
Submitted:
20 May 2025
Posted:
22 May 2025
You are already at the latest version
Abstract
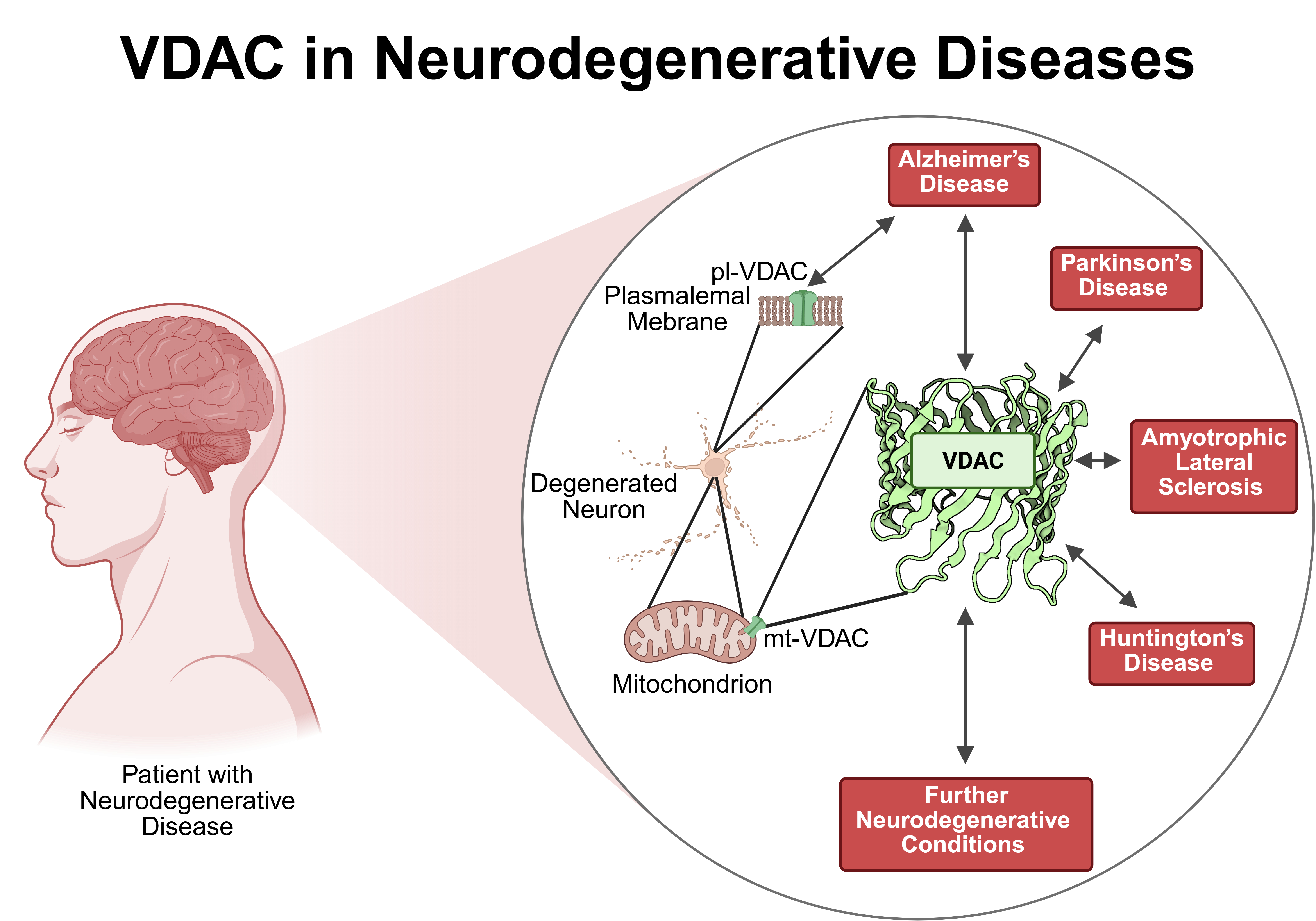
The family of voltage-dependent anion channels (VDAC) comprises three isoforms (VDAC-1, VDAC-2, VDAC-3). VDAC has been extensively described as localized in the outer mitochondrial membrane where it is involved in the exchange of ions, metabolites and ATP/ADP between mitochondria and cytosol. VDAC interacts with disease specific proteins and thus regulates the mitochondrial function and controls the cellular energy resources explaining its involvement in cell death and apoptosis. In addition, VDAC-1 and -2 can also be found at other cellular locations such as in the sarcoplasmic reticulum, in the endoplasmic reticulum as well as in the plasma membrane. Through single channel pore regulation, oligomerization or changed expression levels VDAC is involved in different neurodegenerative diseases such as Alzheimer’s disease, Parkinson’s disease, Amyotrophic lateral sclerosis, Huntington’s disease and others. Here, we critically summarize current discussions about VDAC as a common key player for these diseases. We suggest that VDAC acts as a transmembrane multifunctional regulatory protein which might serve as a pharmacological target for the development of novel drugs against neurodegenerative diseases such application of recombinant antibody technology.
Keywords:
voltage-dependent anion channels
; Alzheimer's disease
; Parkinson’s disease
; amyotrophic lateral sclerosis
; Huntington’s disease
1. Voltage-Dependent Anion Channel Function and Structure
In the year 1976, the voltage-dependent anion channel (VDAC) was discovered in the mitochondria of the unicellular organism Paramecium aurelia [1]. Later in 1979, VDAC was found in mitochondria derived from rat liver [2]. In between, it was found that VDAC – also named as porin - is expressed in all eukaryotes [3] and comprises a superfamily of the three isoforms VDAC-1, VDAC-2 and VDAC-3 [4]. While VDAC-1 and VDAC-3 have 9 exons, VDAC-2 has 10 exons in mammals [5,6]. Furthermore, there are two splice variants for VDAC-1 while there is only one splice variant for VDAC-2 and VDAC-3 [3]. The cDNA sequences of both VDAC-1 and VDAC-2 have a homology of 90% whereas the one for VDAC-3 is 68% between human and mice. All VDAC genes are encoded in the nucleus, the protein is synthesized at ribosomes in the cytosol and finally imported into the outer mitochondrial membrane (OMM) [7,8,9,10]. The three VDAC isoforms differ in their expression level: VDAC-1 is the most widely expressed form in mammals, followed by VDAC-2 and the lowest expression level was found for VDAC-3 [3]. In addition, the three isoforms vary in their tissue distribution: VDAC-1 and VDAC-2 are expressed in different tissues like brain, heart, liver and skeletal muscles [11,12] and VDAC-3 shows expression in liver, lung, spleen, ovary adrenal gland and in testes [13]. The molecular weight of VDAC corresponds to ~ 30 kDa [14]. In the year 2008, the structures of human and murine VDAC-1 were solved showing that VDAC-1 is a β-barrel build of 19 anti-parallel β-sheets sloped in a 46° angle and an α-helix at the N-terminal [5,15,16,17]. Regarding VDAC-2, its structure was determined from zebrafish [18] and there is no three-dimensional structure of VDAC-3 so far. The pore-forming channel has a diameter of 2.5 nm in its open state [5]. As the VDAC is located in the OMM, it exchanges ions, metabolites as well as adenosine triphosphate (ATP) and adenosine diphosphate (ADP) between the cytosol and mitochondria [19]. The inner mitochondrial membrane (IMM) contains many carrier proteins and transporters for the specific transport across the IMM [20]. The VDAC shows a certain ion selectivity such that sodium and potassium ions can pass through the channel [21]. Furthermore, dimers of human VDAC-1 and VDAC-2 function as scramblase-type lipid transporter in the OMM for the transport of phospholipids into the inter-membrane space of mitochondria [22]. The transmembrane potential regulates the VDAC activity which is the reason why these channels are named voltage-dependent. When VDAC-1 was inserted into bilayer lipid membrane in in vitro experiments, the channel had a high conductance associated with an open state enabling free transport of monovalent ions and anionic metabolites at a low membrane potential (± 20 mV). When the transmembrane potential was increased or decreased in the range of ±20 to ±40 mV, the VDAC permeability changed so that the Ca2+ transport increased while the permeability for monovalent ions, metabolites and nucleotides was inhibited [11,23,24]. Furthermore, while in vitro experiments using artificial membranes showed high conductance for VDAC-1 and VDAC-2, the human VDAC-3 showed low conductance, and the membrane potential did not influence its activity [25,26]. The N-terminus of VDAC functions as voltage sensor and its movement leads to conformational changes of the barrel influencing the pore diameter. The channel selectivity is changed by a decreased pore diameter [26,27]. It is the interaction of VDAC with cytoplasmic proteins such as hexokinase (HK), tubulin or α-synuclein (αSyn) preferentially that affects the change from the open to the closed pore rather than changes of the membrane potential in a living cell [20,23,28].
1.1. Extra-Mitochondrial Locations of VDAC
Besides the expression of VDAC in the OMM, VDAC is detected in various other subcellular locations such as the sarcoplasmic reticulum, endoplasmic reticulum [29] and the plasma membrane [30,31,32]. In mice, the first exon of the VDAC-1 pre-mRNA is alternatively spliced resulting in two different mRNAs splice variants, one coding for the VDAC-1 located in the mitochondria (mt-VDAC-1) and one coding for the VDAC-1 found in the plasma membrane (pl-VDAC-1). As a result of alternative splicing, the pl-VDAC-1 mRNA codes for a short hydrophobic signal peptide of 13 amino acids which is required to guide the pl-VDAC-1 via the secretory pathway to the plasma membrane. This signal peptide is cleaved off before the pl-VDAC-1 is inserted into the plasma membrane. Finally, mature mt-VDAC-1 and pl-VDAC-1 have the identical amino acid sequences [33]. Although, human cells also contain the pl-VDAC-1 in their plasma membrane, this is not the result of alternative splicing, but an alternative mechanism of dynamic membrane protein redistribution was proposed [32]. Alternatively, pl-VDAC-1 was found to function as the receptor for human plasminogen kringle 5 (K5) in the cell membrane of human umbilical vein endothelial cells (HUVEC) and K5 may induce the intracellular translocation of VDAC-1 to the plasma membrane [34]. The amino acid residues of mt-VDAC-1 exposed to the cytosol, are found on the extracellular site in case of the pl-VDAC-1 [35,36].
The physiological function of the pl-VDAC-1 has not completely uncovered so far; however, pl-VDAC-1 behaves as a NADH-ferricyanide reductase involved in the normal cellular redox homeostasis [37,38]. Furthermore, pl-VDAC-1 participates in cell volume regulation and in ATP release as shown in murine cells [39]. Importantly, the pl-VDAC-1 participates in apoptosis as described in detail below in 1.3. Interestingly, disease condition can lead to a mistargeting of VDAC-1 to un-conventional cellular locations. High concentration of glucose (glucotoxicity) induces VDAC-1 overexpression in insulin-secreting β-cells in type 2 diabetes (T2D) patients. Consequently, the VDAC-1 becomes mistargeted into the plasma membrane and there is a loss of ATP for β-cells. The antidiabetic drug metformin prevents glucotoxicity-induced VDAC-1 overexpression showing that VDAC-1 serves as a target for disease treatment [40]. Other VDAC isoforms, i.e. VDAC-2 and VDAC-3 show extra-mitochondrial expression as these isoforms were found in the outer dense fibres which is a cytoskeletal part of the sperm flagellum [41]. Furthermore, the plasma membrane or the acrosomal membrane of bovine [42], mouse [43] and human [44] spermatozoa contain VDAC-2 and at least for human spermatozoa it was shown that pl-VDAC-2 is involved in sperm function by enabling Ca2+ transmembrane transport [44]. Other examples of extramitochondrial regulation are the upregulations of VDAC-1 and VDAC-2 in the plasma membrane of human pancreatic cancer cells [45] and of pl-VDAC-2 upon iron deprivation in erythroleukemia K562 cells [46]. The increased expression of pl-VDAC-2 might be explained by a compensatory mechanism because iron deprivation induces hypoxia which in turn leads to peroxidation of membrane lipids [47]. VDAC-mediated transport becomes activated by membrane lipid peroxidation. In addition, the activity of enzymes controlling the redox status can be activated by VDAC-2 [46,48].
