
Submitted:
26 May 2025
Posted:
26 May 2025
You are already at the latest version
Abstract
In biological control of pests using predatory natural enemies, predators not only prey on pests directly but also can affect the population fitness of pests through indirect non-consumptive effects (predation risk effects). However, the impact of predation risk effects varies depending on the mode of stress imposed by natural enemies and the state of the stressed pests. Herein, we exposed aphids (Periphyllus koelreuteriae) at different stages to various cues from the multicolored Asian lady beetle (Harmonia axyridis) to assess the effects of different predation risks on P. koelreuteriae development and reproduction. We found that the effect of predation risk on aphid developmental time was clearly stage-dependent. When 1st instar nymphs were exposed to predator cues, their developmental time was prolonged only in the early stages (1st-3rd instar). Similarly, when 3rd instar nymphs were stressed, only the current stage (3rd instar) showed a developmental delay, while 4th instar nymphs and adult aphids were not significantly affected. Additionally, aphids at different stages perceive predation risk differently. Nymphs cannot recognize visual cues from predators but can perceive odor cues. In contrast, adult aphids are sensitive to a variety of predator cues, including visual ones. Predation risk caused aphids to shorten their adult lifespan and reduced reproductive output, but it increased the proportion of diapause offspring. This study highlights the importance of considering how predation risk affects multiple life stages and physiological changes, which are important for gaining insights into the mechanisms of predator-prey interactions and for comprehensively assessing the ability of lady beetles to regulate aphid populations.
Keywords:
Periphyllus koelreuteriae
; Harmonia axyridis
; predator-prey interaction
; non-consumptive effect
; developmental stage-dependent
1. Introduction
One of the most important interspecific interactions in ecology is the constant struggle between predator and prey. Predator-prey interactions can be divided into two types: consumptive and non-consumptive effects [1,2]. Predators use consumptive effects to reduce prey densities through lethal predation and can also affect prey populations by influencing prey fitness through non-consumptive effects or predation risk [3,4]. While predation risk doesn't directly kill prey, it can have adverse effects. Upon sensing a predation threat, prey alter their behavior and physiology, which may decrease their suitability for survival or reproduction.[5]. Research indicates that the non-consumptive impacts of predators on their prey can occasionally be more significant than the consumptive impacts [6]. Identifying how prey sense predators and the trait modifications that lead to declines in prey numbers is vital for a thorough understanding of predator effects on prey.
Prey uses a range of stimuli, such as visual, auditory, olfactory, and tactile cues, to evaluate the risk of predation in their environment [7,8]. Prey can recognize potential threats and initiate avoidance actions based on visual cues like predator appearance, body shape, color, and movement patterns [9,10]. Alternatively, prey can assess the threat situation based on specific vocalizations of predators [11]. Volatile chemical cues have been reported to be particularly important in prey assessment of predation risk [12]. However, prey must rely more on visual or tactile evaluations to determine predation risk when olfactory cues are unclear or absent [13]. To understand the impact of non-consumptive effects on prey populations, it is critical to identify mechanisms for detecting predation risk.
When animals correctly evaluate predation risk, they can exhibit effective antipredator responses. This defensive strategy usually leads to changes in the prey's behavioral responses, developmental progress, morphological characteristics, physiological traits, and resource allocation [14,15,16]. For example, when there is a threat of predators, the larvae of the potato beetle, Leptinotarsa decemlineata, decrease their feeding activity [12]. Under the threat of predation by turtle lady beetles (Propylea japonica), Drosophila melanogaster exhibits different levels of accelerated development [17]. Offspring of Daphnia cucullata Sars have longer helmets in the presence of predator cues, resulting in better survival rates [18]. Exposure to predators of potato beetles increases the risk of being killed by entomopathogenic nematodes and fungi [19]. When female damselflies (Ischnura cervula) detect predator signals in the water, they decrease their feeding drive and focus more on locating safe habitats [20].
Periphyllus koelreuteriae Takahashi is one of the most damaging pests of Koelreuteria bipinnata Franch in East Asia [21]. It mainly deeds on the tender shoots, leaves, and other young parts of the tree, causing the leaves to curl up, in severe cases, even leading to the death of branches and leaves. At the same time, the aphid also secretes large quantities of honeydew, which induces the host leaves and lower shrubs to develop sooty mold disease, reducing the ornamental value and even polluting pedestrians [22]. In China, this aphid has two damage peaks each year (in spring and autumn). It overwinters as fertilized eggs in the bark crevices and at the tree forks [23,24]. The eggs hatch in early spring of the following year and continue to infest K. bipinnata from March to May. Diapausing aphids begin to appear in the field in late April, and the diapause first instar nymphs oversummer after mid-May, forming a second peak of infestation in late autumn [22]. Currently, chemical control is still the main control measure for this aphid, and the long-term use not only accelerates the development of pesticide resistance but also causes environmental pollution and endangers human health. Biological control is a promising alternative for controlling P. koelreuteriae. The multi-colored Asian ladybird beetle, Harmonia axyridis (Pallas), is an impressive consumer of aphids, abundant and ubiquitous in agroforestry ecosystems, and is often used as a biological agent to suppress aphid populations [25]. However, the predation risk posed by H. axyridis on P. koelreuteriae has not been reported. In order to improve the effectiveness of biological control, it is necessary to understand the effect of predation risk on P. koelreuteriae to maximize the role of lady beetles in aphid control.
Here, we investigated the effect of predation risk from H. axyridis on the development and fecundity of different stages (1st instar nymphs, 3rd instar nymphs and adults) of P. koelreuteriae, including the production of diapause offspring. Four types of predation risks were established: predator visual cues (PV), predator odor cues (PO), predator visual and odor cues (PV+PO), and predator visual, odor, and tactile cues (PV+PO+PT). The results reveal the detection mechanism of P. koelreuteriae in response to the predation risk from H. axyridis. This will help evaluate the population control ability of ladybugs over aphids more comprehensively and provide new ideas for environmentally friendly aphid control.
2. Materials and Methods
2.1. Experimental Insects
Eggs of P. koelreuteriae were collected from K. bipinnata trees at Henan University of Science and Technology (34°62’ N, 112°42’ E) Luoyang, China, during November 2022. Barks with aphid eggs were taken back to the laboratory and stored at 0℃ until the end of February 2023. Small pieces of bark with eggs were placed in petri dishes (d = 90 mm) covered with moistened filter paper at the bottom, and then the petri dishes were placed in an artificial climate chamber (PQX-450B-30H, Ningbo Laifu Technology Co., Ltd., Ningbo, China). The control conditions were set at a temperature of 23 ± 1℃, a relative humidity of 65% ± 5%, and a photoperiod of 15:9 h (L: D). Hatched nymphs were collected daily at 9:00 and 21:00 daily. The nymphs were reared on branches of K. bipinnata that were grown in hydroponically. The tested aphids included three groups: first-instar nymphs, third-instar nymphs, and adults.
