
Submitted:
05 June 2025
Posted:
06 June 2025
You are already at the latest version
Abstract
Artocarpus heterophyllus, familiar as jackfruit, is a tropical fruit highly valued not only for its nutritional content but also for its medicinal properties, including potential antidiabetic effects. In this study, ethanolic extract of unripe Artocarpus heterophyllus (EEAH) exhibited significant antidiabetic activity through multiple mechanisms. EEAH enhanced glucose-stimulated insulin secretion in BRIN-BD11 β-cells via KATP channel modulation and cAMP-mediated pathways, with partial dependence on extracellular calcium, and it also promoted β-cell proliferation. In vitro assays revealed its ability to inhibit starch digestion and glucose diffusion, indicating delayed carbohydrate digestion and absorption. In high-fat-fed (HFF) obese mice, Acute and chronic oral administration of EEAH improved oral glucose tolerance, reduced fasting blood glucose, decreased body weight, and normalized food and fluid intake. Lipid profile analysis showed increased HDL and reduced total cholesterol, LDL, and triglycerides, while higher doses of EEAH also enhanced gut motility. Phytochemical screening revealed the presence of bioactive compounds such as alkaloids, tannins, flavonoids, saponins, steroids and, terpenoids, which are likely responsible for these therapeutic effects. These findings highlight EEAH as a promising natural candidate for adjunctive therapy in managing type 2 diabetes and associated metabolic disorders and emphasize the importance of future multi-omics studies to elucidate its molecular targets and pathways.
Keywords:
insulin resistance
; hyperglycaemia
; type 2 diabetes mellitus
; beta-cell proliferation
; lipids
; Artocarpus heterophyllus
1. Introduction
Diabetes mellitus (DM), is a complex multifactorial metabolic disorder, characterized by excessive glucose in the circulation resulting from an impaired insulin sensitivity, deficient secretion of the anabolic hormone insulin, or both [1,2]. Globally, DM presents a significant public health challenge; the International Diabetes Federation (IDF) estimated that approximately 537 million people had DM in 2021, and this number is projected to rise to 783 million by 2045 [3]. Nearly, 10% of adults worldwide are diagnosed with diabetes mellitus (DM), being a major contributor to increased morbidity and mortality [4].
The development and progression of diabetes fundamentally stem from inadequate insulin production or impaired insulin action, both of which are critical for maintaining metabolic balance. Insulin is an essential peptide hormone produced by β-cells, plays a crucial role in regulating metabolic processes [5,6]. Its synthesis begins with the transcription of the insulin gene into mRNA, followed by translation into pre-proinsulin. This precursor is subsequently processed, first into proinsulin and then further into mature insulin and C-peptide. [6,7]. Insulin promotes glucose uptake in skeletal muscle by facilitating its absorption and conversion to glycogen, and it also stimulates glycogen storage and inhibits excessive glucose production in the liver [8].
Diabetes presents with diverse phenotypes. Type 1 diabetes (T1D) is an autoimmune disorder marked by the destruction of pancreatic β-cells, causing absolute insulin deficiency and dysregulated α-cell activity that leads to excessive glucagon secretion. [1,2,9,10]. In contrast, type 2 diabetes mellitus (T2DM), accounts for 90% of diabetes cases, is a heterogeneous condition primarily driven by impaired insulin secretion from pancreatic β-cells and reduced insulin function due to insulin resistance [11,12]. Acute complications of diabetes include life-threatening conditions such as diabetic ketoacidosis (DKA) and severe hypoglycemia. Chronic hyperglycemia leads to both microvascular damage (retinopathy, nephropathy, and neuropathy) and macrovascular complication, thereby increasing the risk of diabetic foot ulcers and premature mortality [13].
Research indicates that obesity is a major risk factor for the development of type 2 diabetes mellitus (T2DM), with approximately 90% of individuals with the disease being overweight or obese [14]. While diet and exercise aid weight loss in T2DM, they often fall short in sustaining optimal glycemic control [15]. Therefore, a range of antidiabetic medications, including metformin, sulfonylureas, meglitinides, thiazolidinediones, SGLT2 inhibitors, GLP-1 mimetics, and DPP-IV inhibitors, are essential for effective blood glucose management. However, these medications are often unaffordable and inaccessible for individuals in underprivileged communities and can also have undesirable side effects [16]. Consequently, affordable and effective alternatives are needed to mitigate the detrimental effects of uncontrolled diabetes [15,17].
Traditionally, people worldwide have relied on various herbs and medicinal plants to manage and treat diabetes mellitus [15,17]. Artocarpus heterophyllus, generally known as jackfruit, which belongs to member of the Moraceae family and is cultivated in tropical regions particularly in countries such as India, Bangladesh, Sri Lanka, Thailand, Indonesia, the Philippines, Brazil, and parts of East and West Africa [18,19,20,21,22,23,24]. It has been utilized by indigenous people for centuries to treat diverse illnesses including diabetes, inflammation, infection, cancer, and CVD due to its innumerable medicinal properties [19,20]. A. heterophyllus is reported to be abundant in dietary fiber, carbohydrates, minerals, carboxylic acids and vitamins such as thiamine and ascorbic acid [18,21]. Previous studies have shown that A. heterophyllus possesses antioxidant, anti-inflammatory, anti-fungal, anti-diabetic, anti-bacterial, immunomodulatory and wound healing properties [20,22]. Recent studies suggested that A. heterophyllus lowers blood glucose, improves oral glucose tolerance, and body weight [23,24,25]. Based on these studies, our present aim was to investigate the insulinotropic and antidiabetic properties of A. heterophyllus, as well as the underlying mechanisms of action on pancreatic β-cells, using both in vitro and in vivo models.
2. Materials and Methods
2.1. Preparation and Extraction of EEAH
Green Artocarpus heterophyllus (unripe jackfruit) was acquired from Jahangirnagar University, Savar, Dhaka, Bangladesh and botanical accession number 87,070 was endorsed by botanical taxonomist of Bangladesh National Herbarium, Mirpur, Dhaka. The unripe jackfruit was meticulously cleaned, allowed to air dry for several hours and afterwards dried further in an oven at 45°-50° before emanating to ethanolic extract preparation. The dried components were further processed and subsequently ground into a fine powder using a grinding machine. Two hundred grams of the powder were then added to 1 liter of 80% (v/v) ethanol and placed on an orbital shaker at 900 rpm for 48-72 h. The resulting mixture was then filtered using Whatman No. 1 filter paper, and the filtrate was dried under vacuum using a rotary evaporator. The final extracted semi-solid, gummy crude extract of A. Heterophyllus was stored at 4°C until experimental use [26].
2.2. Studies of Insulin Release In Vitro
Clonal pancreatic BRIN-BD11 β-cells were used to analyze the impact of EEAH on insulin release in vitro [27]. With multiple varying concentrations (8–5000 µg/ml) of EEAH, BRIN-BD11 β-cells were incubated at 37°C for 20 min at 5.6 mM glucose [28]. The supernatants were collected and stored at -20 °C for insulin radioimmunoassay [29]. To determine the secretory pathways activated by EEAH, its effects were evaluated in the presence of various insulin secretagogues and inhibitors. These included tolbutamide (a sulfonylurea and KATP channel blocker), IBMX (a phosphodiesterase inhibitor), 30 mM KCl, 10 mM alanine, diazoxide (a KATP channel opener), and verapamil (a voltage-dependent Ca2+ channel blocker). Both KCl and alanine induce plasma membrane depolarization and Ca2+ influx, with alanine primarily achieving this through Na+ co-transport and ATP-generating metabolism. [17,30].
2.3. β-Cell Proliferation In Vitro
To investigate the effects of ethanol extract of A. heterophyllus (EEAH) on β-cell proliferation, BRIN-BD11 cells were treated with EEAH at concentrations of 40 and 200 µg/mL, and glucagon-like peptide-1 (GLP-1) (10-6 M), as a positive control. Ki-67 immunostaining was used to quantify cell proliferation. Briefly, cells were seeded onto coverslips (40,000 cells/coverslip) and incubated with EEAH or GLP-1 for 18 h at 37°C. After washing with phosphate-buffered saline (PBS), cells were fixed with 4% paraformaldehyde. Antigen retrieval was performed using citrate buffer (90°C, 20 min), and non-specific binding was blocked with 1.1% bovine serum albumin (BSA) for 30 min. Cells were then incubated with a Ki-67 primary antibody (1:500; Abcam, ab15580), followed by an Alexa Fluor® 488 secondary antibody (1:400, Invitrogen, A-11008). Coverslips were washed with PBS, mounted onto slides, and visualized using an Olympus System fluorescence microscope equipped with a DP70 camera adapter. Proliferation was quantified as the percentage of Ki-67-positive cells relative to the total number of cells analyzed [31].
2.4. In Vitro Starch Digestion
The effect of EEAH on starch digestion was examined by mixing 100 mg/50ml starch solution (Sigma-Aldrich, St. Louis, MO, USA) with/without EEAH, and acarbose. Following dilution, the mixtures were incubated with 0.01% heat stable α-amylase from Bacillus leicheniformis and 0.1% amyloglucosidase from Rhizopus sp. (Sigma-Aldrich, St. Louis, MO, USA) for 20 and 30 min at 80°C and 60°C respectively. Afterwards, the samples were collected and stored for analysing the concentration of glucose using liquid Glucose Oxidase-phenol 4 aminophenazone (GOD-PAP) method (Randox GL 2623) [28,29]. Alpha glucosidase inhibitor, Acarbose was used as a reference drug.
2.5. In Vitro Glucose Diffusion
The implications of EEAH on glucose diffusion in vitro were examined utilizing a cellulose ester dialysis tube (CEDT) (20 cm x 7.5 mm, Spectra/Por®CE layer, MWCO: 2000, Spectrum, Amsterdam, The Netherlands). In a dialysis tube, 2 mL of 0.9% NaCl was added along with 220 mM glucose in the presence or absence of EEAH. The tubes were tightly sealed and then placed into 50 mL centrifuge tubes (Orange Scientific, Orange, CA, USA) filled with 45 mL of 0.9% NaCl. Subsequently, the tubes were agitated on an orbital shaker at 37°C for 24 hr. Following incubation, the samples were collected. The amounts of glucose diffused into the external solution was measured using GOD/PAP method (Randox GL 2623) [32,33].
