Submitted:
30 May 2025
Posted:
02 June 2025
You are already at the latest version
Abstract
Historic mural paintings are highly susceptible to biodeterioration due to microbial colonization. This study reports, for the first time, the presence of Paracoccus marcusii on a mural painting and investigates the potential role of strain IBR3—isolated from a deteriorated 19th-century mural in a private Italian villa—in the degradation of wall surfaces. The strain exhibited significant enzymatic activity, including casein hydrolysis and calcium carbonate dissolution, revealing its involvement in the degradation of both the organic binders and mineral components commonly present in mural paintings. The study also evaluates the antimicrobial efficacy of Thymus serpyllum essential oil against IBR3, through in vitro tests, membrane integrity assays and ATR-FTIR spectroscopy. These findings underscore the potential of Thymus serpyllum essential oil as a sustainable, non-toxic biocide for cultural heritage conservation, and reveal a novel microbial agent in mural biodeterioration.
Keywords:
Thyme essential oil
; mural paintings
; bacteria
; biodeterioration
; Paracoccus
; cultural heritage
; antimicrobial activity
Introduction
Noble residences, especially those built between the 18th and 19th centuries, are not only symbols of their owners’ social prestige but also important cultural and artistic testimonies. One of the main distinctive characteristics of historical villas is the presence of wall paintings adorning ceilings, rooms and staircases. These artworks often illustrate mythological scenes, ornamental motifs, scenes of everyday life, landscapes, or architectural illusions painted in the trompe-l’œil style. Their preservation represents a key aspect of heritage conservation, as they are integrated into the architecture itself and often provide insight into the lives, tastes, values, and cultural environment of their time [1,2]. Both the aesthetic integrity and historical value of these artworks are threatened over time by abiotic and biotic agents. Fluctuations in humidity and temperature, light exposure and air pollution, and colonization by fungi, algae, bacteria, and insects can lead to irreversible damages that involve fading, cracking, peeling, and discoloration of the mural surface [2,3]. The complexity of the process of biodeterioration requires a deep knowledge of microbial community dynamics, functional diversity, and environmental context. Although microorganisms can colonize mural surfaces independently, their growth is enhanced through microbial interactions. The establishment of a self-sustaining microbial community, where the presence of one group supports the growth and activity of others, accelerates the overall deterioration of the artwork. In the dynamic micro-ecosystem established on the surface of the mural, each member plays a specific role. For instance, cyanobacteria provide organic compounds for heterotrophic bacteria and fungi [4]. Meanwhile, fungi release nutrients that can further support microbial growth. Additionally, the hyphal structure of fungi can physically alter the surface, increasing porosity and creating micro-niches where other microbes can settle and thrive. However, these same processes contribute significantly to the biodeterioration of mural paintings. Some bacterial genera, including Bacillus, Acinetobacter, Rhodococcus, Stenotrophomonas, are well known to contribute to the degradation of painting pigment’s binder, due to their widespread enzymatic activities, or to the reduction of iron of some pigments from the paintings [5,6]. Particularly in subterranean contexts, including tombs, caves, and rock-cut structures, Streptomyces spp. are frequently identified as dominant colonizers, producing a diverse range of metabolites—biopigments, organic and inorganic acids, hydrogen sulphide and enzymes- which cause irreversible damage to paintings [7,8]. The increasing awareness of microbial impact on mural painting underscores the urgent need for sustainable preservation strategies. Eco-friendly approaches, which do not compromise the integrity of cultural heritage materials and do not pose toxicity risks to humans, animals, and the environment, are particularly relevant for the conservation of painted surfaces exposed to microbial biofilm formation and biochemical alteration. Thus, the use of plant extracts has been gaining increasing attention for their ability to inhibit the growth of several fungal and bacterial strains associated with environments such as food, plants, and cultural heritage materials [9,10,11,12,13,14,15]. Extracts of Rosmarinus officinalis exhibited antimicrobial activity against several fungal strains such as Aspergillus clavatus, and bacterial strains including Arthrobacter globiformis and Bacillus cereus at low concentrations [16]. Similarly, Corbu et al. [17] reported the efficacy of Allium sativum extracts against more than 200 biodeteriogenic microbial isolates. Several studies report the inhibitory effect of essential oils against a range of fungi and bacteria [12,13,14,18,19,20,21]. These oils are complex mixtures of bioactive compounds, such as terpenes, phenolics, and aldehydes, that can disrupt microbial cell membranes, interfere with enzymatic activity, and inhibit biofilm formation, making them particularly promising for antimicrobial applications in cultural heritage conservation. The present study aims to: 1) characterize the bacterial strain IBR3, isolated from a decolored mural painting; 2) evaluate the capacity of the essential oil of Thymus serpyllum to inhibit its growth 2) review the importance of Paracoccus species on biodeterioration.
2. Results
2.1. Identification and Characterization of the IBR3 Bacterial Strain
IBR3 colonies, cultured on a TSA plate at 28 °C for 2 d, appeared flat, bright orange and round shaped. Cells were sometimes arranged in clusters and pairs, Gram stain-negative, with no flagella, and non-motile. Phylogenetic analysis, based on 1141 unambiguous bases, revealed that strain IBR3 was a member of the Paracoccus genus. It clustered together with P. marcusii, P. haeundaensis, and P. carotinifaciens (Figure 1).
IBR3 grew on several carbon sources (sucrose, fructose, d-sorbitol, d-glucose, l-arabinose, d-mannose, d-mannitol), producing organic acids from d-glucose. The bacteria hydrolyzed gelatin but no starch, while tests for indole production and urease activity were negative (Table 1). Paracoccus IBR3 also showed proteolytic activity towards casein and calcium carbonate dissolving activity.
2.2. Chemical composition of Thymus serpyllum essential oil
Based on the GC-MS analysis, 10 compounds were identified in Thy-EO. Carvacrol (51.88%) and p-cymene (40.28%) were the most abundant components, followed by thymol (2.45%) (Table 2). Most compounds were present at concentrations below 1.5%
2.3. In Vitro Antibacterial Activity of Thymus serpyllum Essential Oil
The effects of different concentrations (0; 0.1;0.2;0.3;0.4;0.5;1.0 mg/mL) of Thy-EO against IBR3 are shown in Figure 2. Data from the control and tween groups were pooled, as no significant differences were observed between them (Student’s t-test; P < 0.05). All Thy-EO inhibited the bacterial growth. MIC and MCB values were 0.4 mg/mL and 1 mg/mL, respectively. The EC50 value was 0.37 mg/mL.
2.4. Role of Paracoccus IBR3 in Mural Painting Deterioration
Members of the Paracoccus genus are Gram-negative, non-spore-forming bacteria commonly found in various environments, including soil (18% of the Biosample data) and aquatic environments (36% of the Biosample data), all over the world. The genus encompasses more than 83 different species. However, only the species P. cavernae, P. onubensis, and P. tibetensis have been associated with mural painting samples [26,27,28,29].
2.5. Cell Membrane Integrity
Treatments with Thy-EO induced the disruption of the bacterial membrane’s integrity. In bacteria exposed to increasing concentrations of Thy-EO (MIC, 2×MIC, and 4×MIC) the release of both dsDNA and protein content increased in a dose dependent manner (Figure 3).
2.6. Crystal Violet Uptake Assay
A crystal violet uptake assay was used to determine the membrane permeability. The uptake of crystal violet by IBR3 was 22% in the absence of Thy-EO which increased by 26% at 0.4 mg/mL (MIC) of Thy-EO treatments.
