
Submitted:
24 December 2025
Posted:
25 December 2025
You are already at the latest version
Abstract
Metabolic dysfunction-associated steatotic liver disease (MASLD) is a growing health concern, yet the role of non-coding RNAs (ncRNAs), including microRNAs (miRNAs), in its pathogenesis remains poorly understood. In this pilot study, we aimed to identify significantly expressed miRNAs and ncRNAs and correlate transcriptomic patterns of the findings with previously identified coding gene expression profiles to explore potential regulatory mechanisms in MASLD. Participants were selected from an existing study population. We conducted transcriptomic profiling of miRNAs and other ncRNAs in whole-blood samples from African American individuals with MASLD and matched controls (n = 4 per group) as a discovery cohort. A subsequent qRT-PCR validation study was performed in 30 participants, including 14 individuals with MASLD and 16 controls. miRNA sequencing was performed by Zymo, USA followed by miRNA extraction using Zymo-Seq™ miRNA Library Kit. Differentially expressed miRNAs and ncRNAs were analyzed using Ingenuity Pathway Analysis (IPA) to identify associated biological pathways. A total of 1,412 miRNAs and 5,423 other ncRNAs were identified in this study. Among them, 35 miRNAs and 28 other ncRNAs exhibited significant differential expressions (fold change cut off 1.5, p < 0.05). miR-206 was upregulated, potentially compensating for insulin resistance, while miR-1343-5p miR-1299, miR-224-5p, and miR-193a-5p were downregulated, connecting impaired lipid metabolism and fibrosis. The validation study confirmed the upregulation of miR-206 and downregulation of miR-185-3p, miR-224-5p, and miR-218-5p. IPA results identified hepatic fibrosis and cirrhosis pathways enriched with interactions between miRNA and ncRNA. Our findings highlight promising candidates for future biomarker validation and therapeutic targeting. Further large-scale studies are necessary to validate these candidates and elucidate their role in MASLD pathogenesis and ethnic disparities.
Keywords:
microRNAs (miRNAs)
; non-coding RNAs (ncRNAs)
; MASLD
; African Americans
; transcriptomics
1. Introduction
Metabolic dysfunction-associated steatotic liver disease (MASLD), formerly known as non-alcoholic fatty liver disease (NAFLD), is now recognized as the most prevalent chronic liver disease globally, affecting approximately 25-30% of the adult population [1]. The disease spectrum ranges from simple hepatic steatosis (fat accumulation in the liver) to non-alcoholic steatohepatitis (NASH), which can further progress to advanced fibrosis, cirrhosis, and hepatocellular carcinoma (HCC) [2]. MASLD is strongly linked to metabolic comorbidities, including obesity, insulin resistance, type 2 diabetes mellitus (T2DM), hypertension, and dyslipidemia. Importantly, MASLD is not just a liver-centric disease but is increasingly recognized as a hepatic manifestation of systemic metabolic dysfunction [3]. The recent transition from NAFLD to MASLD reflects an evolving understanding that metabolic dysfunction is central to the disease's pathogenesis, irrespective of alcohol consumption [85]. Unlike previous definitions that excluded other liver conditions primarily based on alcohol thresholds, MASLD emphasizes the metabolic underpinnings, including insulin resistance, visceral adiposity, and dysregulated lipid metabolism as key drivers of disease progression [85].
Several pathophysiological mechanisms contribute to MASLD development, including hepatic lipid accumulation, mitochondrial dysfunction, oxidative stress, endoplasmic reticulum stress, chronic low-grade inflammation, and fibrogenesis [86]. Insulin resistance plays a pivotal role by increasing free fatty acid flux to the liver, promoting de novo lipogenesis, and impairing lipid exports. This lipid overload leads to lipotoxicity, which triggers hepatocellular injury and activation of hepatic stellate cells, culminating in fibrosis [87]. Despite its high prevalence, MASLD exhibits considerable heterogeneity across populations, with African Americans (AAs) paradoxically displaying a lower prevalence of hepatic steatosis and slower progression to advanced fibrosis despite a high burden of metabolic risk factors [5,8]. The molecular mechanisms driving these ethnic differences remain poorly understood.
A limited, but increasing, number of studies have underscored the regulatory roles of non-coding RNAs, particularly microRNAs (miRNAs), and other non-coding RNAs (ncRNAs) in the development and progression of MASLD [10,11]. The miRNAs are small non-coding RNA molecules that post-transcriptionally regulate gene expression by targeting mRNAs for degradation or translational repression. Small, non-coding RNA molecules also play a crucial role in regulating gene expression by targeting mRNA transcripts [39] by binding to the 3' untranslated region (3'UTR) of target mRNAs, leading to mRNA degradation or inhibition of translation. This post-transcriptional regulation of gene expression helps control various cellular processes, including cell growth, development, and differentiation [39].
miRNAs play crucial roles both within cells and in the extracellular environment by regulating gene expression intracellularly and functioning as intercellular messengers when secreted into extracellular fluids. They are transported via exosomes or RNA-binding protein complexes such as those involving AGO proteins to modulate gene expression in target cells [40]. Furthermore, an expanding body of evidence demonstrates that ncRNAs operate within competitive endogenous RNA (ceRNA) networks, where lncRNAs or circular RNAs (circRNAs) act as molecular sponges, sequestering specific miRNAs and thereby modulating their availability and function. For example, circ_0057558 and lncRNA MALAT1 have been shown to promote hepatic lipid accumulation and steatosis by sponging miR-206, a key regulator of lipid metabolism and insulin sensitivity [23,24]. Similarly, lncRNAs such as LINC00963 and LINC01234 are implicated in liver fibrosis by modulating fibrogenic signaling through ceRNA mechanisms [37,38]. However, there have been few attempts to coordinate expression patterns of non-coding RNAs with the altered gene expression patterns in MASLD subjects, especially within the same individuals.
To determine the feasibility of addressing this issue, a case-control pilot study of the expression patterns of ncRNAs was conducted using individuals from a previous [7,8] transcriptomic investigation conducted by our group. This population was comprised of African Americans (AA) to address the lack of inclusion of minority participants in prior studies of transcriptomic patterns in MASLD patients. This study utilized whole blood samples which we and others have demonstrated to have substantial utility in establishing transcriptomic patterns that largely overlap patterns observed in hepatic tissue [7,8]. The current pilot study, to the best of our knowledge, reports for the first time the transcriptomic profiling study of miRNA and other ncRNA expressions in whole blood samples from AA individuals with MASLD.
2. Results
2.1. Study Participants
All selected participants (total n=38) were AAs and were part of the previous transcriptomic study from the Washington DC area [7]. The participants were separated into two groups: a control group of individuals without MASLD, and a case group of individuals with early stage MASLD (confirmed hepatic steatosis and exhibited one or more comorbid metabolic features, viz., type 2 diabetes, hypertension, hyperlipidemia, or obesity). A qRT-PCR validation study was then performed in 30 participants, including 14 from the MASLD group and 16 from the control group. MASLD was diagnosed based on standard criteria including confirmed hepatic steatosis (based on their imaging/biopsy records supported by the presence of hepatic steatosis on cross-sectional imaging, liver elastography, and/or histological confirmation by percutaneous liver biopsy) and exhibited one or more comorbid metabolic features, viz., type 2 diabetes, hypertension, hyperlipidemia, or obesity [7]. Table 1 displays the characteristics of each group. No significant differences in age, BMI, or HbA1c were observed in the discovery group; however, a significant difference (p-value 0.01) in HbA1c (%) was observed in the validation cohort, with mean HbA1c levels of 5.35 ± 0.57% in the control group and 6.58 ± 1.56% in the MASLD group. For the purposes of liver steatosis staging, we used the S grade (S0 - S3); the higher the grade, the higher percentage of liver affected by fatty changes.
2.2. miRNA and Noncoding-RNA Sequencing, Differential Expression, Number of Reported Studies
Out of a total of 1412 miRNA identified transcripts, 35 miRNAs in the MASLD cases were significantly differentially expressed when compared to the controls (fold change cutoff 1.5-fold and p-value < 0.05) with 24 downregulated and 11 upregulated (Figure 1A and Table 2). Out of a total of 5423 other ncRNAs transcripts, 28 were significantly differentially expressed with 17 downregulated and 11 upregulated in the MASLD cases compared to the controls (Figure 1B and Table 3).
We also examined the differential miRNA expression of each individual case compared to the control group’s expression status (Figure 2A). We observed miR-1299, miR-193a-5p, miR-185-3p, miR-3960, miR-1343-5p, and miR-224-5p, were significantly downregulated and miR-206 significantly upregulated in all the MASLD subjects.

Table 4.
Micro-RNAs differentially expressed in all MASLD subjects.
| miRNA ID | Fold Change | p-Value | Role in MASLD | |
| miR-206 | 2.22±0.19 | 0.0353 | miR-206 regulates lipid metabolism and fibrosis in MASLD by downregulating FGF21 and modulating the MAPK pathway. | |
| miR-1343-5p | -3.98±2.50 | 0.0020 | miR-1343-5p contributes to MASLD by modulating the PI3K/Akt pathway, promoting hepatic lipid accumulation and inflammation. | |
| miR-224-5p | -2.65±0.52 | 0.0001 | miR-224-5p exacerbates MASLD by activating the TGF-β/Smad pathway, promoting liver fibrosis and inflammation. | |
| miR-1299 | -3.59±1.71 | 0.0055 | miR-1299 plays a role in MASLD by inhibiting the Wnt/β-catenin pathway, thereby reducing hepatic fibrosis and lipid accumulation. | |
| miR-193a-5p | -1.79±0.26 | 0.0031 | miR-193a-5p contributes to MASLD by deactivating the JNK/c-Jun pathway, which reduces inflammation and hepatic injury. | |
| miR-185-3p | -2.59±1.06 | 0.0038 | miR-185-3p mitigates MASLD by inhibiting the NF-κB pathway, reducing inflammation and liver damage. | |
| miR-3960 | -1.64±0.95 | 0.0270 | miR-3960 contributes to MASLD by activating the SIRT1/AMPK pathway, promoting lipid metabolism and reducing hepatic steatosis. | |
2.3. Top Biofunctions, Canonical Pathways, and Network Analysis
The IPA analysis revealed significant biofunctions, including "fibrosis of liver" and "cirrhosis of liver," with significant overlap percentages (p-value <0.05). These biofunctions were associated with 10 distinct miRNAs: miR-100-5p, miR-1273h-5p, miR-130a-3p, miR-133a-3p, miR-135a-5p, miR-143-3p, miR-16-5p, miR-199a-5p, miR-27a-3p, and miR-526a-5p (Figure 2A, 2B).
In addition to the miRNA findings, other ncRNAs were prominently featured in the network analysis (Figure 3). Several long non-coding RNAs (lncRNAs), such as LINC00963, SNHG7, CYTOR, and HORMAD2-AS1, were identified as key regulators in the hepatic fibrosis and cirrhosis pathways. These lncRNAs interacted with critical molecular hubs, including EZH2, AKT, and YAP1, highlighting their regulatory roles in fibrosis-related processes. The network further identified ncRNAs, such as RP11 and LINC01234, as contributing to the modulation of gene expression, emphasizing their potential involvement in liver disease pathogenesis. The canonical pathway analysis (Figure 4) highlighted the "hepatic fibrosis signaling pathway" as a central mechanism linking these ncRNAs and miRNAs to critical molecular and cellular functions. The interactions between miRNAs, lncRNAs, and target genes suggest a tightly regulated network underlying MASLD.
