Submitted:
03 June 2025
Posted:
04 June 2025
You are already at the latest version
Abstract
Calcium (Ca2+) is a macro-mineral essential for the growth, development, yield, and quality of vegetables and fruits. It performs structural, enzymatic, and signaling functions in plants. This review outlines Ca2+ translocation from soil to fruit via the plant xylem network, emphasizing the importance of Ca2+ compartmentalization within fruit cell organelles in developing the Ca2+-deficiency disorder, blossom-end rot (BER). The causes and possible control measures of BER are also discussed. Soil available Ca2+ enters the root apoplast with the water flow and moves towards the xylem via apoplastic or symplastic routes. The transpiration force and growth of organs determine the movement of Ca2+-containing xylem sap to aerial plant parts, including fruits. The final step of fruit-Ca2+ regulation is the partitioning among cellular compartments, which determines susceptibility to Ca2+-deficiency disorders such as BER. Depleting cytosolic and apoplastic Ca2+ due to excessive deposition in organelles such as the vacuole may lead to disintegration of the plasma membrane, resulting in BER, even at high Ca2+ availability at the blossom end of the fruit. BER management requires cultural and physiological practices that ensure Ca2+ translocation to the fruit and proper Ca2+ compartmentalization. The use of BER-resistant and Ca2+-efficient cultivars may also help in BER management. Therefore, a comprehensive understanding of Ca2+ dynamics in plants is crucial for managing BER, reducing production costs, minimizing environmental impact, and enhancing crop productivity.
Keywords:
BER
; Ca2+-deficiency disorder
; cellular organelles
; Ca2+-transportation
; Ca2+-compartmentalization
1. Introduction
Calcium (Ca2+) as a macro-mineral is essential for plant growth and development [1,2,3], and is found within plants in higher quantities than any other divalent inorganic cations. When hydrated, Ca2+ is a big cation with a 41.2 Å ionic radius [4]. Among the macronutrients, Ca2+ is third after nitrogen (N) and potassium (K) in terms of quantity in a plant body, which reflects its essentiality for the plant. The plant requires 1-3 mM Ca2+ for proper growth and development [5]. It plays roles in membrane and cell wall stabilization, cell function, signal transduction, growth and development, gene expression, and stress resistance [6,7,8,9,10]. Ca2+ is required for every cellular compartment, such as cell wall, apoplast, plasma membrane, cytosols, and organelles suspended within cytosols, e.g., vacuoles, endoplasmic reticulum (ER), plastid, Golgi apparatus, and nucleus (Fig.1). Ca2+ concentration varies from 10-7 M to 10-3 M across the cell organelles. Though cytoplasm contains 10-7 M calcium at the resting stage, it increases to 10-5 to 10-4 M in the storage organelles and 10-3 M in the extracellular milieu [11,12]. The Ca2+ content of mitochondrial and nuclear matrices is similar to that of the cytosol. The free Ca2+ content in cytosol and vacuoles is 100–200 nM and 1–10 mM, respectively [13,14], and 60% of the plant Ca2+ remains as calcium pectate. Ca2+ content in plants varies greatly; shoot-Ca2+ ranges from 0.1%-5%, while fruit-Ca2+ varies from 0.2%-0.3% of total dry mass [15]. The proportion of Ca2+ in specific tissues can be more than 10% without affecting plant growth and development [4].
Calcium is essential for cell wall integrity, membrane permeability, and stability and plays a role in the signaling route as a messenger [15,16,17,18]. The available Ca2+ enters the root apoplast with the water flow [19], and moves towards the xylem, following either apoplastic or symplastic routes [20]. Along the xylem water flow, Ca2+ is transported to the leaves and fruits based on their transpiration strength during the daytime, while at night, root pressure does the job [21]. Poor Ca2+ translocation to the fruit or leaf tip can result in Ca2+ deficiency disorders, e.g., blossom-end rot (BER), tip burn, blackheart, brown heart, bitter pit, empty pod, and fruit cracking (Table 1) [20,22,23,24,25,26].
Among numerous Ca2+-deficiency disorders, BER is most prevalent and causes substantial economic losses worldwide. BER often initiates at the fruit blossom part (away from the peduncle) during early fruit growth stages (2-3 weeks following anthesis) [27,28,29,30,31,32,33,34,35,36,37]. Insufficient Ca2+ supply to the rapidly growing fruit tissue causes the disintegration of the plasma membrane and lysis of the middle lamella, resulting in cell plasmolysis and a water-soaked appearance. Subsequent drying develops sunken, brown, and black spots that are limited to the fruit blossom end or can encompass the entire fruit [38]. BER incidence is positively correlated with root’s relative water content, fruit number per plant, potassium (K+) and vitamin C levels of fruits and negatively correlated with plant height, leaf chlorophyll content, total yield, and fruit Ca2+ content in tomato [39,40].
Though agricultural soils are not usually low in Ca2+, Ca2+-deficiency disorders are numerous and cause significant economic losses worldwide. Ca2+ deficiency is usually not manifested by the unavailability of soil Ca2+; instead, it is the soil’s inability to supply sufficient Ca2+ to the affected plant parts [23]. By being phloem immobile, Ca2+ cannot be translocated from the available sources (mature leaves and peduncle-end of fruits) to the deficient sinks (young-growing leaves and blossom-end of fruit). Therefore, Ca2+ fertilization generally does not overcome these physiological disorders, and thus, these disorders (e.g., BER) are complex and challenging to solve. Though the genes linked to calcium deficiency disorders are not well-documented, expression of Ca2+ /H+ antiporters (CAXs) such as CAX1 and sCAX1 [41,42,43,44,45], and calreticulin (CRT) [44,46] may play a role in this regard [47]. There is no straightforward solution for these disorders. Moreover, the uncertainty of the onset of these deficiency disorders complicates the issues regarding their workable solutions [23]. The present study discusses the route of calcium translocation within plants and the causes and control of BER, the most devastating Ca2+-deficiency disorder.
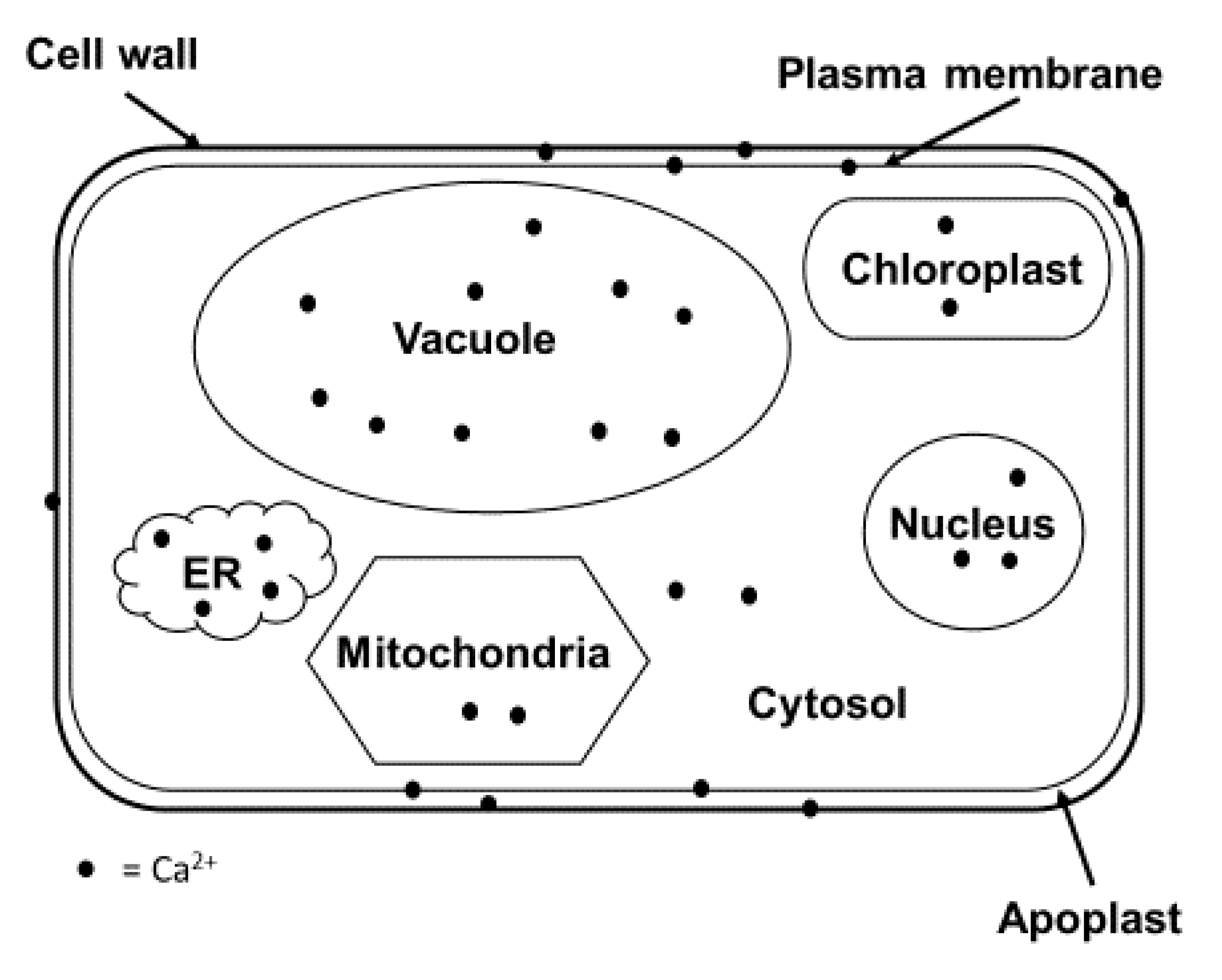
Figure 1.
Calcium compartmentalization within a plant cell. Calcium is required for every part of a plant cell and is an essential mineral. Calcium plays a pivotal role in cell structures such as the cell wall pectin and the plasma membrane, as a signaling molecule in the cytosol, and as a cofactor for several enzymes. Most calcium is found in the vacuoles, which function as an ion balance and pH mediator.
Figure 1.
Calcium compartmentalization within a plant cell. Calcium is required for every part of a plant cell and is an essential mineral. Calcium plays a pivotal role in cell structures such as the cell wall pectin and the plasma membrane, as a signaling molecule in the cytosol, and as a cofactor for several enzymes. Most calcium is found in the vacuoles, which function as an ion balance and pH mediator.
2. Function of Calcium in Plants
Ca2+ performs numerous plant functions [15] as a structural component of cells, enzymatic regulation, and signal transduction [48]. Ca2+ contributes to growth and development via cellular growth, metabolism, and signaling [49].
2.1. Structural Role of Calcium
Calcium maintains cell wall integrity [15,50], cell division and cell elongation [51,52], cell expansion [53], membrane permeability and membrane stability [50,54], and assembly of microtubules [55].
The plant cell wall contains carbohydrates (cellulose, hemicellulose, pectin), proteins, particularly structural ones, and lignin (secondary cell wall). Cell wall Ca2+ mainly represents Ca2+ bindings in the middle lamella that glue adjacent cells and maintain cell wall integrity. Ca2+ is unique among other inorganic elements, which are not usually integral components of cell walls except nitrogen (N). Cell wall Ca2+ generally binds with pectin, a polymer of a diverse group of pectic polysaccharides, including homogalacturonans, rhamnogalacturonan I, and rhamnogalacturonan II. Homogalacturonans are polymers of galacturonic acid in a fashion of α(1–4) linkage. Ca2+ forms a tight linkage between the charged carboxyl (COO-) group of galacturonic acid [56], through which it provides cell wall strength. Pectin forms a gel-like structure by binding with Ca2+ molecules in a reversible fashion that aids in tightening (binding with Ca2+) and loosening (Ca2+ removal) of the cell wall. During the biosynthesis of sugar residues in the Golgi apparatus, the charged carboxyl group can be esterified with methyl, acetyl, or unknown groups that prevent the binding of Ca2+ [56], and keep the cell wall loose. Cell walls are also loosened by the degradation of Ca2+-pectate by polygalacturonase (PG), and the activity of PG is inhibited by high Ca2+ concentration [57]. The PG activity is increased in Ca2+-deficient tissue, leading to middle lamella disintegration, primary cell wall degradation, and cell death. Ca2+ cannot bind to methylated pectin residue. Pectin methyl esterase (PME) removes the methyl group from methylated pectin and opens up free binding sites for Ca2+, where Ca2+ binds to form a strong electrovalent bond [58]. Demethylation of pectin by PME action favors the further degradation of pectin by enzymes like endo-polygalacturonase, exo-polygalacturonase, β-galactosidases, and pectate lyases [59,60].
Besides strengthening the cell wall, Ca2+ plays a role in stabilizing and functioning of the plasma membrane. The plasma membrane is composed of phospholipid bilayers held together by proteins. Ca2+, on the apoplast side of the plasma membrane, binds to the carboxylic group of protein and the phosphate group of phospholipid and thus stabilizes the membrane, allowing a proper membrane selective permeability. The requirement of Ca2+ increases due to an increase in heavy metals [61], aluminum (Al+3), sodium (Na+) [62], and protons (H+) in the external environment. Other cations can replace the Ca2+, but their role is not in proper membrane functioning. Ca2+ in high concentration is required to restrain the unfavorable effects. For plants growing in soil with a higher concentration of other cations, the Ca2+ requirement increased substantially to ensure optimum plant growth and development [63]. Replacement of Ca2+ with Na+, heavy metals, or Al+3 can cause salinity, heavy metal, or aluminum toxicity, respectively [64,65]. Membrane instability is prevalent under freezing, low temperature, and anaerobiosis [4]. Unstable membranes are prone to loss of low molecular weight solutes, such as potassium (K+) and sugars. It can also cause an influx of toxic ions (e.g., the heavy metal Al+3) in the cytosol. A high concentration of free Ca2+ in the apoplast prevents the loss of solutes and helps to avoid potential toxicity from toxic elements. Lack of Ca2+ results in a leaky membrane that causes loss of cell material, impairment of cell metabolism, and subsequent cell death.
