Submitted:
21 May 2025
Posted:
23 May 2025
You are already at the latest version
Abstract
This study investigated the essential oils (EOs) from leaves, bark, and fruits of Mespilodaphne cymbarum (Kunth) Trofimov (Lauraceae), characterizing their chemical composition and evaluating their antimicrobial and antibiofilm activities. EOs were extracted from plants collected in the Amazon during dry and flood seasons and analyzed by gas chromatography coupled to mass spectrometry (GC-MS). Although chemical differences were evident among plant organs and chemotypes, the influence of seasonality was not pronounced. Major metabolites of fruits EO were α- and β-santalene and limonene. Bark EO was rich in phenylpropanoids, including methyl eugenol, myristicin and elemicin. Leaf EOs showed the greatest metabolic diversity, with chemotype-specific variations. Essential oils from leaves and bark demonstrated superior antibacterial and antibiofilm activities compared to fruit oils, especially against Gram-positive bacteria. Differences among plant organs and chemotypes were observed, with leaf oils showing strong inhibition of biofilm formation and bark oils exhibiting broader antibacterial effects. These findings highlight the potential of M. cymbarum EOs as a natural source of bioactive compounds and emphasizing the importance of chemotype and plant organ selection for optimized applications.
Keywords:
bark
; leaves
; fruit
; flooding
; Ocotea cymbarum
; gram‐positive
1. Introduction
The botanical family Lauraceae Juss. is recognized as a source of essential oils (EOs) with numerous biological properties and industrial applications, such as cinnamon (Cinnamomum spp.), laurel (Laurus nobilis), and rosewood (Aniba rosaeodora) [1,2]. Predominantly distributed in tropical regions, Lauraceae encompasses approximately 55 genera and 2,500–3,500 species [3]. Among these, the Ocotea Aubl. complex stands out, comprising around 16 genera and 700 species, many of which hold economic and biological importance, including Nectandra, Aniba, Licaria, and Ocotea [3]. Genetic analyses based on nuclear internal transcribed spacer (ITS) sequences led to the reinstatement of the genus Mespilodaphne Nees & Mart. and the reclassification of Ocotea cymbarum Kunth as Mespilodaphne cymbarum (Kunth) Trofimov [4]. The genus Mespilodaphne includes eight species, distributed across South and Central America, as well as in tropical forests of the Antilles at elevations of up to 2,000 meters [4].
Mespilodaphne cymbarum is typically found throughout the Amazon Region with occurrences in Brazil, Colombia, Venezuela, and Guyana [5]. This species is commonly registered in várzea forests, such as the banks of Solimões River, where it is subjected to periodic flooding [5,6].
Due to its valuable wood, M. cymbarum has been exploited for timber [6,7]. The wood, bark, and leaves of M. cymbarum are known to contain neolignans and have also been reported as sources of essential oils (EOs) [8,9,10]. The neolignan burchelin, isolated from the bark of M. cymbarum, demonstrated potential in vitro activity against both the epimastigote and trypomastigote forms of Trypanosoma cruzi [9]. Another neolignan, biseugenol, was isolated from leaves and showed anti-inflammatory activity, as well as inhibition of angiogenesis and fibrogenesis [10].
Research on the EO of M. cymbarum has focused on the bark, and trunk wood [7,8,11,12,13]. Shukis and Wachs [14] reported the presence of safrole in this species, but studies employing modern analytical techniques failed to confirm this occurrence [7,11,12]. In addition, the metabolites α-phellandrene, p-cymene, and α-pinene were reported from EO of trunk wood M. cymbarum [8]. In contrast, EOs of sassafras from wood/bark cited as major components the monoterpenes α-terpineol (34.9%), α-pinene (18.5%), fenchol (6.3%), and borneol (6.2%) [11]. Recently, the EO from sapwood and heartwood of M. cymbarum was analyzed by SPME and the volatiles with higher relative area percentage were α-copaene (17%), 1,8-cineole (11%), trans-calamene (7.4%), α-calacorene (6.4%), and δ-cadinene (5.5%) [7]. While from EO of M. cymbarum bark, α-selinene (26%), δ-cadinene (19%), terpinen-4-ol (9%), and α-cadinol (6.2%) were described as major metabolites [12].
Residents of Amazonian communities have reported that M. cymbarum produces aromatic oils in multiple plant organs, highlighting its potential for sustainable use by local populations. Given the lack of data on the chemical and biological properties of essential oils from M. cymbarum leaves and fruits, this study provides a detailed characterization of oils from different plant organs and their antimicrobial and antibiofilm activities against six pathogenic bacterial strains. Samples were collected during both dry and flooding seasons in a seasonally flooded forest (várzea) of the central Amazon to explore potential seasonal and chemotypic variations.
2. Results and Discussion
2.1. Yields of the Essential Oils (EOs) from Bark, Leaves and Fruits of M. cymbarum
Our study reported for the first time the yield and chemical composition of the essential oils (EOs) from leaves (Le) and fruits (Fr) of Mespilodaphne cymbarum (Table 1). The highest yield was recorded for fruit EO (3.6%). Two distinct chemotypes were identified based on pilot studies using solid phase microextraction (SPME) and GC-MS on leaves from 14 individuals (Figure S1, Supporting Information). Consequently, the individuals were grouped accordingly before EO extraction, resulting in four experimental groups that reflect both seasonal and chemotypic variation: Le-F-1 (leaves from the flooding season of chemotype-1), Le-D-1 (leaves from the dry season of chemotype-1), Le-F-2 (leaves from the flooding season of chemotype-2), and Le-D-2 (leaves from the dry season of chemotype-2).
The EOs from the leaves showed similar yield among the two chemotypes (1 and 2) and between the dry and flooding seasons. The essential oils from leaves collected in Le-D-1 and Le-F-1 yielded 0.91%, while Le-D-2 and Le-F-2 returned 1.2 and 0.93%, respectively. The bark EOs resulted in lower yield than the other parts, showing yields of 0.6% for Ba-D-1 and 0.5% for Ba-F-1. These results for bark surpassed the bark EO yield (0.1%) reported for the same species [12].
Compared to other species within the Lauraceae family, particularly in the genus Ocotea and closely related taxa, M. cymbarum exhibits remarkably high essential oil yields, especially from its fruits (3.6%) and leaves (~1%). These values are substantially higher than those reported for several Ocotea species, where leaf yields often remain below 1% and bark or stem oils are even more limited [13,15,16,17,18,19]. Moreover, the consistency in leaf EO yields across both dry and flooding seasons in M. cymbarum is noteworthy, suggesting a level of phenotypic or physiological stability that could be advantageous for sustainable harvesting and commercial exploitation.
2.2. Chemical Composition and Statistical Analysis of the Essential Oils (EOs) from Bark, Leaves and Fruits of M. cymbarum
From the EOs, we detected 122 metabolites across different plant organs (Table 1). Principal components analysis (PCA) was performed using 16 samples, distributed as follows: eight replicates from leaves (Le)—comprising two analytical replicates for each leaf EOs groups (Le-D-1, Le-F-1, Le-D-2, Le-F-2), four from bark—two analytical replicates per season (Ba-D-1; Ba-F-1), and four analytical replicates from fruit EO, obtained from two hydrodistillation extractions (Figure 1a). The first two principal components, PC1 (45.4%) and PC2 (34%) explained 79.4% of total variance, revealing three distinct groups (Figure 1a; PERMANOVA, R2 = 0.9078, p < 0.001). PC1 clearly separated leaf EOs from bark and fruit EOs (Figure 1). The metabolites contributing to this separation were V9, V10, V14, V15, V17, V24, V26, V27, V30, V48, V59, V89, V93, V106, V112, V117, V119, V120—detected exclusively in leaves—and V13, V21, V23, V49, V53, V54, and V55 that were higher in this group. In contrast, V3, V22, V75, and V95 were the key volatile compounds contributing to the separation of fruit and bark EOs from leaf EOs along PC1 (Figure 1b).
Leaf EOs exhibited the highest metabolite diversity, containing 105 volatiles, with 34 metabolites detected exclusively in these samples. Sesquiterpene hydrocarbons were the major constituents of all leaf EOs, accounting for approximately 39% in samples from the flooding season, and around 30% in those from the dry season (Table 1). Chemotype-1 contained higher amounts of oxygenated monoterpenes, with 12.6% (Le-F-1) and 15.3% (Le-D-1), as well as monoterpenes hydrocarbons at about 7.6% (Le-F-1) and 8.3% (Le-D-1). On the other hand, chemotype-2 exhibited a higher proportion of oxygenated sesquiterpenes, reaching 14.9 (Le-F-2) and 15.8% (Le-D-2). The complex volatile profile of the leaf EOs resulted in the coelution of several metabolites, which made their identification particularly challenging.
The two chemotypes analyzed in our study showed qualitative differences in their leaf EOs (Table 1; Figure 2). Leaf EOs from chemotype-1 contained 10 metabolites (V26, V31, V35, V38, V66, V78, V79, V85, V108, V117) that were not detected in chemotype-2. Conversely, chemotype-2 presented 17 metabolites (V34, V74, V84, V86, V90, V91, V92, V96, V97, V99 V101, V102, V103, V107, V115, V116, and V118) that were not found in chemotype-1 (Figure S2, Supporting Information). However, some of these compounds from both chemotypes also occurred in fruit or bark EOs (Table 1).
The EOs from leaves of chemotype-1 presented as major metabolites, showing relative area higher than 5%, the volatiles β-caryophyllene (V42) (Le-F-1 = 7.9%; Le-D-1 = 4.7%), α-santalene (V43) (Le-F-1 = 6.2%; Le-D-1 = 6.3%), unknown m/z 234 (V82) (Le-F-1 = 5.4%; Le-D-1 = 5.9%), and cryptone (V21) (approximately 5% in both season). Chemotype-2 presented the metabolites unknown m/z 234 (V82) (Le-F-2 = 11%; Le-D-2 = 9.8%), β-caryophyllene (V42) (Le-F-2 = 9%; Le-D-2 = 7.6%), and δ-cadinene (V67) (Le-F-2 = 5.1%; Le-D-2 = 3.6%), as major volatiles (>5%). Although the compound unknown m/z 234 (V82) is present at high intensity, its fragmentation pattern does not match any metabolites available in the databases or compounds described in the literature. Therefore, its identification would require isolation and further chemical analyses to characterize its molecular structure. However, such characterization is beyond the scope of this study.
Studies examining the chemical composition of EOs from Ocotea and Mespilodaphne have identified monoterpenoids, sesquiterpenoids, and phenylpropanoids as the major classes of volatiles [13,16,20]. Among these, sesquiterpenes have been reported as the predominant class in several Ocotea species [21,22]. The leaf EO of M. quixos from Ecuador was mainly composed of sesquiterpenes (35.6%), oxygenated monoterpenes (24.8%), and monoterpene hydrocarbons (21.7%), with β-caryophyllene (15.1%), cinnamyl acetate (11.4%), and sabinene (7.6%), as the main volatiles [23]. From M. quixos leaf EO, collected in the Amazonian region of Pastaza (Ecuador), it was reported oxygenated monoterpenes as the most abundant compounds, showing 1,8-cineole (39.1%) and α-terpineol (7.6%) as the primary constituents, followed by hydrocarbon monoterpenes such as sabinene (6.46%), α-pinene (6.3%), and p-cymene (6.1%) [24]. Other commonly detected metabolites in M. quixos leaf EOs include the aromatics trans-cinnamaldehyde, trans-methyl cinnamate, and trans-cinnamyl acetate [19,25,26]. In Mespilodaphne veraguensis, oxygenated sesquiterpenes were the predominant class in leaf EO (58.8%), with bulnesol (29.5%), and spathulenol (8.5%) as the main constituents, followed by monoterpene hydrocarbons (27.5%), primarily p-cymene (19.8%) [20]. Chaverri et al. [16] observed in the leaf EO of M. morae, high percentage of monoterpenes, including β-pinene (17.5%), α-pinene (10.4%), and 1,8-cineole (7.3%), and sesquiterpenes, such as bicyclogermacrene (8.8%), germacrene D (7.5%), β-caryophyllene (7.1%).
The pronounced chemical variability observed among Mespilodaphne and Ocotea species, ranging from sesquiterpene to monoterpene dominant profiles, underscores the influence of taxonomic and geographic factors. Notably, β-caryophyllene frequently emerges as a predominant constituent across several species [13,20,23]. This metabolite was detected in both chemotypes. Conversely, the presence of α-santalene (V43) in higher amounts in chemotype-1 was relevant to distinguish this chemotype, since it was detected only in trace in chemotype-2. The presence of α- and β-santalene is not commonly reported in this genus [13]. However, these compounds were detected at low concentrations—approximately 0.36 and 0.11%, respectively—in the flower calyces of M. quixos [27].