2.7. ATR-FTIR Spectroscopy
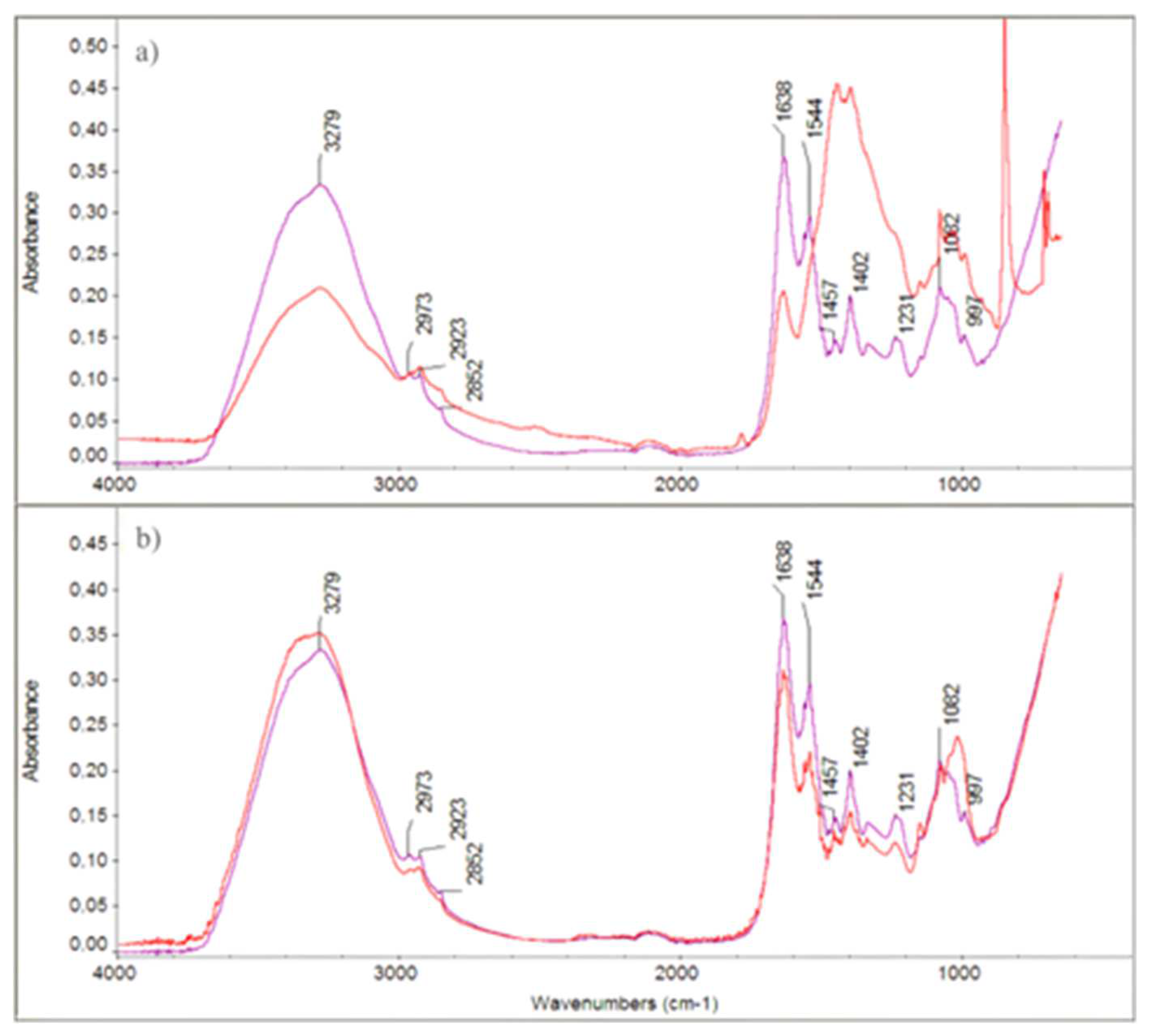
The FTIR spectra of both treated and untreated IBR3 colonies showed notable differences in the OH stretching region (~3279 cm⁻¹), CH stretching vibrations (2973, 2923, 2852 cm⁻¹), and amide I and II bands (1638 and 1544 cm⁻¹), which suggested alterations in membrane proteins and lipids upon treatment. Additionally, changes in peaks at ~1402, 1231, 1082, and 897 cm⁻¹ indicate modifications in polysaccharide and phospholipid structures.
Figure 4.
FTIR spectra of IBR3 strain treated with 0.3 mg/mL (a), 0.4 mg/mL (b) of Thy-EO (red lines), compared to untreated controls (purple lines).
Figure 4.
FTIR spectra of IBR3 strain treated with 0.3 mg/mL (a), 0.4 mg/mL (b) of Thy-EO (red lines), compared to untreated controls (purple lines).
3. Discussions
In this study, a bacterial strain IBR3 with degradation capability of mural painting was isolated and identified on the basis of phenotypic, biochemical and molecular characteristics, as Paracoccus marcusii. To our knowledge, the presence of P. marcusii on a deteriorated mural painting in a private villa represents a novel finding in the context of cultural heritage microbiology. This finding confirms the role of carotenoid-producing bacteria in the degradation of mural paintings, and, therefore, their contribution to the aesthetic damage of the painting [22,30,31]. Paracoccus genus has been previously highlighted as one the most abundant genera in cultural heritage artifacts, including wall paintings of caves and grottoes, books and textiles [32,33,34,35]. However, this genus remains relatively understudied, in fact, according to the NCBI biosample database, only three taxa that have been identified at species level associated with mural paintings: Paracoccus cavernae, isolated from Ardales Cave, Spain [27]; Paracoccus onubensis, obtained in Gruta de las Maravillas, in Huelva, Spain [28]; Paracoccus tibetensis recorded in a 1,500-year-old tomb in Shanxi Province, China [26]. The presence of Paracoccus species on mural walls is not surprising due to their metabolic versatility, including the ability to oxidate sulfur compounds, biofilm to degrade various organic compounds and survive in nutrient-poor environments [29]. In this study, the occurrence of P. marcusii IBR3 suggests that, even lesser-known bacterial taxa, Paracoccus species may play an important role in the alteration of the mural painting surfaces. In fact, the biochemical activities of P. marcusii IBR3 not only facilitate microbial colonization but also actively drive the physical and chemical deterioration of mural paintings. Hydrolysis of casein and gelatin lead to the formation of acidic byproducts, which can destabilize the paint layer by weakening binding agents and thus damaging pigments and underlying plaster. Similarly, the dissolution of calcium carbonate decreases the local pH, promoting acidification that can damage pigments and the mural’s substrate and can lead to the formation of calcium carbonate precipitates, affecting visual and structural integrity. The contribution of P. marcusii to the deterioration of mural artworks, either directly through enzymatic degradation of organic binders or indirectly by altering the local microenvironment, may have been underestimated and requires the development of targeted cleaning and control strategies. Several studies have tested the antimicrobial activity of essential oils against microorganisms associated with biodeterioration in archives, libraries, and museums [36,37,38,39]. Lavin et al. [40] showed that the vapours of the essential oils of T. vulgaris reduced or stopped temporarily the growth of Scopulariopsis sp. and Fusarium spp. isolated from historical archives [40,41], just to cite a few examples. This study reports for the first time the efficacy of the T. serpillum essential oil against P. marcusii. The MIC, MCB and ED50 values (0.4;1.0;0.37 mg/mL) indicated that Thy-EO exhibited effective biological activity at relatively low concentrations, supporting its potential for further antimicrobial investigation. Thy-EO disrupted the P. marcusii IBR3 membrane, leading to the release of cellular content as confirmed based on the absorbance values measured at 260 nm and 280 nm, and the ATR-FTIR analysis. These observations confirmed data from the literature reporting the significant efficacy of T. serpyllum against bacteria and fungi [13,42,43,44,45]. The integration of essential oils into lipid-rich membranes, which is due to their constituents’ hydrophobic nature, allows them to disrupt membrane integrity and increase permeability, particularly in the cell membrane and mitochondria [46,47,48]. Xu et al. [46] showed that a dose of 200 mg/mL of carvacrol and thymol, the main compounds in thyme EOs, was enough to reduce membrane permeability. Similarly, Kauser et al. [49] reported that thymol concentration of 32 µg/mL, 63 µg/mL, and 125 µg/mL suppressed Candida albicans, C. tropicalis and C. glabrata growth, respectively, and altered their morphologies. Moreover, treatments caused leakage of intracellular macromolecules, significant reduced the ergosterol levels and inhibited plasma membrane H+-ATPase (Pma1) activity.