2.4. qRT-PCR Validation of Differentially Expressed miRNAs
To validate the sequencing-derived miRNA signature, we performed qRT-PCR in 30 participants (MASLD: n = 14; Controls: n = 16) who were not part of the above profiling experiments. We targeted seven miRNAs that were consistently dysregulated across all MASLD subjects in the discovery dataset. As shown in Figure 5, the qRT-PCR results demonstrated strong agreement with the discovery data, both in fold-change direction and level of significance. miR-206, which showed an average fold-change of 2.22 ± 0.19 in the discovery dataset, was significantly upregulated in MASLD (p < 0.05). Among the downregulated miRNAs, miR-185-3p (–1.78 ± 1.90), miR-224-5p (–4.07 ± 2.24), and miR-218-5p (–5.01± 4.36) demonstrated significant reductions in MASLD (p < 0.05). Three additional miRNAs—miR-1343-5p (–0.74 ± 1.94), miR-1299 (–0.42 ± 2.73), and miR-193a-5p (–0.43 ± 1.55), displayed consistent downward trends compared to controls, although these did not reach statistical significance in the validation cohort, likely reflecting biological variability and the modest sample size (Figure 5).
3. Discussion
In this pilot study, we present a pilot transcriptomic analysis of miRNA and other ncRNA expression profiles in whole blood samples from AA individuals with early -stage MASLD. By identifying differentially expressed ncRNA species, our work aims to generate hypotheses on regulatory pathways involved in early stage of MASLD.
Integrating sequencing data with qRT-PCR validation, four miRNAs, miR-206, miR-185-3p, miR-224-5p, and miR-218-5p, demonstrated statistically significant dysregulation, highlighting their strong involvement in lipid metabolism, inflammation, and fibrogenesis in MASLD. miR-206, which plays a dual role in lipid metabolism and insulin sensitivity, has been shown to inhibit hepatic lipogenesis and gluconeogenesis, thereby promoting insulin responsiveness [23]. Chen et al., 2021 demonstrated that circ_0057558 promotes hepatic lipid accumulation by targeting miR-206, which in turn regulates the ROCK1/AMPK signaling pathway. Inhibition of miR-206 resulted in increased lipid accumulation, while its overexpression reduced lipid content, suggesting miR-206 plays a protective role in preventing hepatic steatosis and potentially improving insulin sensitivity [23]. Xiang et al., 2022 reported that miR-206 negatively regulates ARNT expression, impacting the PPARα/CD36 pathway, which plays a crucial role in hepatic lipid metabolism, and showed that manipulating miR-206 levels alters lipid accumulation and liver injury severity [24]. Mohammed et al., 2024 identified elevated circulating miR-206 levels in patients with hepatic steatosis and hyperlipidemia, suggesting a systemic role for miR-206 in metabolic regulation, possibly as a compensatory response to metabolic dysfunction [22]. In light of these findings, the consistent upregulation (2.5-fold) of miR-206 in MASLD patients suggests a compensatory role in mitigating hepatic insulin resistance and lipid accumu[20,25lation, aligning with previous findings demonstrating its regulatory effect on the AMPK and PPARα pathways [23,24].
miR-224-5p is known to be involved in multiple regulatory processes such as lipid accumulation, endoplasmic reticulum stress, mitochondrial damage, inflammatory response, autophagy, and hepatic stellate cell activation, potentially influencing the progression of MASLD [20,25]. Upregulated miR-224-5p targets the leptin (LEP) gene and leads to its suppression through dysregulation of the AMPK pathway, which is associated with MASLD progression [18]. Intriguingly, miR-224-5p exhibited significant downregulation in both discovery and validation datasets (–4.07 ± 2.24; p < 0.05), despite prior studies reporting upregulation in non-AA MASLD populations [20,21]. Given that most published MASLD miRNA studies have profiled hepatic tissue or serum/plasma rather than whole blood, differences in biological compartment may partially explain this discordance. Directionality of miRNA dysregulation is not uniformly conserved between liver tissue and peripheral blood, reflecting differences in cellular origin, immune composition, and systemic metabolic state. Indeed, non-MASLD studies have reported context-dependent and compartment-specific regulation of miR-224-5p, with opposing patterns observed between tissue and circulating immune cells. Nevertheless, the consistent downregulation of miR-224-5p across all AA MASLD participants in this study suggests that circulating miRNA signatures may capture regulatory processes distinct from hepatic tissue expression and potentially shaped by ancestry-associated immunometabolic environments [5,26]. Additionally, dysregulation of other miRNAs, such as miR-370-3p and miR-218-5p, further connect pathways linked to lipid metabolism, inflammation, and hepatic fibrosis, underscoring their multifactorial contribution to MASLD pathobiology [41,42]. Both miR-185-3p and miR-218-5p were significantly downregulated by qRT-PCR (p < 0.05), and have regulatory roles in NF-κB–mediated inflammation and hepatic lipogenesis. Future studies involving diverse, multi-ethnic cohorts are essential to validate whether the regulatory patterns of miR-224-5p are indeed influenced by racial or genetic backgrounds and to elucidate potential African American-specific mechanisms.
Downregulation of miR-1343-5p and miR-1299, both known negative regulators of TGF-β and Wnt/β-catenin signaling respectively, may exacerbate fibrogenesis by promoting hepatic stellate cell activation and extracellular matrix deposition [28,37,38]. We observed, downregulation of miR-1343-5p in all four MASLD patients in our study supports our previously published findings, which noted an upregulation of TGF-β and activation of the hepatic fibrosis signaling pathway in these subjects [7]. Another previous study reported that upregulation of circulating miR-1343-5p is a potential biomarker in MASLD in adolescents with severe obesity, [27]. Although miR-1343-5p (–0.74 ± 1.94), and miR-1299 (–0.42 ± 2.73) showed consistent downward trends across discovery and validation, these changes did not reach statistical significance in the validation cohort, likely reflecting biological variability.
The significant downregulation of miR-193a-5p in the discovery cohort study, which has been previously reported as a biomarker for liver fibrosis and cirrhosis [29,30,31,32,33,34], reinforces its potential role as an early indicator of MASLD. The downregulation of miR-193a-5p is thought to inhibit pro-fibrotic gene targets, such as TGFB2, thereby promoting hepatic stellate cell activation, a central event in the fibrotic cascade [35]. This observation suggests a plausible link between miR-193a-5p downregulation and progressive fibrogenesis in MASLD, highlighting its relevance in the early stages of disease pathogenesis. Moreover, given that miR-193a-5p plays a regulatory role in extracellular matrix remodeling and fibrotic signaling, its dysregulation may contribute directly to the hepatic tissue alterations characteristic of MASLD progression [88]. Notably we observe similar downregulation trend in the validation study, but the relationship was not significant. Our prior research identified dysregulation of the TGFB1 and E2F1 genes and associated pathways in peripheral blood samples in the cohort of AA patients with early-stage MASLD [7] used for the current study. We observed the activation of hepatic fibrosis signaling pathways and their potential role in the development of hepatocellular carcinoma, particularly when TGFB1 was upregulated and E2F1 was downregulated [7]. However, the study did not establish a definitive role for TGFB1 and E2F1 regulation in the development of hepatic steatosis or the lower prevalence of MASLD in AAs. Some previous studies have reported an upregulation trend of miR-193a-5p in blood serum and plasma samples, mainly in Caucasian populations [29,30,31].
In addition to miRNAs, our pathway and network analyses identified several lncRNAs involved in fibrosis-related signaling networks. lncRNAs such as LINC00963, SNHG7, CYTOR, and HORMAD2-AS1 interact with pivotal regulatory molecules including EZH2, AKT, and YAP1. EZH2, a histone methyltransferase, has been implicated in the progression of fibrosis, while YAP1, a core component of the Hippo signaling pathway, plays a critical role in liver regeneration and fibrogenesis [37]. Notably, circ_0057558 and lncRNA MALAT1 have been shown to promote hepatic lipid accumulation and metabolic dysfunction by sponging miR-206, a key regulator of lipid metabolism and insulin sensitivity [23,24]. The consistent upregulation of miR-206 in our MASLD cohort may therefore reflect compensatory regulatory feedback within ceRNA networks involving these or other uncharacterized transcript variants. Other studies demonstrate that transcript variants derived from lncRNAs such as LINC01234, LINC01138, and CYTOR can generate multiple isoforms with diverse ceRNA functions, influencing hepatic stellate cell activation, extracellular matrix remodeling, and the fibrotic response—central events in MASLD pathogenesis [37,38,81]. In other study, CYTOR has been shown to participate in YAP1-mediated pathways, enhancing fibrogenic gene expression, while alternative transcript isoforms of WDFY3-AS2 may exert protective effects by dampening pro-fibrotic signaling cascades through competitive miRNA biFnding [83]. Our canonical pathway analysis indicates strong associations between several ncRNAs (LINC00963, HCG18, ST7-AS1, RP11_25D31, CYTOR and LINC01234) and hepatic fibrosis pathways. The notable upregulation of WDFY3-AS2 and LINC02767 suggests that these lncRNAs may contribute to hepatic inflammation and fibrotic remodeling, although further validation is required in larger populations [38].
Although our study offers new hypotheses on the transcriptomic landscape of MASLD in African Americans and demonstrates the utility of whole blood samples for such investigations, it also has some limitations. The small study size restricts the generalizability of our findings and is vulnerable to selection bias. We focused on patients with early stage MASLD, for whom future interventions might prevent the progression of the disease. Future studies should incorporate larger, multi-ethnic cohorts and additional analytical tools to enhance statistical power and validate these preliminary observations. As our analysis was based on whole blood RNA profiles, the findings may not fully recapitulate hepatic cell transcriptomic alterations. Future research therefore should attempt to include matched analyses of liver tissue with the circulating plasma miRNAs to provide a more comprehensive understanding of MASLD pathogenesis. Nonetheless, the strong concordance in expression patterns between discovery and validation, and the statistical validation of four miRNAs, support the robustness of our circulating miRNA signature.
4. Materials and Methods
4.1. Study Particepants and Blood Sample Collection.
In this pilot study, the study participants consisted of eight individuals (control n=4; MASLD n=4), with equal numbers of males and females, who self-identified as AA and were born in the USA. All participants responded to an advertisement through Howard University and Georgetown University Community Newsletter via email and/or flyers and public announcements and were recruited with their informed consent. The protocol was approved by Georgetown-MedStar IRB (MODCR00002260). Participants with MASLD were recruited from the MedStar Georgetown Transplant Institute. We selected only those patients who were at early stages of development MASLD (confirmed hepatic steatosis and exhibited one or more comorbid metabolic features, viz., type 2 diabetes, hypertension, hyperlipidemia, or obesity). Individuals with severe fibrosis or cirrhosis were not included because of the small size of the participant groups and the wide spectrum of tissue features present during different stages of liver disease; we chose to limit subject inclusion to earlier stages of liver disease to increase the homogeneity of disease presentation in the different subjects. Individuals with severe fibrosis or cirrhosis were not included; we chose to limit subject inclusion to earlier stages of liver disease to increase the homogeneity of disease presentation in the different subjects (Table 1). Patients with other potential causes of liver disease, including viral, immunological, iron storage disease, Wilson disease, or alpha 1 antitrypsin deficiency, were excluded from the study. Participants with heavy alcohol use were also excluded from the study. Control participants were those who responded to the same flyers and advertisements described above but did not have MASLD. These individuals were negative for self-reported HCV and HBV and had normal liver enzyme profiles. A questionnaire was provided to all participants to collect demographic and clinical information.
4.2. RNA Extraction and miRNA Library Preparation
Whole blood was collected in a DNA/RNA Shield™ Blood Collection Tube (Manufacturer: Zymo Research, Cat # R1150) during recruitment by experienced phlebotomists. Blood collection tubes were prefilled with 6 ml DNA/RNA Shield™ for direct collection of up to 3 ml whole human blood. DNA/RNA Shield lyses cells, inactivate nucleases and infectious agents (e.g., viruses and pathogens), and is ideal for safe sample storage and transport at ambient temperatures. RNA was extracted from DNA/RNA Shield tubes using the Quick-DNA/RNA™ Blood Tube Kit (Zymo Research, Cat. # R1151) according to the manufacturer's instructions. DNA contamination was removed using an Applied Biosystems Inc. (ABI) DNA-free kit (ThermoFisher, CA, Cat # AM 1906). RNA was quantified using a NanoDrop™ One spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA). The ratio of absorbance at 260 and 280 nm was used to assess the purity of DNA and RNA.