2.6. Animals
Male Swiss albino mice, approximately two weeks old and weighing 20-25g, were obtained from the Animal Resources Facility, ICDDRB, Dhaka, Bangladesh. They were housed at a controlled temperature of 22±3°C and humidity of 55-65%, with ad libitum access to food and water, under an automated 12-h light/dark cycle in the Animal House at Independent University, Bangladesh. Following a one-week acclimation period, the mice were divided into two groups based on body weight: a normal control group (NC) fed a standard diet (10% fat, 30% protein, and 60% carbohydrate) and a high-fat-fed (HFF) group fed a high-fat-fed diet (19.4% powdered normal animal food, 17.5% sugar, 21% beef fat, 39.5% condense milk, 1.1% Vitamin-B complex and 1.5% salt) for 6 to 8 weeks. For subsequent experiments, the HFF group was further subdivided into four groups: a high-fat-fed diet control group (HFF) (Group 2), two treatment groups (Group 3 and 4), and a standard treatment group (Group 5). HFF diet-induced obese type 2 diabetic mice were characterized by fasting blood glucose levels exceeding the normal range (>6.0 mmol/L) and a body weight of 40-45g. The normal control and high-fat-fed diet control groups received saline (5 mL/kg). The treatment groups received 250 and 500 mg/5mL/kg of EEAH, respectively, and the standard treatment group received glibenclamide (5 mg/5mL/kg) twice daily (BID) for 60 days. Each group contained n=6 mice. This study was approved by the Institutional Review Board (IRB) at Independent University, Bangladesh (IUB) on September 1, 2022 for animal experiments. The experiments were conducted in accordance with the animal Welfare Act 2019 of Bangladesh, UK Animals (Scientific Procedures) Act 1986 and EU Directive 2010/63/EU. Experiments were performed under UK Home Office Animal license PIL1822, ensuring ethical animal care and minimizing distress. The groups were divided as follows:
- Group 1: Lean control (saline)
- Group 2: High fat fed diet control (saline)
- Group 3: High fat diet + EEAH (250 mg/5 mL/kg)
- Group 4: High fat diet + EEAH (500 mg/5 mL/kg)
- Group 5: High fat diet + Glibenclamide (5 mg/5 mL/kg)
2.7. Acute and Chronic Oral Glucose Tolerance Test
Oral glucose tolerance tests (OGTT) were performed on high-fat-fed (HFF) diet-induced obese mice at days 0, 15, 30, 45, and 60 of the 60-day study to monitor glucose handling over time. The OGTT conducted on day 0 was carried out prior to the initiation of treatment to assess baseline glucose tolerance, while subsequent tests evaluated the progression and potential therapeutic effects of the test agents. HFF diet mice were starved for 12h prior to the experiment. Blood glucose were measured using tail vein bleeding at particular time intervals prior to (0 min) and after (30, 60, 120 & 180-min) oral gavage of glucose (2.5 gm/5 mL/kg, body weight, control) with/without EEAH (250 and 500 mg/5 mL/kg, body weight) or STD, glibenclamide (5 mg/5 mL/kg, body weight). Blood glucose levels were determined using an Ascencia Contour glucose meter and glibenclamide, an insulin secretagogue, was used as the reference drug [12].
2.8. Chronic Study of Blood Glucose, Body Weight, Food and Fluid Intake
A 60-day long-term study was conducted on high-fat-diet-induced obese mice. Various parameters, including fasting blood glucose, body weight, food and fluid intake, were measured every three days. All groups, including the high-fat-diet-induced obese mice and the normal control group, were fasted for 12 h prior to these measurements. The treatment groups received EEAH at doses of 250 and 500 mg/5 mL/kg, respectively. The positive control group received glibenclamide (5 mg/5 mL/kg), while both the high-fat-diet control group and the normal control group received saline (5 mL/kg). An Ascensia Contour Blood Glucose Meter (Bayer, Newbury, UK) was used to measure blood glucose levels [12,30,34].
2.9. Lipid Profile Test
Throughout the 60-day study, high-fat-diet-induced obese mice received twice-daily oral treatments of either EEAH (250 and 500 mg/5 mL/kg) or glibenclamide (5 mg/ 5 mL/kg). A lipid profile test was conducted as previously described [15]. At the end of study, the mice were euthanized, and blood was collected from the heart using 5 mL syringes into heparinized microcentrifuge tubes (Sarstedt, Nümbrecht, Germany) to prevent coagulation. The blood samples were centrifuged at 12,000 rpm for 5 min, and the resulting plasma serum was separated. High-density lipoprotein (HDL), low-density lipoprotein (LDL), triglyceride (TG), and total cholesterol (TC) levels were then analysed using COD-PAP, GPO-PAP (Elabscience Biotechnology Co., Ltd., TX, USA), and CHOD-PAP (Biolabo SAS, Maizy, France) reagents with an automated analyzer [35,36].
2.10. Gut Motility In Vivo
At the end of the 60-day chronic study, mice were sacrificed and gastrointestinal motility was assessed by measuring the distance traveled by a barium sulfate (BaSO₄) milk solution in mice fasted for 20 h. One hour before administration of the BaSO₄ solution, the treatment groups received EEAH (250 and 500 mg/5 mL/kg), bisacodyl (10 mg/5 mL/kg), or loperamide (5 mg/5 mL/kg). The high-fat-fed (HFF) control and the normal control groups received only the BaSO₄ solution. Fifteen minutes after administration of the BaSO₄ milk solution, the mice were euthanized, and their entire small intestines were extracted. The BaSO₄ transit distance was then measured and expressed as a percentage of the total small intestine length (from the pylorus to the ileocecal junction) [37,38].
2.11. Phytochemical Screening
To ascertain the source of alkaloids, tannins, flavonoids, saponins, steroids, terpenoids, glycosides and reducing sugar, a phytochemical analysis of EEAH was performed. In order to determine the presence of alkaloids, 2 mL of EEAH was acidified with hydrochloric acid (HCl), followed by 1 mL of Dragendroff’s solution. The emergence of a red colour implies the presence of alkaloids. To determine the presence of tannins, 2 ml of EEAH was mixed with 10% lead acetate. A white precipitate formation upon mixing, confirming the presence of tannins. The presence of flavonoids was determined by heating a mixture of 1.5 mL of methanol and 4 mL of EEAH, followed by the addition of metal magnesium and a few drops of HCl to the heated mixture, which resulted in the development of pink colour. To evaluate the presence of saponins, 1 mL of EEAH was added to 9 mL of distilled water, resulting in the formation of a stable foam. The presence of steroids was analysed by adding 2 mL of EEAH with 10 mL of chloroform, followed by 1 mL of acetic anhydride, and 2 mL of sulphuric acid, leading in the formation of bluish-green colour. To test for terpenoids, 1g of EEAH was dissolved in 2ml of chloroform. Then, 3ml of strong sulfuric acid was carefully added to form a distinct layer. A reddish-brown color at the interface indicated the presence of terpenoids. The presence of glycoside was detected by mixing 1 mL of EEAH with a few drops of glacial acetic acid, ferric chloride, followed by concentrated sulfuric acid, which resulted in a bluish-green colour. In order to identify the presence of reducing sugars, 1 mL of EEAH, 1 mL of water and a few drops of Fehling’s reagent were heated, resulting in the formation of red-brick colour. [32,39,40].
2.12. Statistical Analysis
GraphPad Prism version 10 was used for statistical analysis. Data were assessed using a two-tailed unpaired Student's t-test or one-way or two-way analysis of variance (ANOVA) with a Bonferroni post hoc test. Results were presented as mean ± SEM, with statistical significance defined as p<0.05.
3. Results
3.1. Insulin Release with EEAH
At 5.6 mM glucose, EEAH significantly stimulated insulin secretion from BRIN-BD11 β-cells in a concentration-dependent manner across a range of 40–5000 µg/mL (p < 0.05 to p < 0.001; Figure 1A). Alanine (10 mM), served as a positive control, remarkably increased insulin secretion (p < 0.001; Figure 1A).
3.2. Insulin Release with EEAH, Known Modulators and Absence of Extracellular Ca2+
To elucidate the insulinotropic mechanisms of non-cytotoxic concentrations (200 µg/mL) of EEAH, further investigations were conducted (Figure 1B). As depicted in Figure 1B, EEAH-stimulated insulin secretion exhibited potentiation in the presence of key modulators: high glucose (16.7 mM, p < 0.01), the non-selective phosphodiesterase inhibitor IBMX (p < 0.001), and the sulfonylurea receptor 1 (SUR1) agonist tolbutamide (p < 0.001). The synergistic action of IBMX, leading to elevated intracellular cyclic AMP, and tolbutamide, promoting SUR1 closure and subsequent membrane depolarization, amplified both cAMP production and insulin secretion. Conversely, the KATP channel opener diazoxide and the L-type voltage-gated calcium channel (VGCC) antagonist verapamil attenuated EEAH-induced insulin release (p < 0.05-0.01; Figure 1B), as evidenced by reduced insulin secretion when incubated with Ca2+ free buffer (P < 0.05; Figure 1C). Notably, EEAH retained its capacity to stimulate insulin secretion from cells depolarized with 30 mM KCl (p < 0.001; Figure 1B), indicating a mechanism downstream of membrane potential. However, the absence of extracellular Ca2+ diminished, though did not completely abrogate, EEAH's insulin-releasing activity (p < 0.001; Figure 1C), underscoring the partial dependence on extracellular calcium for its full insulinotropic effect.
3.3. Proliferation In Vitro with EEAH
Figure 2A-D displays BRIN BD11 β-cell proliferation images for three conditions: (A) Control, (B) GLP-1 (10-6 M), and (C,D) EEAH (at concentrations of 40 and 200 μg/ml). Quantitative analysis (Figure 2E) revealed that EEAH significantly enhanced BRIN-BD11 β-cell proliferation at both 40 μg/ml (p<0.01) and 200 μg/ml (p<0.001) compared to the control. The positive control, GLP-1 (10-6 M), demonstrated the most potent effect on BRIN-BD11 β-cell growth (p<0.001; Figure 2E).
3.4. Starch Digestion In Vitro with EEAH
Figure 1D illustrates the impact of Acarbose and EEAH on starch digestion. The positive control, Acarbose (62.5–1000 µg/mL), demonstrated a concentration-dependent reduction in starch digestion ranging from 20% to 80% (Data not shown). EEAH also significantly inhibited starch digestion (11–33%, p<0.05–0.001, Figure 1D) at higher concentrations (250–1000 µg/mL).
3.5. Glucose Diffusion In Vitro with EEAH
3.6. Oral Glucose Tolerance Test with EEAH
Following oral gavage of EEAH (250 and 500 mg/5 mL/kg) in conjunction with glucose (2.5 gm/5 mL/kg), significantly (p <0.05-0.001) improved oral glucose tolerance at 30, 60, and 120 min on 0, 15, 30, and 60 days of studies (Figure 3A-D) in HFF diet induced obese mice as compared to high-fat-fed diet induced obese control mice. Treatment with EEAH at doses of 250 and 500 mg/5 mL/kg improved oral glucose tolerance by 1.2-fold and 1.4-fold at 30 and 60 min, respectively, consistently throughout the studies. In comparison to the HFF diet induced obese control mice, the standard drug glibenclamide (5 mg/5 mL/kg) also significantly (p <0.05-0.001) decreased blood glucose levels at 30, 60, and 120 min on days 0, 15, 30, and 60 (Figure 3A-D).
