
Submitted:
22 May 2025
Posted:
23 May 2025
You are already at the latest version
Abstract
Climate change is reshaping plant reproductive processes, particularly at the vulnerable seed germination stage. This study examines the germination responses of four Actinidia species (A. rufa, A. latifolia, A. deliciosa, and A. setosa) under controlled experimental conditions, integrating empirical germination data with classifier modeling to predict species-specific responses under future climate scenarios. Unlike traditional species distribution models (SDMs), our classifier approach incorporates physiological dormancy mechanisms and key environmental cues such as chilling requirements, temperature fluctuations, and drought stress. Results reveal significant interspecific differences: A. rufa exhibited strong ecological plasticity, maintaining stable germination under warming and drought, while A. deliciosa displayed extreme sensitivity to warming, with germination dropping below 25% due to its strict chilling requirement. A. latifolia showed latitude-dependent vulnerability, with southern populations experiencing reduced germination under warming conditions, and A. setosa demonstrated complex dormancy patterns with higher germination at high elevations. The predictive accuracy of our models was validated against long-term field data, underscoring their robustness in forecasting climate-induced germination shifts. These findings highlight the need for targeted breeding programs to develop A. deliciosa cultivars with reduced chilling requirements and suggest A. rufa as a strong candidate for ecological restoration under future warming scenarios. By refining climate impact assessments through physiological modeling, this study provides valuable insights for kiwifruit conservation, agricultural adaptation, and broader plant-climate interactions under global warming.
Keywords:
Climate adaptation
; seed dormancy
; phenological modeling
; ecological resilience
; classifier modeling
; kiwifruit conservation
; Actinidia species
; global warming impact
; bioclimatic stress responses
1. Introduction
Anthropogenic climate change is fundamentally reshaping global ecosystems, driving shifts in species distributions, altering phenological patterns, and disrupting ecological dynamics [1,2]. Since the onset of the Industrial Revolution, rising atmospheric greenhouse gas concentrations have led to significant increases in temperature, alterations in precipitation regimes, and more frequent extreme weather events, imposing complex stressors on plant communities [3,4,5]. Among these challenges, changes in seasonal temperature regimes pose severe threats to plant reproductive success, particularly during the critical stages of seed dormancy release and germination [6,7]. Understanding how plant species respond to climate-driven stressors is essential for predicting future distribution patterns and developing adaptive strategies for conservation and agriculture [8,9].
Seed germination is one of the most climate-sensitive phases in a plant’s life cycle, directly influencing population persistence, recruitment success, and ecosystem resilience under shifting environmental conditions [10,11,12,13]. While significant research has focused on the impact of climate change on flowering and fruiting phenology [14], relatively little attention has focused on climate-induced alterations in seed dormancy mechanisms and germination dynamics [15]. Temperature shifts may disrupt dormancy-breaking cues, alter stratification requirements, and impose physiological constraints on seed viability [16], ultimately affecting species survival and community structure [17]. Given the ecological and economic importance of seed-based regeneration, a deeper understanding of species-specific germination responses to climate variability is urgently needed.
Traditional species distribution models (SDMs) have been widely employed to predict plant responses to climate change based on ecological niche theory [18,19]. However, most SDMs rely on macroclimatic variables and often fail to incorporate species-specific physiological traits [20], such as chilling requirements, dormancy-breaking thresholds, and microhabitat preferences. This limitation can lead to overestimations or underestimations of species’ climatic tolerances and adaptive capacities [8]. Addressing this gap requires an integrative modeling approach that combines empirical germination data with statistical classification frameworks to improve the accuracy of climate impact assessments.
The genus Actinidia (kiwifruit) is an ecologically and commercially significant taxon, widely distributed across East and Southeast Asia [21,22]. Taiwan hosts several indigenous Actinidia species alongside the widely cultivated A. deliciosa (A. Chev.) C.F. Liang & A.R. Ferguson [23,24], making this genus an ideal model for investigating species-specific seed germination responses to climate change. These species occupy diverse altitudinal and latitudinal gradients, allowing for a comprehensive examination of how environmental factors such as winter warming, drought stress, and elevation-dependent dormancy shape germination patterns [25]. Despite their ecological importance, the effects of climate change on Actinidia seed germination remain poorly understood, particularly concerning chilling requirements, dormancy mechanisms, and their resilience to warming and drought conditions.
This study aims to address these knowledge gaps by systematically investigating the seed germination responses of four Actinidia species (Actinidia rufa (Siebold & Zucc.) Planch. ex Miq., A. latifolia (Gardner & Champ.) Merr., A. deliciosa, and A. setosa (H.L. Li) C.F. Liang & A.R. Ferguson) under controlled experimental conditions. By integrating empirical germination data with classifier modeling, we simulate germination dynamics under projected climate scenarios, focusing on temperature-dependent dormancy shifts, drought stress tolerance, and elevational and latitudinal variations in germination success. Our study presents a novel application of classifier models, providing a quantitative framework for assessing species-specific resilience and vulnerability to climate stressors.
By refining our understanding of climate-driven germination responses, this study contributes to the broader field of plant ecophysiology, climate adaptation, and conservation biology. The findings are expected to inform conservation strategies for wild Actinidia populations, guide breeding programs for climate-resilient kiwifruit cultivars, and enhance predictive models of species distributions under future warming scenarios. As global temperatures continue to rise, integrating physiological and ecological modeling approaches will be critical for developing adaptive management strategies to mitigate the impacts of climate change on plant reproductive success.
2. Materials and Methods
2.1. Study Species and Seed Collection
The genus Actinidia (kiwifruit) is widely distributed across East and Southeast Asia, encompassing species with diverse ecological adaptations. This study focuses on four Actinidia species—A. rufa, A. latifolia, A. deliciosa, and A. setosa—to investigate interspecific variation in seed dormancy and germination responses under projected climate change scenarios. These species were selected due to their broad ecological range, economic significance, and distinct physiological adaptations to environmental stressors, making them ideal models for assessing climate-induced shifts in germination dynamics.
Mature fruits were collected during the peak fruiting season (September to November) from natural forests and cultivated orchards across Taiwan. To capture the full range of environmental variability, seeds were sampled across different elevations and latitudes:
- A. setosa: Siyuan Yakou, Yilan County (1,870 m); Cuifeng, Nantou County (2,250 m)
- A. deliciosa: Shengguang, Heping District, Taichung City (2,500 m)
- A. rufa: Near Qingjing Farm, Nantou County (1,800 m)
- A. latifolia: Beidongyan Mountain, Nantou County (1,600 m); Wufeng, Taichung County (200 m); Mudan Township, Pingtung County (400 m)
Fruits were stored at ambient temperature until fully ripened. Seeds were extracted by pulp removal, and non-viable seeds were discarded using the flotation method. The remaining viable seeds were air-dried at 25°C for 48 hours and subsequently stored in airtight containers at 5°C with 33% relative humidity (RH) until further experimentation.
2.2. Experimental Design
To assess seed dormancy and germination responses, a controlled growth chamber experiment was conducted. Three primary environmental stressors—cold stratification, temperature fluctuation, and drought stress—were examined through a series of factorial germination trials.
2.2.1. Cold Stratification Treatments
To evaluate the influence of chilling duration on dormancy release, seeds were subjected to five cold stratification periods (0, 2, 4, 8, and 12 weeks) at 5°C in moist sphagnum moss. Following stratification, seeds were sown in Petri dishes lined with moistened filter paper and incubated at 25/15°C (12-h photoperiod, 60–80 µmol m⁻² s⁻¹ photon irradiance).