Ca2+ stabilizes the cell wall by binding with pectin, the cell membrane, and the proteins and lipids at the membrane surfaces [48,66]. Ca2+ influences vesicles - full of materials and enzymes for cell wall and membrane construction - incorporation into the plasma membrane [53]. Moreover, Ca2+ is required for regulating ion uptake, pH, carbohydrate translocation, the activity of the oxygen-evolving complex, and as a counteraction in the vacuoles for all types of anions [4,16].
2.2. Enzymatic Role of Calcium
Ca2+ can promote or demote enzyme activity essential for cell growth and development. The activity of α-amylase is stimulated by high Ca2+ concentration during starch breakdown in germinating cereal seeds, in which Ca2+ ion stabilizes amylase [67]. However, high Ca2+ concentrations may inhibit enzyme activity [23], as has been shown with cytosolic enzyme fructose-1,6-bisphosphatase (FBPase) (Figure 2), which regulates sucrose synthesis from triosephosphate (TP) in the cytosol. A slight increase in Ca2+ concentration can markedly inhibit the activity of that particular enzyme [4]. Ca2+ is also a cofactor of several enzymes, e.g., 1,4-lactonase, phosphoinositide phospholipase C, N-acetylgalactoseaminyltransferase, and affects the synthesis and transport of those enzymes [68].
2.3. Calcium and Signal Transduction
The interest in Ca2+ in recent years has gained momentum due to its role as a secondary messenger, particularly for developmental and environmental cues [4]. It plays a crucial role as a signaling molecule for signaling pathways [69]. Ca2+ acts as a universal signaling molecule [70], and plays a role in plants' growth, development, and stress management. Environmental stressors initiate cytosolic Ca2+ spikes, activating downstream gene expression and adaptation in adverse conditions [70]. Understanding Ca2+ dynamics may help develop and engineer climate-smart crop varieties [70]. Ca2+ is a stress-response element. Upon sensing stresses, it conveys signals to the downstream protein kinases, leading to phenotypic responses that may result in stress tolerance [71,72,73,74]. It also contributes to immunity by activating immune responses [75]. Characterizing the Ca2+ channels, pumps, and binding proteins is required to comprehend the role of stress signals on Ca2+ homeostasis and adaptive responses [76]. It will improve understanding of how specific stress signals modulate Ca2+ homeostasis to orchestrate adaptive responses [76].
In response to stimuli, Ca2+ transduces signals to the other end upon binding with calmodulin, a calcium protein in the cytosol [17,77,78,79]. Plants maintained a very low (100 to 200 nM) cytosolic [Ca2+], which skyrocketed up to 2 µM at the stimulated state [80]. Plants maintain very low cytosolic [Ca2+ ] to serve as a messenger, to prevent precipitation of inorganic phosphate, and to minimize competition for binding sites with magnesium [4]. The role of Ca2+ as a messenger is possible due to very low cytosolic [Ca2+] and chemistry [20]. Any signal - intensity of light, day length, temperature, salinity, drought, osmotic and oxidative stresses, aluminum toxicity, mechanical injury, anoxia, nodulation, and pathogen attack - exerts an abrupt change in cytosolic [Ca2+] and initiates a Ca2+-signaling pathway [20,78,81]. Besides, the pathway is also activated by various developmental cues, such as germination, cell division and elongation, circadian rhythms, tropic responses, senescence, and apoptosis [4]. The Ca2+-signaling event is location- and time-specific and vital for encoding specific cellular responses [82]. This signaling is subject to judicious regulations as a marked increase in Ca2+-concentration activates Ca+-dependent enzymes, which are harmful to a cell. Therefore, very tight regulation is in place for Ca2+-signaling processes through the coordinated activities of calcium proteins, calcium channels, and efflux systems.
2.3.1. Calcium Proteins
Changes in cytosolic [Ca2+] are detected by specific proteins that either relay or respond to the messages. Upon binding with Ca2+, relay proteins such as calmodulin undergo conformational changes that enable them to interact with a target protein to regulate its function [83,84]. Response proteins such as Ca2+-dependent protein kinases (CDPKs) bind with Ca2+, followed by a conformational change that initiates their intrinsic kinase activity. Cytosolic Ca2+-binding proteins include calmodulins (CaMs), CaM-like proteins, annexins, calcineurin B-like (CBL) proteins, and CDPKs. Calmodulins bind with CaM-binding transcription activators (CAMTAs) and are responsible for gene expression [85,86]. Calmodulins and similar proteins initiate responses to developmental or environmental cues and pathogen attack; CBL to cold, drought, salinity, and wounding; and CDPKs to various stimuli [4]. Plant annexins are associated with cell elongation, membrane repair, the secretory process, salinity, and drought stresses [87]. Several Ca2+-binding proteins, e.g., calreticulin, calnexin, calsequestrin, and BiP (Binding Immunoglobulin protein), are found in the ER and are responsible for protein folding, Ca2+ homeostasis, and modifications at the post-translational stage [4].
2.3.2. Calcium Channels
The membrane-bound calcium channels channel Ca2+ in the cell cytosol from the apoplast space, vacuoles, and ER. The channels are voltage-sensitive and are called depolarization-activated calcium channels (DACCs), hyperpolarization-activated calcium channels (HACCs), and voltage-insensitive calcium channels (VICCs) [84,88,89]. The membrane-bound K+ channel [outward-rectifying (Ca2+ -permeable) K+ channel, KORC] is also considered a calcium-permeable DACC [90]. Calcium channels are activated and perform specific roles to different environmental and developmental signals, such as DACCs, which are activated by stresses such as low temperatures [20,91]; HACCs by pathogen attack, oxidative stresses, cell elongation, and tropism [20,78,92]; and VICCs maintains steady-state cytosolic Ca2+ at resting stage of cell [20]. Ca2+ channels are also found in the tonoplast and ER membrane, allowing Ca2+ to enter the cytosol. Tonoplast-bound channels include HACC, SV (slow-vacuolar), inositol phosphates (IP3, Inositol-1,4,5-triosephosphate; IP6), and cADPR (cyclic ADP-ribose). Tonoplast IP3 may be involved in turgor regulation, cell elongation, tropism, salt stress, and hyperosmotic stress [20,93,94,95], and cADPR in cold adaptations, desiccation tolerance, stomatal behavior, circadian rhythms, and pathogen attack [4]. The IP3, cADPR, and NAADP receptors are also found in the membrane of the ER [20].
2.3.2. Calcium Efflux Systems
Plant cells tightly regulate cytosolic [Ca2+]. Therefore, extra Ca2+ is expelled out to the vacuoles, apoplast, ER, and plastid through active transporters such as Ca+-ATPases and H+/ Ca2+-antiporters (CAX) [20] to aid in proper metabolism in cytoplasm, to restore intra- and extracellular Ca2+-stores, and to remove divalent cations [96,97,98,99,100,101]. As Ca2-transporters, Ca2+-ATPases have high-affinity but low-capacity attributes, and H+/Ca+-antiporters are the opposite [102]. Ca2+-ATPases are located in organelle membranes such as vacuolar membranes (tonoplast), ER membranes, plastid, and cell [103,104,105,106,107,108,109], and ER-type calcium ATPases are found in the Golgi, ER, and endosomes [110,111,112]. CAX are found in the plasma membrane and the vacuolar membrane (tonoplast) [84,102,113,114,115]. Ca2+ serves as the coordinator for cell wall and cytoplasm communication [116].
3. Plant Calcium Uptake by the Root System
The soil Ca2+ may not ensure its availability for the plant unless it has a vigorous system to uptake available Ca2+. The Ca2+ in solution enters with water into the young, un-suberized root. Ca2+ generally enters through the root tip where the suberized endodermis (Casparian strip) [117] is absent, and where the suberized endodermis is broken due to new root growth [52,118]. The Casparian strip is a barrier to apoplastic solute movement, while suberization blocks Ca2+ transfer in endodermal cells [52,118,119]. Thus, the root is the first gateway of entering Ca2+ from the soil solution into the plant system. Intact roots with profuse new growth (volume and size) may exploit more soil volume, which favors higher Ca2+ uptake. Impaired root systems due to hard soil, waterlogged conditions, pathogen attacks, damage by insects and animals, and nematode infestation can reduce Ca2+ uptake.
4. Calcium Uptake Through Foliar Application
Foliar application of Ca2+ can increase leaf- and fruit-Ca2+ and reduce deficiency disorders. Foliar application of Ca2+ strengthens cell walls [120], and tomato leaves absorb 90% of foliar calcium chloride application [121]. Foliar application of Ca2+ @ 0.9% increases fruit Ca2+ and decreases BER in tomatoes [122]. Foliar application of 150% of the recommended dose of calcium nitrate decreases cabbage tip burn [123]. Tip burn of two mini Chinese cabbages (QYH and HN) disappears due to applications of 4-6 mmol.L-1 Ca2+ [124]. Foliar spray of eggshell solutions increases the Ca2+ content on the aerial part of tomato plants and decreases BER in fruits [125]. Calcium foliar application increases defense mechanisms against diseases [120]. Spraying with CaCl2 or Ca(NO3)2 controls blackheart, a Ca2+ deficiency disorder in celery [126,127]. Ca2+-spraying in the form of ‘Calciogreen’ or ‘CaCl2’ or with other calcium formulations either decreases or effectively controls deficiency disorders, including BER in tomato and bell pepper [128,129,130]. However, Ca2+ has minimal mobility within the phloem [131], and thus foliar Ca2+-application may not improve fruit-Ca2+ status [132]. Therefore, foliar absorption and Ca2+ transport are yet to be clarified [121].
5. Calcium Uptake Through the Fruit
One of the leading causes of Ca2+ application is to increase fruit Ca2+ content to reduce deficiency disorders. Soil or foliar applications, decreasing competition at the root zone, and ameliorating plant and environmental issues are all indirect approaches to increasing fruit Ca2+ content. Applying Ca2+ directly to the fruit surface can be another approach. However, it is complex to maintain sufficient fruit Ca2+ [133]. It is noteworthy to recall a four-decade-old comment about the BER complexity - ‘‘the number of possible interactions that can affect Ca2+ uptake and distribution is so great that in the near future, we are unlikely to see the development of cultural practices that will eliminate Ca2+ deficiency, without a direct application of Ca2+ to the susceptible organ’’[134]. Ca2+ applied to the apple fruit surface may penetrate the fruit epidermis [135,136], preferably at 40-50 days after full bloom [137], probably through trichomes and stomata. Young apple fruitlets take up less exogenous Ca2+ than mature fruit [138]; penetration of Ca2+ into the fruit depends on the retention time of the solution on the fruit surface and the concentration of the applied solution [137]. Applying 1.33 g calcium-nitrate/polybag decreases BER in tomatoes [139]. Ca2+ @100 ppm reduces BER incidence in susceptible tomato accession (Solanum lycopersicum lycopersicum) by 5-11% [40]. Bone meal decreases BER in tomato ‘Cobra F1’ by increasing fruit Ca2+ content [140]. Ca2+ application improves the quality of cucumber, Ca2+ content in leaves and peels in pomegranate, and reduces phenolics and flavonoids in cherries [141,142,143]. Instead of fruit calcium (pedicel, proximal half, and distal half) content, Ca2+/ K+ and their relative % in the pedicel are associated with the BER in peppers [144]. BER-resistant peppers express defense responses against calcium-deficient stressors [144]. However, direct Ca2+ application can decrease BER symptoms but cannot control the disorder completely; moreover, exogenous Ca2+ may leak out easily [145,146]. Detection of BER at early stages may lead to appropriate control measures to minimize postharvest losses, as the success rate of detecting BER by fluorescence and image analysis at this stage is above 86% [147].
6. Calcium Translocations
6.1. Calcium Translocations Within the Plant
The soil available Ca2+ enters the root apoplast with the water flow and moves through the xylem following either apoplastic or symplastic routes [19,20]. The apoplastic routes consist of cell walls and intercellular spaces, with Ca2+ traveling following water potential gradients [52,148], and, for the symplastic pathway – made up of cytoplasm – Ca2+ moves through plasmodesmata [52,148]. The apoplastic route is considered the principal route of Ca2+ translocation [20,24,148]. However, the Casparian strip along this route restricts further Ca2+ movement to the xylem. Therefore, Ca2+ enters the root either through the unrubberized endodermis of the root tip [117], or where the endodermis has been broken due to the growth of a new root [24,52,118,148]. Once Ca2+ is in the xylem sap, its further movement within the shoot is regulated by the xylem flow of water, xylem water potential [149,150,151], and cation exchange capacity (CEC) of the xylem cell wall. Ca2+ moves mainly with the xylem flow of water from root to shoot [24]. The canopy's transpiration force and plant growth drive the xylem water flow. Xylem water potential depends on dissolved solutes, and xylem cell wall CEC on available binding sites for Ca2+ in the xylem cell wall. Along with xylem water flow, Ca2+ is transported to the fruits and storage tissues, and this movement is aided by the leaf transpiration strength during the day time and root pressure at night [21].