1.2. VDAC and Mitochondrial Dynamics
Mitochondria are considered as dynamic organelles because of their proliferation by fusion and fission, transport processes and selective degradation [49,50]. VDAC-mediated mitochondrial dynamics result from the interaction with proteins required for fusion like mitfusins and those needed for fission such as dynamin-related proteins (Drp-1) [51]. Furthermore, mitochondrial fusion and fission become affected by altered VDAC expression or activity. Whereas high activity of VDAC leads to mitochondrial depolarization and fragmentation, low VDAC activity may promote fusion and maintenance of a healthy mitochondrial network [52]. The selective degradation of damaged or dysfunctional mitochondria – called mitophagy – is regulated by involving VDAC due to its interactions with proteins such as phosphatase and tensin homologue (PTEN)-induced putative kinase protein 1 (PINK1) and parkin. Furthermore, mitophagy can be impaired by a dysregulated VDAC activity resulting in accumulation of dysfunctional mitochondria [53]. Additionally, VDAC contributes to cellular signalling pathways targeting kinases, phosphatases as well as cytoskeletal components influencing mitochondrial dynamics [54,55]. Consequently, VDAC dysregulation effects signalling leading to disturbed mitochondrial dynamics [56,57]. Overall, mitochondrial dysfunction in several diseases might be better understood by investigating the influence of VDAC on mitochondrial dynamics.
1.3. The Involvement of VDAC in Apoptosis
The development of neurodegenerative diseases is in many cases associated with mitochondrial dysfunction along with caspase-mediated apoptosis [58,59]. Due to this and to point out the role of VDAC regarding apoptosis, the molecular steps of apoptosis are shortly summarised here. The programmed cell death called apoptosis can be divided into the extrinsic pathway and the intrinsic mitochondrial-mediated pathway. External ligands binding to cell surface death receptors like death receptor 4 and 5 (DR4, DR5), tumour necrosis factor (TNFα), TNF-related apoptosis-inducing ligand (TRAIL) receptors, tumour necrosis factor receptor 1 and 2 (TNFR1, TNFR2) and Fas/CD95 activate these receptors. Upon activation, the death signalling complex (DISC) is formed which activates pro-caspase-8 that in turn activates caspases-3 and -7. Finally, this executes the cell death [60,61].
Intracellular signals such as elevated levels of reactive oxygen species (ROS), DNA damage, Ca2+ overload, and chemotherapy drugs result in the activation of the intrinsic mitochondrial-dependent apoptosis. These apoptotic signals change the permeability of the OMM releasing the apoptotic factors cytochrome c (Cyto c) and apoptosis-inducing factor (AIF) into the cytosol. Second mitochondria-derived activator of caspase (SMAC) or the human ortholog direct IAP-binding protein with a low pI (Diablo), endonuclease G (EndoG) and high-temperature requirement protein A2 [62,63]. Cell death proteases (caspases) are activated by these released factors. Cyto c interacts with apoptotic protease activating factor 1 (Apaf-1) forming an oligomeric structure in presence of dATP. The apoptosome is built upon binding of this oligomeric structure with cytoplasmic inactive pro-caspase-9 leading. Thereby pro-caspase-9 becomes activated to caspase-9 that in turn cleaves pro-caspase-3 and -7 obtaining activated effector caspase-3 and -7. The cell is destroyed from within by these activated effector caspases by cleaving cellular substrates and thereby forming apoptotic bodies [62]. Calpains or cathepsins cleave AIF which translocases to the nucleus for activating chromatin degradation and condensation. Furthermore, chromatin DNA is cleaved into nucleosomal fragments by EndoG which was transferred into the nucleus as well. From mitochondria into the cytosol released SMAC antagonises caspase inhibition by interacting with inhibitor of apoptosis proteins (IAPs) [64]. In addition, caspase-8 mediates crosstalk between the extrinsic apoptotic pathway and the intrinsic mitochondrial-mediated apoptotic pathway by cleaving Bid resulting in the truncated protein tBid which is transferred to the mitochondria for activating the intrinsic apoptosis [65]. The apoptogenic proteins cross the OMM via several possible mechanisms. One possibility is the rupture of the OMM enabling non-specific release of apoptotic initiators out of the mitochondrial intermembrane space [66]. Furthermore, the permeability transition pore (PTP) opens upon Ca2+ overload or after overproduction of ROS [67]. Proteins of the B cell lymphoma 2 (Bcl-2) family can be differentiated into anti-apoptotic proteins such as Bcl-2 and Bcl-xL and pro-apoptotic proteins like Bak, Bax, Bid and Bim [68]. An additional mechanism describes how a large channels are built up by Bax and/or Bak oligomers [69,70], hetero-oligomers of Bax and VDAC-1 [71,72] or oligomers of VDAC-1 [73,74,75,76,77,78,79,80,81,82,83,84]. Additionally, apoptosis can be regulated by the interaction of VDAC-1 with anti-apoptotic proteins providing a further level of control over apoptosis [78,85,86]. The interaction of Bcl-2 and Bcl-xL with VDAC-1 mediates protection from apoptosis [87,88]. In detail, the BH4 domain of Bcl-2/Bcl-xL is sufficient and essential for interacting with VDAC-1 and thereby inhibiting apoptosis [89]. VDAC-1 over-expression induces apoptosis in all investigated cell types ranging from human, animal, fish and plant cells [80,90,91,92,93,94]. Consequently, VDAC-1 inhibitors such as 4-acetamido-4-isothiocyanato-stilbene-2,2-disulfonic acid (SITS), 4,4 diisothiocyanostilbene-2,2-disulfonic acid (DIDS), 4,4’ diisothiocyanatodihy-drostilbene-2,2‘-disulfonic acid (H2DIDS) or RuR prevent apoptosis by their direct interaction with VDAC-1 [81,91]. In line with this, apoptosis can also be inhibited by the over-expression of Bcl-2 as well as HKI as these are anti-apoptotic proteins [90,95]. Accordingly, using non-small cell lung cancer cells, cisplatin-induced apoptosis was prevented due to siRNA-mediated reduction in levels of VDAC-1 expression [96].
Besides to the involvement of mitochondrial VDAC in apoptosis, the plasmalemmal VDAC is also involved in apoptosis. The pl-VDAC-1 contributes to apoptosis as demonstrated in neuronal cells such as the human neuroblastoma cell line SK-N-MC, in the mouse hippocampal cell line HT22, and in primary differentiated hippocampal neurons which were stimulated with the protein kinase inhibitor staurosporine. These conditions lead to the activation and opening of pl-VDAC-1 preceding the activation of caspases. The extra-cellular application of anti-VDAC antibodies inhibited apoptosis in these experiments demonstrating the involvement of pl-VDAC-1 [97,98]. In line with this, extracellular applied anti-VDAC antibodies mediated protection from amyloid β peptide-induced apoptosis in HT22 cells and in the mouse cholinergic septal neuronal cell line SN56 [99] and from 15-deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2) induced apoptosis in rat primary cortical neurons [100]. Furthermore, a study using prostate cancer cell line LNCaP led to the suggestion that pl-VDAC-1 might be a positive stimulator of the extrinsic apoptotic pathway [101,102].
1.4. Involvement of pl-VDAC-1 in Neuroprotection
For investigating the role of the small GTPase Ras in a neuronal context, a genetically modified mouse was created, in which human constitutive activated V12-Ha-RAS is expressed in post-mitotic neurons thereby over-activating the endogenous downstream mitogen-activated protein kinase (MAPK) [103]. Several studies confirmed that neurons such as facial motoneurons, the substantia nigra, cortex and hippocampus were protected from chemical or mechanical insults [103,104,105,106,107,108]. However, the detailed molecular mechanism explaining the neuroprotection remained elusive. A proteome study showed that in mice expressing constitutively activated Ras in neurons (=transgenic activation of neuronal Ras) several proteins were changed in their expression level, i.e. the mitochondrial respiratory chain, synaptic protein recycling and vesicle mediated transport were upregulated while microtubulin remodeling proteins and VDAC-1 were downregulated.
Focusing on VDAC-1 demonstrated a selective decrease in the expression level of pl-VDAC-1 while mt-VDAC-1 was unchanged in cortex and hippocampus of this mouse model. Furthermore, the selective decrease of pl-VDAC-1was confirmed in primary cortical cultures derived from this mouse model. The selective inhibition of the MAPK signaling by the inhibitor U0126 led to an attenuated pl-VDAC-1 level in transgenic neurons like in wild type derived neurons demonstrating that activated V12-Ha-RAS/MAPK signaling influences the alternative splicing of VDAC-1 mRNA. Correspondingly, the extracellular application of anti-VDAC antibodies prior to an excitotoxic glutamate stimulation protected wild type primary cortical cultures to the same extent as transgenic V12-Ha-RAS in primary cortical cultures (Figure 1). Taken together, the selective reduction of expression of pl-VDAC-1 is involved in the V12-Ha-RAS mediated neuroprotection observed in this mouse model [109].
2. Alzheimer’s Disease and VDAC
Typical hallmarks of Alzheimer’s disease (AD) are a decline in cognitive function and a progressive memory loss. On the molecular level, AD is characterized by amyloid plaques and neurofibrillary tangles which consist of abnormal hyperphosphorylated tau protein. Tau is required in microtubule stabilization and furthermore its association with synaptic loss and cognitive impairments are seen in AP patients. Although the biological mechanisms resulting in sporadic forms of AD are not fully understood so far, it is proposed that cholinergic dysfunction, mitochondrial dysfunction, inflammation, tau accumulation, amyloid β (Aβ) plaque formation, inflammatory response, lysosomal dysfunction and hormone regulation are involved [110].
In early stages of AD pathogenesis, mitochondrial dysfunction is observed with several pathophysiological events such as disruption of Ca2+ homeostasis, reduced metabolism, lipid peroxidation, an increase in ROS production and finally apoptosis. Interestingly, this impaired brain metabolism develops several decades before dementia becomes obvious [111,112,113,114,115]. The accumulation of Aβ and dysfunctional mitochondria can result in oxidative stress and increased production of ROS [116]. High levels of ROS affect mitochondrial components like membrane lipids, mitochondrial DNA (mtDNA) and components of oxidative phosphorylation [117,118]. Furthermore, ROS can oxidize VDAC making it dysfunctional [119,120,121].
AD post-mortem patient brains showed high levels of VDAC-1 expression in neurites of Aβ deposits which was also observed in amyloid precursor protein (APP) transgenic mice [122,123,124]. Neurons of AD brains show characteristics of apoptosis and the massive loss of neurons in AD is caused by apoptosis [83,125,126]. As mentioned above (see 1.3), VDAC-1 overexpression triggers apoptosis and this might be one of the reasons for the neuronal cell death in AD.