Adults of H. axyridis were collected from oilseed rape (Brassica napus) fields near the campus in early April. They were fed P. koelreuteriae, Myzus persicae and Cinara tujafilina in a rearing box (13.7 cm ◊ 8.2 cm ◊ 5.3 cm, room temperature), with sufficient aphids provided every day. Efforts were made to ensure that the ladybug beetles could feed on different species of aphids to maintain their feeding habits and vitality in the wild. Only adult H. axyridis were used in the experiment, but without age standardization.
2.2. The Effect of H. axyridis Risk on Development and Reproduction of P. koelreuteriae
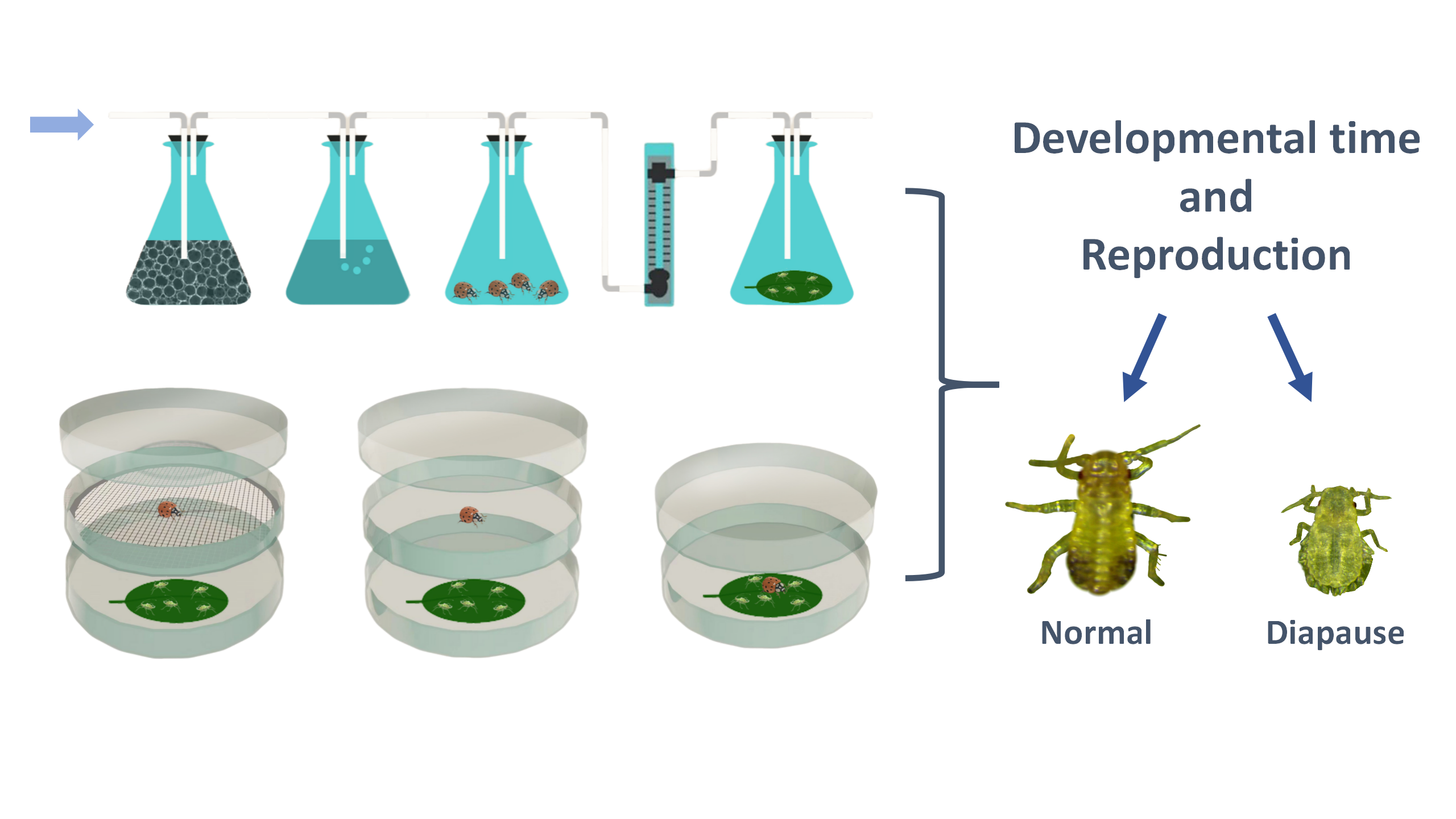
The test apparatus used to detect aphid response to different predator cues was modified according to Hermann et al. (2021) [26]. The effect of ladybug odors on aphid development and reproduction was assessed using an odor source test system (Figure 1A). The system consisted of four triangular flasks (50 mL) connected in sequence. First, air flowed through the first triangular flask containing activated charcoal; then, it was humidified in the second triangular flask containing distilled water. The air was then pumped into the third triangular flask containing ladybug beetles and finally flowed into the last triangular flask containing aphids reared on the leaves. A flow meter controlled the airflow rate at 1.0 L/min. Black cardboard was placed between the two flasks containing ladybugs and aphids to ensure that the aphids could not see the ladybugs and could only sense the volatile odors emitted by them. Prior to each test, one healthy adult ladybug beetle was randomly selected to be placed in the odor source flask and starved for 12 h in advance. 10 aphids were placed in the triangular flask to be exposed to ladybug odors for 12 h.
The effects of PV, PV+PO, and PV+PO+PT on aphids were investigated using a modified double-deck Petri dish system (Figure 1B). Predator visual cues test device: consists of upper and lower Petri dishes. The upper Petri dish contains ladybug beetles, while the lower Petri dish holds aphids. The edges of both dishes are sealed with sealing film. Aphids can see the ladybug beetles in the upper dish through the glass, but cannot perceive the volatile odors of the ladybug beetles. Predator visual and odor cues test device: similarly, it consists of upper and lower Petri dishes, except that the aphids and ladybugs are separated from each other by a metal mesh between the two dishes, allowing the aphids in the lower Petri dish to perceive predator visual and odor cues simultaneously. Predator visual, odor and tactile cues test device: aphids and ladybugs were placed together in one petri dish. Before the test, the mandibles of ladybugs were cut off with a surgical blade, rendering them incapable of feeding on the aphids but not affecting their ability to move around and still have access to the aphids. Meanwhile, aphids were reared individually in separate petri dishes as a control.
Before each experiment, adult ladybugs were starved for 12 h in advance. One ladybug was placed as the predation risk source for each treatment group. 10 freshly laid nymphs (< 6 h old, 1st instar nymphs) or 3rd instar nymphs were exposed to predation risk for 12 h. In contrast, adult aphids (< 6 h after eclosion) were only treated for 6 h to avoid the effects of adult aphid reproduction. After exposure to each predation risk, aphids were individually fed in petri dishes with host leaves; the bottoms of the dishes were moisturized with 0.6% agar, and fresh leaves were replaced daily. The development and survival of each individual was recorded at 12-hour intervals, and reproduction (the number of nymphs in the first 6 days) and adult longevity were recorded after adult emergence. The number of diapause nymphs in the offspring was also recorded. There were a total of 50 initial aphids per treatment. Each treatment was kept at 23 ± 1℃, 65% ± 5% relative humidity, and a 15:9 h (L:D) photoperiod.