Although cryptone (V21) and δ-cadinene (V67) were present in leaf EOs from both chemotypes, cryptone (V21) was greater in chemotype-1, while δ-cadinene (V67) was more abundant in chemotype-2. The occurrence of distinct chemotypes may be influenced by genotypic factors as well as abiotic and/or biotic pressures [28,29]. In M. quixos the presence of two distinct chemotypes, a trans-methyl cinnamate chemotype and trans-caryophyllene and trans-cinnamyl acetate chemotype, were determined, which were influenced by environmental conditions, such as soil composition, heights and shade percentage [26]. Similarly, diverse chemotypes in Nectandra megapotamica were recorded regarding the volatile profile from specimens collected in different geographic locations, but other compound classes, such as non-volatile phenolics, were similar within populations [30].
In the EO of fruits, we found 66 compounds, predominantly sesquiterpenes hydrocarbons (65.1%), such as α-santalene (V43) (26.4%), β-santalene (V51) (12.0%), and trans-α-bergamotene (V44) (8.18%). These compounds are the same found in the sandalwood oil extracted from Santalum album, which are extensively used by the perfumery and fragrance industries, presenting high economic value [31]. Santalenes and santalols have been detected in 32 species, including the Lauraceae Cinnamomum camphora, but in low concentration of about 2.7% [31]. Therefore, in addition to displaying a valuable sesquiterpene profile, the fruit EO of M. cymbarum also showed a high yield (3.6%), highlighting the potential of this resource. Furthermore, we used the fruit pulp, which allowed us to obtain the material while preserving the seeds, enabling a sustainable use of this species. Besides the potential applications of its essential oil, our results suggest that this species may also be a valuable source for the discovery of novel enzymes involved in the biosynthesis of santalenes and santalols.
Monoterpenes hydrocarbons were also abundant in fruit EO, accounting for approximately 28% with limonene (V6) (19.32%) as a prevalent metabolite (Figure 2). Metabolites distinguishing the fruit EOs in the PCA included V8, V11, V60, V73, V88, V110, V113, which were exclusively found in this sample, while V1, V3, V6, V43, V45, and V51 were present at higher percentages in fruit EO (Table 1; Figure 1b).
Studies analyzing the EO of fruits from Ocotea and Mespilodaphne are scarce [15,32]. The fruit EO from Ocotea duckei also recorded high amounts of limonene (30.1%), with other major volatiles being α-pinene (12.2%) and β-pinene (9.9%) [15]. Silva et al. [32] evaluated the EO of fruits at different ripening stages (unripe and ripe), and identified caryophyllene oxide ranging from 52.1% (unripe) to 27.9% (ripe), β-chenopodiol (17%) which was detected only in unripe fruits, and bicyclogermacrene that varied between 9.9% (unripe) to 6.9% (ripe).
The chemical distinction of bark EOs was explained by the metabolites V41, V68, V77, V98, V111, V114, V121, and V122, which were exclusive to these samples, along with V18, V20, V22, and V46 which were found at higher concentrations in the bark EOs (Figure 1b; Table 1). In total, 64 volatile compounds were detected in bark EOs. Phenylpropanoids were found only in bark EOs, being the dominant chemical class regardless of season, accounting for 56.7% (Ba-F-1) and 62.0% (Ba-D-1). The most abundant phenylpropanoids were methyl eugenol (V41) (37.5% in Ba-F-1, and 43.6% in Ba-D-1), followed by elemicin (V77) (11.9% in Ba-F-1, and 8.8% in Ba-D-1), and myristicin (V68) (7.4% in Ba-F-1, and 9.6% in Ba-D-1). Sesquiterpenes hydrocarbons represent 15.6% (Ba-F-1) and 12.4% (Ba-D-1), with δ-cadinene (V67) as the main compound with 7.4% in flooding-season and 6.3% in dry-season. Oxygenated sesquiterpenes correspond to 11.8 (Ba-F-1) and 9.7% (Ba-D-1), while oxygenated monoterpenes showed similar proportions (~11%) in both seasons. Overall, the chemical profiles of bark EOs are qualitatively consistent between seasons (Table 1, Figure 2).
Essential oils from bark and wood have been the most studied oil from M. cymbarum and have presented chemical variations. Zoghbi et al. [12] analyzed bark EO extracted from M. cymbarum collected in Pará state, Brazil, and reported high proportion of sesquiterpene hydrocarbons, such as α-selinene (25.8%) and δ-cadinene (18.6%). On the other hand, Avila et al. [11] identified the monoterpenes α-terpineol (34.9%), α-pinene (18.5%), fenchol (6.3%), and borneol (6.2%) as the major components of wood/bark EO obtained in plants from Arauca, Colombia. Recently, volatiles extracted by SPME from the sapwood and heartwood of M. cymbarum from revealed α-copaene (17%), 1,8-cineole (11%), trans-calamene (7.4%), α-calacorene (6.4%), and δ-cadinene (5.5%) as the main constituents, each with a relative area greater than 5% [7]. In M. morae, the major components of bark EO were the oxygenated monoterpene 1,8-cineole (12.8%) and the hydrocarbon sesquiterpene β-caryophyllene (6.1%) [16]. Bark EO of M. quixos was rich in phenylpropanoid derivatives, including trans-cinnamaldehyde (44.7%) and trans-methyl cinnamate (26.2%) as the dominant constituents [19].
The chemical data were applied to produce a molecular networking (Figure S3, Supporting Information), which assisted the visualization of the chemical similarities between the metabolites. Oxygenated monoterpenes formed a distinct group, predominantly marked by metabolites from the leaf Eos, while the phenylpropanoid cluster was clearly separated and composed exclusively of metabolites from the bark EOs. The fruit EOs were more prominently represented in the cluster of hydrocarbon monoterpenes. Oxygenated sesquiterpenes formed another cluster connected to the hydrocarbon sesquiterpenes and included compounds from all EO types.
These inter- and intraspecific chemical variations, including the biosynthesis of secondary metabolites, can be influenced by genotypes, biotic and abiotic factors such as herbivory, pathogens, plant age, phenological stages, temperature, drought, flooding, light exposure, shading, altitude, soil type, salinity, and ultraviolet radiation [22,33]. The biosynthesis of volatile compounds varies by plant organ, with EOs from bark, leaves, and fruits each exhibiting distinct chemical profiles. In general, we did not observe qualitative differences between the EOs from flooding and dry seasons. However, some metabolites showed relative area percentage variation depending on the collection period (Figure 2) For instance, the total content of hydrocarbon sesquiterpenes identified were higher in the EOs of flooding season, whereas hydrocarbon monoterpenes, oxygenated monoterpenes and oxygenated sesquiterpenes were present in greater proportions in the leaf EOs from dry season (Table 1).
Conversely, the leaves EO of Ocotea lancifolia was influenced by seasonality, presenting a higher quantity of oil during the spring (1.03%) and summer (0.96%; w/w%) compared to winter and autumn (0.56 and 0.6%, respectively) [32]. Seasonal differences in EO yield were also observed in N. megapotamica collected in southern Brazil, with the highest oil content (0.59%) found in young leaves during spring [34]. However, no significant seasonal variation was detected in samples collected in São Paulo, and the EO production in Nectandra lanceolata (0.17%) during spring and in Nectandra grandiflora (0.2%) during autumn were similar [35]. Nectandra grandiflora from Rio Grande do Sul also exhibited the highest EO content during spring [36].
It’s worth noting that in the southern regions of Brazil, seasons are markedly distinct, with temperatures and daylight hours varying throughout the year [37]. On the other hand, the Amazon region is near the equator line, and experiences less pronounced variations in temperature and daylight [37]. Even precipitation is more consistent throughout the year with certain areas, such as the várzea forest, undergoing periodic flooding as a stressor [37]. However, flooding did not appear to affect the oil yield in M. cymbarum.
2.3. Antibiofilm and Antibacterial Activity of Essential Oils from M. cymbarum
We investigated the impact of essential oils (EOs) from leaves, bark, and fruits of M. cymbarum on bacterial growth and biofilm formation in three Gram-positive bacterial strains: S. epidermidis (ATCC 35984), S. aureus (ATCC 25904), and M. luteus (ATCC 4698), and three Gram-negative, E. coli (ATCC 25922), P. aeruginosa (ATCC 27853), and P. aeruginosa (PAO1). In general, the EOs extracted from leaves and bark exhibited higher activity at higher concentrations (200 and 100 µg/ml) compared to fruit EO, demonstrating superior efficacy against Gram-positive than Gram-negative strains (Figure 3; Figure S4). The cell envelope of Gram-positive bacteria lacks an outer membrane and features a thick layer of peptidoglycan along with teichoic acid, which allows the penetration of hydrophobic molecules [38,39]. In contrast, Gram-negative bacteria present a thinner peptidoglycan layer, and present an outer membrane composed primarily of lipopolysaccharides, within a double layer of phospholipids [38,39]. Then, certain metabolites can inhibit the growth of Gram-positive bacteria, but result in weaker effect on gram-negative, particularly due to their hydrophobic properties, enabling penetration within the thick peptidoglycan cell wall from gram-positives [40].
In terms of this, the effectiveness of EOs can vary between Gram-positive and Gram-negative that is commonly linked to the EOs volatile composition [40]. For instance, thymol, a volatile compound associated with the antimicrobial properties of several EOs, has demonstrated greater effectiveness against Gram-positive bacteria due to its ability to disturb lipids on the plasma membrane [41]. The metabolite β-caryophyllene exhibited strong antibacterial activity, with minimum inhibitory concentrations (MIC) ranging from 3 to 14 μM, showing a more pronounced effect on gram-positive bacteria [42].
Figure 3.
Antibacterial and antibiofilm activities of the essential oils from leaves, bark and fruits of Mespilodaphne cymbarum against three Gram-positive bacterial strains. Control treatment contain the vehicle dimethyl sulfoxide 2% (DMSO 2%); treatment with the essential oils (EOs): Le-D-1 (leaves from the dry season of chemotype-1); Le-F-1 (leaves from the flooding season of chemotype-1); Le-D-2 (leaves from the dry season of chemotype-2); Le-F-2 (leaves from the flooding season of chemotype-2); Ba-D-1 (bark from the dry season of chemotype-1); Ba-F-1 (bark from the flooding season of chemotype-1); Fr-1 (fruits from the flooding season of chemotype-1). (*) Significant activity compared with the control by paired t-test (*p < 0.05).
Figure 3.
Antibacterial and antibiofilm activities of the essential oils from leaves, bark and fruits of Mespilodaphne cymbarum against three Gram-positive bacterial strains. Control treatment contain the vehicle dimethyl sulfoxide 2% (DMSO 2%); treatment with the essential oils (EOs): Le-D-1 (leaves from the dry season of chemotype-1); Le-F-1 (leaves from the flooding season of chemotype-1); Le-D-2 (leaves from the dry season of chemotype-2); Le-F-2 (leaves from the flooding season of chemotype-2); Ba-D-1 (bark from the dry season of chemotype-1); Ba-F-1 (bark from the flooding season of chemotype-1); Fr-1 (fruits from the flooding season of chemotype-1). (*) Significant activity compared with the control by paired t-test (*p < 0.05).