Experiments in the HT22 cell line and in the mouse cholinergic septal neuronal cell line SN56 showed that the application of anti-VDAC antibodies protected from neurotoxicity demonstrating the involvement of plasmalemmal VDAC in AD. Furthermore, pl-VDAC-1 interacts with membrane-related isoform of estrogen receptor α (mERα) forming a complex in caveolae [99]. The pl-VDAC along with mERα builds a complex with scaffolding protein caveolin-1 in caveolae of human hippocampus and cortex. AD brains show an accumulation of VDAC in caveolae of dystrophic neurites of senile plaques [124]. Both, the mitochondrial and the plasmalemmal VDAC-1 are involved in Aβ mediated neurotoxicity according to the following model: Extra-cellular Aβ oligomers interacts with the N-terminus of the pl-VDAC-1. The interaction of VDAC-1 with Aβ might involve positive charges of VDAC-1 in its N-terminal domain, negative charges in Aβ, hydrophobic interaction, electrostatic interaction and interactions via the GXXXG motif. VDAC-1 provides one GXXXG motif in its α-helix at the N-terminus [127] and Aβ contains three GXXXG motifs [128]. There is the hypothesis that the GXXXG motif of Aβ interacts with the GXXXG motif of VDAC-1 in AD [129,130]. The interaction of Aβ with pl-VDAC-1 results in VDAC-1 oligomerization and building a large pore consisting of Aβ/VDAC-1 heteromers. Aβ enter the cell through this large pore. Intracellular, Aβ interacts with the mt-VDAC-1 resulting in detachment of hexokinase 1 (HKI) and induction of VDAC-1 oligomerization along with Aβ forming large heteromeric Aβ/VDAC-1 pores. These pores enable cytochrome c release into the cytoplasm and thereby induce apoptosis (Figure 2) [83]. Consequently, a reduced VDAC-1 expression as shown in a mouse model, protects from degenerative changes [131]. Additionally, siRNA mediated silencing of VDAC-1 expression inhibited Aβ entry into the cytosol and protected from Aβ-induced toxicity [83]. Furthermore, as the newly developed small molecule VBIT-4 inhibits VDAC-1 [81], VBIT-4 prevented Aβ-induced VDAC-1 overexpression and apoptotic cell death in neuronal cultures. In addition, using an AD mouse model with VDAC-1 overexpression in neurons surrounded by Aβ plaques, application of VBIT-4 protected from pathophysiological changes such as neurometabolic dysfunction, neuroinflammation and neuronal cell death. Interestingly, behavioral assessments of this mouse model showed that VBIT-4 prevented cognitive decline. With respect to AD treatment, VDAC-1 is an interesting target and VBIT-4 might be an encouraging drug candidate [132]. However, a recent study showed that VBIT-4 might be toxic to healthy cells, at least demonstrated for breast adenocarcinoma (MCF-7) cells. High concentration of VBIT-4 induced the suppression of mitochondrial respiration, increased the H2O2 and ROS production and lead to cell death thus demanding for cautionary application in clinical settings [133].
There is another possible link between AD and pl-VDAC-1: AD patients show an increased formation of prostaglandin D2 (PGD2) [134] and its metabolite 15-deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2) exerts a high neurotoxicity [135]. The pl-VDAC-1 was found as a membrane target for 15d-PGJ2 [100]. Extracellular application of anti-VDAC-1 antibodies protects from 15d-PGJ2-mediated neurotoxicity in rat primary cortical neurons [100].
Recently, it was shown that dimers of VDAC-1 with VDAC-2 function as a phospholipid scramblase [22]. Apoptotic cells lose their lipid asymmetry because phosphatidylserine (PS) becomes externalized to the external leaflet [136]. This externalization of PS could be explained by the phospholipid scramblase activity of dimeric VDAC in the plasma membrane under apoptotic conditions as proposed by Rockenfeller [137]. In the context of AD, the author extended this hypothesis proposing that VDAC dimers could insert APP into the plasma membrane [137].
An additional link emerges from increased glycogen synthase kinase 3 (GSK3β) in AD resulting in a non-characteristic APP processing leading to an elevated Aβ production and hyperphosphorylated tau [138]. Furthermore, GSK3β phosphorylates mt-VDAC-1 causing the detachment of HK which alters the cellular metabolism (Figure 2) [139] by reducing the ATP supply for glycolysis and glucose metabolism, making the cells more prone for apoptosis. As mentioned above, HK detachment can also be induced by Aβ resulting in VDAC-1 oligomerization which enables the release of cytochrome c and subsequent activation of apoptosis [83].
In AD, an irregular hyperphosphorylation of tau protein was found which results in its aggregation and the formation of neurofibrillary tangles. [140]. Phosphorylated tau interacts with VDAC-1 as found in the brains of AD mice and AD patients [122,141]. The mitochondrial function is influenced by the interaction of phosphorylated tau with VDAC [142]. In AD, phosphorylated tau protein binds to VDAC and thereby closes the channel yielding to impaired transport of ions and metabolites across the mitochondrial membrane (Figure 2) [56,141]. This decreases the ATP production and causes oxidative stress contributing to neurodegeneration and cognitive decline [143,144]. Interestingly, a VDAC-1 heterozygous mouse model (VDAC-1 +/-) showed reduced mRNA level for AD-related genes such as Aβ, APP and tau demonstrating that a reduced VDAC-1 expression level protects from AD-mediated neurotoxicity [131].
Furthermore, in AD VDAC-1 interacts with the translocator protein (TSPO) which is located in the OMM. TSPO is an 18 kDa protein participating in several process such as mitochondrial metabolism, cholesterol import, cell proliferation, inflammation, oxidative stress and apoptosis [145]. It forms a complex with mt-VDAC-1 (Figure 2) which could probably occur via GXXXG motifs as the N-terminus of VDAC containing one GXXXG motif (see above) and TSPO providing three GXXXG motifs [146]. The complex of VDAC-1 with TSPO promotes the overproduction of ROS [146]. Interestingly, TSPO is overexpressed as well (as described for VDAC-1) in brains of AD patients [147]. In addition, overexpression of TSPO is seen in all AD models [148,149,150,151,152,153].
Post-transcriptional gene expression regarding various cellular processes including neuronal function can be regulated by small noncoding RNAs called microRNAs (miRs) [154,155]. The VDAC-1 expression level can be regulated by several miRNAs [156,157,158,159] while specifically miR-29a showed an association with AD [160]. The analysis of post-mortem brains from patients suffering from sporadic AD showed a loss of miR-29a [161]. In mouse, the knockdown of miR-29 in brain resulted in massive cell death in hippocampus and cerebellum which was reasoned by an increased VDAC-1 expression level. In miR-29 knockdown cells, apoptosis could be partly inhibited by down-regulation of VDAC-1 expression. The authors draw the conclusion that VDAC-1 expression levels can be explained by miR-29 which plays an important role regarding cellular survival of neurons in the brain [160]. However, a miR-29 knockdown may influence more targets than just VDAC-1 and miR-29 controls cell survival via VDAC-1 in astrocytes [162]. In conclusion, due to the influence of miRNAs on the expression level of VDAC-1, miRNAs seem to be a promising target for developing therapeutic approaches regarding different neurodegenerative diseases like AD.
Finally, there is a link between AD and T2D involving VDAC. Several correlations between AD and T2D have been described demonstrating that T2D is a risk factor for AD and vice versa [163,164,165]. Persons with T2D have approximately a 1.5-fold higher relative risk for AD according to a meta-analysis of longitudinal studies [166]. Furthermore, another study showed that the risk of dementia is doubled in T2D patients [167]. The antidiabetic drug metformin was proposed to have a neuroprotective potential because it reduces the risk of AD onset [167]. The neuroprotective potential of metformin can be mechanistically explained by its inhibition of hyperinsulinemia which in absence of inhibition participates in Aβ plaque formation and yields to onset of AD [168]. Furthermore, using a tau transgenic mouse model, tau phosphorylation was reduced by metformin as demonstrated in primary neurons [169]. The connection between AD and T2D is further strengthened by the analysis of post-mortem brains, showing a defective insulin signaling [170,171]. This link led to the suggestion that AD could be considered as “type 3 diabetes” [172,173]. Diabetes induces changes in insulin signaling, glucose metabolism, vascular function and modifying Aβ/tau metabolism resulting in neurodegeneration [174,175,176]. A common feature in AD and T2D is the overexpression of VDAC-1 as found in affected regions of brains from AD patients [122,123,177] and in β-cells of T2D [178,179]. This VDAC-1 overexpression mediates apoptosis in AD and T2D. Interestingly, a study used mice to investigate metformin mediated side effects showing increased VDAC-1 levels connected to the formation of dimers and trimers of VDAC-1 along with mitochondrial dysfunction in cortex and toxic amyloid pre-fibrillar aggregates were directly induced by metformin. In total, these effects increase the risk for the onset of AD [180].
Taken together, plasmalemmal and mitochondrial located VDACs are strongly involved in the pathology of AD because of increased expression levels, their potential to generate oligomers and due to their interactions with Aβ, GSK3β, phosphorylated tau and TSPO.
3. Parkinson’s Disease and VDAC
Coming after AD, the second most neurodegenerative disease is Parkinson’s disease (PD) [181]. PD is characterised by motor symptoms such as resting tremor, muscular rigidity, dystonia, postural instability and dyskinesias and non-motor symptoms as anxiety, depression, hallucination, hyposmia, fatigue, sleep disorders, cognitive impairment and diarrhoea [182,183,184,185]. PD arises from several factors like genetics, aging and environmental factors. The main pathogenic genes of PD comprise α-synuclein, leucine-rich repeat kinase 2 (LRRK2), phosphatase and tensin homologue (PTEN)-induced putative kinase 1, parkin RBR E3 ubiquitin protein ligase (PRKN), DJ-1 (Parkinsonism associated deglycase, PARK7), glucosylceramidase (GBA) and vacuolar protein sorting-35 (VPS35) [186]. Motor symptoms become obvious after a majority (about 70%) of dopaminergic (DA) neurons have died in the substantia nigra pars compacta (SNpc). Consequently, dopamine is depleted in the striatum and results in the impairment of the thalamo-corticobasal ganglia circuits [187,188,189]. Gold standard for the treatment of PD is the administration of the physiological precursor L-3,4-dihydroxyphenylalanine (L-DOPA) to substitute the striatal dopamine loss. Other pharmacological treatments use inhibitors of catechol-o-methyltransferase (COMT) and monoamine oxidase (MAO)-B, or amantadine. At later stage, another therapeutic option is the deep brain stimulation of the internal part of the globus pallidus (GPi) or of the subthalamic nucleus (STN). Unfortunately, none of the therapeutic options can neither cure nor stop the progression of PD. [190]. Furthermore, motor complications develop as side effect of long-term treatment with L-DOPA which are referred as L-DOPA induced dyskinesia. [191]. Additionally, the non-motor symptoms cannot be counteracted by L-DOPA.
Pathological hallmarks of PD are the loss of DA neurons and the occurrence of lewy bodies (LBs) which are cytoplasmatic neuronal inclusions consisting mainly of the protein αSyn [192]. Although, the accumulation of misfolded αSyn, and its neurotoxic effects and interference with mitochondrial functions have been described in detail in excellent reviews such as [193], we shortly summarize key aspects here.
αSyn misfolds by a not fully understood process into amyloid fibrils which accumulate intra-cellularly [194]. The small 14 kDa protein αSyn is ubiquitously expressed, especially at presynaptic terminals. The N-terminal of αSyn forms an α-helix structure enabling the interaction with lipid membranes [195]. The central domain of αSyn is built of a highly hydrophobic motif which is required for the aggregation of αSyn [196]. The C-terminus of αSyn consists of proline residues and negatively charged amino acids in a region enabling various post-translational modifications such as phosphorylation at serine 129 (S129) which was first identified in extracts from LBs of PD patients. [197]. The phosphorylation of αSyn at S129 influences the interaction between αSyn and lipid membrane and strengthen its binding to metal ions and further proteins resulting in protein aggregation [198]. Pathological misfolding of αSyn starts with the acquirement of a structure rich in β-sheets followed by self-assembly into intermediates of metastable oligomers which finally accumulate as fibrils within LBs [199]. In dependence to the disease stage, αSyn aggregates were found in different brain regions of PD patients leading to the proposal that αSyn might transmits from cell-to-cell via a mechanism like prions. [200,201]. The neurotoxicity of αSyn oligomers is mediated by modifying biological membranes in their stability and by interaction with mitochondrial proteins which become influenced in their function [202]. Consequently, toxic effects are triggered by αSyn oligomers in a cascade such as membrane permeabilization, mitochondrial dysfunction, increased Ca2+ influx, oxidative stress, loss of proteostasis, and finally cell death [203,204,205]. Mitochondrial dysfunction is accompanied by the transition of monomeric αSyn to pathological oligomers [193].