2.3. Statistics
Percentage data (proportion of diapause offspring) were arcsin-square root transformed. Data normality was checked with the Shapiro-Wilk test, and variance homogeneity was assessed using Levene’s test. The effects of predator cues on developmental time, total fecundity and the proportion of diapause offspring were analyzed by one-way ANOVA with post-hoc Turkey’s HSD test. The reproductive traits were also analyzed using two-way ANOVA with stressed aphid stage and stress type as factors. All statistical analyses were conducted using SPSS Statistics 25 software.
3. Results
3.1. The Effects of Different Predation Risk on the Development of Aphids
When 1st instar nymphs were exposed to different predator cues, the developmental time was significantly prolonged only during the early stages (1-3) (Table 1). There were significant effects on the developmental duration of the 1st instar nymphs (N1), 2nd instar nymphs (N2), 3rd instar nymphs (N3), and total nymph stages after the exposure of 1st instar nymphs to different predation risks (N1: F4,105=20.904, P<0.001; N2: F4,105=8.072, P<0.001; N3: F4,105=4.611, P=0.002; Total nymph stage: F4,105=24.343, P<0.001). However, the developmental duration of 4th instar nymphs (N4) and adult longevity were not significantly affected (N4: F4,105=1.101, P=0.360; Adult: F4,105=1.956, P=0.107). In particular, the developmental durations of N1, N2 and N3 in the PV+PO+PT group were significantly longer than those in the control and PV groups, but there was no difference between the PV group and the control group. In addition, there was no significant difference in the developmental durations between the PO group and the PV+PO group.
When the 3rd instar nymphs were exposed to different predation cues, only the developmental duration of the 3rd instar nymphs showed significant differences (F4,85=7.518, P<0.001), but there was no effect on the 4th instar nymphs and adults (N4: F4,85=0.290, P=0.883; Adult: F4,85=1.447, P=0.216). The developmental duration of the 3rd instar nymphs in the PV+PO+PT group was significantly longer than those in the control and PV groups, but not significantly different from that of the PO and PV+PO groups (Table 2).
When adult aphids were exposed to different predation cues, all four predation risk treatment groups of ladybug beetles significantly shortened the lifespan of adult aphids (F4,85=8.578, P<0.001). The lifespan of adult aphids in the PV+PO+PT group was significantly shorter than that in the PV and PO groups, but not significantly different from that in the PV+PO group. Although adult longevity in the PO group was not significantly different from that in the PV group, it was significantly shorter in the PV+PO group than in the PV group (Table 3).
3.2. The Effects of Different Predation Risk on the Reproduction of Aphids
The fecundity and proportion of diapause offspring in P. koelreuteriae varied depending on the stage of stressed aphids and stress type (Table 4, Figure 2). Overall, the more mature the aphids were, the less they reproduced, but the greater the proportion of diapause offspring. Regardless of the type of stress, reproduction and the proportion of diapause offspring during adult aphid stress were significantly different from those in the nymph-stress group and the control group (Figure 2).
Regardless of which predation risk cues the 1st instar nymphs were exposed to, their fecundity and the proportion of diapause offspring were not significantly different from that of the control group (Figure 2).
Compared with the control group, the predator visual cues showed no difference in the reproduction of the 3rd instar nymphs; however, the other three predation risk cues significantly reduced reproduction (F4,85=13.844, P<0.001). Additionally, there was no significant difference among these three groups (Figure 2A). Similarly, there was no significant difference in the proportion of diapause offspring between the PV group and the control group. However, the diapause ratio of offspring in the other predation risk groups significantly increased (F4,85=8.158, P<0.001), and there was no significant difference among the three groups (Figure 2B).
When adult aphids were exposed to different predation cues, all four predation risk treatment groups of ladybug beetles significantly reduced their fecundity (F4,85=21.868, P<0.001). There was no significant difference between the PO, PV+PO, and PV+PO+PT groups, but all were significantly lower than the PV group (Figure 2A). Similarly, all four predation risk treatment groups of ladybug beetles significantly increased the diapause ratio of offspring (F4,85=19.364, P<0.001). The PV+PO and PV+PO+PT groups were significantly higher than the PV group, but not significantly different from the PO group (Figure 2B).
4. Discussion
This study indicates that the non-predatory effects of H. axyridis on P. koelreuteriae are related not only to different predation risk cues from ladybug beetles but also to the life stages of aphids under stress. The strongest stress effects for aphids were observed in response to multiple cues from predators, with the stress induced by predator odor cues being stronger than that caused by predator visual cues. Aphids at different developmental stages responded differently to the predation risk. The more mature the aphids were, the more sensitive they became to this risk. The 1st and 3rd instar nymphs did not sense predator visual cues until they reached adulthood. When exposed to predator cues, the development time of nymphs was prolonged, while the adult lifespan was shortened. Additionally, the fecundity of aphids under the stress of predation risk decreased, while the proportion of diapause offspring increased.
When confronted with a predator threat, the anti-predator strategies of prey usually lead to changes in their growth rates [16]. This study showed that the development time of P. koelreuteriae nymphs exposed to predation risk cues was significantly prolonged. The development time of 1st-3rd instar stages was significantly prolonged when 1st instar nymphs were exposed to multiple predator cues; 3rd instar nymphs exhibited developmental delay only when their current developmental stage (3rd instar) was stressed. Studies have shown that the growth rate of insets is not only related to feeding but also to metabolic efficiency. Peach aphids reduce phloem feeding time in the presence of predators [27]. Predation risk can reduce the growth rate of Lestes sponsa larvae by decreasing their food digestion efficiency and metabolic rate [28]. However, there are also contrasting results. For example, butterfly larvae (Pararge aegeria) allocate more energy resources to growth and development in order to develop rapidly when facing predation risks [29]. This difference may be related to energy allocation strategies.
Studies have shown that under predation risk, prey make trade-offs between responding to predation threats and their lifespan [30,31]. In this study, aphid lifespan was shortened when faced with predation risk. The results are in line with previous findings: as predation pressure increases, the development time of Tyrophagus putrescentiae larvae is shortened, while the adult lifespan is also shortened [32]. The lifespan of Spodoptera litura was similarly shortened under bat predation risk [33]. Long-term exposure to the predation threat from Menochilus sexmaculatus significantly shortened the adult lifespan of cotton aphids (Aphis gossypii) [34]. Additionally, the adult lifespan of Bactrocera dorsalis exposed to the predation risk from Hierodula patellifera was also shortened [35]. These results suggest that adults may devote more resources and energy to defense rather than development, thereby altering their developmental time.
