Submitted:
28 May 2025
Posted:
28 May 2025
You are already at the latest version
Abstract
Swertiamarin (SW) is a natural iridoid glycoside primarily isolated from Gentianaceae family and has gained significant attention in natural product research in recent years. Pharmacological studies have demonstrated that swertiamarin possesses a wide range of pharmacological activities including antioxidant, anti-inflammatory, anti-tumor, antidiabetic, and neuroprotective activities, and its mechanism of action involves multiple signaling pathways such as Nrf2, NF-κB, and MAPK. In this study, we systematically reviewed the progress of the pharmacokinetic characteristics, pharmacological effects, and molecular mechanisms of swertiamarin. This review aims to provide a scientific basis and reference for the clinical application and drug development of Swertiamarin.
Keywords:
Swertiamarin
; Gentianaceae
; pharmacokinetic properties
; pharmacological effects
; molecular mechanisms
1. Introduction
Swertiamarin(SW) is a natural iridoid glycoside primarily isolated from Gentianaceae plants [1]. As a medicinal plant of natural origin, extracts from Gentianaceae possess various pharmacological activities, including anti-inflammatory and antioxidant, hepatoprotective, and anti-diabetic effects [2,3,4,5,6,7,8,9,10]. One of its main active components, swertiamarin, also plays a significant therapeutic role in treating diabetes mellitus, cardiovascular diseases, and neurological disorders. Furthermore, research on its pharmacokinetic properties and toxicological safety continues to advance. Previous reviews have highlighted its role in hypoglycemic [11] and metabolic diseases [12,13]. In this paper, we will focus on a systematic review of the pharmacokinetic properties, pharmacological effects, and molecular mechanisms associated with swertiamarin, providing a reference for its clinical application and development.
Figure 1.
Chemical structure of swertiamarin.
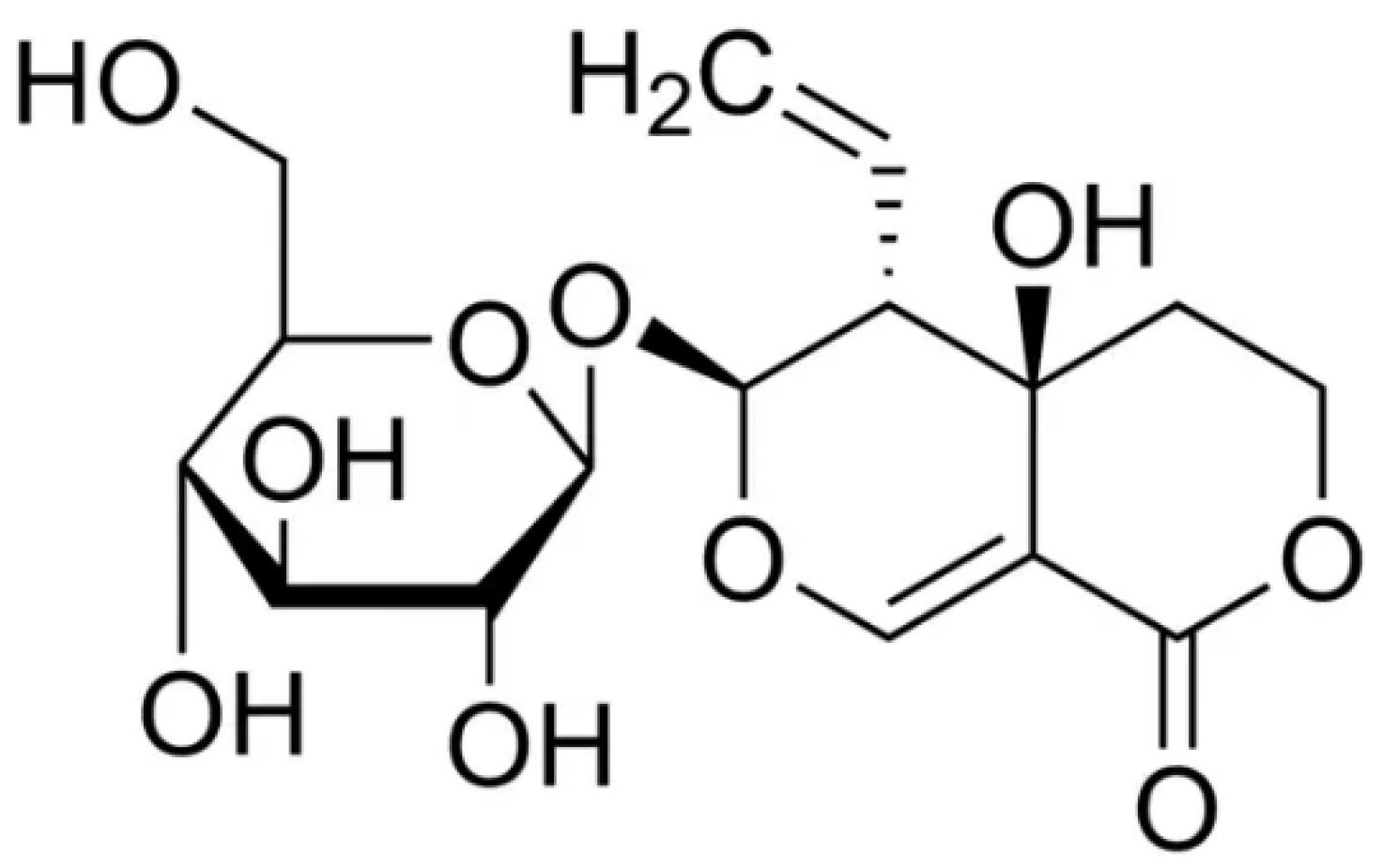
2. Pharmacokinetic properties
The analysis of pharmacokinetic data showed that SW is rapidly absorbed into the circulatory system after oral administration of SW (20 mg/kg) in SD rats, reaching peak concentrations in approximately 0.95 hours, and has a short half-life, the t1/2z (h) is 1.104 ± 0.22, Vz/F (L/kg) is 9.637 ± 4.322, CLz/F (L/h/kg) is 5.638 ± 2.151, AUC (μg/L h) is 3593.7 ± 985.4, MRT0–∞ (h) is 1.929 ± 0.364, Tmax (h) is 0.945 ± 0.136, Cmax (μg/L) is 1920.1 ± 947.0 [14]. The absolute bioavailability of SW was low at 10.3%, which may be attributed to its poor permeability through the intestinal epithelial membrane and first-pass effect in the liver. Following a single oral dose of SW (50 mg/kg) to rats was rapidly distributed throughout the tissues and peaked at 1h. Its concentrations are relatively high in the kidney and liver suggesting that SW is rapidly metabolized in the liver and excreted by the kidneys, while its levels in the brain were notably low due to its high hydrophilicity, which hinders its ability to cross the blood-brain barrier [15,16]. The nine prototype constituents were rapidly absorbed into the circulation system and distributed to liver and lung tissues after oral administration of Jingyin Granules to rats, and the levels of SW in serum, liver, and lung were the highest compared with the other eight components 1 h after administration [17].
The pharmacokinetic (PK) parameters of SW are also affected by various combinations of drug administration. Studies on the pharmacokinetics of the three SW (20 mg/kg) administration types in rats showed that QYDT and co-administration of SW and oleanolic acid significantly decreased peak plasma concentration and AUC, as well as increased CL and Vz/F compared with Oral administration of SW alone. These data indicate that oleanolic acid or another component of QYDT might reduce the oral bioavailability of SW in rats [18]. Furthermore, analysis of the PK parameters of four components (gentiopicroside, geniposide, baicalin, and swertiamarin) in rat plasma after oral administration of the Chinese herbal prescription LDXGT (10 g/kg, p.o.) showed that SW exhibited the lowest level of AUC (2.5 ± 0.1 min µg/mL) and fell below the lower limit of quantification (LLOQ) [16]. Additionally, pathological conditions and the compatibility effects of other ingredients in Chinese herbal formulations could also affect the pharmacokinetics of SW. There were significant differences in PK parameters between rats with CFA-induced arthritis and normal rats orally administered with Huo Luo Xiao Ling Dan (HLXLD), the AUC 0-t, AUC0–∞ and Cmax of the three secoiridoid glycosides (gentiopicroside, swertiamarin, sweroside) decreased to varying degrees after oral administration of HLXLD as compared with the normal control group, while the clearance of these glycosides increased significantly. The results indicate that the three components were are poorly absorbed and rapidly metabolized in the RA model. Meanwhile, the three secoiridoid glycosides in the HLXLD group had a longer Tmax compared with a single-herb extract of GME, suggesting slower absorption and distribution processes in the HLXLD group [19].
Mean plasma concentration-time curves of SW in rats after oral administration of three different doses of Swertia cincta solution showed that the AUC increased in three different ranges with the increase in the orally administered dose [20]. Studies have shown that there are significant gender differences in the pharmacokinetic properties of SW in rats. The Cmax, AUC0-t, AUC0–∞ and t1/2 values were significantly higher in female rats than in male rats after oral administration of SW(1.9 mg/kg), which indicated that SW is absorbed to a greater extent and eliminated more slowly in females compared with males [21].
After a single oral administration of 4 mL/kg of the Swertia pseudochinensis extract to rats, the concentration-time curve showed double peaks distribution of SW in rat plasma, which may be related to such factors as hepatic and intestinal circulation and the physicochemical nature of the compound. The excretion data indicate that six analytes, including SW, are excreted primarily in the urine. The excretion of SW and the other five analytes via d by bile, urine, and feces was very limited, suggesting that these analytes were mainly excreted as metabolite forms [22]. Metabolic analyses in Wistar rats after oral administration of SW showed that SW was initially hydrolyzed by β-glycosidase in the intestine and due to the instability of the aglycone it spontaneously reacted with ammonia to form gentianine. Gntianine was further converted to the nitrogen-containing metabolite gentiandiol in the liver and ultimately excreted through urine [23]. the endophyte could convert swertiamarin into a range of active metabolites, among which the metabolite M01 ((gentianine) has anti-diabetic potential due to its nitrogen-substituted structure [24,25].In addition, the aglycone of SW hydrolyzed by bacterial β-glucosidase was readily converted to erythrocentaurin(ECR). Some ECR is further reduced to 3,4-dihydro-5-(hydroxymethyl) isochroman-1-one (HMIO) by both liver and intestinal bacteria. Therefore, SW can be metabolized to dihydroisocoumarin and alkaloid compounds in vivo to exert pharmacological effects [26,27]. The Cmax and t1/2 of SW are very short, indicating that it is rapidly metabolized in vivo, and its oral bioavailability is between 6.2–8.0 %. Further studies showed that a total of 6 metabolites were isolated and identified in serum, urine, bile, and feces of SD male rats after oral administration of SW, and all metabolites were classified into the cleavage products of de-hydroxylation of aglycones, the isomerization product after dehydration of aglycones, the aglycon heterocyclic product, which first formed aglycones before the next metabolism [28]. Another study showed that 49 metabolites altogether including archetype compound of SW, were found in SD male rats following oral administration. and the in vivo biotransformation process mainly involved phases I reactions such as reduction, dehydration, hydroxylation, and phase II reactions such as sulphonation and N-acetylcysteine (NAC) formation. as well as drug metabolic cluster (DMC) centered on gentianine or other com-pounds were also detected [29].
3. Pharmacological Effects
3.1. Hepatoprotective Effects
An earlier study showed that, in a rat model of D-galactosamine-induced acute liver injury, SW (100 and 200mg/kg p.o.) treatment for 8 days attenuated pathological morphological changes such as liver tissue necrosis and bile duct proliferation caused by hepatic injury, restored the biochemical indexes towards normal levels, thus alleviating D-GalN caused hepatotoxicity, and enhanced the levels of antioxidant enzymes (CAT, SOD, GSH) and decreased levels of TBARS to exert hepatoprotective effects in rats [30]. SW obviously attenuated the inflammatory response of CCl4-induced liver injury in rats. as evidenced by a significant reduction in MPO activity, iNOS activity, NO levels, and the content of pro-inflammatory factors IL-1β, TNF-α, and IL-6 in the liver after SW treatment, as well as remarkably repressing hepatic oxidative stress, inducing the expression of hepatic detoxification enzymes CYPs, hepatic efflux transporter proteins (Bsep, Mrp2, Mrp3, Mrp40) and PDZK1. In addition, co-treatment of SW with CCl4 notably up-regulated the expression of Nrf2, HO-1, and NQO1. These results suggested that the protective effect of SW against CCl4-induced liver injury might be linked with the activation of the Nrf2/HO-1 pathway [31]. In CCl4-induced liver injury mice, SW supplementation exhibited lower liver weight and liver index, mitigating cell degeneration, inflammatory cell infiltration, and collagenous fibers deposition, mice treated with SW significantly reversed the elevation of ALT, AST, MDA, and Hyp levels induced by CCl4, multi-omics analysis. Furthermore, multi-omics analysis revealed that SW could ameliorate CCl4-induced liver toxicity through regulating gut microbiota, and its metabolites [32]. SW also attenuated carbon tetrachloride-induced apoptosis in rat hepatocytes and hepatic stellate cells activation via increasing Bcl-2 levels and inhibiting the expression and activation of caspase-3, and this anti-apoptotic effect may be related to the regulation of the PI3K/Akt signaling pathway [33]. In addition, SW ameliorated nicotine-induced damage in SD rats by regulating oxidative stress [34].
SW is an iridoid glycoside compound that is unstable to heat and prone to decomposition and transformation [35]. The heat-transformed products (HTPS) of SW were mainly responsible for hepatoprotective effects. Sweritranslactone D, a novel secoiridoid dimer isolated from the HTPS, exhibited more potent activity to alleviate liver injury than N-acetyl-L-cysteine in the L-O2 AP-induced cell model [36]. Another study demonstrated that protective effects of SW or HTPS against acetaminophen (APAP)-induced hepatotoxicity in vivo and in vitro, the treatment of SW or HTPS alleviated apoptosis in APAP-induced L-O2 cells and significantly reduced the liver injury index histological abnormalities in APAP-induced mice. Furthermore, SW or HTPS improved APAP-induced liver damage by inhibiting inflammatory reactions and oxidative stress, regulating Nrf-2/NF-κB signaling pathway [37].
Liver fibrosis is an abnormal wound-healing response caused by many types of acute and chronic liver injury, which may further progress to cirrhosis, liver failure, and liver cancer [38]. In vitro, SW significantly inhibited Ang II-induced primary rat hepatic stellate cells (HSCs) proliferation and activation, down-regulating TGF-β1 mRNA level. In vivo, SW significantly improved DMN-induced liver function injury, and ameliorated liver fibrosis by reducing collagen deposition, inhibiting α-smooth muscle actin(α-SMA) expression and hydroxyproline (Hyp) content in rat liver. This anti-hepatic fibrosis effect is mainly mediated by modulation of the RAS including down-regulation of Ang II and AT1R expression and inhibition of extracellular signal-regulated kinase (ERK) and c-jun phosphorylation [39].
Nonalcoholic fatty liver disease (NAFLD) is one of the most common chronic liver diseases, with a worldwide prevalence of up to 25% [40]. In fructose-fed mice, administrated with swertiamarin (25, 50, and 100 mg/kg) significantly decreased serum TG, GLU, UA, AST and ALT levels, SW ameliorated fructose-induced liver injury by inhibiting hepatic ballooning degeneration and fatty deposition, and reduced liver tissue TG levels. Further studies have shown that SW could attenuate NAFLD and metabolic alterations in fructose-fed mice by inhibiting hepatic inflammatory responses and XO activity, enhancing the antioxidant defense system, reducing hepatic steatosis, and down-regulating the expression of lipogenesis-controlling factors SREBP-1, FAS, and ACC1 [41]. In OA-induced insulin resistance HepG2 cells, SW attenuated hepatic glycemic load, lipid accumulation, ROS, and insulin resistance by targeting AMPK and PPAR-α, modulating fat metabolic enzymes such as FAS, ACC-1, PEPCK, and the expression levels of insulin signal proteins as ser-307 IRS1, IR-β, PI(3)K, p-AKT. Which provides a potential source of drugs for the treatment of hepatic steatosis [42].