We used Zymo-Seq™ miRNA Library Kit (Catalog Numbers: R3006, R3007) to generate small RNA libraries. To acquire miRNAs, present in total RNA or cell-free RNA extracted from biofluids we followed manufacturer's instructions and protocol. Briefly the protocol was with five major steps, which are adapter ligation and blocking, circularization and dimer removal, reverse transcription, index PCR, and library purification. At the time of index PCR pre-mixed forward and reverse primers with the following sequence:
Forward Primer Sequence:
5′-AATGATACGGCGACCACCGAGATCTACACNNNNNNNN(NN)ACAC TCTTTCCCTACACGACGCTCTTCCGATCT-3′
Reverse Primer Sequence:
5′-CAAGCAGAAGACGGCATACGAGATNNNNNNNN(NN)GTGACTGGA GTTCCTTGGCACCCGAGAATTCCA-3′
4.3. Sequencing and Data Analysis
The Sequencing was performed by Zymo Research (Irvine, California, US). Data analysis was performed according to instruction mentioned in the Zymo-Seq™ miRNA Library Kit (Catalog Numbers: R3006, R3007). To read the Zymo-Seq™ miRNA libraries we used bioinformatics tools (QIAGEN CLC Genomics Workbench) designed for Illumina’s TruSeq Small RNA libraries. Prior to sequence alignment, sequenced reads processed with adapter trimming. For the trimming we used sequence of TGGAATTCTCGGGTGCCAAGG. In the final analysis data was extracted in the form of fold change and p-values keeping false discovery rate (FDR) at the level of p<0.05.
4.4. Ingenuity Pathway Analysis (IPA)
IPA was utilized to explore complex biofunctions within a biological system, identifying functional roles, molecular processes, and key networks associated with significantly differentially expressed genes in participants with MASLD. From the differential expression of miRNAs and other ncRNAs datasets described above, the identification of cellular processes and pathways by IPA (Qiagen, USA) was performed according to the methods described in our earlier study [15,16,17]. Briefly, datasets comprising miRNAs and other ncRNAs identifiers and corresponding expression values (fold-change) from the sequencing data were imported into IPA. Differentially expressed identifiers (miRNAs and other ncRNAs) were mapped to related changes in biofunctions [16]. The networks were generated algorithmically based on their connectivity. Using IPA, we identified the top network by amalgamating a large set of differentially expressed miRNAs and other ncRNAs with the goal of uncovering the most extensive array of relationships among the focus genes [13]. A score (P-score = − log10 (p-value)) according to the fit of the set of supplied genes and a list of biological functions stored in the Ingenuity Knowledge Base are generated [15] Networks were “named” on the most prevalent functional group(s) present. Canonical pathway analysis identified function-specific genes that were significantly present within the networks.
4.5. qRT-PCR Validation Study
In the qRT-PCR validation study, 30 participants were included (14 from the MASLD group, 16 from the control group). We used Applied Biosystem® pre-configured TaqMan array card (96 well array card format) to examine the expression of genes of interest, viz., 18s (manufacturing control), GAPDH (internal control), miR-206, miR-1343-5p, miR-224-5p, miR1299, miR-193a-5p, miR185-3p, and miR-218-5p as identified based on the global expression data and list of significant miRNA expressed in all samples. Six samples per plate were analyzed by High Throughput FAST Real-Time PCR (Applied Biosystems, CA). qRT-PCR was carried out on a QuantStudio 7 Pro PCR system (Applied Biosystems, CA). The qRT-PCR mixture contained 5 μL of TaqMan™ Fast Advanced Master mix (Part # 4444557, Applied Biosystems), 5 μL of cDNA diluted sample (as recommended by Taqman advanced miRNA assays protocol, Publication Number 100027897). All qRT-PCR reactions were performed in triplicates.
5. Conclusions
Our study is among the first to generate hypotheses on the significance of miRNA and ncRNA expression patterns in MASLD among AAs. The pathway analyses reveal correlations between these ncRNA patterns and the transcriptomic patterns of coding genes, particularly in pathways involving TGFB1 and E2F signaling. The findings also suggest that miR-206 upregulation may represent a protective response to insulin resistance. miR-206, miR-370-3p, and miR-193a-5p downregulation could contribute to MASLD pathogenesis via impaired lipid metabolism and fibrosis promotion. Other ncRNAs such as LINC00963, SNHG7, and CYTOR are implicated in hepatic fibrosis signaling. These findings highlight promising candidates for future biomarker investigation and therapeutic targeting. Further large-scale studies, including longitudinal transcriptomic profiling, will be essential to elucidate the role of ncRNAs in MASLD progression and ethnic disparities.
Author Contributions
Conceptualization, S.G., B.K., T.M. and C.L.; writing T.M.; formal analysis, C.L. and T.M.; resources, G.N.B., R.Q., C.H., C.S., and G.M.; methodology, T.M.; writing—review and editing, G.N.B., C.L., BK, Z.A.S., and S.G.
Funding
This study was funded by a P20 grant (CA262617-01) (PI: Ghosh) from the NCI of Health (NIH), and by U54 (MD007597-31-5959; PI/PD: Southerland, Lead PI: Ghosh) from the National Institutes of Health (NIH).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board (or Ethics Committee) of Georgetown-MedStar IRB (protocol code MODCR00002260, approval date on 2024-07-18).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors would like to extend their sincere thanks to all the respondents who voluntarily participated in the study and provided valuable responses during the recruitment process. The authors are also grateful to Dr. Alexander Kroemer, Dr. Ziman Zang, and Dr. Digvijay Patil from MedStar Georgetown Transplant Institute, Georgetown University School of Medicine, Washington, DC, for their scientific review and valuable suggestions on the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Younossi, Z.M.; Stepanova, M.; Afendy, M.; Fang, Y.; Younossi, Y.; Mir, H.; Srishord, M. Changes in the Prevalence of the Most Common Causes of Chronic Liver Diseases in the United States From 1988 to 2018. Clin. Gastroenterol. Hepatol. 2019, 17, 2239–2246. [Google Scholar] [CrossRef]
- Chalasani, N.; Younossi, Z.; LaVine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the American Association for the Study of Liver Diseases. Hepatology 2021, 67, 328–357. [Google Scholar] [CrossRef] [PubMed]
- Diehl, A.M.; Day, C. Cause, Pathogenesis, and Treatment of Nonalcoholic Steatohepatitis. New Engl. J. Med. 2020, 383, 2063–2072. [Google Scholar] [CrossRef]
- Browning, J.D.; Szczepaniak, L.S.; Dobbins, R.; Nuremberg, P.; Horton, J.D.; Cohen, J.C.; Grundy, S.M.; Hobbs, H.H. Prevalence of hepatic steatosis in an urban population in the United States: Impact of ethnicity. Hepatology 2004, 40, 1387–1395. [Google Scholar] [CrossRef]
- Rich, N.E.; Oji, S.; Mufti, A.R.; Browning, J.D.; Parikh, N.D.; Odewole, M.; Mayo, H.; Singal, A.G. Racial and Ethnic Disparities in Nonalcoholic Fatty Liver Disease Prevalence, Severity, and Outcomes in the United States: A Systematic Review and Meta-analysis. Clin. Gastroenterol. Hepatol. 2022, 20, 1021–1031. [Google Scholar] [CrossRef]
- Abdelmalek, M.F.; Suzuki, A.; Guy, C.; Unalp-Arida, A.; Colvin, R.; Johnson, R.J.; Diehl, A.M. Ethnic differences in the histological severity of nonalcoholic fatty liver disease. Hepatology 2019, 50, 792–799. [Google Scholar]
- Mondal, T.; Smith, C.I.; Loffredo, C.A.; Quartey, R.; Moses, G.; Howell, C.D.; Korba, B.; Kwabi-Addo, B.; Nunlee-Bland, G.; Rucker, L.R.; et al. Transcriptomics of MASLD Pathobiology in African American Patients in the Washington DC Area †. Int. J. Mol. Sci. 2023, 24, 16654. [Google Scholar] [CrossRef]
- Mondal, T.; Loffredo, C.A.; Simhadri, J.; Nunlee-Bland, G.; Korba, B.; Johnson, J.; Cotin, S.; Moses, G.; Quartey, R.; Howell, C.D.; et al. Insights on the pathogenesis of type 2 diabetes as revealed by signature genomic classifiers in an African American population in the Washington, DC area. Diabetes/Metabolism Res. Rev. 2023, 39, e3589. [Google Scholar] [CrossRef]
- Sookoian, S.; Pirola, C.J. Genetic predisposition in nonalcoholic fatty liver disease. Clin. Mol. Hepatol. 2018, 23, 1–12. [Google Scholar] [CrossRef]
- Klett, D.; Moehle, C.; García-Rodríguez, J.L. MicroRNAs in NAFLD: Progress and perspectives. Biomolecules 2018, 8, 156. [Google Scholar]
- Pirola, C.J.; Sookoian, S. Noncoding RNAs in nonalcoholic fatty liver disease: Molecular insights and therapeutic implications. Nature Reviews Gastroenterology & Hepatology 2020, 17, 123–138. [Google Scholar]
- Estep, M.; Armistead, D.; Hossain, N.; Elarainy, H.; Goodman, Z.; Baranova, A.; Chandhoke, V.; Younossi, Z.M. Differential expression of miRNAs in the visceral adipose tissue of patients with non-alcoholic fatty liver disease. Aliment. Pharmacol. Ther. 2010, 32, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-H.; Ampuero, J.; Gil-Gómez, A.; Montero-Vallejo, R.; Rojas, Á.; Muñoz-Hernández, R.; Gallego-Durán, R.; Romero-Gómez, M. miRNAs in patients with non-alcoholic fatty liver disease: A systematic review and meta-analysis. J. Hepatol. 2021, 74, 76–89. [Google Scholar] [CrossRef]
- Arrese, M.; Cabrera, D.; Kalergis, A.M.; Feldstein, A.E. Innate Immunity and Inflammation in NAFLD/NASH. Dig. Dis. Sci. 2016, 61, 1294–1303. [Google Scholar] [CrossRef]
- Sur, T.K.; Mondal, T.; Noreen, Z.; Johnson, J.; Nunlee-Bland, G.; Loffredo, C.A.; Korba, B.E.; Chandra, V.; Jana, S.S.; Kwabi-Addo, B.; et al. Developing non-invasive molecular markers for early risk assessment of Alzheimer's disease. Biomarkers Neuropsychiatry 2025, 12. [Google Scholar] [CrossRef]