Many studies have also shown a trade-off between predation risk and reproduction. When prey reallocates more energy to risk-avoidance behaviors than to reproduction, the non-consumptive effects of predation reduce prey abundance. Both 3rd instar nymphs and adults in this study exhibited reduced fecundity under predation pressure. This result is consistent with our initial expectation that investment in offspring would be diminished by the detection of predation risk. Several previous studies have shown that prey experiencing predation risk reduced reproduction. For example, M. persicae exhibited a 25% reduction in population size when peach aphids decreased the time spent feeding on phloem in the presence of H. axyridis odor cues [27]. Phytophagous mites, such as T. putrescentiae, under predation pressure, reduce fecundity and allocate resources to anti-predation strategies [36]. Sitobion miscanthi also reduced net fecundity due to the presence of isolated predators [37]. Female B. tryoni lay fewer eggs in the presence of predator olfactory cues [38]. However, there is also an increase in predation risk to prey reproduction. Exposing the potato aphid Macrosiphum euphorbiae Thomas to Hippodamia convergens Guérin-Méneville, which were rendered non-lethal by mouthpart manipulation, resulted in significantly higher numbers of nymphs [39]. Female B. dorsalis treated for predation risk had a reduced developmental time but a higher reproductive rate than the control treatment [35]. When threatened by long-term predation, the lifespan of the aphid Rhopalosiphum padi was shortened, but the reproduction rate increased [40]. This may be due to the habituation of prey to predator cues, moderating the effect of risk on prey reproduction.
There is growing evidence that maternal exposure to predation risk affects the phenotype of prey offspring [40]. Prey mothers exposed to predation risk sometimes alter the morphology and/or anti-predator behaviors of their offspring to reduce predation risk. In the presence of predator risk, pea aphids (Acyrthosiphon pisum) and peach aphids (M. persicae) increase the production of winged aphid offspring [26,41]. The reproductive capacity of this morphotype is lower than that of the wingless offspring. It is mainly responsible for long-distance transmission and enhances the ability of offspring to evade predators [42]. In this study, compared with the control treatment, aphids exposed to predation risk increased the proportion of diapause offspring. Diapause aphids differed significantly from normal aphids in appearance and behavior, with diapause aphids having transparent bodies and being sedentary (neither eating nor moving) at the edges of leaves, while normal aphids had black speckles on their bodies and were active feeders in the centers of leaves. Compared with normal aphids, diapause aphids have a lower probability of being preyed upon by predators. In fact, caterpillars are more vulnerable to predation when actively feeding than when stationary [43]. The increase in the proportion of diapause offspring suggests transgenerational plasticity regulation. Although it is unclear how the condition information about conditions experienced by mothers is transmitted to offspring, several studies have focused on epigenetic mechanisms [44,45]. Future studies using molecular approaches may reveal the mechanisms by which predators exert transgenerational effects on the morphology and behavior of aphid offspring.
This study confirmed the dominant role of chemical cues in risk perception. The stress effect of odor cues alone was significantly stronger than that of visual cues, which is consistent with the conclusions of most aphid studies. Volatile and contact chemical cues play a decisive role in whether aphids choose to settle on plants [46]. Ladybug tracks left on leaves cause aphid avoidance and reduce host plant colonization by aphids through contact and olfactory cues, or a combination of both [47]. Vision plays a central role in predator detection in most animals active in light environments, even those with simple visual systems [48]. Several studies have shown that aphids use vision to detect approaching predators. Behavioral responses such as twitching, kicking, crouching, and dropping from the plant have been observed in studies testing aphid responses to artificial visual stimuli or passing shadows [49,50,51]. However, it has recently been found that visual cues (hand-held objects and expanding shapes on the screen) can only elicit aphid antennal movements, but not defense responses [52]. Due to their low visual acuity, aphids may struggle to reliably recognize visual predators [53]. Visual cues may be the least important way for aphids to detect the approach of a predator. Notably, visual cues, when acting alone, affected only adult aphids in this study, suggesting that there may be developmental stage-specific signal integration mechanisms for their risk perception.
Although detecting moments of predator contact relies on the sense of touch, aphids detecting distant approaching predators must rely on the perception of visual, chemical, and plant-borne vibrational cues. Pea aphids can perceive both predators and non-predators through vibrational and visual cues, utilizing multimodal cues to detect approaching seven-spotted ladybug predators [52]. The major breakthrough of this study is the revelation of the synergistic effect of multimodal signals, with the combination of odor, visual and touch cues having the strongest effect. The mechanism by which aphids detect predators may involve a complex interaction among multiple sensory cues. We cannot accurately determine the integration of information from multiple sensory modalities by aphids. This is because it is inherently challenging for any animal to gain insight into multisensory integration and risk assessment through behavioral assays [54]. Animals may also adjust their responses according to their internal state [55,56], and the absence of a response does not necessarily mean that the animal has failed to detect a certain cue [54].
This study revealed a significant developmental stage dependence of predation risk cues on the regulation of development time in P. koelreuteriae. 1st instar nymphs only prolonged their developmental time in the early stages (1st-3rd instars) when exposed to multiple predator cues. 3rd instar nymphs showed a developmental delay only when their current developmental stage (3rd instar) was stressed, while 4th instar nymphs and adults were not significantly affected. Adult aphids exhibited a comprehensive stress response. The increased sensitivity of adult aphids to risk cues may reflect evolutionary adaptations. According to dynamic energy budget theory [57], adult aphids have completed their developmental investment and can allocate more resources to stress resistance responses. This is consistent with the shortened lifespan and reproduction suppression, but increased diapause offspring under stress in this study. Pea aphids no longer remember their experience 60 min after exposure to cues from conspecifics and ladybird beetles, and they are no longer responsive to predation risk [58]. In fact, animal memory tends to disappear over time [59,60]. Further research is needed to clarify how the priming effect of predator risk stress dissipates.
5. Conclusions
This study systematically reveals the multilevel regulation of aphid life history strategies by the non-consumption effects of predation risk, providing new insights into the ecological regulation of pests. For example, artificial risk cues were released during the sensitive developmental stage (adult aphid stage) for biological control. The main limitations of this study include the failure to quantify the stress intensity of different cues and the inability to detect transgenerational effects. It is recommended that follow-up studies combine behavioral ecology and epigenetic approaches to explore the dose-response relationships and transgenerational memory mechanisms of risk cues. Additionally, one caveat of the current study is that it was conducted in small arenas, resulting in highly concentrated cues and limited space, which may not accurately represent this system in nature.
Author Contributions
Conceptualization, H.Y., and Z.H.; methodology, H.Y. and L.W.; formal analysis, H.Y. and J.D.; investigation, J.D. and L.W; data curation, H.Y. and P.Z.; writing—original draft preparation, H.Y. and J.D.; writing—review and editing, H.Y., J.H., Z.H. and D.L.; supervision, H.Y. and D.L.; project administration, H.Y.; funding acquisition, H.Y., and Z.H. All authors have read and agreed to the published version of the manuscript.