6.2. Leaf or Fruit?
What determines whether the xylem sap containing Ca2+ will move toward the leaf or the fruit? It is the transpiration force and organ growth [149,150,151]. Transpiration from leaf and fruit surfaces triggers water flow towards them. The leaf, being a strong transpiring organ and a higher mass accumulator than fruit [53,149], results in most of the Ca2+ being deposited in the leaf. Fruit sap uptake can be facilitated by either reducing leaf transpiration or enhancing stomatal closure, leading to better Ca2+ uptake and thus minimizing BER [24,149,150,152]. Ca2+ content does not decrease in the leaves by being phloem-immobile; instead, it may increase due to dehydration during senescence [153,154].
6.3. Calcium Translocation Within the Fruit
Xylem sap Ca2+ enters the fruit through the peduncle and is distributed within the fruit based mainly on the xylem network. Being phloem immobile, Ca2+ accumulation within the fruit depends on fruit transpiration. Fruit transpiration rate is lower than that of leaves, resulting in a low Ca2+ supply to the fruit. High N causes fruit expansion, leading to reduced Ca2+ availability to fruit through dilution [33,155], resulting in BER. Though high [K+] and [Mg2+] may replace plasma membrane Ca2+, they cannot substitute the function of Ca2+ in the membrane [156,157], which may also lead to loss of membrane permeability and make the fruit susceptible to Ca2+ deficiency disorders. Compared to the total fruit Ca2+, the relative Ca2+ contents, such as the ratios N/Ca2+, K+/Ca2+, Mg2+/Ca2+, (K+ +Mg2+)/Ca2+, are better predictors of Ca2+ deficiency disorders such as BER [158,159,160]. Fruits can also regulate Ca2+ translocation by altering aquaporin activity and cell wall properties [161].
7. Calcium Compartmentalization Within the Cell
Partitioning of Ca2+ within cellular compartments is the final step of Ca2+ regulation. Fruit sensitivity to Ca2+ deficiency disorders is triggered by modifying cellular Ca2+-partitioning [41,162]. Ca2+ compartmentalization is regulated by the capacity of binding Ca2+ to the cell wall and the presence of Ca2+ channels, ATPases, and exchangers in the membranes of organelles [163]. The cellular plasma membrane is located between the apoplast and cytosol. Apoplastic Ca2+ includes water-soluble plasma membrane and cell wall Ca2+ [53]. Water-soluble Ca2+ stabilizes the plasma membrane by binding phosphate and carboxylate from phospholipids and proteins, respectively, and keeps it functional [48,65]. A certain threshold of water-soluble Ca2+ is always maintained in the apoplast to avoid membrane damage and leakiness [65,164] and replacement of Ca2+ with other ions can damage the membrane [60,165]. Cell wall Ca2+ binds with the pectin matrix to obtain the rigidity of the cell wall. Newly synthesized pectic polysaccharides are highly methyl-esterified. Removal of a methyl group by pectin methyl esterases (PMEs) creates a carboxylate group with which Ca2+ binds strongly [57,166].
The Ca2+ concentration of organelles varies greatly, and the cell maintains a certain Ca2+ threshold level for its function. The vacuole is the biggest store of Ca2+, maintaining 1-10 mM Ca2+ [20,167]. Other Ca2+ storage sites are the ER (1-5 mM) [168], chloroplast (0.1-10 µM) [169,170], mitochondria (0.2-1.2 µM) [169,170], nucleus (0.1-0.2 µM) [171], and cytosol (100-200 nM) [80] (Figure 1).
8. Calcium Deficiency Disorders
Ca2+ deficiency in crop plants can cause numerous disorders that are responsible for significant crop losses. An economically crucial Ca2+-related disorder is BER. Symptoms of BER include the development of dry, brown/black, sunken spots at the blossom end of fruits, leading to rotting that may cover a significant part of fruits in peppers, tomatoes, watermelon, eggplant, and squash [20,22,24,172] (Figure 3). Ca2+ deficiency leads to physiological disorders in tomatoes, peppers, apples, and watermelons [173,174]. Ca2+ deficiency causes cell death in the apical meristem [175], and weakens the cell wall, leading to disease and pest susceptibility [176].
Tip burn is another relevant physiological disorder. It is characterized by necrosis of rapidly growing young leaves in cabbage, Chinese cabbage, Brussels sprouts, lettuce, chervil, chicory, escarole, onion, fennel, and potatoes [23,25,177,178,179,180] (Table 1). Other disorders include bitter pit – the development of brown/black depressed spots on the blossom end of fruit – in apples [20,134,158]; blackheart – collapsing of young leaf tissue that turned black, usually at the center (heart) of the plant – in celery [126,181]; brown heart – necrosis of the tip of young leaves that cover the entire leaf later – in leafy vegetables [20]; empty pod – poor or no development of seed kernel results in empty pod/shell – in peanut [20]; and fruit cracking – splitting of skin or cuticle – in apple, tomatoes, and cherry [20] (Table 1). Besides deficiency, Ca2+-toxicity is reported in crop plants such as gold spot/yellowish flecks – tiny flecks develop around the calyx and shoulder of fruit – in tomato [184], and Ca2+-toxicity halted germination and growth of vegetables [23].
Figure 3.
Blossom-end rot (BER) in tomato and bell pepper fruits. Blossom-end rot affects the distal end of the fruit and occurs during the first few weeks of fruit growth. BER is considered a calcium deficiency disorder that other environmental conditions can exacerbate. In advanced stages, dry, sunken, black/brown symptoms appear that can cover the entire blossom part of the fruit. Upper panel: BER in tomato; middle panel: development of BER in bell pepper while in the plant; and bottom panel: different stages of development of BER symptoms in bell pepper (from left to right: very low, low, moderate, high, and very high BER). Photos are from the first author’s experiments conducted in Athens, GA, USA, from December 2015 to April 2018.
Figure 3.
Blossom-end rot (BER) in tomato and bell pepper fruits. Blossom-end rot affects the distal end of the fruit and occurs during the first few weeks of fruit growth. BER is considered a calcium deficiency disorder that other environmental conditions can exacerbate. In advanced stages, dry, sunken, black/brown symptoms appear that can cover the entire blossom part of the fruit. Upper panel: BER in tomato; middle panel: development of BER in bell pepper while in the plant; and bottom panel: different stages of development of BER symptoms in bell pepper (from left to right: very low, low, moderate, high, and very high BER). Photos are from the first author’s experiments conducted in Athens, GA, USA, from December 2015 to April 2018.

Tip burn is another relevant physiological disorder. It is characterized by necrosis of rapidly growing young leaves in cabbage, Chinese cabbage, Brussels sprouts, lettuce, chervil, chicory, escarole, onion, fennel, and potatoes [23,25,177,178,179,180] (Table 1). Other disorders include bitter pit – the development of brown/black depressed spots on the blossom end of fruit – in apples [20,134,158]; blackheart – collapsing of young leaf tissue that turned black, usually at the center (heart) of the plant – in celery [126,181]; brown heart – necrosis of the tip of young leaves that cover the entire leaf later – in leafy vegetables [20]; empty pod – poor or no development of seed kernel results in empty pod/shell – in peanut [20]; and fruit cracking – splitting of skin or cuticle – in apple, tomatoes, and cherry [20] (Table 1). Besides deficiency, Ca2+-toxicity is reported in crop plants such as gold spot/yellowish flecks – tiny flecks develop around the calyx and shoulder of fruit – in tomato [184], and Ca2+-toxicity halted germination and growth of vegetables [23].

Table 1.
Calcium deficiency disorders of crops.
| Deficiency symptoms | Crops | Description | Reference |
| Blossom-end rot | Bell pepper, tomato, watermelon, eggplant, Squash | Blossom-end rot in fruit and vegetables develops dry, brown/black, sunken spots, leading to rotting that may cover most of the fruit. | [20,22,24,172,182,183] |
| Blackheart | Celery | Young leaf tissue collapsed and turned black, usually at the center (heart) of celery. | [126,181] |
| Bitter pit | Apple | Development of brown/black depressed spots on the fruits. | [20,134,158] |
| Empty pod | Peanut | Poor or no development of the seed kernel results in an empty pod/shell of the peanut | [20] |
| Tip burn | Cabbage, Chinese cabbage, other cabbages | The tips of rapidly growing young leaves become necrotic | [23,25,178] |
| Brussels sprouts, lettuce |
Necrosis of the tip of rapidly growing young leaves | [23,179,180] | |
| Chervil | Tip of rapidly growing young leaves become necrotic | [23,177] | |
| Chicory, escarole, onion, fennel, potatoes | Necrosis of tip of rapidly growing young leaves | [23] | |
| Brown heart | Leafy vegetables | Necrosis of tip of young leaves that cover the entire leaf later | [20] |
| Fruit cracking | Tomato, cherry, apple | Splitting of skin or cuticle | [20] |
8.1. Genesis of Blossom-End Rot Development
BER is a costly Ca2+ deficiency disorder that decreases the yield and quality of the produce, leading to significant economic losses. The Ca2+ content of the Earth's crust is 3.64%, higher than most minerals [185]. The inadequacy of soil Ca2+ for plant growth is rare [1]. Soil Ca2+ is found in bound, exchangeable, and soluble forms. Bound Ca2+ occurs in the form of Ca2+-minerals, e.g., calcite, dolomite, and apatite [1,185], and is not readily available to the plant. Exchangeable Ca2+ remains bound with the soil in negative sites (cation exchange capacity, CEC). It can be available to the plants based on soil pH and the presence of other competing cations. The soil solution Ca2+ is readily available for the plants to be taken up. Soil solution Ca2+ depends on the weathering of parent rock material, the mineralization of primary minerals and soil organic matter, soil pH, fertilization, and diffusion along the gradient [2]. Just the presence of Ca2+ in the soil may not ensure its availability to the plant. Ca2+ depends on water availability, the competition of Ca2+ with other cations, medium pH, salinity, root growth, anoxia, root zone temperature, and root damage by pathogens, insects, and nematodes [186]. Ca2+ can be supplied to the plants by applying Ca2+ in the soil and on leaf and fruit surfaces.
Cations decrease while anions increase Ca2+ uptake by the plants. The presence of cations such as K+, manganese (Mn2+), magnesium (Mg2+), ammonium (NH4+), Al 3+, and Na+ antagonize [24,34,187], and anions such as nitrate (NO3-) and phosphate (PO43−) synergize Ca2+ uptake by the plant root system. Although soil contains about 10 times more Ca2+ than K+, the uptake of Ca2+ is lower than that of K+ [65], which might be due to the higher valency of Ca2+ [15]. NH4+ competes with Ca2+ to be taken up by the plants. Moreover, high N fertilization promotes shoot growth, which diverts absorbed Ca2+ to the leaf instead of the fruit because of the higher leaf transpiration rate than the fruit [53,188]. At high soil calcium availability, fruit Ca2+-deficiency disorder may not appear. However, at a low calcium availability, BER can appear due to depletion of apoplastic Ca2+ content. BER may also occur at high calcium availability due to improper Ca2+ compartmentalization (Figure 4).
8.2. Incidence of BER Based on Variety, Season, and Truss
The incidence of BER may vary from variety to variety, as reported from cultivated peppers and tomatoes. Ca2+ deficiency, differential fruit growth rate, and variation in xylem development are considered the basis of this variability [189]. Ca2+-efficient cultivars absorbed Ca2+ more efficiently than Ca2+-inefficient cultivars when the availability of Ca2+ is low on the substrate. Thus, Ca2+-efficient cultivars could be selected against BER, but the difficulty is that they yield poorly. However, no significant difference in BER susceptibility has been found between efficient and inefficient varieties [189]. Tomato varieties ‘Calypso’ and ‘Spectra’ showed higher incidences of BER than ‘Counter’ [190]; ‘Petomech II’ higher than ‘IPA-L’ [191]; ‘Celebrity’ higher than ‘Rutgers,’ ‘Mountain Pride’, and ‘Mountain Spring’ [192]; STEP 158 (breeding line) higher than ‘Rutgers’ and ‘Doublerich’ [193], and ‘Boludo’ higher than ‘Daniela’ [194]. Elongated tomato varieties are more susceptible to BER [195]. BER never occurs in small-fruit and wild tomato cultivars [53]. Lack or excess of minerals may cause deficiency disorders [196,197]. A low fruit apoplastic calcium, which results in leaky cell membranes, leads to BER development [198]. BER occurs in tomatoes if the calcium concentrations of the lateral ends of green fruits are < 0.2 µmol g-1 [199]. The fruit growth rate of the BER-susceptible tomato accession (Solanum lycopersicum lycopersicum) is higher than the BER-resistant one (Solanum lycopersicum cerasiforme), however, it is not clear whether this may contribute to BER development [186]. Phytohormones may also affect BER incidence. Foliar spray of ABA reduces BER incidence by increasing calcium availability [38,199], and gibberellins increase it through increasing oxidative stresses in plants [200]. ABA increases, and GA decreases calcium accumulation [144]. Thus, applying ABA or GA inhibitors (paclobutrazol and prohexadione-Ca2+) may decrease BER in peppers [144]. Transport and homeostasis of calcium ions are crucial for preventing BER in peppers [144].
Pepper varieties with larger final fruit sizes and faster growth rates, such as ‘Marconi’ and ‘J27’, had higher sensitivity to BER than ‘Jerid’, which produces a smaller final fruit size [202]. No incidence of BER has been reported from wild-type tomatoes (small fruit size). This observation indicates that BER might be associated with larger fruits under favorable growth conditions that influence rapid fruit growth. ‘Marmande’ tomatoes had a higher BER incidence than cherry, cocktail, or round tomatoes [203]. Pygmy fruits, having no rapid growth phase, usually do not develop BER [204]. BER-affected tomatoes ripen earlier and are smaller than healthy fruit [205].