- Mondal, T.; Noreen, Z.; Loffredo, C.A.; Johnson, J.; Bhatti, A.; Nunlee-Bland, G.; Quartey, R.; Howell, C.D.; Moses, G.; Nnanabu, T.; et al. Transcriptomic Analysis of Alzheimer’s Disease Pathways in a Pakistani Population. J. Alzheimer's Dis. Rep. 2024, 8, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Mondal, T.; Johnson, J.; Sur, T.K.; Loffredo, C.A.; Cotin, S.T.; Sahota, J.; Korbe, B.E.; Nunlee-Blnad, G.; Ghosh, S.G. Metabolic Dysfunction and Alzheimer’s Disease Risks in African Americans. Alzheimer's Dement. 2025, 20, e086476. [Google Scholar] [CrossRef]
- Vulf, M.; Shunkina, D.; Komar, A.; Bograya, M.; Zatolokin, P.; Kirienkova, E.; Gazatova, N.; Kozlov, I.; Litvinova, L. Analysis of miRNAs Profiles in Serum of Patients With Steatosis and Steatohepatitis. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef]
- Leti, F.; Malenica, I.; Doshi, M.; Courtright, A.; Van Keuren-Jensen, K.; Legendre, C.; Still, C.D.; Gerhard, G.S.; DiStefano, J.K. High-throughput sequencing reveals altered expression of hepatic microRNAs in nonalcoholic fatty liver disease–related fibrosis. Transl. Res. 2015, 166, 304–314. [Google Scholar] [CrossRef]
- Cheung, O.; Puri, P.; Eicken, C.; Contos, M.J.; Mirshahi, F.; Maher, J.W.; Kellum, J.M.; Min, H.; Luketic, V.A.; Sanyal, A.J. Nonalcoholic steatohepatitis is associated with altered hepatic MicroRNA expression. Hepatology 2008, 48, 1810–1820. [Google Scholar] [CrossRef]
- Mehta, R.; Otgonsuren, M.; Younoszai, Z.; Allawi, H.; Raybuck, B.; Younossi, Z. Circulating miRNA in patients with non-alcoholic fatty liver disease and coronary artery disease. BMJ Open Gastroenterol. 2016, 3, e000096. [Google Scholar] [CrossRef]
- Mohammed, A.; Shaker, O.G.; Khalil, M.A.F.; Abu-El-Azayem, A.K.; Samy, A.; Fathy, S.A.; AbdElguaad, M.M.K.; Mahmoud, F.A.M.; Erfan, R. Circulating miR-206, miR-181b, and miR-21 as promising biomarkers in hypothyroidism and their relationship to related hyperlipidemia and hepatic steatosis. Front. Mol. Biosci. 2024, 11, 1307512. [Google Scholar] [CrossRef]
- Chen, X.; Tan, Q.-Q.; Tan, X.-R.; Li, S.-J.; Zhang, X.-X. Circ_0057558 promotes nonalcoholic fatty liver disease by regulating ROCK1/AMPK signaling through targeting miR-206. Cell Death Dis. 2021, 12, 809. [Google Scholar] [CrossRef]
- Xiang, J.; Deng, Y.-Y.; Liu, H.-X.; Pu, Y. LncRNA MALAT1 Promotes PPARα/CD36-Mediated Hepatic Lipogenesis in Nonalcoholic Fatty Liver Disease by Modulating miR-206/ARNT Axis. Front. Bioeng. Biotechnol. 2022, 10. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, H.; Sun, Q.; Liu, X.; Wu, Z.; Wang, X.; Fang, W.; Ma, Z. miR-224-5p-enriched exosomes promote tumorigenesis by directly targeting androgen receptor in non-small cell lung cancer. Mol. Ther. - Nucleic Acids 2021, 23, 1217–1228. [Google Scholar] [CrossRef]
- Saini, A.; Rutledge, B.; Damughatla, A.R.; Rasheed, M.; Naylor, P.; Mutchnick, M. Manifestation and Progression of Metabolic Dysfunction-Associated Steatotic Liver Disease in a Predominately African American Population at a Multi-Specialty Healthcare Organization. Healthcare 2024, 12, 1478. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-J.; O Baumert, B.; Stratakis, N.; A Goodrich, J.; Wu, H.-T.; He, J.-X.; Zhao, Y.-Q.; Aung, M.T.; Wang, H.-X.; Eckel, S.P.; et al. Circulating microRNA expression and nonalcoholic fatty liver disease in adolescents with severe obesity. World J. Gastroenterol. 2024, 30, 332–345. [Google Scholar] [CrossRef]
- Stolzenburg, L.R.; Harris, A. Microvesicle-mediated delivery of miR-1343: impact on markers of fibrosis. Cell Tissue Res. 2018, 371, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; Leary, P.J.; Govaere, O.; Barter, M.J.; Charlton, S.H.; Cockell, S.J.; Tiniakos, D.; Zatorska, M.; Bedossa, P.; Brosnan, M.J.; et al. Increased serum miR-193a-5p during non-alcoholic fatty liver disease progression: Diagnostic and mechanistic relevance. JHEP Rep. 2021, 4, 100409. [Google Scholar] [CrossRef]
- Zhang, X.; Mens, M.M.J.; Abozaid, Y.J.; Bos, D.; Murad, S.D.; de Knegt, R.J.; Ikram, M.A.; Pan, Q.; Ghanbari, M. Circulatory microRNAs as potential biomarkers for fatty liver disease: the Rotterdam study. Aliment. Pharmacol. Ther. 2021, 53, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, J.T.; Sohal, A.; Handa, P.; Maliken, B.D.; Kim, T.-K.; Wang, K.; Gochanour, E.; Li, Y.; Rose, J.B.; Nelson, J.E.; et al. Serum miRNA profiles are altered in patients with primary sclerosing cholangitis receiving high-dose ursodeoxycholic acid. JHEP Rep. 2023, 5, 100729. [Google Scholar] [CrossRef]
- Behrooz, M.; Hajjarzadeh, S.; Kahroba, H.; Ostadrahimi, A.; Bastami, M. Expression pattern of miR-193a, miR122, miR155, miR-15a, and miR146a in peripheral blood mononuclear cells of children with obesity and their relation to some metabolic and inflammatory biomarkers. BMC Pediatr. 2023, 23, 1–12. [Google Scholar] [CrossRef]
- Estep, M.; Armistead, D.; Hossain, N.; Elarainy, H.; Goodman, Z.; Baranova, A.; Chandhoke, V.; Younossi, Z.M. Differential expression of miRNAs in the visceral adipose tissue of patients with non-alcoholic fatty liver disease. Aliment. Pharmacol. Ther. 2010, 32, 487–497. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Bu, X.; Zhang, X.; Cui, L. Identification of a novel miRNA-based recurrence and prognosis prediction biomarker for hepatocellular carcinoma. BMC Bioinform. 2022, 23, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, T.; Yang, Y.; Cho, W.C.; Flynn, R.J.; Harandi, M.F.; Song, H.; Luo, X.; Zheng, Y. Interplays of liver fibrosis-associated microRNAs: Molecular mechanisms and implications in diagnosis and therapy. Genes Dis. 2022, 10, 1457–1469. [Google Scholar] [CrossRef]
- Chen, X.; Tao, X.; Wang, M.; Cannon, R.D.; Chen, B.; Yu, X.; Qi, H.; Saffery, R.; Baker, P.N.; Zhou, X.; et al. Circulating extracellular vesicle-derived miR-1299 disrupts hepatic glucose homeostasis by targeting the STAT3/FAM3A axis in gestational diabetes mellitus. J. Nanobiotechnology 2024, 22, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Meng, J.; Shen, X.; Wang, M.; Yu, Y.; Shi, L.; Li, Y.-L.; Hassan, H.M.; Li, H.; He, Z.-X.; et al. Formononetin Mitigates Liver Fibrosis via Promoting Hepatic Stellate Cell Senescence and Inhibiting EZH2/YAP Axis. J. Agric. Food Chem. 2024, 72, 22606–22620. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, X.; Gui, S.; Yang, F.; Cao, Z.; Cheng, R.; Xia, X.; Li, C. The Roles and Mechanisms of lncRNAs in Liver Fibrosis. Front. Pharmacol. 2021, 12, 779606. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- O'BRien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Wu, G.; Zhang, Y.; Liang, B.; Yin, L.; Gao, M.; Zhang, H.; Xu, Y.; Han, X.; Qi, Y.; Liu, F.; et al. miR-218-5p promotes hepatic lipogenesis through targeting Elovl5 in non-alcoholic fatty liver disease. Biochem. Pharmacol. 2024, 226, 116411. [Google Scholar] [CrossRef]
- Yanni, J.; D’sOuza, A.; Wang, Y.; Li, N.; Hansen, B.J.; Zakharkin, S.O.; Smith, M.; Hayward, C.; Whitson, B.A.; Mohler, P.J.; et al. Silencing miR-370-3p rescues funny current and sinus node function in heart failure. Sci. Rep. 2020, 10, 11279. [Google Scholar] [CrossRef]
- Allahverdi, A.; Arefian, E.; Soleimani, M.; Ai, J.; Nahanmoghaddam, N.; Yousefi-Ahmadipour, A.; Ebrahimi-Barough, S. MicroRNA-4731-5p delivered by AD-mesenchymal stem cells induces cell cycle arrest and apoptosis in glioblastoma. J. Cell. Physiol. 2020, 235, 8167–8175. [Google Scholar] [CrossRef]
- Gozuacik, D.; Akkoc, Y.; Ozturk, D.G.; Kocak, M. Autophagy-Regulating microRNAs and Cancer. Front. Oncol. 2017, 7, 65. [Google Scholar] [CrossRef]
- Tang, W.; Rao, Y.; Pi, L.; Li, J. A review on the role of MiR-193a-5p in oncogenesis and tumor progression. Front. Oncol. 2025, 15, 1543215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.-B.; Du, Y.; Tian, Y.; Ji, Z.-G.; Yang, P.-Q. MiR-1299 functions as a tumor suppressor to inhibit the proliferation and metastasis of prostate cancer by targeting NEK2. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 530–538. [Google Scholar] [PubMed]
- Lafferty, M.J.; Aygün, N.; Patel, N.K.; Krupa, O.; Liang, D.; Wolter, J.M.; Geschwind, D.H.; de la Torre-Ubieta, L.; Stein, J.L. MicroRNA-eQTLs in the developing human neocortex link miR-4707-3p expression to brain size. eLife 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Kong, B; Zhao, S; Kang, X; Wang, B. MicroRNA-133a-3p inhibits cell proliferation, migration and invasion in colorectal cancer by targeting AQP1 [retracted in: Oncol Lett. 2024 Nov 20;29(2):66. https://doi.org/10.3892/ol.2024.14812.]. Oncol Lett. 2021, 22(3), 649. [Google Scholar] [CrossRef]
- Cheng, F.; Yang, M.-M.; Yang, R.-H. MiRNA-365a-3p promotes the progression of osteoporosis by inhibiting osteogenic differentiation via targeting RUNX2. 2019, 23, 7766–7774. [Google Scholar] [CrossRef]
- Hu, Y.; Dingerdissen, H.; Gupta, S.; Kahsay, R.; Shanker, V.; Wan, Q.; Yan, C.; Mazumder, R. Identification of key differentially expressed MicroRNAs in cancer patients through pan-cancer analysis. Comput. Biol. Med. 2018, 103, 183–197. [Google Scholar] [CrossRef]
- Dai, Q.; Shi, R.; Zhang, G.; Wang, Y.; Ye, L.; Peng, L.; Guo, S.; He, J.; Yang, H.; Jiang, Y. miR-539-5p targets BMP2 to regulate Treg activation in B-cell acute lymphoblastic leukemia through TGF-β/Smads/MAPK. Exp. Biol. Med. 2024, 249, 10111. [Google Scholar] [CrossRef]
- Qian, X.; Wang, Y.; Hu, W.; Xu, X.; Gao, L.; Meng, Y.; Yan, J. MiR-369-5p inhibits the proliferation and migration of hepatocellular carcinoma cells by down-regulating HOXA13 expression. Tissue Cell 2022, 74, 101721. [Google Scholar] [CrossRef]
- Zhang, N.; Wei, X.; Xu, L. miR-150 promotes the proliferation of lung cancer cells by targeting P53. FEBS Lett. 2013, 587, 2346–2351. [Google Scholar] [CrossRef]