3.7. Chronic Study of Fasting Blood Glucose, Body Weight, Food and Fluid Intake with EEAH
Oral administration of EEAH (250 and 500 mg/5 mL/kg) and glibenclamide (5 mg/5 mL/kg) twice-daily showed remarkable reductions in fasting blood glucose levels in HFF diet induced obese mice during the study period of 60 days (Figure 4A). In comparison to HFF alone, EEAH at doses of 250 and 500 mg/5 mL/kg considerably (p < 0.05-0.001) lowered blood glucose levels from day 27 and onwards, with 500 mg/5 mL/kg dose showing greater potency than 250 mg/5 mL/kg dose. Additionally, the standard drug glibenclamide (5 mg/5 mL/kg) consistently decreased (p < 0.05-0.001; Figure 4A) blood glucose throughout the study period.
Oral gavage of EEAH (250 and 500 mg/5 mL/kg) and glibenclamide (5 mg/5 mL/kg) twice daily improved body weight in HFF mice during 60-day study period (Figure 4B). Compared to the HFF group alone, EEAH at doses of 250 and 500 mg/5 mL/kg gradually (p < 0.05-0.001) decreased body weight from day 6 and onwards. However, 500 mg/5 mL/kg dose showed a more consistent effect from day 18 and onwards. In addition, glibenclamide (5 mg/5 mL/kg) showed a remarkable (p < 0.05-0.001; Figure 4B) improvement in the body weight of HFF diet-induced obese mice from day 36 and onwards.
Oral administration of EEAH (250 and 500 mg/5 mL/kg) twice daily for 60 days significantly (p < 0.05-0.001; Figure 4C,D) improved food and fluid intake consistently from day 18 compared to HFF diet-induced obese control mice. Similarly, glibenclamide (5 mg/5 mL/kg) noticeably (p < 0.01-0.001; Figure 4C, D) decreased food and fluid intake consistently throughout the studies.
3.8. Lipid Profiling with EEAH
Long-term treatment (twice daily for 60 days) with EEAH (250 and 500 mg/5 mL/kg) or glibenclamide (5 mg/5 mL/kg) effectively (p < 0.05-0.01) elevated HDL cholesterol, while significantly (p < 0.05-0.001; Figure 5A-D) reduced total cholesterol, LDL and triglyceride levels, respectively, in high-fat-fed diet induced obese mice. High dose of EEAH at 500 mg/5 mL/kg showed the most potent antihyperglycemic effect. A sulfonylurea drug, glibenclamide (5 mg/5 mL/kg), also increased (p < 0.05, Figure 5A) HDL concentrations and reduced (p < 0.05-0.01; Figure 5B-D) total cholesterol, LDL as well as triglyceride levels compared to high-fat-fed diet induced obese mice alone.
3.9. Gut Motility with EEAH
Following 60 days of treatment with EEAH at 250 mg/5 mL/kg, no significant changes was observed in gastrointestinal motility (Figure 5E). However, EEAH at 500 mg/5 mL/kg significantly (p < 0.001; Figure 4E) ameliorated gut motility. Additionally, treatment with the stimulant laxative bisacodyl (10 mg/5 mL/kg) and the sulfonylurea drug glibenclamide considerably (p < 0.001 and p <0.05; Figure 4E) stimulated gut motility, whereas the antidiarrheal agent loperamide (5 mg/5 mL/kg) was associated with decrease (p < 0.01, Figure 4E) in intestinal motility.
3.10. Phytochemical Screening with EEAH
To identify potential anti-diabetic phytochemicals, present in unripe A. heterophyllus ethanolic extract, additional phytochemical screening was performed. The presence of alkaloids, tannins, flavonoids, saponins, terpenoids and steroids has been identified in EEAH (Table 1).
4. Discussion
Medicinal plants exhibit natural antioxidant properties which serve as an effective source of herbal glucose lowering medicine, particularly due to their bioactive phytochemicals such as flavonoids, saponins, tannins, phenolics, and alkaloids. These compounds can help in enhancing endocrine pancreatic function by the stimulation of insulin secretion or modulating intestinal glucose absorption [41]. This study investigated the antidiabetic potential of unripe Artocarpus heterophyllus (EEAH) extract via comprehensive in vitro and in vivo analyses to elucidate its mechanisms in regulation of blood glucose homeostasis.
The ethanolic extract of EEAH demonstrated a significant insulinotropic effect by enhancing insulin secretion from pancreatic β-cells under basal glucose conditions in a dose-dependent manner. This stimulatory activity was comparable to that of known insulin secretagogues which indicates a potent ability of EEAH to modulate β-cell function. Importantly, EEAH maintained its insulin-releasing capacity in the presence of tolbutamide and high extracellular potassium, suggesting that its mechanism of action may involve both KATP channel-dependent and -independent pathways. The suppression of EEAH-induced insulin secretion by diazoxide supports the involvement of KATP channel closure in its effect [42]. Furthermore, the reduction of insulin release by verapamil implicates voltage-dependent Ca²⁺ influx as a crucial mediator of EEAH action. The synergistic enhancement of insulin secretion with IBMX co-treatment implies the potential involvement of cAMP-mediated pathways [43]. Collectively, these findings demonstrate that EEAH stimulates insulin secretion through a multimodal mechanism, including closure of KATP channels, activation of voltage-dependent Ca²⁺ channels, and modulation of the cAMP signaling pathway.
The significant proliferative effect of EEAH on BRIN-BD11 β-cells at both lower and higher concentrations compared to control also represents a potentially therapeutically relevant finding for diabetes management. This proliferative response mirrored the effect of GLP-1, positive control which exhibits its action through established cAMP/PKA pathway activation [44]. This differential response indicates that bioactive compounds in EEAH, likely including polyphenols and flavonoids, may stimulate this action via this pathway [45]. EEAH may stimulate the proliferation of insulin-producing β-cells by activating key pathways such as PI3K/Akt, Wnt/β-catenin, or JAK-STAT, which are known to be involved in this regenerative process [46,47].
The pathogenesis of diabetes mellitus is generally multifactorial, key contributions often coming from enzymatic starch hydrolysis mediated by α-amylase and α-glucosidase as well as subsequent glucose absorption and diffusion across the intestinal epithelium. In this context, the ethanol extract of unripe Artocarpus heterophyllus (EEAH) exhibited a significant concentration-dependent inhibitory effect on enzymatic starch degradation, suggesting it may have the potential to attenuate postprandial hyperglycemia. This inhibitory activity may be attributed to the presence of bioactive phytoconstituents such as flavonoids and phenolic compounds known for their capacity to suppress α-amylase activity [48,49,50]. Additionally, EEAH significantly also impeded glucose diffusion and intestinal absorption in a concentration-dependent manner, indicating its possibility in limiting glucose bioavailability in the gastrointestinal tract. These findings align with previous literature documenting the antidiabetic potential of plant-based extracts through enzymatic and absorptive interference mechanisms [51].
Obesity which is known as a critical predisposing factor in case of T2DM, is often linked to elevated levels of non-esterified fatty acids (NEFAs) being released from adipose tissue, notorious in impairing insulin sensitivity and disrupting pancreatic β-cell function [52]. In the present investigation, EEAH resulted in a significant improvement in oral glucose tolerance of high-fat-fed (HFF) obese mice. These findings are in concordance with earlier plant based anti-diabetic studies, demonstrating that A. heterophyllus and other phytochemical-rich plant extracts can enhance glycemic control and preserve β-cell functionality in HFF-induced rodent models [53].
In the present study, chronic oral administration of the EEAH also demonstrated a significant antihyperglycemic effect in high-fat-fed (HFF) diet-induced obese mice during the 60-day treatment period. Twice-daily administration of EEAH has effectively led to a time-dependent reduction in blood glucose levels, with the higher dose exerting an earlier and more sustained glycemic control. These findings indicate a dose-responsive antihyperglycemic potential of EEAH, comparable to the standard antidiabetic agent, glibenclamide, which consistently reduced blood glucose throughout the study duration. The glycemic-lowering effect of EEAH may be attributed to its capacity to enhance pancreatic β-cell responsiveness, potentiate insulin secretion, and possibly modulate insulin signaling pathways [54]. These observations align with a previous study of A. heterophyllus, where similar blood glucose lowering trend was observed [54]. In both studies glibenclamide was used as a positive control that lowers blood glucose by binding to SUR1 on pancreatic β-cells, closing KATP channels, leading to calcium influx and stimulating insulin secretion [56]. Therefore, the bioactive compounds present in A. heterophyllus might be modulating through this pathway as it showed a comparable glucose lowering trend compared to control.
EEAH administration was also associated with a gradual and sustained reduction in body weight in obese mice, indicating its potential role in ameliorating obesity-linked metabolic disturbances. Excessive caloric intake combined with inadequate energy expenditure disrupts lipid metabolism, leading to obesity and associated metabolic disorders like diabetes and non-alcoholic fatty liver disease (NAFLD). The effect on body weight by EEAH may reflect improved energy homeostasis and reduced adiposity, potentially through modulation of lipid metabolism and appetite regulation [57,58]. The presence of polyphenol also contributes to this appetite suppression and weight loss [59]. In line with this, EEAH also significantly improved both food and fluid intake patterns, which are typically disrupted in diabetic and obese states. These observations are consistent with earlier reports on A. heterophyllus [60].
Both tested doses of EEAH showed a marked increase in HDL-cholesterol levels while displayed a concurrent decrease in total cholesterol, LDL-cholesterol, and triglyceride levels. This lipid-modulating potential may be attributed to the extract’s ability to enhance lipoprotein catabolism and promote reverse cholesterol transport, aligning with previous findings on plant-derived compounds with antihyperlipidemic activity [61]. The observed increase in HDL may further be supported by improved hormonal regulation, potentially involving thyroid-related pathways that facilitate the hepatic clearance of LDL [62].
EEAH treatment over 60 days resulted a significant improvement in gastrointestinal motility at higher dose. This suggests that EEAH may be helpful in reducing glucose absorption by accelerating transit time through the gastrointestinal tract. The increased motility could limit the duration available for nutrient absorption, contributing to the extract’s antihyperglycemic activity [63]. Disaccharides like sucrose require breakdown into monosaccharides for absorption due to the absence of specific carriers in the GI tract; thus, the increased sucrose content following EEAH treatment likely reflects reduced sucrose digestion [64]. Such effects are likely linked to the presence of dietary fibers or bioactive compounds within EEAH that modulate gut function, similar to other plant-based treatments known to influence glucose regulation via digestive mechanisms [63,64].