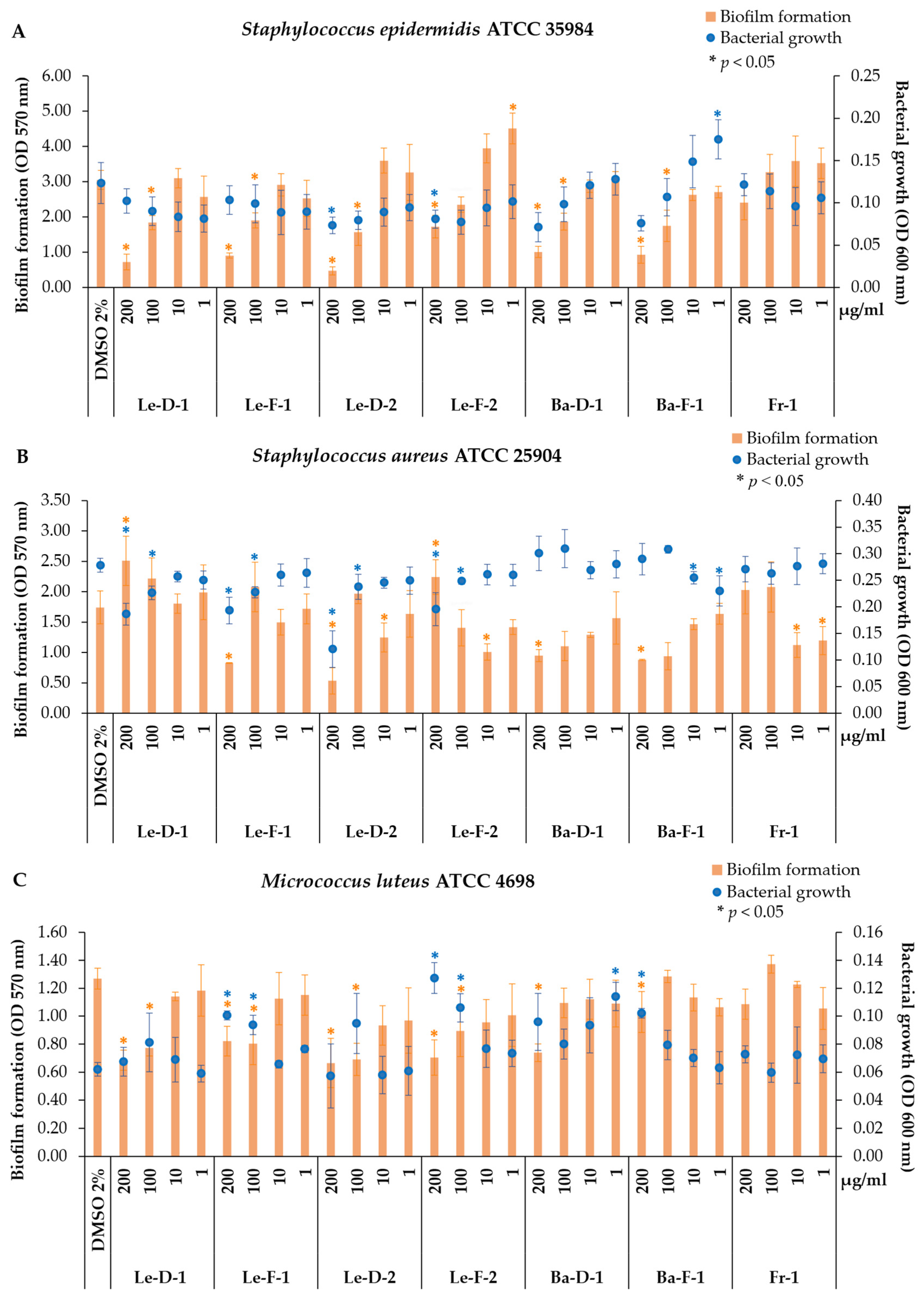
Considered a commensal organism of the skin microbiota, S. epidermidis is a Gram-positive, and coagulase-negative bacterium [43,44]. Under balanced conditions, S. epidermidis is often benign, even providing protection against other pathogens, including S. aureus [44]. However, S. epidermidis can also act as an opportunistic pathogen, frequently related to nosocomial infections due to its ability to produce biofilm, being one of the most common contaminants on medical devices, catheters, and implants [43,44]. In addition, this strain in skin conditions is frequently found in biofilms and is associated with atopic dermatitis [44].
Therefore, the Le-D-1 and Le-F-1 inhibited the biofilm formation of S. epidermidis, while the samples Le-D-2, Le-F-2, Ba-D-1, and Ba-F-1 affected the bacterial growth. The biofilm production is a key virulence factor in S. epidermidis, protecting this organism from immune defense cells, and reducing the action of antibiotics [43]. Since biofilm integrity is crucial for bacterial resistance, compounds capable of disrupting this matrix can be important in the treatment of infections [45].
Considering the major volatile compounds, chemotype-2 exhibited higher percentages of δ-cadinene (V67) and the unknown m/z 234 (V82), whereas chemotype-1 was characterized by higher levels of α-santalene (V43) and cryptone (V21) (Table 1). Interestingly, although the fruit essential oil displayed the highest percentage of α-santalene (V43), it showed no significant activity against S. epidermidis. These results suggest that α-santalene (V43) does not play a critical role in inhibiting biofilm formation in this strain.
Similarly, both chemotypes contain comparable amounts of β-caryophyllene (V42), yet they displayed distinct biological activities. This suggests that β-caryophyllene may not be solely responsible for the EOs’ effectiveness or that it could act synergistically with specific compounds unique to each chemotype. In contrast, cryptone (V21) is higher in chemotype-1, while the unknown m/z 234 (V82) and δ-cadinene (V67) is higher in chemotype-2, thus they could be related with the distinct activity of each oil. A moderate activity against S. aureus (ATCC 25932) for essential oil from Eucalyptus odorata, which showed around 20% of cryptone, was also described [46]. Another possibility is the synergistic effect of metabolites, since we found a notable difference between chemotype-1 and -2, as well as some volatiles exclusively of each chemotype (Table 01, Figure 2).
In contrast, bark EOs presented methyl eugenol (V41) (approx. 40%), followed by elemicin (V77) (Ba-F-1 = 12.1%; Ba-D-1 = 9%), myristicin (V68) (Ba-F-1 = 7.5%; Ba-D-1 = 10.5%), and δ-cadinene (V67) (Ba-F-1 = 7.6%; Ba-D-1 = 6%). The EO from Malaleuca bracteata showed approximately 88% methyl eugenol and demonstrated moderate activity against S. epidermidis with MIC value of 500 µg/ml [47]. Interestingly, the authors of the same study also evaluated the pure methyl eugenol and its MIC against S. epidermidis was higher than that of the EO (1000 µg/ml), suggesting a possible synergism among the volatiles present in this EO [47]. Another species with chemotypes rich in methyl eugenol (≈39% ) is Ocimum basilicum, which demonstrated bactericidal activity against S. epidermidis with the minimum bactericidal concentration (MBC) of approximately 416 µg/ml [48]. Additionally, this EO showed better results against Gram-positive bacteria compared to Gram-negative bacteria [48].
In the assays against S. aureus (ATCC 25904), all leaf EOs at higher concentrations (200 and 100 μg/ml) significantly decreased bacterial growth to some extent: Le-D-1 (33 and 19%), Le-F-1 (31 and 18%), Le-D-2 (57 and 14%), and Le-F-2 (30 and 11%) (Figure 3B). Leaf EOs of M. cymbarum from both chemotypes reduced S. aureus growth in different values. Among the major metabolites in these EOs, we identified β-caryophyllene, which is known for several biological properties, such as anti-inflammatory, antioxidant, cytotoxic, antifungal, and antimicrobial [42,49,50]. The structure of β-caryophyllene facilitates the penetration of cell membranes, potentiating the effects of other drugs [49]. Other essential oils containing high levels of β-caryophyllene, as well as this isolated metabolite, exhibited varying degrees of activity, being described as potent against S. aureus (MTCC 7405) with MIC of 3 μM [42], but weak activity against S. aureus (ATCC 25923) with MIC of 1 mg/ml [50]. Moreover, the EO of Murraya paniculata showed higher biological activity compared to the isolated β-caryophyllene, indicating a synergistic effect of its chemical components [50].
Additionally, antibiofilm activity was observed at the highest concentrations of bark EOs, with reductions of 45 (Ba-D-1) and 49% (Ba-F-1), and at the lowest concentrations of fruit EO, decreasing biofilm formation by 35% at 10 μg/ml and 31% at 1 μg/ml (Figure 3B). Bark EOs, rich in phenylpropanoids, particularly methyl eugenol (V41), demonstrated typical antibiofilm activity by reducing biofilm formation without inhibiting bacterial growth. EO from L. nobilis from plants collected in Sousse (Tunisia) presented as major volatiles 1,8-cineole (30.8%), methyl eugenol (15.6%), and α-terpinyl acetate (14.5%), and evidenced low antibacterial activity (MIC= 31.25 mg/ml), and moderate biofilm inhibition up to 70% at concentration of 1.95 mg/ml [51].
Another Gram-positive strain evaluated in our study was M. luteus, a bacterium of the mammalian skin microbiome, that is an obligate aerobic species, nonmotile, non-spore-forming, and catalase and oxidase-positive [52]. Generally, M. luteus is not considered harmful to humans, but it can act as an opportunistic pathogen in certain cases, particularly in immunocompromised individuals, causing pneumonia, bacteremia, endocarditis, peritonitis, ventriculitis, and septic arthritis [53]. This species is capable of producing biofilms and colonizing, for instance, prosthetic material, leading to infections such as endocarditis [53].
The EOs of M. cymbarum presented antibiofilm activity against M. luteus (ATCC 4698), reducing biofilm formation. No significant activity against M. luteus was observed for Fr-1, but all leaf EOs demonstrated antibiofilm activity at higher concentrations (Figure 3C), diminishing the biofilm formation by 30 to 47 %. Additionally, Ba-D-1 and Ba-F-1 reduced biofilm formation at 200 µg/ml by 42 and 19%, respectively. This antibiofilm potential could be explored as a therapeutic strategy against this pathogen. Since biofilms form complex barriers that hinder antibiotic efficacy, compounds capable of disrupting this matrix could serve as valuable adjuncts in infection treatment [40].
In contrast, the EOs of M. cymbarum were not active against the gram-negative bacterial strains, except for P. aeruginosa (ATCC 27853) (Figure S4; Supplementary Information). The leaf EOs from dry season (Le-D-1 and Le-D-2) at 200 µg/ml reduced P. aeruginosa (ATCC 27853) growth by 42 and 43%, respectively (Figure S4-A). Biofilm formation was inhibited by Ba-F-1 by approximately 19% at 10 µg/ml and by Fr-1 by about 22% at 100 µg/ml and 20% at 10 µg/ml (Figure S4-A). The EOs from M. cymbarum did not decrease the bacterial growth of P. aeruginosa (PAO1) and E. coli (ATCC 25922), even stimulated biofilm formation in some concentrations (Figure S4-B and C). Several studies have reported that EOs tend to be less effective against gram-negative than gram-positive bacteria, presumably due to differences in their membrane composition [40,48].
4. Materials and Methods
4.1. Study Area
The samplings of M. cymbarum were carried out in the Mamirauá Sustainable Development Reserve, which covers approximately 1,124,000 hectares in the Central Amazon, at the confluence of the Solimões, Japurá, and Auati-Paraná rivers [54]. The climate is tropical humid with annual average temperatures ranging from 28 to 30°C. The flood pulse is monomodal with high water levels (flooding) from May to July, peaking in June, and a dry period from September to November reaching its lowest levels in October [54]. It is important to note that in tropical humid climates, rainfall occurs throughout the year, but it is more abundant during specific periods, leading to occurrence of flooding.
4.2. Processing the Vegetal Material
We collected leaves and bark from 14 individuals with different diameters at breast height (DBH) to minimize the influence of plant age and ensure the inclusion of samples from different developmental stages (Table S1, Supplementary Information). The access to Brazilian genetic heritage was registered in the National Management System for Genetic Heritage and Associated Traditional Knowledge (SisGen) under the number A8B202A. Both leaves and bark were collected during the dry (September, 2021) and flooding (April, 2022) seasons. Fruits were only available during the flooding season (April, 2022) and we successfully collected fruits from eight specimens of chemotype-1 (n=8). The plant materials (bark and leaves) were dried in a controlled environment at 24°C. Due to their fleshy nature, we subjected the fruits to drying in an air-circulating oven at 45°C until they reached a consistent weight. Then, the dried materials were powdered by a knife mill. Additionally, fertile specimens were deposited in the Herbarium of Campo Grande, MS, under the catalog number (CGMS—83228). The plant identification was confirmed by professor Flávio Macedo Alves, a specialist in Lauraceae.
4.3. Extraction and Yield of Essential Oils (EOs) from M. cymbarum
The EOs were extracted by hydrodistillation for 4 h using a Clevenger apparatus. Leaves from different individuals were grouped based on the two chemotypes identified from a pilot study (data shown on Figure S1). Then, the samples were extracted separately, resulting in four distinct sample groups: leaves from the dry season of chemotype-1 (Le-D-1); and chemotype-2 (Le-D-2); and leaves from the flood season of chemotype-1 (Le-F-1); and chemotype-2 (Le-F-2). The fruits were available only during the flood season and from chemotype-1 (Fr-1). Bark EOs were exclusively from chemotype-1, collected in both the dry (Ba-D-1) and flood (Ba-F-1) seasons, as insufficient bark material was available for chemotype-2. The EOs were filtered through anhydrous sodium sulfate to remove water residues and stored in amber glass containers at -18°C until analysis. The extracted oil volume was used to calculate the yield as a percentage relative to the dry plant weight (% v/w).
4.4. Analysis of the Essential Oils (EOs) by Gas Chromatography Coupled to Mass Spectrometry (GC-MS)
The EOs were analyzed using a Shimadzu model QP2010 gas chromatograph coupled to a mass spectrometry (GC-MS), which was equipped with an RTx-5MS chromatographic column (30 m x 0.25 mm i.d., 0.25 µm film thickness) and an ionization energy of 70 eV. The temperature program was initiated at 60°C increasing at a rate of 3°C/min up to 240°C. The injector and interface temperatures were set at 250°C. Helium was used as carrier gas with a linear velocity of 41.6 cm/s and a pressure of 79.7 K Pa. Each EO was prepared at 10 mg/ml and 1 µl was injected into the GC-MS applying a split ratio of 1:10. The samples were injected in duplicates and all the samples were used to prepare a quality control sample, which was injected during the analysis. A series of C8–C40 n-alkane standards were injected and used to calculate the retention indices.