Monomeric αSyn directly interacts via its C-terminus with VDAC. There are two possibilities for the interaction of αSyn with VDAC which first regulate the permeability of OMM or secondly mediate cytotoxicity. In the first option, monomeric αSyn can bind into the pore of VDAC in the OMM leading to a sterically blocked channel and thereby inhibiting the flux of ATP and ADP. Consequently, the adenine-nucleotide translocator (ANT) of the IMM is disturbed by a substrate unbalance. This depletes the substrate of ATP synthase, the mitochondrial potential decreases resulting in an impairment of oxidative phosphorylation. The VDAC blockage by αSyn is reversible, concertation and voltage-dependent and might be part of a regulatory mechanism for mitochondrial respiration (Figure 3). In the second option, under stress conditions, monomeric αSyn is translocated through the pore of VDAC into the intermembrane space (IMS). There, it can directly interact with the complexes of the electron transport chain (ETC) embedded in the IMM [206]. αSyn directly targets complex I [205,207,208], complexes II and III [209] and complex IV [210]. The interaction of αSyn with the ETC results in mitochondrial dysfunction due to increased production of ROS (Figure 3). Consequently, monomeric αSyn is oxidized in the cytosol leading to oligomerisation of αSyn. Oligomeric αSyn associates with the OMM causing mitochondrial dysfunction and thereby finally mediating neurotoxicity (Figure 3) [206,211]. In addition, the composition of the OMM lipids may influence how αSyn bind to the OMM and thereby influences the complex formation of αSyn with VDAC as recently discussed in a review [212]. Furthermore, when αSyn binds to VDAC, the Ca2+ permeability of VDAC becomes modulated leading to an increased Ca2+ flux through VDAC (Figure 3) [213]. Comparing the binding affinities between αSyn and VDAC-1 or VDAC-3, respectively showed a 10-to 100-fold lower affinity to VDAC-3 than to VDAC-1 [214]. The adeno-viral mediated overexpression of αSyn in the substantia nigra of rat brains resulted in neuronal death of dopaminergic neurons due to an interaction of αSyn with mt-VDAC-1 and its subsequent activation of PTP [215]. In agreement with this, the analysis of a transgenic mouse model which expresses human αSyn A53T and shows a PD-like phenotype revealed that αSyn A53T interacts with neuronal mt-VDAC and with the PTP modulator cyclophilin D [216]. Remarkably, the accumulation of αSyn in nigral neurons of postmortem PD patients induced a decreased expression level of VDAC-1 compared to the age-matched control group. This could be confirmed in a rat model by the viral mediated expression of human mutant αSyn A30P leading to a decreased VDAC-1 level in striatal fibers and nigral neurons [217]. The outcome of an altered dopamine homeostasis was investigated using the human neuroblastoma cell line SH-SY5Y and showed reduced VDAC-1 and VDAC-2 protein levels while their mRNA levels remained unchanged suggesting a dopamine-induced increased protein degradation mediated by mitochondrial proteases. The authors also found a reduced VDAC-3 level but they focused on VDAC-1 and VDAC-2 arguing that in general VDAC-3 shows a low expression level in SH-SY5Y cells [218]. Similar results were observed using the human neuroblastoma cell line NMB as the administration of dopamine induced apoptosis along with a reduction on the mRNA level for VDAC-1, VDAC-2 and VDAC-3. Interestingly, transient transfection mediated expression of human VDAC or human VDAC-2 protected from dopamine-mediated neurotoxicity in NMB cells [219]. It needs to be further investigated why the toxic effect of a 24 hour treatment of dopamine (1µg/ml) is reduced after transfection of cells with VDAC or VDAC-2 while in other degenerative models (see below) VDAC acts as a pro-apoptotic protein. Certainly, this calls for a critical pre-evaluation of the therapeutic application in PD when targeting VDACs.
Contrastingly and in line with the notion that VDAC could be considered as a protein with pro-apoptotic activity, an increased VDAC mRNA and protein level was observed in SH-SY5Y cells upon rotenone treatment. Rotenone is used in cellular PD models as it inhibits mitochondrial complex I leading to translocation of Cyto c [220]. Furthermore, the expression of VDAC-1 is increased upon 1-methyl-4-phenylpyridinium (MPP+) [157,221] and 6-hydroxydopamine (6-OHDA) [222] induction in PD cell models. Recently, the 6-OHDA induced VDAC-1 upregulation could be reduced by administration of vitamin D in a male rat model of PD [223]. Interestingly, there might be a common control for the expression of VDAC-1 and αSyn respectively, because both mRNAs are targets of the same miR-7 [157,224].
A changed expression of several miRs has been connected to the pathogenesis of PD [225]. The analysis of postmortem PD brain samples showed a reduction in miR-7 in comparison to samples of a healthy control group. Furthermore, using a mouse model for inducing loss of miR-7 resulted in increased expression of αSyn along with a reduction of nigral DA neurons and reduced striatal dopamine content [226]. In SH-SY5Y cells, miR-7 overexpression reduced VDAC-1 expression and thereby protected the cells from MPP+-induced initiation of apoptosis such as Cyto c release and calcium efflux [157]. Interestingly, the synthetic cholesterol-like compound olesoxime (cholest-4-en-3-one, oxime, TRO19622) binds into the OMM and thereby inhibits the translocation of αSyn through VDAC thus mediating neuroprotection [227,228]. Recently, a small membrane-binding peptide called HK2p mediated neuroprotection by inhibiting the complex formation between VDAC and αSyn. Furthermore, HK2p induces detachment of αSyn from the OMM resulting in an open state of VDAC allowing ATP/ADP exchange and restoring the mitochondrial potential [229]. Recently, the role of the antioxidant and anti-inflammatory phytoalexin resveratrol was investigated in the context of VDAC-1 in the pathogenesis of PD. Using the A53T mouse model, the administration of resveratrol resulted in decreased expression of VDAC-1 and αSyn on protein level in DA neurons. Consequently, the opening of PTP was prevented by resveratrol in DA neurons. Interestingly, there was no change in the expression levels of VDAC-2 and VDAC-3 in DA neurons. Furthermore, resveratrol improved the cognitive and motor abilities in this model as shown by animal behavioral tests [230].
Another protein involved in PD is LRRK2. As shown in ETNA (embryonic neuronal precursors) cells and in SH-SY5Y cells, overexpression of wild type LRRK2 and its mutants R1441C, G2019S and Y1699C mediated mitochondrial-dependent neuronal apoptosis [231]. LRRK2 directly interacts with three proteins of the PTP, such as ANT, VDAC and ubiquitous mitochondrial creatine kinase (uMtCK) as demonstrated in PC12 cells which is a pheochromocytoma cell line derived from the rat adrenal medulla. The processing of immature uMtCK can be inhibited by both LRRK2 and its G2019S mutant due to suppressing the translocation of uMtCK into mitochondria and thereby retaining the preprotein uMtCK on the OMM. Furthermore, the expression of wild type and G2019S LRRK2 supported the interaction between VDAC and ANT which is important for the opening of the PTP, allowing the release of Cyto c resulting in neuronal apoptosis (Figure 3). [232]. However, the exact molecular composition of the PTP is still a matter of ongoing research although this pore has been studied for over 50 years as recently reviewed in [67].
Mutations in the genes of PINK1 and PRKN cause recessive forms of PD [233,234,235,236]. PINK1 and PRKN are important regarding mitochondrial quality control. Under cellular stress conditions when mitochondria are depolarized, the phosphorylated serine/threonine kinase PINK1 localizes at the OMM recruiting the E3 ligase PRKN to the OMM as well. Upon phosphorylation by PINK1, PRKN ubiquitinates several proteins of the OMM and thereby inducing mitophagy, which is a variant of autophagy for removing damaged mitochondria. [237,238]. At least VDAC-1 is essential and required for the PINK1/PRKN-induced mitophagy. Furthermore, VDAC-1 becomes poly-ubiquitylated by PRKN. Several mutations of PRKN impairment induce mitophagy and thereby damaged mitochondria are not removed by mitophagy enabling the release of Cyto c resulting in apoptosis (Figure 3) [239]. In addition, it was demonstrated that PRKN directly interacts with all isoforms of VDAC (VDAC-1, -2 and -3) at the OMM mediating mitophagy [53]. Recently, cryo-electron microscopy was used solving a structure at a 3.1 Å resolution of dimeric human PINK1 bound to a symmetric array of a central VDAC-2 dimer while each VDAC-2 protein was surrounded by TOM5 and TOM20 [240]. VDAC-1 can become mono- and polyubiquitinated by PRKN. This leads to different physiological outcomes as VDAC-1 monoubiquitination prevents apoptosis while VDAC-1 polyubiquitination induces mitophagy, demonstrating the central role of VDAC-1 in the antagonistically regulated apoptosis and mitophagy in response to the PINK1–PRKN pathway. Interestingly, the mutant PRKN T415N prevents monoubiquitination of VDAC-1 and consequently apoptosis is not inhibited. Nevertheless, polyubiquitination of VDAC-1 by PRKN T415N is not affected. PRKN T415N belongs to one of different mutations of PRKN found in PD patients [241]. Interestingly, silencing of PINK1 induced mitophagy associated with a changed mitochondrial morphology, enhanced production of ROS, a loss of the mitochondrial membrane potential and opening of PTP as shown in mouse dopaminergic MN9D cells [242]. In a PD mouse model, the administration of the drug idebenone upregulated the expression of mt-VDAC-1, thus activating PINK1/PRKN-mediated mitophagy so that damaged mitochondria were removed. Consequently, damage of dopaminergic neurons was reduced and behavioral disorders were improved [243].
The protein DJ-1 is encoded by the gene PARK7 which belongs to several genes associated with familial forms of early-onset PD. DJ-1 functions as molecular chaperone, redox sensor, antioxidant scavenger and transcriptional regulator. DJ-1 can be located in the cytosol, in the nucleus and upon oxidative stress it is translocated to mitochondria allowing to develop its pleiotropic functions [244]. Furthermore, DJ-1 is involved in maintaining the function and integrity of the mitochondrial network controlling the mitochondrial Ca2+ homeostasis via the regulation of the interaction between the endoplasmic reticulum (ER) and mitochondria [245]. Moreover, DJ-1 is an essential and important component of the IP3R3-Grp75-VDAC-1 complex consisting of the ER Ca2+ channel inositol 1,4,5-trisphosphate receptor (IP3R3), the mitochondrial chaperone glucose-regulated protein 75 (Grp75) and VDAC-1. This complex is located at the mitochondria-associated membrane (MAM), in particular the VDAC-1 in the OMM while IP3R3 and GRP75 are inserted in the membrane of the ER. This DJ1-IP3R3-Grp75-VDAC-1 complex enables efficient interorganelle transfer of Ca2+ [246,247]. The IP3R3-Grp75-VDAC-1 disrupts upon ablation of DJ-1 resulting in reduced interaction of the ER with mitochondria, disturbed Ca2+ efflux from the ER and accumulation of IP3R3 at the MAM (Figure 3). The PD-associated mutant DJ-1 L166P shows reduced interaction in the DJ1-IP3R3-Grp75-VDAC-1 complex. Furthermore, sporadic PD patients showed reduced levels of DJ-1 in the substantia nigra associated with lower ER-mitochondria interaction and reduced IP3R3-DJ-1 interaction. In summary, mutated or reduced DJ-1 disturbs the association of ER with mitochondria and consequently takes part in the pathogenesis of PD [246].
Taken together, several PD-related proteins such as αSyn, LRRK2, PINK1/PRKN and DJ-1 directly interact with VDAC and thereby include VDAC as a possible candidate for executing the pathogenic effects in PD brain.