Phytochemical analysis of the ethanol extract of unripe A. heterophyllus (EEAH) revealed the presence of several bioactive constituents, including alkaloids, tannins, flavonoids, saponins, steroids and terpenoids, Tannins are known to improve glucose uptake, block adipocyte differentiation, inhibit inflammation, oxidative stress and improve insulin sensitivity [65,66]. Saponins, particularly steroidal types, have been reported to mimic insulin action and improve glucose metabolism by modulating activity of adipokines [67]. Flavonoids, including well-studied compounds such as catechin and epicatechin, exert antioxidant effects and improve glycemic control by targeting pathways involved in insulin resistance and glucose homeostasis [68]. Alkaloids and terpenoids also play significant roles in modulating carbohydrate metabolism, enhancing GLUT-4 translocation and increasing pancreatic β-cell function [69]. The presence of these diverse phytochemicals in EEAH suggests a multifaceted mechanism underlying its antidiabetic potential, warranting further investigation into their individual and combined therapeutic contributions.
5. Conclusions
This study highlights the therapeutic potential of the ethanol extract of unripe A. heterophyllus (EEAH) as a promising candidate for the management of type 2 diabetes mellitus. EEAH demonstrated significant insulinotropic, antihyperglycemic and hypolipidemic effects, improved gastrointestinal motility, and contained a diverse array of phytochemicals known to modulate glucose metabolism, insulin sensitivity, and lipid profiles. These findings suggest a multimodal mechanism of action including enhancement of insulin secretion, modulation of nutrient absorption, and possible interaction with signaling pathways involved in metabolic regulation. Given the complex interplay of phytochemicals within EEAH and their broad biological targets, future research should prioritize the isolation and characterization of bioactive phytomolecules responsible for the observed antidiabetic activity and the elucidation of their detailed mechanisms of action, ultimately paving the way for the development of targeted and effective therapeutic strategies. To further unravel these complex interactions, subsequent investigations using multi-omics approaches are essential. These integrative studies will provide a comprehensive understanding of the molecular mechanisms underpinning EEAH's antidiabetic action and aid in identifying specific biomarkers of efficacy and safety, ultimately supporting the development of EEAH as a standardized, evidence-based nutraceutical or adjunct therapy for diabetes management in humans.
Author Contributions
P.R.F., P.A., and Y.H.A.A.-W. were responsible for the conceptualization and design of this research and jointly oversaw its execution; P.A., S.S.I., M.S.R.M., A.A. contributed to the experimental work and data analysis; the interpretation of the findings, figure preparation, and initial manuscript drafting were undertaken by P.A., A.D.R., J.T.K., A.A., and J.M.A.H.; P.R.F., P.A. and Y.H.A.A.-W. were involved in editing the revised manuscript. All authors have read and approved the final version of this manuscript.
Funding
This research was conducted without external financial support.
Institutional Review Board Statement
Animal experiments for this study were ethically approved by the Institutional Review Board (IRB) at Independent University, Bangladesh (IUB) on September 1, 2022. All procedures adhered strictly to national and international animal welfare guidelines, including the Animal Welfare Act 2019 of Bangladesh, the UK Animals (Scientific Procedures) Act 1986, and EU Directive 2010/63/EU. Experiments were conducted under UK Home Office Animal license PIL1822, ensuring high standards of animal care and minimizing distress.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data generated and analyzed during this study are not publicly accessible due to ethical or legal constraints. Requests for access to the data can be directed to the corresponding author and will be considered on a reasonable basis.
Acknowledgments
The authors gratefully acknowledge the Ulster University Strategic Research Funding and Independent University, Bangladesh (IUB), Dhaka, for the provision of laboratory resources that facilitated this research.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Banday, M. Z.; Sameer, A. S.; Nissar, S. Pathophysiology of Diabetes: An Overview. Avicenna J. Med. 2020, 10, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Ansari, P.; Samia, J. F.; Khan, J. T.; Rafi, M. R.; Rahman, M. S.; Rahman, A. B.; Abdel-Wahab, Y. H. A.; Seidel, V. Protective Effects of Medicinal Plant-Based Foods Against Diabetes: A Review on Pharmacology, Phytochemistry, and Molecular Mechanisms. Nutrients 2023, 15, 3266. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M. J.; Al-Mamun, M.; Islam, M. R. Diabetes Mellitus, the Fastest Growing Global Public Health Concern: Early Detection Should be Focused. Health Sci Rep. 2024, 7, e2004. [Google Scholar] [CrossRef] [PubMed]
- Hannan, J. M. A.; Nipa, N.; Toma, F. T.; Talukder, A.; Ansari, P. Acute Anti-Hyperglycaemic Activity of Five Traditional Medicinal Plants in High Fat Diet Induced Obese Rats. Front. Biosci. (Schol. Ed) 2023, 15, 5. [Google Scholar] [CrossRef]
- Rahman, M. S.; Hossain, K. S.; Das, S.; Kundu, S.; Adegoke, E. O.; Rahman, M. A.; Hannan, M. A.; Uddin, M. J.; Pang, M.-G. Role of Insulin in Health and Disease: An Update. Int. J. Mol. Sci. 2021, 22, 6403. [Google Scholar] [CrossRef]
- Fu, Z.; Gilbert, E. R.; Liu, D. Regulation of Insulin Synthesis and Secretion and Pancreatic Beta-Cell Dysfunction in Diabetes. Curr. Diabetes Rev. 2013, 9, 25–53. [Google Scholar] [CrossRef]
- Vakilian, M.; Tahamtani, Y.; Ghaedi, K. A Review on Insulin Trafficking and Exocytosis. Gene 2019, 706, 52–61. [Google Scholar] [CrossRef]
- Petersen, M. C.; Shulman, G. I. Mechanisms of Insulin Action and Insulin Resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef]
- Subramanian, S.; Khan, F.; Hirsch, I. B. New Advances in Type 1 Diabetes. BMJ 2024, 384, e075681. [Google Scholar] [CrossRef]
- Lucier, J.; Mathias, P. M. Type 1 Diabetes. In StatPearls [Internet] 2024. Available online: https://www.ncbi.nlm.nih.gov/books/NBK507713/.
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K. B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- Ansari, P.; Flatt, P. R.; Harriott, P.; Abdel-Wahab, Y. H. A. Insulin Secretory and Antidiabetic Actions of Heritiera fomes Bark Together with Isolation of Active Phytomolecules. PloS one 2022, 17, e0264632. [Google Scholar] [CrossRef] [PubMed]
- Farmaki, P.; Damaskos, C.; Garmpis, N.; Garmpi, A.; Savvanis, S.; Diamantis, E. Complications of the Type 2 Diabetes Mellitus. Curr. Cardiol. Rev. 2020, 16, 249–251. [Google Scholar] [CrossRef] [PubMed]
- Olokoba, A. B.; Obateru, O. A.; Olokoba, L. B. Type 2 Diabetes Mellitus: A Review of Current Trends. Oman Med. J. 2012, 27, 269–73. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, B. S. Q.; Vadakekut, E. S.; Mahdy, H. Gestational Diabetes. In StatPearls [Internet]; StatPearls Publishing: 2024. Available online: https://www.ncbi.nlm.nih.gov/books/NBK545196/.
- Polonsky, W. H.; Henry, R. R. Poor Medication Adherence in Type 2 Diabetes: Recognizing the Scope of the Problem and its Key Contributors. Patient Prefer. Adherence 2016, 10, 1299–1307. [Google Scholar] [CrossRef]
- Herges, J. R.; Neumiller, J. J.; McCoy, R. G. Easing the Financial Burden of Diabetes Management: A Guide for Patients and Primary Care Clinicians. Clin. Diabetes 2021, 39, 427–436. [Google Scholar] [CrossRef]
- Ajiboye, B. O.; Adeleke Ojo, O.; Adeyonu, O.; Imiere, O.; Emmanuel Oyinloye, B.; Ogunmodede, O. Ameliorative Activity of Ethanolic Extract of Artocarpus heterophyllus Stem Bark on Alloxan-induced Diabetic Rats. Adv. Pharm. Bull. 2018, 8, 141–147. [Google Scholar] [CrossRef]
- Roy, J. R.; Julius, A.; Chinnapan, V. Health Benefits and Future Prospects of Artocarpus heterophyllus. Res. J. Pharm. Tech. 2023, 16, 4443–4446. [Google Scholar] [CrossRef]
- Ranasinghe, R. A. S. N.; Maduwanthi, S. D. T.; Marapana, R. A. U. J. Nutritional and Health Benefits of Jackfruit (Artocarpus heterophyllus Lam.): A Review. Int. J. Food Sci. 2019, 2019, 4327183. [Google Scholar] [CrossRef]
- Omar, H. S.; El-Beshbishy, H. A.; Moussa, Z.; Taha, K. F.; Singab, A. N. Antioxidant Activity of Artocarpus heterophyllus Lam. (Jack Fruit) Leaf Extracts: Remarkable Attenuations of Hyperglycemia and Hyperlipidemia in Streptozotocin-Diabetic Rats. ScientificWorldJournal 2011, 11, 788–800. [Google Scholar] [CrossRef]
- Prakash, O.; Kumar, R.; Mishra, A.; Gupta, R. Artocarpus heterophyllus (Jackfruit): An Overview. Pharmacogn. Rev. 2009, 3, 353–358. [Google Scholar]
