integration provides a holistic view of biological responses to drought stress. Enables identification and manipulation of genes linked to drought tolerance, | [165,166,167] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