Cholestasis is a pathological state caused by impaired bile secretion and excretion, prolonged cholestasis can also lead to liver failure and cirrhosis, increasing the risk of liver fibrosis and liver cancer [43,44]. Metabolomics analyses in the ANIT-induced cholestasis rat model revealed that SW altered multiple metabolomic pathways related to cholestasis, including primary bile acid synthesis, retinol metabolism, glycerophospholipid metabolism, and sphingolipid metabolism. SW also reversed changes in multiple protein targets, such as CYP7A1, BSEP, NTCP, SHP, and MRP2, mainly focusing on the farnesoid X receptor (FXR). These results suggested that SW can mitigate ANIT-induced cholestasis by activating FXR and bile acid excretion pathways [45]. In the ANIT-induced rat model of cholestatic hepatitis, the total iridoid and xanthone extract from Swertia mussotii Franch (TIXS) promoted bile secretion and lowered the activities of the serum enzymes ALT, AST, and ALP, as well as the levels of the bilirubins TBIL, DBIL, and UCBIL, thus exerting a therapeutic effect on cholestatic hepatitis. Swertiamarin and swertianolin are the active constituents of TIXS [46]. Bile duct ligation (BDL) rats administrated with swertiamarin showed low levels of ALT, AST, TNF-α, IL-1, IL-6, and SW reduced the toxic bile salt concentrations in the serum of cholestatic rats, including chenodeoxycholic acid (CDCA) and deoxycholic acid (DCA) [47]. These studies have elucidated the protective role of SW in cholestasis.
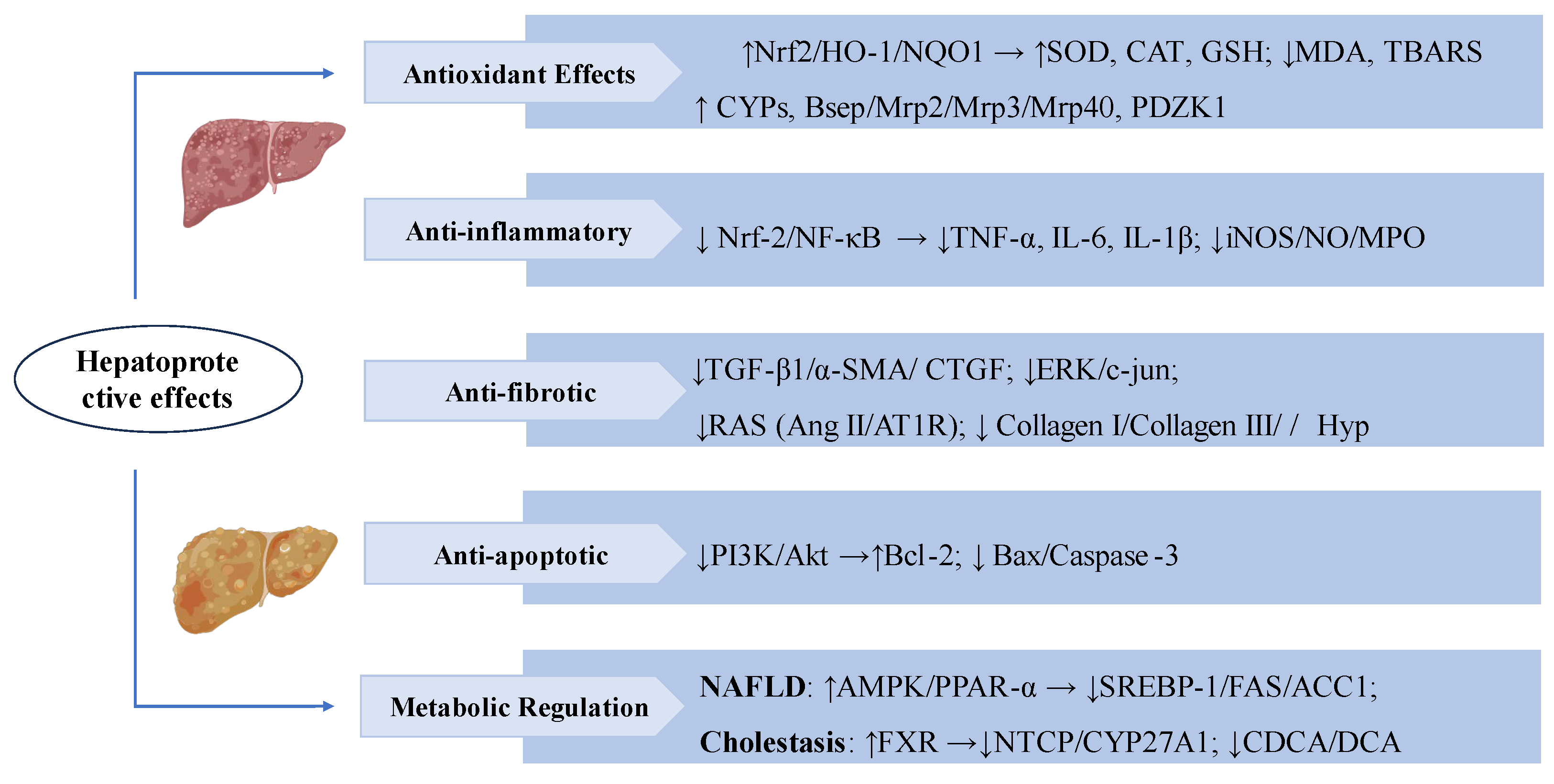
Figure 2.
Swertiamarin exerts multifaceted hepatoprotective effects by targeting key molecular pathways. (1) Antioxidant: SW activates the Nrf2/HO-1/NQO1 pathway to enhance antioxidant enzymes (SOD, CAT, GSH) and reduce oxidative stress markers (MDA, TBARS). It upregulates cytochrome P450 (CYPs) and bile transport proteins, promoting toxin metabolism and excretion. (2) Anti-inflammatory: SW suppresses Nrf-2/NF-κB signaling pathway, thereby decreasing the release of pro-inflammatory cytokines (TNF-α, IL-6, IL-1β) and reducing the expression of inflammatory mediators (iNOS/NO/MPO). (3) Anti-fibrotic: SW inhibits the TGF-β1/α-SMA/CTGF signaling cascade, leading to reduced collagen deposition (Collagen I/III) and Hyp level. And blocks the RAS system (Ang II/AT1R) and ERK/c-jun signaling, thereby slowing the progression of hepatic fibrosis. (4) Anti-apoptotic: SW modulates the PI3K/Akt pathway to upregulate the anti-apoptotic protein Bcl-2 while downregulating pro-apoptotic factors (Bax/Caspase-3). (5) Metabolic regulation: SW improves NAFLD via AMPK/PPAR-α/SREBP-1 and alleviates cholestasis by activating FXR, inhibiting NTCP/CYP27A1 and toxic bile acids (CDCA/DCA).
Figure 2.
Swertiamarin exerts multifaceted hepatoprotective effects by targeting key molecular pathways. (1) Antioxidant: SW activates the Nrf2/HO-1/NQO1 pathway to enhance antioxidant enzymes (SOD, CAT, GSH) and reduce oxidative stress markers (MDA, TBARS). It upregulates cytochrome P450 (CYPs) and bile transport proteins, promoting toxin metabolism and excretion. (2) Anti-inflammatory: SW suppresses Nrf-2/NF-κB signaling pathway, thereby decreasing the release of pro-inflammatory cytokines (TNF-α, IL-6, IL-1β) and reducing the expression of inflammatory mediators (iNOS/NO/MPO). (3) Anti-fibrotic: SW inhibits the TGF-β1/α-SMA/CTGF signaling cascade, leading to reduced collagen deposition (Collagen I/III) and Hyp level. And blocks the RAS system (Ang II/AT1R) and ERK/c-jun signaling, thereby slowing the progression of hepatic fibrosis. (4) Anti-apoptotic: SW modulates the PI3K/Akt pathway to upregulate the anti-apoptotic protein Bcl-2 while downregulating pro-apoptotic factors (Bax/Caspase-3). (5) Metabolic regulation: SW improves NAFLD via AMPK/PPAR-α/SREBP-1 and alleviates cholestasis by activating FXR, inhibiting NTCP/CYP27A1 and toxic bile acids (CDCA/DCA).
3.2. Anti-Diabetic Effects
Diabetes is a highly prevalent disease that was originally classified into three main categories: type 1 diabetes, type 2 diabetes, gestational diabetes mellitus, and specific types of diabetes due to other causes, It is estimated that type 2 diabetes accounts for 90–95% of all cases of diabetes [48]. Previous studies have demonstrated that the plant extract of Enicostemma littorale possesses hypoglycemic effects, which were mainly attributed to it containing the active ingredient SW. SW and its derivatives can exert its anti-diabetic effects through modulation of multiple targets, including adiponectin, PPARγ, GLUT4, GLUT2, Phosphoenolpyruvate carboxykinase (PEPCK), Glucokinase (GK), HMG-CoA Reductase (HMGR), Acetyl-CoA Carboxylase, 5-HT2 [49], and other molecular targets [11]. In vivo, the combination of SW and quercetin (CSQ) significantly reduced the levels of fasting blood glucose, TG, TC, LDL in STZ-induced T2DM Wistar rats, improved lipid metabolism abnormalities, recovered insulin secretion and regulated carbohydrate metabolic enzymes, CSQ treatment provided antioxidant effect by increasing the levels of serum SOD, CAT, GSH, GPx and decreasing the levels of lipid peroxide, and probably increased the regeneration of the pancreatic islets and the secretion of insulin in STZ-induced diabetes [50]. In nicotinamide-streptozotocin (NA-STZ)-induced diabetic rats, SM inhibited HMG-CoA reductase activity in dyslipidemic conditions, improved insulin sensitivity, and modulated carbohydrate and fat metabolism by modulating PPAR-γ and other transcription factors. SW could be an effective therapeutic agent for TIIDM [51]. SW increased glucose consumption by activating the PI3K/AKT pathway in HepG2 cells, thereby ameliorating dexamethasone-induced insulin resistance and exerting a hypoglycemic effect [52]. SW treatment for 28 days significantly reduced serum urea and creatinine levels, restored glucose metabolizing enzymes, promoted islet regeneration, and ameliorated diabetes-induced pancreatic, cardiac, hepatic, and renal lesions in STZ-induced diabetic rats [53].
As swertiamarin has a very short plasma half-life, the anti-diabetic effect of swertiamarin may be related to its active metabolite, gentianine. Treatment with gentianine induced adipogenesis and differentiation by upregulating the gene expression of PPAR-γ, GLUT-4, and adiponectin. Adiponectin mRNA expression was also significantly increased by swertiamarin treatment, thereby enhancing insulin sensitivity and exerting anti-diabetic effects [24]. In streptozotocin-induced diabetic rats fed a high-fat diet, the treatment with SW significantly decreased the level of glucose to 142.8 mg/dl and also reduced the level of serum TG and cholesterol. Following treatment with SW significantly ameliorated renal function and renal fibrosis in rats by preventing hypertrophy in the kidneys and decreasing the levels of kidney index, blood creatinine, and blood uric acid. SW mitigates the levels of AGEs in the serum and kidney in diabetic rats. Additionally, it more remarkably reduced MDA levels in the serum and kidneys of rats, indicating that it effectively alleviates oxidative stress induced by hyperglycemia and AGEs, the treatment with SW also could reduce the levels of advanced glycation end products (RAGE), p38 MAPK, and NADPH, blocking the binding of AGEs with RAGE [54]. Lomatogonium rotatum (LR) is an important herb containing the active ingredient SW, which prevented high-fat, high-glucose diet and STZ-induced diabetes in rats by altering the levels of serum metabolites such as vitamin B6, mevalonate-5P, and promoting the release of insulin and GLP-1 [55].
Diabetes can cause complications in multiple systems and organs, and the main risks include retinopathy, nephropathy (DN), neuropathy, and cardiovascular disease [56]. Numerous studies have indicated that SW also has an excellent curative effect on diabetic complications. SW treatment (50 mg/kg, i.p.) reduced serum triglyceride, cholesterol, and low-density lipoprotein (LDL) levels in diabetic animals and significantly increased the insulin sensitivity index, thereby improving the dyslipidaemic condition developed due to the insulin resistance [57]. SW exerted anti-glycosylation effects by inhibiting the formation of fructose-induced AGEs and the binding of AGEs with RAGE, SW inhibited the expression of inflammatory factors IL-6, TNF-α and IL-1β in MG-treated NRK-52E cells and reduced oxidative stress in renal cells by up-regulating the levels of Nrf-2 and HO-1, which reduced TGF-β activation and improved EMT in MG-induced NRK-52E. The inhibition of the RAGE/MAPK/TGF-β pathway may be the mechanism by which SW prevents DN in MG-induced kidney cells [58]. Oral administration of swertiamarin (50 mg/kg) significantly reduced serum urea and creatinine levels, and ameliorated glomerular injury and other parameters associated with the development of DN in type 1 diabetic rats [59]. Diabetic peripheral neuropathy (DPN) is one of the common complications of diabetes mellitus, with a reported incidence of 13-26% [60,61]. In a streptozotocin (STZ)-induced rat model of DPN, SW maintained inflammatory factor homeostasis by inhibiting the signaling pathway of the NOXS/ROS/NLRP3 inflammatory cascade response to reduce hyperalgesia and effectively protected the nerves for the treatment of DPN [62]. In Zucker fa/fa rats, treatment with SW significantly reduced serum glucose, serum cholesterol, TG, and NEFA levels, and lowered serum MMP-9, MMP-3 levels, and serum urea levels, and it had a significant protective effect against diabetes-induced cardiovascular complications such as atherosclerosis and nephropathy [63].
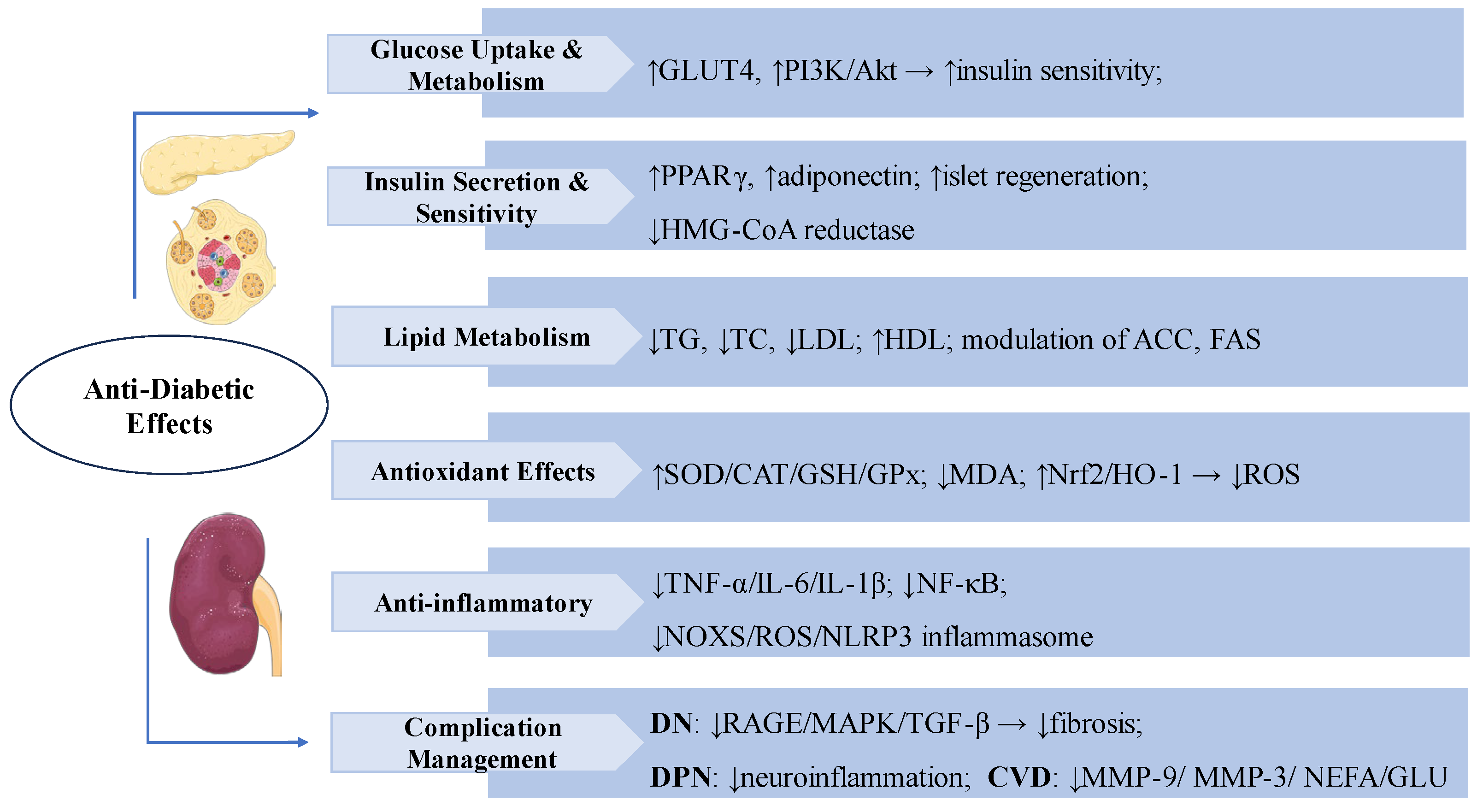
Figure 3.