Funding
The research was supported by the National Natural Science Foundation of China (31901872), the Key Science and Technology Program of Henan Province (232102111032, 232301420028), and the Young Backbone Teachers Project of Henan University of Science and Technology (2023).
Data Availability Statement
The data presented in this study are available in the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Orrock, J.L.; Grabowski, J.H.; Pantel, J.H.; Peacor, S.D.; Peckarsky, B.L.; Sih, A.; Werner, E.E. Consumptive and nonconsumptive effects of predators on metacommunities of competing prey. Ecology 2008, 89, 2426–2435. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, E.G.; Johnson, C.N. Predator interactions, mesopredator release and biodiversity conservation. Ecology Letters 2009, 12, 982–998. [Google Scholar] [CrossRef] [PubMed]
- Hawlena, D.; Schmitz, O.J. Herbivore physiological response to predation risk and implications for ecosystem nutrient dynamics. Proc. Natl. Acad. Sci. U.S.A. 2010, 107, 15503–15507. [Google Scholar] [CrossRef] [PubMed]
- Culshaw-Maurer, M.; Sih, A.; Rosenheim, J.A. Bugs scaring bugs: enemy-risk effects in biological control systems. Ecology Letters 2020, 23, 1693–1714. [Google Scholar] [CrossRef]
- Peacor, S.D.; Peckarsky, B.L.; Trussell, G.C.; Vonesh, J.R. Costs of predator-induced phenotypic plasticity: A graphical model for predicting the contribution of nonconsumptive and consumptive effects of predators on prey. Oecologia 2013, 171, 1–10. [Google Scholar] [CrossRef]
- Preisser, E.L.; Bolnick, D.I.; Benard, M.F. Scared to death? The effects of intimidation and consumption in predator–prey interactions. Ecology 2005, 86, 501–509. [Google Scholar] [CrossRef]
- Kral, K. Visually guided search behavior during walking in insects with different habitat utilization strategies. J Insect Behav 2019, 32, 290–305. [Google Scholar] [CrossRef]
- Zanuzzo, F.S.; de, C. Bovolato, A.L.; Pereira, R.T.; Valença-Silva, G.; Barcellos, L.J.G.; Barreto, R.E. Innate response based on visual cues of sympatric and allopatric predators in Nile tilapia. Behavioural Processes 2019, 164, 109–114. [Google Scholar] [CrossRef]
- Grubb, T.C. Antipredator defenses in birds and mammals. The Auk 2006, 123, 601–605. [Google Scholar] [CrossRef]
- Munoz, N.E.; Blumstein, D.T. Multisensory perception in uncertain environments. Behavioral Ecology 2012, 23, 457–462. [Google Scholar] [CrossRef]
- Hettena, A.M.; Munoz, N.; Blumstein, D.T. Prey responses to predator’s sounds: A review and empirical study. Ethology 2014, 120, 427–452. [Google Scholar] [CrossRef]
- Hermann, S.L.; Thaler, J.S. Prey perception of predation risk: Volatile chemical cues mediate non-consumptive effects of a predator on a herbivorous insect. Oecologia 2014, 176, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.E.; Magnavacca, G. Predator inspection behaviour in a characin fish: an interaction between chemical and visual information? Ethology 2003, 109, 739–750. [Google Scholar] [CrossRef]
- Werner, E.E.; Peacor, S.D. A review of trait-mediated indirect interactions in ecological communities. Ecology 2003, 84, 1083–1100. [Google Scholar] [CrossRef]
- Koch, N.; Lynch, B.; Rochette, R. Trade-off between mating and predation risk in the marine snail, littorina plena. Invertebrate Biology 2007, 126, 257–267. [Google Scholar] [CrossRef]
- Urban, M.C. The growth–predation risk trade-off under a growing gape-limited predation threat. Ecology 2007, 88, 2587–2597. [Google Scholar] [CrossRef]
- Li, Y.P.; Ge, F. Effects of predatory stress imposed by Harnonia axyridis on the development and fecundity of Drosophila melanogaster. Chinese Journal of Ecology 2010, 29, 554–559. [Google Scholar] [CrossRef]
- Laforsch, C.; Tollrian, R. Inducible defenses in multipredator environments: Cyclomorphosis in Daphnia cucullata. Ecology 2004, 85, 2302–2311. [Google Scholar] [CrossRef]
- Miller, T.; Crossley, M.S.; Fu, Z.; Meier, A.R.; Crowder, D.W.; Snyder, W.E. Exposure to predators, but not intraspecific competitors, heightens herbivore susceptibility to entomopathogens. Biological Control 2020, 151, 104403. [Google Scholar] [CrossRef]
- Siepielski, A.M.; Fallon, E.; Boersma, K. Predator olfactory cues generate a foraging–predation trade-off through prey apprehension. R. Soc. open sci. 2016, 3, 150537. [Google Scholar] [CrossRef]
- Zhang, G.X.; Zhong, T.S. Economic Insect Fauna of China, Fasc. 25. Homoptera: Aphidomorpha. 25th ed.; Science Press, Beijing, 1983.
- Li, D.X.; Ren, J.; Du, D.; Wang, Q.C. Life tables of the laboratory population of Periphyllus koelreuteriae (Hemiptera: Chaitophoridae) at different temperatures. Acta Entomolohica Sinica 2015, 58, 154–159. [Google Scholar] [CrossRef]
- Wang, N.C.; Li, Z.H.; Liu, G.L.; Ye, B.H.; Dong, J.X.; Ren, P. Studies on the bionomics of Periphyllus koelreuteriae and its control. Journal of Shandong Agricultural University 1990, 21, 47–50. [Google Scholar]
- Gu, P.; Zhou, L.Q.; Xu, Z. Biological characteristics and control of Periphyllus koelreuteriae (Takahashi) in Shanghai district. Journal of Shanghai Jiaotong University 2004, 22, 389–392. [Google Scholar]
- Michaud, J.P. Coccinellids in biological control. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); Wiley, 2012; pp. 488–519 ISBN 978-1-4051-8422-9.