- Ansari, P.; Hannan, J. M. A.; Flatt, P. R.; Abdel-Wahab, Y. H. A. Anti-Diabetic Properties of the Ethanolic Extract of Unripe Artocarpus heterophyllus Fruit Regulates Glucose Homeostasis in High-Fat-Fed Diet Induced Obese Mice. In Endocrine Abstracts 2023, 94. [Google Scholar] [CrossRef]
- Chackrewarthy, S.; Thabrew, M. I.; Weerasuriya, M. K.; Jayasekera, S. Evaluation of the Hypoglycemic and Hypolipidemic Effects of an Ethylacetate Fraction of Artocarpus heterophyllus (Jak) Leaves in Streptozotocin-Induced Diabetic Rats. Pharmacogn. Mag. 2010, 6, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Ansari, P.; Khan, J. T.; Chowdhury, S.; Reberio, A. D.; Kumar, S.; Seidel, V.; Abdel-Wahab, Y. H. A.; Flatt, P. R. Plant-Based Diets and Phytochemicals in the Management of Diabetes Mellitus and Prevention of Its Complications: A Review. Nutrients 2024, 16, 3709. [Google Scholar] [CrossRef] [PubMed]
- Ansari, P.; Azam, S.; Seidel, V.; Abdel-Wahab, Y. H. A. In Vitro and In Vivo Antihyperglycemic Activity of the Ethanol Extract of Heritiera fomes Bark and Characterization of Pharmacologically Active Phytomolecules. J. Pharm. Pharmacol. 2022, rgac010. [Google Scholar] [CrossRef]
- O'Harte, F. P.; Ng, M. T.; Lynch, A. M.; Conlon, J. M.; Flatt, P. R. Novel Dual Agonist Peptide Analogues Derived from Dogfish Glucagon Show Promising In Vitro Insulin Releasing Actions and Antihyperglycaemic Activity in Mice. Mol. Cell. Endocrinol. 2016, 431, 133–144. [Google Scholar] [CrossRef]
- Ansari, P.; Flatt, P. R.; Harriott, P.; Abdel-Wahab, Y. H. A. Evaluation of the Antidiabetic and Insulin Releasing Effects of A. squamosa, Including Isolation and Characterization of Active Phytochemicals. Plants 2020, 9, 1348. [Google Scholar] [CrossRef]
- Ansari, P.; Khan, J. T.; Soultana, M.; Hunter, L.; Chowdhury, S.; Priyanka, S. K.; Paul, S. Flatt, P. R.; Abdel-Wahab, Y. H. A. Insulin Secretory Actions of Polyphenols of Momordica charantia Regulate Glucose Homeostasis in Alloxan-Induced Type 2 Diabetic Rats. RPS Pharm. Pharmacol. Rep. 2024, 3, rqae005. [Google Scholar] [CrossRef]
- Ansari, P.; Hannan, J. M. A.; Seidel, V.; Abdel-Wahab, Y. H. A. Polyphenol-Rich Leaf of Annona squamosa Stimulates Insulin Release from BRIN-BD11 Cells and Isolated Mouse Islets, Reduces (CH2O)n Digestion and Absorption, and Improves Glucose Tolerance and GLP-1 (7-36) Levels in High-Fat-Fed Rats. Metabolites 2022, 12, 995. [Google Scholar] [CrossRef]
- Zhu, W.; Tanday, N.; Lafferty, R. A.; Flatt, P. R.; Irwin, N. Novel Enzyme-Resistant Pancreatic Polypeptide Analogs Evoke Pancreatic Beta-Cell Rest, Enhance Islet Cell Turnover, and Inhibit Food Intake in Mice. Biofactors 2024, 50, 1101–1112. [Google Scholar] [CrossRef]
- Ansari, P.; Islam, S. S.; Akther, S.; Khan, J. T.; Shihab, J. A.; Abdel-Wahab, Y. H. Insulin Secretory Actions of Ethanolic Extract of Acacia arabica Bark in High Fat-Fed Diet-Induced Obese Type 2 Diabetic Rats. Biosci. Rep. 2023, 43, BSR20230329. [Google Scholar] [CrossRef]
- Gallagher, A. M.; Flatt, P. R.; Duffy, G.; Abdel-Wahab, Y. H. A. The Effects of Traditional Antidiabetic Plants on In Vitro Glucose Diffusion. Nutr. Res. 2003, 23, 413–424. [Google Scholar] [CrossRef]
- Ansari, P.; Hannan, J. M. A.; Choudhury, S. T.; Islam, S. S.; Talukder, A.; Seidel, V.; Abdel-Wahab, Y. H. A. Antidiabetic Actions of Ethanol Extract of Camellia sinensis Leaf Ameliorates Insulin Secretion, Inhibits the DPP-IV Enzyme, Improves Glucose Tolerance, and Increases Active GLP-1 Levels in High-Fat-Diet-Fed Rats. Medicines 2022, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- López-Soldado, I.; Guinovart, J. J.; Duran, J. Increasing Hepatic Glycogen Moderates the Diabetic Phenotype in Insulin-Deficient Akita Mice. J. Biol. Chem. 2021, 296, 100498. [Google Scholar] [CrossRef] [PubMed]
- Khan, J. T.; Richi, A. E.; Riju, S. A.; Jalal, T.; Orchi, R. J.; Singh, S.; Bhagat, P.; Abdel-Wahab, Y. H. A.; Ansari, P. Evaluation of Antidiabetic Potential of Mangifera indica Leaf in Streptozotocin-Induced Type 2 Diabetic Rats: Focus on Glycemic Control and Cholesterol Regulation. Endocrines 2024, 5, 137–152. [Google Scholar] [CrossRef]
- Hannan, J. M. A.; Ali, L.; Rokeya, B.; Khaleque, J.; Akhter, M.; Flatt, P. R.; Abdel-Wahab, Y. H. Soluble Dietary Fibre Fraction of Trigonella Foenum-graecum (Fenugreek) Seed Improves Glucose Homeostasis in Animal Models of Type 1 and Type 2 Diabetes by Delaying Carbohydrate Digestion and Absorption, and Enhancing Insulin Action. Br. J. Nutr. 2007, 97, 514–521. [Google Scholar] [CrossRef]
- Hannan, J. M. A.; Ansari, P.; Haque, A.; Sanju, A.; Huzaifa, A.; Rahman, A.; Ghosh, A.; Azam, S. Nigella sativa Stimulates Insulin Secretion from Isolated Rat Islets and Inhibits the Digestion and Absorption of (CH2O)n in the Gut. Biosci. Rep. 2019, 39, BSR20190723. [Google Scholar] [CrossRef]
- Ansari, P.; Badhan, S.; Azam, S.; Sultana, N.; Anwar, S.; Mohamed Abdurahman, M. S.; Hannan, J. M. A. Evaluation of Antinociceptive and Anti-inflammatory Properties of Methanolic Crude Extract of Lophopetalum javanicum (Bark). J. Basic Clin. Physiol. Pharmacol. 2016, 27, 379–385. [Google Scholar] [CrossRef]
- Credo, D.; Machumi, F.; Masimba, P. J. Phytochemical Screening and Evaluation of Ant-Diabetic Potential of Selected Medicinal Plants used Traditionally for Diabetes Management in Tanzania. Int. J. Res. Pharm. Chem. 2018, 8, 405–411. [Google Scholar]
- Kooti, W.; Farokhipour, M.; Asadzadeh, Z.; Ashtary-Larky, D.; Asadi-Samani, M. The Role of Medicinal Plants in the Treatment of Diabetes: A Systematic Review. Electron. Physician 2016, 8, 1832–1842. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Harvat, T.; Kinzer, K.; Zhang, L.; Feng, F.; Qi, M.; Oberholzer, J. Diazoxide, a K(ATP) Channel Opener, Prevents Ischemia-Reperfusion Injury in Rodent Pancreatic Islets. Cell Transplant. 2015, 24, 25–36. [Google Scholar] [CrossRef]
- Sharp, G. W. The Adenylate Cyclase-Cyclic AMP System in Islets of Langerhans and its Role in the Control of Insulin Release. Diabetologia 1979, 16, 287–96. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Zong, Y.; Ma, Y.; Tian, Y.; Pang, Y.; Zhang, C.; Gao, J. Glucagon-like Peptide-1 Receptor: Mechanisms and Advances in Therapy. Signal Transduct. Target. Ther. 2024, 9, 234. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Q.; Kang, S. G.; Huang, K.; Tong, T. Dietary Bioactive Ingredients Modulating the cAMP Signaling in Diabetes Treatment. Nutrients 2021, 13, 3038. [Google Scholar] [CrossRef] [PubMed]
- García-Aguilar, A.; Guillén, C. Targeting Pancreatic Beta Cell Death in Type 2 Diabetes by Polyphenols. Front. Endocrinol. 2022, 13, 1052317. [Google Scholar] [CrossRef]
- Stewart, A. F.; Hussain, M. A.; García-Ocaña, A.; Vasavada, R. C.; Bhushan, A.; Bernal-Mizrachi, E.; Kulkarni, R. N. Human β-cell Proliferation and Intracellular Signaling: Part 3. Diabetes 2015, 64, 1872–1885. [Google Scholar] [CrossRef]
- Jiang, W. J.; Peng, Y. C.; Yang, K. M. Cellular Signaling Pathways Regulating β-cell Proliferation as a Promising Therapeutic Target in the Treatment of Diabetes. Exp. Ther. Med. 2018, 16, 3275–3285. [Google Scholar] [CrossRef]
- Dias, D. T. M.; Palermo, K. R.; Motta, B. P.; Kaga, A. K.; Lima, T. F. O.; Brunetti, I. L.; Baviera, A. M. Rutin Inhibits the in Vitro Formation of Advanced Glycation Products and Protein Oxidation More Efficiently than Quercetin. Rev. Cienc. Farm. Basica Apl. 2021, 42, e718. [Google Scholar] [CrossRef]
- Falla, N. M.; Demasi, S.; Caser, M.; Scariot, V. Phytochemical Profile and Antioxidant Properties of Italian Green Tea, a New High Quality Niche Product. Horticulturae 2021, 7, 91. [Google Scholar] [CrossRef]
- Ansari, P.; Flatt, P. R.; Harriott, P.; Abdel-Wahab, Y. H. Anti-hyperglycaemic and Insulin-Releasing Effects of Camellia sinensis Leaves and Isolation and Characterization of Active Compounds. Br. J. Nutr. 2020, 126, 1149–1163. [Google Scholar] [CrossRef]
- Takahama, U.; Hirota, S. Interactions of Flavonoids with α-Amylase and Starch Slowing down Its Digestion. Food Funct. 2018, 9, 677–687. [Google Scholar] [CrossRef]
- Wickramasinghe, A. S. D.; Kalansuriya, P.; Attanayake, A. P. Herbal Medicines Targeting the Improved β-Cell Functions and β-Cell Regeneration for the Management of Diabetes Mellitus. Evid. Based Complement. Alternat. Med. 2021, 2021, 2920530. [Google Scholar] [CrossRef] [PubMed]
- Winzell, M. S.; Ahrén, B. The High-Fat Diet-Fed Mouse: A Model for Studying Mechanisms and Treatment of Impaired Glucose Tolerance and Type 2 Diabetes. Diabetes 2004, 53 (Suppl 3), S215–9. [Google Scholar] [CrossRef] [PubMed]
- Dwitiyanti, D.; Rachmania, R. A.; Efendi, K.; Septiani, R.; Jihadudin, P. In Vivo Activities and In Silico Study of Jackfruit Seeds (Artocarpus heterophyllus Lam.) on the Reduction of Blood Sugar Levels of Gestational Diabetes Rate Induced by Streptozotocin. Open Access Maced. J. Med. Sci. 2019, 7, 3819–3826. [Google Scholar] [CrossRef]
- Costello, R. A.; Nicolas, S.; Shivkumar, A. Sulfonylureas. In StatPearls [Internet]; StatPearls Publishing: 2025. Available online: https://www.ncbi.nlm.nih.gov/books/NBK513225/.