In summary, this study confirms that Thy-EO had potent inhibitory activity against P. marcusii IBR3 by inducing extensive membrane destruction, leading to inhibited bacterial growth. The use of the essential oil of T. serpyllum is an eco-friendly alternative to conventional chemical treatments to protect mural paintings from deterioration caused by microbial agents. Further research is needed for its long-term effects and optimal application methods to enhance the protection of mural paintings from biological degradation.
4. Material and methods
4.1. Isolation and Characterization of a Bacterial Strain
Samples were collected in August 2024 from a mural painting located in the main room of a private Villa, dated late 1800s, in Central Italy. The painting depicted a rural landscape. Three samples were collected in three small square plots (10 cm) with evident signs of deterioration, due to pink patina, by means of sterile swabs. The swabs were combined in the same tube to make a composite sample. Bacteria were isolated by using serial dilution technique and the taxa were cultured on Tryptic Soy Agar (TSA) media. Plates were incubated at 37°C for 24-48 hours in aerobic condition. Single orange colonies were selected from plates a 10⁻³ dilution labeled 10-3 and sub-cultured for 24-48 hours. Strains were grouped according to their morphologies. The isolate IBR3 was chosen as representative of the most abundant colonies and later used in the following analysis. The bacteria were stained by Gram method and observed by optical microscope under the oil-immersion lens (Nikon Eclipse Ei trinocular (NIKON, Osaka, Japan)) [50]. Motility test was carried out using the protocol described by Reller & Mirrett [51]. Bacterial growth on different carbon sources was assessed following the method of Maheswaran & Forchhammer [52]. A M9 minimal medium was used with one of the following different carbon sources, each at 0.4%: D-glucose, D-fructose, D-sucrose, D-mannose, D-mannitol, L- arabinose, and D-sorbitol. The proteolytic activity was evaluated using Milk Nutrient Agar (Milk-NA) and R2A Agar media, supplemented with 0.4% gelatin, as described by Krakovà et al. [53]. Starch hydrolysis was tested using the method of Dhawale et al. [54]. Calcium carbonate (CaCO₃) dissolution was tested on agar plates following the protocol by Cacchio et al. [55], while organic acid production was evaluated in Czapek-Dox liquid medium, as described by Jurado et al. [56]. Indole production from tryptophan was evaluated using the protocol of Doğan et al. [57], while the urease activity was tested in Urea Agar according to Hussain Qadri et al. [58].
4.2. Molecular Identification
A pure IBR3 fresh colony was collected from a TSA plate using a sterile syringe needle and transferred into the already prepared PCR reaction mixture [59]. The PCR amplification of the 16s rRNA genes was done using 27F (5′-AGAGTTTGATCCTGGCTCAC-3′) and 1492R (5′-CGGTTACCTTGTTACGACTT-3′) primers, according to Sharma et al. [60]. After the quality check, PCR products were sequenced by Eurofins Scientific (Milan, Italy). The resulting chromatograms were visually inspected, and regions of low quality were removed. The NCBI database and the BLAST search tool were used to identify the genus and species of the bacteria. Phylogenetic trees were reconstructed using the neighbor-joining algorithms with the Kimura 2-parameter, implemented in MEGA 11 software [61,62]. Bootstrap analysis was based on 1000 replications. Sequences of other Paracoccus species used in the phylogenetic analysis were retrieved from GenBank (Table 3).
4.3. Essential Oil of Thymus serpyllum
The essential oil of T. serpyllum (Thy-EO) was provided by Inalme SRL (Catania, Italy). Thy-EO was submitted for GC–MS analysis. The analyses were performed using the Konik instrumentation equipped with a 5000C gas chromatograph coupled to a mass spectrometer detector MS Q2 (Sant Cugat del Vallés, Barcelona, Spain). The gas chromatograph operated with a Merck (Darmstadt, Germany) fused-silica (30 m × 0.25 mm i.d.) The gas chromatograph operated with a Macherey-Nagel (Düren, Germany) fused-silica (30 m × 0.25 mm i.d.) column coated with 5% Diphenyl/ 95% Dimethylpolysiloxano (SLB-5ms, film thickness 0.25 μm) and under the following conditions: injector temperature at 220 °C; transfer line at 240 °C; oven temperature programmed from 60 °C to 246 °C, at 3 °C/min; carrier gas—He at 1.0 mL/min. The mass spectrometer operated in electron impact (EI) mode at 70 eV with a scan range from 45 to 450 amu (2 scan/min).
4.4. Growth Inhibition Assays
The microbial growth of the strain IBR3 was evaluated using the 96-well plate microdilution assay. Thy-EO (40 mg) was added to 0.5% Tween 80 and the volume was made to 2 mL with sterile Tryptic Soy Broth (TSB) to provide a stock solution containing 20 mg/mL of oil. Serial twofold dilutions of the Thy-EO stock were made with TSB to yield final concentrations ranging from 0.1 to 1 mg/mL. A bacterial solution (20 μL; 9x107 CFU/mL) was transferred to the microplate wells and mixed with the micropipette. Positive (A: TSB plus bacterial cells; B: TSB, 0.5% Tween 80 and bacterial cells) and negative controls (TSB) were included in the plates. The plates were sealed and incubated aerobically at 28°C, with orbital shaking at 150 rpm. After 12 hours, the results were measured using a SpectraMax ABS PLUS UV-VIS spectrophotometer (OD600 nm). The MIC value was defined as the lowest concentration of Thy-EO that inhibited the growth of the microorganism. To evaluate the MBC, a volume of 10 μL of each well was spread onto TSA and incubated aerobically at 28°C for 24 h. The MCB was determined as the concentration with no visible growth on the plate. Bacterial cultures without treatments served as positive controls. Effective dose (EC50) values, which is the concentration of a drug that gives half-maximal response, was calculated by probit analysis using the Graphpad Instat® software (San Diego, CA, USA) [63].