2.2.2. Temperature and Warming Treatments
To simulate the impact of projected climate warming in a subtropical region, seeds were incubated under four temperature regimes reflecting anticipated climate scenarios:
- No warming (15/6°C): This serves as the control group, representing the current or baseline climate conditions. It allows researchers to compare how seed germination performs under the current climate compared to warmer scenarios.
- Mild warming (20/10°C): This level simulates a slight increase in temperature, reflecting a potential future climate scenario with minimal warming.
- Moderate warming (25/15°C): This represents a more significant temperature increase, corresponding to a scenario with moderate climate warming.
- Extreme warming (30/20°C): This level simulates a substantial temperature rise, indicative of a future climate with significant warming. It helps researchers understand how seeds might respond to more extreme temperature conditions.
2.2.3. Drought Stress Treatments
To assess the effects of moisture limitation on germination, seeds were subjected to different dry storage durations before rehydration and germination testing:
- Mild drought: 2 weeks of dry storage (RH 33%)
- Moderate drought: 4–8 weeks of dry storage
- Severe drought: ≥12 weeks of dry storage
After the designated storage period, seeds were rehydrated and incubated under the control temperature regime (25/15°C) to assess germination recovery.
2.3. Germination Assessment and Data Collection
Germination was monitored weekly for 8 weeks, and a seed was recorded as germinated upon radicle emergence (>2 mm). Each treatment was replicated three to four times, with 50–100 seeds per replicate.
To minimize experimental bias:
- Germination conditions were maintained in a controlled-environment growth chamber (relative humidity: 70%; light/dark cycle: 12/12 h; photon irradiance: 60–80 µmol m⁻² s⁻¹).
- Watering and seed hydration conditions were standardized to prevent variation in moisture levels.
- Germination percentages were calculated as:
2.4. Classifier Modeling and Model Validation
A logistic regression-based classifier model was developed using R version 4.3.3 [26] to assess the probability of seed germination under different environmental conditions. Logistic regression is particularly suited for binary outcome prediction (germination: 1, non-germination: 0), allowing for robust quantification of species-specific germination responses to climate-driven stressors.
2.4.1. Model Structure and Predictors
The general form of the logistic regression classifier model is:
where g(germination) is the probability of germination, Xi represents explanatory variables (e.g., temperature, chilling duration, altitude, latitude, drought duration), and βi are the estimated coefficients.
Six logistic regression models were developed to examine specific germination drivers:
- Chilling Duration Model: Influence of cold stratification on germination success.
- Winter Warming Model: Effect of increasing temperature on A. setosa, A. deliciosa, and A. rufa germination.
- Altitudinal Model: Relationship between germination percentage and elevation for A. setosa and A. latifolia.
- Latitudinal Model: Impact of latitude on A. latifolia germination under warming conditions.
- Drought Model: Effect of dry storage duration on germination probability for A. latifolia, A. deliciosa, and A. rufa.
- Combined Stress Model: Interactive effects of warming and drought on A. deliciosa and A. rufa.
2.4.2. Model Validation with Field Data
To validate the models’ accuracy and ecological applicability, predicted germination responses were compared with long-term field distribution data of Actinidia species across Taiwan [27]. The observed species distributions under varying climatic conditions were analyzed to assess whether model predictions aligned with real-world germination success and recruitment patterns. A strong correspondence between modeled predictions and field observations would support the models’ utility for forecasting climate-induced germination shifts.
3. Results
3.1. Seed Germination Responses Across Different Species and Elevations
The germination responses of the four Actinidia species exhibited substantial variation across elevations and environmental conditions, as illustrated in Table 1. A. latifolia seeds collected from the low-elevation site in southern Taiwan (Mudan, Pingtung County, 400 m) showed a notably low germination percentage of 43.0%, whereas those from the low-elevation site in central Taiwan (Wufeng, Taichung County, 200 m) demonstrated a significantly higher germination rate of 96.3%. This stark contrast suggests that regional climatic differences or genetic adaptations may influence germination success at similar elevations. Seeds from the higher-elevation site (Beidongyan Mountain, Nantou County, 1,600 m) exhibited a germination rate of 94.8%, aligning closely with the central low-elevation population.
For A. setosa, which primarily inhabits higher elevations, the initial germination percentage were relatively low, with 8.0% at 1,870 m and 10.7% at 2,250 m. Cold stratification at 5°C moderately improved germination to 16.0% and 14.0%, respectively, indicating that A. setosa exhibits deep dormancy, which is only partially alleviated by chilling exposure.
A. rufa, collected from an elevation of 1,800 m, displayed inherently high germination percentage (89.3%), with a marginal increase to 90.7% following cold stratification. These results suggest that A. rufa seeds do not require prolonged chilling exposure to break dormancy, indicating a high level of ecological adaptability.
In contrast, A. deliciosa exhibited a strong dependence on chilling for dormancy release. Seeds collected from the highest elevation (2,500 m) initially showed a very low germination rate of 3.3%. However, after cold stratification, germination increased drastically to 93.3%, underscoring this species’ strong requirement for chilling accumulation before germination.
3.2. Impact of Low Temperature on Germination
The effect of cold stratification on seed germination was further evaluated using classifier model analysis (Figure 1). Low-temperature exposure significantly enhanced the germination of A. deliciosa (p < 0.001), demonstrating its strict chilling requirement. In contrast, germination in A. rufa, A. setosa, and A. latifolia was not significantly influenced by chilling exposure (p > 0.05), confirming that these species do not rely on extended cold exposure for dormancy release. The simulation results revealed that eight weeks of chilling were sufficient to break dormancy in A. deliciosa, while A. setosa exhibited persistent dormancy even under prolonged chilling conditions.
3.3. Effects of Winter Warming on Germination
Under simulated winter warming conditions, significant species-specific differences in germination responses were observed (Figure 2). A. deliciosa showed a significant reduction in germination as temperatures increased (p < 0.01), emphasizing its reliance on cold exposure for germination. In contrast, A. rufa benefited from warmer conditions, with germination percentage increasing significantly (p < 0.001), suggesting that this species is adapted to milder winter conditions. While A. setosa exhibited a slight decline in germination under warming conditions, this effect was not statistically significant (p > 0.1). These results indicate that A. deliciosa is highly vulnerable to climate-induced warming, whereas A. rufa may exhibit increased germination success under future warming scenarios.
3.4. Altitudinal and Latitudinal Variability in Germination
The effects of altitude on germination responses were examined in Figure 3. A. setosa seeds from lower elevations exhibited a pronounced decline in germination, while those from higher elevations maintained relatively stable germination percentage. This suggests that populations at lower elevations may be more sensitive to climate fluctuations, possibly due to genetic differentiation or local adaptation. In contrast, A. latifolia did not show a significant correlation between altitude and germination success (p > 0.05), indicating that its germination is influenced more by other environmental factors.
The influence of latitude on germination success was assessed in Figure 4. A. latifolia populations from southern Taiwan exhibited significantly lower germination percentage under warming conditions (p < 0.001), whereas populations from northern Taiwan remained largely unaffected. This suggests that southern populations are more vulnerable to climate-induced temperature increases, likely due to differences in their thermal tolerance thresholds or genetic adaptations. These findings highlight the importance of considering latitude-dependent responses in conservation and breeding strategies for A. latifolia.