- Hussain, A.; Cho, J. S.; Kim, J. S.; Lee, Y. I. Protective Effects of Polyphenol Enriched Complex Plants Extract on Metabolic Dysfunctions Associated with Obesity and Related Nonalcoholic Fatty Liver Diseases in High Fat Diet-Induced C57BL/6 Mice. Molecules 2021, 26, 302. [Google Scholar] [CrossRef]
- Vrânceanu, M.; Hegheş, S. C.; Cozma-Petruţ, A.; Banc, R.; Stroia, C. M.; Raischi, V.; Miere, D.; Popa, D. S.; Filip, L. Plant-Derived Nutraceuticals Involved in Body Weight Control by Modulating Gene Expression. Plants 2023, 12, 2273. [Google Scholar] [CrossRef]
- Boccellino, M.; D'Angelo, S. Anti-Obesity Effects of Polyphenol Intake: Current Status and Future Possibilities. Int. J. Mol. Sci. 2020, 21, 5642. [Google Scholar] [CrossRef]
- Tripathi, K.; Kumar, P.; Kumar, R.; Saxena, R.; Kumar, A.; Badoni, H.; Goyal, B.; Mirza, A. A. Efficacy of Jackfruit Components in Prevention and Control of Human Disease: A Scoping Review. J. Educ. Health Promot. 2023, 12, 361. [Google Scholar] [CrossRef]
- Mokhtari, I.; Shahat, A. A.; Noman, O. M.; Milenkovic, D.; Amrani, S.; Harnafi, H. Effects of Cynara scolymus L. Bract Extract on Lipid Metabolism Disorders Through Modulation of HMG-CoA Reductase, Apo A-1, PCSK-9, p-AMPK, SREBP-2, and CYP2E1 Expression. Metabolites 2024, 14, 728. [Google Scholar] [CrossRef]
- Sinha, R. A.; Singh, B. K.; Yen, P. M. Direct Effects of Thyroid Hormones on Hepatic Lipid Metabolism. Nat. Rev. Endocrinol. 2018, 14, 259–269. [Google Scholar] [CrossRef]
- Ansari, P.; Hannan, J. M. A.; Seidel, V.; Abdel-Wahab, Y. H. A. Polyphenol-Rich Leaf of Annona squamosa Stimulates Insulin Release from BRIN-BD11 Cells and Isolated Mouse Islets, Reduces (CH2O)n Digestion and Absorption, and Improves Glucose Tolerance and GLP-1 (7-36) Levels in High-Fat-Fed Rats. Metabolites 2022, 12, 995. [Google Scholar] [CrossRef]
- Kiela, P. R.; Ghishan, F. K. Physiology of Intestinal Absorption and Secretion. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 145–59. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Kim, J. K.; Li, Y.; Li, J.; Liu, F.; Chen, X. Tannic Acid Stimulates Glucose Transport and Inhibits Adipocyte Differentiation in 3T3-L1 Cells. J. Nutr. 2005, 135, 165–71. [Google Scholar] [CrossRef] [PubMed]
- Sahakyan, G.; Vejux, A.; Sahakyan, N. The Role of Oxidative Stress-Mediated Inflammation in the Development of T2DM-Induced Diabetic Nephropathy: Possible Preventive Action of Tannins and Other Oligomeric Polyphenols. Molecules 2022, 27, 9035. [Google Scholar] [CrossRef] [PubMed]
- Elekofehinti, O. O.; Ejelonu, O. C.; Kamdem, J. P.; Akinlosotu, O. B.; Adanlawo, I. G. Saponins as Adipokines Modulator: A Possible Therapeutic Intervention for Type 2 Diabetes. World J. Diabetes 2017, 8, 337–345. [Google Scholar] [CrossRef]
- Martín, M. Á.; Ramos, S. Dietary Flavonoids and Insulin Signaling in Diabetes and Obesity. Cells 2021, 10, 1474. [Google Scholar] [CrossRef]
- Singh, S.; Bansal, A.; Singh, V.; Chopra, T.; Poddar, J. Flavonoids, Alkaloids and Terpenoids: A New Hope for the Treatment of Diabetes Mellitus. J. Diabetes Metab. Disord. 2022, 21, 941–950. [Google Scholar] [CrossRef]
Figure 1.
Effects of Ethanol extract of unripe A. heterophyllus (EEAH) on insulin secretion from (A) clonal pancreatic BRIN BD11 β-cells alone, (B) with known modulators or inhibitors, (C) with or without extracellular calcium, (D) starch digestion, and (E) glucose diffusion. Percentage of glucose liberation from starch digestion and glucose diffusion in vitro were represented in the scatter dot plot. Values are mean ± SEM; n = 8 for insulin secretion and n = 4 for starch digestion and glucose diffusion. *p<0.05, **p<0.01, *** p < 0.001 compared to control. ϕ p < 0.05 and ϕϕϕ p < 0.001 compared to 5.6 mM glucose with EEAH. Δp<0.05, ΔΔp<0.01, ΔΔΔ p < 0.001 compared to respective incubation without EEAH. EEAH, ethanol extract of A. heterophyllus (unripe fruit).
Figure 1.
Effects of Ethanol extract of unripe A. heterophyllus (EEAH) on insulin secretion from (A) clonal pancreatic BRIN BD11 β-cells alone, (B) with known modulators or inhibitors, (C) with or without extracellular calcium, (D) starch digestion, and (E) glucose diffusion. Percentage of glucose liberation from starch digestion and glucose diffusion in vitro were represented in the scatter dot plot. Values are mean ± SEM; n = 8 for insulin secretion and n = 4 for starch digestion and glucose diffusion. *p<0.05, **p<0.01, *** p < 0.001 compared to control. ϕ p < 0.05 and ϕϕϕ p < 0.001 compared to 5.6 mM glucose with EEAH. Δp<0.05, ΔΔp<0.01, ΔΔΔ p < 0.001 compared to respective incubation without EEAH. EEAH, ethanol extract of A. heterophyllus (unripe fruit).
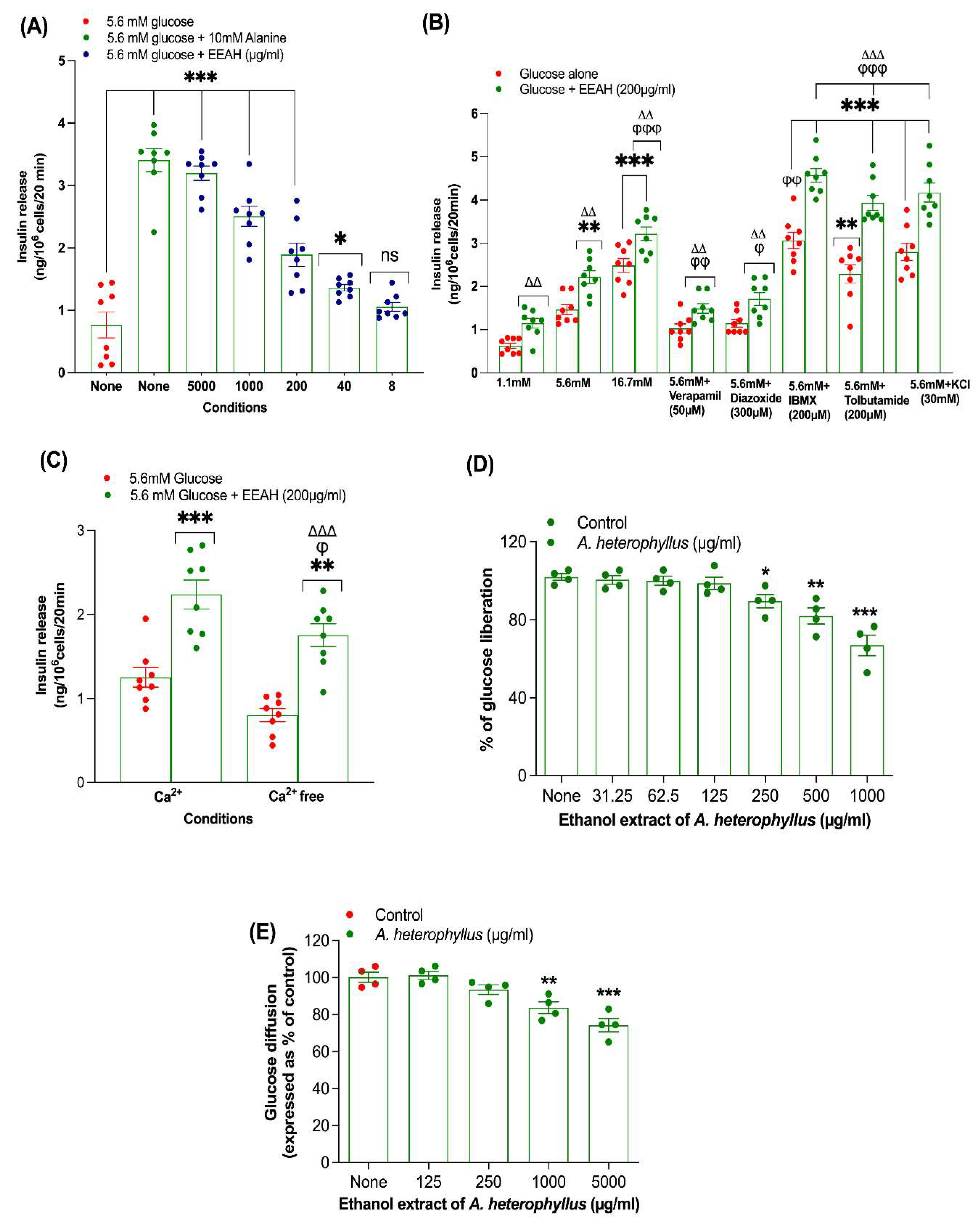
Figure 2.
Effects of Ethanol extract of unripe A. heterophyllus (EEAH) on (A-E) BRIN-BD11 β-cells proliferation. BRIN-BD11 cells were cultured (18 h) with EEAH (40 and 200 μg/ml) or GLP-1 (10-6 M) and proliferation assessed by Ki-67 staining. Representative images depict cells stained with DAPI (blue) and Ki-67 (red), with arrows indicating Ki-67-positive (proliferating) cells. Figure 2A-D shows BRIN BD11 β-cell proliferation images under four conditions: (A) Control, (B) GLP-1 (10⁻⁶ M), (C) EEAH at 40 μg/ml, and (D) EEAH at 200 μg/ml. Percentage of Ki-67+ve cells were represented in the scatter dot plot. All values are mean ± SEM with n = 6. **, *** p < 0.01-0.001 compared with control. ΔΔ, ΔΔΔ p < 0.01-0.001 compared with GLP-1 (10-6 M).
Figure 2.