- Ma, J.; Mannoor, K.; Gao, L.; Tan, A.; Guarnera, M.A.; Zhan, M.; Shetty, A.; Stass, S.A.; Xing, L.; Jiang, F. Characterization of microRNA transcriptome in lung cancer by next-generation deep sequencing. Mol. Oncol. 2014, 8, 1208–1219. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Flores, F.; Kaneko-Tarui, T.; Beyer, W.; Katz, J.; Chu, T.; Catalano, P.; Sadovsky, Y.; Hivert, M.-F.; O’tIerney-Ginn, P. Placental miR-3940-3p Is Associated With Maternal Insulin Resistance in Late Pregnancy. J. Clin. Endocrinol. Metab. 2021, 106, 3526–3535. [Google Scholar] [CrossRef]
- Xue, S.; Liu, K.; Zhao, L.; Zhou, L.; Gao, X.; Liu, L.; Liu, N.; He, J. The role of miR-369-3p in proliferation and differentiation of preadipocytes in Aohan fine-wool sheep. Arch. Anim. Breed. 2023, 66, 93–102. [Google Scholar] [CrossRef]
- Mushtaq, I.; Hsieh, T.-H.; Chen, Y.-C.; Kao, Y.-H.; Chen, Y.-J. MicroRNA-452-5p regulates fibrogenesis via targeting TGF-β/SMAD4 axis in SCN5A-knockdown human cardiac fibroblasts. iScience 2024, 27, 110084. [Google Scholar] [CrossRef] [PubMed]
- Ferraldeschi, M.; Romano, S.; Giglio, S.; Romano, C.; Morena, E.; Mechelli, R.; Annibali, V.; Ubaldi, M.; Buscarinu, M.C.; Umeton, R.; et al. Circulating hsa-miR-323b-3p in Huntington's Disease: A Pilot Study. Front. Neurol. 2021, 12, 657973. [Google Scholar] [CrossRef]
- Sun, S.; Wang, X.; Xu, X.; Di, H.; Du, J.; Xu, B.; Wang, Q.; Wang, J. MiR-433-3p suppresses cell growth and enhances chemosensitivity by targeting CREB in human glioma. Oncotarget 2017, 8, 5057–5068. [Google Scholar] [CrossRef]
- Meng, L.; Du, Y.; Deng, B.; Duan, Y. miR-379-5p regulates the proliferation, cell cycle, and cisplatin resistance of oral squamous cell carcinoma cells by targeting ROR1. 2023, 15, 1626–1639. [Google Scholar] [PubMed]
- Josson, S.; Gururajan, M.; Hu, P.; Shao, C.; Chu, G.C.-Y.; Zhau, H.E.; Liu, C.; Lao, K.; Lu, C.-L.; Lu, Y.-T.; et al. miR-409-3p/-5p Promotes Tumorigenesis, Epithelial-to-Mesenchymal Transition, and Bone Metastasis of Human Prostate Cancer. Clin. Cancer Res. 2014, 20, 4636–4646. [Google Scholar] [CrossRef]
- Wang, J.; Tan, J.; Qi, Q.; Yang, L.; Wang, Y.; Zhang, C.; Hu, L.; Chen, H.; Fang, X. miR-487b-3p Suppresses the Proliferation and Differentiation of Myoblasts by Targeting IRS1 in Skeletal Muscle Myogenesis. Int. J. Biol. Sci. 2018, 14, 760–774. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, X.; Dou, L.; Huang, X.; Zhu, K.; Guo, J.; Yan, M.; Wang, S.; Man, Y.; Tang, W.; et al. miR-154-5p Functions as an Important Regulator of Angiotensin II-Mediated Heart Remodeling. Oxidative Med. Cell. Longev. 2019, 2019, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Bao, D.; Ye, Z.; Cao, B.; Jin, G.; Lu, Z.; Chen, J. miR-3195 suppresses the malignant progression of osteosarcoma cells via targeting SOX4. J. Orthop. Surg. Res. 2023, 18, 809. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Song, X.; Wang, K.; Zheng, B.; Lin, Q.; Yu, M.; Xie, L.; Chen, L.; Song, X. Plasma exosomal miR-320d, miR-4479, and miR-6763-5p as diagnostic biomarkers in epithelial ovarian cancer. Front. Oncol. 2022, 12, 986343. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Wang, C.; Liu, Z. miR-196a-5p promotes metastasis of colorectal cancer via targeting IκBα. BMC Cancer 2019, 19, 30. [Google Scholar] [CrossRef]
- Jia, F.; Zhang, L.; Jiang, Z.; Tan, G.; Wang, Z. FZD1/KLF10-hsa-miR-4762-5p/miR-224-3p-circular RNAs axis as prognostic biomarkers and therapeutic targets for glioblastoma: a comprehensive report. BMC Med Genom. 2023, 16, 21. [Google Scholar] [CrossRef]
- Li, Z.; Lu, J.; Zeng, G.; Pang, J.; Zheng, X.; Feng, J.; Zhang, J. MiR-129-5p inhibits liver cancer growth by targeting calcium calmodulin-dependent protein kinase IV (CAMK4). Cell Death Dis. 2019, 10, 789. [Google Scholar] [CrossRef]
- Khalilian, S.; Imani, S.Z.H.; Ghafouri-Fard, S. Emerging roles and mechanisms of miR-206 in human disorders: a comprehensive review. Cancer Cell Int. 2022, 22, 412. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, S.; Liu, D.; Wang, Z.; Zhu, Y.; Li, J.; Xu, K.; Li, F.; Wen, H.; Yang, R. Exosomal miR-4645-5p from hypoxic bone marrow mesenchymal stem cells facilitates diabetic wound healing by restoring keratinocyte autophagy. Burn. Trauma 2024, 12, tkad058. [Google Scholar] [CrossRef]
- Chen, E.-B.; Zhou, Z.-J.; Xiao, K.; Zhu, G.-Q.; Yang, Y.; Wang, B.; Zhou, S.-L.; Chen, Q.; Yin, D.; Wang, Z.; et al. The miR-561-5p/CX3CL1 Signaling Axis Regulates Pulmonary Metastasis in Hepatocellular Carcinoma Involving CX3CR1+ Natural Killer Cells Infiltration. Theranostics 2019, 9, 4779–4794. [Google Scholar] [CrossRef]
- Nakano, T.; Chen, C.-L.; Chen, I.-H.; Tseng, H.-P.; Chiang, K.-C.; Lai, C.-Y.; Hsu, L.-W.; Goto, S.; Lin, C.-C.; Cheng, Y.-F. Overexpression of miR-4669 Enhances Tumor Aggressiveness and Generates an Immunosuppressive Tumor Microenvironment in Hepatocellular Carcinoma: Its Clinical Value as a Predictive Biomarker. Int. J. Mol. Sci. 2023, 24, 7908. [Google Scholar] [CrossRef] [PubMed]
- Sathipati, S.Y.; Tsai, M.-J.; Aimalla, N.; Moat, L.; Shukla, S.K.; Allaire, P.; Hebbring, S.; Beheshti, A.; Sharma, R.; Ho, S.-Y. An evolutionary learning-based method for identifying a circulating miRNA signature for breast cancer diagnosis prediction. NAR Genom. Bioinform. 2024, 6, lqae022. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Najafi, S.; Hussen, B.M.; Ganjo, A.R.; Taheri, M.; Samadian, M. DLX6-AS1: A Long Non-coding RNA With Oncogenic Features. Front. Cell Dev. Biol. 2022, 10, 746443. [Google Scholar] [CrossRef] [PubMed]
- Schröder, S.; Fuchs, U.; Gisa, V.; Pena, T.; Krüger, D.M.; Hempel, N.; Burkhardt, S.; Salinas, G.; Schütz, A.-L.; Delalle, I.; et al. PRDM16-DT is a novel lncRNA that regulates astrocyte function in Alzheimer’s disease. Acta Neuropathol. 2024, 148, 32. [Google Scholar] [CrossRef]
- Zhao, Q.; Cumming, H.; Cerruti, L.; Cunningham, J.M.; Jane, S.M. Site-specific Acetylation of the Fetal Globin Activator NF-E4 Prevents Its Ubiquitination and Regulates Its Interaction with the Histone Deacetylase, HDAC1. J. Biol. Chem. 2004, 279, 41477–41486. [Google Scholar] [CrossRef]
- Xian, J.; Wu, W.; Chen, X.; Bao, H.; Zhang, S.; Sheng, X.; Chen, S. SNORD99 promotes endometrial cancer development by inhibiting GSDMD-mediated pyroptosis through 2'-O-methylation modification. J. Cell. Mol. Med. 2024, 28, e18500. [Google Scholar] [CrossRef]
- Shang, X.; Song, X.; Wang, K.; Yu, M.; Ding, S.; Dong, X.; Xie, L.; Song, X. SNORD63 and SNORD96A as the non-invasive diagnostic biomarkers for clear cell renal cell carcinoma. Cancer Cell Int. 2021, 21, 56. [Google Scholar] [CrossRef]
- Liang, J.; Wen, J.; Huang, Z.; Chen, X.-P.; Zhang, B.-X.; Chu, L. Small Nucleolar RNAs: Insight Into Their Function in Cancer. Front. Oncol. 2019, 9, 587. [Google Scholar] [CrossRef]
- Jiang, J.; Hu, H.; Chen, Q.; Zhang, Y.; Chen, W.; Huang, Q.; Chen, X.; Li, J.; Zhong, M. Long non-coding RNA SNHG29 regulates cell senescence via p53/p21 signaling in spontaneous preterm birth. Placenta 2021, 103, 64–71. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, J.; Liu, X.; Li, S.; Wang, Q.; Chen, D.; Hu, Z.; Yu, T.; Ding, J.; Li, J.; et al. The LINC01138 drives malignancies via activating arginine methyltransferase 5 in hepatocellular carcinoma. Nat. Commun. 2018, 9, 1572. [Google Scholar] [CrossRef]
- Yang, Y.; Ren, M.; Song, C.; Li, D.; Soomro, S.H.; Xiong, Y.; Zhang, H.; Fu, H. LINC00461, a long non-coding RNA, is important for the proliferation and migration of glioma cells. Oncotarget 2017, 8, 84123–84139. [Google Scholar] [CrossRef]
- Lin, X.; Ding, J.-M.; Zheng, X.-Z.; Chen, J.-G. Immunity-related long noncoding RNA WDFY3-AS2 inhibited cell proliferation and metastasis through Wnt/β-catenin signaling in oral squamous cell carcinoma. Arch. Oral Biol. 2023, 147, 105625. [Google Scholar] [CrossRef]
- John, S.; Bhowmick, K.; Park, A.; Huang, H.; Yang, X.; Mishra, L. Recent advances in targeting obesity, with a focus on TGF-β signaling and vagus nerve innervation. Bioelectron. Med. 2025, 11, 10. [Google Scholar] [CrossRef]
- Iruzubieta, P.; Jimenez-Gonzalez, C.; Cabezas, J.; Crespo, J. From NAFLD to MASLD: transforming steatotic liver disease diagnosis and management. Metab. Target Organ Damage 2025, 5. [Google Scholar] [CrossRef]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Sunny, N.E.; Parks, E.J.; Browning, J.D.; Burgess, S.C. Excessive Hepatic Mitochondrial TCA Cycle and Gluconeogenesis in Humans with Nonalcoholic Fatty Liver Disease. Cell Metab. 2011, 14, 804–810. [Google Scholar] [CrossRef]
- Wang, S.; Chen, Y.; Lei, G.; Ma, X.; An, L.; Wang, H.; Song, Z.; Lin, L.; He, Q.; Xu, R.; et al. Serum Exosome–Derived microRNA-193a-5p and miR-381-3p Regulate Adenosine 5'-Monophosphate–Activated Protein Kinase/Transforming Growth Factor Beta/Smad2/3 Signaling Pathway and Promote Fibrogenesis. Clin. Transl. Gastroenterol. 2024, 15, e00662. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Transcriptomic Profiling miRNA and other ncRNAs from whole blood. (A) Volcano plot showing differential expression micro-RNA (log2 of fold-change; x axis) and statistical significance of this change (log10 of significance; y axis) in comparison of MASLD cases compared to the control group. (B) Volcano plot showing differential expression other ncRNAs (log2 of fold-change; x axis) and statistical significance of this change (log10 of significance; y axis). Colored points represent differentially expressed miRNAs and other ncRNAs (cutoff FDR 0.05) with magnitude of change 1.5 that are either overexpressed (blue) or under expressed (green). Most significant are labeled.