4. Amyotrophic Lateral Sclerosis and VDAC
The progressive adult-onset neurodegenerative disease amyotrophic lateral sclerosis (ALS; also called Lou Gehrig’s disease) is based on the loss of upper and lower motor neurons in the spinal cord and brain [248]. Typically, the onset occurs in the age of 50 to 60 years characterized by progressive paralysis and finally these patients will die within 2-5 years upon onset by respiratory failure [249]. Curative treatments are not available for ALS so far [250]. While 90 % of ALS cases are sporadic (sALS) without any genetic background, the remaining 10 % of cases show an autosomal dominant trait (familial ALS, fALS). In more than three decades of research, over 40 different ALS associated genes were identified among them for example copper-zinc superoxide dismutase (SOD1), transactive response DNA-binding protein 43 (TDP-43) and chromosome 9 open reading frame 72 (C9orf72) [251]. ALS shows several cellular deficits such as oxidative stress, mitochondrial dysfunction, glutamate excitotoxicity, axonal transport dysregulation, endosomal and vesicular transport impairment, aberrant RNA metabolism and impaired protein homeostasis [252]. Mutations in the gene coding for the cytoplasmic SOD1 are associated with 20% of the fALS [253]. Disturbances in the redox properties of SOD1 yield to atypical structural changes of the SOD1 protein resulting in a gain of toxic properties which mediates cell death of motor neurons in both sALS and fALS [254]. As shown in mouse motor-neuron-like NSC-34 cells, mutated SOD1 misfolds and accumulates in the cytosol while there is a correlation between the mutation severity and the degree of protein misfolding. In addition, the mutation severity of SOD1 correlates with its property to associate with mitochondria [255]. This association of misfolded SOD1 with the OMM results in mitochondrial dysfunction and further on in cellular toxicity via two different mechanisms: First, misfolded SOD1 binds to mt-VDAC-1 at the OMM and changes its channel conductance for adenine nucleotides disturbing the cellular energy supply [256]. Second, the import of proteins into mitochondria is suppressed by misfolded SOD1 yielding to a changed protein composition [257]. The mutants of human SOD1 G93A and SOD1 H46R interact with the cytoplasmic part of mt-VDAC-1. This interaction was observed with reconstituted purified components and on isolated mitochondria derived from spinal cord of an ALS rat model expressing mutant human SOD1. Spinal cord mitochondria from these ALS rats show reduced ADP passage through the OMM. As shown by reconstitution experiment using a lipid bilayer, the channel conductance was inhibited by direct binding of mutant SOD1 to VDAC-1. Using a peptide array, the residues 28-61 of SOD1 were identified for the interaction with VDAC-1 [258]. Furthermore, mice expressing SOD1 G37R showed a reduced VDAC-1 activity and an accelerating onset of fatal paralysis yielding to a reduced life span [256]. Moreover, there is an interaction between mutant SOD1 G93A and Bcl-2 altering the interaction between Bcl-2 and VDAC-1 resulting in a reduced OMM permeability. The complex formation between SOD1 G93A and Bcl-2 can be inhibited by small SOD1-like peptides preventing mitochondrial hyperpolarization and mediating protection from cell death as demonstrated in NSC-34 cells. [259]. A further study using the SOD1-G93A mouse model unraveled that the interaction of mutant SOD1 with Bcl-2 induces a conformational change of Bcl-2 uncovering its toxic BH3 domain and thereby converting it into a toxic protein [260]. The interaction of SOD G93A and SOD1 G85R with VDAC-1 could be narrowed down to the N-terminus of VDAC-1. Interestingly, wild type SOD1 does not interact with VDAC-1. Expression of SOD1 G93A or SOD1 G37R respectively in NSC-34 cells and expression of SOD1 G93A in mouse embryonic stem cell-derived motor neurons resulted in cell death which could be reduced upon the application of small cell-penetrating peptides mimicking the VDAC-1 N-terminus. These VDAC-1 N-terminal derived peptides interact with mutant SOD1 preventing the interaction with mt-VDAC-1 and thus protected from mitochondrial dysfunction [261].
While under physiological condition HKI binds to mt-VDAC-1 and protects from apoptosis [90,91], mutant SOD1 G93A competes with HKI for the same binding site on mt-VDAC-1 and consequently this SOD1-VDAC-1 association mediates mitochondrial dysfunction. Interestingly, application of peptides derived from the N-terminus of HKI recovered the cell viability and resulted in an increased mt-VDAC-1 expression and reduced mutant SOD1 accumulation at mitochondria in NSC-34 cells expressing SOD1 G93A [262,263]. A proteomic approach was applied on NSC-34 cell investigating how the proteome changes upon the expression of SOD1 G93A. The results showed changes in several mitochondrial proteins, among them a reduced expression of mt-VDAC-1 and mt-VDAC-2. In addition, changes of the post-translational modifications of mt-VDAC-2 were found suggesting its involvement in regulation of apoptosis and mutant SOD1-mediated neurotoxicity [264]. A further study revealed changes in the posttranslational modifications of VDAC-1 derived from NSC-34 cells expressing SOD1 G93A showing that VDAC-1 has selective deamidations of asparagine and glutamine as well as over-oxidation of methionine and cysteines. These altered posttranslational modifications may result in important structural changes of mt-VDAC-1 influencing the energetic metabolism of motor neurons in ALS [265].
Recently, in NSC-34 cells expressing human SOD1 G93A changes of the post-translational modifications of VDAC-3 were found such as succination events, deamidation, and over-oxidation. In more detail, VDAC-3 with deamidation of Asn215 embedded in artificial membranes showed an altered single channel behaviour. This might affect the protective property of VDAC-3 against ROS which is essential in the context of ALS [266]. An ALS mouse model expressing SOD1 G93A was treated with the VDAC inhibitor olesoxime resulting in delayed muscle denervation and delayed death of motor neurons [267]. The administration of olesoxime was tested in a clinical phase II – III trail in ALS patients but it failed to show beneficial effects [268]. As recently demonstrated, mitochondria isolated from spinal cord of mutant SOD1 G93A rats and mice respectively showed oligomerisation of VDAC-1. The small molecules VBIT-12 is an inhibitor for VDAC-1 oligomerization and thereby prevents apoptosis, like VBIT-4 as mentioned above (see 2). Administration of VBIT-12 in mutant SOD1 G93A mice improved muscle endurance although the survival was not extended [258].
Interestingly, spinal motor neurons derived from the SOD1 G93A mouse model showed an increased level of VDAC-1 before the onset of symptoms [258]. An upregulated VDAC-1 expression due to an adeno-associated virus (AAV) 2/5 injected into the spinal cord of pre-symptomatic neonatal pups of SOD1 G93A mouse restored the mitochondrial respiratory profile. This can be explained by enhanced activity of key regulators of mitochondrial maintenance and function such as sirtuins (Sirt), the respiratory chain complex I and the receptor subunit of the translocase of the outer membrane (TOM) complex, Tom20 [269]. Recently, blood samples from ALS patients were used for analyzing transcriptomic data with the help of an ARACNe-AP (Algorithm for the Reconstruction of Accurate Cellular Networks - Adaptive Partitioning) algorithm unrevealing four subnetworks of hub genes including one subnetwork with VDAC-3 being one hub gene. Each hub gene showed a connection to p53-mediated pathways which might be linked to ALS neuroinflammation [270].
Another protein involved in ALS is the heterogeneous nuclear ribonucleoprotein TDP-43. A mouse model of mutant TDP-43 and iPSC-derived motor neurons from ALS patients showed that TDP-43 induced the release of mtDNA into the cytosol through the PTP. This could be prevented by VBIT-4, the VDAC-1 oligomerization inhibitor [271] because mtDNA can be released through a channel of oligomeric VDAC-1 into the cytosol. [74]. Furthermore, knockout of VDAC-1 in mouse embryonic fibroblasts (MEFs) over-expressing TDP-43 inhibited the expression of the innate immune-related factors Ifnb1 (Interferon Beta 1) and TNF [271]. A direct interaction between TDP-43 and VDAC-1 was shown by a proteomic screen investigating mitochondrial interacting proteins of TDP-43 in a mouse model of motor neuron disease [272]. These studies suggest regarding ALS pathogenesis that TDP-43 may mislocalize into the mitochondria and there interact with VDAC-1 and thus enables the release of mtDNA into the cytosol.
As mentioned above (see 3), there is a protein complex built of VDAC-1 and GRP75 and further proteins for the crosstalk between ER and mitochondria at the MAM [247]. Recently, mechanisms of early ER stress were investigated using iPSC-derived motor neurons from C9orf72-ALS/FTD patients who have a monogenic form of ALS due to a hexanucleotide repeat expansion in their C9orf72 gene along with symptoms of frontotemporal dementia (FTD). These patients derived motor neurons showed elevated levels of GRP75 and VDAC-1 respectively along with increased IP3R-VDAC-1 interaction [273]. In addition, studies of spinal cord of C9-500 mice showed an increase of the IP3R-VDAC-1 interaction at post-natal day 125 (P125) and a decrease at P240 correlating to the GRP75 expression level. Furthermore, AAV-forced expression of GRP75 in C9-500 mice protected from ER stress, mitochondrial function was normalized and the IP3R-VDAC-1 interaction increased [273]. In conclusion, neurons in C9orf72-ALS/FTD are vulnerable to ER–mitochondrial dysfunction and the critical endogenous GRP75 protein shows neuroprotective properties influencing the IP3R-VDAC-1 interaction.
Taken together, all three isoforms of VDAC are involved in ALS, although most studies focus on VDAC-1. ALS related proteins such as mutated SOD1 and TDP-43 directly interact with mt-VDAC-1 while there are no studies showing any functional connection of pl-VDAC-1 in the pathogenesis of ALS. The interaction of mutated SOD1 with mt-VDAC-1 changes the channel conductance resulting in disturbed cellular energy. Although the application of small peptide, inhibitors such as VBIT-4, VBIT-12 or olesoxime mediated beneficial and promising effect in the ALS models, unfortunately no drug arises for a treatment of ALS. As more than 40 genes are associated to ALS, maybe further ALS relevant VDAC-interacting proteins might be discovered in future.
5. Huntington’s Disease and VDAC
The fatal human neurodegenerative disorder Huntington’s disease (HD) is caused by a CAG repeat expansion in exon 1 of the huntingtin gene (HTT). HD is inherited in an autosomal dominant manner. Several CAG repeats are translated into N-terminal polyglutamine residues resulting in a mutant HTT protein forming aggregates in different cells including neurons [274]. Aggregates of mutant HTT protein induce atrophy in subthalamic nuclei and in the basal ganglia, aberrations in the subcortical white matter and finally result in the development of neurologic symptoms [275]. These symptoms comprise involuntary movements, such as dystonia, chorea, bruxism, bradykinesia and rigidity as well as psychiatric symptoms along with cognitive decline resulting in dementia [276]. Different stages of disease are characterised by several complex cascades involving impaired proteostasis, excitotoxicity, oxidative stress, mitochondrial dysfunction, transcriptional dysregulation and neuroinflammation [277]. So far, there is no therapy available to treat HD. Worldwide 2.7 per 100,000 persons are affected by HD while Europe shows a higher rate with 10 per 100,000 individuals [274].