Effects of Ethanol extract of unripe A. heterophyllus (EEAH) on (A-E) BRIN-BD11 β-cells proliferation. BRIN-BD11 cells were cultured (18 h) with EEAH (40 and 200 μg/ml) or GLP-1 (10-6 M) and proliferation assessed by Ki-67 staining. Representative images depict cells stained with DAPI (blue) and Ki-67 (red), with arrows indicating Ki-67-positive (proliferating) cells. Figure 2A-D shows BRIN BD11 β-cell proliferation images under four conditions: (A) Control, (B) GLP-1 (10⁻⁶ M), (C) EEAH at 40 μg/ml, and (D) EEAH at 200 μg/ml. Percentage of Ki-67+ve cells were represented in the scatter dot plot. All values are mean ± SEM with n = 6. **, *** p < 0.01-0.001 compared with control. ΔΔ, ΔΔΔ p < 0.01-0.001 compared with GLP-1 (10-6 M).
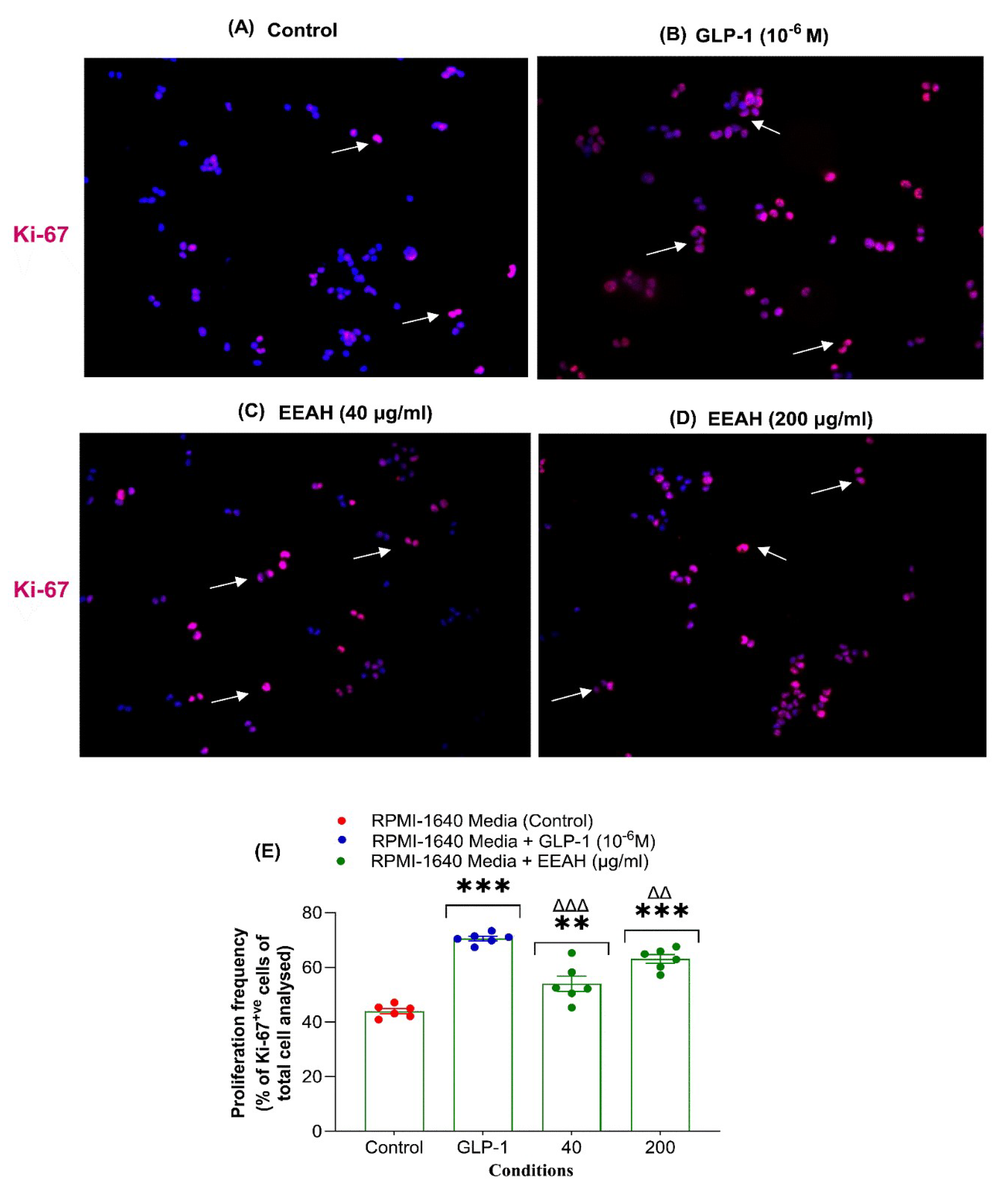
Figure 3.
Acute and chronic effects of ethanol extract of unripe A. heterophyllus (EEAH) in high-fat-fed mice on oral glucose tolerance at (A) 0, (B) 15, (C) 30 and (D) 60 days respectively expressed as line graphs. High-fat-fed mice fasted for overnight were used to measure the blood glucose from the tail tips prior to and after receiving glucose (2.5 gm/5 ml/kg, body weight, control) with or without EEAH (250 and 500 mg/5 ml/kg, body weight) or glibenclamide (5 mg/5 ml/kg). Values are the mean ± SEM for n = 6 mice. *, ** p < 0.05-0.01 compared to lean control (saline), Δ, ΔΔ, ΔΔΔ p < 0.05-0.001 compared to high-fat-fed diet control mice. Glibenclamide was used as a positive control.
Figure 3.
Acute and chronic effects of ethanol extract of unripe A. heterophyllus (EEAH) in high-fat-fed mice on oral glucose tolerance at (A) 0, (B) 15, (C) 30 and (D) 60 days respectively expressed as line graphs. High-fat-fed mice fasted for overnight were used to measure the blood glucose from the tail tips prior to and after receiving glucose (2.5 gm/5 ml/kg, body weight, control) with or without EEAH (250 and 500 mg/5 ml/kg, body weight) or glibenclamide (5 mg/5 ml/kg). Values are the mean ± SEM for n = 6 mice. *, ** p < 0.05-0.01 compared to lean control (saline), Δ, ΔΔ, ΔΔΔ p < 0.05-0.001 compared to high-fat-fed diet control mice. Glibenclamide was used as a positive control.
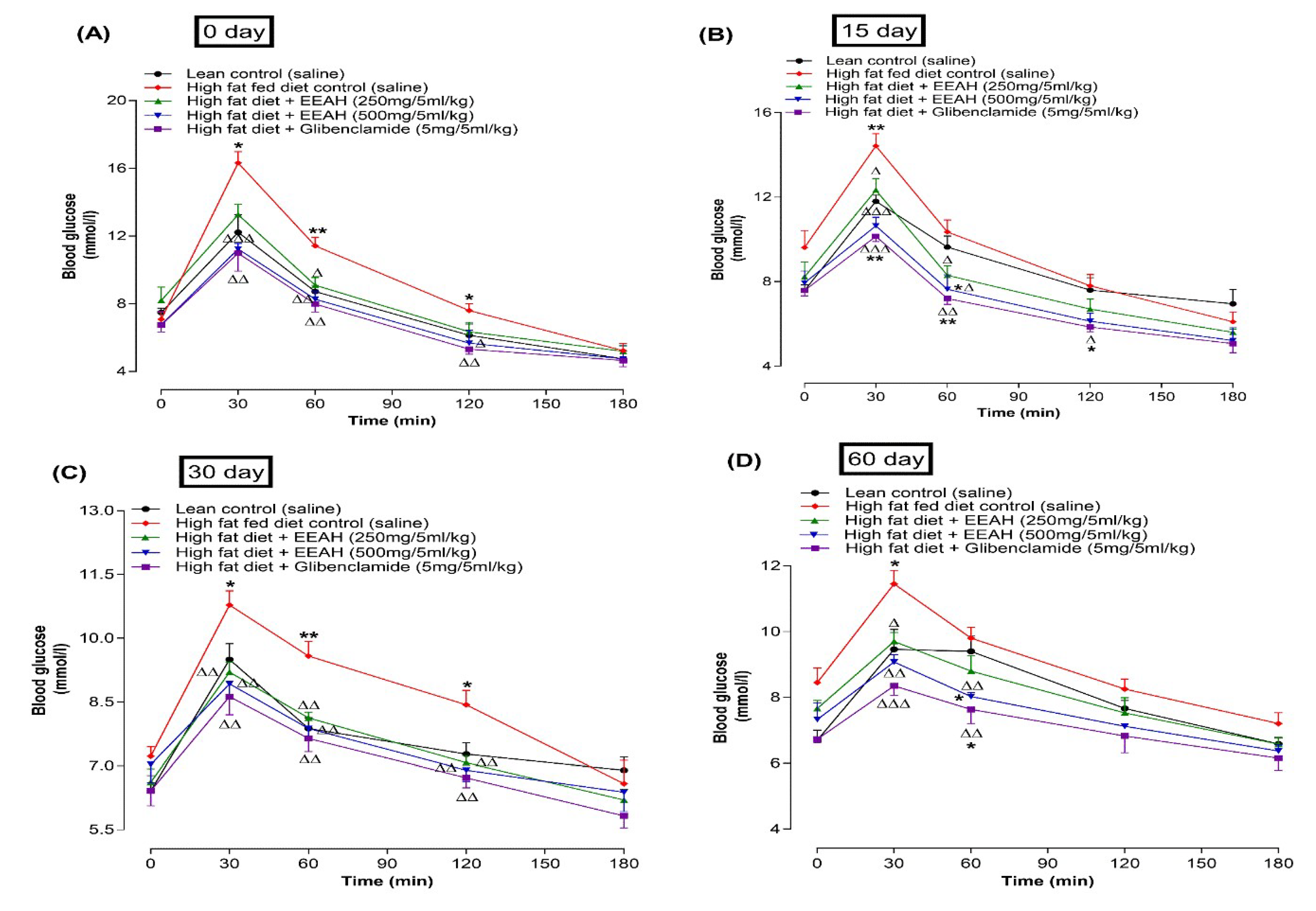
Figure 4.
Effects of 60-days chronic treatment (twice-daily) with ethanol extract of unripe A. heterophyllus (EEAH) on (A) body weight, (B) fasting blood glucose, (C) food intake, (D) fluid intake in high-fat-fed mice represented as line graphs. Parameters were measured at 3-day intervals prior to and after oral administration of either EEAH (250 and 500 mg/5 mL/kg, body weight) or glibenclamide (5 mg/5 ml/kg, body weight) twice daily. Values are the mean ± SEM for n = 6 mice. *, **, *** p < 0.05-0.001 compared to control (saline). Δ, ΔΔ, ΔΔΔ p < 0.05-0.001 compared to high-fat-fed diet control mice. As a positive control, glibenclamide was used.
Figure 4.