- Hermann, S.L.; Bird, S.A.; Ellis, D.R.; Landis, D.A. Predation risk differentially affects aphid morphotypes: Impacts on prey behavior, fecundity and transgenerational dispersal morphology. Oecologia 2021, 197, 411–419. [Google Scholar] [CrossRef]
- Kansman, J.T.; Nersten, C.E.; Hermann, S.L. Smelling danger: lady beetle odors affect aphid population abundance and feeding, but not movement between plants. Basic and Applied Ecology 2023, 71, 1–8. [Google Scholar] [CrossRef]
- Stoks, R. Food Stress and predator-induced stress shape developmental performance in a damselfly. Oecologia 2001, 127, 222–229. [Google Scholar] [CrossRef]
- Gotthard, K. Increased risk of predation as a cost of high growth rate: an experimental test in a butterfly. Journal of Animal Ecology 2000, 69, 896–902. [Google Scholar] [CrossRef]
- Duong, T.M.; McCauley, S.J. Predation risk increases immune response in a larval dragonfly (Leucorrhinia intacta). Ecology 2016, 97, 1605–1610. [Google Scholar] [CrossRef]
- Schwenke, R.A.; Lazzaro, B.P.; Wolfner, M.F. Reproduction–immunity trade-offs in insects. Annual Review of Entomology 2016, 61, 239–256. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, Z.-Q. Level-dependent effects of predation stress on prey development, lifespan and reproduction in mites. Biogerontology 2022, 23, 515–527. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Y.; Wang, Z.; Lin, T.; Feng, J.; Jiang, T. Effects of predation risks of bats on the growth, development, reproduction, and hormone levels of Spodoptera litura. Front. Ecol. Evol. 2023, 11, 1126253. [Google Scholar] [CrossRef]
- Lin, X.; Cui, X.; Tang, J.; Zhu, J.; Li, J. Predation risk effects of lady beetle Menochilus sexmaculatus (Fabricius) on the melon aphid, aphis Gossypii glover. Insects 2024, 15, 13. [Google Scholar] [CrossRef]
- Liu, X.; Wen, J.; Geng, X.; Xiao, L.; Zou, Y.; Shan, Z.; Lu, X.; Fu, Y.; Fu, Y.; Cao, F. The impact of predation risks on the development and fecundity of Bactrocera dorsalis Hendel. Insects 2024, 15, 322. [Google Scholar] [CrossRef]
- Wei, X.; Liu, J.; Zhang, Z.-Q. Predation stress experienced as immature mites extends their lifespan. Biogerontology 2023, 24, 67–79. [Google Scholar] [CrossRef]
- Wang, L.; Atlihan, R.; Chai, R.; Dong, Y.; Luo, C.; Hu, Z. Assessment of non-consumptive predation risk of Coccinella septempunctata (Coleoptera: Coccinellidae) on the population growth of Sitobion miscanthi (Hemiptera: Aphididae). Insects 2022, 13, 524. [Google Scholar] [CrossRef] [PubMed]
- Kempraj, V.; Park, S.J.; Taylor, P.W. Forewarned is forearmed: Queensland fruit flies detect olfactory cues from predators and respond with predator-specific behaviour. Sci Rep 2020, 10, 7297. [Google Scholar] [CrossRef]
- Kersch-Becker, M.F.; Thaler, J.S. Plant resistance reduces the strength of consumptive and non-consumptive effects of predators on aphids. Journal of Animal Ecology 2015, 84, 1222–1232. [Google Scholar] [CrossRef]
- Tariel, J.; Plénet, S.; Luquet, É. Transgenerational plasticity in the context of predator-prey interactions. Front. Ecol. Evol. 2020, 8, 548660. [Google Scholar] [CrossRef]
- Hirano, A.; Yoshida, T.; Choh, Y. Maternal exposure to predation risk increases winged morph and antipredator dispersal of the pea aphid, Acyrthosiphon pisum (Hemiptera: Aphididae). Appl Entomol Zool 2022, 57, 227–235. [Google Scholar] [CrossRef]
- Huberty, A.F.; Denno, R.F. Trade-off in investment between dispersal and ingestion capability in phytophagous insects and its ecological implications. Oecologia 2006, 148, 226–234. [Google Scholar] [CrossRef]
- Bernays, E.A. Feeding by lepidopteran larvae is dangerous. Ecological Entomology 1997, 22, 121–123. [Google Scholar] [CrossRef]
- Duempelmann, L.; Skribbe, M.; Bühler, M. Small RNAs in the transgenerational inheritance of epigenetic information. Trends in Genetics 2020, 36, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Norouzitallab, P.; Baruah, K.; Vanrompay, D.; Bossier, P. Can epigenetics translate environmental cues into phenotypes? Science of The Total Environment 2019, 647, 1281–1293. [Google Scholar] [CrossRef]
- Webster, B. The role of olfaction in aphid host location. Physiological Entomology 2012, 37, 10–18. [Google Scholar] [CrossRef]
- Ninkovic, V.; Feng, Y.; Olsson, U.; Pettersson, J. Ladybird footprints induce aphid avoidance behavior. Biological Control 2013, 65, 63–71. [Google Scholar] [CrossRef]
- Cronin, T.W.; Godin, J.G.J. The visual ecology of predator-prey interactions. In: Barbosap, castellanosi (eds) ecology of predator-prey interactions. Oxford University Press: New York, 2005. [Google Scholar]
- Dixon, A.F.G.; McKay, S. Aggregation in the sycamore aphid Drepanosiphum platanoides (Schr.) (Hemiptera: Aphididae) and its relevance to the regulation of population growth. The Journal of Animal Ecology 1970, 39, 439. [Google Scholar] [CrossRef]
- Hartbauer, M. Collective defense of Aphis Nerii and Uroleucon hypochoeridis (Homoptera, Aphididae) against natural enemies. PLoS ONE 2010, 5, e10417. [Google Scholar] [CrossRef]
- Ben-Ari, M.; Inbar, M. Aphids link different sensory modalities to accurately interpret ambiguous cues. Behavioral Ecology 2014, 25, 627–632. [Google Scholar] [CrossRef]
- Gish, M. Aphids detect approaching predators using plant-borne vibrations and visual cues. J Pest Sci 2021, 94, 1209–1219. [Google Scholar] [CrossRef]
- Döring, T.F.; Spaethe, J. Measurements of eye size and acuity in aphids (Hemiptera: Aphididae). Entomol Gen 2009, 32, 77–84. [Google Scholar] [CrossRef]
- Lima, S.L.; Steury, T.D. Perception of predation risk: the foundation of nonlethal predator-prey interactions. In: Ecology Of Predator-Prey Interactions; Barbosa, P., Castellanos, I., Eds.; Oxford University PressNew York, NY, 2005; pp. 166–188 ISBN 978-0-19-517120-4.
- Herberholz, J.; Marquart, G.D. Decision making and behavioral choice during predator avoidance. Front. Neurosci. 2012, 6. [Google Scholar] [CrossRef] [PubMed]
- Schadegg, A.C.; Herberholz, J. Satiation level affects anti-predatory decisions in foraging juvenile crayfish. J Comp Physiol A 2017, 203, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Kooijman, S.A.L.M. Dynamic Energy Budget Theory for Metabolic Organisation; 3rd ed.; Cambridge university press: Cambridge, 2010; ISBN 978-0-521-13191-9.
- Tamai, K.; Choh, Y. Previous exposures to cues from conspecifics and ladybird beetles prime antipredator responses in pea aphids Acyrthosiphon pisum (Hemiptera: Aphididae). Appl Entomol Zool 2019, 54, 277–283. [Google Scholar] [CrossRef]
- Brown, G.E.; Ferrari, M.C.O.; Malka, P.H.; Oligny, M.-A.; Romano, M.; Chivers, D.P. Growth rate and retention of learned predator cues by juvenile rainbow trout: faster-growing fish forget sooner. Behav Ecol Sociobiol 2011, 65, 1267–1276. [Google Scholar] [CrossRef]
- Ferrari, M.C.O.; Vrtělová, J.; Brown, G.E.; Chivers, D.P. Understanding the role of uncertainty on learning and retention of predator information. Anim Cogn 2012, 15, 807–813. [Google Scholar] [CrossRef]
Figure 1.