4.5. Data Processing
We aligned the GC-MS data using MetAlign 3.0 software and reduced the entrances with MSClust, resulting in 349 entrances. Then, we remove the duplicate entrances, those with low probability of being identified as compounds (cent.factor < 0.9), and peaks with ion intensity lower than 10,000. Next, we cross this data with the chromatographic data processed in software GC Solution Version 4.20 (Shimadzu), in which peak areas were integrated and only peaks with a relative area greater than 0.3% were selected, resulting in a total of 122 features. For peak integration, we applied a slope parameter based on the sample type: for leaves, the slope was set to max height/1000, and for bark and fruits, it was set to max height/2000. For statistical analysis, we excluded all peaks with a relative area below 0.3%.
4.6. Molecular Networking
The GC-MS data of the EOs from bark, leaves, and fruits were used to create a molecular network at Global Natural Products Social Molecular Networking (GNPS) platform (https://gnps.ucsd.edu, accessed on 23 April 2025) [57]. The MS/MS fragment ions within +/- 17 Da of the precursor m/z were removed. Additionally, the MS/MS spectra were refined by retaining only the top six fragment ions in the +/- 50 Da window throughout the spectrum, and the MS/MS fragment ion tolerance was 1 Da. A molecular network was built by retaining only edges with a cosine score greater than 0.7 and more than six matched peaks. Moreover, edges between two nodes were included only if each node was among the other’s top 10 most similar nodes. The maximum size of each molecular family was limited to 100 nodes. The library spectra were filtered using the same method as the input data. Only the matches with a score above 0.5 and at least 6 matched peaks were kept to the network spectra and library spectra. The network and library spectra results are available in the following link https://gnps.ucsd.edu/ProteoSAFe/status.jsp?task=3892fb335b6d47b2a2f76a0631165574. The molecular network was visualized and edited using Cytoscape software, version 3.10.2 [58].
4.7. Bacterial Strains
Six bacterial strains were used, including three Gram-positive—Staphylococcus epidermidis (ATCC 35984), Staphylococcus aureus (ATCC 25904), and Micrococcus luteus (ATCC 4698), and three Gram-negative—Escherichia coli (ATCC 25922), Pseudomonas aeruginosa (ATCC 27853), and Pseudomonas aeruginosa (PAO1).
4.8. Bacterial Growth and Biofilm Formation Assays
The evaluation of EOs on bacterial growth and biofilm formation was carried out using crystal violet microplate assay, adapted from Trentin et al. [59]. The EOs were initially diluted in 100% dimethyl sulfoxide (DMSO) and afterwards diluted to a final concentration of 2% DMSO for use in the assays. These solutions were then diluted to achieve final concentrations of 200, 100, 10 and 1 µg/ml, and 4 µl of each sample was added in a 96-well microplate. Wells containing 4 µl of DMSO 2% were used as evidence for growth control. Subsequently, 80 µl of sterile water and 40 µl of tryptone soy broth (TSB) culture medium (Oxoid Ltd., England) were added, except for the experiment with S. aureus ATCC 25904 that was carried out with Brain Heart Infusion Broth culture medium (BHI) supplemented with 1% glucose. Then, a bacterial suspension with an absorbance of 0.15 ± 0.01 (OD 600 nm) was added and the final absorption was read. The plates were incubated at 37°C for 24 h.
After 24 h, the absorbance (OD600 nm) was taken to evaluate bacterial growth. Subsequently, the volume of the wells was removed, washed three times with saline, and incubated for 1 h to dry the adhered biofilm in a 60°C oven. Following the addition of 200 µl of crystal violet, the samples were incubated for 15 min. The excess stain was removed through vigorous washing, and 200 µl of absolute ethanol was then added to solubilize the retained dye. Therefore, a new absorbance reading (OD 570 nm) was performed to quantify the violet crystal adhered to the biofilm. The results were compared with the control for the presence of DMSO 2%. Inhibition percentages for both bacterial growth and biofilm formation were calculated using the following formula:
% inhibition = [(OD control − OD sample)/OD control] x 100.
4.9. Statistical Analysis
The peak intensity matrix was statistically analyzed using R software version 4.2.3 (https://www.r-project.org/) [60]. The data were log-transformed (base 10) and scaled to minimize the influence of extreme values. Principal Component Analysis (PCA) was performed on the volatile intensities of the essential oils (EOs) extracted from leaves, bark, and fruits, using the “stats” [60] and “factoextra” packages [61]. To assess differences among EOs from bark, leaves, and fruits, a Permutational Analysis of Variance (PERMANOVA) was conducted using the “vegan” package [62]. The analysis was based on Bray-Curtis dissimilarities, with 999 permutations and a significance level set at p < 0.05.
Hierarchical Clustering Analysis (HCA), along with a heatmap, was carried out in MetaboAnalyst 6.0 [63] using the matrix of relative area (%) for each volatile compound. Initially, we analyzed the EOs from bark, fruits, and leaves. Then, we conducted HCA and generated a heatmap for the leaf EOs from both chemotypes to illustrate the distinctions between these groups exhibited in the supplementary information.
The potential antimicrobial and antibiofilm activities were evaluated using a paired t-test to compare each sample and concentration against the control (containing 2% DMSO). Absorbance values were used, and a significance threshold of p < 0.05 was applied. These analyses were conducted in Excel.
5. Conclusions
The EOs of M. cymbarum extracted from leaves, bark, and fruits exhibited distinct chemical compositions, resulting in different potentials against the evaluated pathogenic bacterial strains. Although the fruit EO demonstrated the weakest activity against the tested bacterial strains, yielded a high oil content and featured an interesting chemical profile, including considerable amounts of valuable sesquiterpenes, such as α- and β-santalene, α-trans-bergamotene, and α-santalol. The leaf and bark EOs showed higher activity, particularly against Gram-positive bacteria, acting either on bacterial growth and/or biofilm formation. Notably, the biological activity varied between the two leaf EO chemotypes, likely reflecting their chemical divergence. While major constituents are often associated with antimicrobial activity, it is plausible that minor compounds contribute synergistically to the overall effect. The enhanced activity observed in leaf EOs may also be linked to their greater chemical complexity, as they contained a higher number of volatile constituents compared to bark and fruit EOs.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Information about the 14 individuals of Mespilodaphne cymbarum collected in the Mamirauá Sustainable Development Reserve. Figure S1: Heatmap and Hierarchical Clustering Analysis (HCA) of the volatiles extracted by SPME from the leaves of 14 individuals of M. cymbarum, obtained in the flooding (F) and dry (D) seasons. Figure S2: Heatmap and Hierarchical Clustering Analysis (HCA) of the volatiles from the essential oils (EOs) extracted by hydrodistillation from the leaves of M. cymbarum. Figure S3: Molecular network of the essential oils extracted from the leaves, bark and fruits of M. cymbarum. Figure S4: Antimicrobial and antibiofilm activity of the essential oils (EOs) from leaves, bark and fruits of M. cymbarum against three gram-negative bacterial strains.
Author Contributions
Conceptualization, A.G.B., D.G., C.A.C. and D.B.S.; methodology, A.G.B., D.G., J.C.Q., J.S, and K.F.S.; formal analysis, A.G.B., J.S., A.J.M.; investigation, A.G.B., D.G., J.C.Q., J.S ; resources, D.G., A.J.M., C.A.C., D.B.S.; writing—original draft preparation, A.G.B. writing—review and editing, K.F.S, D.G., A.J.M., D.B.S., C.A.C; visualization, C.A.C, A.J.M., D.B.S.; supervision, D.B.S.; project administration, D.G., J.C.Q. A.G.B.; funding acquisition, D.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Fundação de Amparo à Pesquisa do Estado do Amazonas (State Funding Agency of Amazonas—FAPEAM), through Call No. 003/2020 (Programa de Apoio à Interiorização em Pesquisa e Inovação Tecnológica no Amazonas).
Data Availability Statement
The molecular network is available in the following link https://gnps.ucsd.edu/ProteoSAFe/status.jsp?task=3892fb335b6d47b2a2f76a0631165574.
Acknowledgments
The authors are grateful to the Federal University of Mato Grosso do Sul (UFMS), Fundação de Apoio ao Desenvolvimento do Ensino, Ciência e Tecnologia do estado de Mato Grosso do Sul (FUNDECT) (process number 71/032.390/2022), Instituto Nacional de Ciência e Tecnologia em Áreas Úmidas (INAU), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, process number 312194/2023-4), State Funding Agency of Amazonas (FAPEAM, No. 003/2020) and Mamirauá Institute for Sustainable Development (IDSM). The authors also thank Erivan Lima de Castro and Mario Jorge Lima de Castro for their valuable assistance in locating, identifying, and collecting the specimens, and Prof. Dr. Flávio Macedo Alves for the taxonomic identification of the species.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| EO/ EOs | Essential oil/ Essential oils |
| GC-MS | Gas chromatography-mass spectrometry |
| DMSO | Dimethyl sulfoxide |
| Le-D-1 | Leaves from chemotype-1 collected in the dry season |
| Le-F-1 | Leaves from chemotype-1 collected in the flooding season |
| Le-D-2 | Leaves from chemotype-2 collected in the dry season |
| Le-F-2 | Leaves from chemotype-2 collected in the flooding season |
| Ba-D-1 | Bark from chemotype-1 collected in the dry season |
| Ba-F-1 | Bark from chemotype-1 collected in the flooding season |
| Fr-1 | Fruit from chemotype-1 collected in the flooding season |
| ATCC | American Type Culture Collection |
| PAO1 | Pseudomonas aeruginosa resistant to chloramphenicol |
References
- Damasceno, C.S.B.; Fabri Higaki, N.T.; Dias, J. de F.G.; Miguel, M.D.; Miguel, O.G. Chemical Composition and Biological Activities of Essential Oils in the Family Lauraceae: A Systematic Review of the Literature. Planta Med. 2019, 85, 1054–1072. [CrossRef]
- Farias, K.S.; Alves, F.M.; Santos-Zanuncio, V.S.; de Sousa, P.T.J.; Silva, D.B.; Carollo, C.A. Global Distribution of the Chemical Constituents and Antibacterial Activity of Essential Oils in Lauraceae Family: A Review. S. Afr. J. Bot. 2023, 155, 214–222. [CrossRef]
- Trofimov, D.; Cadar, D.; Schmidt-Chanasit, J.; Rodrigues de Moraes, P.L.; Rohwer, J.G. A Comparative Analysis of Complete Chloroplast Genomes of Seven Ocotea Species (Lauraceae) Confirms Low Sequence Divergence within the Ocotea Complex. Sci Rep 2022, 12, 1120. [CrossRef]
- Trofimov, D.; de Moraes, P.L.R.; Rohwer, J.G. Towards a Phylogenetic Classification of the Ocotea Complex (Lauraceae): Classification Principles and Reinstatement of Mespilodaphne. Botanical Journal of the Linnean Society 2019, 190, 25–50. [CrossRef]
- Souza, D.A.T.; de Lima, A.A.; Wrege, M.S.; de Aguiar, A.V.; Bezerra, C. de S.; Meneses, C.H.S.G.; Lopes, R.; Ramos, S.L.F.; Paranatinga, I.L.D.; Lopes, M.T.G. Impacts of Climate Change on the Natural Distribution of Species of Lowland High and Low in the Amazon. Revista Arvore 2024, 48.
- da Silva Marinho, T.A.; Piedade, M.T.F.; Wittmann, F. Distribution and Population Structure of Four Central Amazonian High-Várzea Timber Species. Wetl. Ecol. Manag. 2010, 18, 665–677. [CrossRef]
- Muñoz-Acevedo, A.; López-Camacho, R.; González, M.C.; Castillo, O.J.; Cervantes-Díaz, M.; Celis, M. Prospecting for Non-Timber Forest Products by Chemical Analysis of Four Species of Lauraceae from the Amazon Region of Colombia. J Wood Sci 2024, 70, 33. [CrossRef]
- Diaz, A.M.P.; Gottlieb, H.E.; Gottlieb, O.R. Dehydrodieugenols from Ocotea cymbarum. Phytochemistry 1980, 19, 681–682. [CrossRef]
- Cabral, M.M.O.; Barbosa-Filho, J.M.; Maia, G.L.A.; Chaves, M.C.O.; Braga, M.V.; De Souza, W.; Soares, R.O.A. Neolignans from Plants in Northeastern Brazil (Lauraceae) with Activity against Trypanosoma cruzi. Experimental Parasitology 2010, 124, 319–324. [CrossRef]
- Ferreira, B.A.; de Moura, F.B.R.; Gomes, K.S.; da Silva Souza, D.C.; Lago, J.H.G.; Araújo, F. de A. Biseugenol from Ocotea cymbarum (Lauraceae) Attenuates Inflammation, Angiogenesis and Collagen Deposition of Sponge-Induced Fibrovascular Tissue in Mice. Inflammopharmacol 2023. [CrossRef]
- Ávila, W.A.D.; Suárez, L.E.C.; Fernando, J. Composición química del aceite esencial de Ocotea cymbarum Kunth (cascarillo y/o sasafrás) de la región Orinoquia. Rev. Cubana de Plantas Medicinales 2016, 21(3), 248-260,.