Swertiamarin exerts comprehensive anti-diabetic effects through multiple mechanisms: (1) Glucose uptake and metabolism: SW enhances glucose metabolism by promoting GLUT4 expression and activating the PI3K/Akt pathway to improve insulin sensitivity, while stimulating pancreatic islet regeneration. (2) Insulin Secretion & Sensitivity: SW improves insulin secretion and sensitivity by upregulating PPARγ and adiponectin expression, and inhibiting HMG-CoA reductase activity. (3) Lipid Metabolism: SW modulates lipid metabolism by reducing TG, TC, and LDL levels while increasing HDL, and regulates key enzymes including ACC and FAS. (4) Antioxidant Effects: SW elevates SOD, CAT, GSH, and GPx while decreasing MDA through Nrf2/HO-1 pathway activation to reduce ROS. (5) Swertiamarin demonstrates anti-inflammatory effects by suppressing TNF-α, IL-6, IL-1β, NF-κB signaling, and the NOX4/ROS/NLRP3 inflammasome. (6) Complication management: SW manages diabetic complications by inhibiting RAGE/MAPK/TGF-β pathways, reducing neuroinflammation, MMP-9, MMP-3, NEFA, and glucose levels.
Figure 3.
Swertiamarin exerts comprehensive anti-diabetic effects through multiple mechanisms: (1) Glucose uptake and metabolism: SW enhances glucose metabolism by promoting GLUT4 expression and activating the PI3K/Akt pathway to improve insulin sensitivity, while stimulating pancreatic islet regeneration. (2) Insulin Secretion & Sensitivity: SW improves insulin secretion and sensitivity by upregulating PPARγ and adiponectin expression, and inhibiting HMG-CoA reductase activity. (3) Lipid Metabolism: SW modulates lipid metabolism by reducing TG, TC, and LDL levels while increasing HDL, and regulates key enzymes including ACC and FAS. (4) Antioxidant Effects: SW elevates SOD, CAT, GSH, and GPx while decreasing MDA through Nrf2/HO-1 pathway activation to reduce ROS. (5) Swertiamarin demonstrates anti-inflammatory effects by suppressing TNF-α, IL-6, IL-1β, NF-κB signaling, and the NOX4/ROS/NLRP3 inflammasome. (6) Complication management: SW manages diabetic complications by inhibiting RAGE/MAPK/TGF-β pathways, reducing neuroinflammation, MMP-9, MMP-3, NEFA, and glucose levels.
3.3. Neuroprotective Effects
The incidence of neurodegenerative diseases is increasing with the aging population. Recently, SW demonstrated neuroprotective effects in vivo and in vitro by rotenone-induced Parkinson’s disease (PD) mouse model and LPS-induced C6 glial cells, SW(10–100 μg/mL) treatment showed a significant reduction in pro-inflammatory cytokines IL-1β, IL-6, and TNF-α levels in LPS-induced C6 glial cell activation, SW also alleviated motor impairment caused by rotenone-induced neurotoxicity motor impairment, substantially inhibited the activation of microglial and astrocytes in the SN. SW might exert these neuroprotective effects by impeding the accumulation of α-syn in the striatum and substantia nigra, increasing TH immunoreactivity in the striatum and the number of TH+ neurons in the substantia nigra [64,65]. In the Caenorhabditis elegans model, SW enhanced neurotransmission by modulating nAChR and AChE activities, attenuated ROS levels, and increased antioxidant enzymes levels by up-regulating sod-3 and gst-4 levels, thereby ameliorating cholinergic dysfunction and exerting a protective effect against neurodegeneration [66]. In the 3-NP-induced Huntington’s disease (HD) rat model, administration of SW attenuated 3-NP-induced memory deficits, behavioral abnormalities, and biochemical parameters alterations, partially reduced ChE levels in the cerebral cortex and hippocampus of HD rats, and significantly ameliorated mitochondrial complexes enzyme activities and oxidative damage in various brain regions. Therefore, it may be an effective neuroprotective agent for HD and other degenerative diseases [67].
Moreover, SW has significant anti-inflammatory activity, which has a potential protective role on neuro-inflammation. In BV-2 cells treated with LPS, SW (10, 25, 50 μg/mL) differentially inhibited the inflammatory cytokines secretion of IL-1β, IL-6, IL-18, and TNF-α in BV-2 cells. The proteomics analysis results suggested that the potential bio-processings regulated by SW were mainly involved in the cellular response to carbon monoxide, strand displacement, palmitoleoyltransferase activity, D2 dopamine receptor binding, and RNA polymerase II transcription cofactor activity [68]. SW also inhibited apoptosis and ROS production via the TLR4/PARP1/NF-κB pathway, thereby protecting human neuronal SH-SY5Y cells from OGDR-stimulated damage [69]. In α-syn-expressing worms, the expression of fat-5 and fat-7 lipid levels were significantly reduced after SW treatment, thereby reducing α-syn deposition. At the same time, it increased the mitochondrial viability, decreased the levels of ROS, reduced apoptosis, significantly elevated DA levels, and modulated dopamine-dependent behavior in NI 5901 worms. In the BZ555 worms induced by the neurotoxin 6-OHDA, SW remarkably protected against the neurotoxin-induced dopaminergic neuronal degeneration. The neuroprotective effect of SW may upregulate the expression of SKN-1 and GST-4 through the MAPK pathway [70]. SW may also exert antidepressant effects by modulating 5-HT2 receptors [49].
Stroke is one of the cerebrovascular diseases with high global morbidity and mortality, thrombolysis is an effective treatment for ischemic stroke, while cerebral ischemia-reperfusion injury(CIRI) is a major complication of treatment [71]. In mice middle cerebral artery occlusion (MCAO) model, at 24 h after reperfusion, pre-treatment of SW((25, 100, or 400 mg/kg)) dose-dependently markedly reduced infarct volume, reversed the change of pathological produced by CIRI to some extent and significantly inhibited the I/R-induced neuronal apoptosis by increased the levels of the anti-apoptotic protein Bcl-2 and decreased Bax levels. SW also protected against I/R-induced oxidative stress damage by reducing MDA levels and upregulating the antioxidant enzymes (GSH-PX, SOD, and CAT) activities in vivo. In addition, SW treatment decreased cell death and intracellular ROS levels induced by OGD/R in primary cultured hippocampal neurons, and it promoted Nrf2 nuclear translocation from Keap1-Nrf2 complex, increased the expression of Nrf2, HO-1, and NQO1 in the mouse MACO model and the primary hippocampal neuronal cell OGD/R model. These results illustrated that the neuroprotective effects of SW against I/R injury may depend on anti-oxidative stress mediated by activating the Nrf2 pathway, it may be a promising protective agent against ischemic brain damage [72].
In the pilocarpine (PILO)-treated mice, SW pretreatment significantly delayed the first convulsion and reduced the incidence of persistent status epilepticus and mortality, as well as reduced neuronal loss and neuronal damage in hippocampal CA1 and CA3 regions, and inhibited astrocyte activation and inflammatory responses [73]. SW also stimulated GABA receptors and significantly inhibited PTZ-induced convulsion seizures in albino mice [74], which has the potential to prevent and attenuate epileptic seizures.
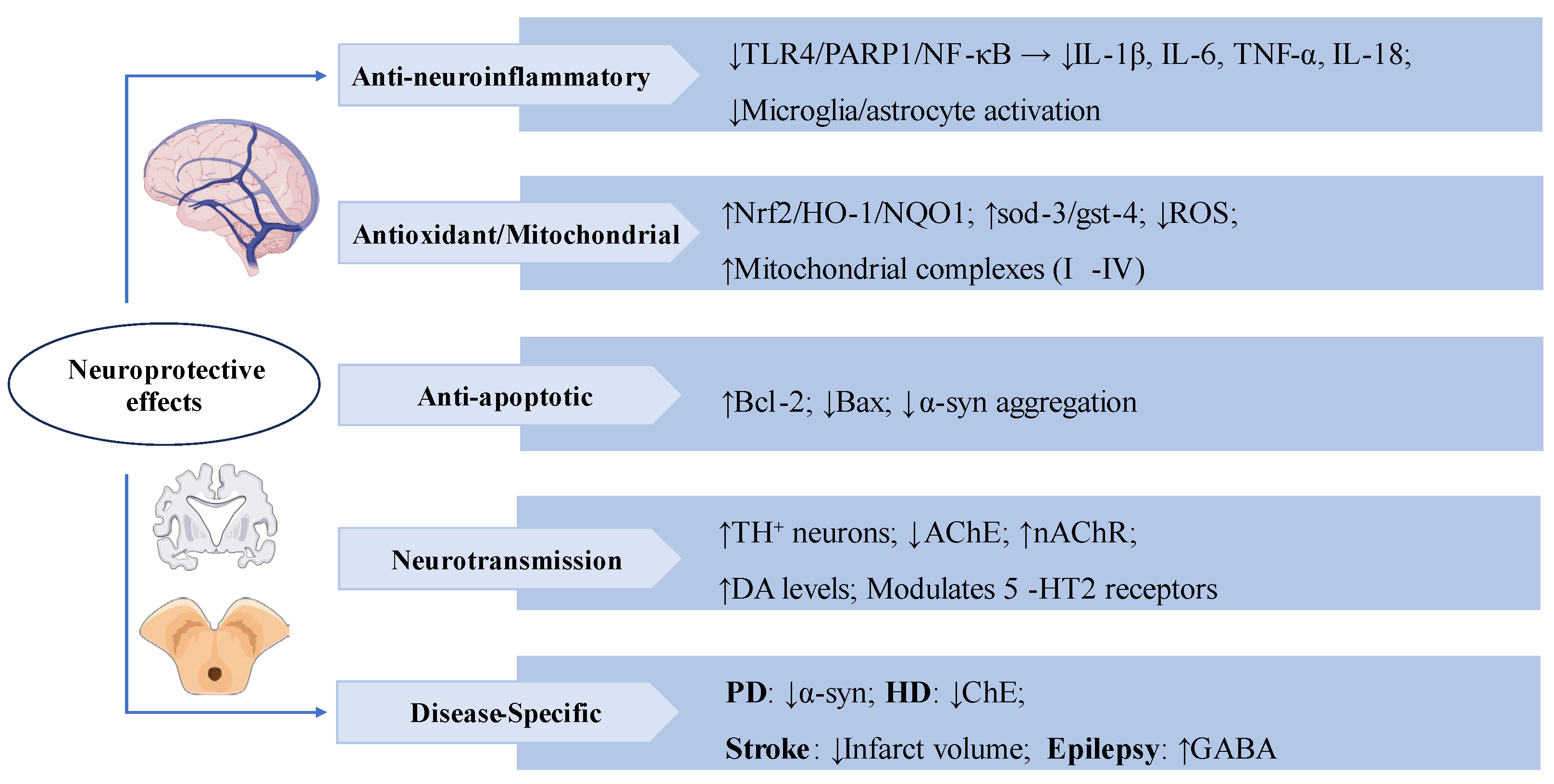
Figure 4.
Swertiamarin exerts multi-target neuroprotective effects through anti-neuroinflammatory, antioxidant, and neuromodulatory mechanisms. (1) Anti-neuroinflammatory: SW inhibits TLR4/PARP1/NF-κB signaling to reduce pro-inflammatory cytokines and suppresses microglia/astrocyte activation. (2) Antioxidant/Mitochondrial: SW demonstrates potent antioxidant activity by activating the Nrf2/HO-1/NQO1 pathway, upregulating sod-3/gst-4, reducing ROS, and enhancing mitochondrial complexes I-IV. (3) Anti-apoptotic: SW upregulates Bcl-2 and downregulates Bax, while inhibiting α-synuclein aggregation. (4) Neurotransmission: SW modulates neurotransmission by increasing tyrosine hydroxylase-positive (TH+) neurons and DA levels, inhibiting AChE and nAChRs, and regulating 5-HT2 receptors. (5) Disease-specific benefits include: reducing α-synuclein in Parkinson’s disease (PD), inhibiting cholinesterase (ChE) in Huntington’s disease (HD), decreasing infarct volume in stroke, and enhancing GABA activity in epilepsy.
Figure 4.
Swertiamarin exerts multi-target neuroprotective effects through anti-neuroinflammatory, antioxidant, and neuromodulatory mechanisms. (1) Anti-neuroinflammatory: SW inhibits TLR4/PARP1/NF-κB signaling to reduce pro-inflammatory cytokines and suppresses microglia/astrocyte activation. (2) Antioxidant/Mitochondrial: SW demonstrates potent antioxidant activity by activating the Nrf2/HO-1/NQO1 pathway, upregulating sod-3/gst-4, reducing ROS, and enhancing mitochondrial complexes I-IV. (3) Anti-apoptotic: SW upregulates Bcl-2 and downregulates Bax, while inhibiting α-synuclein aggregation. (4) Neurotransmission: SW modulates neurotransmission by increasing tyrosine hydroxylase-positive (TH+) neurons and DA levels, inhibiting AChE and nAChRs, and regulating 5-HT2 receptors. (5) Disease-specific benefits include: reducing α-synuclein in Parkinson’s disease (PD), inhibiting cholinesterase (ChE) in Huntington’s disease (HD), decreasing infarct volume in stroke, and enhancing GABA activity in epilepsy.
3.4. Regulation of Lipid Metabolism
Disorders of lipid metabolism are one of the risk factors for many cardiovascular diseases such as hyperlipidemia, atherosclerosis, and coronary heart disease [75,76]. In high cholesterol diet-induced hypercholesterolemic rats. Administration of SW (50 and 75 mg/kg, p.o.) significantly reduced serum TC, TG, LDL, VLDL levels and inhibited the activity HMG-CoA reductase enzyme in rats, as well as increased fecal excretion of the bile acid and total sterols by enhancing the activity of the 17-α-hydroxylase enzyme, the may be one of the mechanisms of its anti-hyperlipidaemic effect [77]. In P-407-induced hyperlipidaemic rats, Administration of SW (50 mg/kg, oral) after 15 and 24 h significantly reduced the ratios of LDL/HDL serum cholesterol [78]. Another study demonstrated that SW also could decrease fat storage in vivo, knockdown of kat-1 inhibited the lipid-lowering effect of SW, whereas SW treatment significantly increased kat-1 expression. These results indicate that SW regulates lipid homeostasis and exerts lipid-lowering effects dependent on kat-1 [79]. These results demonstrated the potential activity of SW in lipid-lowering and anti-atherosclerosis. The aqueous root extracts containing SW inhibited PDGF-induced activation of ERK1/2 and proliferation of rat aortic smooth muscle cells (RASMC), which provide new drug candidates for the treatment of atherosclerosis [80].