3.5. Influence of Winter Drought on Germination
Winter drought conditions had a notable impact on seed germination across species (Figure 5). A. deliciosa exhibited an unexpected increase in germination under prolonged drought exposure, suggesting that dormancy release in this species may be triggered by desiccation stress. Conversely, A. latifolia showed a sharp decline in germination with extended drought exposure (p < 0.001), indicating its high sensitivity to moisture deficits. The germination of A. rufa remained largely unaffected by drought stress, further reinforcing its classification as a highly resilient species. These findings suggest that A. latifolia may require targeted conservation strategies to mitigate the negative impacts of prolonged drought conditions.
3.6. Combined Effects of Winter Warming and Drought Stress
The combined effects of winter warming and drought stress were assessed in Figure 6, revealing the most severe impacts on A. deliciosa. Under concurrent warming and drought conditions, A. deliciosa germination percentage dropped to below 25% (p < 0.001), highlighting its extreme sensitivity to compounded climate stressors. In contrast, A. rufa exhibited minimal changes in germination, reinforcing its ecological adaptability and potential as a climate-resilient species. The pronounced vulnerability of A. deliciosa suggests that climate change mitigation strategies for kiwifruit cultivation must prioritize adaptive management approaches to maintain germination success.
3.7. Validation of Model Predictions
To evaluate the reliability of the classifier models, predicted germination responses were compared with long-term field distribution data for Actinidia species in Taiwan [27,28]. The observed distribution patterns aligned strongly with model projections, confirming the robustness of the approach. A. rufa, identified as the most resilient species in experimental conditions, was also found to have the broadest natural distribution range, spanning elevations from 50 to 2,200 m. Similarly, the high sensitivity of A. deliciosa to warming and drought stress was reflected in its restricted high-altitude distribution, indicating that future climate warming may significantly reduce its viable habitat. A. latifolia’s latitude-dependent germination responses were also validated by field observations, with southern populations exhibiting lower recruitment success compared to their northern counterparts. The strong correlation between model predictions and real-world distributions underscores the potential application of classifier modeling in forecasting climate change impacts on plant species. These findings provide crucial insights for conservation strategies, breeding programs, and climate adaptation measures for kiwifruit species under future climate scenarios.
4. Discussion
This study provides critical insights into the differential seed germination responses of four Actinidia species under climate-induced stressors, highlighting the intricate interplay between environmental conditions and the mechanisms underlying seed dormancy. By integrating empirical germination data with classifier modeling, we have demonstrated that species exhibit distinct resilience patterns to warming and drought, underscoring the importance of considering species-specific physiological traits in climate impact assessments.
Among the studied species, A. rufa exhibited the highest resilience to climate change stressors, maintaining stable germination percentage under both warming and drought conditions. This suggests that A. rufa possesses an ecological dormancy mechanism [29] that enables germination once favorable conditions return, making it an ideal candidate for conservation and ecological restoration efforts. Its widespread natural distribution across various elevations further supports its adaptability, reinforcing the species’ potential to persist under future climate scenarios.
In contrast, A. deliciosa emerged as the most climate-sensitive species, with germination percentage plummeting under warming conditions. The sharp decline in germination success under reduced chilling exposure indicates a strong dependency on cold stratification for dormancy release [30,31,32,33,34]. Given the projected rise in winter temperatures, the primary breeding goal for A. deliciosa should prioritize the development of cultivars with reduced chilling requirements, rather than improving cold tolerance. Selecting and breeding genotypes that require less chilling for dormancy release would enhance the species’ adaptability to a warming climate and ensure the sustainability of kiwifruit production in regions experiencing reduced winter chill accumulation.
A. latifolia demonstrated a latitude-dependent germination response, with populations south of Tropic of Cancer (23.5°N) exhibiting a pronounced decline in germination success under warming conditions. This finding suggests that populations from higher latitudes possess greater thermal resilience, potentially due to localized adaptations to cooler environments [35]. Furthermore, A. latifolia exhibited high sensitivity to drought, with extended dry periods leading to significant reductions in germination. The correlation between population distribution and moisture availability aligns with previous ecological observations, emphasizing the necessity of maintaining adequate habitat humidity to ensure species persistence.
A. setosa presented the most complex dormancy behavior, with seed germination responses strongly influenced by altitude. Notably, A. setosa germination was significantly constrained by seed collection altitude, with higher-elevation populations exhibiting greater germination success. However, despite the variation in germination percentage, cold stratification did not effectively break dormancy in this species, suggesting that its dormancy mechanisms may be more physiologically complex than previously assumed. This altitudinal pattern [36] highlights the potential role of genetic and environmental interactions in dormancy regulation, and the implications for population survival under future climate scenarios should be carefully considered. As climate change accelerate warming trends, populations at lower elevations may experience increased germination failure, necessitating conservation interventions to safeguard genetic diversity.
The predictive models developed in this study were validated against long-term field observations of Actinidia species distributions in Taiwan [27,28], demonstrating strong alignment between empirical data and model projections. Traditional species distribution models (SDMs) often fail to incorporate physiological processes [20] such as dormancy-breaking requirements, potentially leading to overgeneralized predictions [17]. Our study highlights the necessity of integrating physiological and ecological data into predictive frameworks to enhance the accuracy of climate impact assessments. The robust model validation underscores its applicability for forecasting species distribution shifts and informing conservation strategies under future climate scenarios.
Beyond its implications for Actinidia conservation, this study provides broader insights into the effects of climate change on seed germination dynamics. The findings underscore the need for species-specific conservation planning, as responses to warming and drought are highly variable among species. A. rufa emerges as a strong candidate for habitat restoration and assisted migration initiatives, while A. deliciosa requires targeted breeding programs to develop low-chilling-requirement cultivars. Additionally, the identification of latitude- and altitude-specific vulnerabilities in A. latifolia and A. setosa offers valuable guidance for refining conservation priorities and developing adaptive management strategies.
Future research should explore the long-term impacts of climate change on Actinidia seedling survival and establishment. While germination success is a critical factor, seedling survival and growth under climate-induced stressors may determine overall population persistence. In particular, the phenomenon of ‘living dead trees’ [2], where mature individuals persist but seedling recruitment fails due to climate stress, warrants further investigation. Such studies should integrate research on bud dormancy, flowering phenology, and other life-stage transitions to formulate comprehensive conservation and agricultural strategies. Collaborating with plant phenology gardens and phenology networks would provide a robust framework for long-term monitoring and enhance the efficiency of climate adaptation research [1], ensuring that mitigation strategies are based on the most comprehensive data available.
In conclusion, our study provides a detailed assessment of Actinidia seed germination responses to climate change, offering novel empirical data and predictive modeling insights. The findings contribute to the broader field of plant ecophysiology and climate adaptation, emphasizing the necessity of integrating physiological traits into climate impact assessments. As global temperatures continue to rise, the insights gained from this study will inform conservation efforts, agricultural sustainability, and future research on plant-climate interactions.
5. Conclusions
This study provides a comprehensive assessment of seed germination responses in four Actinidia species under climate change-induced stressors, revealing distinct interspecific differences in resilience to warming and drought. By integrating empirical germination data with classifier modeling, we demonstrate that these species exhibit unique adaptive strategies that shape their survival and potential distribution under future climate scenarios.