- Zoghbi, M. G. B.; Andrade, e H A; Santos, A S; Silva, M H L; Maia, J G S. Constituintes Voláteis de Espécies de Lauraceae com Ocorrência na Floresta Nacional de Caxiuanã—Melgaço—PA. In: Estação Científica Ferreira Penna—Dez Anos de Pesquisa, 2003, Belém, 2003.
- Passos, B.G.; De Albuquerque, R.D.D.G.; Muñoz-Acevedo, A.; Echeverria, J.; Llaure-Mora, A.M.; Ganoza-Yupanqui, M.L.; Rocha, L. Essential Oils from Ocotea Species: Chemical Variety, Biological Activities and Geographic Availability. Fitoterapia 2022, 156, 105065. [CrossRef]
- Shukis, A.J.; Wachs, Herman. Determination of Safrole in the Oil of Ocotea cymbarum. A Cryoscopic Method. Anal. Chem. 1948, 20, 248–249. [CrossRef]
- Barbosa-Filho, J.M.; Cunha, R.M.; Dias, C.S.; Athayde-Filho, P.F.; Silva, M.S.; Da-Cunha, E.V.L.; Machado, M.I.L.; Craveiro, A.A.; Medeiros, I.A. GC-MS Analysis and Cardiovascular Activity of the Essential Oil of Ocotea Duckei. Rev. Brasileira Farmacognosia 2008, 18, 37–41. [CrossRef]
- Chaverri, C.; Díaz, C.; Cicció, J.F. Chemical Analysis of Essential Oils from Ocotea gomezii W.C. Burger and Ocotea morae Gómez-Laur. (Lauraceae) Collected at “Reserva Biológica Alberto M. Brenes” in Costa Rica and Their Cytotoxic Activity on Tumor Cell Lines. J. Braz. Chem. Soc. 2011, 22, 741–745. [CrossRef]
- Silva, J.K.; Da Trindade, R.; Moreira, E.C.; Maia, J.G.S.; Dosoky, N.S.; Miller, R.S.; Cseke, L.J.; Setzer, W.N. Chemical Diversity, Biological Activity, and Genetic Aspects of Three Ocotea Species from the Amazon. International Journal of Molecular Sciences 2017, 18, 1081. [CrossRef]
- Yamaguchi, K.K.L.; Pedrosa, T.D.N.; de Vasconcellos, M.C.; Lima, E.S.; Veiga-Junior, V.F. Ocotea (Lauraceae) Amazonian Essential Oils Chemical Composition and Their Tyrosinase Inhibition to Use in Cosmetics. Bol.latinoam. y del Caribe de Plant.med.y aromat. 2020, 19, 519–526. [CrossRef]
- Gilardoni, G.; Montalván, M.; Vélez, M.; Malagón, O. Chemical and Enantioselective Analysis of the Essential Oils from Different Morphological Structures of Ocotea quixos (Lam.) Kosterm. Plants 2021, 10. [CrossRef]
- Takaku, S.; Haber, W.A.; Setzer, W.N. Leaf Essential Oil Composition of 10 Species of Ocotea (Lauraceae) from Monteverde, Costa Rica. Biochem. Syst. Ecol. 2007, 35, 525–532. [CrossRef]
- Rambo, M.A.; Soares, K.D.; Danielli, L.J.; Lana, D.F.D.; Bordignon, S.A. de L.; Fuentefria, A.M.; Apel, M.A. Biological Activities of Essential Oils from Six Genotypes of Four Ocotea Species. Braz. J. Pharm. Sci. 2022, 58, e181097. [CrossRef]
- Xavier, J.K.A.M.; Alves, N.S.F.; Setzer, W.N.; da Silva, J.K.R. Chemical Diversity and Biological Activities of Essential Oils from Licaria, Nectrandra and Ocotea Species (Lauraceae) with Occurrence in Brazilian Biomes. Biomolecules 2020, 10, 869. [CrossRef]
- Sacchetti, G.; Guerrini, A.; Noriega, P.; Bianchi, A.; Bruni, R. Essential Oil of Wild Ocotea quixos (Lam.) Kosterm. (Lauraceae) Leaves from Amazonian Ecuador. Flavour Fragrance J. 2006, 21, 674–676. [CrossRef]
- Radice, M.; Pietrantoni, A.; Guerrini, A.; Tacchini, M.; Sacchetti, G.; Chiurato, M.; Venturi, G.; Fortuna, C. Inhibitory Effect of Ocotea Quixos (Lam.) Kosterm. and Piper Aduncum L. Essential Oils from Ecuador on West Nile Virus Infection. Plant Biosyst. 2019, 153, 344–351. [CrossRef]
- Arteaga-Crespo, Y.; Ureta-Leones, D.; García-Quintana, Y.; Montalván, M.; Gilardoni, G.; Malagón, O. Preliminary Predictive Model of Termiticidal and Repellent Activities of Essential Oil Extracted from Ocotea quixos Leaves against Nasutitermes corniger (Isoptera: Termitidae) Using One-Factor Response Surface Methodology Design. Agronomy 2021, 11. [CrossRef]
- Valarezo, E.; Vullien, A.; Conde-Rojas, D. Variability of the Chemical Composition of the Essential Oil from the Amazonian Ishpingo Species (Ocotea quixos). Molecules 2021, 26, 3961. [CrossRef]
- Bruni, R.; Medici, A.; Andreotti, E.; Fantin, C.; Muzzoli, M.; Dehesa, M.; Romagnoli, C.; Sacchetti, G. Chemical Composition and Biological Activities of Ishpingo Essential Oil, a Traditional Ecuadorian Spice from Ocotea quixos (Lam.) Kosterm. (Lauraceae) Flower Calices. Food Chem. 2004, 85, 415–421. [CrossRef]
- He, X.; Wang, S.; Shi, J.; Sun, Z.; Lei, Z.; Yin, Z.; Qian, Z.; Tang, H.; Xie, H. Genotypic and Environmental Effects on the Volatile Chemotype of Valeriana jatamansi Jones. Front. Plant Sci. 2018, 9. [CrossRef]
- Karban, R.; Wetzel, W.C.; Shiojiri, K.; Ishizaki, S.; Ramirez, S.R.; Blande, J.D. Deciphering the Language of Plant Communication: Volatile Chemotypes of Sagebrush. New Phytologist 2014, 204, 380–385. [CrossRef]
- Farias, K.D.S.; Delatte, T.; Arruda, R.D.C.D.O.; Alves, F.M.; Silva, D.B.; Beekwilder, J.; Carollo, C.A. In Depth Investigation of the Metabolism of Nectandra megapotamica Chemotypes. PLoS ONE 2018, 13, e0201996. [CrossRef]
- El Hachlafi, N.; Benkhaira, N.; Mssillou, I.; Touhtouh, J.; Aanniz, T.; Chamkhi, I.; El Omari, N.; Khalid, A.; Abdalla, A.N.; Aboulagras, S.; et al. Natural Sources and Pharmacological Properties of Santalenes and Santalols. Industrial Crops and Products 2024, 214, 118567. [CrossRef]
- Silva, D.T.; Pinheiro, C.G.; Bianchini, N.H.; Batista, B.F.; Diefenthaeler, J.; Brião Muniz, M.D.F.; Heinzmann, B.M. Microbiological Damage Influences the Content, Chemical Composition and the Antifungal Activity of Essential Oils in a Wild-Growing Population of Ocotea lancifolia (Schott) Mez. J. Essent. Oil Res. 2018, 30, 265–277. [CrossRef]
- Escobar-Bravo, R.; Lin, P.-A.; Waterman, J.M.; Erb, M. Dynamic Environmental Interactions Shaped by Vegetative Plant Volatiles. Nat. Prod. Rep. 2023, 40, 840–865. [CrossRef]
- Amaral, L.D.P.; Tondolo, J.S.M.; Schindler, B.; Silva, D.T.D.; Pinheiro, C.G.; Longhi, S.J.; Mallmann, C.A.; Heinzmann, B.M. Seasonal Influence on the Essential Oil Production of Nectandra Megapotamica (Spreng.) Mez. Braz. arch. biol. technol. 2015, 58, 12–21. [CrossRef]
- Ferraz, E.D.O.; Vieira, M.A.R.; Ferreira, M.I.; Fernandes Junior, A.; Marques, M.O.M.; Minatel, I.O.; Albano, M.; Sambo, P.; Lima, G.P.P. Seasonality Effects on Chemical Composition, Antibacterial Activity and Essential Oil Yield of Three Species of Nectandra. PLoS ONE 2018, 13, e0204132. [CrossRef]
- Silva, D.T.D.; Bianchini, N.H.; Amaral, L.D.P.; Longhi, S.J.; Heinzmann, B.M. Análise do Efeito da Sazonalidade Sobre o Rendimento do Óleo Essencial das Folhas de Nectandra grandiflora Nees. Rev. Árvore 2015, 39, 1065–1072. [CrossRef]
- Lima, R.F. de; Aparecido, L.E. de O.; Torsoni, G.B.; Rolim, G. de S. Climate Change Assessment in Brazil: Utilizing the Köppen-Geiger (1936) Climate Classification. Rev. bras. meteorol. 2024, 38, e38230001. [CrossRef]
- Nourbakhsh, F.; Lotfalizadeh, M.; Badpeyma, M.; Shakeri, A.; Soheili, V. From Plants to Antimicrobials: Natural Products against Bacterial Membranes. Phytotherapy Research 2022, 36, 33–52. [CrossRef]
- Ruhal, R.; Kataria, R. Biofilm Patterns in Gram-Positive and Gram-Negative Bacteria. Microbiological Research 2021, 251, 126829. [CrossRef]
- Aljaafari, M.N.; AlAli, A.O.; Baqais, L.; Alqubaisy, M.; AlAli, M.; Molouki, A.; Ong-Abdullah, J.; Abushelaibi, A.; Lai, K.-S.; Lim, S.-H.E. An Overview of the Potential Therapeutic Applications of Essential Oils. Molecules 2021, 26, 628. [CrossRef]
- Cáceres, M.; Hidalgo, W.; Stashenko, E.; Torres, R.; Ortiz, C. Essential Oils of Aromatic Plants with Antibacterial, Anti-Biofilm and Anti-Quorum Sensing Activities against Pathogenic Bacteria. Antibiotics 2020, 9, 147. [CrossRef]
- Dahham, S.S.; Tabana, Y.M.; Iqbal, M.A.; Ahamed, M.B.K.; Ezzat, M.O.; Majid, A.S.A.; Majid, A.M.S.A. The Anticancer, Antioxidant and Antimicrobial Properties of the Sesquiterpene β-Caryophyllene from the Essential Oil of Aquilaria Crassna. Molecules 2015, 20, 11808–11829. [CrossRef]
- Otto, M. Staphylococcus epidermidis — the “accidental” Pathogen. Nat Rev Microbiol 2009, 7, 555–567. [CrossRef]
- Severn, M.M.; Horswill, A.R. Staphylococcus epidermidis and Its Dual Lifestyle in Skin Health and Infection. Nat Rev Microbiol 2023, 21, 97–111. [CrossRef]
- Verderosa, A.D.; Totsika, M.; Fairfull-Smith, K.E. Bacterial Biofilm Eradication Agents: A Current Review. Frontiers in Chemistry 2019, 7.