Figure 1.
Transcriptomic Profiling miRNA and other ncRNAs from whole blood. (A) Volcano plot showing differential expression micro-RNA (log2 of fold-change; x axis) and statistical significance of this change (log10 of significance; y axis) in comparison of MASLD cases compared to the control group. (B) Volcano plot showing differential expression other ncRNAs (log2 of fold-change; x axis) and statistical significance of this change (log10 of significance; y axis). Colored points represent differentially expressed miRNAs and other ncRNAs (cutoff FDR 0.05) with magnitude of change 1.5 that are either overexpressed (blue) or under expressed (green). Most significant are labeled.
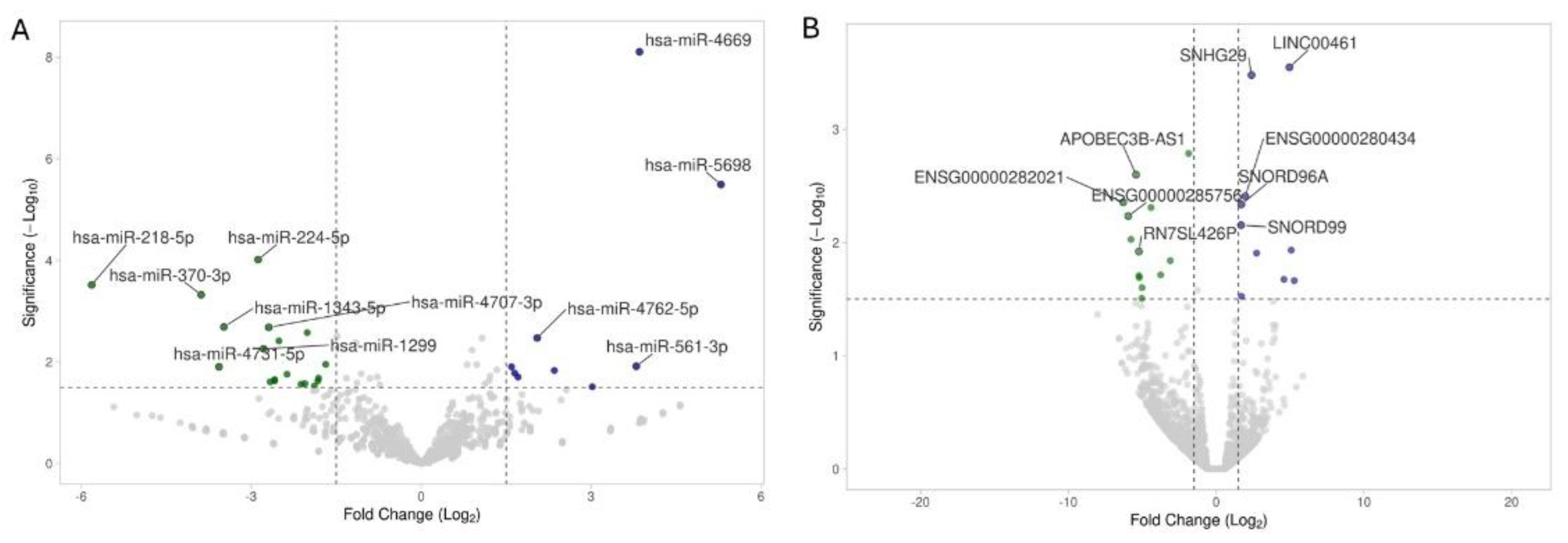
Figure 2.
Volcano plot showing differential expression miRNAs (log2 of fold-change; x-axis) and statistical significance of this change (log10 of significance; y-axis) in comparison of each MASLD case compared to the control group. Colored points represent differentially expressed miRNAs (cutoff FDR 0.05) with magnitude of change 1.5 that are either overexpressed (red) or under expressed (blue). Common miRNAs are labeled.
Figure 2.
Volcano plot showing differential expression miRNAs (log2 of fold-change; x-axis) and statistical significance of this change (log10 of significance; y-axis) in comparison of each MASLD case compared to the control group. Colored points represent differentially expressed miRNAs (cutoff FDR 0.05) with magnitude of change 1.5 that are either overexpressed (red) or under expressed (blue). Common miRNAs are labeled.
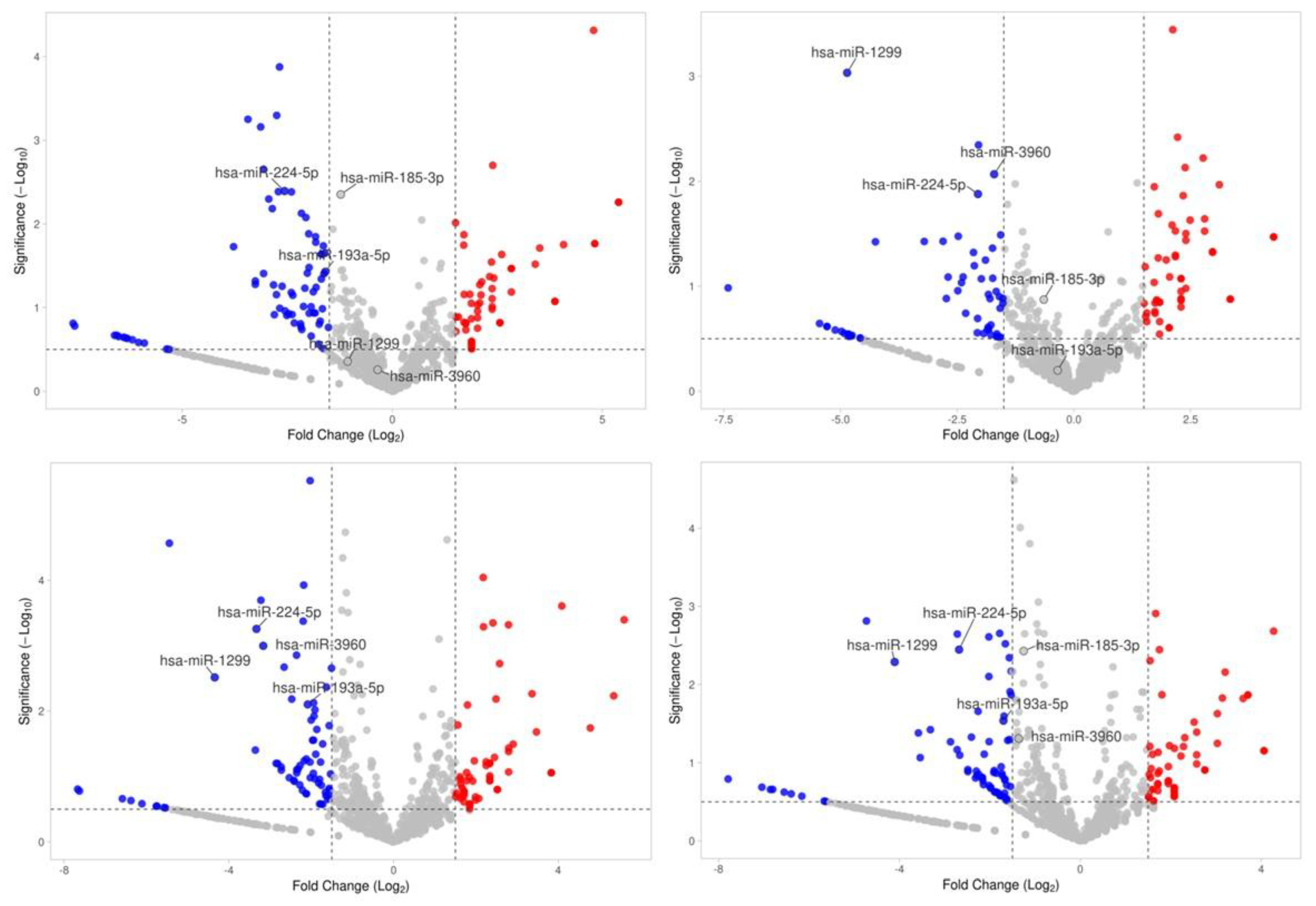
Figure 3.
Ingenuity Pathway Analysis (IPA)-identified differentially expressed miRNA connected to fibrosis and cirrhosis of the liver. (A) Differentially expressed miRNAs connected with fibrosis. (B) Differentially expressed miRNA connected with cirrhosis.
Figure 3.
Ingenuity Pathway Analysis (IPA)-identified differentially expressed miRNA connected to fibrosis and cirrhosis of the liver. (A) Differentially expressed miRNAs connected with fibrosis. (B) Differentially expressed miRNA connected with cirrhosis.
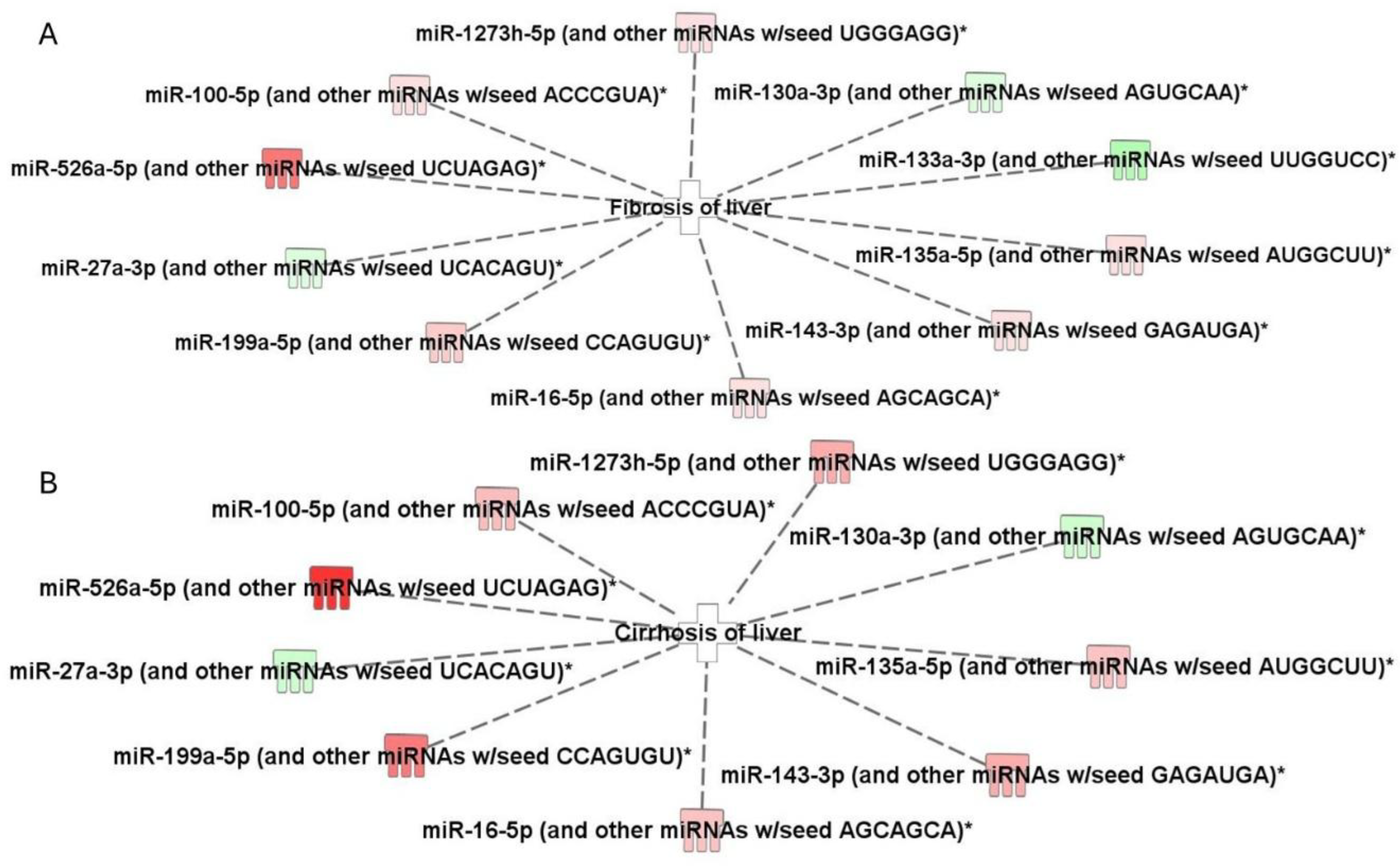
Figure 4.