Our findings highlight that A. rufa is the most resilient among the studied species, maintaining stable germination percentage across diverse environmental conditions. This adaptability suggests that A. rufa could serve as a key species for ecological restoration efforts in response to climate change. Conversely, A. deliciosa, the only cultivated species in this study, is highly vulnerable to warming and drought stress due to its strong dependency on chilling for dormancy release. Given the projected reduction in winter chill accumulation, the primary breeding goal for A. deliciosa should focus on developing cultivars with low chilling requirements rather than increasing cold tolerance. Targeted breeding programs incorporating genotypes with lower chilling demands will be critical to sustaining kiwifruit production under future climatic conditions.
A. latifolia exhibits latitude-dependent germination success, with populations from southern Taiwan displaying reduced germination under warming scenarios. This highlights the importance of maintaining genetic diversity across latitudinal gradients to enhance the species’ resilience. Meanwhile, A. setosa demonstrate the most complex dormancy behavior, with germination percentage highly dependent on elevation. While seeds from higher elevations exhibit greater germination success, low-temperature stratification fails to effectively break dormancy, indicating that A. setosa possesses physiological dormancy mechanisms that require further investigation. As lower-elevation populations face increasing germination failure under climate warming, conservation efforts must prioritize habitat protection and genetic resource preservation.
The validation of our predictive models against long-term field observations confirms their robustness in forecasting species distribution shifts under climate change. Unlike traditional species distribution models, which often overlook physiological processes, our approach integrate dormancy-breaking requirements and germination dynamics, thereby improving the accuracy of climate impact assessments. These findings underscore the necessity of incorporating physiological traits into predictive frameworks to enhance conservation planning and climate adaptation strategies.
Future research should extend beyond studying germination alone, and investigate the long-term impacts of climate change on Actinidia seedling survival and establishment. While high germination percentage are essential for population persistence, successful seedling recruitment and growth ultimately determine species resilience. The phenomenon of ‘living dead trees,’ where mature individuals persist but fail to recruit new seedlings, warrants further investigation, particularly in the context of shifting temperature and precipitation regimes. Additionally, integrating seed dormancy research with phenological observations through collaborations with plant phenology gardens and phenology networks could significantly enhance our understanding of climate-induced reproductive shifts in Actinidia species. Such efforts will be instrumental in refining conservation priorities, guiding adaptive management practices, and informing agricultural strategies for kiwifruit cultivation under changing climatic conditions.
In conclusion, this study provides critical empirical data and predictive modeling insights that advance our understanding of seed germination responses to climate change. Our findings emphasize the importance of species-specific conservation strategies, targeted breeding initiatives, and ecosystem-based management approaches to safeguard kiwifruit biodiversity and agricultural sustainability. As global temperatures continue to rise, these insights will play a pivotal role in shaping future research and conservation efforts aimed at mitigating climate change impacts on plant reproductive success.
Author Contributions
T.-Y. Hsieh and C.-T. Chien conceptualized the study and designed the experiments. F. Li contributed to statistical analyses and modeling. All authors participated in data collection, interpretation, and manuscript preparation. Shih-Li Huang supervised the study and finalized the manuscript. All authors have read and approved the final version of the manuscript.
Acknowledgments
We thank Fujian Polytechnic Normal University and the Taiwan Forestry Research Institute for providing essential support and laboratory facilities. Special thanks go to Yen-Wei Chang, Chang-Yen Chen, Wen-Yu Hsu, Yu-Han Tsai, and Shun-Ying Chen (Taiwan Forestry Research Institute) for their excellent technical assistance with seed collection, data recording, and germination experiments. We also acknowledge valuable insights from the anonymous reviewers for constructive feedback that improved the manuscript.
References
- Hsieh, T.-Y.; Chiou, C.-R. Phytophenology and its applications in climate change research: review and future perspectives. Quarterly Journal of Chinese Forestry 2013, 46, 391–410. [Google Scholar]
- Hsieh, T.-Y. Impacts of Climate Change on Species, Ecosystems, and their Management Strategies. Taiwan Journal of Forest Science 2016, 31, 227–255. [Google Scholar]
- Dang, C.; Shao, Z.; Huang, X.; Zhuang, Q.; Cheng, G.; Qian, J. Climate warming-induced phenology changes dominate vegetation productivity in Northern Hemisphere ecosystems. Ecological Indicators 2023, 151, 110326. [Google Scholar] [CrossRef]
- Gao, X.; Zhao, D. Impacts of climate change on vegetation phenology over the Great Lakes Region of Central Asia from 1982 to 2014. Science of the Total Environment 2022, 845, 157227. [Google Scholar] [CrossRef]
- Hsieh, T.-Y.; Yang, C.-J.; Li, F.; Chiou, C.-R. Numerical Ecology and Social Network Analysis of the Forest Community in the Lienhuachih Area of Taiwan. Diversity 2023, 15, 60. [Google Scholar] [CrossRef]
- Rathore, M.K.; Sharma, L.K. Efficacy of species distribution models (SDMs) for ecological realms to ascertain biological conservation and practices. Biodiversity and Conservation 2023, 32, 3053–3087. [Google Scholar] [CrossRef]