- Elaissi, A.; Rouis, Z.; Salem, N.A.B.; Mabrouk, S.; ben Salem, Y.; Salah, K.B.H.; Aouni, M.; Farhat, F.; Chemli, R.; Harzallah-Skhiri, F.; et al. Chemical Composition of 8 Eucalyptus Species’ Essential Oils and the Evaluation of Their Antibacterial, Antifungal and Antiviral Activities. BMC Complement Altern Med 2012, 12, 81. [CrossRef]
- Goswami, P.; Verma, S.K.; Chauhan, A.; Venkatesha, K.; Verma, R.S.; Singh, V.R.; Darokar, M.P.; Chanotiya, C.S.; Padalia, R.C. Chemical Composition and Antibacterial Activity of Melaleuca bracteata Essential Oil from India: A Natural Source of Methyl Eugenol. Natural Product Communications 2017, 12, 1934578X1701200. [CrossRef]
- Joshi, R. Chemical Composition and Antimicrobial Activity of the Essential Oil of Ocimum basilicum L. (Sweet Basil) from Western Ghats of North West Karnataka, India. Ancient Sci Life 2014, 33, 149. [CrossRef]
- Francomano, F.; Caruso, A.; Barbarossa, A.; Fazio, A.; La Torre, C.; Ceramella, J.; Mallamaci, R.; Saturnino, C.; Iacopetta, D.; Sinicropi, M.S. β-Caryophyllene: A Sesquiterpene with Countless Biological Properties. Applied Sciences 2019, 9, 5420. [CrossRef]
- Selestino Neta, M.C.; Vittorazzi, C.; Guimarães, A.C.; Martins, J.D.L.; Fronza, M.; Endringer, D.C.; Scherer, R. Effects of β-Caryophyllene and Murraya paniculata Essential Oil in the Murine Hepatoma Cells and in the Bacteria and Fungi 24-h Time–Kill Curve Studies. Pharmaceutical Biology 2017, 55, 190–197. [CrossRef]
- Merghni, A.; Marzouki, H.; Hentati, H.; Aouni, M.; Mastouri, M. Antibacterial and Antibiofilm Activities of Laurus nobilis L. Essential Oil against Staphylococcus aureus Strains Associated with Oral Infections. Current Research in Translational Medicine 2016, 64, 29–34. [CrossRef]
- Dib, J.R.; Liebl, W.; Wagenknecht, M.; Farías, M.E.; Meinhardt, F. Extrachromosomal Genetic Elements in Micrococcus. Appl Microbiol Biotechnol 2013, 97, 63–75. [CrossRef]
- Rodriguez-Nava, G.; Mohamed, A.; Yanez-Bello, M.A.; Trelles-Garcia, D.P. Advances in Medicine and Positive Natural Selection: Prosthetic Valve Endocarditis Due to Biofilm Producer Micrococcus luteus. IDCases 2020, 20, e00743. [CrossRef]
- Ramalho, E.E.; Macedo, J.; Vieira, T.M.; Valsecchi, J.; Calvimontes, J.; Marmontel, M.; Queiroz, H.L. Ciclo Hidrológico nos Ambientes de Várzea da Reserva de Desenvolvimento Sustentável Mamirauá—Médio Rio Solimões, Período De 1990 A 2008. Scientific Magazine UAKARI 2009, 5(1), 61-87.
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography Mass Spectroscopy; Allured Publishing Corporation: Carol Stream, Ill, 2007; ISBN 978-1-932633-21-4.
- Babushok, V.I.; Linstrom, P.J.; Zenkevich, I.G. Retention Indices for Frequently Reported Compounds of Plant Essential Oils. Journal of Physical and Chemical Reference Data 2011, 40, 043101. [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat Biotechnol 2016, 34, 828–837. [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [CrossRef]
- Trentin, D. da S.; Giordani, R.B.; Zimmer, K.R.; da Silva, A.G.; da Silva, M.V.; Correia, M.T. dos S.; Baumvol, I.J.R.; Macedo, A.J. Potential of Medicinal Plants from the Brazilian Semi-Arid Region (Caatinga) against Staphylococcus epidermidis Planktonic and Biofilm Lifestyles. Journal of Ethnopharmacology 2011, 137, 327–335. [CrossRef]
- R Core Team (2023). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2003, https://www.R-project.org/.
- Kassambara A, Mundt F (2020). factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R package version 1.0.7, 2020, https://CRAN.R-project.org/package=factoextra.
- Oksanen J, Simpson G, Blanchet F, Kindt R, Legendre P, Minchin P, O’Hara R, Solymos P, Stevens M, Szoecs E, Wagner H, Barbour M, Bedward M, Bolker B, Borcard D, Carvalho G, Chirico M, De Caceres M, Durand S, Evangelista H, FitzJohn R, Friendly M, Furneaux B, Hannigan G, Hill M, Lahti L, McGlinn D, Ouellette M, Ribeiro Cunha E, Smith T, Stier A, Ter Braak C, Weedon J (2022). vegan: Community Ecology Package. R package version 2.6-4, 2022, https://CRAN.R-project.org/package=vegan.
- Pang, Z.; Lu, Y.; Zhou, G.; Hui, F.; Xu, L.; Viau, C.; Spigelman, A.F.; MacDonald, P.E.; Wishart, D.S.; Li, S.; et al. MetaboAnalyst 6.0: Towards a Unified Platform for Metabolomics Data Processing, Analysis and Interpretation. Nucleic Acids Research 2024, 52, W398–W406. [CrossRef]
Figure 1.
Principal Component Analysis (PCA) of the essential oils (EOs) from leaves, bark and fruits from Mespilodaphne cymbarum (a) score plot illustrating three distinct groups of bark (Ba), fruit (Fr) and leaves (Le) (PERMANOVA, R2 = 0.9078; p < 0.001); (b) biplot exhibiting all volatiles from the EOs.
Figure 1.
Principal Component Analysis (PCA) of the essential oils (EOs) from leaves, bark and fruits from Mespilodaphne cymbarum (a) score plot illustrating three distinct groups of bark (Ba), fruit (Fr) and leaves (Le) (PERMANOVA, R2 = 0.9078; p < 0.001); (b) biplot exhibiting all volatiles from the EOs.
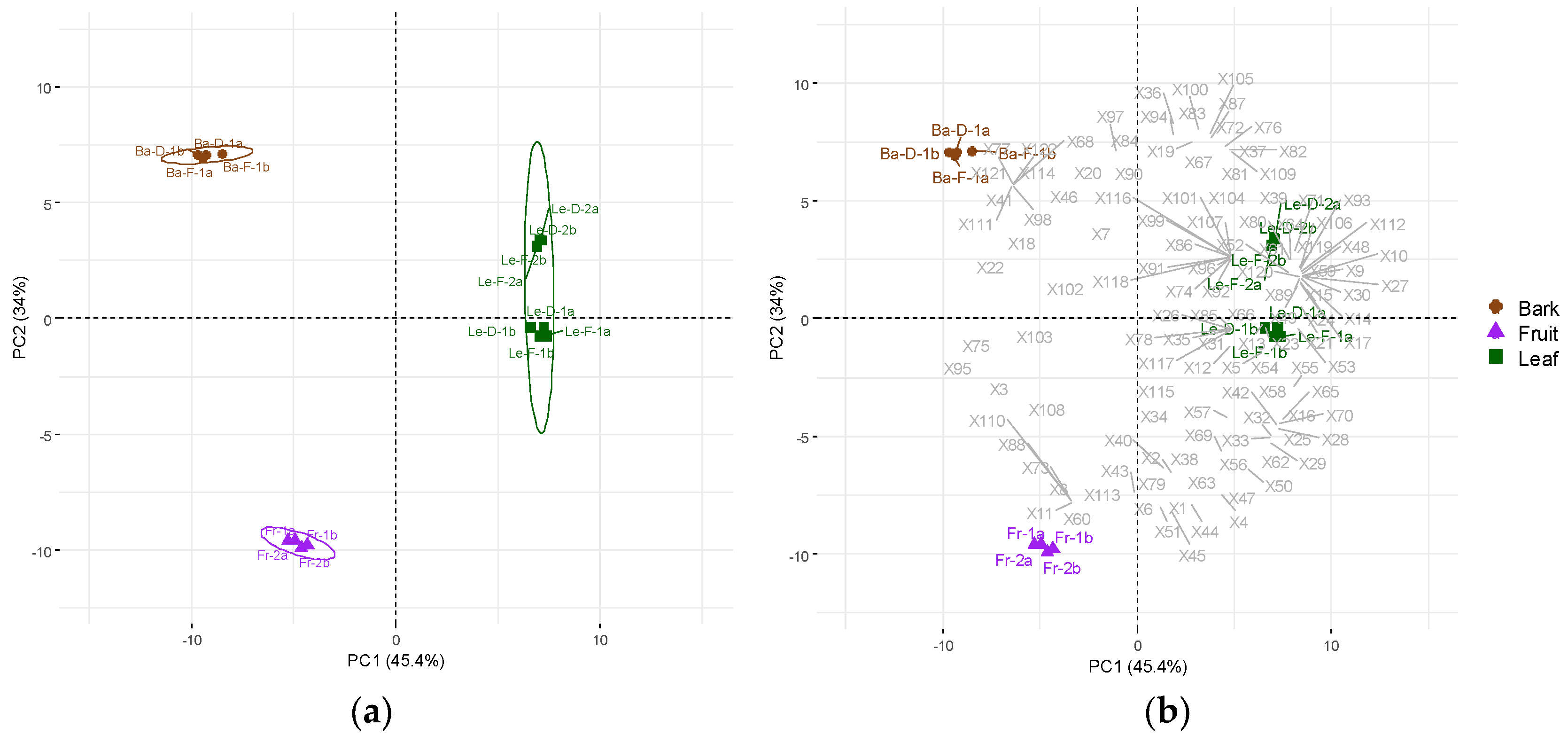
Figure 2.
Heatmap and Hierarchical Clustering Analysis (HCA) showing the relative area (%) of each volatile compound of the essential oils (EOs) from bark (Ba), leaves (Le) and fruits (Fr) of Mespilodaphne cymbarum. Bark and leaf samples were collected in both the dry (D) and flooding (F) seasons. A, B, C, and D represent the analytical replicates and 1 and 2 are the chemotypes.
Figure 2.
Heatmap and Hierarchical Clustering Analysis (HCA) showing the relative area (%) of each volatile compound of the essential oils (EOs) from bark (Ba), leaves (Le) and fruits (Fr) of Mespilodaphne cymbarum. Bark and leaf samples were collected in both the dry (D) and flooding (F) seasons. A, B, C, and D represent the analytical replicates and 1 and 2 are the chemotypes.
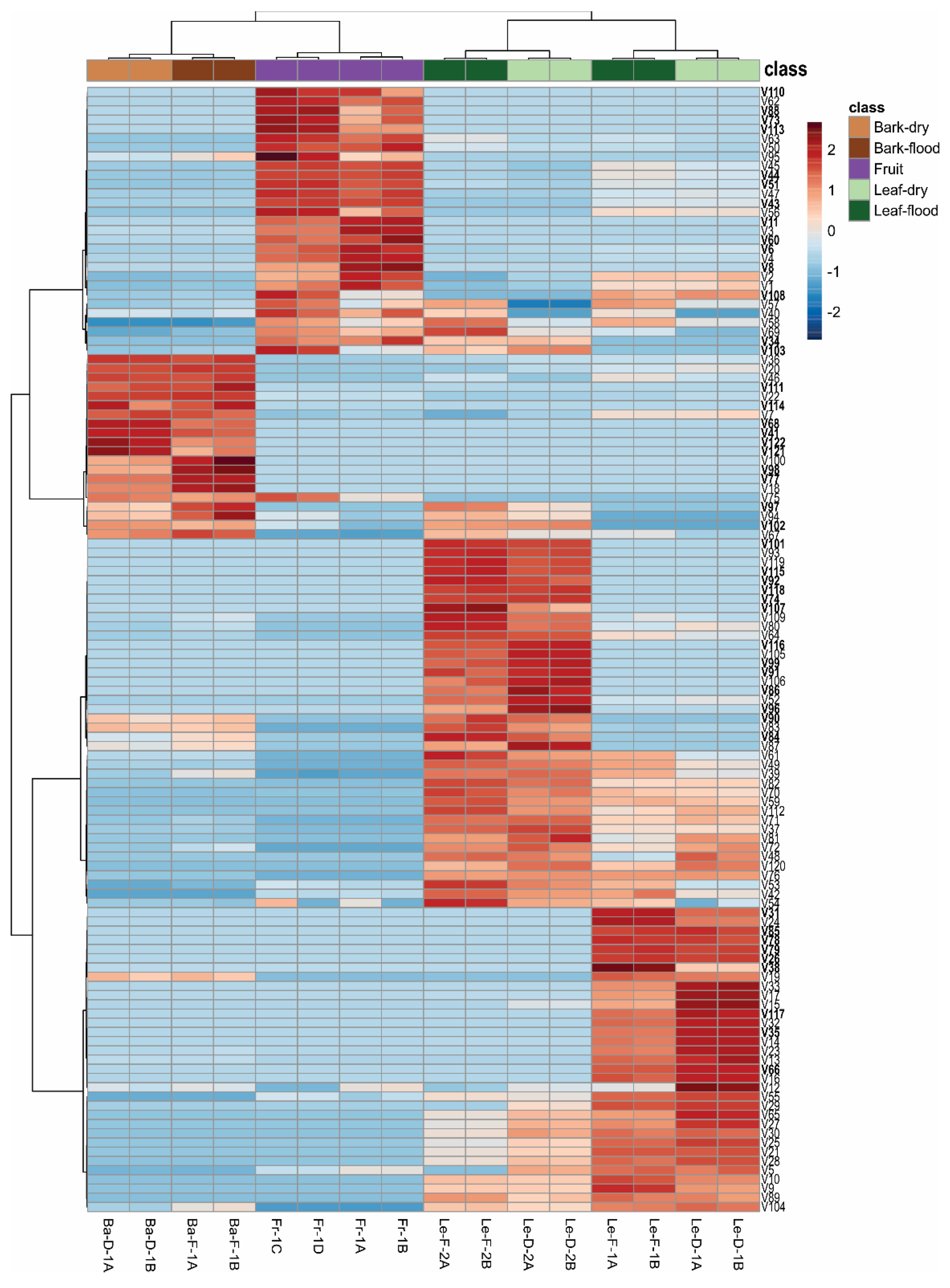

Table 1.