The onset of BER can vary based on trusses and seasons. Frequent incidence of BER was observed with the first truss [36] followed by a subsequent decrease [31,206,207], or increase [207,208]. Basal fruits of a truss had more severe BER than the others [28]. Based on seasons, BER incidence can increase or decrease from the first to the upper trusses [151,190].
9. Control of BER
BER, a critical physiological disorder for several vegetables, has been studied for over a century, although the mechanism is unclear. Most researchers agree that this condition is a Ca2+-deficiency disorder, and supplying sufficient Ca2+ to the fruit may prevent the symptom development. However, the Ca2+ route from soil to the target organ, i.e., fruit, is not straightforward. Many factors are in action on the journey of Ca2+ from the soil to the fruit. Moreover, it is not the effect of a single factor, rather a combined effect of one or more factors [24], such as low soluble soil Ca2+, high Mg2+, NH4+, and K+ concentrations, high salinity, inconsistent soil moisture (high, low or fluctuating), rapid fruit growth rate, poor xylem network towards the blossom end of the fruit, high temperature, and high or low transpiration of the target organ [24]. Cultural management approaches should favor Ca2+ translocation to the fruit to control BER. However, the transport of Ca2+ to the fruit may not ensure BER control, as cellular Ca2+ partitioning is the final and most crucial control level for this disorder [163]. Reports show severe BER incidence in the distal part of the fruit despite high Ca2+-concentration [211]. Moreover, no BER symptoms when the Ca2+-supply was low in the case of slow-growing plants [212] (Figure 3). The relatively high Ca2+ concentration in the BER-affected fruit might be explained through abnormal cellular partitioning, such as transport of abundant Ca2+ to the vacuoles, which may cause Ca2+ deficiency in other sub-cellular organelles and may develop BER. Moreover, an unanticipated change in cellular Ca2+ concentration response to environmental stimuli or hormonal effects may cause localized Ca2+ deficiency, leading to BER development [163].
BER occurs due to abiotic stresses, high temperature, drought, salinity, water logging, higher transpiration, production of ROS, and low availability of ascorbic acid [195,214]. Control of BER is complicated due to the involvement of many changing and unpredictable abiotic factors, and proper management can only reduce the incidence rate [36]. Spraying calcium on the fruit during the developmental stage reduces BER incidence [53]. Spraying should be started at the early stage of the fruit and, of course, before the onset of BER, and it needs to be continued for the entire development stage. However, spraying may not be effective [195]. Balanced fertilizer and avoiding vigorous foliage growth may help reduce BER [195]. Shade net may also reduce BER incidence [215,216,217,218,219]. However, the incidence of BER in tomatoes is positively correlated with fruit plant-1, K content, root relative water content, firmness, vitamin C, titratable acidity, and peroxidase, and negatively correlated with calcium content [39]. A high calcium concentration (e.g., 20 mmol.L-1) decreases growth [plant height, diameter, biomass production (leaf, stem, root, and total)], physiological attributes [photosynthesis, stomatal conductance, transpiration, and chlorophyll content (chlorophyll a; chlorophyll b)], enzyme levels (superoxide dismutase, catalase, and peroxidase), and water use efficiency in poplar seedlings [49]. Therefore, correctly understanding calcium nutrition helps reduce crop cultivation costs, minimize environmental pollution, and boost crop production [220].
Cultural and physiological crop management that ensures Ca2+ transportation to the fruit and appropriate cellular distribution may reduce the incidence of BER. The selection of BER-resistant and Ca2+-efficient cultivars may also help to reduce BER incidence. Controlling or skipping calcium deficiency disorders may include modifications of watering, light, temperature, transpiration, and application of mulching and growth regulators. Using resistant varieties and customization of planting and harvesting (early planting and harvesting) may help skip calcium deficiency disorders [23]. Understanding the molecular mechanisms of BER may help in better managing the disorder [221].With the combination of appropriate management practices and BER-resistant cultivars, the incidence of this disorder may be eliminated in the future [53].
10. Conclusions
Calcium is a crucial element for plant growth and development. Though soils worldwide are not typically deficient in Ca2+, Ca2+-deficiency disorders in crops are numerous and cause substantial yield loss. Among the Ca2+-deficiency disorders, BER is widely prevalent worldwide. The complex route of Ca2+ from soil to the appropriate cellular compartments, such as the cytosol depends on multiple factors, e.g., soil (moisture availability, the competition of Ca2+ with other cations, pH, anoxia, and salinity), plant [genotypes, growth habit (dwarf, tall), xylem network, root and shoot growth, yield, root damaged by pathogens, insects, and nematodes], and environment [temperature (air and root zone), relative humidity, vapor pressure deficit, and transpiration] that renders it difficult to control BER. Moreover, the cellular Ca2+ compartmentalization, particularly in the vacuoles, depletes cytosolic Ca2+ levels and may disintegrate the plasma membrane, leading to BER development despite having high Ca2+ content in the blossom end of the fruit. Therefore, possible ways of minimizing and controlling BER include i) an integrated approach that ameliorates soil, plant, and environmental factors towards supplying sufficient Ca2+ into the cells; ii) appropriate cultural and physiological management of crops; iii) a favorable environment; and iv) BER-resistant and Ca2+-efficient cultivars. Other approaches that include agronomic, physiological, breeding, and molecular methods may also contribute to minimizing BER occurrence.
Author Contributions
MYK: Conceptualization; Writing – original draft, review & editing; JCD: Writing – review & editing, Supervision, Conceptualization
Funding
This review is part of the first author's Ph.D. program supported by the United States Agency for International Development as part of the Feed the Future initiative under the CGIAR Fund, award number BFS-G-11-00002, and the predecessor fund, the Food Security and Crisis Mitigation II grant, award number EEM-G-00-04-00013.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Keiser, J. R., & Mullen, R. E. (1993). Calcium and relative humidity effects on soybean seed nutrition and seed quality. Crop Science, 33(6), 1345-1349. [CrossRef]
- McLaughlin, S. B., & Wimmer, R. (1999). Tansley Review No. 104 Calcium physiology and terrestrial ecosystem processes. The New Phytologist, 142(3), 373-417. [CrossRef]
- Zartdinova, R., & Nikitin, A. (2023). Calcium in the life cycle of legume root nodules. Indian Journal of Microbiology, 63(4), 410-420. [CrossRef]
- Hawkesford, Malcolm, Walter Horst, Thomas Kichey, Hans Lambers, Jan Schjoerring, Inge Skrumsager Møller, and Philip White. 2012. 'Chapter 6 - Functions of Macronutrients A2 - Marschner, Petra.' in Marschner's Mineral Nutrition of Higher Plants (Third Edition) (Academic Press: San Diego).
- Saito, S., & Uozumi, N. (2020). Calcium-regulated phosphorylation systems controlling uptake and balance of plant nutrients. Frontiers in Plant Science, 11, 44. [CrossRef]
- Kim, M. C., Chung, W. S., Yun, D. J., & Cho, M. J. (2009). Calcium and calmodulin-mediated regulation of gene expression in plants. Molecular plant, 2(1), 13-21. [CrossRef]
- Feng, D., Wang, X., Gao, J., Zhang, C., Liu, H., Liu, P., & Sun, X. (2023). Exogenous calcium: Its mechanisms and research advances involved in plant stress tolerance. Frontiers in Plant Science, 14, 1143963. [CrossRef]
- Hu, W., Liu, J., Liu, T., Zhu, C., Wu, F., Jiang, C., ... & Zheng, H. (2023). Exogenous calcium regulates the growth and development of Pinus massoniana detecting by physiological, proteomic, and calcium-related gene expression analysis. Plant Physiology and Biochemistry, 196, 1122-1136. [CrossRef]
- Hu, W., Tian, S. B., Di, Q., Duan, S. H., & Dai, K. (2018). Effects of exogenous calcium on mesophyll cell ultrastructure, gas exchange, and photosystem II in tobacco (Nicotiana tabacum Linn.) under drought stress. Photosynthetica, 56(4), 1204-1211. [CrossRef]
- Sun, X., Pan, B., Wang, Y., Xu, W., & Zhang, S. (2020). Exogenous calcium improved resistance to Botryosphaeria dothidea by increasing autophagy activity and salicylic acid level in pear. Molecular Plant-Microbe Interactions, 33(9), 1150-1160. [CrossRef]
- Alonso, M. T., Torres-Vidal, P., Calvo, B., Rodriguez, C., Delrio-Lorenzo, A., Rojo-Ruiz, J., ... & Patel, S. (2023). Use of aequorin-based indicators for monitoring Ca2+ in acidic organelles. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 1870(6), 119481. [CrossRef]
- Vaughan, M. K. (2020). Arginine vasotocin and vertebrate reproduction. In The Pineal Gland (pp. 125-163). CRC Press.
- Lecourieux, D., Mazars, C., Pauly, N., Ranjeva, R., & Pugin, A. (2002). Analysis and effects of cytosolic free calcium increases in response to elicitors in Nicotiana plumbaginifolia cells. The Plant Cell, 14(10), 2627-2641. [CrossRef]
- Carafoli, E. (2005). Calcium–a universal carrier of biological signals: delivered on 3 July 2003 at the Special FEBS Meeting in Brussels. The FEBS journal, 272(5), 1073-1089. [CrossRef]
- Marschner, Horst. 1995. '8 - Functions of Mineral Nutrients: Macronutrients.' In, Mineral Nutrition of Higher Plants (Second Edition) (Academic Press: London).
- Kulik, L. V., Epel, B., Lubitz, W., & Messinger, J. (2007). Electronic structure of the Mn4O x Ca cluster in the S0 and S2 States of the oxygen-evolving complex of photosystem II based on pulse 55Mn-ENDOR and EPR spectroscopy. Journal of the American Chemical Society, 129(44), 13421-13435. [CrossRef]
- McAinsh, M. R., & Hetherington, A. M. (1998). Encoding specificity in Ca2+ signalling systems. Trends in Plant Science, 3(1), 32-36. [CrossRef]
- Simon, E. W. (1978). The symptoms of calcium deficiency in plants. New phytologist, 80(1), 1-15. [CrossRef]
- Barber, S. A. (1995). Soil nutrient bioavailability: a mechanistic approach. John Wiley & Sons.Barickman, T. C., Kopsell, D. A., & Sams, C. E. (2014). Foliar applications of abscisic acid decrease the incidence of blossom-end rot in tomato fruit. Scientia Horticulturae, 179, 356-362. [CrossRef]
- White, P.J., Broadley, M.R., 2003. Calcium in plants. Annals of Botany, 92(4), 487-511. [CrossRef]
- Morard, P., Lacoste, L., & Silvestre, J. (2000). Effects of calcium deficiency on nutrient concentration of xylem sap of excised tomato plants. Journal of Plant Nutrition, 23(8), 1051-1062. [CrossRef]
- Díaz-P´erez, J.C., Hook, J.E., 2017. Plastic-mulched bell pepper (Capsicum annuum L.) plant growth and fruit yield and quality as influenced by irrigation rate and calcium fertilization. HortScience 52, 774–781. [CrossRef]
- Olle, M., & Bender, I. (2009). Causes and control of calcium deficiency disorders in vegetables: A review. The Journal of Horticultural Science and Biotechnology, 84(6), 577-584. [CrossRef]
- Taylor, M. D., & Locascio, S. J. (2004). Blossom-end rot: a calcium deficiency. Journal of Plant nutrition, 27(1), 123-139. [CrossRef]
- Aloni, B., Pashkar, T., Libel, R., 1986. The possible involvement of gibberellins and calcium in tipburn of Chinese cabbage: study of intact plants and detached leaves. Plant Growth Regulation, 4, 3-11. [CrossRef]
- Geraldson, C. M. (1956). The use of calcium for control of blossom-end rot of Tomatoes. Proceedings of the Florida State Horticultural Society, 68: 197-202.
- Adams, P., & El-Gizawy, A. M. (1988). Effect of calcium stress on the calcium status of tomatoes grown in NFT. Acta Horticulturae, 222: 15-22.
- Banuelos, G. S., Offermann, G. P., & Seim, E. C. (1985). High relative humidity promotes blossom-end rot on growing tomato fruit. HortScience, 20(5), 894-895.
- Barke, R. E. (1968). Absorption and translocation of calcium foliar sprays in relation to the incidence of blossom-end rot in tomatoes. Queensland Journal of Agricultural and Animal Sciences, 25(4), 179-197.
- Cho, I. H., Lee, E. H., Kim, T. Y., Woo, Y. H., & Kwon, Y. S. (1998). Effects of high humidity on occurrence of tomato blossom-end rot. Journal of the Korean Society for Horticultural Science (Korea Republic), 39(3). 47-249.
- El-Gizawy, A. M., & Adams, P. (1985, June). Effect of temporary calcium stress on the calcium status of tomato fruit and leaves. In Symposium on Nutrition, Growing Techniques and Plant Substrates 178: 37-44. 10.17660/ActaHortic.1986.178.3.
- Gutteridge, C. G., & Bradfield, E. G. (1983). Root pressure stops blossom-end rot. Grower 100(6): 25-26.
- Saure, M. C. (2001). Blossom-end rot of tomato (Lycopersicon esculentum Mill.)—a calcium-or a stress-related disorder? Scientia Horticulturae, 90(3-4), 193-208. [CrossRef]
- Spurr, A. R. (1959). Anatomical aspects of blossom-end rot in the tomato with special reference to calcium nutrition. Hilgardia 28 (12): 269-295.