- Chien, C.-T.; Kuo-Huang, L.-L.; Lin, T.-P. Changes in ultrastructure and abscisic acid level, and response to applied gibberellins in Taxus mairei seeds treated with warm and cold stratification. Annals of Botany 1998, 81, 41–47. [Google Scholar] [CrossRef]
- Thuiller, W.; Münkemüller, T.; Lavergne, S.; Mouillot, D.; Mouquet, N.; Schiffers, K.; Gravel, D. A road map for integrating eco-evolutionary processes into biodiversity models. Ecology letters 2013, 16, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Mou, S.S.; Haus, M.J.; Hayden, Z.D.; Patterson, E.L.; Saha, D. Climate-driven challenges in weed management for ornamental crop production in the United States: a review. Frontiers in Agronomy 2025, 7, 1556418. [Google Scholar] [CrossRef]
- Hudson, A.R.; Ayre, D.J.; Ooi, M.K. Physical dormancy in a changing climate. Seed Science Research 2015, 25, 66–81. [Google Scholar] [CrossRef]
- Solarik, K.A.; Gravel, D.; Ameztegui, A.; Bergeron, Y.; Messier, C. Assessing tree germination resilience to global warming: a manipulative experiment using sugar maple (Acer saccharum). Seed Science Research 2016, 26, 153–164. [Google Scholar] [CrossRef]
- Whitmore, T. Potential impact of climatic change on tropical rain forest seedlings and forest regeneration. Climatic Change 1998, 39, 429–438. [Google Scholar] [CrossRef]
- Klupczyńska, E.A.; Pawłowski, T.A. Regulation of seed dormancy and germination mechanisms in a changing environment. International journal of molecular sciences 2021, 22, 1357. [Google Scholar] [CrossRef]
- Zu, K.; Chen, F.; Li, Y.; Shrestha, N.; Fang, X.; Ahmad, S.; Nabi, G.; Wang, Z. Climate change impacts flowering phenology in Gongga Mountains, Southwest China. Plant Diversity 2023. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Bu, Z.J.; Poschlod, P.; Yusup, S.; Zhang, J.Q.; Zhang, Z.X. Seed dormancy types and germination response of 15 plant species in temperate montane peatlands. Ecology and Evolution 2024, 14, e11671. [Google Scholar] [CrossRef]
- Lamont, B.B.; Pausas, J.G. Seed dormancy revisited: Dormancy-release pathways and environmental interactions. Functional Ecology 2023, 37, 1106–1125. [Google Scholar] [CrossRef]
- Giménez-Benavides, L.; Escudero, A.; García-Camacho, R.; García-Fernández, A.; Iriondo, J.; Lara-Romero, C.; Morente-López, J. How does climate change affect regeneration of Mediterranean high-mountain plants? An integration and synthesis of current knowledge. Plant Biology 2018, 20, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Chiou, C.-R.; Hsieh, T.-Y.; Chien, C.-C. Plant bioclimatic models in climate change research. Botanical studies 2015, 56, e26. [Google Scholar] [CrossRef]
- A. Lee-Yaw, J.; L. McCune, J.; Pironon, S.; N. Sheth, S. Species distribution models rarely predict the biology of real populations. Ecography 2022, 2022, e05877. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, J.; García Molinos, J.; Mammola, S.; Bede-Fazekas, Á.; Feng, X.; Kitazawa, D.; Assis, J.; Qiu, T.; Lin, Q. Incorporating physiological knowledge into correlative species distribution models minimizes bias introduced by the choice of calibration area. Marine Life Science & Technology 2024, 6, 349–362. [Google Scholar]
- Ferguson, A.R. Kiwifruit: the wild and the cultivated plants. In Advances in food and nutrition research; Elsevier, 2013; Volume 68, pp. 15–32. [Google Scholar]
- Li, H.-L. A taxonomic review of the genus Actinidia. Journal of the Arnold Arboretum 1952, 33, 1–61. [Google Scholar] [CrossRef]
- Hsieh, T.-Y.; Ku, S.-M.; Chien, C.-T.; Liou, Y.-T. Classifier modeling and numerical taxonomy of Actinidia (Actinidiaceae) in Taiwan. Botanical Studies 2011, 52, 337–357. [Google Scholar]
- Nee, C.C.; Tsay, T.T. Kiwifruit in Taiwan. In Proceedings of the Acta Horticulturae; 1991; pp. 175–182. [Google Scholar]
- Chien, C.T.; Chen, S.Y.; Yang, J.C. Effect of stratification and drying on the germination and storage of Purnus campanulata seeds. Taiwan Journal of Forest Science 2002, 17, 413–420. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing; Vienna, Austria, 2024. [Google Scholar]
- Hsieh, T.-Y. Distribution of Indigenous Actinidia in Taiwan. Horticulture NCHU 2011, 36, 1–8. [Google Scholar]
- Hsieh, T.-Y. Taxonomy and distribution of indigenous Actinidia in Taiwan; National Chung-Hsing University: Taichung, Taiwan, 2011. [Google Scholar]
- Lang, G.A. Dormancy: a new universal terminology. 1987.
- Esfandiari, A.; Norling, C.; Kaji, R.; McLachlan, A.; Mathew, L.; Fleming, M.; Morgan, E.; Nadarajan, J. Variations in Seed Dormancy Occurrence and Their Classifications in Thirteen Actinidia Species. Seeds 2024, 3, 179–195. [Google Scholar] [CrossRef]
- Smith, R.L.; Toy, S.J. Effects of stratification and alternating temperatures on seed germination of chinese gooseberry Actinidia chinensis Planch. In Proceedings of the Proceedings of the American Society for Horticultural Science, 1967; pp. 409–412.
- Lawes, G.; Anderson, D. Influence of temperature and gibberellic acid on kiwifruit (Actinidia chinensis) seed germination. New Zealand Journal of Experimental Agriculture 1980, 8, 277–280. [Google Scholar] [CrossRef]
- Geneve, R.L. Impact of temperature on seed dormancy. HortScience 2003, 38, 336–340. [Google Scholar] [CrossRef]
- Celik, H.; Zenginbal, H.; Özcan, M. Enhancing germination of kiwifruit seeds with temperature, medium and gibberellic acid. Horticultural Science 2006, 33, 39–45. [Google Scholar] [CrossRef]
- Hsieh, T.Y.; Nee, C.C.; Chien, C.T. Seed germination of Taiwanese Actinidia latifolia (Gardn. and Champ.) Merr. Taiwan J For Sci 2004, 19, 173–176. [Google Scholar]
- Marris, E. The escalator effect. Nature Reports Climate Change 2007, 1, 94–96. [Google Scholar] [CrossRef]
Figure 1.
Simulations of the chilling duration in winter impacts on the seed germination of A. deliciosa, A. rufa, A. setosa, and A. latifolia under different chilling accumulated weeks.
Figure 1.
Simulations of the chilling duration in winter impacts on the seed germination of A. deliciosa, A. rufa, A. setosa, and A. latifolia under different chilling accumulated weeks.
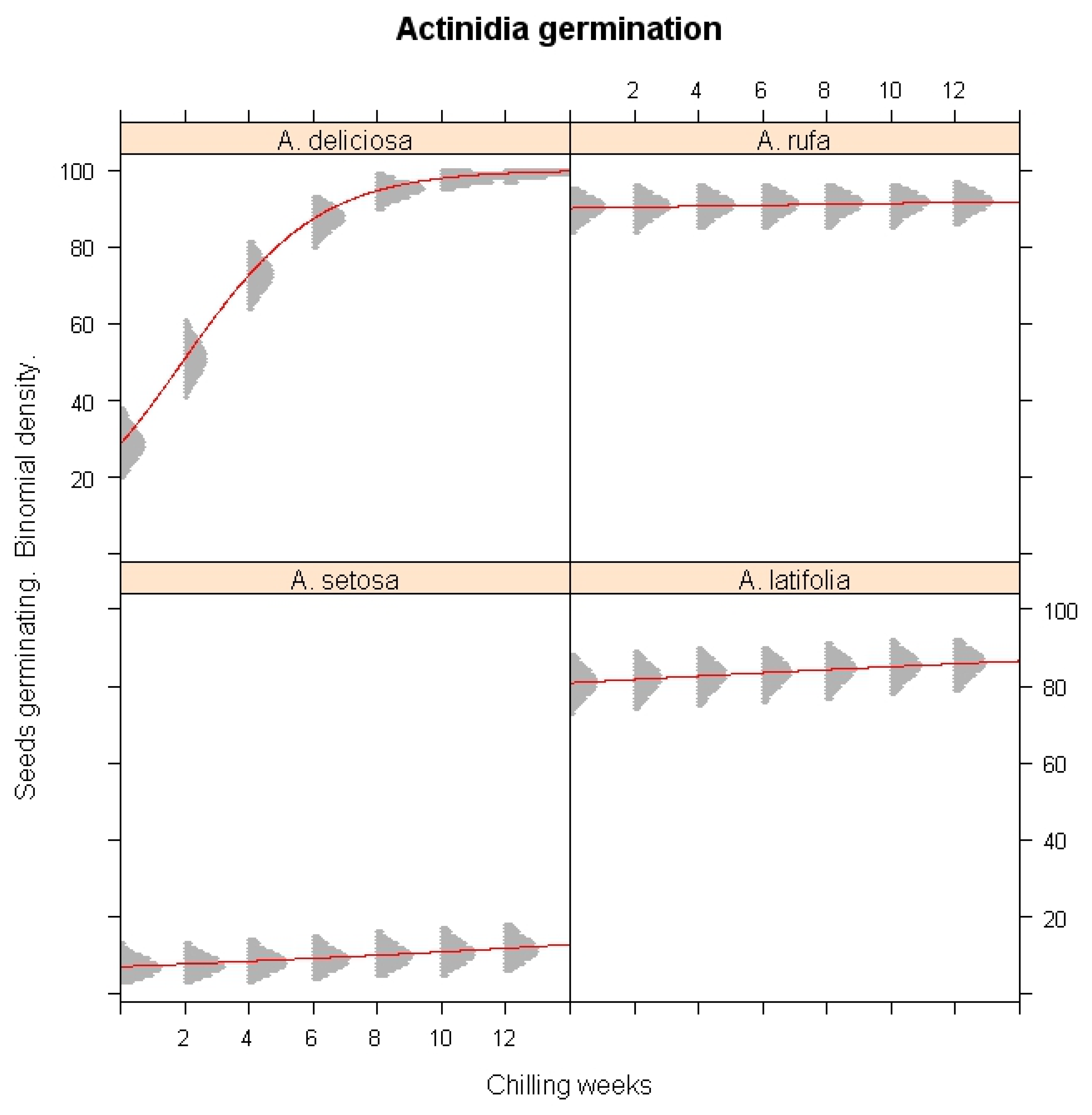
Figure 2.