Figure 4. Network of differentially expressed (p<0.05) other ncRNAs in the MASLD participants, relative to control group. The network was generated using Ingenuity Pathway Analysis (IPA) from QIAGEN, USA.
Figure 4.
Figure 4. Network of differentially expressed (p<0.05) other ncRNAs in the MASLD participants, relative to control group. The network was generated using Ingenuity Pathway Analysis (IPA) from QIAGEN, USA.
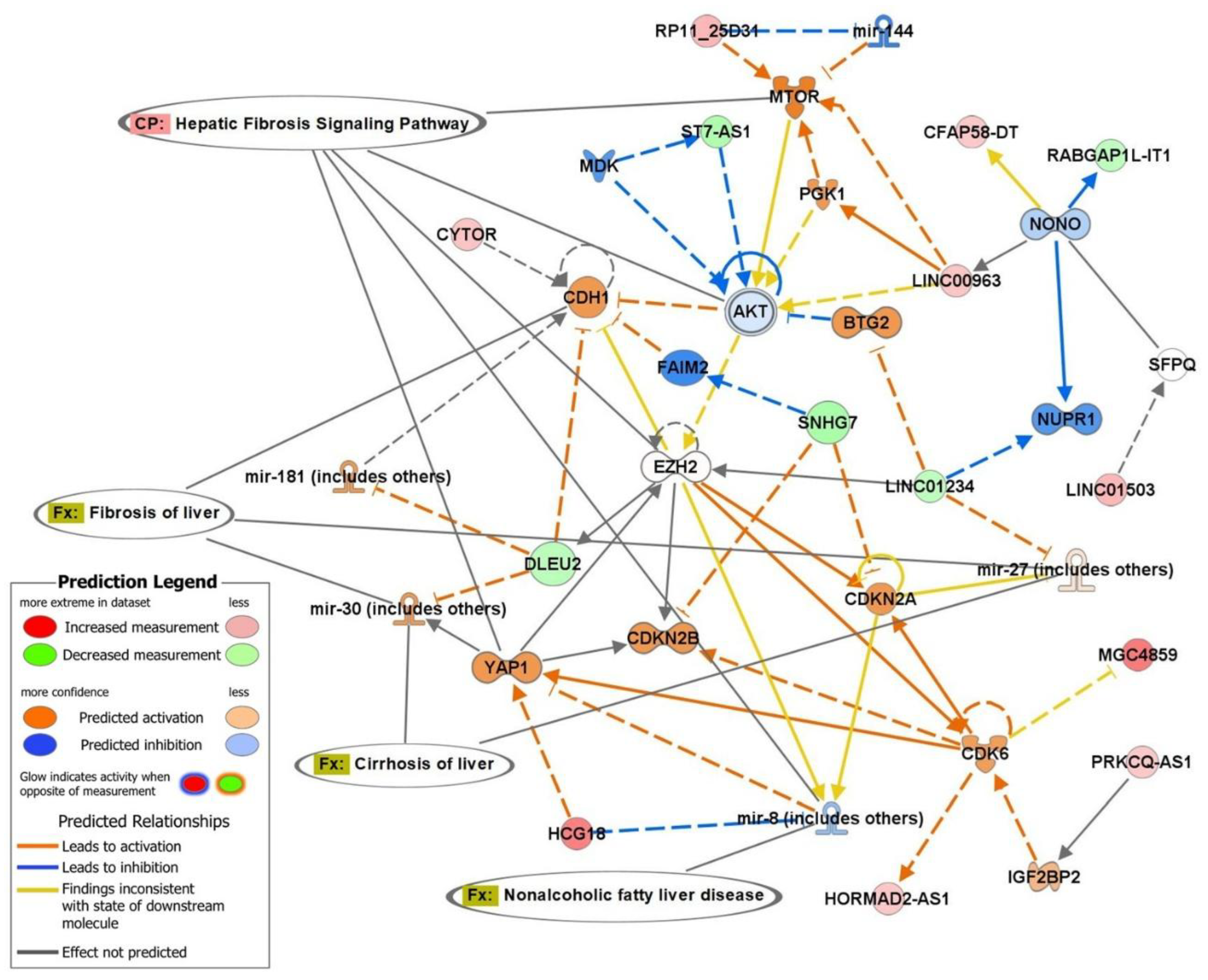
Figure 5.
Quantitative real-time PCR (qRT-PCR) of major miRNA which were significantly up or down regulated all MASLD sample (observed by global miRNA assay) compared to the control. For the validation study, we included 30 additional participants (14 from the MASLD group, 16 from the control group) and we used Applied Biosystem® pre-configured TaqMan array card (96 well array card format) to examine the expression of target of interest, viz., 18s (manufacturing control), GAPDH (internal control), miR-206, miR-1343-5p, miR-224-5p, miR1299, miR-193a-5p, miR185-3p, and miR-218-5p as identified based on the previous data. We selected only those patients who were at early stages of development MASLD (confirmed hepatic steatosis and exhibited one or more comorbid metabolic features, viz., type 2 diabetes, hypertension, hyperlipidemia, or obesity). Six samples per plate were analyzed by High Throughput FAST Real-Time PCR (Applied Biosystems, CA). qRT-PCR was carried out on a QuantStudio 7 Pro PCR system (Applied Biosystems, CA). Significance was determined using unpaired t-tests (p<0.05), and results are presented as mean ± SEM from triplicate experiments.
Figure 5.
Quantitative real-time PCR (qRT-PCR) of major miRNA which were significantly up or down regulated all MASLD sample (observed by global miRNA assay) compared to the control. For the validation study, we included 30 additional participants (14 from the MASLD group, 16 from the control group) and we used Applied Biosystem® pre-configured TaqMan array card (96 well array card format) to examine the expression of target of interest, viz., 18s (manufacturing control), GAPDH (internal control), miR-206, miR-1343-5p, miR-224-5p, miR1299, miR-193a-5p, miR185-3p, and miR-218-5p as identified based on the previous data. We selected only those patients who were at early stages of development MASLD (confirmed hepatic steatosis and exhibited one or more comorbid metabolic features, viz., type 2 diabetes, hypertension, hyperlipidemia, or obesity). Six samples per plate were analyzed by High Throughput FAST Real-Time PCR (Applied Biosystems, CA). qRT-PCR was carried out on a QuantStudio 7 Pro PCR system (Applied Biosystems, CA). Significance was determined using unpaired t-tests (p<0.05), and results are presented as mean ± SEM from triplicate experiments.
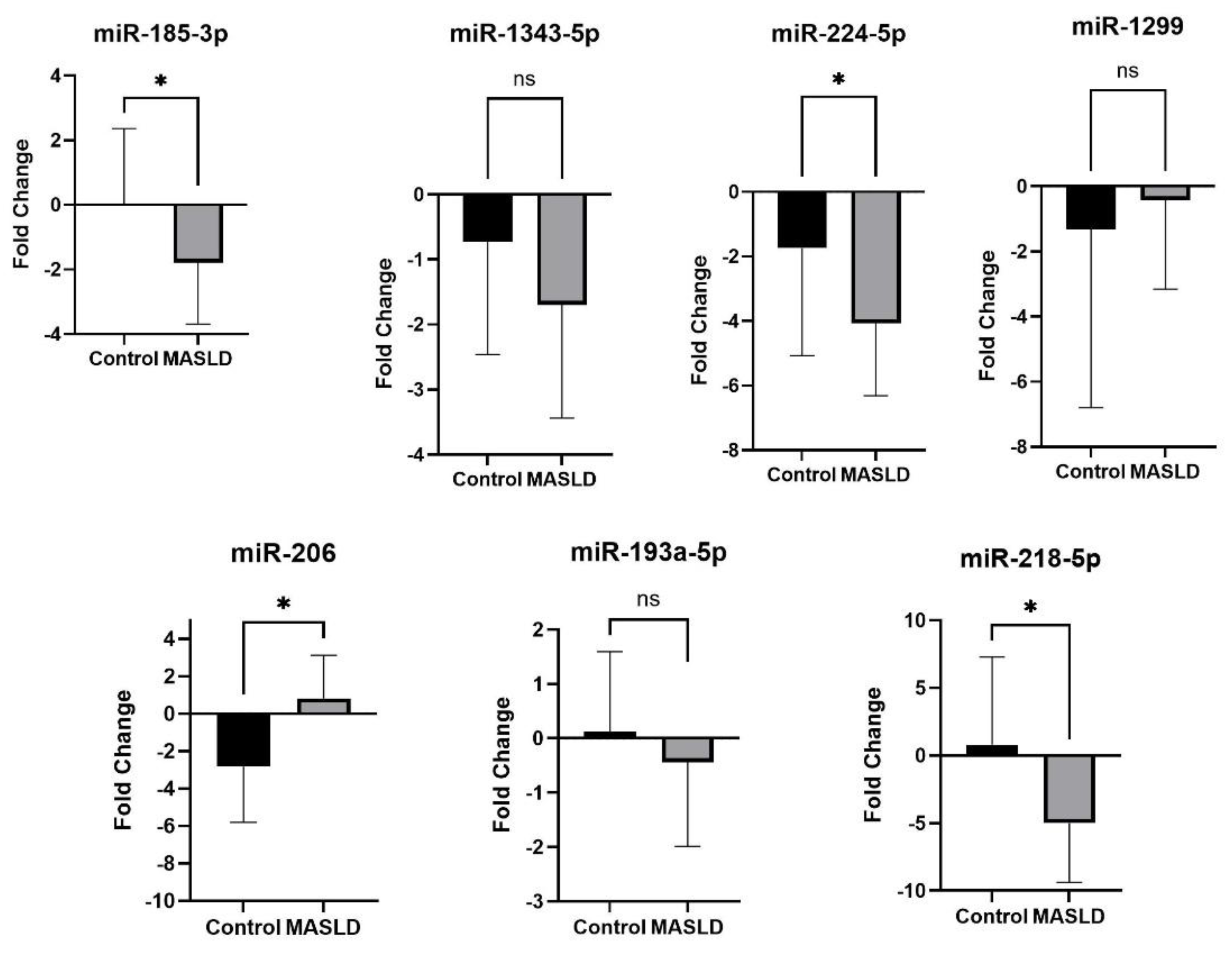
Table 1.