Obesity is one of the global public health problems, and it is closely related to diabetes and cardiovascular disease [81]. In high-fat diet (HFD)- induced obese C57BL/6 mice, SW attenuated HFD-induced weight gain, glucose intolerance, and obesity-related insulin resistance by enhanced insulin signaling. Treatment with SW notably increased lipolysis and reduced adipocyte hypertrophy and macrophage infiltration in the epididymal white adipose tissue (eWAT). SW supplementation decreased the expression of inflammatory cytokines (TNF-α and IL-1β) and chemokines (Ccl2 and Ccl5) in LPS-stimulated macrophages and in HFD-fed mice, which also further indicated SW improved obesity-associated chronic inflammation and hepatic steatosis by suppressing activation of the p38 MAPK and NF-κB pathways [82]. Another study showed that SW treatment for 10 weeks significantly reduced body weight and enhanced energy expenditure by promoting BAT activation and fat browning in diet-induced obese mice, as well as improved the irregular fiber structure and excess lipid deposition and oxidative metabolism by increasing fatty acid oxidation in skeletal muscle [83]. The above studies highlight the potential clinical utility of SW in the prevention of obesity and related metabolic disorders.
Figure 5.
Swertiamarin demonstrates comprehensive lipid-modulating and metabolic regulatory effects through multiple mechanisms. (1) Anti-hypercholesterolemic: SW inhibits HMG-CoA reductase to reduce total cholesterol (TC), triglycerides (TG), and LDL/VLDL levels, while simultaneously enhancing bile acid excretion through 17-α-hydroxylase activation. (2) Regulation of lipid homeostasis: SW reduces lipid accumulation by elevating the kat-1 level. (3) SW reduces pro-inflammatory mediators and chemokines by suppressing p38 MAPK/NF-κB pathways. (4) Anti-obesity and metabolic enhancement: SW increases the activation of BAT to promote thermogenesis and ameliorates glucose intolerance and obesity-related insulin resistance by enhancing insulin signaling.
Figure 5.
Swertiamarin demonstrates comprehensive lipid-modulating and metabolic regulatory effects through multiple mechanisms. (1) Anti-hypercholesterolemic: SW inhibits HMG-CoA reductase to reduce total cholesterol (TC), triglycerides (TG), and LDL/VLDL levels, while simultaneously enhancing bile acid excretion through 17-α-hydroxylase activation. (2) Regulation of lipid homeostasis: SW reduces lipid accumulation by elevating the kat-1 level. (3) SW reduces pro-inflammatory mediators and chemokines by suppressing p38 MAPK/NF-κB pathways. (4) Anti-obesity and metabolic enhancement: SW increases the activation of BAT to promote thermogenesis and ameliorates glucose intolerance and obesity-related insulin resistance by enhancing insulin signaling.
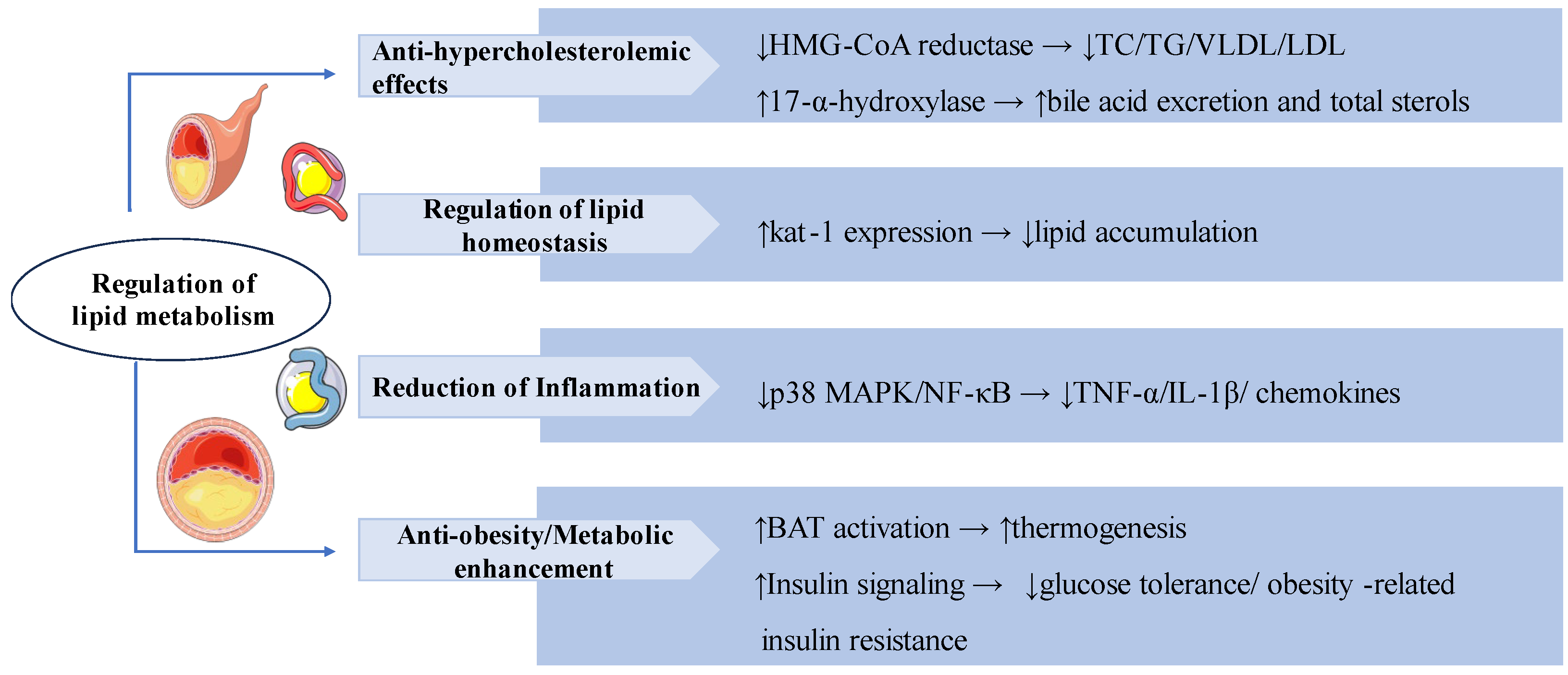
3.5. Anti-Inflammatory and Immunomodulation
In SRBC-immunized mice, SW treatment (2, 5, and 10 mg/kg b.w.) significantly increased antibody titer, plaque-forming cells, and immune organ weights, and attenuated delayed-type hypersensitivity (DTH). In vitro, SW significantly inhibited the release of free radicals in PHA-induced neutrophils and ameliorated the expression of pro-inflammatory factors in LPS-induced macrophages, suggesting that SW exerted its anti-inflammatory effects by modulating humoral and cell-mediated immune responses, inhibiting pro-inflammatory mediators (Th 1) and inducing anti-inflammatory mediators (Th 2) [84]. As a potential natural AKT inhibitor, SW could directly target the structural domain of AKT-PH to inhibit AKT activation and down-regulate the phosphorylation level of AKT, which further inhibits the production of downstream inflammatory molecules, thus presenting significant anti-inflammatory activity in cells and animal models of acute lung injury [85].
In the Plasmodium berghei-infected Swiss albino mouse model, oral administration of SW (200 mg/kg) reduced parasite load and the severity of malaria infection in mice. SW also stimulated the production of immunomodulatory mediators in vivo and modulated the levels of IFN-γ, IL-10, and TNF-α to exert anti-malarial protection, and could be used as an immunomodulatory agent for the combination treatment of malaria infection [86]. Another study has shown that rutin and swertiamarin have dose-dependent chemosuppressant effects on early malaria infection, and they possess potential antimalarial activity in a 1:1 ratio combination [87].
Chlorogenic acid (CA) and SW as key compounds in Honeysuckle Extract Preparation (HEP), which synergistically inhibited pro-inflammatory cytokines IL-1 and IL-6 and PGE2 production via targeting the PI3 K-AKT and p38 MAPK pathways thereby alleviating LPS-induced fever [88]. In a study of the anti-inflammatory activity of four Gentiana species, SW as one of the key chemical components showed significant inflammatory inhibition in LPS-induced RAW264.7 cells [3]. SW also can exert antiedematogenic effects through anti-inflammatory and free radical scavenging in rat paw edema models [89].
Figure 6.
Swertiamarin exhibits broad immunomodulatory and anti-inflammatory properties through multiple mechanisms. (1) Immunomodulation: SW enhances humoral and cellular immunity by increasing antibody titers, plaque-forming cells, immune organ weights, and reducing delayed-type hypersensitivity (DTH) responses and Th1/Th2 balance shifting. (2) Anti-inflammation: SW inhibits AKT-PH domain binding to suppress p-AKT activation, consequently reducing pro-inflammatory cytokines. And synergistically modulates both PI3K-AKT and p38 MAPK pathways to decrease IL-1, IL-6, and PGE2 production. (3) Anti-malarial protection: SW reduces parasite load and regulates cytokine profiles for antimalarial effects. (4) Free Radical Scavenging: SW reduces ROS in PHA-stimulated neutrophils and edema in paw models.
Figure 6.
Swertiamarin exhibits broad immunomodulatory and anti-inflammatory properties through multiple mechanisms. (1) Immunomodulation: SW enhances humoral and cellular immunity by increasing antibody titers, plaque-forming cells, immune organ weights, and reducing delayed-type hypersensitivity (DTH) responses and Th1/Th2 balance shifting. (2) Anti-inflammation: SW inhibits AKT-PH domain binding to suppress p-AKT activation, consequently reducing pro-inflammatory cytokines. And synergistically modulates both PI3K-AKT and p38 MAPK pathways to decrease IL-1, IL-6, and PGE2 production. (3) Anti-malarial protection: SW reduces parasite load and regulates cytokine profiles for antimalarial effects. (4) Free Radical Scavenging: SW reduces ROS in PHA-stimulated neutrophils and edema in paw models.
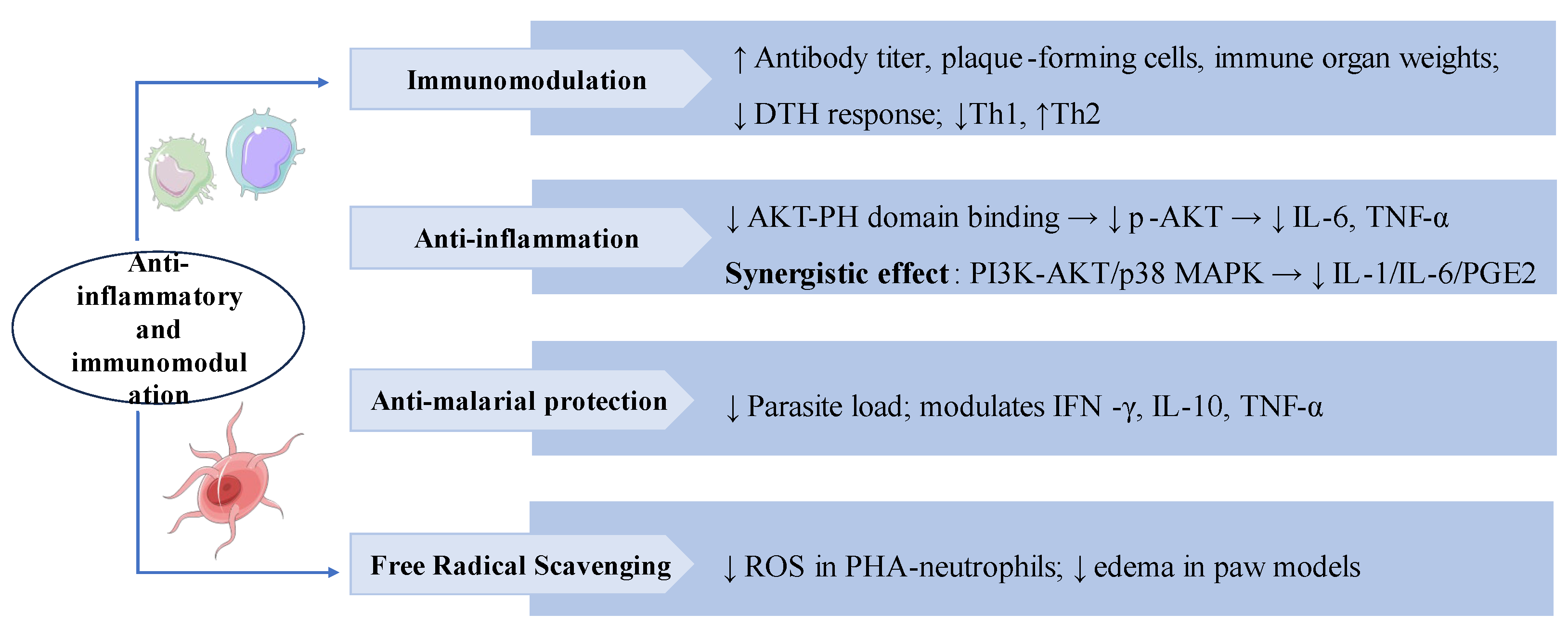
3.6. Anti-Tumor Effects
Studies have demonstrated that SW possesses anti-tumor activity. SW significantly inhibited the viability and invasion of HepG2 liver cancer cells and promoted apoptosis. In vivo, SW also notably suppressed the growth of SK-Hep-1 cells xenografted in nude mice. Bioinformatics analysis has identified PI3K, JUN, and STAT3 as predictive targets through which SW exerts its anti-tumor effects in hepatocellular carcinoma (HCC) [90]. Further investigations have shown that SW inhibited HepG2 and Huh7 liver cancer cells proliferation, migration, and invasion by downregulating FRAT1 and partially suppressing the FRAT1/Wnt/β-catenin signaling axis [91]. These data demonstrated the therapeutic potential of SW for HCC. SW also showed significant antiproliferative activity on the HEp-2 and HT-29 cancer cell lines [74]. Gentiana kurroo root extract blocked the G0/G1 cell cycle and protected DNA from oxidative damage, inducing apoptosis in a human pancreatic cancer cell line (Miapaca-2) by disrupting the mitochondrial membrane potential, with SW as one of the major components [92]. Swertiamarin B, a novel secoiridoid compound isolated from Swertia mussotii Franch, effectively inhibited the proliferation of gastric cancer cells [93]. In studies of the human peripheral blood mononuclear cells (PBMCs), the expression levels of PARP1, Bax/Bcl -2 were elevated after SW treatment suggesting that SW may activate apoptosis and necroptosis to eliminate malignant cells [94]. SW bonded strongly to both human serum albumin (HSA) and α-1 acid glycoprotein (AGP), inhibited proliferation, and induced apoptosis of the neuroblastoma (SK-N-AS) cell line [95].
Figure 7.
Swertiamarin exhibits broad anti-tumor activity across multiple cancer types. (1) HCC: SW inhibits tumor proliferation, migration, and invasion by suppressing the FRAT1/Wnt/β-catenin signaling axis. (2) Miapaca-2: SW induces G0/G1 cell cycle arrest, promotes apoptosis while protecting normal DNA integrity. (3) Gastric cancer cells: SW demonstrates significant anti-proliferative effects. (4) PBMCs: SW upregulates pro-apoptotic markers, including PARP1 and the Bax/Bcl-2 ratio. (5) Protein Binding: SW binds to HSA and AGP to inhibit proliferation and induce apoptosis in SK-N-AS cells..
Figure 7.
Swertiamarin exhibits broad anti-tumor activity across multiple cancer types. (1) HCC: SW inhibits tumor proliferation, migration, and invasion by suppressing the FRAT1/Wnt/β-catenin signaling axis. (2) Miapaca-2: SW induces G0/G1 cell cycle arrest, promotes apoptosis while protecting normal DNA integrity. (3) Gastric cancer cells: SW demonstrates significant anti-proliferative effects. (4) PBMCs: SW upregulates pro-apoptotic markers, including PARP1 and the Bax/Bcl-2 ratio. (5) Protein Binding: SW binds to HSA and AGP to inhibit proliferation and induce apoptosis in SK-N-AS cells..