Experimental apparatus used to detect aphid response to predator cues. A: Aphid response to predator odor. B: Aphid responses to predator visual, odor, and tactile cues. PO: predator odor cues; PV: predator visual cues; PV+PO: predator visual and odor cues; PV+PO+PT: predator visual, odor, and tactile cues
Figure 1.
Experimental apparatus used to detect aphid response to predator cues. A: Aphid response to predator odor. B: Aphid responses to predator visual, odor, and tactile cues. PO: predator odor cues; PV: predator visual cues; PV+PO: predator visual and odor cues; PV+PO+PT: predator visual, odor, and tactile cues
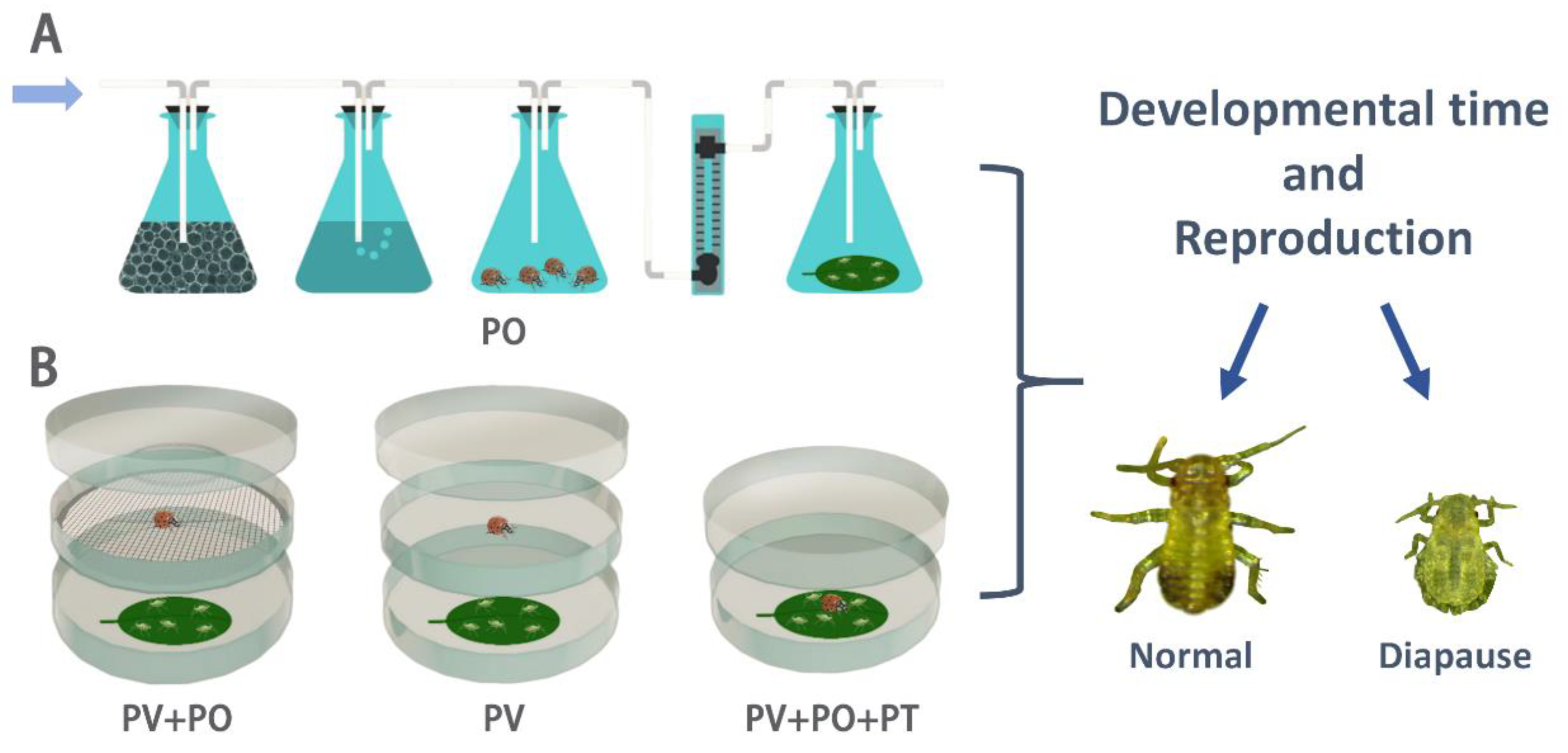
Figure 2.
Fecundity (A) and proportion of diapause offspring (B) of Periphyllus koelreuteriae after exposure to predator cues. Different letters above the bars indicate significant differences between different treatments (p < 0.05), based on the result of Tukey’s HSD.
Figure 2.
Fecundity (A) and proportion of diapause offspring (B) of Periphyllus koelreuteriae after exposure to predator cues. Different letters above the bars indicate significant differences between different treatments (p < 0.05), based on the result of Tukey’s HSD.