4.5. Membrane Integrity of IBR3
The integrity of the IBR3 cell membrane was investigated by measuring the absorbance of cellular constituents at 260 nm and 280 nm as described by Cao et al. [64]. Specifically, 5% of the overnight culture was added to fresh TSB and cultured at 37 °C until an OD600 nm of 0.12-0.14 was reached. Then, increasing concentrations of Thy-EO, equal to MIC, 2×MIC, and 4×MIC values, were incubated with the IBR3 suspension at 37 °C, shaking at 150rpm for 2 h. Triton X-100 1% and TSB were used as positive and negative controls, respectively. At time 0 and after 2 hours the samples were centrifuged at 3000 rpm for 10 min, and supernatants were read using a SpectraMax ABS PLUS UV-Vis spectrophotometer at 260 nm and 280 nm.
4.6. Crystal Violet Assay for IBR3 Membrane Permeability
The variation in membrane permeability of the IBR3 strain was evaluated using the crystal violet uptake assay, as described by Vaara & Vaara [42]. Bacterial cells grown in TSB to a concentration of 10⁸ CFU/mL were harvested by centrifugation at 4,500 × g for 5 min at 4 °C. The resulting pellet was washed twice with PBS, (pH 7.4), resuspended in PBS containing Thy-EO at 0.4 mg/mL, and incubated at 37 °C for 30 min. Control samples were prepared under identical conditions but without Thy-EO treatment. Following incubation, the cells were centrifuged at 9,300 × g for 5 min and resuspended in PBS containing 10 µg/mL of crystal violet. The suspension was incubated at 37 °C for 10 min with gentle shaking, then centrifuged at 13,400 × g for 15 min. The absorbance of the supernatant was measured at 590 nm using a SpectraMax ABS PLUS UV-Vis spectrophotometer. The OD of the original crystal violet solution (without bacterial exposure) was taken as 100%. The percentage of crystal violet uptake by the samples was calculated using the following formula:
4.7. ATR-FTIR Spectroscopy
Thy-EO at different concentrations (0.3; 0.4; 0.8; 1.2 mg/mL) was added to a cell suspension of pure IBR3 culture in exponential phase (OD=0.4). Treatment was performed for 3 h at 30 °C [44]. Cells were then washed three times with 0.9 NaCl solution and centrifuged at 11.000 rpm for 2 min and dried at environmental temperature (25 °C) [66]. Control samples consisted in untreated IBR3 pure culture. All IR spectra (4000–600 cm−1) were obtained by attenuated total reflection (ATR) technique, using an Agilent FTIR Cary 630 model spectrometer equipped with a diamond ATR sampling accessory. One hundred and twenty-eight scans were taken with 4 cm−1 resolution.
4.8. Role of Paracoccus in Mural Painting Deterioration
In order to better understand the involvement of Paracoccus in mural painting deterioration, records from the NCBI BioSample database have been analyzed, identifying relevant strains and their reported environmental contexts.
4.9. Statistical Analysis
All the experiments were performed three times with triplicates. The data were presented as means ± standard errors. Data were tested for normality with the Shapiro-Wilk test, therefore, ANOVA with Tukey’s multiple comparison of means test was conducted (p value ≤ 0.05), using the Graphpad Instat® software (San Diego, CA, USA).
Author Contributions
Investigation, , A.M.V, M.N, C.P., and V.V.; Conceptualization, A.M.V Methodology, C.P., and V.V, and A.M.V.; Resources and Supervision A.M.V; Data Curation, A.M.V.; Writing—Original Draft Preparation A.M.V; Writing—Review and Editing, M.N., C.P., V.V., C.A, and A.M.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article
Acknowledgments
The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Abbreviation
The following abbreviations are used in this manuscript:
Thy-EO : Thymus serpyllum essential oil
References
- Suphaphimol, N.; Suwannarach, N.; Purahong, W.; Jaikang, C.; Pengpat, K.; Semakul, N.; Yimklan, S.; Jongjitngam, S.; Jindasu, S.; Thiangtham, S.; et al. Identification of Microorganisms Dwelling on the 19th Century Lanna Mural Paintings from Northern Thailand Using Culture-Dependent and -Independent Approaches. Biology 2022, 11, 228. [CrossRef]
- Pei, S.; Wu, F.; Chen, Y.; Ma, W.; He, D.; Zhang, Q.; Gu, J.-D.; Wang, W.; Tian, T.; Feng, H. Mechanisms of Lead-Containing Pigment Discoloration Caused by Naumannella Cuiyingiana AFT2T Isolated from 1500 Years Tomb Wall Painting of China. Int. Biodeterior. Biodegrad. 2023, 185, 105689. [CrossRef]
- He, D.; Wu, F.; Ma, W.; Zhang, Y.; Gu, J.-D.; Duan, Y.; Xu, R.; Feng, H.; Wang, W.; Li, S.-W. Insights into the Bacterial and Fungal Communities and Microbiome That Causes a Microbe Outbreak on Ancient Wall Paintings in the Maijishan Grottoes. Int. Biodeterior. Biodegrad. 2021, 163, 105250. [CrossRef]
- Hoffmann, L. Caves and Other Low-Light Environments: Aerophitic Photoautotrophic Microorganisms. In Encyclopedia of Environmental Microbiology; Bitton, G., Ed.; Wiley, 2003 ISBN 978-0-471-35450-5.