- Wada, T., Ikeda, H., Ikeda, M., & Furukawa, H. (1996). Effects of foliar application of calcium solutions on the incidence of blossom-end rot of tomato fruit. Journal of the Japanese Society for Horticultural Science, 65(3), 553-558. [CrossRef]
- Westerhout, J. (1962). Relation of fruit development to the incidence of blossom-end rot of tomatoes. Netherlands Journal of Agricultural Science, 10(3), 223-234. [CrossRef]
- Wui, M., & Takano, T. (1995). Effect of Temperature and Concentration of Nutrient Solution during the Stage of the Fruit Development on the Incidence of Blossom-End Rot in Fruits of Tomato, Lycopersicon esculentum L. Environment Control in Biology, 33(1), 7-14.
- De Freitas, S. T., McElrone, A. J., Shackel, K. A., & Mitcham, E. J. (2014). Calcium partitioning and allocation and blossom-end rot development in tomato plants in response to whole-plant and fruit-specific abscisic acid treatments. Journal of Experimental Botany, 65(1), 235-247. [CrossRef]
- Abdelkader, M., Elkhawaga, F. A., Suliman, A. A., Puchkov, M., Kuranova, K. N., Mahmoud, M. H., & Abdelkader, M. F. (2024). Understanding the Regular Biological Mechanism of Susceptibility of Tomato Plants to Low Incidences of Blossom-End Rot. Horticulturae, 10(6), 648. [CrossRef]
- Kabir, M. Y., Díaz-Pérez, J. C., Doyle, J. W., Berenguer, E. I., van der Knaap, E., & Nambeesan, S. U. (2017, September). The Effect of Calcium Application and Irrigation on Development of Blossom-end Rot in Tomato. In 2017 ASHS Annual Conference. ASHS.
- De Freitas, S. T., Padda, M., Wu, Q., Park, S., & Mitcham, E. J. (2011a). Dynamic alternations in cellular and molecular components during blossom-end rot development in tomatoes expressing sCAX1, a constitutively active Ca2+/H+ antiporter from Arabidopsis. Plant Physiology, 156(2), 844-855. [CrossRef]
- Gao, H., Wu, X., Zorrilla, C., Vega, S. E., & Palta, J. P. (2020). Fractionating of calcium in tuber and leaf tissues explains the calcium deficiency symptoms in potato plant overexpressing CAX1. Front. Plant Sci., 10, 1793. [CrossRef]
- Park, S., Kim, C. K., Pike, L. M., Smith, R. H., & Hirschi, K. D. (2004). Increased calcium in carrots by expression of an Arabidopsis H+/Ca 2+ transporter. Molecular Breeding, 14, 275-282. [CrossRef]
- Wu, Q., Shigaki, T., Han, J. S., Kim, C. K., Hirschi, K. D., & Park, S. (2012). Ectopic expression of a maize calreticulin mitigates calcium deficiency-like disorders in sCAX1-expressing tobacco and tomato. Plant Mol. Biol., 80, 609-619. [CrossRef]
- Zorrilla, C., Schabow, J. E., Chernov, V., & Palta, J. P. (2019). CAX1 vacuolar antiporter overexpression in potato results in calcium deficiency in leaves and tubers by sequestering calcium as calcium oxalate. Crop Science, 59(1), 176-189. [CrossRef]
- Wyatt, S. E., Tsou, P. L., & Robertson, D. (2002). Expression of the high capacity calcium-binding domain of calreticulin increases bioavailable calcium stores in plants. Transgenic Research, 11, 1-10. [CrossRef]
- Kuronuma, T., & Watanabe, H. (2021). Identification of the Causative Genes of Calcium Deficiency Disorders in Horticulture Crops: A Systematic Review. Agric, 11, 906. [CrossRef]
- Hirschi, K. D. (2004). The calcium conundrum. Both versatile nutrient and specific signal. Plant Physiology, 136(1), 2438-2442. [CrossRef]
- Weng, X., Li, H., Ren, C., Zhou, Y., Zhu, W., Zhang, S., & Liu, L. (2022). Calcium regulates growth and nutrient absorption in poplar seedlings. Front. Plant Sci., 13, 887098. [CrossRef]
- Kitano, M., Araki, T., Yoshida, S., & Eguchi, H. (1999). Dependence of calcium uptake on water absorption and respiration in roots of tomato plants (Lycopersicon esculentum Mill.). Biotronics: Environment Control and Environmental Biology || 28 |, p121-130. http://133.5.207.201/index.html.
- Hewitt, E. J. (1963). The essential nutrient elements: Requirements and interactions. Plant Physiology, 137-360.
- White, P. J. (2001). The pathways of calcium movement to the xylem. Journal of Experimental Botany, 52(358), 891-899. [CrossRef]
- Ho, L. C., & White, P. J. (2005). A cellular hypothesis for the induction of blossom-end rot in tomato fruit. Annals of Botany, 95(4), 571-581. [CrossRef]
- Epstein, E. (1972). Mineral Nutrition of Plants: Principles and Perspectives. (New York: John Wiley & Sons).
- Fuller, G. M., Ellison, J. J., McGill, M., Sordahl, L. A., & Brinkley, B. R. (1975). Studies on the inhibitory role of calcium in the regulation of microtubule assembly in vitro and in vivo. Microtubules and microtubule inhibitors, 379-390.
- Taiz, L, and E Zeiger. 2011. 'Plant Physiology Online Fifth Edition [WWW Document]', URL http://5e. plantphys. net.
- Wehr, J. B., Menzies, N. W., & Blamey, F. P. C. (2004). Inhibition of cell-wall autolysis and pectin degradation by cations. Plant Physiology and Biochemistry, 42(6), 485-492. [CrossRef]
- Ralet, M. C., Dronnet, V., Buchholt, H. C., & Thibault, J. F. (2001). Enzymatically and chemically de-esterified lime pectins: characterisation, polyelectrolyte behaviour and calcium binding properties. Carbohydrate Research, 336(2), 117-125. [CrossRef]
- Goulao, L. F., Santos, J., de Sousa, I., & Oliveira, C. M. (2007). Patterns of enzymatic activity of cell wall-modifying enzymes during growth and ripening of apples. Postharvest Biology and Technology, 43(3), 307-318. [CrossRef]
- Massiot, P., Baron, A., & Drilleau, J. F. (1994). Characterisation and enzymatic hydrolysis of cell-wall polysaccharides from different tissue zones of apple. Carbohydrate Polymers, 25(3), 145-154. [CrossRef]
- Wallace, A., Frolich, E., & Lunt, O. R. (1966). Calcium requirements of higher plants. Nature, 209(5023), 634-634. [CrossRef]
- Lahaye, P. A., & Epstein, E. (1971). Calcium and salt toleration by bean plants. Physiologia Plantarum, 25(2), 213-218. [CrossRef]
- Asher, C. J., & Edwards, D. G. (1983). Modern solution culture techniques. In Inorganic plant nutrition (pp. 94-119). Berlin, Heidelberg: Springer Berlin Heidelberg. [CrossRef]
- Cramer, G. R. (2002). Sodium-calcium interactions under salinity stress. In Salinity: Environment-plants-molecules (pp. 205-227). Dordrecht: Springer Netherlands.
- Horst, W. J., Wang, Y., & Eticha, D. (2010). The role of the root apoplast in aluminium-induced inhibition of root elongation and in aluminium resistance of plants: a review. Annals of Botany, 106(1), 185-197. [CrossRef]
- Kirkby, E. A., & Pilbeam, D. J. (1984). Calcium as a plant nutrient. Plant, Cell & Environment, 7(6), 397-405. [CrossRef]
- Lovegrove, A., & Hooley, R. (2000). Gibberellin and abscisic acid signalling in aleurone. Trends in Plant Science, 5(3), 102-110. [CrossRef]
- Bush, D. S., Cornejo, M. J., Huang, C. N., & Jones, R. L. (1986). Ca2+-stimulated secretion of α-amylase during development in barley aleurone protoplasts. Plant Physiology, 82(2), 566-574. [CrossRef]
- Luan, S., & Wang, C. (2021). Calcium signaling mechanisms across kingdoms. Annual Review of Cell and Developmental Biology, 37(1), 311-340. [CrossRef]
- Naz, M., Afzal, M. R., Raza, M. A., Pandey, S., Qi, S., Dai, Z., & Du, D. (2024). Calcium (Ca2+) signaling in plants: A plant stress perspective. South African Journal of Botany, 169, 464-485. [CrossRef]
- Bu, Y., Fu, W., Chen, J., Takano, T., & Liu, S. (2021). Description of AtCAX4 in response to abiotic stress in Arabidopsis. International Journal of Molecular Sciences, 22(2), 856. [CrossRef]
- Pirayesh, N., Giridhar, M., Khedher, A. B., Vothknecht, U. C., & Chigri, F. (2021). Organellar calcium signaling in plants: An update. biochimica et biophysica Acta (bbA)-molecular Cell Research, 1868(4), 118948. [CrossRef]
- Srivastava, A. K., Shankar, A., Nalini Chandran, A. K., Sharma, M., Jung, K. H., Suprasanna, P., & Pandey, G. K. (2020). Emerging concepts of potassium homeostasis in plants. Journal of Experimental Botany, 71(2), 608-619. [CrossRef]
- Xiao, Y., Ma, C., Li, M., Zhangzhong, L., Song, P., & Li, Y. (2023). Interaction and adaptation of phosphorus fertilizer and calcium ion in drip irrigation systems: the perspective of emitter clogging. Agricultural Water Management, 282, 108269. [CrossRef]
- Verma, S., Negi, N. P., Narwal, P., Kumari, P., Kisku, A. V., Gahlot, P., ... & Kumar, D. (2022). Calcium signaling in coordinating plant development, circadian oscillations and environmental stress responses in plants. Environmental and Experimental Botany, 201, 104935. [CrossRef]
- Ghosh, S., Bheri, M., & Pandey, G. K. (2021). Delineating calcium signaling machinery in plants: tapping the potential through functional genomics. Current Genomics, 22(6), 404-439. [CrossRef]
- Fedrizzi, L., Lim, D., & Carafoli, E. (2008). Calcium and signal transduction. Biochemistry and Molecular Biology Education, 36(3), 175-180. [CrossRef]
- Lecourieux, D., Ranjeva, R., & Pugin, A. (2006). Calcium in plant defence-signalling pathways. New Phytologist, 171(2), 249-269. [CrossRef]
- Paliyath, G., & Thompson, J. E. (1987). Calcium-and calmodulin-regulated breakdown of phospholipid by microsomal membranes from bean cotyledons. Plant Physiology, 83(1), 63-68. [CrossRef]
- Rudd, J. J., & Franklin-Tong, V. E. (1999). Calcium signaling in plants. Cellular and Molecular Life Sciences CMLS, 55, 214-232. [CrossRef]
- McAinsh, M. R., & Pittman, J. K. (2009). Shaping the calcium signature. New Phytologist, 181(2), 275-294. [CrossRef]
- Putney Jr, J. W. (1998). Calcium signaling: Up, down, up, down.... What's the point? Science, 279(5348), 191-192. [CrossRef]
- Luan, S., Kudla, J., Rodriguez-Concepcion, M., Yalovsky, S., & Gruissem, W. (2002). Calmodulins and calcineurin B–like proteins: Calcium sensors for specific signal response coupling in plants. The Plant Cell, 14(suppl_1), S389-S400. [CrossRef]
- Sanders, D., Pelloux, J., Brownlee, C., & Harper, J. F. (2002). Calcium at the crossroads of signaling. The Plant Cell, 14(suppl_1), S401-S417. [CrossRef]
- Bouché, N., Scharlat, A., Snedden, W., Bouchez, D., & Fromm, H. (2002). A novel family of calmodulin-binding transcription activators in multicellular organisms. Journal of Biological Chemistry, 277(24), 21851-21861. [CrossRef]
- Doherty, C. J., Van Buskirk, H. A., Myers, S. J., & Thomashow, M. F. (2009). Roles for Arabidopsis CAMTA transcription factors in cold-regulated gene expression and freezing tolerance. The Plant Cell, 21(3), 972-984. [CrossRef]
- Laohavisit, A., & Davies, J. M. (2009). Multifunctional annexins. Plant Science, 177(6), 532-539. [CrossRef]
- Miedema, H., Bothwell, J. H., Brownlee, C., & Davies, J. M. (2001). Calcium uptake by plant cells–channels and pumps acting in concert. Trends in Plant Science, 6(11), 514-519. [CrossRef]
- White, P. J. (2000). Calcium channels in higher plants. Biochimica et Biophysica Acta (BBA)-Biomembranes, 1465(1-2), 171-189. [CrossRef]
- White, P. J., Bowen, H. C., Demidchik, V., Nichols, C., & Davies, J. M. (2002). Genes for calcium-permeable channels in the plasma membrane of plant root cells. Biochimica et Biophysica Acta (BBA)-Biomembranes, 1564(2), 299-309. [CrossRef]
- White, P. J. (2009). Depolarization-activated calcium channels shape the calcium signatures induced by low-temperature stress. New Phytologist, 6-8. http://www.jstor.org/stable/40302000.
- Miedema, H., Demidchik, V., Véry, A. A., Bothwell, J. H., Brownlee, C., & Davies, J. M. (2008). Two voltage-dependent calcium channels co-exist in the apical plasma membrane of Arabidopsis thaliana root hairs. New Phytologist, 179(2), 378-385. [CrossRef]
- Amtmann, A., & Blatt, M. R. (2009). Regulation of macronutrient transport. New Phytologist, 181(1). 10.1111/j.1469-8137.2008.02666.x.