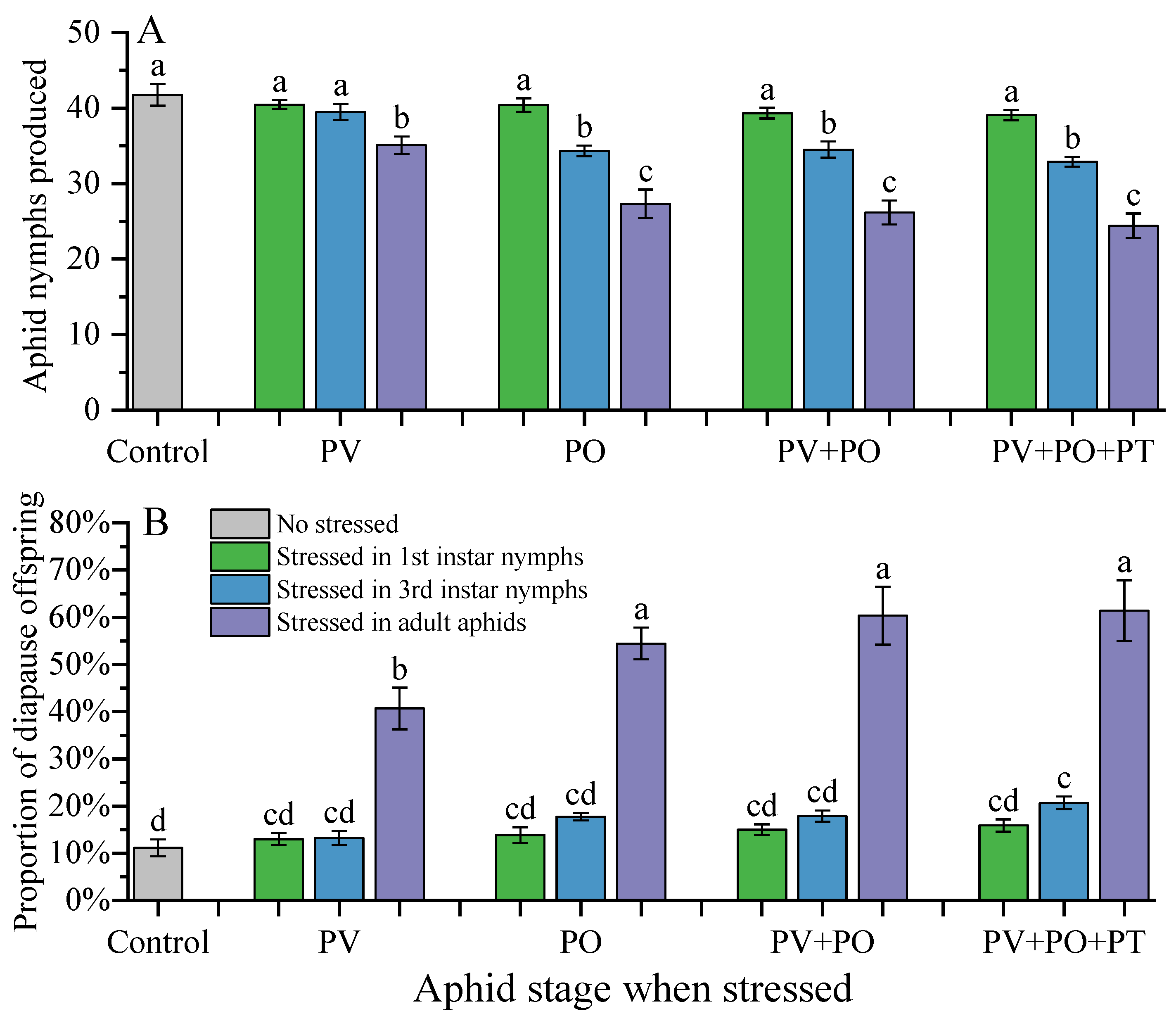

Table 1.
Developmental time of each insect stage when 1st instar nymphs were exposed to different predator cues.
Table 1.
Developmental time of each insect stage when 1st instar nymphs were exposed to different predator cues.
| Predator risk | N1 | N2 | N3 | N4 | Total nymph stage | Adult longevity |
| CK | 3.14 ± 0.13 c | 2.05 ± 0.09 c | 1.82 ± 0.09 bc | 1.85 ± 0.09 a | 8.84 ± 0.20 c | 9.72 ± 0.15 a |
| PV | 3.20 ± 0.11 c | 2.09 ± 0.08 c | 1.87 ± 0.09 bc | 1.86 ± 0.09 a | 9.03 ± 0.24 c | 9.34 ± 0.21 a |
| PO | 3.64 ± 0.15 b | 2.39 ± 0.09 b | 2.11 ± 0.07 ab | 1.95 ± 0.09 a | 10.09 ± 0.23 b | 9.95 ± 0.34 a |
| PV+PO | 3.65 ± 0.12 b | 2.43 ± 0.10 ab | 2.14 ± 0.08 ab | 2.02 ± 0.11 a | 10.25 ± 0.20 b | 9.33 ± 0.37 a |
| PV+PO+PT | 4.66 ± 0.16 a | 2.66 ± 0.09 a | 2.24 ± 0.08 a | 2.09 ± 0.12 a | 11.65 ± 0.26 a | 8.86 ± 0.35 a |
CK: control; PV: predator visual cues; PO: predator odor cues; PV+PO: predator visual and odor cues; PV+PO+PT: predator visual, odor, and tactile cues; N1: 1st instar nymph; N2: 2nd instar nymph; N3: 3rd instar nymph; N4: 4th instar nymph. Different letters within the same column indicated significant differences between the treatments (p < 0.05), based on the result of Tukey’s HSD.
Table 2.
Developmental time of each insect stage when 3rd instar nymphs were exposed to different predator cues.
Table 2.
Developmental time of each insect stage when 3rd instar nymphs were exposed to different predator cues.
| Predator risk | N3 | N4 | Adult longevity |
| CK | 1.89 ± 0.10 c | 1.83 ± 0.11 a | 9.75 ± 0.18 a |
| PV | 2.00 ± 0.09 bc | 1.86 ± 0.11 a | 9.03 ± 0.20 a |
| PO | 2.31 ± 0.08 ab | 1.92 ± 0.07 a | 9.08 ± 0.27 a |
| PV+PO | 2.33 ± 0.11 ab | 1.94 ± 0.09 a | 9.47 ± 0.31 a |
| PV+PO+PT | 2.56 ± 0.11 a | 1.94 ± 0.08 a | 9.14 ± 0.28 a |
CK: control; PV: predator visual cues; PO: predator odor cues; PV+PO: predator visual and odor cues; PV+PO+PT: predator visual, odor, and tactile cues; N3: 3rd instar nymph; N4: 4th instar nymph. Different letters within the same column indicated significant differences between the treatments (p < 0.05), based on the result of Tukey’s HSD.
Table 3.
The influence of adult aphids exposed to different predator cues on their lifespan.
| Predator risk | Adult longevity |
| CK | 9.75 ± 0.18 a |
| PV | 8.78 ± 0.31 b |
| PO | 8.25 ± 0.22 bc |
| PV+PO | 7.61 ± 0.23 cd |
| PV+PO+PT | 7.22 ± 0.29 d |
CK: control; PV: predator visual cues; PO: predator odor cues; PV+PO: predator visual and odor cues; PV+PO+PT: predator visual, odor, and tactile cues. Different letters within the same column indicated significant differences between the treatments (p < 0.05), based on the result of Tukey’s HSD.
Table 4.
Two-way ANOVA for the effects of different treatments on fecundity and proportion of diapause offspring in Periphyllus koelreuteriae.
Table 4.
Two-way ANOVA for the effects of different treatments on fecundity and proportion of diapause offspring in Periphyllus koelreuteriae.
| Trait | source | df | Mean-Square Value (MS) | F-Values | p |
| Fecundity | Stressed aphid stage | 2 | 1625.215 | 104.908 | <0.001 |
| Stress type | 3 | 259.581 | 16.756 | <0.001 | |
| Stressed aphid stage× Stress type | 6 | 55.928 | 3.610 | 0.002 | |
| Error | 132 | 15.492 | |||
| Proportion of diapause offspring | Stressed aphid stage | 2 | 2.367 | 191.616 | <0.001 |
| Stress type | 3 | 0.077 | 6.238 | 0.001 | |
| Stressed aphid stage× Stress type | 6 | 0.023 | 1.845 | 0.095 | |
| Error | 132 | 0.012 |
“×” is an interactive token. Significant values (p < 0.05) are printed in bold.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