The effect of olesoxime was investigated in the BACHD rat model of HD, in which full length HTT along with 97 CAA/CAG repeats are overexpressed. The administration of olesoxime reduced cleavage of mutant HTT, accumulation of mutant HTT fragments and calpain activation and results in improved mitochondrial function. Besides these neuropathological improvements, olesoxime treatment resulted in behavioural improvements as well. In respect to VDAC, olesoxime treated BACHD rats showed increased expression of mt-VDAC-1 and mt-VDAC-2 in cortex and striatum which was not observed in BACHD rats without olesoxime treatment [278]. In a further study, the HD mouse model R6/2 which expresses mutant HTT with 120 to 128 glutamines was investigated to assess mitochondrial function and the rate of oxygen consumption in vivo. The striatum showed a reduced oxygen consumption because of less mitochondria while the cortical oxygen consumption was lowered by disturbances of energetic pathways. Several mitochondrial proteins were compared in their expression level with wild type mice and thereby a lower mt-VDAC-1 expression was found in the striatum while there was no change in cortex [279]. In line with this, mouse striatum-derived STHdh cells expressing mutant HTT with 111 glutamine residues either homozygous or heterozygous were investigated towards their mitochondrial morphology, the mitochondrial disulfide relay system and their function in HD. Besides several other findings, these cells showed a reduced VDAC expression while this reduction was stronger in cells homzygously expressing mutant HTT than in heterozygous expressing cells [280]. Furthermore, the R6/2 mouse model was analysed by a proteomic approach showing that six proteins were oxidized over the course of disease, among them VDAC-1 [281]. In order to study the role of VDAC in the pathogenesis of HD, VDAC was extracted from PC12 model cells expressing HTT or mutant HTT with 74 repeats of glutamine respectively. VDAC was reconstituted into artificial membranes for further analysis. Although the ratio between the three VDAC isoforms were not changed, only VDAC-1 showed a changed open state conductance and a changed voltage-dependence upon the expression of mutant HTT but not of wild-type HTT expression [282]. A recent study showed that extracellular vesicles derived from HD cells contained mtDNA and several mitochondrial proteins, among them VDAC-1. These HD cells were neurons with a striatal phenotype obtained by reprogramming of neuronal stem cells derived from induced-pluripotent stem cells which were derived from reprogrammed human fibroblasts of individuals considered as HD carrier. Stress induces mitochondria to generate mitochondrial-derived vesicles which fuse with the endolysosomal system forming multivesicular bodies that are released as extracellular vesicles from the cells [283]. Recently, rats were treated with the HD model substance 3-nitropropionic acid (3-NP) resulting in impaired mitochondrial respiration and mitochondrial dynamics as well as in induced ER-stress. Proteins such as VDAC-1 and Grp75, which are involved in the mitochondria – ER communication (see 3), showed elevated expression levels. Application of bezafibrate inhibited these 3-NP mediated changes including a normalised expression level of VDAC-1 and Grp75 [284]. In a recent study, rats were treated with 3-NP and then tested how the antioxidant flavonoid morin hydrate (MH) may counteract the 3-NP mediated HD phenotype. MH showed neuroprotective properties because motor dysfunction, degeneration of striatal neuros, ER stress, apoptosis and mitophagy were reduced. Thereby, MH mediated inactivation of VDAC-1 by its phosphorylation [285].
Taken together, data from different cellular and animal models show an involvement of VDAC-1 and VDAC-2 in HD. Substances such as olesoxime, bezafibrate and MH mediated beneficial effects towards HD in these model systems and thereby involved VDAC direct or indirect. However, there is no data about the expression level of VDAC in HD patients available so far. Furthermore, no functional relationship between the plasmalemmal localized VDAC and HD has been demonstrated so far.
6. Further Neurodegenerative Conditions and VDAC
The neurodegenerative lysosomal storage disease neuronal ceroid lipofuscinosis (NCL) – also known as Batten disease - comprises a group of 13 subtypes of NCL. Distinct genes are associated with each NCL subtype having mutations coding for lysosmal enzymes, transmembrane proteins or secretory proteins. This autosomal-recessive disorder manifests in infants by symptoms such as vision impairment, epilepsy, motor and cognitive dysfunctions, and premature death. Although, there are different treatment approaches such as pharmacological therapy, enzyme replacement therapy, stem cell therapy and gene therapy, all approaches have shown limited efficacy so far [286]. In one study two different genetic mouse models of NCL were investigated showing a progressive breakdown of synapses and axons in the cortex and thalamus of these mice. The expression levels for a group of proteins involved in axonal and synaptic vulnerability were quantified demonstrating individual expression profiles. Interestingly, the two proteins pituitary tumor-transforming gene 1 (PTTG1) product and VDAC-1 showed significant changes in their expression level in the thalamus at time-points of pre or early onset of symptoms in both mouse lines. Therefore, VDAC-1 was suggested to be used as an early biomarker for NCL [287].
Wernicke’s encephalopathy (WE) is a neurological disorder caused by thiamine (vitamin B1) deficiency characterised by eye movement abnormalities, acute confusion and ataxia [288]. WE can develop into a chronic syndrome called Wernicke-Korsakoff syndrome (WKS) [289]. One study showed the investigation of a WKS rat model regarding thiamine deficiency on spatial learning and differential protein expression. The expression level of seven thalamic proteins were significantly changed, among them VDAC. The VDAC expression was decreased when the rats showed worst performance in the behavioural test, called Morris Water Maze. While the VDAC expression was decreased on protein level, the mRNA level was unchanged suggesting that post-transcriptional processes may result in the decreased expression [290].
Recently, old male mice showed a reduced length and areas of their hippocampal mitochondria, increased neurodegeneration, a decline in recognition memory and changed protein expression including a decline in VDAC-1 in the hippocampus [291].
7. Conclusions
In summary, VDAC is involved in neurodegenerative diseases such as AD, PD, ALS and HD as well as additional neurodegenerative conditions. Typically, VDAC physically interacts with disease specific proteins such as Aβ, GSK3β, phosphorylated tau, TSPO, αSyn, LRRK2, PINK1/PRKN, DJ-1, mutated SOD-1, TDP-43, and polyglutamine mutated HTT resulting in modulation of mitochondrial permeability and participating in apoptosis (Figure 4). Furthermore, the expression level of VDAC might also be regulated by miRs like miR-29a and miR-7 or changed upon disease conditions. In line with the abundance of the different VDAC isoforms, most studies deal with VDAC-1 while investigations on VDAC-2 or VDAC-3 are less numerous. Although, VDAC is a prominent protein of the OMM, it is found in other cellular location e. g. in the plasma membrane. Only AD showed an involvement of pl-VDAC-1 while for all other neurodegenerative diseases reports are not available so far. The VDAC might be an interesting target for developing treatment options as evidenced by several inhibiting interventions using small molecules such as VBIT-4 and VBIT-12, olesoxime, resveratrol, idebenone, bezafibrate, morin hydrate, application of interfering peptides, antibodies or metformin, the latter being an established drug for the treatment of T2D. Taken together, VDAC may turn out to be a major player regulating neuronal survival in neurodegenerative diseases and therefore may provide an emerging platform for so far unrecognized therapeutical treatment options.
Author Contributions
Conceptualization, S.N. and R.H.; writing—original draft preparation, S.N.; writing—review and editing, S.N. and R.H.; visualization, S.N.; supervision, S.N. and R.H., project administration, R.H.; funding acquisition, R.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The graphical abstract was created with BioRender (https://BioRender.com; accessed on 15th May 2025).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schein, S.J.; Colombini, M.; Finkelstein, A. Reconstitution in planar lipid bilayers of a voltage-dependent anion-selective channel obtained from paramecium mitochondria. J Membr Biol 1976, 30, 99–120. [Google Scholar] [CrossRef] [PubMed]
- Colombini, M. A candidate for the permeability pathway of the outer mitochondrial membrane. Nature 1979, 279, 643–645. [Google Scholar] [CrossRef] [PubMed]
- Craigen, W.J.; Graham, B.H. Genetic strategies for dissecting mammalian and drosophila voltage-dependent anion channel functions. J Bioenerg Biomembr 2008, 40, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Sampson, M.J.; Ross, L.; Decker, W.K.; Craigen, W.J. A novel isoform of the mitochondrial outer membrane protein vdac3 via alternative splicing of a 3-base exon. Journal of Biological Chemistry 1998, 273, 30482–30486. [Google Scholar] [CrossRef]
- Colombini, M. Vdac structure, selectivity, and dynamics. Biochim Biophys Acta 2012, 1818, 1457–1465. [Google Scholar] [CrossRef]
- Raghavan, A.; Sheiko, T.; Graham, B.H.; Craigen, W.J. Voltage-dependant anion channels: Novel insights into isoform function through genetic models. Biochim Biophys Acta 2012, 1818, 1477–1485. [Google Scholar] [CrossRef]
- Freitag, H.; Janes, M.; Neupert, W. Biosynthesis of mitochondrial porin and insertion into the outer mitochondrial membrane of neurospora crassa. Eur J Biochem 1982, 126, 197–202. [Google Scholar] [CrossRef]
- Hartl, F.U.; Pfanner, N.; Nicholson, D.W.; Neupert, W. Mitochondrial protein import. Biochim Biophys Acta 1989, 988, 1–45. [Google Scholar] [CrossRef]
- Herrmann, J.M.; Bykov, Y. Protein translocation in mitochondria: Sorting out the toms, tims, pams, sams and mia. FEBS Lett 2023, 597, 1553–1554. [Google Scholar] [CrossRef]
- Benz, R. Solute transport through mitochondrial porins in vitro and in vivo. Biomolecules 2024, 14. [Google Scholar] [CrossRef]
- Messina, A.; Reina, S.; Guarino, F.; De Pinto, V. Vdac isoforms in mammals. Biochim Biophys Acta 2012, 1818, 1466–1476. [Google Scholar] [CrossRef] [PubMed]