- Kim, M. D., Kim, Y. H., Kwon, S. Y., Yun, D. J., Kwak, S. S., & Lee, H. S. (2010). Enhanced tolerance to methyl viologen-induced oxidative stress and high temperature in transgenic potato plants overexpressing the CuZnSOD, APX and NDPK2 genes. Physiologia Plantarum, 140(2), 153-162. [CrossRef]
- Moran, N. (2007). Osmoregulation of leaf motor cells. FEBS letters, 581(12), 2337-2347. [CrossRef]
- Harper, J. F. (2001). Dissecting calcium oscillators in plant cells. Trends in Plant Science, 6(9), 395-397. [CrossRef]
- Klüsener, B., Boheim, G., Liss, H., Engelberth, J., & Weiler, E. W. (1995). Gadolinium-sensitive, voltage-dependent calcium release channels in the endoplasmic reticulum of a higher plant mechanoreceptor organ. The EMBO Journal, 14(12), 2708-2714. [CrossRef]
- Hirschi, K. D. (2001). Vacuolar H+/Ca2+ transport: who's directing the traffic?. Trends in plant science, 6(3), 100-104. [CrossRef]
- Wu, Z., Liang, F., Hong, B., Young, J. C., Sussman, M. R., Harper, J. F., & Sze, H. (2002). An endoplasmic reticulum-bound Ca2+/Mn2+ pump, ECA1, supports plant growth and confers tolerance to Mn2+ stress. Plant Physiology, 130(1), 128-137. [CrossRef]
- Blatt, M. R. (2000). Cellular signaling and volume control in stomatal movements in plants. Annual Review of Cell and Developmental Biology, 16(1), 221-241. [CrossRef]
- Ritchie, S. M., Swanson, S. J., & Gilroy, S. (2002). From common signalling components to cell-specific responses: insights from the cereal aleurone. Physiologia Plantarum, 115(3), 342-351. [CrossRef]
- Evans, D. E., & Williams, L. E. (1998). P-type calcium ATPases in higher plants–biochemical, molecular and functional properties. Biochimica et Biophysica Acta (BBA)-Reviews on Biomembranes, 1376(1), 1-25. [CrossRef]
- Huang, L., Berkelman, T., Franklin, A. E., & Hoffman, N. E. (1993). Characterization of a gene encoding a Ca (2+)-ATPase-like protein in the plastid envelope. Proceedings of the National Academy of Sciences, 90(21), 10066-10070. [CrossRef]
- Harper, J. F., Hong, B., Hwang, I., Guo, H. Q., Stoddard, R., Huang, J. F., ... & Sze, H. (1998). A novel calmodulin-regulated Ca2+-ATPase (ACA2) from Arabidopsis with an N-terminal autoinhibitory domain. Journal of Biological Chemistry, 273(2), 1099-1106. [CrossRef]
- Bonza, M. C., Morandini, P., Luoni, L., Geisler, M., Palmgren, M. G., & De Michelis, M. I. (2000). At-ACA8 encodes a plasma membrane-localized calcium-ATPase of Arabidopsis with a calmodulin-binding domain at the N terminus. Plant Physiology, 123(4), 1495-1506. [CrossRef]
- George, L., Romanowsky, S. M., Harper, J. F., & Sharrock, R. A. (2008). The ACA10 Ca2+-ATPase regulates adult vegetative development and inflorescence architecture in Arabidopsis. Plant Physiology, 146(2), 716. [CrossRef]
- Schiøtt, M., Romanowsky, S. M., Bækgaard, L., Jakobsen, M. K., Palmgren, M. G., & Harper, J. F. (2004). A plant plasma membrane Ca2+ pump is required for normal pollen tube growth and fertilization. Proceedings of the National Academy of Sciences, 101(25), 9502-9507. [CrossRef]
- Geisler, M., Frangne, N., Gomes, E., Martinoia, E., & Palmgren, M. G. (2000). The ACA4 gene of Arabidopsis encodes a vacuolar membrane calcium pump that improves salt tolerance in yeast. Plant Physiology, 124(4), 1814-1827. [CrossRef]
- Li, L., Kim, B. G., Cheong, Y. H., Pandey, G. K., & Luan, S. (2006). A Ca2+ signaling pathway regulates a K+ channel for low-K response in Arabidopsis. Proceedings of the National Academy of Sciences, 103(33), 12625-12630. [CrossRef]
- Liang, F., Cunningham, K. W., Harper, J. F., & Sze, H. (1997). ECA1 complements yeast mutants defective in Ca2+ pumps and encodes an endoplasmic reticulum-type Ca2+-ATPase in Arabidopsis thaliana. Proceedings of the National Academy of Sciences, 94(16), 8579-8584. [CrossRef]
- Mills, R. F., Doherty, M. L., López-Marqués, R. L., Weimar, T., Dupree, P., Palmgren, M. G., ... & Williams, L. E. (2008). ECA3, a Golgi-localized P2A-type ATPase, plays a crucial role in manganese nutrition in Arabidopsis. Plant Physiology, 146(1), 116-128. [CrossRef]
- Li, X., Chanroj, S., Wu, Z., Romanowsky, S. M., Harper, J. F., & Sze, H. (2008). A distinct endosomal Ca2+/Mn2+ pump affects root growth through the secretory process. Plant Physiology, 147(4), 1675-1689. [CrossRef]
- Cheng, N. H., Pittman, J. K., Shigaki, T., Lachmansingh, J., LeClere, S., Lahner, B., ... & Hirschi, K. D. (2005). Functional association of Arabidopsis CAX1 and CAX3 is required for normal growth and ion homeostasis. Plant Physiology, 138(4), 2048-2060. [CrossRef]
- Hirschi, K. D., Korenkov, V. D., Wilganowski, N. L., & Wagner, G. J. (2000). Expression of Arabidopsis CAX2 in tobacco. Altered metal accumulation and increased manganese tolerance. Plant Physiology, 124(1), 125-134. [CrossRef]
- Luo, G. Z., Wang, H. W., Huang, J., Tian, A. G., Wang, Y. J., Zhang, J. S., & Chen, S. Y. (2005). A putative plasma membrane cation/proton antiporter from soybean confers salt tolerance in Arabidopsis. Plant Molecular Biology, 59, 809-820. [CrossRef]
- Hepler, P. K., & Winship, L. J. (2010). Calcium at the cell wall-cytoplast interface. Journal of Integrative Plant Biology, 52(2), 147-160. [CrossRef]
- De Kreij, C. (1996). Interactive effects of air humidity, calcium and phosphate on blossom-end rot, leaf deformation, production and nutrient contents of tomato. Journal of plant nutrition, 19(2), 361-377. [CrossRef]
- Clarkson, D. T. (1993). Roots and the delivery of solutes to the xylem. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 341(1295), 5-17. [CrossRef]
- Moore, C. A., Bowen, H. C., Scrase-Field, S., Knight, M. R., & White, P. J. (2002). The deposition of suberin lamellae determines the magnitude of cytosolic Ca2+ elevations in root endodermal cells subjected to cooling. The Plant Journal, 30(4), 457-465. [CrossRef]
- Park, S., & Waterland, N. L. (2021). Evaluation of calcium application methods on delaying plant wilting under water deficit in bedding plants. Agronomy, 11(7), 1383. [CrossRef]
- Santos, E., Montanha, G. S., Agostinho, L. F., Polezi, S., Marques, J. P. R., & de Carvalho, H. W. P. (2023). Foliar calcium absorption by tomato plants: Comparing the effects of calcium sources and adjuvant usage. Plants, 12(14), 2587. [CrossRef]
- Haleema, B., Shah, S. T., Basit, A., Hikal, W. M., Arif, M., Khan, W., ... & Fhatuwani, M. (2024). Comparative Effects of Calcium, Boron, and Zinc Inhibiting Physiological Disorders, Improving Yield and Quality of Solanum lycopersicum. Biology, 13(10), 766. [CrossRef]
- Li, Y., Ma, J., Gao, X., Tie, J., Wu, Y., Tang, Z., ... & Yu, J. (2022). Exogenous brassinosteroids alleviate calcium deficiency-induced tip-burn by maintaining cell wall structural stability and higher photosynthesis in mini Chinese Cabbage. Frontiers in Plant Science, 13, 999051. [CrossRef]
- Yang, J., Ma, J., Zhang, W., Gao, X., Wang, X., Chen, W., ... & Hu, L. (2025). Quality Response of Two Mini Chinese Cabbage Cultivars to Different Calcium Levels. Foods, 14(5), 872. [CrossRef]
- Geraldson, C.M., 1952. Studies on control of blackheart of celery. In Proceedings of the Florida State Horticultural Society, 65, 171-172.
- Vergara, C., Araujo, K. E. C., Santos, A. P., de Oliveira, F. F., de Souza Silva, G., de Oliveira Miranda, N., ... & de Medeiros, J. F. (2024). Use of crushed eggshell to control tomato blossom-end rot. Research, Society and Development, 13(5), e2213545667-e2213545667. [CrossRef]
- Geraldson, C. M. (1954). The control of blackheart of celery. In Proceedings of the American Society for Horticultural Science, 353-58.
- Kleemann, M. (2000). Effects of salinity, nutrients and spraying with cacl2 solution on the development of calcium deficiency in chervil (Anthriscus cerefolium (L.) Hoffm.) and curled parsley (Petroselinum crispum (Mill.) nym. Convar. Crispum). In Integrated View of Fruit and Vegetable Quality (pp. 41-53). CRC Press.
- Schmitz-Eiberger, M., Haefs, R., & Noga, G. (2002). Calcium deficiency-influence on the antioxidative defense system in tomato plants. Journal of Plant Physiology, 159(7), 733-742. [CrossRef]
- Parađiković, N., Lončarić, Z., Bertić, B., & Vukadinović, V. (2004). Influence of Ca-foliar application on yield and quality of sweet pepper in glasshouse conditions. Poljoprivreda, 10(2), 24-27. https://hrcak.srce.hr/22919.
- Raven, J. A. (1977). H+ and Ca2+ in phloem and symplast: relation of relative immobility of the ions to the cytoplasmic nature of the transport paths. New Phytologist, 79(3), 465-480. [CrossRef]
- Kohl, W. (1966). Die calciumverteilung in apfeln und ihre veranderung wahrend des wachstums. Die Gartenbauwissenschaft, 31: 513-47.
- Perring, M. A., & Jackson, C. H. (1975). The mineral composition of apples. Calcium concentration and bitter pit in relation to mean mass per apple. Journal of the Science of Food and Agriculture, 26(10), 1493-1502. [CrossRef]
- Bangerth, F. (1979). Calcium-related physiological disorders of plants. Annual Review of Phytopathology, 17(1), 97-122.
- Chamel, A. R. (1989). Permeability characteristics of isolated 'Golden Delicious' apple fruit cuticles with regard to calcium. Journal of the American Society for Horticultural Science, 114 (5): 804-809.
- Glenn, G. M., & Poovaiah, B. W. (1985). Cuticular permeability to calcium compounds in ‘Golden Delicious’ apple fruit. Journal of the American Society for Horticultural Science, 110(2), 192-195.
- Schlegel, T. K., & Schönherr, J. (2002). Penetration of calcium chloride into apple fruits as affected by stage of fruit development. Acta Horticulturae, 594: 527-533.
- Wojcik, P. (2001). Effect of calcium chloride sprays at different water volumes on “Szampion” apple calcium concentration. Journal of Plant Nutrition, 24(4-5), 639-650.
- Sarijan, A., Limbongan, A. A., Ekowati, N. Y., & Kusumah, R. (2025, March). Calcium nitrate application on tomatoes to increase blossom-end rot disease resistance. In IOP Conference Series: Earth and Environmental Science (Vol. 1471, No. 1, p. 012025). IOP Publishing. [CrossRef]
- Assaha, D. V., & Petang, L. Y. (2024). Bone meal enhances the growth and yield of the tomato cultivar Cobra F1 by increasing fruit Ca content and alleviating blossom-end rot. Asian Journal of Agriculture, 8(1). [CrossRef]
- Madani, B., Mirshekari, A., Sofo, A., & Tengku Muda Mohamed, M. (2016). Preharvest calcium applications improve postharvest quality of papaya fruits (Carica papaya L. cv. Eksotika II). Journal of Plant Nutrition, 39(10), 1483-1492. [CrossRef]
- Parsa, Z., Roozbehi, S., Hosseinifarahi, M., Radi, M., & Amiri, S. (2021). Integration of pomegranate peel extract (PPE) with calcium sulphate (CaSO4): A friendly treatment for extending shelf-life and maintaining postharvest quality of sweet cherry fruit. Journal of Food Processing and Preservation, 45(1), e15089. [CrossRef]
- Zhai, J., Gao, Y., Zhang, X. W., Han, L. J., Bi, H. A., Li, Q. M., & Ai, X. (2019). Effects of silicon and calcium on photosynthesis, yield and quality of cucumber in solar-greenhouse. Acta Hortic. Sin., 46: 701–713. 10.16420/j.issn.0513-353x.2018-0687.