Simulations of warming winter impacts on the seed germination of A. deliciosa, A. rufa and A. setosa under different temperature regimes.
Figure 2.
Simulations of warming winter impacts on the seed germination of A. deliciosa, A. rufa and A. setosa under different temperature regimes.
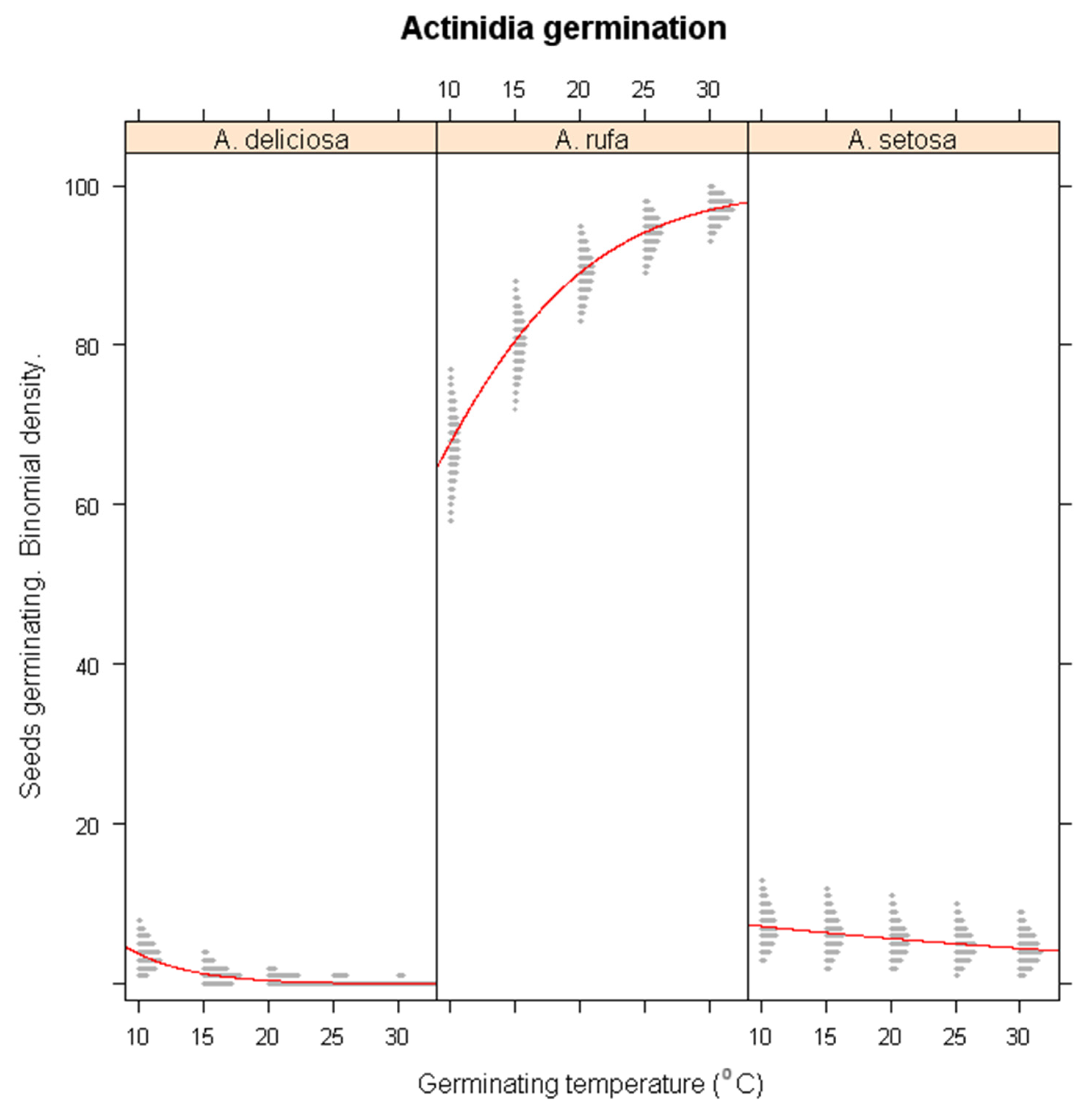
Figure 3.
Simulations of warming winter impacts on the seed germination of A. setosa and A. latifolia along altitude gradients of its habiats.
Figure 3.
Simulations of warming winter impacts on the seed germination of A. setosa and A. latifolia along altitude gradients of its habiats.
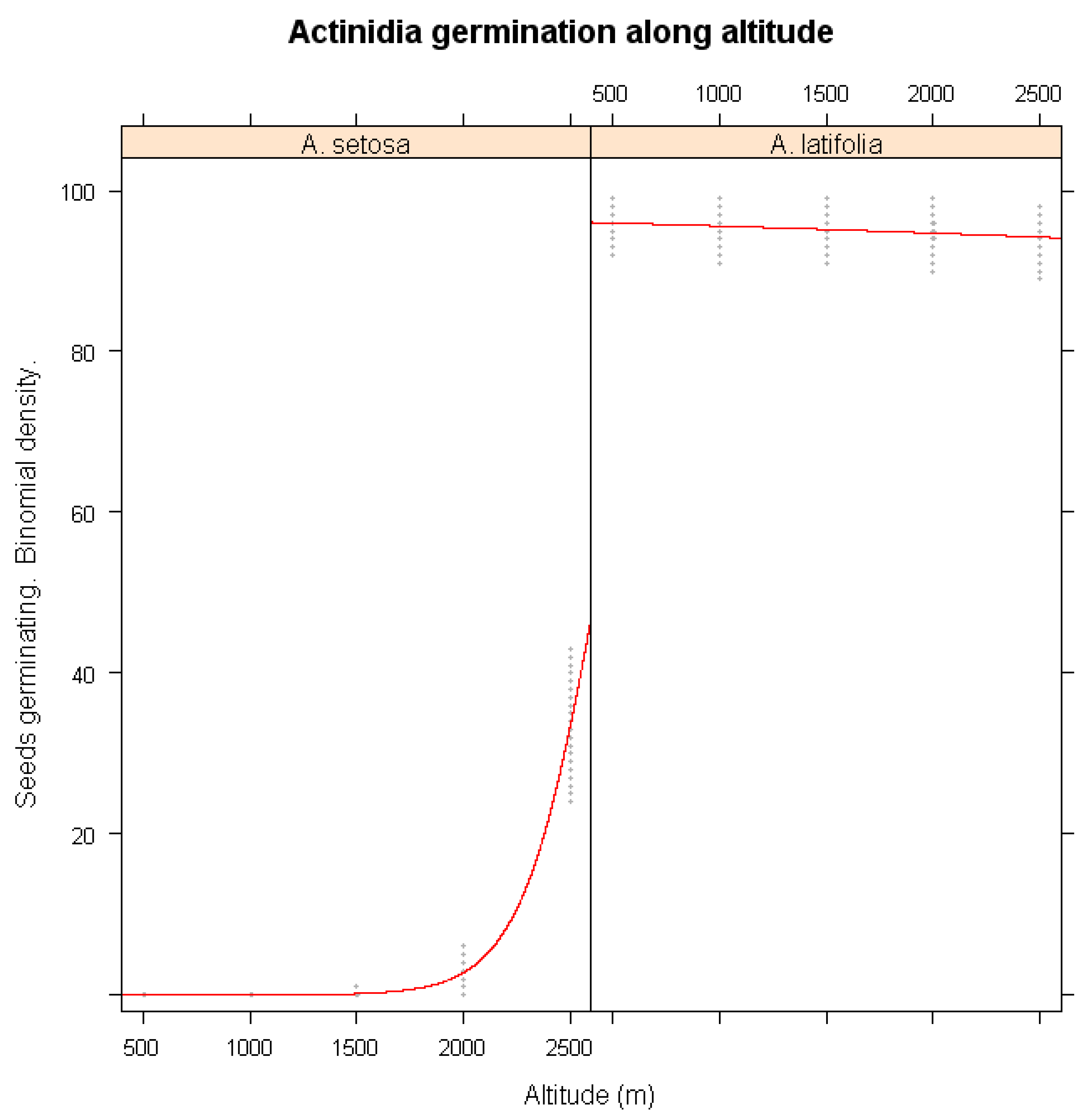
Figure 4.