Chemical composition and yield (%) of the essential oils (EOs) extracted from leaves (flooding and dry), bark (flooding and dry), and fruits of Mespilodaphne cymbarum, collected in the Mamirauá Sustainable Development Reserve, Uarini, AM, Brazil. These EOs were analyzed by GC-MS using a RTx-5MS chromatographic column.
Table 1.
Chemical composition and yield (%) of the essential oils (EOs) extracted from leaves (flooding and dry), bark (flooding and dry), and fruits of Mespilodaphne cymbarum, collected in the Mamirauá Sustainable Development Reserve, Uarini, AM, Brazil. These EOs were analyzed by GC-MS using a RTx-5MS chromatographic column.
| VOC | IR* | Compound Name | Le-F-1 | Le-D-1 | Le-F-2 | Le-D-2 | Fr-1 | Ba-F-1 | Ba-D-1 | Class** |
|---|---|---|---|---|---|---|---|---|---|---|
| V1 | 938 | α-Pinene | 1.71 ±0.021 | 1.88 ±0.247 | 0.1 ±0.007 | 0.38 ±0.007 | 3.32 ±0.583 | 0.21 ±0.021 | 0.08 ±0.001 | HM |
| V2 | 979 | β-Pinene | 0.83 ±0.007 | 0.87 ±0.078 | <0.1 | 0.26 ±0.007 | 1.2 ±0.289 | 0.13 ±0.007 | 0.08 ±0.007 | HM |
| V3 | 990 | Myrcene | - | - | - | - | 0.6 ±0.134 | <0.1 | <0.1 | HM |
| V4 | 1004 | α-Phellandrene | 0.11 ±0.007 | 0.11 ±0.007 | <0.1 | <0.1 | 1.76 ±0.246 | - | - | HM |
| V5 | 1026 | p-Cymene | 2.64 ±0.078 | 2.59 ±0.198 | 0.2 ±0.007 | 2.04 ±0.042 | 0.96 ±0.237 | <0.1 | <0.1 | HM |
| V6 | 1031 | Limonene | 2.37 ±0.042 | 2.89 ±0.233 | 0.33 ±0.007 | 1.38 ±0.028 | 19.32 ±2.79 | 0.17 ±0 | 0.22 ±0.014 | HM |
| V7 | 1034 | 1,8-Cineole | 1.73 ±0.007 | 1.9 ±0.106 | 0.22 ±0.007 | 0.76 ±0.021 | 0.62 ±0.114 | 3.33 ±0.177 | 3.44 ±0.184 | OM |
| V8 | 1050 | trans-β-Ocimene | - | - | - | - | 0.54 ±0.179 | - | - | HM |
| V9 | 1074 | cis-Linalool oxide | 0.38 ±0.007 | 0.28 ±0.007 | 0.21 ±0.002 | 0.21 ±0.007 | - | - | - | OM |
| V10 | 1088 | trans-Linalool oxide | 0.27 ±0.007 | 0.22 ±0.001 | 0.17 ±0.001 | 0.16 ±0.001 | - | - | - | OM |
| V11 | 1088 | Terpinolene | - | - | - | - | 0.5 ±0.074 | - | - | HM |
| V12 | 1098 | Linalool | 0.11 ±0.007 | 0.34 ±0.007 | <0.1 | 0.1 ±0.007 | <0.1 | 0.1 ±0.007 | 0.08 ±0.014 | OM |
| V13 | 1141 | trans-Pinocarveol | 0.86 ±0.021 | 1.13 ±0.113 | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 | OM |
| V14 | 1144 | cis-Verbenol | 0.26 ±0.007 | 0.37 ±0.001 | <0.1 | <0.1 | - | - | - | OM |
| V15 | 1148 | trans-Verbenol | 0.63 ±0.014 | 1.3 ±0.014 | <0.1 | 0.2 ±0.007 | - | - | - | OM |
| V16 | 1153 | NI (m/z 138) | 0.67 ±0.035 | 0.8 ±0.007 | <0.1 | <0.1 | <0.1 | - | - | |
| V17 | 1165 | Pinocarvone | 0.27 ±0.007 | 0.49 ±0.001 | <0.1 | <0.1 | - | - | - | OM |
| V18 | 1169 | Borneol | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 | 0.88 ±0.049 | 0.65 ±0.396 | OM |
| V19 | 1170 | p-Mentha-1,5-dien-8-ol | 0.55 ±0.021 | 0.5 ±0.007 | <0.1 | <0.1 | - | <0.1 | 0.35 ±0.042 | OM |
| V20 | 1179 | Terpinen-4-ol | 0.55 ±0.007 | 0.76 ±0.007 | 0.18 ±0.007 | 0.41 ±0.021 | 0.18 ±0.031 | 2.07 ±0.014 | 1.97 ±0.021 | OM |
| V21 | 1188 | Cryptone | 5.03 ±0.014 | 5.08 ±0.134 | 1.94 ±0.028 | 2.64 ±0.035 | <0.1 | <0.1 | <0.1 | Ke |
| V22 | 1192 | α-Terpineol | 0.42 ±0.014 | 0.64 ±0.007 | 0.23 ±0.007 | 0.29 ±0.007 | 0.75 ±0.049 | 4.5 ±0.078 | 4.38 ±0.042 | OM |
| V23 | 1197 | Myrtenal | 0.72 ±0.014 | 1.1 ±0.007 | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 | OM |
| V24 | 1210 | Verbenone | 1.29 ±0.021 | 0.86 ±0.007 | <0.1 | <0.1 | - | - | - | OM |
| V25 | 1221 | trans-Carveol | 0.62 ±0.007 | 0.7 ±0.007 | 0.22 ±0.007 | 0.34 ±0.014 | <0.1 | - | - | OM |
| V26 | 1226 | NI | 0.57 ±0.007 | 0.56 ±0.007 | - | - | - | - | - | |
| V27 | 1243 | p-Isopropylbenzaldehyde (p-Cumic aldehyde) | 1.94 ±0.057 | 2.58 ±0.028 | 0.95 ±0.035 | 1.62 ±0.049 | - | - | - | OM |
| V28 | 1247 | Carvone | 0.8 ±0.028 | 0.89 ±0.007 | 0.35 ±0.007 | 0.52 ±0.014 | <0.1 | - | - | OM |
| V29 | 1276 | p-Menth-1-en-7-al | 0.63 ±0.007 | 0.67 ±0.001 | <0.1 | 0.26 ±0.014 | <0.1 | - | - | OM |
| V30 | 1292 | p-Cymen-7-ol | 0.61 ±0.028 | 0.64 ±0.014 | 0.32 ±0.007 | 0.52 ±0.035 | - | - | - | OM |
| V31 | 1308 | NI | 0.74 ±0.014 | 0.59 ±0.007 | - | - | - | - | - | |
| V32 | 1331 | NI | 1.04 ±0.014 | 1.39 ±0.014 | <0.1 | <0.1 | <0.1 | - | - | |
| V33 | 1334 | NI | 0.61 ±0.007 | 1.05 ±0.028 | <0.1 | <0.1 | <0.1 | - | - | |
| V34 | 1337 | δ-Elemene | - | - | 0.26 ±0.007 | 0.26 ±0.014 | 0.37 ±0.045 | - | - | HS |
| V35 | 1339 | NI | 1.57 ±0.007 | 2.12 ±0.014 | - | - | - | - | - | |
| V36 | 1351 | α-Cubebene | 0.18 ±0.007 | 0.21 ±0.014 | <0.1 | <0.1 | - | 1.1 ±0.049 | 1.12 ±0.014 | HS |
| V37 | 1368 | Cyclosativene | 0.32 ±0.007 | 0.33 ±0.007 | 0.62 ±0.007 | 0.68 ±0.007 | - | <0.1 | <0.1 | HS |
| V38 | 1370 | NI | 1.07 ±0.035 | 0.35 ±0.001 | - | - | <0.1 | - | - | |
| V39 | 1376 | α-Copaene | 1.39 ±0.021 | 0.91 ±0.014 | 1.65 ±0.021 | 1.7 ±0.014 | 0.26 ±0.024 | 0.95 ±0.049 | 0.53 ±0.014 | HS |
| V40 | 1391 | β-Elemene | 0.18 ±0.007 | <0.1 | 0.22 ±0.007 | <0.1 | 0.32 ±0.051 | <0.1 | <0.1 | HS |
| V41 | 1411 | Methyl eugenol | - | - | - | - | - | 37.49 ±1.29 | 43.61 ±0.49 | Ph |
| V42 | 1419 | β-Caryophyllene | 7.91 ±0.926 | 4.77 ±0.106 | 8.99 ±0.071 | 7.61 ±0.113 | 2.97 ±0.093 | - | - | HS |
| V43 | 1421 | α-Santalene | 6.17 ±0.085 | 6.29 ±0.226 | <0.1 | <0.1 | 26.41 ±0.50 | 1.69 ±0.028 | 1.11 ±0.028 | HS |
| V44 | 1437 | trans-α-Bergamotene | 2.85 ±0.042 | 1.94 ±0.014 | 0.79 ±0.007 | 0.33 ±0.007 | 8.18 ±0.181 | 0.27 ±0.021 | 0.2 ±0.014 | HS |
| V45 | 1439 | α-Guaiene | 0.68 ±0.007 | 0.45 ±0.001 | 0.21 ±0.001 | <0.1 | 1.95 ±0.054 | <0.1 | <0.1 | HS |
| V46 | 1444 | 6,9-Guaiadiene | 0.29 ±0.007 | 0.15 ±0.021 | 0.16 ±0.001 | <0.1 | <0.1 | 0.79 ±0.042 | 0.79 ±0.014 | HS |
| V47 | 1448 | epi-β-Santalene | 0.3 ±0.007 | 0.37 ±0.049 | <0.1 | <0.1 | 1.61 ±0.107 | - | - | HS |
| V48 | 1451 | Allo-Aromadendrene | <0.1 | 0.3 ±0.035 | 0.31 ±0.007 | 0.28 ±0.021 | - | - | - | HS |
| V49 | 1454 | α-Humulene | 0.72 ±0.007 | 0.45 ±0.021 | 0.92 ±0.007 | 0.85 ±0.021 | <0.1 | 0.16 ±0.007 | <0.1 | HS |
| V50 | 1458 | trans-β-Farnesene | 0.24 ±0.007 | 0.14 ±0.021 | 0.35 ±0.007 | 0.22 ±0.001 | 1.47 ±0.115 | - | - | HS |
| V51 | 1462 | β-Santalene | 3.77 ±0.014 | 2.74 ±0.021 | 2.05 ±0.007 | 1.24 ±0.014 | 12.02 ±0.64 | 0.78 ±0.049 | 0.45 ±0.021 | HS |
| V52 | 1470 | Naphthalene, 1,2,3,4,6,7,8,8a-octahydro-1,8a-dimethyl-7-(1-methylethenyl)-, (1R,7S,8aS)- | 0.58 ±0.007 | 0.9 ±0.001 | 2.89 ±0.014 | 3.73 ±0.085 | <0.1 | <0.1 | <0.1 | HS |
| V53 | 1477 | γ-Muurolene | 1.45 ±0.014 | 0.71 ±0.007 | 2.19 ±0.007 | 1.75 ±0.071 | 0.65 ±0.092 | 0.25 ±0.007 | <0.1 | HS |
| V54 | 1480 | α-Amorphene | 0.31 ±0.021 | <0.1 | 0.54 ±0.007 | 0.36 ±0.007 | 0.14 ±0.158 | <0.1 | 0.07 ±0 | HS |
| V55 | 1485 | β-Selinene | 2.85 ±0.021 | 3.14 ±0.007 | 1.49 ±0.021 | 1.34 ±0.021 | 0.84 ±0.241 | 0.06 ±0.014 | 0.05 ±0 | HS |
| V56 | 1488 | Eremophilene | 0.3 ±0.007 | 0.27 ±0.007 | <0.1 | <0.1 | 0.66 ±0.18 | - | - | HS |
| V57 | 1491 | β-cadinene | 0.45 ±0.014 | 0.27 ±0.001 | 0.44 ±0.007 | <0.1 | 0.42 ±0.132 | 0.2 ±0.014 | 0.16 ±0.014 | HS |
| V58 | 1494 | Viridiflorene | 1.09 ±0.007 | 0.73 ±0.014 | 1.3 ±0.014 | 0.67 ±0.014 | 1 ±0.202 | 0.19 ±0.035 | 0.17 ±0.007 | HS |
| V59 | 1498 | NI | 0.64 ±0.021 | 0.58 ±0.028 | 0.9 ±0.021 | 0.74 ±0.007 | - | - | - | |