- Gonzalez, I.; Laiz, L.; Hermosin, B.; Caballero, B.; Incerti, C.; Saiz-Jimenez, C. Bacteria Isolated from Rock Art Paintings: The Case of Atlanterra Shelter (South Spain). J. Microbiol. Methods 1999, 36, 123–127. [CrossRef]
- Pavić, A.; Ilić-Tomić, T.; Pačevski, A.; Nedeljković, T.; Vasiljević, B.; Morić, I. Diversity and Biodeteriorative Potential of Bacterial Isolates from Deteriorated Modern Combined-Technique Canvas Painting. Int. Biodeterior. Biodegrad. 2015, 97, 40–50. [CrossRef]
- Saiz-Jimenez, I.G., C. Actinomycetes in Hypogean Environments. Geomicrobiol. J. 1999, 16, 1–8. [CrossRef]
- Gorbushina, A.A.; Heyrman, J.; Dornieden, T.; Gonzalez-Delvalle, M.; Krumbein, W.E.; Laiz, L.; Petersen, K.; Saiz-Jimenez, C.; Swings, J. Bacterial and Fungal Diversity and Biodeterioration Problems in Mural Painting Environments of St. Martins Church (Greene–Kreiensen, Germany). Int. Biodeterior. Biodegrad. 2004, 53, 13–24. [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; Feo, V.D. Essential Oils and Antifungal Activity. Pharmaceuticals 2017, 10, 86. [CrossRef]
- Palla, F.; Bruno, M.; Mercurio, F.; Tantillo, A.; Rotolo, V. Essential Oils as Natural Biocides in Conservation of Cultural Heritage. Molecules 2020, 25, 730. [CrossRef]
- Liao, W.; Badri, W.; Dumas, E.; Ghnimi, S.; Elaissari, A.; Saurel, R.; Gharsallaoui, A. Nanoencapsulation of Essential Oils as Natural Food Antimicrobial Agents: An Overview. Appl. Sci. 2021, 11, 5778. [CrossRef]
- Vettraino, A.M.; Zikeli, F.; Scarascia Mugnozza, G.; Vinciguerra, V.; Tabet, D.; Romagnoli, M. Lignin Nanoparticles Containing Essential Oils for Controlling Phytophthora Cactorum Diseases. For. Pathol. 2022, 52, e12739. [CrossRef]
- Vettraino, A.M.; Zikeli, F.; Humar, M.; Biscontri, M.; Bergamasco, S.; Romagnoli, M. Essential Oils from Thymus Spp. as Natural Biocide against Common Brown- and White-Rot Fungi in Degradation of Wood Products: Antifungal Activity Evaluation by in Vitro and FTIR Analysis. Eur. J. Wood Prod. 2023, 81, 747–763. [CrossRef]
- Zikeli, F.; Vettraino, A.M.; Biscontri, M.; Bergamasco, S.; Palocci, C.; Humar, M.; Romagnoli, M. Lignin Nanoparticles with Entrapped Thymus Spp. Essential Oils for the Control of Wood-Rot Fungi. Polymers 2023, 15, 2713. [CrossRef]
- Isola, D.; Capobianco, G.; Tovazzi, V.; Pelosi, C.; Trotta, O.; Serranti, S.; Lanteri, L.; Zucconi, L.; Spizzichino, V. Biopatinas on Peperino Stone: Three Eco-Friendly Methods for Their Control and Multi-Technique Approach to Evaluate Their Efficacy. Microorganisms 2025, 13, 375. [CrossRef]
- Corbu, V.M.; Gheorghe-Barbu, I.; Marinas, I.C.; Avramescu, S.M.; Pecete, I.; Geanǎ, E.I.; Chifiriuc, M.C. Eco-Friendly Solution Based on Rosmarinus Officinalis Hydro-Alcoholic Extract to Prevent Biodeterioration of Cultural Heritage Objects and Buildings. Int J Mol Sci 2022, 23, 11463. [CrossRef]
- Corbu, V.M.; Gheorghe, I.; Marinaș, I.C.; Geană, E.I.; Moza, M.I.; Csutak, O.; Chifiriuc, M.C. Demonstration of Allium Sativum Extract Inhibitory Effect on Biodeteriogenic Microbial Strain Growth, Biofilm Development, and Enzymatic and Organic Acid Production. Molecules 2021, 26, 7195. [CrossRef]
- Geweely, N.S.; Afifi, H.A.; Ibrahim, D.M.; Soliman, M.M. Efficacy of Essential Oils on Fungi Isolated from Archaeological Objects in Saqqara Excavation, Egypt. Geomicrobiol. J. 2019, 36, 148–168. [CrossRef]
- Senbua, W.; Wichitwechkarn, J. Molecular Identification of Fungi Colonizing Art Objects in Thailand and Their Growth Inhibition by Local Plant Extracts. 3 Biotech 2019, 9, 356. [CrossRef]
- Albasil, M.D.; Mahgoub, G.; El Hagrassy, A.; Reyad, A.M. Evaluating the Antimicrobial Activity of Essential Oils in the Conservation of Mural Paintings. Conserv. Sci. Cult. Herit. : 21 2021. [CrossRef]
- Mohamed, A.A.; Ahmed, M.A.; Korayem, A.S.; Abu-Hussien, S.H.; Rashidy, W.B. Antifungal, Toxicological, and Colorimetric Properties of Origanum Vulgare, Moringa Oleifera, and Cinnamomum Verum Essential Oils Mixture against Egyptian Prince Yusuf Palace Deteriorative Fungi. BMC Biotechnol 2025, 25, 4. [CrossRef]
- Harker, M.; Hirschberg, J.; Oren, A. Paracoccus Marcusii Sp. Nov., an Orange Gram-Negative Coccus. Int. J. Syst. Bacteriol. 1998, 48, 543–548. [CrossRef]
- Wang, Y.-S.; Yan, Z.-F.; Lin, P.; Gao, W.; Yi, T.-H. Paracoccus Pueri Sp. Nov., Isolated from Pu’er Tea. Antonie Van Leeuwenhoek 2018, 111, 1535–1542. [CrossRef]
- Lee, J.H.; Kim, Y.S.; Choi, T.-J.; Lee, W.J.; Kim, Y.T. Paracoccus Haeundaensis Sp. Nov., a Gram-Negative, Halophilic, Astaxanthin-Producing Bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1699–1702. [CrossRef]
- Tsubokura, A.; Yoneda, H.; Mizuta, H. Paracoccus Carotinifaciens Sp. Nov., a New Aerobic Gram-Negative Astaxanthin-Producing Bacterium. Int. J. Syst. Evol. Microbiol. 1999, 49, 277–282. [CrossRef]
- Zhu, S.; Zhao, Q.; Zhang, G.; Jiang, Z.; Sheng, H.; Feng, H.; An, L. Paracoccus Tibetensis Sp. Nov., Isolated from Qinghai-Tibet Plateau Permafrost. Int. J. Syst. Evol. Microbiol. 2013, 63, 1902–1905. [CrossRef]
- Dominguez-Moñino, I.; Jurado, V.; Hermosin, B.; Saiz-Jimenez, C. Paracoccus Cavernae Sp. Nov., Isolated from a Show Cave. Int. J. Syst. Evol. Microbiol. 2016, 66, 2265–2270. [CrossRef]
- Gutierrez-Patricio, S.; Gonzalez-Pimentel, J.L.; Miller, A.Z.; Hermosin, B.; Saiz-Jimenez, C.; Jurado, V. Paracoccus Onubensis Sp. Nov., a Novel Alphaproteobacterium Isolated from the Wall of a Show Cave. Int J Syst Evol Microbiol 2021, 71, 004942. [CrossRef]
- Puri, A.; Bajaj, A.; Singh, Y.; Lal, R. Harnessing Taxonomically Diverse and Metabolically Versatile Genus Paracoccus for Bioplastic Synthesis and Xenobiotic Biodegradation. J Appl Microbiol 2022, 132, 4208–4224. [CrossRef]
- Gomoiu, I.; Mohanu, D.; Radvan, R.; Dumbravician, M.; Neagu, S.E.; Cojoc, L.R.; Enache, M.I.; Chelmus, A.; Mohanu, I. Environmental Impact on Biopigmentation of Mural Painting. Acta Phys. Pol. A 2017, 131, 48–51. [CrossRef]
- Cojoc, L.R.; Enache, M.I.; Neagu, S.E.; Lungulescu, M.; Setnescu, R.; Ruginescu, R.; Gomoiu, I. Carotenoids Produced by Halophilic Bacterial Strains on Mural Paintings and Laboratory Conditions. FEMS Microbiol. Lett. 2019, 366, fnz243. [CrossRef]
- Ma, Y.; Zhang, H.; Du, Y.; Tian, T.; Xiang, T.; Liu, X.; Wu, F.; An, L.; Wang, W.; Gu, J.-D.; et al. The Community Distribution of Bacteria and Fungi on Ancient Wall Paintings of the Mogao Grottoes. Sci Rep 2015, 5, 7752. [CrossRef]
- Pietrzak, K.; Puchalski, M.; Otlewska, A.; Wrzosek, H.; Guiamet, P.; Piotrowska, M.; Gutarowska, B. Microbial Diversity of Pre-Columbian Archaeological Textiles and the Effect of Silver Nanoparticles Misting Disinfection. J. Cult. Herit. 2017, 23, 138–147. [CrossRef]
- Saridaki, A.; Katsivela, E.; Glytsos, T.; Tsiamis, G.; Violaki, E.; Kaloutsakis, A.; Kalogerakis, N.; Lazaridis, M. Identification of Bacterial Communities on Different Surface Materials of Museum Artefacts Using High Throughput Sequencing. J. Cult. Herit. 2022, 54, 44–52. [CrossRef]