Simulations of warming winter impacts on the seed germination of A. latifolia along latitude gradients of its habiats.
Figure 4.
Simulations of warming winter impacts on the seed germination of A. latifolia along latitude gradients of its habiats.
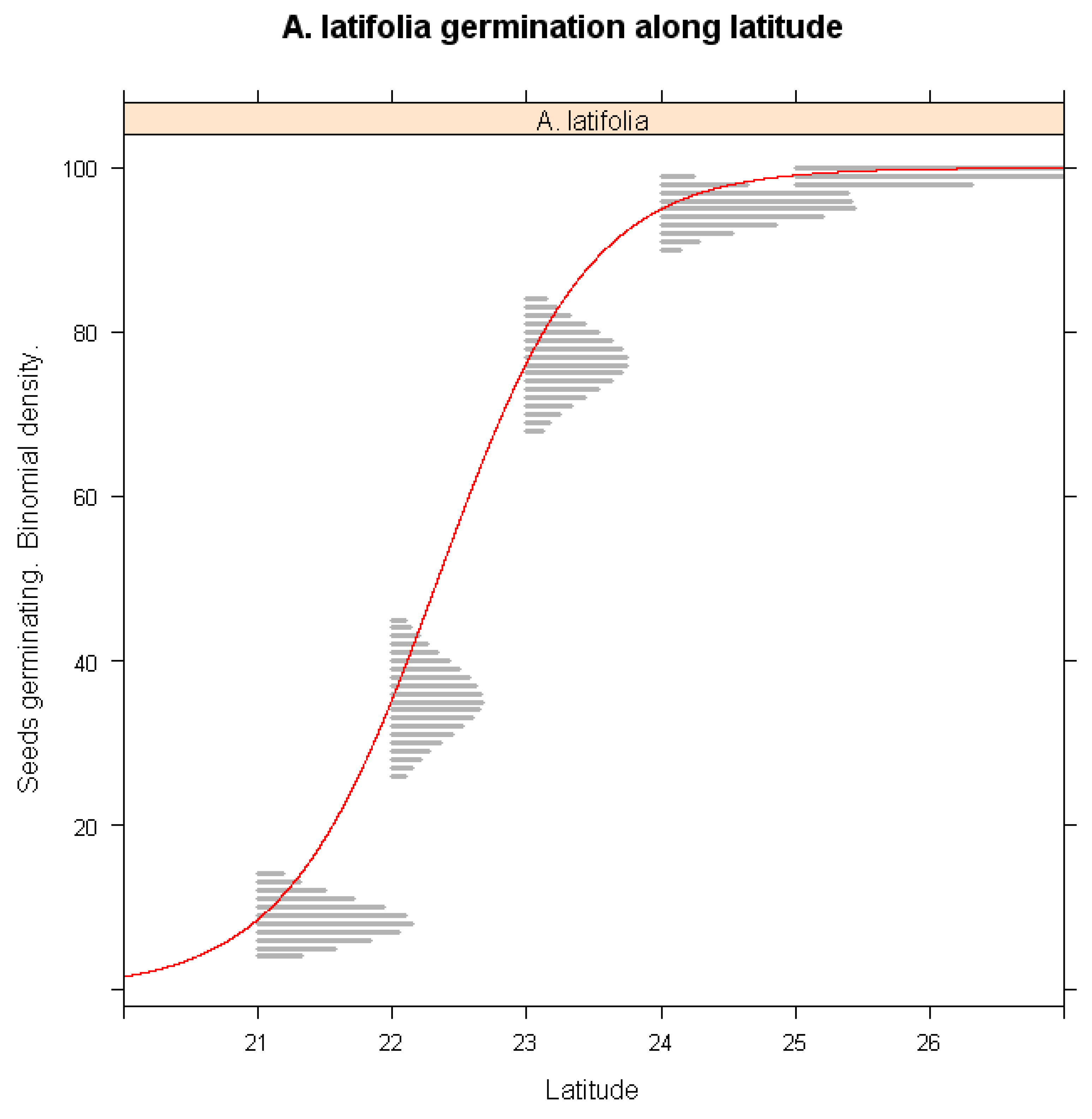
Figure 5.
Predicted seed germination percentages of three Actinidia species under different durations of winter drought.
Figure 5.
Predicted seed germination percentages of three Actinidia species under different durations of winter drought.