Effects of 60-days chronic treatment (twice-daily) with ethanol extract of unripe A. heterophyllus (EEAH) on (A) body weight, (B) fasting blood glucose, (C) food intake, (D) fluid intake in high-fat-fed mice represented as line graphs. Parameters were measured at 3-day intervals prior to and after oral administration of either EEAH (250 and 500 mg/5 mL/kg, body weight) or glibenclamide (5 mg/5 ml/kg, body weight) twice daily. Values are the mean ± SEM for n = 6 mice. *, **, *** p < 0.05-0.001 compared to control (saline). Δ, ΔΔ, ΔΔΔ p < 0.05-0.001 compared to high-fat-fed diet control mice. As a positive control, glibenclamide was used.
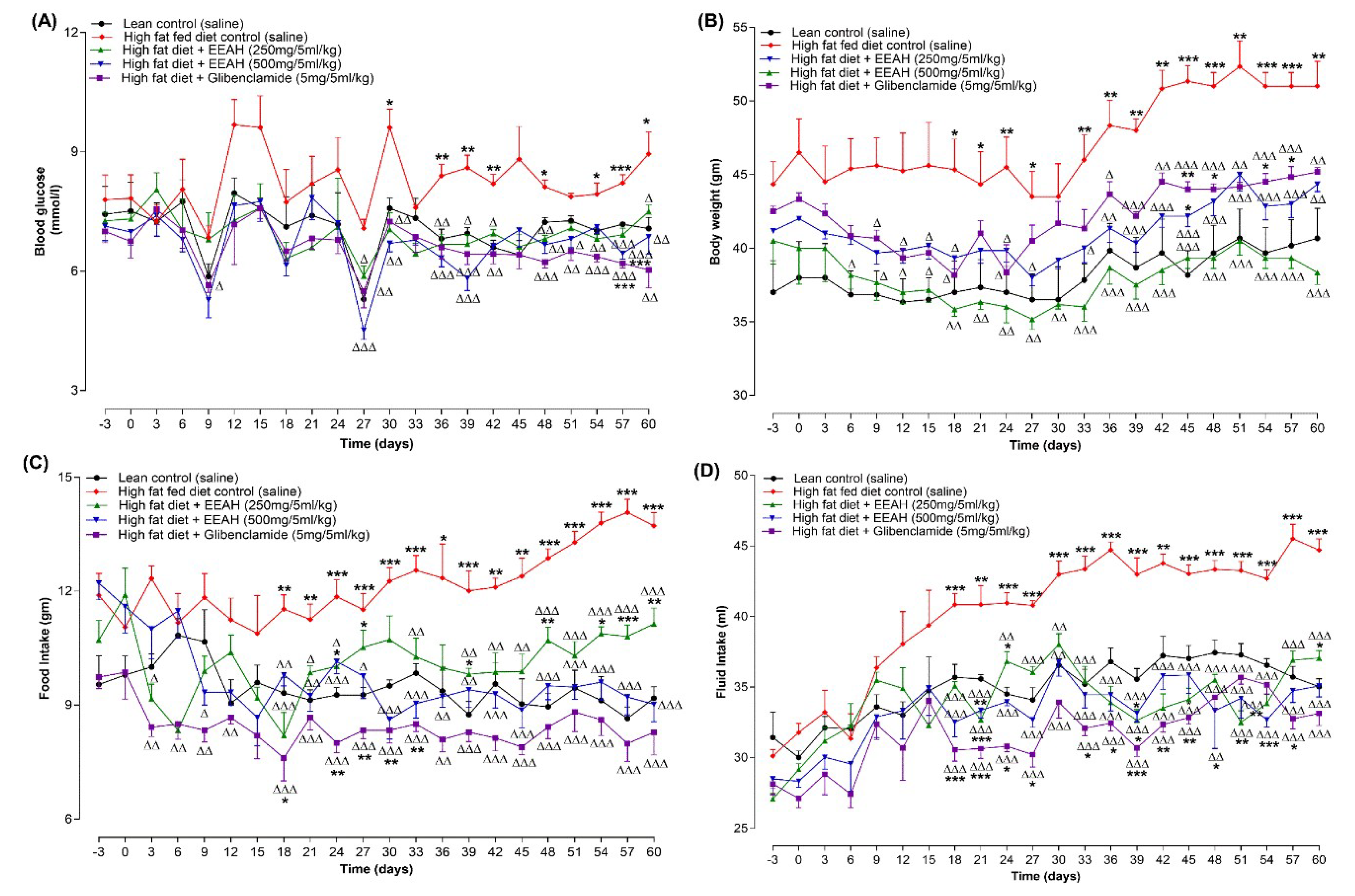
Figure 5.
Effects of 60-days (twice-daily) oral administration of ethanol extract of unripe A. heterophyllus (EEAH) on (A) HDL cholesterol, (B) total cholesterol, (C) triglyceride and (D) LDL cholesterol and (E) gut motility in lean mice and high-fat-fed mice expressed as scatter dot plot. Parameters were measured on day 60 following twice-daily treatment with/without EEAH (250 and 500 mg/5 ml/kg, body weight) or glibenclamide (5 mg/5 ml/kg, body weight) or loperamide (5 mg/5 ml/kg) or bisacodyl (10 mg/5 ml/kg). Gastrointestinal motility was assessed by measuring BaSO4 transit length in 20h fasted mice after ingestion of BaSO4 solution. Values are expressed as mean ± SEM with n = 6 mice. *, **, *** p < 0.05-0.001 compared to control (saline) and Δ, ΔΔ p < 0.05-0.01 compared to high-fat-fed diet control mice. As a positive control, glibenclamide, Loperamide and bisacodyl were used.
Figure 5.
Effects of 60-days (twice-daily) oral administration of ethanol extract of unripe A. heterophyllus (EEAH) on (A) HDL cholesterol, (B) total cholesterol, (C) triglyceride and (D) LDL cholesterol and (E) gut motility in lean mice and high-fat-fed mice expressed as scatter dot plot. Parameters were measured on day 60 following twice-daily treatment with/without EEAH (250 and 500 mg/5 ml/kg, body weight) or glibenclamide (5 mg/5 ml/kg, body weight) or loperamide (5 mg/5 ml/kg) or bisacodyl (10 mg/5 ml/kg). Gastrointestinal motility was assessed by measuring BaSO4 transit length in 20h fasted mice after ingestion of BaSO4 solution. Values are expressed as mean ± SEM with n = 6 mice. *, **, *** p < 0.05-0.001 compared to control (saline) and Δ, ΔΔ p < 0.05-0.01 compared to high-fat-fed diet control mice. As a positive control, glibenclamide, Loperamide and bisacodyl were used.
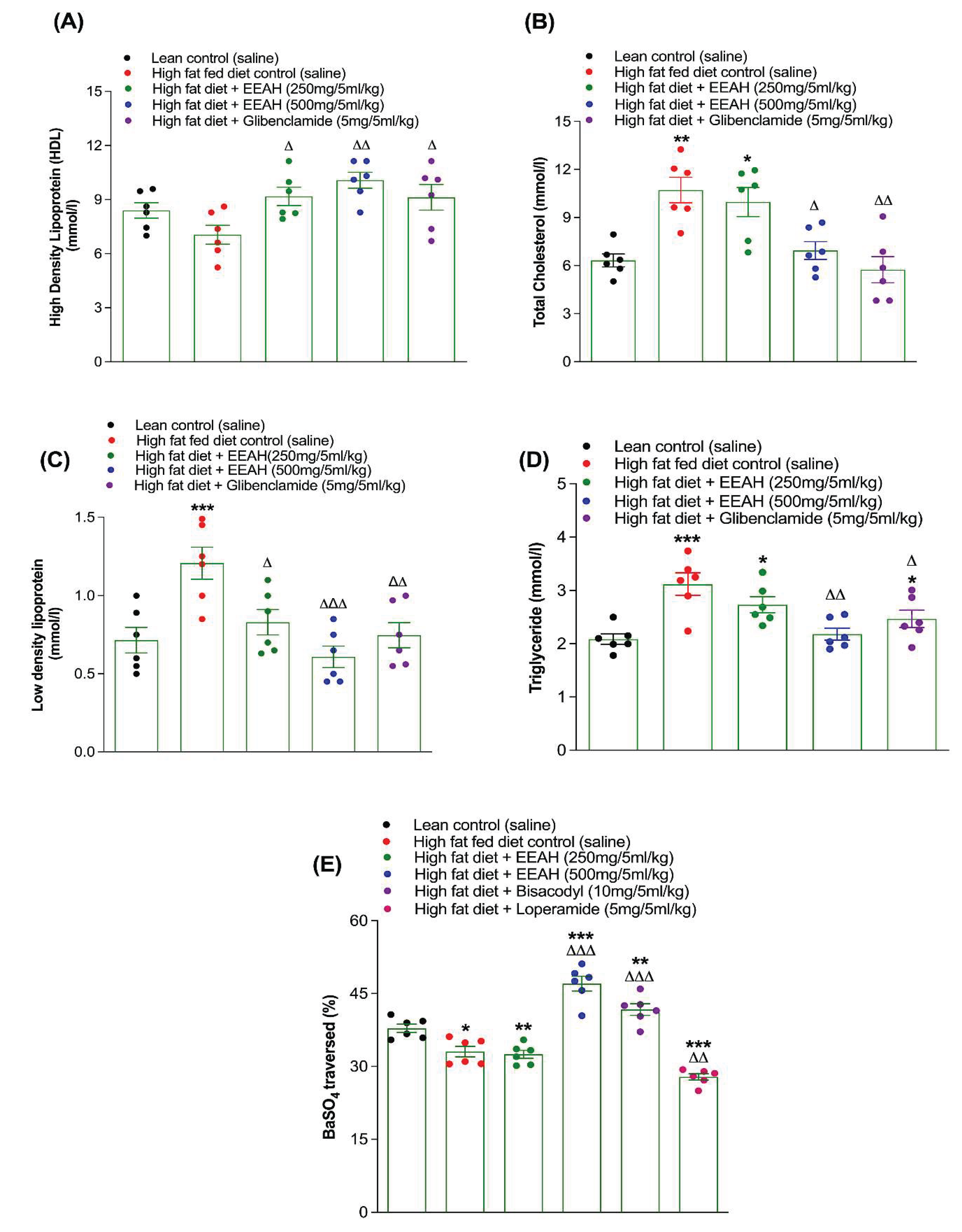

Table 1.
Phytochemical screening of ethanol extract of A. heterophyllus (EEAH) unripe fruit.
| Group Test | Observation |
|---|---|
| Alkaloids | + |
| Tannins | + |
| Flavonoids | + |
| Saponins | + |
| Steroids | + |
| Terpenoids | + |
| Glycoside | ‒ |
| Reducing Sugar | ‒ |
‘+’ indicates present; ‘-’ indicates absent.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