- Pavlović, J.; Puškárová, A.; Planý, M.; Farkas, Z.; Rusková, M.; Kvalová, K.; Kraková, L.; Bučková, M.; Pangallo, D. Colored Stains: Microbial Survey of Cellulose-Based and Lignin Rich Papers. Int. J. Biol. Macromol. 2023, 241, 124456. [CrossRef]
- Borrego, S.; Saravia, S.G.D.; Oderlaise Valdés; Isbel Vivar; Battistoni, P.; Guiamet, P. Biocidal activity of two essential oils on the fungal biodeterioration of historical documents. Int. J. Conserv. Sci. 2016, 7, 369-380. [CrossRef]
- Bosco, F.; Mollea, C.; Demichela, M.; Fissore, D. Application of Essential Oils to Control the Biodeteriogenic Microorganisms in Archives and Libraries. Heritage 2022, 5, 2181–2195. [CrossRef]
- Russo, R.; Palla, F. Plant Essential Oils as Biocides in Sustainable Strategies for the Conservation of Cultural Heritage. Sustainability 2023, 15, 8522. [CrossRef]
- Fernandes, L.; Paiva, D.S.; Pereira, E.; Rufino, A.C.; Landim, E.; Marques, M.P.; Cabral, C.; Portugal, A.; Mesquita, N. Evaluating the Antifungal Activity of Volatilized Essential Oils on Fungi Contaminating Artifacts from a Museum Collection. Appl. Sci. 2025, 15, 2378. [CrossRef]
- Lavin, P.; De Saravia, S.G.; Guiamet, P. Scopulariopsis Sp. and Fusarium Sp. in the Documentary Heritage: Evaluation of Their Biodeterioration Ability and Antifungal Effect of Two Essential Oils. Microb Ecol 2016, 71, 628–633. [CrossRef]
- Casiglia, S.; Bruno, M.; Scandolera, E.; Senatore, F.; Senatore, F. Influence of Harvesting Time on Composition of the Essential Oil of Thymus Capitatus (L.) Hoffmanns. & Link. Growing Wild in Northern Sicily and Its Activity on Microorganisms Affecting Historical Art Crafts. Arab. J. Chem. 2019, 12, 2704–2712. [CrossRef]
- Vaara, M.; Vaara, T. Outer Membrane Permeability Barrier Disruption by Polymyxin in Polymyxin-Susceptible and -Resistant Salmonella Typhimurium. Antimicrob Agents Chemother 1981, 19, 578–583. [CrossRef]
- Huleihel, M.; Pavlov, V.; Erukhimovitch, V. The Use of FTIR Microscopy for the Evaluation of Anti-Bacterial Agents Activity. J. Photochem. Photobiol. B: Biol. 2009, 96, 17–23. [CrossRef]
- Devi, K.P.; Nisha, S.A.; Sakthivel, R.; Pandian, S.K. Eugenol (an Essential Oil of Clove) Acts as an Antibacterial Agent against Salmonella Typhi by Disrupting the Cellular Membrane. J. Ethnopharmacol. 2010, 130, 107–115. [CrossRef]
- Di Rosario, M.; Continisio, L.; Mantova, G.; Carraturo, F.; Scaglione, E.; Sateriale, D.; Forgione, G.; Pagliuca, C.; Pagliarulo, C.; Colicchio, R.; et al. Thyme Essential Oil as a Potential Tool Against Common and Re-Emerging Foodborne Pathogens: Biocidal Effect on Bacterial Membrane Permeability. Microorganisms 2024, 13, 37. [CrossRef]
- Xu, J.; Zhou, F.; Ji, B.-P.; Pei, R.-S.; Xu, N. The Antibacterial Mechanism of Carvacrol and Thymol against Escherichia Coli. Lett Appl Microbiol 2008, 47, 174–179. [CrossRef]
- Yap, P.S.X.; Yusoff, K.; Lim, S.-H.E.; Chong, C.-M.; Lai, K.-S. Membrane Disruption Properties of Essential Oils—A Double-Edged Sword? Processes 2021, 9, 595. [CrossRef]
- Al-Mijalli, S.H.; El Hachlafi, N.; Jeddi, M.; Abdallah, E.M.; Assaggaf, H.; Qasem, A.; Lee, L.-H.; Law, J.W.-F.; Aladhadh, M.; Alnasser, S.M.; et al. Unveiling the Volatile Compounds and Antibacterial Mechanisms of Action of Cupressus Sempervirens L., against Bacillus subtilis and Pseudomonas Aeruginosa. Biomed. Pharmacother. 2023, 167, 115609. [CrossRef]
- Kauser, S.; Raj, N.; Ahmedi, S.; Manzoor, N. Mechanistic Insight into the Membrane Disrupting Properties of Thymol in Candida Species. Microbe 2024, 2, 100045. [CrossRef]
- Coico, R. Gram Staining. CP Microbiol. 2006, 00. [CrossRef]
- Reller, L.B.; Mirrett, S. Motility-Indole-Lysine Medium for Presumptive Identification of Enteric Pathogens of Enterobacteriaceae. J Clin Microbiol 1975, 2, 247–252. [CrossRef]
- Maheswaran, M.; Forchhammer, K. Carbon-Source-Dependent Nitrogen Regulation in Escherichia Coli Is Mediated through Glutamine-Dependent GlnB Signalling. Microbiology 2003, 149, 2163–2172. [CrossRef]
- Kraková, L.; Chovanová, K.; Selim, S.A.; Šimonovičová, A.; Puškarová, A.; Maková, A.; Pangallo, D. A Multiphasic Approach for Investigation of the Microbial Diversity and Its Biodegradative Abilities in Historical Paper and Parchment Documents. Int. Biodeterior. Biodegrad. 2012, 70, 117–125. [CrossRef]
- Dhawale, M.R.; Wilson, J.J.; Khachatourians, G.G.; Ingledew, W.M. Improved Method for Detection of Starch Hydrolysis. Appl Env. Microbiol 1982, 44, 747–750. [CrossRef]
- Cacchio, P.; Contento, R.; Ercole, C.; Cappuccio, G.; Martinez, M.P.; Lepidi, A. Involvement of Microorganisms in the Formation of Carbonate Speleothems in the Cervo Cave (L’Aquila-Italy). Geomicrobiol. J. 2004, 21, 497–509. [CrossRef]
- Jurado, V.; Gonzalez-Pimentel, J.L.; Hermosin, B.; Saiz-Jimenez, C. Biodeterioration of Salón de Reinos, Museo Nacional Del Prado, Madrid, Spain. Appl. Sci. 2021, 11, 8858. [CrossRef]
- Doğan, B.; Yıldız, Z.; Aksöz, N.; Eninanç, A.B.; Dağ, İ.; Yıldız, A.; Doğan, H.H.; Yamaç, M. Flask and Reactor Scale Production of Plant Growth Regulators by Inonotus Hispidus: Optimization, Immobilization and Kinetic Parameters. Prep. Biochem. Biotechnol. 2023, 53, 1210–1223. [CrossRef]
- Hussain Qadri, S.M.; Zubairi, S.; Hawley, H.P.; Mazlaghani, H.H.; Ramirez, E.G. Rapid Test for Determination of Urea Hydrolysis. Antonie Van Leeuwenhoek 1984, 50, 417–423. [CrossRef]
- Woodman, M.E.; Savage, C.R.; Arnold, W.K.; Stevenson, B. Direct PCR of Intact Bacteria (Colony PCR). CP Microbiol. 2016, 42. [CrossRef]
- Sharma, M.; Schmid, M.; Rothballer, M.; Hause, G.; Zuccaro, A.; Imani, J.; Kämpfer, P.; Domann, E.; Schäfer, P.; Hartmann, A.; et al. Detection and Identification of Bacteria Intimately Associated with Fungi of the Order Sebacinales. Cell. Microbiol. 2008, 10, 2235–2246. [CrossRef]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide Sequences. J Mol Evol 1980, 16, 111–120. [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [CrossRef]
- Finney, D.J. Statistical Logic in the Monitoring of Reactions to Therapeutic Drugs. Methods Inf Med 1971, 10, 237–245. [CrossRef]
- Cao, S.; Du, R.; Zhao, F.; Xiao, H.; Han, Y.; Zhou, Z. The Mode of Action of Bacteriocin CHQS, a High Antibacterial Activity Bacteriocin Produced by Enterococcus Faecalis TG2. Food Control 2019, 96, 470–478. [CrossRef]
- Li, N.; Luo, M.; Fu, Y.; Zu, Y.; Wang, W.; Zhang, L.; Yao, L.; Zhao, C.; Sun, Y. Effect of Corilagin on Membrane Permeability of Escherichia Coli, Staphylococcus Aureus and Candida Albicans. Phytother. Res. 2013, 27, 1517–1523. [CrossRef]
- Yang, H.; Shi, H.; Feng, B.; Wang, L.; Chen, L.; Alvarez-Ordóñez, A.; Zhang, L.; Shen, H.; Zhu, J.; Yang, S.; et al. Protocol for Bacterial Typing Using Fourier Transform Infrared Spectroscopy. STAR Protoc. 2023, 4, 102223. [CrossRef]
Figure 1.