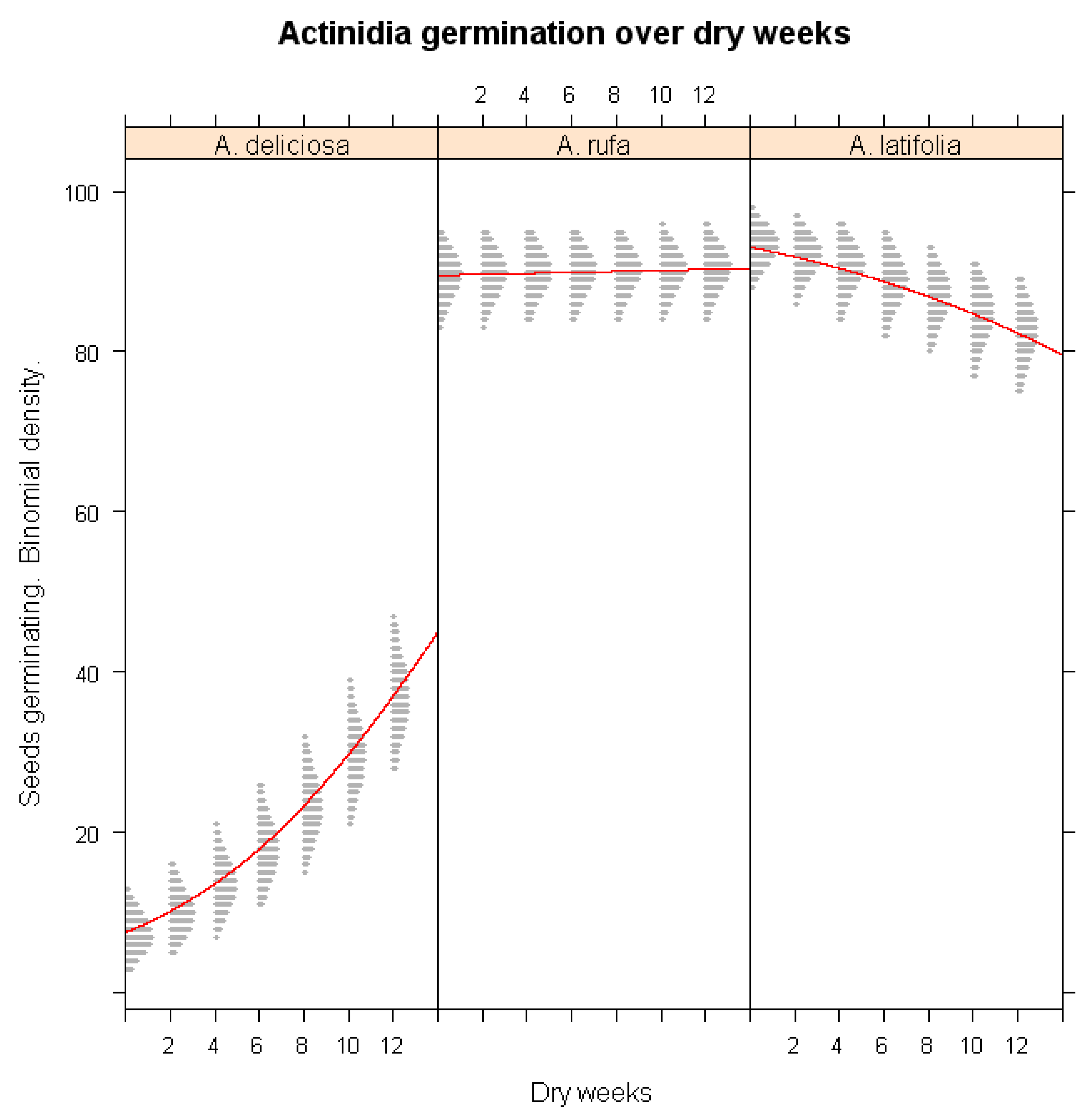
Figure 6.
Predicted seed germination percentages of Actinidia deliciosa and A. rufa under severe warm winter conditions following different drought durations.
Figure 6.
Predicted seed germination percentages of Actinidia deliciosa and A. rufa under severe warm winter conditions following different drought durations.
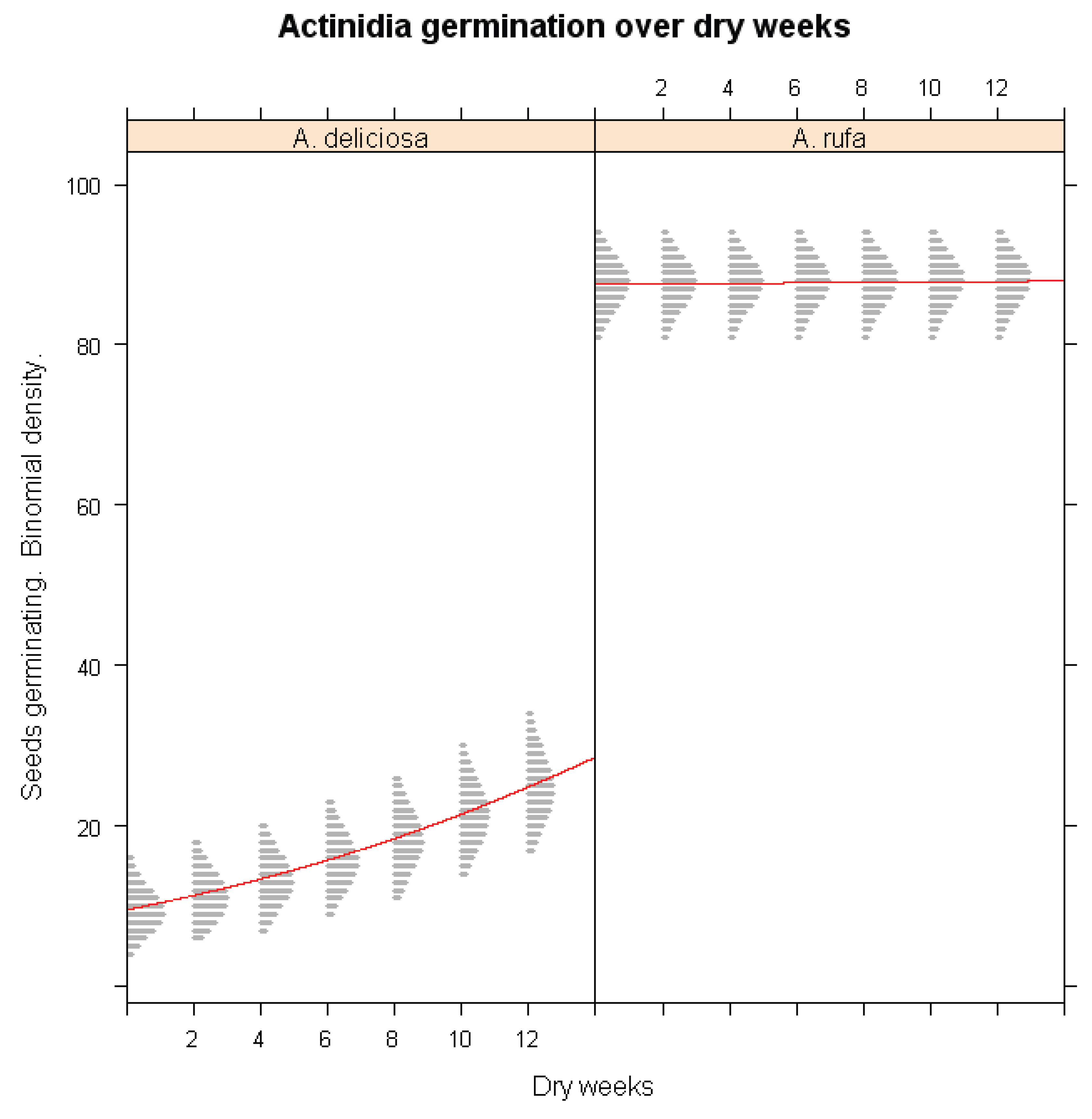

Table 1.
Information of the 4 species and seed germination percentages.
| Species | Collection sites and elevations |
Maximum seed germination (%) | Maximum seed germination after cold stratification (%) |
| Actinidia latifolia | Mutan, Pingtung County; elevation 400m Wufeng, Taichung County elevation 200m Peitungyenshan, Nantou County elevation 1600m |
43.0 96.3 94.8 |
51.5 94.3 94.6 |
| Actinidia setosa | Tsuifeng, Nantou County elevation 2250m |
10.7 |
14.0 |
| Siyuan-Yakou, Yilan County elevation 1870m |
8 | 16 | |
| Actinidia rufa | Qingjing, Nantou County elevation 1800m |
89.3 |
90.7 |
| Actinidia deliciosa | Shengguang, Heping Dist., Taichung City elevation 2500m |
3.3 | 93.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