- Shoshan-Barmatz, V.; Nahon-Crystal, E.; Shteinfer-Kuzmine, A.; Gupta, R. Vdac1, mitochondrial dysfunction, and alzheimer’s disease. Pharmacol Res 2018, 131, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.H.; Kim, S.K.; Kim, E.H.; Kim, S.W.; Sohn, S.H.; Lee, S.C.; Choi, S.; Pyo, S.; Rhee, D.K. Korean red ginseng up-regulates c21-steroid hormone metabolism via cyp11a1 gene in senescent rat testes. J Ginseng Res 2011, 35, 272–282. [Google Scholar] [CrossRef]
- Heslop, K.A.; Milesi, V.; Maldonado, E.N. Vdac modulation of cancer metabolism: Advances and therapeutic challenges. Front Physiol 2021, 12, 742839. [Google Scholar] [CrossRef] [PubMed]
- Hiller, S.; Garces, R.G.; Malia, T.J.; Orekhov, V.Y.; Colombini, M.; Wagner, G. Solution structure of the integral human membrane protein vdac-1 in detergent micelles. Science 2008, 321, 1206–1210. [Google Scholar] [CrossRef]
- Bayrhuber, M.; Meins, T.; Habeck, M.; Becker, S.; Giller, K.; Villinger, S.; Vonrhein, C.; Griesinger, C.; Zweckstetter, M.; Zeth, K. Structure of the human voltage-dependent anion channel. Proc Natl Acad Sci U S A 2008, 105, 15370–15375. [Google Scholar] [CrossRef]
- Ujwal, R.; Cascio, D.; Colletier, J.P.; Faham, S.; Zhang, J.; Toro, L.; Ping, P.; Abramson, J. The crystal structure of mouse vdac1 at 2.3 a resolution reveals mechanistic insights into metabolite gating. Proc Natl Acad Sci U S A 2008, 105, 17742–17747. [Google Scholar] [CrossRef]
- Schredelseker, J.; Paz, A.; Lopez, C.J.; Altenbach, C.; Leung, C.S.; Drexler, M.K.; Chen, J.N.; Hubbell, W.L.; Abramson, J. High resolution structure and double electron-electron resonance of the zebrafish voltage-dependent anion channel 2 reveal an oligomeric population. J Biol Chem 2014, 289, 12566–12577. [Google Scholar] [CrossRef]
- Benz, R. Permeation of hydrophilic solutes through mitochondrial outer membranes: Review on mitochondrial porins. Biochim Biophys Acta 1994, 1197, 167–196. [Google Scholar] [CrossRef]
- Shoshan-Barmatz, V.; Maldonado, E.N.; Krelin, Y. Vdac1 at the crossroads of cell metabolism, apoptosis and cell stress. Cell Stress 2017, 1, 11–36. [Google Scholar] [CrossRef]
- Najbauer, E.E.; Becker, S.; Giller, K.; Zweckstetter, M.; Lange, A.; Steinem, C.; de Groot, B.L.; Griesinger, C.; Andreas, L.B. Structure, gating and interactions of the voltage-dependent anion channel. Eur Biophys J 2021, 50, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Jahn, H.; Bartos, L.; Dearden, G.I.; Dittman, J.S.; Holthuis, J.C.M.; Vacha, R.; Menon, A.K. Phospholipids are imported into mitochondria by vdac, a dimeric beta barrel scramblase. Nat Commun 2023, 14, 8115. [Google Scholar] [CrossRef] [PubMed]
- Rostovtseva, T.K.; Bezrukov, S.M.; Hoogerheide, D.P. Regulation of mitochondrial respiration by vdac is enhanced by membrane-bound inhibitors with disordered polyanionic c-terminal domains. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Colombini, M. Vdac closure increases calcium ion flux. Biochim Biophys Acta 2007, 1768, 2510–2515. [Google Scholar] [CrossRef]
- Checchetto, V.; Reina, S.; Magri, A.; Szabo, I.; De Pinto, V. Recombinant human voltage dependent anion selective channel isoform 3 (hvdac3) forms pores with a very small conductance. Cell Physiol Biochem 2014, 34, 842–853. [Google Scholar] [CrossRef]
- Sander, P.; Gudermann, T.; Schredelseker, J. A calcium guard in the outer membrane: Is vdac a regulated gatekeeper of mitochondrial calcium uptake? Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Shuvo, S.R.; Ferens, F.G.; Court, D.A. The n-terminus of vdac: Structure, mutational analysis, and a potential role in regulating barrel shape. Biochim Biophys Acta 2016, 1858, 1350–1361. [Google Scholar] [CrossRef]
- Magri, A.; Reina, S.; De Pinto, V. Vdac1 as pharmacological target in cancer and neurodegeneration: Focus on its role in apoptosis. Front Chem 2018, 6, 108. [Google Scholar] [CrossRef]
- Shoshan-Barmatz, V.; Israelson, A. The voltage-dependent anion channel in endoplasmic/sarcoplasmic reticulum: Characterization, modulation and possible function. J Membr Biol 2005, 204, 57–66. [Google Scholar] [CrossRef]
- Thinnes, F.P.; Gotz, H.; Kayser, H.; Benz, R.; Schmidt, W.E.; Kratzin, H.D.; Hilschmann, N. [identification of human porins. I. Purification of a porin from human b-lymphocytes (porin 31hl) and the topochemical proof of its expression on the plasmalemma of the progenitor cell]. Biol Chem Hoppe Seyler 1989, 370, 1253–1264. [Google Scholar] [CrossRef]
- Bathori, G.; Parolini, I.; Tombola, F.; Szabo, I.; Messina, A.; Oliva, M.; De Pinto, V.; Lisanti, M.; Sargiacomo, M.; Zoratti, M. Porin is present in the plasma membrane where it is concentrated in caveolae and caveolae-related domains. J Biol Chem 1999, 274, 29607–29612. [Google Scholar] [CrossRef] [PubMed]
- De Pinto, V.; Messina, A.; Lane, D.J.; Lawen, A. Voltage-dependent anion-selective channel (vdac) in the plasma membrane. FEBS Lett 2010, 584, 1793–1799. [Google Scholar] [CrossRef] [PubMed]
- Buettner, R.; Papoutsoglou, G.; Scemes, E.; Spray, D.C.; Dermietzel, R. Evidence for secretory pathway localization of a voltage-dependent anion channel isoform. Proc Natl Acad Sci U S A 2000, 97, 3201–3206. [Google Scholar] [CrossRef]
- Li, L.; Yao, Y.C.; Gu, X.Q.; Che, D.; Ma, C.Q.; Dai, Z.Y.; Li, C.; Zhou, T.; Cai, W.B.; Yang, Z.H.; et al. Plasminogen kringle 5 induces endothelial cell apoptosis by triggering a voltage-dependent anion channel 1 (vdac1) positive feedback loop. J Biol Chem 2014, 289, 32628–32638. [Google Scholar] [CrossRef]
- Bahamonde, M.I.; Fernandez-Fernandez, J.M.; Guix, F.X.; Vazquez, E.; Valverde, M.A. Plasma membrane voltage-dependent anion channel mediates antiestrogen-activated maxi cl- currents in c1300 neuroblastoma cells. J Biol Chem 2003, 278, 33284–33289. [Google Scholar] [CrossRef] [PubMed]
- Bathori, G.; Parolini, I.; Szabo, I.; Tombola, F.; Messina, A.; Oliva, M.; Sargiacomo, M.; De Pinto, V.; Zoratti, M. Extramitochondrial porin: Facts and hypotheses. J Bioenerg Biomembr 2000, 32, 79–89. [Google Scholar] [CrossRef]
- Baker, M.A.; Lane, D.J.; Ly, J.D.; De Pinto, V.; Lawen, A. Vdac1 is a transplasma membrane nadh-ferricyanide reductase. J Biol Chem 2004, 279, 4811–4819. [Google Scholar] [CrossRef]
- Baker, M.A.; Ly, J.D.; Lawen, A. Characterization of vdac1 as a plasma membrane nadh-oxidoreductase. Biofactors 2004, 21, 215–221. [Google Scholar] [CrossRef]
- Okada, S.F.; O’Neal, W.K.; Huang, P.; Nicholas, R.A.; Ostrowski, L.E.; Craigen, W.J.; Lazarowski, E.R.; Boucher, R.C. Voltage-dependent anion channel-1 (vdac-1) contributes to atp release and cell volume regulation in murine cells. J Gen Physiol 2004, 124, 513–526. [Google Scholar] [CrossRef]
- Zhang, E.; Mohammed Al-Amily, I.; Mohammed, S.; Luan, C.; Asplund, O.; Ahmed, M.; Ye, Y.; Ben-Hail, D.; Soni, A.; Vishnu, N.; et al. Preserving insulin secretion in diabetes by inhibiting vdac1 overexpression and surface translocation in beta cells. Cell Metab 2019, 29, 64–77.e66. [Google Scholar] [CrossRef]
- Hinsch, K.D.; De Pinto, V.; Aires, V.A.; Schneider, X.; Messina, A.; Hinsch, E. Voltage-dependent anion-selective channels vdac2 and vdac3 are abundant proteins in bovine outer dense fibers, a cytoskeletal component of the sperm flagellum. J Biol Chem 2004, 279, 15281–15288. [Google Scholar] [CrossRef] [PubMed]
- Menzel, V.A.; Cassara, M.C.; Benz, R.; de Pinto, V.; Messina, A.; Cunsolo, V.; Saletti, R.; Hinsch, K.D.; Hinsch, E. Molecular and functional characterization of vdac2 purified from mammal spermatozoa. Biosci Rep 2009, 29, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Paradowska, A.; Bohring, C.; Krause, E.; Krause, W. Identification of evolutionary conserved mouse sperm surface antigens by human antisperm antibodies (asa) from infertile patients. Am J Reprod Immunol 2006, 55, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wang, P.; Wang, Z.; Zhang, W. The use of anti-vdac2 antibody for the combined assessment of human sperm acrosome integrity and ionophore a23187-induced acrosome reaction. PLoS One 2011, 6, e16985. [Google Scholar] [CrossRef]
- Ning, L.; Pan, B.; Zhao, Y.P.; Liao, Q.; Zhang, T.P.; Chen, G.; Wang, W.B.; Yang, Y.C. [immuno-proteomic screening of human pancreatic cancer associated membrane antigens for early diagnosis]. Zhonghua Wai Ke Za Zhi 2007, 45, 34–38. [Google Scholar] [CrossRef]
- Valis, K.; Neubauerova, J.; Man, P.; Pompach, P.; Vohradsky, J.; Kovar, J. Vdac2 and aldolase a identified as membrane proteins of k562 cells with increased expression under iron deprivation. Mol Cell Biochem 2008, 311, 225–231. [Google Scholar] [CrossRef]
- Stark, G. Functional consequences of oxidative membrane damage. J Membr Biol 2005, 205, 1–16. [Google Scholar] [CrossRef]
- Budzinska, M.; Galganska, H.; Wojtkowska, M.; Stobienia, O.; Kmita, H. Effects of vdac isoforms on cuzn-superoxide dismutase activity in the intermembrane space of saccharomyces cerevisiae mitochondria. Biochem Biophys Res Commun 2007, 357, 1065–1070. [Google Scholar] [CrossRef]
- Zhou, H.; Ren, J.; Toan, S.; Mui, D. Role of mitochondrial quality surveillance in myocardial infarction: From bench to bedside. Ageing Res Rev 2021, 66, 101250. [Google Scholar] [CrossRef]
- Bertholet, A.M.; Delerue, T.; Millet, A.M.; Moulis, M.F.; David, C.; Daloyau, M.; Arnaune-Pelloquin, L.; Davezac, N.; Mils, V.; Miquel, M.C.; et al. Mitochondrial fusion/fission dynamics in neurodegeneration and neuronal plasticity. Neurobiology of disease 2016, 90, 3–19. [Google Scholar] [CrossRef]
- Nardin, A.; Schrepfer, E.; Ziviani, E. Counteracting pink/parkin deficiency in the activation of mitophagy: A potential therapeutic intervention for parkinson’s disease. Curr Neuropharmacol 2016, 14, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zhou, H.; Sun, Y.; Fu, H.; Ran, Y.; Yang, B.; Yang, F.; Bjorklund, M.; Xu, S. Miro-1 interacts with vdac-1 to regulate mitochondrial membrane potential in caenorhabditis elegans. EMBO Rep 2023, 24, e56297. [Google Scholar] [CrossRef]
- Sun, Y.; Vashisht, A.A.; Tchieu, J.; Wohlschlegel, J.A.; Dreier, L. Voltage-dependent anion channels (vdacs) recruit parkin to defective mitochondria to promote mitochondrial autophagy. J Biol Chem 2012, 287, 40652–40660. [Google Scholar] [CrossRef]
- Duncan, M.R.; Fullerton, M.; Chaudhuri, M. Tim50 in trypanosoma brucei possesses a dual specificity phosphatase activity and is critical for mitochondrial protein import. J Biol Chem 2013, 288, 3184–3197. [Google Scholar] [CrossRef]