- Ahn, J., Park, M., Kim, J., Huq, E., Kim, J., & Kim, D. H. (2024). Physiological and transcriptomic analyses of healthy and blossom-end-rot (BER)-defected fruit of chili pepper (Capsicum annuum. L.). Horticulture, Environment, and Biotechnology, 1-10. [CrossRef]
- Ferguson, I. B., & Watkins, C. B. (1989). Bitter pit in apple fruit. Horticultural Reviews, 11, 289-355. [CrossRef]
- Ferguson, I. B., & Watkins, C. B. (1983). Cation distribution and balance in apple fruit in relation to calcium treatments for bitter pit. Scientia Horticulturae, 19(3-4), 301-310. [CrossRef]
- Huang, Z., Takemoto, T., Omwange, K. A., Orsino, M., Konagaya, K., & Kondo, N. (2025). Early detection of Blossom-End Rot in green peppers using fluorescence and normal color images in visible region. Food Control, 172, 111156. [CrossRef]
- Karley, A. J., & White, P. J. (2009). Moving cationic minerals to edible tissues: potassium, magnesium, calcium. Current opinion in plant biology, 12(3), 291-298. [CrossRef]
- De Freitas, S. T., Shackel, K. A., & Mitcham, E. J. (2011b). Abscisic acid triggers whole-plant and fruit-specific mechanisms to increase fruit calcium uptake and prevent blossom end rot development in tomato fruit. Journal of Experimental Botany, 62(8), 2645-2656. [CrossRef]
- Ho, L.C., 1989. Environmental effects on the diurnal accumulation of 45Ca by young fruit and leaves of tomato plants. Annals of Botany, 63(2), 281-288. [CrossRef]
- Ho, L. C., Belda, R., Brown, M., Andrews, J., & Adams, P. (1993). Uptake and transport of calcium and the possible causes of blossom-end rot in tomato. Journal of Experimental Botany, 44(2), 509-518. [CrossRef]
- Tadesse, T., Nichols, M.A., Hewett, E.W., Fisher, K.J., 2001. Relative humidity around the fruit influences the mineral composition and incidence of blossom-end rot in sweet pepper fruit. The Journal of Horticultural Science and Biotechnology, 76(1), 9-16. [CrossRef]
- Fischer, A. M. (2007). Nutrient remobilization during leaf senescence. Annual Reviews of Senescence Processes in Plants, 26, 87-107.
- Marschner, P. (2011). Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic press: Cambridge, MA, USA; ISBN 9780123849052.
- Bar-Tal, A., Aloni, B., Karni, L., Oserovitz, J., Hazan, A., Itach, M., ... & Rosenberg, R. (2001). Nitrogen nutrition of greenhouse pepper. I. Effects of nitrogen concentration and NO3: NH4 ratio on yield, fruit shape, and the incidence of blossom-end rot in relation to plant mineral composition. HortScience, 36(7), 1244-1251.
- Schönherr, J., & Bukovac, M. J. (1973). Ion exchange properties of isolated tomato fruit cuticular membrane: exchange capacity, nature of fixed charges and cation selectivity. Planta, 109, 73-93. [CrossRef]
- Yermiyahu, U., Nir, S., Ben-Hayyim, G., & Kafkafi, U. (1994). Quantitative competition of calcium with sodium or magnesium for sorption sites on plasma membrane vesicles of melon (Cucumis melo L.) root cells. The Journal of Membrane Biology, 138, 55-63. [CrossRef]
- De Freitas, S. T., do Amarante, C. V., Labavitch, J. M., & Mitcham, E. J. (2010). Cellular approach to understand bitter pit development in apple fruit. Postharvest Biology and Technology, 57(1), 6-13. [CrossRef]
- Dris, R., Niskanen, R., & Fallahi, E. (1998). Nitrogen and calcium nutrition and fruit quality of commercial apple cultivars grown in Finland. Journal of Plant Nutrition, 21(11), 2389-2402. [CrossRef]
- Lanauskas, J., & Kvikliene, N. (2006). Effect of calcium foliar application on some fruit quality characteristics of ‘Sinap Orlovskij’apple. Agronomy Research, 4(1), 31-36.
- Gilliham, M., Dayod, M., Hocking, B. J., Xu, B., Conn, S. J., Kaiser, B. N., ... & Tyerman, S. D. (2011). Calcium delivery and storage in plant leaves: exploring the link with water flow. Journal of Experimental Botany, 62(7), 2233-2250. [CrossRef]
- Park, S., Cheng, N. H., Pittman, J. K., Yoo, K. S., Park, J., Smith, R. H., & Hirschi, K. D. (2005). Increased calcium levels and prolonged shelf life in tomatoes expressing Arabidopsis H+/Ca2+ transporters. Plant Physiology, 139(3), 1194-1206.
- De Freitas, S. T., & Mitcham, E. I. (2012). 3 factors involved in fruit calcium deficiency disorders. Horticultural Reviews, 40(1), 107-146. [CrossRef]
- Picchioni, G. A., Watada, A. E., Conway, W. S., Whitaker, B. D., & Sams, C. E. (1998). Postharvest calcium infiltration delays membrane lipid catabolism in apple fruit. Journal of Agricultural and Food Chemistry, 46(7), 2452-2457. [CrossRef]
- Lund, Z. F. (1970). The effect of calcium and its relation to several cations in soybean root growth. Soil Science Society of America Journal, 34(3), 456-459. [CrossRef]
- Caffall, K. H., & Mohnen, D. (2009). The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydrate Research, 344(14), 1879-1900. [CrossRef]
- Bush, D. S. (1995). Calcium regulation in plant cells and its role in signaling. Annual Review of Plant Biology, 46(1), 95-122. [CrossRef]
- Kendall, J. M., Dormer, R. L., & Campbell, A. K. (1992). Targeting aequorin to the endoplasmic reticulum of living cells. Biochemical and Biophysical Research Communications, 189(2), 1008-1016. [CrossRef]
- Johnson, C. H., Knight, M. R., Kondo, T., Masson, P., Sedbrook, J., Haley, A., & Trewavas, A. (1995). Circadian oscillations of cytosolic and chloroplastic free calcium in plants. Science, 269(5232), 1863-1865. [CrossRef]
- Logan, D. C., & Knight, M. R. (2003). Mitochondrial and cytosolic calcium dynamics are differentially regulated in plants. Plant Physiology, 133(1), 21-24. [CrossRef]
- Pauly, N., Knight, M. R., Thuleau, P., Graziana, A., Muto, S., Ranjeva, R., & Mazars, C. (2001). The nucleus together with the cytosol generates patterns of specific cellular calcium signatures in tobacco suspension culture cells. Cell Calcium, 30(6), 413-421. [CrossRef]
- Voyle, G. (2021). Blossom end rot causes and cures in garden vegetables. Michigan State University Extension, MI, USA.
- de Freitas, S. T., Amarante, C. D., & Mitcham, E. J. (2016). Calcium deficiency disorders in plants. In: Postharvest Ripening Physiology of Crops, 477-502. CRC Press: Boca Raton, FL, USA. ISBN 9781498703819.
- Hocking, B., Tyerman, S. D., Burton, R. A., & Gilliham, M. (2016). Fruit calcium: transport and physiology. Frontiers in plant science, 7, 569. [CrossRef]
- Ren, H., Zhao, X., Li, W., Hussain, J., Qi, G., & Liu, S. (2021). Calcium signaling in plant programmed cell death. Cells, 10(5), 1089. [CrossRef]
- Ojha, R. K., & Jha, S. K. (2021). Role of mineral nutrition in management of plant diseases. Farmers’ Prosperity through Improved Agricultural Technologies, 241-261.
- Kleemann, M. (1999). Development of calcium deficiency symptoms in chervil (Antriscus cerefolium (L.) Hoffm.) and curled parsley (Petroselinum crispum (Mill.) Nym. convar crispum). Zeszyty Problemowe Postępów Nauk Rolniczych, (468).
- Aloni, B., 1986. Enhancement of leaf tip burn by restricting root growth in Chinese cabbage plants. Journal of Horticultural Science, 61(4), 509-513. [CrossRef]
- Collier, G.F., Tibbitts, T.W., 1982. Tipburn of lettuce. Horticultural Reviews, 4, 49-65. [CrossRef]
- Cox, E.F., McKee, J.M.T., Dearman, A.S., 1976. The effect of growth rate on tipburn occurrence in lettuce. Journal of Horticultural Science, 51(3), 297-309. [CrossRef]
- Bible, B.B., Stiehl, B., 1986. Effect of atmospheric modification on the incidence of blackheart and the cation content of celery. Scientia Horticulturae, 28(1-2), 19-28. [CrossRef]
- Adams, P., & Holder, R. (1992). Effects of humidity, Ca and salinity on the accumulation of dry matter and Ca by the leaves and fruit of tomato (Lycopersicon esculentum). Journal of Horticultural Science, 67(1), 137-142. [CrossRef]
- De Kreij, C., Janse, J., Van Goor, B. J., & Van Doesburg, J. D. J. (1992). The incidence of calcium oxalate crystals in fruit walls of tomato (Lycopersicon esculentum Mill.) as affected by humidity, phosphate and calcium supply. Journal of Horticultural Science, 67(1), 45-50. [CrossRef]
- Mengel, K., & Kirkby, E. A. (1987). Principles of Plant Nutrition. (No. Ed. 4, p. 687pp), , ISBNs: 978-1-4020-0008-9, 978-9-40-101009-2 . [CrossRef]
- Kabir, M. Y. (2019). Abiotic Factors Affecting Plant Physiology and Fruit Yield and Physiological Disorders in Bell Pepper and Tomato (Doctoral dissertation, University of Georgia). 219 p.
- Horst, W. J. (1987). Aluminium tolerance and calcium efficiency of cowpea genotypes. Journal of plant nutrition, 10(9-16), 1121-1129.
- Ho, L. C., Hand, D. J., & Fussell, M. (1997, May). Improvement of tomato fruit quality by calcium nutrition. In International Symposium on Growing Media and Hydroponics 481 (pp. 463-468). 10.17660/ActaHortic.1999.481.53.
- Ho, L. C., Adams, P., Li, X. Z., Shen, H., Andrews, J., & Xu, Z. H. (1995). Responses of Ca-efficient and Ca-inefficient tomato cultivars to salinity in plant growth, calcium accumulation and blossom-end rot. Journal of Horticultural Science, 70(6), 909-918. [CrossRef]
- Adams, P., & Ho, L. C. (1992). The susceptibility of modern tomato cultivars to blossom-end rot in relation to salinity. Journal of Horticultural Science, 67(6), 827-839. [CrossRef]
- Cardoso, M. O., Queiróz, M. D., & Souza, R. F. (1995). Incidence of blossom-end rot in five tomato cultivars grown in soil with three calcium levels. Horticultura Brasileira, 13 (2): 172-175.
- Sperry, W. J., Davis, J. M., & Sanders, D. C. (1996). Soil moisture and cultivar influence cracking, blossom-end rot, zippers, and yield of staked fresh-market tomatoes. HortTechnology, 6(1), 21-24.
- Maynard, D. N., Barham, W. S., & McCombs, C. L. (1957). The effect of calcium nutrition of tomatoes as related to the incidence and severity of blossom-end rot. In Proc. Amer. Soc. Hort. Sci, 318-22.
- Magán, J. J., Gallardo, M., Thompson, R. B., & Lorenzo, P. (2008). Effects of salinity on fruit yield and quality of tomato grown in soil-less culture in greenhouses in Mediterranean climatic conditions. Agricultural Water Management, 95(9), 1041-1055. [CrossRef]
- Srivastava, S., Singh, L., Debnath, B., Shukla, M., Kumar, S., Rana, M., & Meshram, S. (2022). Blossom end-rot (BER): an abiotic devastating disorder of tomato. Climate Change and Environmental Sustainability, 10(1), 15-22. [CrossRef]
- Kumar, V., Darvhankar, M., & Srivastava, S. (2019). Evaluation of Trichoderma species and fungicides on nutritional quality and yield of tomato. Pl. Cell Biotechnol. Mol. Biol, 20, 331-338.
- Das, K. A., Bhosale, R. K., Chavan, S. S., Rana, M., & Srivastava, S. (2023). Effect of physiological factors and different growth media on Alternaria solani under in vitro condition and eco-friendly management of early blight of tomato (Solanum lycopersicum L.). Ann Biol 39(1):66-72.
- Zhou, L., Fu, Y., & Yang, Z. (2009). A genome-wide functional characterization of Arabidopsis regulatory calcium sensors in pollen tubes. Journal of Integrative Plant Biology, 51(8), 751-761. [CrossRef]
- Yoshida, Y., Irie, N., Vinh, T. D., Ooyama, M., Tanaka, Y., Yasuba, K. I., & Goto, T. (2014). Incidence of blossom-end rot in relation to the water-soluble calcium concentration in tomato fruits as affected by calcium nutrition and cropping season. Journal of the Japanese Society for Horticultural Science, 83(4), 282-289. [CrossRef]
- Barickman, T. C., Kopsell, D. A., & Sams, C. E. (2014). Abscisic acid increases carotenoid and chlorophyll concentrations in leaves and fruit of two tomato genotypes. Journal of the American Society for Horticultural Science, 139(3), 261-266. [CrossRef]
- De Freitas, S. T., Martinelli, F., Feng, B., Reitz, N. F., & Mitcham, E. J. (2018). Transcriptome approach to understand the potential mechanisms inhibiting or triggering blossom-end rot development in tomato fruit in response to plant growth regulators. Journal of Plant Growth Regulation, 37, 183-198. [CrossRef]
- R'him, T., & Jebari, H. (2008). Blossom-end rot in relation to morphological parameters and calcium content in fruits of four pepper varieties (Capsicum annuum L.). Biotechnologie, Agronomie, Société et Environnement,12 (4): 361-366.
- Grasselly, D., Rosso, L., Holgard, S., Cottet, V., Navez, B., Jost, M., & Berier, A. (2008). Soilless culture of tomato: effect of salinity of the nutrient solution. Infos-Ctifl, 239: 41-45.