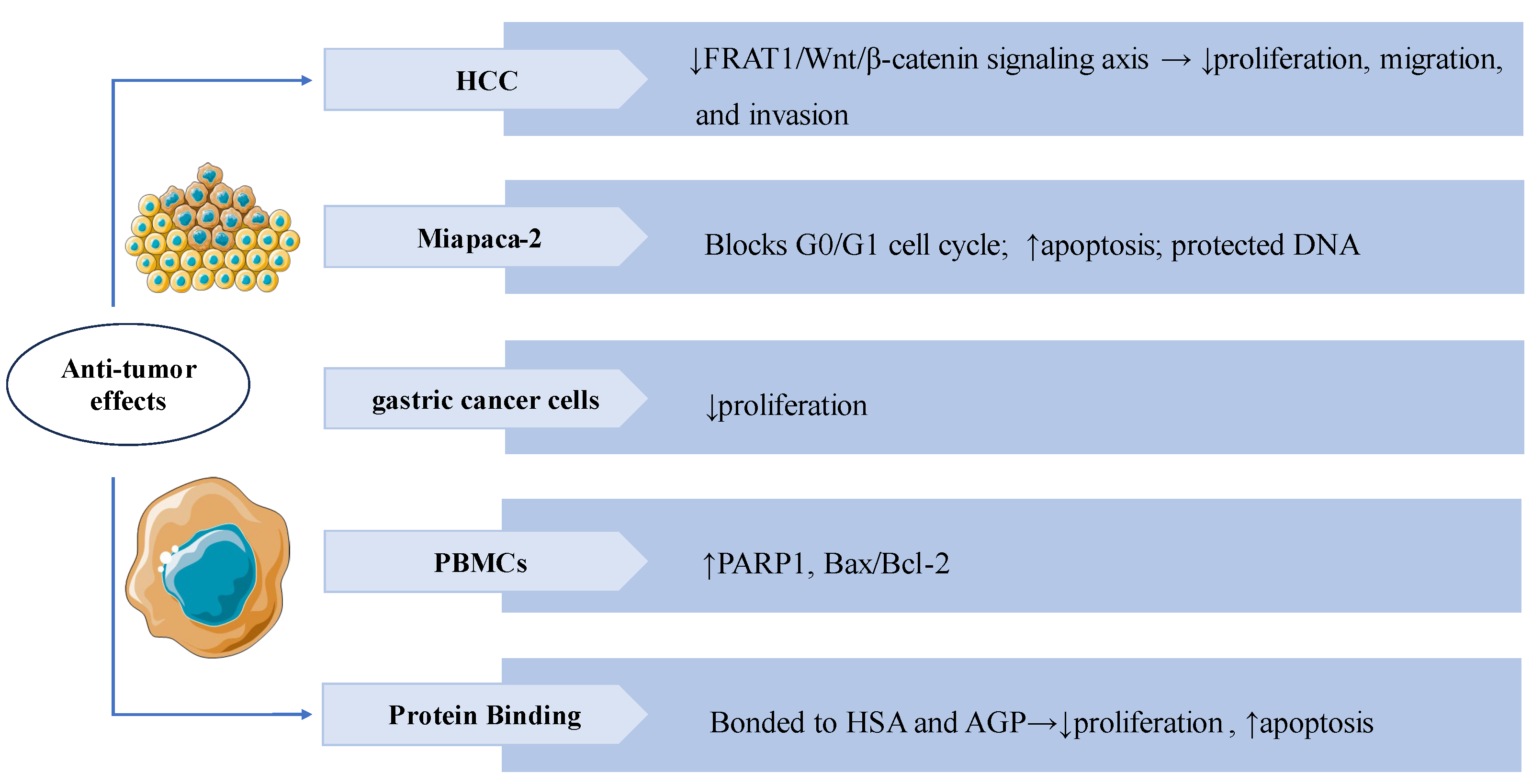
3.7. Bone and Joint Protective Effects
Rheumatoid arthritis (RA) is a chronic autoimmune disease characterized by synovial inflammation. [96]. SW also has a significant therapeutic effect on RA. In IL-1β induced AA-FLS cells (adjuvant-induced arthritic rat fibroblast-like synoviocytes), SW treatment with significantly inhibited proliferation and NO production, increased mRNA and protein expression levels of apoptosis mediator caspase 3 in a dose-dependent manner, while decreasing the levels of the inflammatory mediator iNOS, COX-2, PGE2, TNF-α, and IL-6, and also modulating the balance of MMPs/TIMPs, the treatment significantly suppressed the release of osteoclastogenic mediators RANKL and p38 MAPKα. These results indicated that SW can alleviate IL-1β-induced AA-FLS from joint destruction by exerting anti-inflammatory, pro-apoptotic, and anti-osteoclastogenic effects. [97]. Bone erosion is the main clinical manifestation of RA [98]. In vivo, treatment with SW significantly reversed pathological alterations caused by adjuvant (FCA) stimulation in the animals, including calcium and phosphorus levels, bone collagen levels, TRAP, ACP, and ALP levels. In vitro, SW significantly promoted the proliferation of osteoblast cells and the release of ALP, reduced the TRAP-positive cells in osteoclast cells, and modulated the levels of TNF-α, IL-6, IL-1β, MMPs, and NF-κB p65 release in osteoblast and osteoclast co-culture system, SW also downregulated expression of TRAP, RANKL, RANK levels and upregulated the expression of OPG levels in vivo and vitro. These results suggested that SW prevents bone erosion by regulating RANKL/RANK/OPG signaling to control the release of pro-inflammatory cytokines, and pro-angiogenic markers [99]. In adjuvant-induced arthritic animals, SW treatment modulated RBC, WBC, and ESR levels, reduced the release of acute phase proteins, lysosomal enzymes, protein-bound carbohydrates, and urinary degradation products, and elevated levels of the anti-inflammatory factors IL-4 and IL-10. SW treatment also significantly inhibited the release of NF-κB p65, p-IκBα, p-JAK2, and p-STAT3 levels on both arthritic animals and LPS-induced RAW 264.7 macrophage cells. These results indicated that the anti-arthritic effect of SW is closely related to the NF-κB/IκB and JAK2/STAT3 signaling [100]. The above studies also revealed that SW might have the potential to treat RA.
Figure 8.
Swertiamarin demonstrates comprehensive bone-protective and immunomodulatory effects through multi-target mechanisms. (1) Anti-inflammatory: SW suppresses pro-inflammatory cytokines (TNF-α, IL-6, IL-1β) and iNOS/COX-2/PGE2 production, and downregulates NF-κB/RB and JAK2/STAT3 signaling pathways while modulating MMPs/TIMPs balance. (2) Anti-osteoclastogenic: SW inhibits osteoclast formation by reducing RANKL/RANK signaling and TRAP/ACP activity. (3) Pro-Osteoblastic: SW promotes osteoblast growth and ALP expression. (4) Immune modulation and pro-apoptotic effect: SW modulates immune function by normalizing RBC/WBC counts and ESR levels while enhancing anti-inflammatory cytokines (IL-4/IL-10), and activates caspase-3 mediated apoptosis.
Figure 8.
Swertiamarin demonstrates comprehensive bone-protective and immunomodulatory effects through multi-target mechanisms. (1) Anti-inflammatory: SW suppresses pro-inflammatory cytokines (TNF-α, IL-6, IL-1β) and iNOS/COX-2/PGE2 production, and downregulates NF-κB/RB and JAK2/STAT3 signaling pathways while modulating MMPs/TIMPs balance. (2) Anti-osteoclastogenic: SW inhibits osteoclast formation by reducing RANKL/RANK signaling and TRAP/ACP activity. (3) Pro-Osteoblastic: SW promotes osteoblast growth and ALP expression. (4) Immune modulation and pro-apoptotic effect: SW modulates immune function by normalizing RBC/WBC counts and ESR levels while enhancing anti-inflammatory cytokines (IL-4/IL-10), and activates caspase-3 mediated apoptosis.
3.8. Prostate-Protective Effects
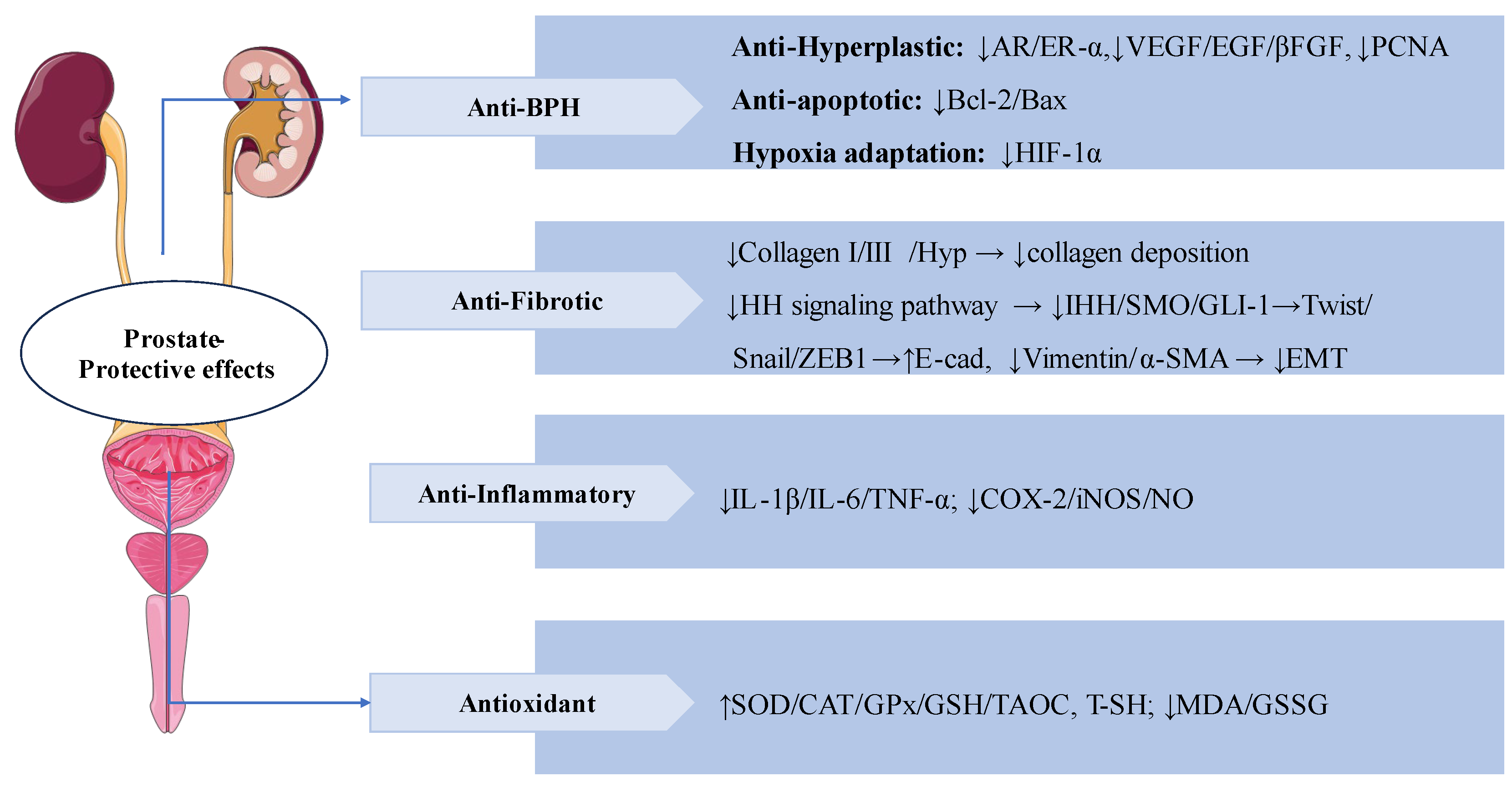
SW also has significant prostate-protective effects. Benign prostatic hyperplasia (BPH) is a common prostate disease in middle-aged and older men [101]. In testosterone-induced BPH rats, treatment with SW (16 mg/kg/d, p.o.) for 28 days significantly reduced PI levels and PACP activity, as well as ameliorated testosterone-induced morphological alterations such as collagen deposition, and epithelial cells expansion in the prostate. In addition, treatment with SW significantly alleviated testosterone-induced over-expressions of VEGF, EGF, βFGF, PCNA, AR and ER-α, HIF-1α, and reduced the ratio of Bcl-2/Bax. SW also obviously increased the levels of T-SH, GSH, COX-2, and SOD, and decreased the activity of GSSG and MDA. SW treatment down-regulated Twist, Vimentin, α-SMA levels, and up-regulated E-cadherin levels thereby attenuating prostatic EMT. These results suggest that SW has anti-prostatic hyperplasia, anti-inflammatory, and antioxidant effects [102]. In vivo, SW (32 and 16 mg/kg/d) relieved CS-induced prostatic collagen deposition and morphological changes by inhibiting the expression levels of Hyp, Col-I, and Col-III in rats. SW also significantly ameliorated CS-induced prostate local inflammation by decreasing the levels of pro-inflammatory mediators (IL-1β, IL-6, TNF-α and NO) and the activities of COX-2 and iNOS, as well as it also relived oxidative stress by elevating the activities of antioxidant enzymes (SOD, CAT, and GPx), increasing the levels of TAOC, T-SH and GSH, decreasing the contents of MDA and GSSG. In vitro, SW inhibited CS-induced RWPE-1 and WPMY-1 human prostate cells proliferation. SW also significantly reduced the expression levels of α-SMA, IHH, SMO, GLI-1, Snail, ZEB 1, and TGF-β1 while elevating the levels of E-cad in rat prostate tissues and human prostate cells, thereby inhibiting prostatic epithelial-mesenchymal transition(EMT) and the activation of HH signaling pathway to ameliorate CS-induced prostatic fibrosis [103].
Figure 9.
Swertiamarin demonstrates multi-target therapeutic effects for prostate disorders through four key mechanisms. (1) BPH: SW plays anti-hyperplastic action by modulating androgen/estrogen receptors (AR/ER-α) and growth factors (VEGF/EGF/βFGF) and cell proliferation markers (PCNA); And Balances apoptosis (Bcl-2/Bax ratio) and inhibits hypoxia adaptation (HIF-1α). (2) Anti-fibrotic protection: SW reduces collagen deposition and prevents EMT by HH pathway. (3) Anti-inflammatory and antioxidant: SW lowers pro-inflammatory mediators while enhancing antioxidant defenses and reducing oxidative stress.
Figure 9.
Swertiamarin demonstrates multi-target therapeutic effects for prostate disorders through four key mechanisms. (1) BPH: SW plays anti-hyperplastic action by modulating androgen/estrogen receptors (AR/ER-α) and growth factors (VEGF/EGF/βFGF) and cell proliferation markers (PCNA); And Balances apoptosis (Bcl-2/Bax ratio) and inhibits hypoxia adaptation (HIF-1α). (2) Anti-fibrotic protection: SW reduces collagen deposition and prevents EMT by HH pathway. (3) Anti-inflammatory and antioxidant: SW lowers pro-inflammatory mediators while enhancing antioxidant defenses and reducing oxidative stress.
3.9. Other Therapeutic Effects
Radiation-induced intestinal injury (RIII) is one of the intestinal complications caused by radiotherapy for abdominal or pelvic tumors [104]. A study showed that SW has significant protective effects against RIII both in vitro and in vivo. In radiation-induced cellular and mouse intestinal injury models, Prophylactic administration of SW attenuated radiation damage to cells by reducing ROS and superoxide anion levels. SW also increased the survival rate, improved the morphological injury of the small intestine, and reduced the relative abundance of primary bile acids in irradiated mice. Further research elucidated that SW can prevent RIII by reducing DNA damage and inhibiting the activation of the cGAS-STING pathway [105]. SW exhibited both peripheral and central antinociceptive activity dose-dependently in three different mice pain models [106]. Molecular docking studies showed SW also has good binding interactions with the cyclooxygenase-2 protein [107].