Participants Characteristics.
| Discovery Cohort | Validation Cohort | ||||||
| Control (n=4) | MASLD (n=4) | P-value | Control (n=16) | MASLD (n=14) | P-value | ||
| Age (years) | 61±4.83 | 53±5.94 | 0.08 | 53±12.97 | 46±8.36 | 0.08 | |
| Male/Female | 2/2 | 2/2 | - | 6/10 | 6/8 | - | |
| BMI (kg/m2) | 25.22±1.53 | 27.8±4.25 | 0.29 | 30.61±8.38 | 30.26±7.81 | 0.90 | |
| Hba1c (%) | 5.26±0.37 | 5.65±0.07 | 0.27 | 5.35±0.57 | 6.58±1.56 | 0.01 | |
| LDL (Optimal range <100 mg/dL) * | - | 137.34±26.65 | - | - | 102.5±49.17 | - | |
| HDL (Optimal range 40-70 mg/dL) * | - | 50.34±18.82 | - | - | 48.93±17.94 | - | |
| Triglyceride (Optimal range <150 mg/dL)* | - | 124±85.08 | - | - | 138.64±75.11 | - | |
| FibroScan** | - | Patient 1 – F0; Patient 2 – F2-F3; Patient 3 – F0-F1; Patient 4 – F0-F1 | - | - | Patient 5 – F2; Patient 6 – F4; Patient 7 – F0-F1; Patient 8 –F0-F1; Patient 9 – F3; Patient 10 – F2; Patient 11 – F2; Patient 12 – F0-F1; Patient 13 – F0; Patient 14 – F0; Patient 15 – F0-F1; Patient 16 – F2; Patient 17 – F2; Patient 18 – F0 | - | |
| Steatosis Stage*** | - | Patient 1 – S3; Patient 2 – S3; Patient 3 – S1; Patient 4 – S3 | - | - | Patient 5 – S1-S2; Patient 6 – S1-S2; Patient 7 – S3; Patient 8 – S0; Patient 9 – S0; Patient 10 – S3; Patient 11 – S2; Patient 12 – S2; Patient 13 – S2-S3; Patient 14 – S0; Patient 15 – S0; Patient 16 – S3; Patient 17 – S3; Patient 18 – S0 | - | |
*LDL, HDL, triglyceride, FibroScan, and steatosis data were not collected for control participants because these parameters were within normal reference ranges according to their medical records.**F0-Normal, F1-Mild fibrosis, F3-Moderate fibrosis, and F4-Severe fibrosis. ***S0- steatosis less than 11% (normal), S1- steatosis 11% to 33%, S2- steatosis 34% to 66%, S3- steatosis greater than 67%.
Table 2.
Differentially expressed miRNAs in MASLD subjects compared to controls.
| miRNA | Fold Change | P-Value | Biological Functions |
| hsa-miR-218-5p | -5.82 | 0.0003 | Regulates placental development, airway inflammation, and hepatic lipogenesis; targets TGFβ2, SMAD2, TLR4, Elovl5. [41] |
| hsa-miR-370-3p | -3.88 | 0.0005 | Regulates VSMC phenotype, glioblastoma suppression, and sinus node dysfunction in heart failure. [42] |
| hsa-miR-4731-5p | -3.57 | 0.0125 | Tumor suppressor in glioblastoma, melanoma, and NSCLC; impacts viability, EMT, and apoptosis. [43] |
| hsa-miR-1343-5p | -3.48 | 0.0020 | Reduces TGF-β signaling and fibrosis via exosomal delivery; therapeutic potential in lung disease. [28] |
| hsa-miR-224-5p | -2.88 | 0.0001 | Promotes EMT in hepatocellular carcinoma, regulates autophagy in breast cancer, and modulates cardiovascular inflammation. [44] |
| hsa-miR-193a-5p | -2.80 | 0.0031 | Tumor suppressor; inhibits proliferation and metastasis in ovarian and prostate cancers. [45] |
| hsa-miR-1299 | -2.78 | 0.0055 | Tumor suppressor; inhibits NEK2 in prostate cancer, also regulates RHOT1 and PDL1 in other cancers. [46] |
| hsa-miR-4707-3p | -2.69 | 0.0021 | Modulates cell fate in human neocortex development. [47] |
| hsa-miR-133a-3p | -2.67 | 0.0247 | Tumor suppressor in colorectal cancer; inhibits angiogenesis. [48] |
| hsa-miR-365a-3p | -2.59 | 0.0236 | Promotes lung cancer via PI3K/AKT; affects osteogenesis by targeting RUNX2. [49] |
| hsa-miR-4664-5p | -2.59 | 0.0223 | Detected in breast cancer; potential cancer biomarker. [50] |
| hsa-miR-539-5p | -2.51 | 0.0039 | Inhibits pancreatic cancer proliferation; regulates Tregs in leukemia. [51] |
| hsa-miR-369-5p | -2.37 | 0.0175 | Inhibits hepatocellular carcinoma by targeting HOXA13. [52] |
| hsa-miR-150-3p | -2.12 | 0.0275 | Antitumor in lung cancer; enhances neuronal proliferation. [53] |
| hsa-miR-1185-1-3p | -2.05 | 0.0267 | Biomarker for weight loss response; associated with lung cancer. [54] |
| hsa-miR-3940-3p | -2.01 | 0.0026 | Promotes granulosa cell proliferation; linked to insulin resistance in pregnancy. [55] |
| hsa-miR-369-3p | -1.90 | 0.0373 | Anti-inflammatory; inhibits preadipocyte proliferation and differentiation. [56] |
| hsa-miR-452-5p | -1.89 | 0.0297 | Regulates fibrosis and promotes cancer progression. [57] |
| hsa-miR-323b-3p | -1.86 | 0.0363 | Upregulated in Huntington’s disease; involved in neurodegeneration. [58] |
| hsa-miR-433-3p | -1.82 | 0.0236 | Suppresses glioma growth; enhances chemotherapy sensitivity. [59] |
| hsa-miR-379-5p | -1.81 | 0.0209 | Plays a role in regulating cellular processes, particularly in cancer development and progression. [60] |
| hsa-miR-409-5p | -1.71 | 0.0480 | Promotes tumor growth, EMT, and bone metastasis in prostate cancer. [61] |
| hsa-miR-487b-3p | -1.69 | 0.0112 | Negative regulator of skeletal myogenesis; suppresses C2C12 myoblast proliferation. [62] |
| hsa-miR-154-5p | -1.65 | 0.0451 | Triggers cardiac oxidative stress and inflammation; tumor suppressor in glioblastoma. [63] |
| hsa-miR-3195 | 1.60 | 0.0125 | Suppresses osteosarcoma progression by targeting SOX4; linked to prostate cancer. [64] |
| hsa-miR-6758-5p | 1.65 | 0.0165 | Specific function remains unknown. |
| hsa-miR-4479 | 1.7 | 0.0198 | Potential biomarker in cancer; roles in immunosuppression and metastasis. [65] |
| hsa-miR-196a-5p | 1.7 | 0.0437 | Oncogene; promotes invasion, metastasis, and proliferation in many cancers. [66] |
| hsa-miR-4762-5p | 2.0 | 0.0034 | Detected in breast cancer tissues; role in tumorigenesis is under study. [67] |
| hsa-miR-129-5p | 2.35 | 0.0147 | Tumor suppressor; inhibits proliferation in hepatocellular carcinoma. [68] |
| hsa-miR-206 | 2.56 | 0.0353 | Involved in cancers, neurodegenerative, and cardiovascular diseases; tumor suppressor. [69] |
| hsa-miR-4645-5p | 3.02 | 0.0309 | Facilitates diabetic wound healing by restoring keratinocyte autophagy. [70] |
| hsa-miR-561-3p | 3.80 | 0.0122 | Modulates CX3CL1 signaling in hepatocellular carcinoma; suppresses metastasis. [71] |
| hsa-miR-4669 | 3.85 | <0.0001 | Enhances tumor aggressiveness creates immunosuppressive environment in liver cancer. [72] |
| hsa-miR-5698 | 5.29 | <0.0001 | Identified as breast cancer biomarker; functions not well characterized. [73] |
The above list of miRNAs includes those that are differentially expressed in the MASLD group (n=4) compared to the control group (n=4), with a fold change cutoff of ±1.5 (or at least 1.5-fold) and a p-value < 0.05.
Table 3.
Differentially expressed other ncRNAs compared to controls.
| Other ncRNA | Fold Change | P-Value | Biological Functions |
| Homo_sapiens_tRNA-Leu-AAG-1 | -8.03 | 0.043 | Encodes a tRNA specific for leucine with the AAG anticodon, essential for protein synthesis. |
| ENSG00000282021 | -6.29 | 0.004 | Specific function remains unknown. |
| ENSG00000285756 | -5.95 | 0.006 | Specific function remains unknown. |
| DLX6-AS1 | -5.76 | 0.009 | Long non-coding RNA implicated in promoting tumor cell proliferation, migration, invasion, and epithelial-mesenchymal transition in various cancers. [74] |
| FMNL1-DT | -5.44 | 0.034 | Specific function remains unknown. |
| APOBEC3B-AS1 | -5.42 | 0.003 | Specific function remains unknown. |
| RN7SL426P | -5.23 | 0.012 | Specific function remains unknown. |
| ENSG00000254639 | -5.23 | 0.020 | Specific function remains unknown. |
| RSF1-IT1 | -5.20 | 0.020 | Specific function remains unknown. |
| ENSG00000273064 | -5.07 | 0.036 | Specific function remains unknown. |
| PRDM16-DT | -5.03 | 0.031 | Long non-coding RNA involved in regulating astrocyte function and implicated in colorectal cancer metastasis and drug resistance. [75] |
| RNU6-70P | -5.02 | 0.025 | Specific function remains unknown. |
| Homo_sapiens_tRNA-Gly-GCC-5 | -4.41 | 0.005 | Encodes a tRNA specific for glycine with the GCC anticodon, essential for protein synthesis. |
| U8 | -3.75 | 0.019 | Specific function remains unknown. |
| NFE4 | -3.11 | 0.014 | Transcription factor involved in regulating fetal γ-globin gene expression. Acetylation of NFE4 prevents its ubiquitination and modulates its interaction with histone deacetylase HDAC1, influencing gene activation. [76] |
| Homo_sapiens_tRNA-Met-CAT-6 | -1.95 | 0.037 | Encodes transfer RNA for methionine with anticodon CAT, essential for initiating protein synthesis. |
| Homo_sapiens_tRNA-Asp-GTC-2 | -1.86 | 0.002 | Encodes transfer RNA for aspartic acid with anticodon GTC, facilitating incorporation of aspartic acid during protein synthesis. |
| SNORD99 | 1.69 | 0.007 | Small nucleolar RNA involved in 2'-O-methylation of ribosomal RNA. Overexpression promotes endometrial cancer development by inhibiting GSDMD-mediated pyroptosis. [77] |
| SNORD96A | 1.71 | 0.005 | Small nucleolar RNA implicated in ribosomal RNA modification. Elevated levels in plasma serve as a non-invasive diagnostic biomarker for clear cell renal cell carcinoma (ccRCC). [78] |
| SNORD48 | 1.71 | 0.030 | Small nucleolar RNA involved in post-transcriptional modification of other small nuclear RNAs. Associated with prostate and hematologic cancers. [79] |
| ENSG00000280434 | 1.97 | 0.004 | Specific function remains unknown. |
| SNHG29 | 2.40 | 0.000 | Long non-coding RNA that regulates cell senescence via p53/p21 signaling and promotes glioblastoma progression through the miR-223-3p/CTNND1 axis. [80] |
| LINC01138 | 2.74 | 0.012 | Long intergenic non-coding RNA that acts as an oncogenic driver by interacting with PRMT5, enhancing its stability, and promoting tumorigenicity in hepatocellular carcinoma. [81] |
| ENSG00000253374 | 3.86 | 0.033 | Specific function remains unknown. |
| RN7SL33P | 4.58 | 0.021 | Specific function remains unknown. |
| LINC00461 | 4.96 | 0.000 | Long non-coding RNA important for glioma progression, affecting cell proliferation, migration, and invasion via MAPK/ERK and PI3K/AKT signaling pathways. [82] |
| ENSG00000286834 | 5.09 | 0.012 | Specific function remains unknown. |
| WDFY3-AS2 | 5.28 | 0.022 | Long non-coding RNA that acts as a tumor suppressor by inhibiting cell proliferation and metastasis through the Wnt/β-catenin signaling pathway in oral squamous cell carcinoma. [83] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