- Robbins, W. R. (1937). Relation of nutrient salt concentration to growth of the tomato and to the incidence of blossom-end rot of the fruit. Plant Physiology, 12(1), 21. [CrossRef]
- Dekock, P. C., Hall, A., Inkson, R. H., & Alan Robertson, R. (1979). Blossom-end rot in tomatoes. Journal of the Science of Food and Agriculture, 30(5), 508-514. [CrossRef]
- Adams, P., & Ho, L. C. (1993). Effects of environment on the uptake and distribution of calcium in tomato and on the incidence of blossom-end rot. Plant and soil, 154, 127-132. [CrossRef]
- Pill, W. G., & Lambeth, V. N. (1981). Effects of soil water regime and nitrogen form on blossom-end rot, yield, water relations, and elemental composition of tomato. Journal of the American Society for Horticultural Science,105 (5): 730-734.
- Paiva, E. A. S., Sampaio, R. A., & Martinez, H. E. P. (1998). Composition and quality of tomato fruit cultivated in nutrient solutions containing different calcium concentrations. Journal of Plant Nutrition, 21(12), 2653-2661. [CrossRef]
- van Goor, B. J. (1974). Influence of restricted water supply on blossom-end rot and ionic composition of tomatoes grown in nutrient solution. Communications in soil science and plant analysis, 5(1), 13-24. [CrossRef]
- Franco, J. A., Perez-Saura, P. J., Ferná Ndez, J. A., Parra, M., & Garcia, A. L. (1999). Effect of two irrigation rates on yield, incidence of blossom-end rot, mineral content and free amino acid levels in tomato cultivated under drip irrigation using saline water. The Journal of Horticultural Science and Biotechnology, 74(4), 430-435. [CrossRef]
- Franco, J., Bañón, S., & Madrid, R. (1994). Effects of a protein hydrolysate applied by fertigation on the effectiveness of calcium as a corrector of blossom-end rot in tomato cultivated under saline conditions. Scientia Horticulturae, 57(4), 283-292. [CrossRef]
- Marcelis, L. F. M., & Ho, L. C. (1999). Blossom-end rot in relation to growth rate and calcium content in fruits of sweet pepper (Capsicum annuum L.). Journal of Experimental Botany, 50(332), 357-363. [CrossRef]
- Chiu, T. F., & Bould, C. (1976). Effects of shortage of calcium and other cations on 45Ca mobility, growth and nutritional disorders of tomato plants (Lycopersicon esculentum). Journal of the Science of Food and Agriculture, 27(10), 969-977. [CrossRef]
- Saure, M. C. (2014). Why calcium deficiency is not the cause of blossom-end rot in tomato and pepper fruit–a reappraisal. Scientia Horticulturae, 174, 151-154. [CrossRef]
- Gent, M. P. (2007). Effect of degree and duration of shade on quality of greenhouse tomato. HortScience, 42(3), 514-520. [CrossRef]
- Díaz-Perez, J.C., 2014. Bell pepper (Capsicum annum L.) crop as affected by shade level: fruit yield, quality, and postharvest attributes, and incidence of phytophthora blight (caused by Phytophthora capsici Leon.). HortScience 49 (7), 891–900. [CrossRef]
- Kabir, M.Y., Díaz-Pérez, J.C., Nambeesan, S.U. (2020). Effect of shade levels on plant growth, physiology, and fruit yield in bell pepper (Capsicum annuum L.). In: XI International Symposium on Protected Cultivation in Mild Winter Climates and I International Symposium on Nettings and 1268, pp. 311–318. [CrossRef]
- Kabir, M.Y., Nambeesan, S.U., Bautista, J., Díaz-Pérez, J.C. (2022). Plant water status, plant growth, and fruit yield in bell pepper (Capsicum annum L.) under shade nets. Sci. Hortic. 303, 111241 . [CrossRef]
- Kabir, M. Y., Nambeesan, S. U., & Díaz-Pérez, J. C. (2024). Shade nets improve vegetable performance. Scientia Horticulturae, 334, 113326. [CrossRef]
- Jing, T., Li, J., He, Y., Shankar, A., Saxena, A., Tiwari, A., ... & Awasthi, M. K. (2024). Role of calcium nutrition in plant Physiology: Advances in research and insights into acidic soil conditions-A comprehensive review. Plant Physiology and Biochemistry, 108602. [CrossRef]
- Topcu, Y., Nambeesan, S. U., & van der Knaap, E. (2022). Blossom-end rot: a century-old problem in tomato (Solanum lycopersicum L.) and other vegetables. Molecular Horticulture, 2(1), 1. [CrossRef]
Figure 2.
Calcium plays structural, enzymatic, and signaling functions. Activities of α-amylase are seen during seed germination; fructose 1,6 bisphosphatase (FBPase) is a regulatory enzyme in the sucrose biosynthesis pathway; polygalacturonase (PG) hydrolyzes the alpha-1,4 glycosidic bonds between galacturonic acid residues of pectin.
Figure 2.
Calcium plays structural, enzymatic, and signaling functions. Activities of α-amylase are seen during seed germination; fructose 1,6 bisphosphatase (FBPase) is a regulatory enzyme in the sucrose biosynthesis pathway; polygalacturonase (PG) hydrolyzes the alpha-1,4 glycosidic bonds between galacturonic acid residues of pectin.
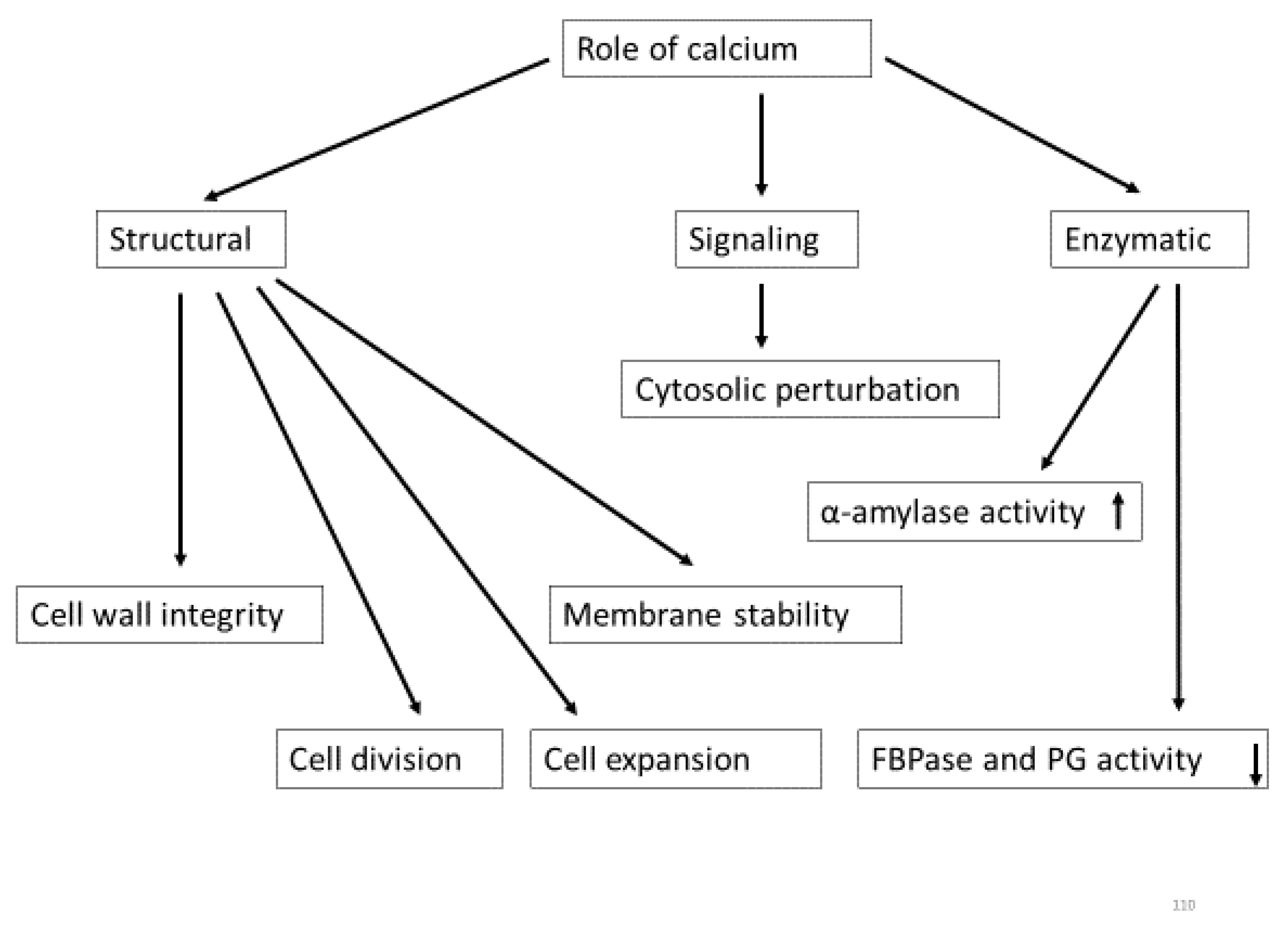
Figure 4.
Calcium availability affects the development of calcium deficiency disorder blossom-end rot (BER). At high soil calcium availability, fruit Ca2+-deficiency disorder may not appear. However, at low calcium availability, deficiency disorder appears through depletion of apoplastic calcium and subsequent membrane leakiness. However, BER may occur at high calcium availability due to improper calcium compartmentalization.
Figure 4.
Calcium availability affects the development of calcium deficiency disorder blossom-end rot (BER). At high soil calcium availability, fruit Ca2+-deficiency disorder may not appear. However, at low calcium availability, deficiency disorder appears through depletion of apoplastic calcium and subsequent membrane leakiness. However, BER may occur at high calcium availability due to improper calcium compartmentalization.
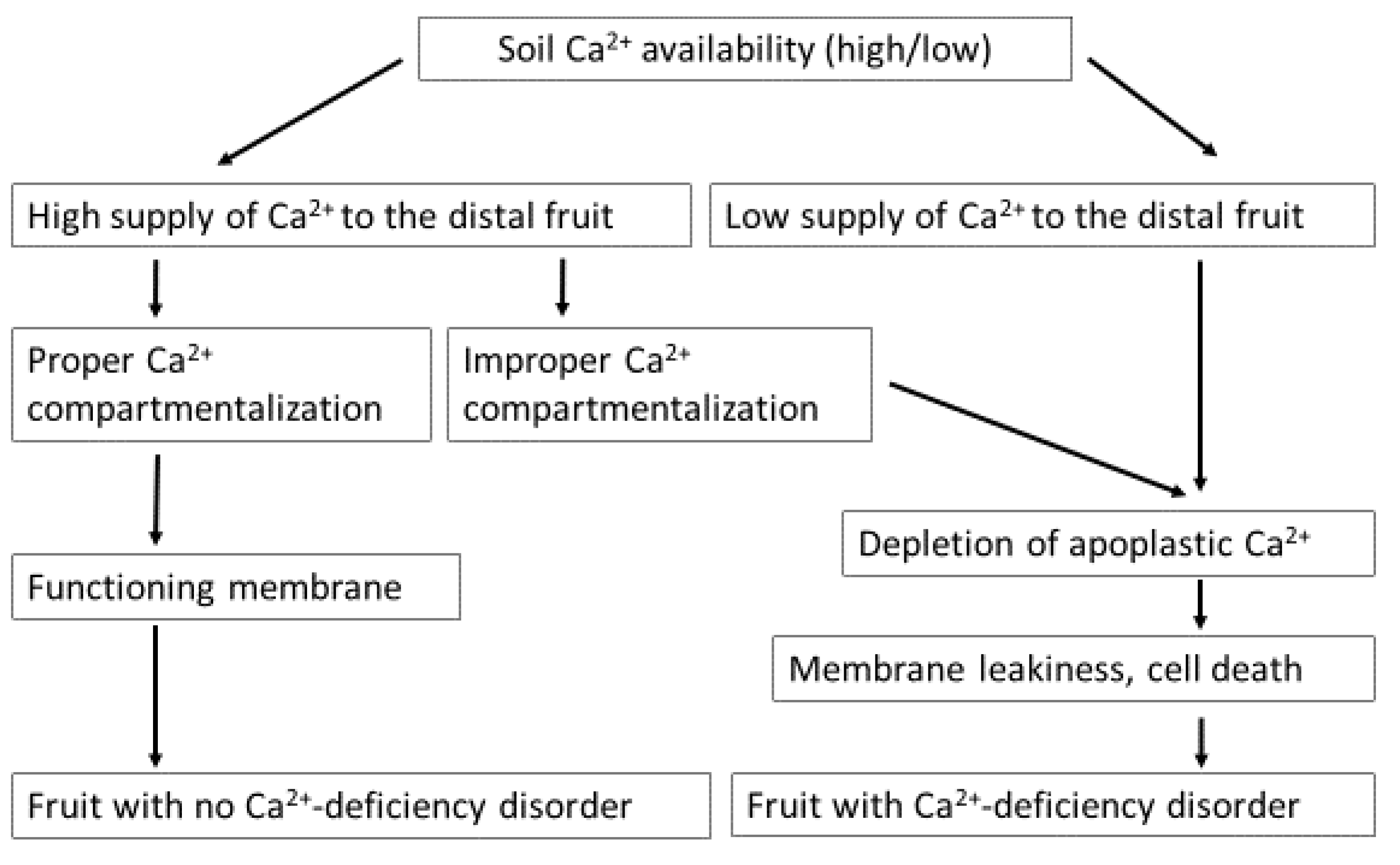
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