SW possessed significant anti-bacterial [108] and anti-fungal activity, including P. aeruginosa, S. aureus, and Trichoderma viride, the minimum inhibitory concentrations (MIC) of SW for S. typhi and K. pneumoniae was 1pg/mL [109,110]. In zebrafish infected with Salmonella typhi, SW significantly reduced bacterial load in zebrafish, attenuated the symptoms of infection such as spinal deformity, and reduced mortality, therefore it may be a promising anti-typhoid drug [111].
In human granulosa-luteal cells from insulin-resistant polycystic ovarian syndrome patients (PCOS - IR), administration of SW significantly reversed the expression of insulin resistance-related factors, including INSR-β, PI(3)K, p-Akt, PKC-β, PPARγ, IRS ( Ser307) and IGF systems, as well as modulated lipid metabolism-related genes, re-established steroidogenesis, restored 17β-HSD and 3 β-HSD enzyme activities and hormone levels, which can be used as an insulin sensitizer to alleviate PCOS-IR [112]. SW stimulated gastric emptying and gastrointestinal motility by inhibiting dopamine D2 receptor and may be a potential treatment for functional dyspepsia [113]. Swertiamarin down-regulated the expression levels of α-SMA, E-cadherin, LOX, COL5A2, and CTGF in TGF-β1-stimulated A549 cells and inhibited the epithelial-mesenchymal transition process exerting an anti-pulmonary fibrosis effect [114,115].
SW had significant cardioprotective effects. In isoproterenol-induced myocardial infarction (MI) rats, pre-treatment with SW notably alleviated myocardial fibrosis and inflammatory cell infiltration in MI rats. It also restored the levels of several cardiac marker enzymes, oxidative stress markers, antioxidant enzymes, Na+/K+ Potassium and Ca2+ATPases, and pro-inflammatory cytokines [116]. The aqueous extract of Enicostemma littorale (E. littorale) showed significant intervention in ISO-induced cardiac hypertrophy in rats by modulating the levels of key factors of cardiac metabolism such as glucose, protein, lipid metabolism, and cardiac enzyme activities, and one of the components in the aqueous extract, SW, may have an anti-hypertrophic potential [117].
4. Toxicology
The toxicity of SW has not been adequately and systematically studied, and the available studies suggest that it is relatively safe at appropriate doses, but more data are needed to fully assess its toxicity for long-term use. A toxicity study of SW in zebrafish showed low or no toxicity at low concentrations (40 µM), while high doses (243 µM) caused organ malformations and developmental abnormalities in zebrafish embryos, as well as significant changes in zebrafish liver enzymology and antioxidant activity [118].
5. Conclusions and Future Perspectives
The pharmacokinetic profile of SW in rats is characterized by rapid absorption, low bioavailability, rapid metabolism, and clearance. It is mainly metabolized in the intestinal tract and the liver, with its metabolites excreted through the urine. SW is affected by a number of factors such as gender, dosing combination, and dosage, and thus exhibits different pharmacokinetic profiles.
In addition, semi-synthetic analogues of SW have also become a new direction of research with the continuous advancement of research. Modifications to chemical structures can further enhance their biological activities and metabolic properties in specific aspects. Among several semi-synthetic SW analogues,these compounds showed higher docking scores for different GLUTs and better antidiabetic effects in terms of glucose uptake and insulin secretion in the NIT-1 cell line compared to Glibenclamide and SW [119]. Additionally, several other SW analogues have been shown to inhibit dipeptidyl peptidase IV (DPP-IV) enzyme activity, indicating their potential as targeted antidiabetic agents [120]. Moreover, several new glycosides also exhibit anti-HBV activity [121].
As an active compound of natural origin, swertiamarin (SW) exhibits a broad spectrum of pharmacological activities mediated through multi-target interactions and multi-mechanistic pathways. Current research reveals its therapeutic potential in hepatoprotection, glycemic regulation, neuroprotection, and anti-inflammatory applications, while also demonstrating promising antitumor, antidiabetic, and osteoarticular protective properties(Figure 10). Oxidative stress and inflammatory response are closely related to a variety of chronic diseases, SW can regulate classical signaling pathways such as NF-κB, MAPK, and PI3K/Akt to exert anti-inflammatory and antioxidant effects, thus achieving multiple biological effects. For example, SW can exert hepatoprotective and neuroprotective effects by inhibiting Nrf2-mediated oxidative stress. SW can ameliorate APAP-induced liver injury, OGDR-stimulated human neuronal SH-SY5Y cell injury, obesity-associated chronic inflammation, and arthritis through the NF-κB signaling pathway; SW also attenuated the LPS-induced fever and insulin resistance via regulating PI3K-Akt and p38 MAPK pathways.
In addition, SW may have a dual role in promoting or inhibiting apoptosis. For example, swertiamarin can promote apoptosis in HepG2 hepatocellular carcinoma cells, human peripheral blood mononuclear cells (PBMCs), and neuroblastoma (SK-N-AS) cell lines to exert antitumor effects. On the other hand, it inhibits apoptosis in hepatocytes to ameliorate hepatic injury and inhibits I/R-induced neuronal apoptosis to exert neuroprotective effects.
Future studies should focus on clarifying its pharmacological mechanisms and molecular targets, refining its pharmacokinetic profile, evaluating its toxicological safety, and exploring its efficacy and feasibility in the clinic. With more comprehensive research, swertiamarin is anticipated to serve as an adjunctive therapeutic agent for various chronic diseases.
Figure 10.
A Summary of the pharmacological effects of swertiamarin. Swertiamarin(SW) exhibits a broad spectrum of pharmacological activities including hepatoprotection, glycemic regulation, neuroprotection, and anti-inflammatory, antitumor,antidiabetic, and osteoarticular protective properties.
Figure 10.
A Summary of the pharmacological effects of swertiamarin. Swertiamarin(SW) exhibits a broad spectrum of pharmacological activities including hepatoprotection, glycemic regulation, neuroprotection, and anti-inflammatory, antitumor,antidiabetic, and osteoarticular protective properties.
Declaration of competing interest
These authors have no conflict of interest to declare.
Funding
This research was supported by the National Natural Science Foundation of China (82260727; 22367018), Yunnan Provincial Science and Technology Department (202101AY070001-010; 202201AY070001-005), the Innovation Team Construction Project of Kunming Medical University (CXTD202203), and the First-Class Discipline Team of Kunming Medical University (2024XKTDTS13).
References
- Chi, X.; Zhang, F.; Gao, Q.; Xing, R.; Chen, S. A Review on the Ethnomedicinal Usage, Phytochemistry, and Pharmacological Properties of Gentianeae (Gentianaceae) in Tibetan Medicine. Plants 2021, 10, 2383. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Gadimli, A.I.; Isaev, J.I.; Kashchenko, N.I.; Prokopyev, A.S.; Kataeva, T.N.; Chirikova, N.K.; Vennos, C. Caucasian Gentiana Species: Untargeted LC-MS Metabolic Profiling, Antioxidant and Digestive Enzyme Inhibiting Activity of Six Plants. Metabolites 2019, 9, 271. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, R.; Zhang, J.; Su, H.; Yang, Q.; Wulu, J.; Li, J.; Zhang, Z.; Lv, Z. Comparative analysis of phytochemical profile and antioxidant and anti-inflammatory activity of four Gentiana species from the Qinghai-Tibet Plateau. J. Ethnopharmacol. 2024, 326, 117926. [Google Scholar] [CrossRef]
- Mahendran, G.; Thamotharan, G.; Sengottuvelu, S.; Narmatha Bai, V. RETRACTED: Anti-diabetic activity of Swertia corymbosa (Griseb.) Wight ex C.B. Clarke aerial parts extract in streptozotocin induced diabetic. Diabet. rats. J. Ethnopharmacol. 2014, 151, 1175–1183. [Google Scholar] [CrossRef]
- Phoboo, S.; Pinto Mda, S.; Barbosa, A.C.; Sarkar, D.; Bhowmik, P.C.; Jha, P.K.; Shetty, K. Phenolic-linked biochemical rationale for the anti-diabetic properties of Swertia chirayita (Roxb. ex Flem.) Karst. Phytother. Res.: PTR 2013, 27, 227–235. [Google Scholar] [CrossRef]
- Patel, M.B.; Mishra, S.H. Hypoglycemic activity of C-glycosyl flavonoid from Enicostemma hyssopifolium. Pharm. Biol. 2011, 49, 383–391. [Google Scholar] [CrossRef]
- Lad, H.; Bhatnagar, D. Amelioration of oxidative and inflammatory changes by Swertia chirayita leaves in experimental arthritis. Inflammopharmacology 2016, 24, 363–375. [Google Scholar] [CrossRef]
- Hudecová, A.; Kusznierewicz, B.; Rundén-Pran, E.; Magdolenová, Z.; Hasplová, K.; Rinna, A.; Fjellsbø, L.M.; Kruszewski, M.; Lankoff, A.; Sandberg, W.J.; Refsnes, M.; Skuland, T.; Schwarze, P.; Brunborg, G.; Bjøras, M.; Collins, A.; Miadoková, E.; Gálová, E.; Dusinská, M. Silver nanoparticles induce premutagenic DNA oxidation that can be prevented by phytochemicals from Gentiana asclepiadea. Mutagenesis 2012, 27, 759–769. [Google Scholar] [CrossRef]
- Dai, K.; Yi, X.J.; Huang, X.J.; Muhammad, A.; Li, M.; Li, J.; Yang, G.Z.; Gao, Y. Hepatoprotective activity of iridoids, seco-iridoids and analog glycosides from Gentianaceae on HepG2 cells via CYP3A4 induction and mitochondrial pathway. Food Funct. 2018, 9, 2673–2683. [Google Scholar] [CrossRef]
- Mihailović, V.; Katanić, J.; Mišić, D.; Stanković, V.; Mihailović, M.; Uskoković, A.; Arambašić, J.; Solujić, S.; Mladenović, M.; Stanković, N. Hepatoprotective effects of secoiridoid-rich extracts from Gentiana cruciata L. against carbon tetrachloride induced liver damage in rats. Food Funct. 2014, 5, 1795–1803. [Google Scholar] [CrossRef]
- Patel, N.; Tyagi, R.K.; Tandel, N.; Garg, N.K.; Soni, N. The Molecular Targets of Swertiamarin and its Derivatives Confer Anti- Diabetic and Anti-Hyperlipidemic Effects. Curr. Drug Targets 2018, 19, 1958–1967. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Sashidhara, K.V. Lipid lowering agents of natural origin: An account of some promising chemotypes. Eur. J. Med. Chem. 2017, 140, 331–348. [Google Scholar] [CrossRef] [PubMed]
- Leong, X.Y.; Thanikachalam, P.V.; Pandey, M.; Ramamurthy, S. A systematic review of the protective role of swertiamarin in cardiac and metabolic diseases. Biomed. Pharmacother. = Biomed. Pharmacother. 2016, 84, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Li, H.L.; Peng, X.J.; He, J.C.; Feng, E.F.; Xu, G.L.; Rao, G.X. Development and validation of a LC-ESI-MS/MS method for the determination of swertiamarin in rat plasma and its application in pharmacokinetics. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2011, 879, 1653–1658. [Google Scholar] [CrossRef]
- Li, H.L.; He, J.C.; Bai, M.; Song, Q.Y.; Feng, E.F.; Rao, G.X.; Xu, G.L. Determination of the plasma pharmacokinetic and tissue distributions of swertiamarin in rats by liquid chromatography with tandem mass spectrometry. Arzneim. -Forsch. 2012, 62, 138–144. [Google Scholar] [CrossRef]
- Lu, C.M.; Lin, L.C.; Tsai, T.H. Determination and pharmacokinetic study of gentiopicroside, geniposide, baicalin, and swertiamarin in Chinese herbal formulae after oral administration in rats by LC-MS/MS. Molecules 2014, 19, 21560–21578. [Google Scholar] [CrossRef]
- Yong-Li, W.; Cong-Cong, Z.; Feng, Z.; Jian, H.; Dan-Dan, W.; Yong-Fang, Z.; Wei, L.; Guang-Bo, G.E.; Jian-Guang, X.U. [Chemical profiling and tissue distribution study of Jingyin Granules in rats using UHPLC-Q-Exactive Orbitrap HR-MS]. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Medica 2020, 45, 5537–5554. [Google Scholar]
- Xu, G.L.; Li, H.L.; He, J.C.; Feng, E.F.; Shi, P.P.; Liu, Y.Q.; Liu, C.X. Comparative pharmacokinetics of swertiamarin in rats after oral administration of swertiamarin alone, Qing Ye Dan tablets and co-administration of swertiamarin and oleanolic acid. J. Ethnopharmacol. 2013, 149, 49–54. [Google Scholar] [CrossRef]
- Wu, Y.; Ai, Y.; Wang, F.; Ma, W.; Bian, Q.; Lee, D.Y.; Dai, R. Simultaneous determination of four secoiridoid and iridoid glycosides in rat plasma by ultra performance liquid chromatography-tandem mass spectrometry and its application to a comparative pharmacokinetic study. Biomed. Chromatogr. BMC 2016, 30, 97–104. [Google Scholar] [CrossRef]
- Ye, X.; Zhang, T.; Han, H. Metabolite Profiling of Swertia cincta Extract in Rats and Pharmacokinetics Study of Three Bioactive Compounds Using UHPLC-MS/MS. Planta Medica 2023, 89, 333–346. [Google Scholar] [CrossRef]
- Feng, B.; Zhu, H.; Guan, J.; Zhao, L.; Gu, J.; Yin, L.; Fawcett, J.P.; Liu, W. A rapid and sensitive UFLC-MS/MS method for the simultaneous determination of gentiopicroside and swertiamarin in rat plasma and its application in pharmacokinetics. J. Pharm. Pharmacol. 2014, 66, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Sheng, N.; Zhi, X.; Yuan, L.; Zhang, Z.; Jia, P.; Zhang, X.; Zhang, L.; Wang, X. Pharmacokinetic and excretion study of three secoiridoid glycosides and three flavonoid glycosides in rat by LC-MS/MS after oral administration of the Swertia pseudochinensis extract. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2014, 967, 75–83. [Google Scholar] [CrossRef]