Neighbour-joining tree based on the 16S rRNA gene sequences, showing the phylogenetic position of strain IBR3 among closely related members of the genus Paracoccus. The related species, strain name and accession number were collected from NCBI, GenBank database. Bootstrap values are shown at branch points (1000 bootstrap replicates).
Figure 1.
Neighbour-joining tree based on the 16S rRNA gene sequences, showing the phylogenetic position of strain IBR3 among closely related members of the genus Paracoccus. The related species, strain name and accession number were collected from NCBI, GenBank database. Bootstrap values are shown at branch points (1000 bootstrap replicates).
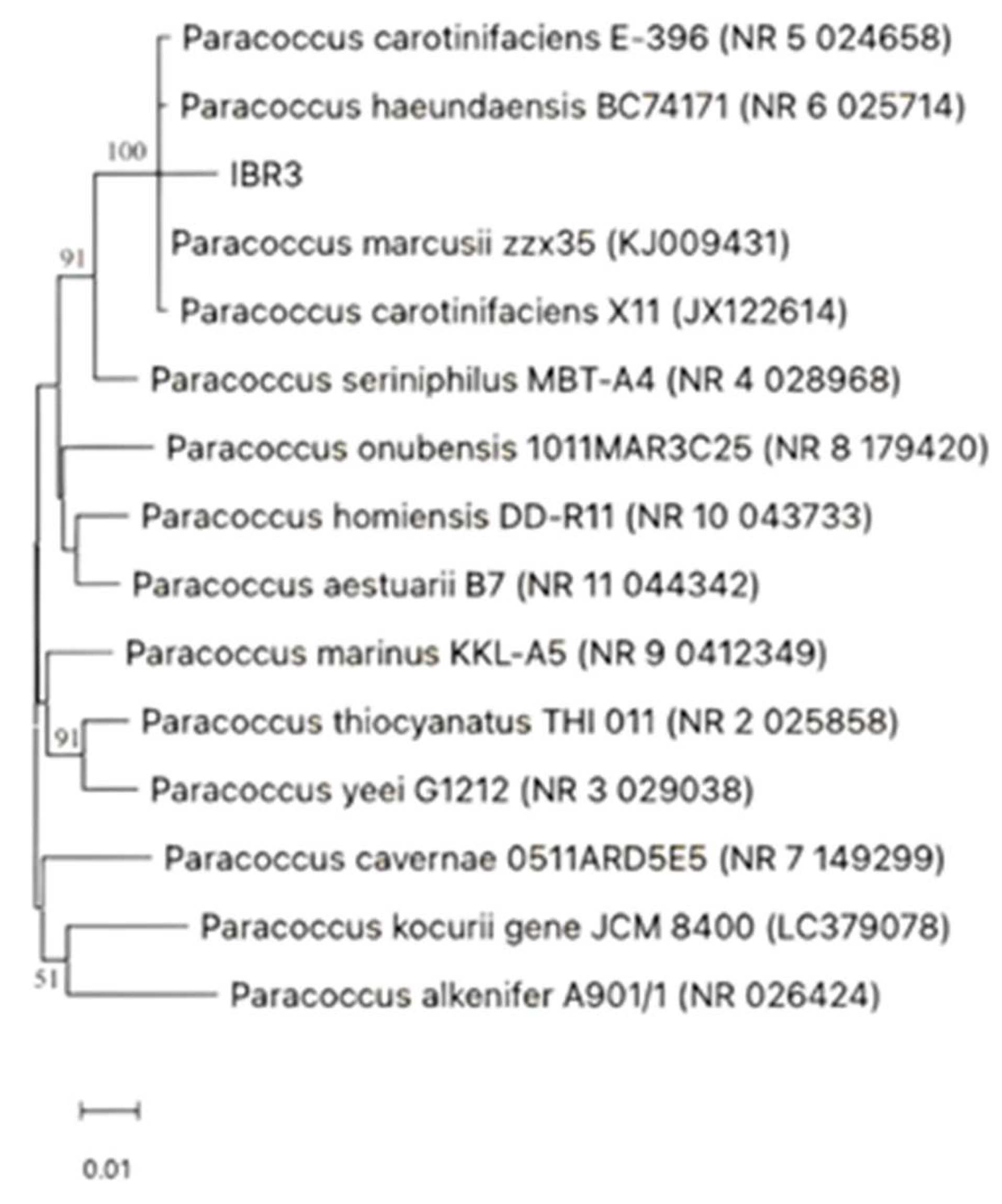
Figure 2.
Paracoccus IBR3 growth at different Thy-EO concentrations, incubation at at 28°C for 12 hours.
Figure 2.
Paracoccus IBR3 growth at different Thy-EO concentrations, incubation at at 28°C for 12 hours.