| V60 | 1498 | α-Selinene | - | - | - | - | 0.38 ±0.065 | - | - | HS |
| V61 | 1499 | α-Muurolene | 0.51 ±0.007 | 0.23 ±0.001 | 0.78 ±0.049 | 0.56 ±0.014 | <0.1 | 0.17 ±0.007 | 0.16 ±0.007 | HS |
| V62 | 1505 | α-Bulnesene | <0.1 | <0.1 | <0.1 | <0.1 | 0.39 ±0.056 | - | - | HS |
| V63 | 1509 | β-Bisabolene | 0.43 ±0.014 | 0.26 ±0.021 | 0.66 ±0.007 | 0.26 ±0.001 | 2.12 ±0.221 | <0.1 | 0.01 ±0 | HS |
| V64 | 1514 | γ-Cadinene | 1.28 ±0.028 | 0.88 ±0.014 | 3.1 ±0.014 | 2.93 ±0.007 | <0.1 | 0.34 ±0.007 | 0.21 ±0.007 | HS |
| V65 | 1517 | NI | 0.63 ±0.007 | 0.93 ±0.014 | 0.32 ±0.007 | 0.51 ±0.014 | <0.1 | - | - | |
| V66 | 1521 | NI | 2.61 ±0.007 | 3.02 ±0.035 | - | - | - | - | - | |
| V67 | 1525 | δ-Cadinene | 3.37 ±0.049 | 1.93 ±0.021 | 5.14 ±0.014 | 3.62 ±0.028 | 0.52 ±0.119 | 7.41 ±0.332 | 6.29 ±0.304 | HS |
| V68 | 1528 | Myristicin | - | - | - | - | - | 7.37 ±0.205 | 9.6 ±0.219 | Ph |
| V69 | 1534 | trans-γ-Bisabolene | 0.98 ±0.014 | 0.37 ±0.007 | 2.61 ±0.028 | 1.21 ±0.021 | 1.97 ±0.234 | 0.43 ±0.028 | 0.21 ±0.014 | HS |
| V70 | 1540 | α-Cadinene | 0.27 ±0.021 | 0.22 ±0.014 | 0.45 ±0.021 | 0.38 ±0.021 | <0.1 | - | - | HS |
| V71 | 1543 | NI | 1.26 ±0.007 | 1.46 ±0.113 | 2.06 ±0.021 | 2.14 ±0.007 | <0.1 | 0.32 ±0.064 | 0.19 ±0 | |
| V72 | 1545 | α-Calacorene | 0.49 ±0.028 | 0.75 ±0.057 | 0.79 ±0.007 | 0.87 ±0.078 | - | 0.3 ±0.042 | 0.16 ±0.007 | HS |
| V73 | 1545 | trans-α-bisabolene | - | - | - | - | 0.42 ±0.113 | - | - | HS |
| V74 | 1547 | NI | - | - | 2.99 ±0.014 | 3.08 ±0.035 | - | - | - | |
| V75 | 1551 | α-Elemol | <0.1 | <0.1 | <0.1 | <0.1 | 0.54 ±0.318 | 0.7 ±0.042 | 0.79 ±0.028 | OS |
| V76 | 1558 | NI | 3.35 ±0.007 | 3.36 ±0.071 | 3.32 ±0.028 | 3.45 ±0.007 | - | 0.5 ±0.028 | 0.54 ±0.014 | |
| V77 | 1564 | Elemicin | - | - | - | - | - | 11.89 ±0.17 | 8.82 ±0.014 | Ph |
| V78 | 1565 | NI | 1.53 ±0.028 | 1.47 ±0.057 | - | - | - | - | - | |
| V79 | 1567 | NI | 0.83 ±0.014 | 0.8 ±0.007 | - | - | <0.1 | - | - | |
| V80 | 1579 | Spathulenol | 0.19 ±0.014 | 0.3 ±0.028 | 0.83 ±0.014 | 0.66 ±0.014 | <0.1 | <0.1 | <0.1 | OS |
| V81 | 1584 | Caryophyllene oxide | 1.39 ±0.205 | 2.92 ±0.113 | 2.88 ±0.007 | 3.96 ±0.389 | - | <0.1 | 0.22 ±0.007 | OS |
| V82 | 1589 | NI (m/z = 234) | 5.38 ±0.191 | 5.87 ±0.134 | 10.99 ±0.057 | 9.84 ±0.134 | - | 0.47 ±0.042 | 0.58 ±0.007 | |
| V83 | 1603 | Rosifoliol | 0.26 ±0.021 | 0.3 ±0.007 | 1.41 ±0.071 | 1.11 ±0.049 | - | 0.87 ±0.049 | 0.91 ±0.049 | OS |
| V84 | 1605 | NI | - | - | 1.04 ±0.007 | 0.83 ±0.099 | - | 0.45 ±0.035 | 0.24 ±0.021 | |
| V85 | 1605 | NI | 0.85 ±0.021 | 0.86 ±0.035 | - | - | - | - | - | |
| V86 | 1608 | NI | - | - | 0.54 ±0.014 | 0.74 ±0.078 | - | - | - | |
| V87 | 1611 | Humulene epoxide II | <0.1 | <0.1 | 0.46 ±0.021 | 0.79 ±0.035 | - | 0.29 ±0.007 | 0.2 ±0.028 | OS |
| V88 | 1614 | Tetradecanal | - | - | - | - | 0.21 ±0.065 | - | - | |
| V89 | 1614 | Viridiflorol | 0.53 ±0.028 | 0.49 ±0.035 | 0.47 ±0.002 | 0.36 ±0.028 | - | - | - | OS |
| V90 | 1618 | 1,10-di-epi-Cubenol | - | - | 1.29 ±0.156 | 1.19 ±0.064 | - | 0.81 ±0.042 | 0.73 ±0.134 | OS |
| V91 | 1620 | NI | - | - | 1.84 ±0.17 | 2.06 ±0.042 | - | - | - | |
| V92 | 1622 | epi-γ-Eudesmol | - | - | 0.77 ±0.014 | 0.68 ±0.035 | - | - | - | OS |
| V93 | 1623 | NI (m/z=236) | <0.1 | <0.1 | 1.04 ±0.014 | 0.96 ±0.021 | - | - | - | |
| V94 | 1630 | 1-epi-Cubenol | <0.1 | <0.1 | 0.52 ±0.007 | 0.39 ±0.007 | 0.2 ±0.082 | 0.84 ±0.156 | 0.48 ±0.021 | OS |
| V95 | 1634 | γ-Eudesmol | - | - | - | - | 0.5 ±0.258 | 0.23 ±0.064 | 0.1 ±0.014 | OS |
| V96 | 1638 | Hinesol | - | - | 0.65 ±0.021 | 1.17 ±0.035 | - | - | - | OS |
| V97 | 1643 | 1,2,3,4,4a,7,8,8a-octahydro-1,6-dimethyl-4-(1-methylethyl)-1-Naphthalenol, | - | - | 0.33 ±0.007 | 0.18 ±0.014 | - | <0.1 | <0.1 | OS |
| V98 | 1645 | epi-α-Cadinol | - | - | - | - | - | 1.19 ±0.064 | 0.61 ±0.014 | OS |
| V99 | 1645 | NI | - | - | 0.59 ±0.028 | 0.7 ±0.028 | - | - | - | |
| V100 | 1649 | α-Muurolol (=Torreyol) | <0.1 | <0.1 | <0.1 | <0.1 | - | 0.33 ±0.057 | 0.17 ±0.014 | OS |
| V101 | 1651 | NI | - | - | 3.01 ±0.042 | 2.87 ±0.001 | - | - | - | |
| V102 | 1652 | β-Eudesmol | - | - | 2.43 ±0.057 | 2.62 ±0.092 | 0.62 ±0.398 | 2.27 ±0.12 | 2.52 ±0.028 | OS |
| V103 | 1655 | α-Eudesmol | - | - | 0.5 ±0.071 | 0.73 ±0.021 | 0.62 ±0.419 | <0.1 | <0.1 | OS |
| V104 | 1657 | α-Cadinol | 2.74 ±0.028 | 2.9 ±0.092 | 2.07 ±0.049 | 1.8 ±0.028 | <0.1 | 1.53 ±0.085 | 0.73 ±0.028 | OS |
| V105 | 1660 | NI | <0.1 | <0.1 | 1.28 ±0.028 | 1.58 ±0.021 | - | <0.1 | <0.1 | |
| V106 | 1664 | NI | <0.1 | <0.1 | 0.4 ±0.057 | 0.56 ±0.035 | - | - | - | |
| V107 | 1671 | NI | - | - | 0.49 ±0.014 | 0.27 ±0.049 | - | - | - | |
| V108 | 1671 | β-Bisabolol | 0.46 ±0.042 | 0.51 ±0.014 | - | - | 0.47 ±0.242 | <0.1 | 0.07 ±0.007 | OS |
| V109 | 1676 | Cadalene | 0.19 ±0.078 | 0.13 ±0.028 | 0.91 ±0.014 | 0.7 ±0.007 | - | 0.17 ±0.035 | 0.08 ±0.007 | HS |
| V110 | 1678 | cis-α-Santalol | - | - | - | - | 0.78 ±0.161 | - | - | OS |
| V111 | 1678 | Bulnesol | - | - | - | - | - | 1.69 ±0.247 | 1.47 ±0.099 | OS |
| V112 | 1680 | NI | 1.04 ±0.099 | 1.41 ±0.014 | 2.08 ±0.007 | 1.64 ±0.035 | - | - | - | |
| V113 | 1680 | NI | - | - | - | - | 0.6 ±0.171 | - | - | |
| V114 | 1680 | NI | - | - | - | - | - | 0.65 ±0.085 | 0.6 ±0.163 | |
| V115 | 1687 | α-Bisabolol | - | - | 0.61 ±0.014 | 0.55 ±0.014 | <0.1 | - | - | OS |
| V116 | 1708 | NI | - | - | 0.76 ±0.001 | 0.87 ±0.014 | - | - | - | |
| V117 | 1710 | NI | 0.85 ±0.014 | 1.12 ±0.057 | - | - | - | - | - | |
| V118 | 1729 | NI | - | - | 0.62 ±0.014 | 0.62 ±0.014 | - | - | - | |
| V119 | 1745 | NI | <0.1 | <0.1 | 0.86 ±0.021 | 0.76 ±0.021 | - | - | - | |
| V120 | 1751 | NI | 0.48 ±0.007 | 0.67 ±0.021 | 0.52 ±0.014 | 0.69 ±0.007 | - | - | - | |
| V121 | 1764 | Guaiol acetate | - | - | - | - | - | 0.28 ±0.057 | 0.47 ±0.028 | OS |
| V122 | 1806 | NI | - | - | - | - | - | 1.2 ±0.071 | 1.8 ±0.134 | |
| Hydrocarbon monoterpene | 7.6 | 8.3 | 0.6 | 4.1 | 28.2 | 0.6 | 0.5 | HM | ||
| Hydrocarbon sesquiterpene | 39.5 | 29.9 | 39.9 | 31.7 | 65.1 | 15.6 | 12.4 | HS | ||
| Ketone | 5.0 | 5.1 | 1.9 | 2.6 | 0.0 | 0.0 | 0.0 | Ke | ||
| Oxygenated monoterpene | 12.6 | 15.3 | 2.9 | 5.4 | 1.7 | 11.3 | 10.9 | OM | ||
| Oxygenated sesquiterpene | 5.1 | 6.9 | 14.9 | 15.8 | 3.8 | 11.8 | 9.7 | OS | ||
| Phenylpropanoid | - | - | - | - | - | 56.7 | 62.0 | Ph | ||
| Not identified (unknown) | 26.7 | 29.4 | 36.4 | 35.6 | 0.9 | 3.7 | 4.0 | NI | ||
| Total (%) | 96.08 | 94.45 | 96.28 | 94.90 | 99.60 | 99.75 | 99.55 | |||
| Yield (v/w %) | 0.91 | 0.91 | 0.93 | 1.2 | 3.6 | 0.6 | 0.5 | |||
*IR -retention index calculated; NI -not identified (unknown); **: Chemical Class; Le-F-1—leaves from the flooding season of chemotype-1; Le-D-1—leaves from the dry season of chemotype-1; Le-F-2—leaves from the flooding season of chemotype-2; Le-D-2—leaves from the dry season of chemotype-2; Ba-D-1 –bark from the dry season of chemotype-1; Ba-F-1—bark from the flooding season of chemotype-1; Fr-1—fruits from the flooding season of chemotype-1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