- Wang, Z.G.; Wang, X.J.; Sun, H.; Chen, L.; Ma, C.M. Determination of novel nitrogen-containing metabolite after oral administration of swertiamarin to rats. J. Asian Nat. Prod. Res. 2012, 14, 176–181. [Google Scholar] [CrossRef]
- Vaidya, H.; Goyal, R.K.; Cheema, S.K. Anti-diabetic activity of swertiamarin is due to an active metabolite, gentianine, that upregulates PPAR-γ gene expression in 3T3-L1 cells. Phytother. Res.: PTR 2013, 27, 624–627. [Google Scholar] [CrossRef]
- Xu, L.L.; Liu, C.; Han, Z.Z.; Han, H.; Yang, L.; Wang, Z.T. Microbial Biotransformation of Iridoid Glycosides from Gentiana Rigescens by Penicillium Brasilianum. Chem. Biodivers. 2020, 17, e2000676. [Google Scholar] [CrossRef]
- Wang, S.; Tang, S.; Sun, Y.; Wang, H.; Wang, X.; Zhang, H.; Wang, Z. Highly sensitive determination of new metabolite in rat plasma after oral administration of swertiamarin by liquid chromatography/time of flight mass spectrometry following picolinoyl derivatization. Biomed. Chromatogr.: BMC 2014, 28, 939–946. [Google Scholar] [CrossRef]
- Wu, X.; Tang, S.; Jin, Y.; Zhang, Y.; Hattori, M.; Wang, X.; Zhang, H.; Wang, Z. New analytical method for the study of metabolism of swertiamarin in rats after oral administration by UPLC-TOF-MS following DNPH derivatization. Biomed. Chromatogr.: BMC 2015, 29, 1184–1189. [Google Scholar] [CrossRef]
- Shi, M.; Xiong, K.; Zhang, T.; Han, H. Pharmacokinetics and metabolic profiles of swertiamarin in rats by liquid chromatography combined with electrospray ionization tandem mass spectrometry. J. Pharm. Biomed. Anal. 2020, 179, 112997. [Google Scholar] [CrossRef]
- Ma, B.; Lou, T.; Wang, T.; Li, R.; Liu, J.; Yu, S.; Guo, Y.; Wang, Z.; Wang, J. Comprehensive metabolism study of swertiamarin in rats using ultra high-performance liquid chromatography coupled with Quadrupole-Exactive Orbitrap mass spectrometry. Xenobiotica; Fate Foreign Compd. Biol. Syst. 2021, 51, 455–466. [Google Scholar] [CrossRef]
- Jaishree, V.; Badami, S. Antioxidant and hepatoprotective effect of swertiamarin from Enicostemma axillare against D-galactosamine induced acute liver damage in rats. J. Ethnopharmacol. 2010, 130, 103–106. [Google Scholar] [CrossRef]
- Wu, T.; Li, J.; Li, Y.; Song, H. Antioxidant and Hepatoprotective Effect of Swertiamarin on Carbon Tetrachloride-Induced Hepatotoxicity via the Nrf2/HO-1 Pathway. Cell. Physiol. Biochem. : Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 41, 2242–2254. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xiao, S.; Dai, X.; Tang, Z.; Wang, Y.; Ali, M.; Ataya, F.S.; Sahar, I.; Iqbal, M.; Wu, Y.; Li, K. Multi-omics analysis and the remedial effects of Swertiamarin on hepatic injuries caused by CCl(4). Ecotoxicol. Environ. Saf. 2024, 282, 116734. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, K.; Wu, T.; Song, H. Swertiamarin ameliorates carbon tetrachloride-induced hepatic apoptosis via blocking the PI3K/Akt pathway in rats. Korean J. Physiol. Pharmacol.: Off. J. Korean Physiol. Soc. Korean Soc. Pharmacol. 2019, 23, 21–28. [Google Scholar] [CrossRef]
- Kolure, R.; Vinaitheerthan, N.; Thakur, S.; Godela, R.; Doli, S.B.; Santhepete Nanjundaiah, M. Protective effect of Enicostemma axillare - Swertiamarin on oxidative stress against nicotine-induced liver damage in SD rats. Ann. Pharm. Fr. 2024, 82, 792–799. [Google Scholar] [CrossRef]
- Liu, L.; Xu, G.L.; Ma, X.X.; Khan, A.; Tan, W.H.; Yang, Z.Y.; Zhou, Z.H. Sweritranslactones A-C: Unusual Skeleton Secoiridoid Dimers via [4 + 2] Cycloaddition from Swertiamarin. J. Org. Chem. 2017, 82, 13263–13267. [Google Scholar] [CrossRef]
- Zou, Y.D.; Ma, X.X.; Du, S.N.; Qi, P.X.; He, F.Y.; Yang, Z.Y.; Tan, W.H.; Khan, A.; Zhou, Z.H.; Liu, L. Sweritranslactone D, a hepatoprotective novel secoiridoid dimer with tetracyclic lactone skeleton from heat-transformed swertiamarin. Fitoterapia 2021, 151, 104879. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhou, Q.; Xia, R.; Zhang, P.; Xie, Y.; Yang, Z.; Khan, A.; Zhou, Z.; Tan, W.; Liu, L. Swertiamarin or heat-transformed products alleviated APAP-induced hepatotoxicity via modulation of apoptotic and Nrf-2/NF-κB pathways. Heliyon 2023, 9, e18746. [Google Scholar] [CrossRef]
- Hernandez-Gea, V.; Friedman, S.L. Pathogenesis of liver fibrosis. Annu. Rev. Pathol. 2011, 6, 425–456. [Google Scholar] [CrossRef]
- Li, S.; Wang, Q.; Tao, Y.; Liu, C. Swertiamarin Attenuates Experimental Rat Hepatic Fibrosis by Suppressing Angiotensin II-Angiotensin Type 1 Receptor-Extracellular Signal-Regulated Kinase Signaling. J. Pharmacol. Exp. Ther. 2016, 359, 247–255. [Google Scholar] [CrossRef]
- Cotter, T.G.; Rinella, M. Nonalcoholic Fatty Liver Disease 2020: The State of the Disease. Gastroenterology 2020, 158, 1851–1864. [Google Scholar] [CrossRef]
- Yang, Y.; Li, J.; Wei, C.; He, Y.; Cao, Y.; Zhang, Y.; Sun, W.; Qiao, B.; He, J. Amelioration of nonalcoholic fatty liver disease by swertiamarin in fructose-fed mice. Phytomedicine: Int. J. Phytother. Phytopharm. 2019, 59, 152782. [Google Scholar] [CrossRef] [PubMed]
- Patel, T.P.; Rawal, K.; Soni, S.; Gupta, S. Swertiamarin ameliorates oleic acid induced lipid accumulation and oxidative stress by attenuating gluconeogenesis and lipogenesis in hepatic steatosis. Biomed. Pharmacother. 2016, 83, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, C.; Ziani, S.; Nelson, L.J.; Ávila, M.A.; Nevzorova, Y.A.; Cubero, F.J. Fibrotic Events in the Progression of Cholestatic Liver Disease. Cells 2021, 10, 1107. [Google Scholar] [CrossRef]
- Padda, M.S.; Sanchez, M.; Akhtar, A.J.; Boyer, J.L. Drug-induced cholestasis. Hepatology 2011, 53, 1377–1387. [Google Scholar] [CrossRef]
- Shi, M.; Tang, J.; Zhang, T.; Han, H. Swertiamarin, an active iridoid glycoside from Swertia pseudochinensis H. Hara, protects against alpha-naphthylisothiocyanate-induced cholestasis by activating the farnesoid X receptor and bile acid excretion pathway. J. Ethnopharmacol. 2022, 291, 115164. [Google Scholar] [CrossRef]
- Tian, C.; Zhang, T.; Wang, L.; Shan, Q.; Jiang, L. The hepatoprotective effect and chemical constituents of total iridoids and xanthones extracted from Swertia mussotii Franch. J. Ethnopharmacol. 2014, 154, 259–266. [Google Scholar] [CrossRef]
- Zhang, L.; Cheng, Y.; Du, X.; Chen, S.; Feng, X.; Gao, Y.; Li, S.; Liu, L.; Yang, M.; Chen, L.; Peng, Z.; Yang, Y.; Luo, W.; Wang, R.; Chen, W.; Chai, J. Swertianlarin, an Herbal Agent Derived from Swertia mussotii Franch, Attenuates Liver Injury, Inflammation, and Cholestasis in Common Bile Duct-Ligated Rats. Evid.-Based Complement. Altern. Med.: eCAM 2015, 2015, 948376. [Google Scholar] [CrossRef]
- Elsayed, N.A.; Aleppo, G.; Bannuru, R.R.; Bruemmer, D.; Collins, B.S.; Ekhlaspour, L.; Gaglia, J.L.; Hilliard, M.E.; Johnson, E.L.; Khunti, K.; Lingvay, I.; Matfin, G.; Mccoy, R.G.; Perry, M.L.; Pilla, S.J.; Polsky, S.; Prahalad, P.; Pratley, R.E.; Segal, A.R.; Seley, J.J.; Selvin, E.; Stanton, R.C.; Gabbay, R.A. 2. Diagnosis and Classification of Diabetes: Standards of Care in Diabetes—2024. Diabetes Care 2024, 47, S20–S42. [Google Scholar]
- Sonawane, R.D.; Deore, V.B.; Patil, S.D.; Patil, C.R.; Surana, S.J.; Goyal, R.K. Role of 5-HT2 receptors in diabetes: Swertiamarin seco-iridoid glycoside might be a possible 5-HT2 receptor modulator. Physiol. Behav. 2015, 144, 66–72. [Google Scholar] [CrossRef]
- Jaishree, V.; Narsimha, S. Swertiamarin and quercetin combination ameliorates hyperglycemia, hyperlipidemia and oxidative stress in streptozotocin-induced type 2 diabetes mellitus in wistar rats. Biomed. Pharmacother. = Biomed. Pharmacother. 2020, 130, 110561. [Google Scholar] [CrossRef]
- Patel, T.P.; Soni, S.; Parikh, P.; Gosai, J.; Chruvattil, R.; Gupta, S. Swertiamarin: An Active Lead from Enicostemma littorale Regulates Hepatic and Adipose Tissue Gene Expression by Targeting PPAR- γ and Improves Insulin Sensitivity in Experimental NIDDM Rat Model. Evid. -Based Complement. Altern. Med.: Ecam 2013, 2013, 358673. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Long, J.; Dong, L.; Zhang, J.; Wang, A.; Zhang, Y.; Yan, D. Uncovering the key pharmacodynamic material basis and possible molecular mechanism of Xiaoke formulation improve insulin resistant through a comprehensive investigation. J. Ethnopharmacol. 2024, 323, 117752. [Google Scholar] [CrossRef]
- Dhanavathy, G. Immunohistochemistry, histopathology, and biomarker studies of swertiamarin, a secoiridoid glycoside, prevents and protects streptozotocin-induced β-cell damage in Wistar rat pancreas. J. Endocrinol. Investig. 2015, 38, 669–684. [Google Scholar] [CrossRef]
- Parwani, K.; Patel, F.; Bhagwat, P.; Dilip, H.; Patel, D.; Thiruvenkatam, V.; Mandal, P. Swertiamarin mitigates nephropathy in high-fat diet/streptozotocin-induced diabetic rats by inhibiting the formation of advanced glycation end products. Arch. Physiol. Biochem. 2024, 130, 136–154. [Google Scholar] [CrossRef]
- Dai, L.L.; Cho, S.B.; Li, H.F.; ALS, *!!! REPLACE !!!*; Ji, X.P.; Pan, S.; Bao, M.L.; Bai, L.; Ba, G.N.; Fu, M.H. Lomatogonium rotatum extract alleviates diabetes mellitus induced by a high-fat, high-sugar diet and streptozotocin in rats. World J. Diabetes 2023, 14, 846–861. [Google Scholar] [CrossRef]
- Forbes, J.M.; Cooper, M.E. Mechanisms of diabetic complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef]
- Vaidya, H.; Prajapati, A.; Rajani, M.; Sudarsanam, V.; Padh, H.; Goyal, R.K. Beneficial effects of swertiamarin on dyslipidaemia in streptozotocin-induced type 2 diabetic rats. Phytother. Res.: PTR 2012, 26, 1259–1261. [Google Scholar] [CrossRef]
- Parwani, K.; Patel, F.; Patel, D.; Mandal, P. Protective Effects of Swertiamarin against Methylglyoxal-Induced Epithelial-Mesenchymal Transition by Improving Oxidative Stress in Rat Kidney Epithelial (NRK-52E) Cells. Molecules 2021, 26, 2748. [Google Scholar] [CrossRef]
- Sonawane, R.D.; Vishwakarma, S.L.; Lakshmi, S.; Rajani, M.; Padh, H.; Goyal, R.K. Amelioration of STZ-induced type 1 diabetic nephropathy by aqueous extract of Enicostemma littorale Blume and swertiamarin in rats. Mol. Cell. Biochem. 2010, 340, 1–6. [Google Scholar] [CrossRef]
- Yang, K.; Wang, Y.; Li, Y.W.; Chen, Y.G.; Xing, N.; Lin, H.B.; Zhou, P.; Yu, X.P. Progress in the treatment of diabetic peripheral neuropathy. Biomed. Pharmacother. = Biomed. Pharmacother. 2022, 148, 112717. [Google Scholar] [CrossRef]
- Selvarajah, D.; Kar, D.; Khunti, K.; Davies, M.J.; Scott, A.R.; Walker, J.; Tesfaye, S. Diabetic peripheral neuropathy: Advances in diagnosis and strategies for screening and early intervention. Lancet Diabetes Endocrinol. 2019, 7, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yao, J.; Yao, X.; Lao, J.; Liu, D.; Chen, C.; Lu, Y. [Swertiamarin alleviates diabetic peripheral neuropathy in rats by suppressing NOXS/ ROS/NLRP3 signal pathway]. Nan Fang Yi Ke Da Xue Xue Bao = J. South. Med. Univ. 2021, 41, 937–941. [Google Scholar]
- Vaidya, H.; Giri, S.; Jain, M.; Goyal, R. Decrease in serum matrix metalloproteinase-9 and matrix metalloproteinase-3 levels in Zucker fa/fa obese rats after treatment with swertiamarin. Exp. Clin. Cardiol. 2012, 17, 12–16. [Google Scholar]
- Sharma, M.; Malim, F.M.; Goswami, A.; Sharma, N.; Juvvalapalli, S.S.; Chatterjee, S.; Kate, A.S.; Khairnar, A. Neuroprotective Effect of Swertiamarin in a Rotenone Model of Parkinson’s Disease: Role of Neuroinflammation and Alpha-Synuclein Accumulation. ACS Pharmacol. Transl. Sci. 2023, 6, 40–51. [Google Scholar] [CrossRef]
- Darwish, S.F.; Elbadry, A.M.M.; Elbokhomy, A.S.; Salama, G.A.; Salama, R.M. The dual face of microglia (M1/M2) as a potential target in the protective effect of nutraceuticals against neurodegenerative diseases. Front. Aging 2023, 4, 1231706. [Google Scholar] [CrossRef]
- Pandey, T.; Smita, S.S.; Mishra, A.; Sammi, S.R.; Pandey, R. Swertiamarin, a secoiridoid glycoside modulates nAChR and AChE activity. Exp. Gerontol. 2020, 138, 111010. [Google Scholar] [CrossRef]
- Ss, S. To Study the Effect of Swertiamarin in Animal Model of Huntington’s Disease. Open Access J. Pharm. Res. 2018, 2, 000163. [Google Scholar] [CrossRef]
- Wang, G.; Quan, J.; Su, N.; Li, P.; Yu, Q. Proteomic Analysis of Swertiamarin-treated BV-2 Cells and Possible Implications in Neuroinflammation. J. Oleo Sci. 2022, 71, 395–400. [Google Scholar] [CrossRef]
- Wu, J.; Yang, F.; Li, X.; Wu, T.; Liu, L.; Song, H. Swertiamarin protects neuronal cells from oxygen glucose deprivation/reoxygenation via TLR4/PARP1/NF-κB pathway. Die Pharm. 2019, 74, 481–484. [Google Scholar]
- Pandey, T.; Shukla, A.; Trivedi, M.; Khan, F.; Pandey, R. Swertiamarin from Enicostemma littorale, counteracts PD associated neurotoxicity via enhancement α-synuclein suppressive genes and SKN-1/NRF-2 activation through MAPK pathway. Bioorganic Chem. 2021, 108, 104655. [Google Scholar] [CrossRef]