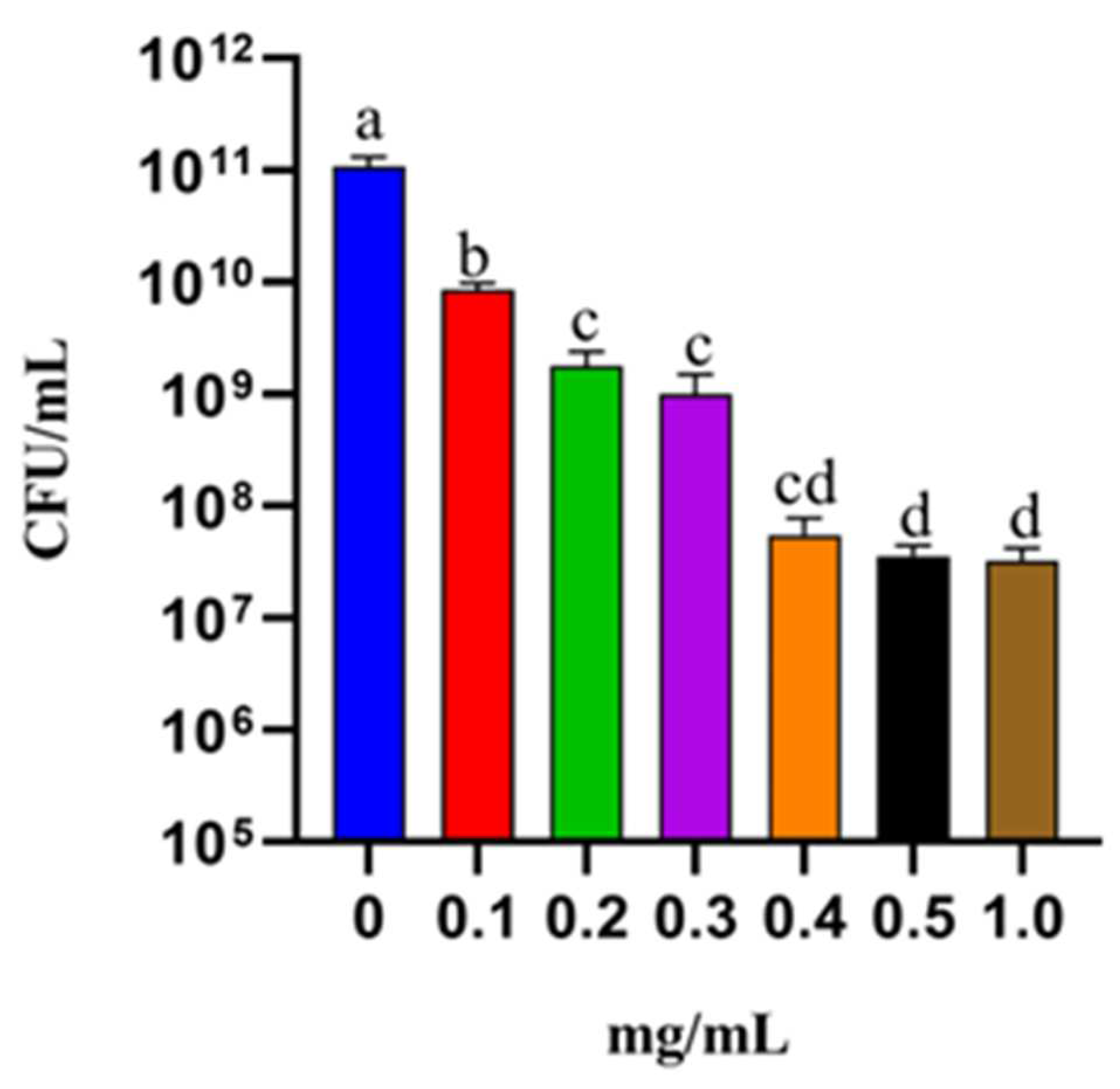
Figure 3.
Optical density values at 260/280 nm of UV-absorbing material were measured following treatment of IBR3 with Thy-EO at MIC, 2× MIC, 4× MIC, and Triton X-100 (1%) at 0 and 2 hours.
Figure 3.
Optical density values at 260/280 nm of UV-absorbing material were measured following treatment of IBR3 with Thy-EO at MIC, 2× MIC, 4× MIC, and Triton X-100 (1%) at 0 and 2 hours.


Table 1.
Comparison of the properties of the strain IBR3 and closely related strains: P. marcusii; P. haeundaensis; P. carotinifaciens.
Table 1.
Comparison of the properties of the strain IBR3 and closely related strains: P. marcusii; P. haeundaensis; P. carotinifaciens.
| Characteristic |
Paracoccus IBR3 |
P. marcusii MH1T |
P. haeundaensis BC74171T |
P. carotinifaciens E-396T |
|---|---|---|---|---|
| Morphology | Cocci to short rods, 1.0 μm in size, growing in single, pairs or short chains. | Cocci to short rods, 1.0-1.3 μm in size, growing in single, pairs or short chains. | rod-shaped, | rod-shaped, |
| 0.3–0.7 μm in diameter and 0.8–2.5 μm in length. | 0.3-0.75 μm in diameter and 1.0-5.0 μm in length. | |||
| Motility | - | - | + | + |
| Orange to red pigment | + | + | + | + |
| Growth at 40 °C | - | - | - | - |
| Growth on: | ||||
| Sucrose | + | + | - | + |
| Fructose | + | + | - | + |
| D-Sorbitol | + | + | - | + |
| D-Glucose | + | + | - | + |
| L- Arabinose | + | + | + | - |
| D-Mannose | + | + | - | + |
| D-Mannitol | + | + | - | + |
| Production of: | ||||
| Urease | - | - | - | - |
| Indole from tryptophan | - | - | - | - |
| Organic acids from D-glucose | + | + | NR | + |
| Hidrolysis of: | ||||
| Starch | - | - | + | - |
| Gelatin | W | W | NR | - |
| +, Positive; -, negative; W, weakly positive; NR, not reported. | ||||
| Data for P. marcusii were obtained from Harker et al. [22] and Wang et al. [23], data for P. haeundaensis BC74171ᵀ were from Lee et al. [24] and data for P. carotinifaciens E-396ᵀ were from Tsubokura et al. [25]. | ||||
Table 2.
Chemical composition of Thymus serpyllum EO based on the GC-MS analysis.
| Components | Relative concentration (%) |
Retention time (min) |
|---|---|---|
| α-Pinene | 0.82 | 15.082 |
| Fenchene | 0.34 | 15.889 |
| Camphene | 2.06 | 16.018 |
| trans-p-Menthane | 0.17 | 17.366 |
| β-Pinene | 0.71 | 17.443 |
| cis-p-Menthane | 0.22 | 18.104 |
| p-Cymene | 40.28 | 19.657 |
| p-Cymen-8-ol | 0.09 | 28.087 |
| Thymol | 2.45 | 32.980 |
| Carvacrol | 51.88 | 33.478 |
Table 3.
NCBI GeBank ID of Paracoccus species used in the phylogenetic analysis.
| Paracoccus strain | NCBI ID |
|---|---|
| P. carotinifaciens E-396 | NR 5 024658 |
| P. haeundaensis BC74171 | NR 6 025714 |
| P. marcusi zzx35 | KJ009431 |
| P. carotinifaciens X11 | JX122614 |
| P. seriniphylus MBT-A4 | NR 4 028968 |
| P. onubensis 1011MAR3C25 | NR 8 179420 |
| P. homiensis DD-R11 | NR 10 043733 |
| P. aestuarii B7 | NR 11 044342 |
| P. marinus KKL-A5 | NR 9 0412349 |
| P. thiocyanatus THI 011 | NR 2 025858 |
| P. yeei G1212 | NR 3 029038 |
| P. cavernae 0511ARD5E5 | NR 7 149299 |
| P. kocurii gene JCM 8400 | LC379078 |
| P. alkenifer A 901/1 | NR 026424 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