- Zeng, M.; Zhang, R.; Yang, Q.; Guo, L.; Zhang, X.; Yu, B.; Gan, J.; Yang, Z.; Li, H.; Wang, Y.; Jiang, X.; Lu, B. Pharmacological therapy to cerebral ischemia-reperfusion injury: Focus on saponins. Biomed. Pharmacother. 2022, 155, 113696. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wei, W.; Lan, X.; Liu, N.; Li, Y.; Ma, H.; Sun, T.; Peng, X.; Zhuang, C.; Yu, J. Neuroprotective Effect of Swertiamain on Cerebral Ischemia/Reperfusion Injury by Inducing the Nrf2 Protective Pathway. ACS Chem. Neurosci. 2019, 10, 2276–2286. [Google Scholar] [CrossRef]
- Deng, X.H.; Zhang, X.; Wang, J.; Ma, P.S.; Ma, L.; Niu, Y.; Sun, T.; Zhou, R.; Yu, J.Q. Anticonvulsant Effect of Swertiamarin Against Pilocarpine-Induced Seizures in Adult Male Mice. Neurochem. Res. 2017, 42, 3103–3113. [Google Scholar] [CrossRef]
- Vaijanathappa, J.; Puttaswamygowda, J.; Bevanhalli, R.; Dixit, S.; Prabhakaran, P. Molecular docking, antiproliferative and anticonvulsant activities of swertiamarin isolated from Enicostemma axillare. Bioorganic Chem. 2020, 94, 103428. [Google Scholar] [CrossRef]
- Li, F.; Jiang, M.; Ma, M.; Chen, X.; Zhang, Y.; Zhang, Y.; Yu, Y.; Cui, Y.; Chen, J.; Zhao, H.; Sun, Z.; Dong, D. Anthelmintics nitazoxanide protects against experimental hyperlipidemia and hepatic steatosis in hamsters and mice. Acta Pharm. Sin. B 2022, 12, 1322–1338. [Google Scholar] [CrossRef]
- Xu, L.; Yang, Q.; Zhou, J. Mechanisms of Abnormal Lipid Metabolism in the Pathogenesis of Disease. Int. J. Mol. Sci. 2024, 25, 8465. [Google Scholar] [CrossRef]
- Vaidya, H.; Rajani, M.; Sudarsanam, V.; Padh, H.; Goyal, R. Swertiamarin: A lead from Enicostemma littorale Blume. for anti-hyperlipidaemic effect. Eur. J. Pharmacol. 2009, 617, 108–112. [Google Scholar] [CrossRef]
- Vaidya, H.; Rajani, M.; Sudarsanam, V.; Padh, H.; Goyal, R. Antihyperlipidaemic activity of swertiamarin, a secoiridoid glycoside in poloxamer-407-induced hyperlipidaemic rats. J. Nat. Med. 2009, 63, 437–442. [Google Scholar] [CrossRef]
- Wang, J.; He, J. Swertiamarin decreases lipid accumulation dependent on 3-ketoacyl-coA thiolase. Biomed. Pharmacother. = Biomed. Pharmacother. 2019, 112, 108668. [Google Scholar] [CrossRef]
- Wei, S.; Chen, G.; He, W.; Chi, H.; Abe, H.; Yamashita, K.; Yokoyama, M.; Kodama, H. Inhibitory effects of secoiridoids from the roots of Gentiana straminea on stimulus-induced superoxide generation, phosphorylation and translocation of cytosolic compounds to plasma membrane in human neutrophils. Phytother. Res.: PTR 2012, 26, 168–173. [Google Scholar] [CrossRef]
- Piché, M.E.; Tchernof, A.; Després, J.P. Obesity Phenotypes, Diabetes, and Cardiovascular Diseases. Circ. Res. 2020, 126, 1477–1500. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, D.; Zhu, Y.; Cai, S.; Liang, X.; Tang, Y.; Jin, S.; Ding, C. Swertiamarin supplementation prevents obesity-related chronic inflammation and insulin resistance in mice fed a high-fat diet. Adipocyte 2021, 10, 160–173. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, H.; Ma, M.; Li, D.; Zhang, O.; Cai, S.; Wang, Y.; Chen, D.; Jin, S.; Ding, C.; Xu, L. Swertiamarin ameliorates diet-induced obesity by promoting adipose tissue browning and oxidative metabolism in preexisting obese mice. Acta Biochim. Et Biophys. Sin. 2022, 55, 131–142. [Google Scholar] [CrossRef]
- Saravanan, S.; Pandikumar, P.; Prakash Babu, N.; Hairul Islam, V.I.; Thirugnanasambantham, K.; Gabriel Paulraj, M.; Balakrishna, K.; Ignacimuthu, S. In vivo and in vitro immunomodulatory potential of swertiamarin isolated from Enicostema axillare (Lam.) A. Raynal that acts as an anti-inflammatory agent. Inflammation 2014, 37, 1374–1388. [Google Scholar] [CrossRef]
- Zhang, M.; Ma, X.; Xu, H.; Wu, W.; He, X.; Wang, X.; Jiang, M.; Hou, Y.; Bai, G. A natural AKT inhibitor swertiamarin targets AKT-PH domain, inhibits downstream signaling, and alleviates inflammation. FEBS J. 2020, 287, 1816–1829. [Google Scholar] [CrossRef]
- Patel, N.; Zinzuvadia, A.; Prajapati, M.; Tyagi, R.K.; Dalai, S. Swertiamarin-mediated immune modulation/adaptation confers protection against Plasmodium berghei. Future Microbiol. 2022, 17, 931–941. [Google Scholar] [CrossRef]
- Shitlani, D.; Choudhary, R.; Pandey, D.P.; Bodakhe, S.H. Ameliorative antimalarial effects of the combination of rutin and swertiamarin on malarial parasites. Asian Pac. J. Trop. Dis. 2016, 6, 453–459. [Google Scholar] [CrossRef]
- Wang, H.; Tian, L.; Han, Y.; Ma, X.; Hou, Y.; Bai, G. Mechanism Assay of Honeysuckle for Heat-Clearing Based on Metabolites and Metabolomics. Metabolites 2022, 12, 121. [Google Scholar] [CrossRef]
- Vaijanathappa, J.; Badami, S. Antiedematogenic and free radical scavenging activity of swertiamarin isolated from Enicostemma axillare. Planta Medica 2009, 75, 12–17. [Google Scholar] [CrossRef]
- Tang, H.; Ke, Y.; Ren, Z.; Lei, X.; Xiao, S.; Bao, T.; Shi, Z.; Zou, R.; Wu, T.; Zhou, J.; Geng, C.A.; Wang, L.; Chen, J. Bioinformatics analysis of differentially expressed genes in hepatocellular carcinoma cells exposed to Swertiamarin. J. Cancer 2019, 10, 6526–6534. [Google Scholar] [CrossRef]
- Xiao, S.; Tang, H.; Bai, Y.; Zou, R.; Ren, Z.; Wu, X.; Shi, Z.; Lan, S.; Liu, W.; Wu, T.; Zhang, C.; Wang, L. Swertiamarin suppresses proliferation, migration, and invasion of hepatocellular carcinoma cells via negative regulation of FRAT1. Eur. J. Histochem. : EJH 2020, 64, 3169. [Google Scholar] [CrossRef] [PubMed]
- Wani, B.A.; Ramamoorthy, D.; Rather, M.A.; Arumugam, N.; Qazi, A.K.; Majeed, R.; Hamid, A.; Ganie, S.A.; Ganai, B.A.; Anand, R.; Gupta, A.P. Induction of apoptosis in human pancreatic MiaPaCa-2 cells through the loss of mitochondrial membrane potential (ΔΨm) by Gentiana kurroo root extract and LC-ESI-MS analysis of its principal constituents. Phytomedicine: Int. J. Phytother. Phytopharm. 2013, 20, 723–733. [Google Scholar] [CrossRef]
- Wang, H.; Huang, H.; Lv, J.; Jiang, N.; Li, Y.; Liu, X.; Zhao, H. Iridoid compounds from the aerial parts of Swertia mussotii Franch with cytotoxic activity. Nat. Prod. Res. 2021, 35, 1544–1549. [Google Scholar] [CrossRef]
- Valenta Šobot, A.; Drakulić, D.; Todorović, A.; Janić, M.; Božović, A.; Todorović, L.; Filipović Tričković, J. Gentiopicroside and swertiamarin induce non-selective oxidative stress-mediated cytotoxic effects in human peripheral blood mononuclear cells. Chem.-Biol. Interact. 2024, 111103. [Google Scholar] [CrossRef]
- Dubey, S.; Kallubai, M.; Subramanyam, R. Comparative binding of Swertiamarin with human serum albumin and α-1 glycoprotein and its cytotoxicity against neuroblastoma cells. J. Biomol. Struct. Dyn. 2020, 38, 5266–5276. [Google Scholar] [CrossRef]
- Smolen, J.S.; Aletaha, D.; Mcinnes, I.B. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef]
- Saravanan, S.; Islam, V.I.; Thirugnanasambantham, K.; Pazhanivel, N.; Raghuraman, N.; Paulraj, M.G.; Ignacimuthu, S. Swertiamarin ameliorates inflammation and osteoclastogenesis intermediates in IL-1β induced rat fibroblast-like synoviocytes. Inflamm. Res.: Off. J. Eur. Histamine Res. Soc. 2014, 63, 451–462. [Google Scholar] [CrossRef]
- Komatsu, N.; Takayanagi, H. Mechanisms of joint destruction in rheumatoid arthritis - immune cell-fibroblast-bone interactions. Nat. Rev. Rheumatol. 2022, 18, 415–429. [Google Scholar] [CrossRef]
- Hairul-Islam, M.I.; Saravanan, S.; Thirugnanasambantham, K.; Chellappandian, M.; Simon Durai Raj, C.; Karikalan, K.; Gabriel Paulraj, M.; Ignacimuthu, S. Swertiamarin, a natural steroid, prevent bone erosion by modulating RANKL/RANK/OPG signaling. Int. Immunopharmacol. 2017, 53, 114–124. [Google Scholar] [CrossRef]
- Saravanan, S.; Islam, V.I.; Babu, N.P.; Pandikumar, P.; Thirugnanasambantham, K.; Chellappandian, M.; Raj, C.S.; Paulraj, M.G.; Ignacimuthu, S. Swertiamarin attenuates inflammation mediators via modulating NF-κB/I κB and JAK2/STAT3 transcription factors in adjuvant induced arthritis. Eur. J. Pharm. Sci.: Off. J. Eur. Fed. Pharm. Sci. 2014, 56, 70–86. [Google Scholar] [CrossRef]
- Liedtke, V.; Stöckle, M.; Junker, K.; Roggenbuck, D. Benign prostatic hyperplasia - A novel autoimmune disease with a potential therapy consequence? Autoimmun. Rev. 2024, 23, 103511. [Google Scholar] [CrossRef]
- Wu, X.; Gu, Y.; Li, L. The anti-hyperplasia, anti-oxidative and anti-inflammatory properties of Qing Ye Dan and swertiamarin in testosterone-induced benign prostatic hyperplasia in rats. Toxicol. Lett. 2017, 265, 9–16. [Google Scholar] [PubMed]
- Chen, J.; Liu, J.; Lei, Y.; Liu, M. The anti-inflammation, anti-oxidative and anti-fibrosis properties of swertiamarin in cigarette smoke exposure-induced prostate dysfunction in rats. Aging 2019, 11, 10409–10421. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Liang, Y.; Tian, S.; Jin, J.; Zhao, Y.; Fan, H. Radiation-Induced Intestinal Injury: Injury Mechanism and Potential Treatment Strategies. Toxics 2023, 11, 1011. [Google Scholar] [CrossRef]
- Zhao, Z.; He, D.; Wang, J.; Xiao, Y.; Gong, L.; Tang, C.; Peng, H.; Qiu, X.; Liu, R.; Zhang, T.; Li, J. Swertiamarin relieves radiation-induced intestinal injury by limiting DNA damage. Mol. Cell. Biochem. 2024, 2277–2290. [Google Scholar]
- Jaishree, V.; Badami, S.; Rupesh Kumar, M.; Tamizhmani, T. Antinociceptive activity of swertiamarin isolated from Enicostemma axillare. Phytomedicine: Int. J. Phytother. Phytopharm. 2009, 16, 227–232. [Google Scholar] [CrossRef]
- Chabane, S.; Boudjelal, A.; Bouaziz-Terrachet, S.; Spinozzi, E.; Maggi, F.; Petrelli, R.; Tail, G. Analgesic effect of Centaurium erythraea and molecular docking investigation of the major component swertiamarin. Nat. Prod. Res. 2023, 1–7. [Google Scholar] [CrossRef]
- Kumarasamy, Y.; Nahar, L.; Cox, P.J.; Jaspars, M.; Sarker, S.D. Bioactivity of secoiridoid glycosides from Centaurium erythraea. Phytomedicine: Int. J. Phytother. Phytopharm. 2003, 10, 344–347. [Google Scholar] [CrossRef]
- Siler, B.; Misić, D.; Nestorović, J.; Banjanac, T.; Glamoclija, J.; Soković, M.; Cirić, A. Antibacterial and antifungal screening of Centaurium pulchellum crude extracts and main secoiridoid compounds. Nat. Prod. Commun. 2010, 5, 1525–1530. [Google Scholar]
- Perumal, S.; Gopal Samy, M.; Subramanian, D. In vitro and in silico screening of novel typhoid drugs from endangered herb (Enicostema axillare). J. Biomol. Struct. Dyn. 2023, 41, 2926–2936. [Google Scholar]
- Perumal, S.; Samy, M.G.; Subramanian, D. Effect of novel therapeutic medicine swertiamarin from Enicostema axillare in zebrafish infected with Salmonella typhi. Chem. Biol. Drug Des. 2022, 100, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Belani, M.A.; Shah, P.; Banker, M.; Gupta, S.S. Investigating the potential role of swertiamarin on insulin resistant and non-insulin resistant granulosa cells of poly cystic ovarian syndrome patients. J. Ovarian Res. 2023, 16, 55. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Sumiyoshi, M. Effects of Swertia japonica extract and its main compound swertiamarin on gastric emptying and gastrointestinal motility in mice. Fitoterapia 2011, 82, 827–833. [Google Scholar] [CrossRef]
- Chang, J.; Zou, S.; Xu, S.; Xiao, Y.; Zhu, D. Screening of Inhibitors against Idiopathic Pulmonary Fibrosis: Few-shot Machine Learning and Molecule Docking based Drug Repurposing. Curr. Comput. -Aided Drug Des. 2024, 20, 134–144. [Google Scholar] [CrossRef]
- Chang, J.; Zou, S.; Xiao, Y.; Zhu, D. Identification and validation of targets of swertiamarin on idiopathic pulmonary fibrosis through bioinformatics and molecular docking-based approach. BMC Complement. Med. Ther. 2023, 23, 352. [Google Scholar] [CrossRef]
- Wang, T.; Wu, S.; Ibrahim Ia, A.; Fan, L. Cardioprotective Role of Swertiamarin, a Plant Glycoside Against Experimentally Induced Myocardial Infarction via Antioxidant and Anti-inflammatory Functions. Appl. Biochem. Biotechnol. 2023, 195, 5394–5408. [Google Scholar] [CrossRef]
- Doss, V.A.; Kuberapandian, D. Evaluation of Anti-hypertrophic Potential of Enicostemma littorale Blume on Isoproterenol Induced Cardiac Hypertrophy. Indian J. Clin. Biochem.: IJCB 2021, 36, 33–42. [Google Scholar] [CrossRef]
- Perumal, S.; Gopal Samy, M.V.; Subramanian, D. Developmental toxicity, antioxidant, and marker enzyme assessment of swertiamarin in zebrafish (Danio rerio). J. Biochem. Mol. Toxicol. 2021, 35, e22843. [Google Scholar] [CrossRef]
- Kumar, S.; Niguram, P.; Jairaj, V.; Chauhan, N.; Jinagal, S.; Sagar, S.; Sindhu, R.K.; Chandra, A. Exploring the potential of semi-synthetic Swertiamarin analogues for GLUT facilitation and insulin secretion in NIT-1 cell lines: A molecular docking and in-vitro study. Nat. Prod. Res. 2024, 1–5. [Google Scholar] [CrossRef]
- Kumar, S.; Niguram, P.; Bhat, V.; Jinagal, S.; Jairaj, V.; Chauhan, N. Synthesis, molecular docking and ADMET prediction of novel swertiamarin analogues for the restoration of type-2 diabetes: An enzyme inhibition assay. Nat. Prod. Res. 2022, 36, 2197–2207. [Google Scholar] [CrossRef]
- Jie, X.X.; Geng, C.A.; Huang, X.Y.; Ma, Y.B.; Zhang, X.M.; Zhang, R.P.; Chen, J.J. Five new secoiridoid glycosides and one unusual lactonic enol ketone with anti-HBV activity from Swertia cincta. Fitoterapia 2015, 102, 96–101. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