- Gupta, R.; Ghosh, S. Phosphorylation of voltage-dependent anion channel by c-jun n-terminal kinase-3 leads to closure of the channel. Biochem Biophys Res Commun 2015, 459, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, M.; Alvir, R.V.; Alvir, R.V.; Bunquin, L.E.; Pradeepkiran, J.A.; Reddy, P.H. A partial reduction of vdac1 enhances mitophagy, autophagy, synaptic activities in a transgenic tau mouse model. Aging Cell 2022, 21, e13663. [Google Scholar] [CrossRef]
- Saini, N.; Lakshminarayanan, S.; Kundu, P.; Sarin, A. Notch1 modulation of cellular calcium regulates mitochondrial metabolism and anti-apoptotic activity in t-regulatory cells. Front Immunol 2022, 13, 832159. [Google Scholar] [CrossRef] [PubMed]
- Petrozzi, L.; Ricci, G.; Giglioli, N.J.; Siciliano, G.; Mancuso, M. Mitochondria and neurodegeneration. Biosci Rep 2007, 27, 87–104. [Google Scholar] [CrossRef]
- Radi, E.; Formichi, P.; Battisti, C.; Federico, A. Apoptosis and oxidative stress in neurodegenerative diseases. J Alzheimers Dis 2014, 42 (Suppl. 3), S125–S152. [Google Scholar] [CrossRef]
- Schneider, P.; Tschopp, J. Apoptosis induced by death receptors. Pharm Acta Helv 2000, 74, 281–286. [Google Scholar] [CrossRef]
- Ashkenazi, A. Targeting the extrinsic apoptotic pathway in cancer: Lessons learned and future directions. J Clin Invest 2015, 125, 487–489. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lai, Y.; Hua, Z.C. Apoptosis and apoptotic body: Disease message and therapeutic target potentials. Biosci Rep 2019, 39. [Google Scholar] [CrossRef] [PubMed]
- Glover, H.L.; Schreiner, A.; Dewson, G.; Tait, S.W.G. Mitochondria and cell death. Nat Cell Biol 2024. [Google Scholar] [CrossRef]
- Kilbride, S.M.; Prehn, J.H. Central roles of apoptotic proteins in mitochondrial function. Oncogene 2013, 32, 2703–2711. [Google Scholar] [CrossRef] [PubMed]
- Repnik, U.; Turk, B. Lysosomal-mitochondrial cross-talk during cell death. Mitochondrion 2010, 10, 662–669. [Google Scholar] [CrossRef]
- Vander Heiden, M.G.; Chandel, N.S.; Li, X.X.; Schumacker, P.T.; Colombini, M.; Thompson, C.B. Outer mitochondrial membrane permeability can regulate coupled respiration and cell survival. Proc Natl Acad Sci U S A 2000, 97, 4666–4671. [Google Scholar] [CrossRef]
- Endlicher, R.; Drahota, Z.; Stefkova, K.; Cervinkova, Z.; Kucera, O. The mitochondrial permeability transition pore-current knowledge of its structure, function, and regulation, and optimized methods for evaluating its functional state. Cells 2023, 12. [Google Scholar] [CrossRef]
- Vogler, M.; Braun, Y.; Smith, V.M.; Westhoff, M.A.; Pereira, R.S.; Pieper, N.M.; Anders, M.; Callens, M.; Vervliet, T.; Abbas, M.; et al. The bcl2 family: From apoptosis mechanisms to new advances in targeted therapy. Signal Transduct Target Ther 2025, 10, 91. [Google Scholar] [CrossRef]
- Antignani, A.; Youle, R.J. How do bax and bak lead to permeabilization of the outer mitochondrial membrane? Curr Opin Cell Biol 2006, 18, 685–689. [Google Scholar] [CrossRef]
- Lovell, J.F.; Billen, L.P.; Bindner, S.; Shamas-Din, A.; Fradin, C.; Leber, B.; Andrews, D.W. Membrane binding by tbid initiates an ordered series of events culminating in membrane permeabilization by bax. Cell 2008, 135, 1074–1084. [Google Scholar] [CrossRef]
- Banerjee, J.; Ghosh, S. Bax increases the pore size of rat brain mitochondrial voltage-dependent anion channel in the presence of tbid. Biochem Biophys Res Commun 2004, 323, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, S.; Tsujimoto, Y. Proapoptotic bh3-only bcl-2 family members induce cytochrome c release, but not mitochondrial membrane potential loss, and do not directly modulate voltage-dependent anion channel activity. Proc Natl Acad Sci U S A 2000, 97, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Keinan, N.; Tyomkin, D.; Shoshan-Barmatz, V. Oligomerization of the mitochondrial protein voltage-dependent anion channel is coupled to the induction of apoptosis. Mol Cell Biol 2010, 30, 5698–5709. [Google Scholar] [CrossRef]
- Kim, J.; Gupta, R.; Blanco, L.P.; Yang, S.; Shteinfer-Kuzmine, A.; Wang, K.; Zhu, J.; Yoon, H.E.; Wang, X.; Kerkhofs, M.; et al. Vdac oligomers form mitochondrial pores to release mtdna fragments and promote lupus-like disease. Science 2019, 366, 1531–1536. [Google Scholar] [CrossRef]
- Zalk, R.; Israelson, A.; Garty, E.S.; Azoulay-Zohar, H.; Shoshan-Barmatz, V. Oligomeric states of the voltage-dependent anion channel and cytochrome c release from mitochondria. Biochem J 2005, 386, 73–83. [Google Scholar] [CrossRef]
- Abu-Hamad, S.; Arbel, N.; Calo, D.; Arzoine, L.; Israelson, A.; Keinan, N.; Ben-Romano, R.; Friedman, O.; Shoshan-Barmatz, V. The vdac1 n-terminus is essential both for apoptosis and the protective effect of anti-apoptotic proteins. J Cell Sci 2009, 122, 1906–1916. [Google Scholar] [CrossRef] [PubMed]
- Shoshan-Barmatz, V.; Mizrachi, D. Vdac1: From structure to cancer therapy. Front Oncol 2012, 2, 164. [Google Scholar] [CrossRef]
- Shoshan-Barmatz, V.; Mizrachi, D.; Keinan, N. Oligomerization of the mitochondrial protein vdac1: From structure to function and cancer therapy. Prog Mol Biol Transl Sci 2013, 117, 303–334. [Google Scholar] [CrossRef]
- Keinan, N.; Pahima, H.; Ben-Hail, D.; Shoshan-Barmatz, V. The role of calcium in vdac1 oligomerization and mitochondria-mediated apoptosis. Biochim Biophys Acta 2013, 1833, 1745–1754. [Google Scholar] [CrossRef]
- Weisthal, S.; Keinan, N.; Ben-Hail, D.; Arif, T.; Shoshan-Barmatz, V. Ca(2+)-mediated regulation of vdac1 expression levels is associated with cell death induction. Biochim Biophys Acta 2014, 1843, 2270–2281. [Google Scholar] [CrossRef]
- Ben-Hail, D.; Begas-Shvartz, R.; Shalev, M.; Shteinfer-Kuzmine, A.; Gruzman, A.; Reina, S.; De Pinto, V.; Shoshan-Barmatz, V. Novel compounds targeting the mitochondrial protein vdac1 inhibit apoptosis and protect against mitochondrial dysfunction. J Biol Chem 2016, 291, 24986–25003. [Google Scholar] [CrossRef] [PubMed]
- Geula, S.; Naveed, H.; Liang, J.; Shoshan-Barmatz, V. Structure-based analysis of vdac1 protein: Defining oligomer contact sites. J Biol Chem 2012, 287, 2179–2190. [Google Scholar] [CrossRef] [PubMed]
- Smilansky, A.; Dangoor, L.; Nakdimon, I.; Ben-Hail, D.; Mizrachi, D.; Shoshan-Barmatz, V. The voltage-dependent anion channel 1 mediates amyloid beta toxicity and represents a potential target for alzheimer disease therapy. J Biol Chem 2015, 290, 30670–30683. [Google Scholar] [CrossRef]
- Huang, L.; Han, J.; Ben-Hail, D.; He, L.; Li, B.; Chen, Z.; Wang, Y.; Yang, Y.; Liu, L.; Zhu, Y.; et al. A new fungal diterpene induces vdac1-dependent apoptosis in bax/bak-deficient cells. J Biol Chem 2015, 290, 23563–23578. [Google Scholar] [CrossRef] [PubMed]
- Shoshan-Barmatz, V.; De, S.; Meir, A. The mitochondrial voltage-dependent anion channel 1, ca(2+) transport, apoptosis, and their regulation. Front Oncol 2017, 7, 60. [Google Scholar] [CrossRef]
- Shoshan-Barmatz, V.; Krelin, Y.; Shteinfer-Kuzmine, A. Vdac1 functions in ca(2+) homeostasis and cell life and death in health and disease. Cell Calcium 2018, 69, 81–100. [Google Scholar] [CrossRef]
- Arbel, N.; Shoshan-Barmatz, V. Voltage-dependent anion channel 1-based peptides interact with bcl-2 to prevent antiapoptotic activity. J Biol Chem 2010, 285, 6053–6062. [Google Scholar] [CrossRef]
- Arbel, N.; Ben-Hail, D.; Shoshan-Barmatz, V. Mediation of the antiapoptotic activity of bcl-xl protein upon interaction with vdac1 protein. J Biol Chem 2012, 287, 23152–23161. [Google Scholar] [CrossRef]
- Shimizu, S.; Konishi, A.; Kodama, T.; Tsujimoto, Y. Bh4 domain of antiapoptotic bcl-2 family members closes voltage-dependent anion channel and inhibits apoptotic mitochondrial changes and cell death. Proc Natl Acad Sci U S A 2000, 97, 3100–3105. [Google Scholar] [CrossRef]
- Abu-Hamad, S.; Zaid, H.; Israelson, A.; Nahon, E.; Shoshan-Barmatz, V. Hexokinase-i protection against apoptotic cell death is mediated via interaction with the voltage-dependent anion channel-1: Mapping the site of binding. J Biol Chem 2008, 283, 13482–13490. [Google Scholar] [CrossRef]
- Zaid, H.; Abu-Hamad, S.; Israelson, A.; Nathan, I.; Shoshan-Barmatz, V. The voltage-dependent anion channel-1 modulates apoptotic cell death. Cell Death Differ 2005, 12, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.; Pandey, N.; Maitra, A.; Brahmachari, S.K.; Pillai, B. A role for voltage-dependent anion channel vdac1 in polyglutamine-mediated neuronal cell death. PLoS One 2007, 2, e1170. [Google Scholar] [CrossRef] [PubMed]
- Godbole, A.; Varghese, J.; Sarin, A.; Mathew, M.K. Vdac is a conserved element of death pathways in plant and animal systems. Biochim Biophys Acta 2003, 1642, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.J.; Dong, C.W.; Du, C.S.; Zhang, Q.Y. Characterization and expression analysis of paralichthys olivaceus voltage-dependent anion channel (vdac) gene in response to virus infection. Fish Shellfish Immunol 2007, 23, 601–613. [Google Scholar] [CrossRef]
- Azoulay-Zohar, H.; Israelson, A.; Abu-Hamad, S.; Shoshan-Barmatz, V. In self-defence: Hexokinase promotes voltage-dependent anion channel closure and prevents mitochondria-mediated apoptotic cell death. Biochem J 2004, 377, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Tajeddine, N.; Galluzzi, L.; Kepp, O.; Hangen, E.; Morselli, E.; Senovilla, L.; Araujo, N.; Pinna, G.; Larochette, N.; Zamzami, N.; et al. Hierarchical involvement of bak, vdac1 and bax in cisplatin-induced cell death. Oncogene 2008, 27, 4221–4232. [Google Scholar] [CrossRef]
- Elinder, F.; Akanda, N.; Tofighi, R.; Shimizu, S.; Tsujimoto, Y.; Orrenius, S.; Ceccatelli, S. Opening of plasma membrane voltage-dependent anion channels (vdac) precedes caspase activation in neuronal apoptosis induced by toxic stimuli. Cell Death Differ 2005, 12, 1134–1140. [Google Scholar] [CrossRef]
- Akanda, N.; Tofighi, R.; Brask, J.; Tamm, C.; Elinder, F.; Ceccatelli, S. Voltage-dependent anion channels (vdac) in the plasma membrane play a critical role in apoptosis in differentiated hippocampal neurons but not in neural stem cells. Cell Cycle 2008, 7, 3225–3234. [Google Scholar] [CrossRef]
- Marin, R.; Ramirez, C.M.; Gonzalez, M.; Gonzalez-Munoz, E.; Zorzano, A.; Camps, M.; Alonso, R.; Diaz, M. Voltage-dependent anion channel (vdac) participates in amyloid beta-induced toxicity and interacts with plasma membrane estrogen receptor alpha in septal and hippocampal neurons. Mol Membr Biol 2007, 24, 148–160. [Google Scholar] [CrossRef]
- Koma, H.; Yamamoto, Y.; Okamura, N.; Yagami, T. A plausible involvement of plasmalemmal voltage-dependent anion channel 1 in the neurotoxicity of 15-deoxy-delta(12,14) -prostaglandin j2. Brain Behav 2020, 10, e01866. [Google Scholar] [CrossRef]
- Liu, Z.; Bengtsson, S.; Krogh, M.; Marquez, M.; Nilsson, S.; James, P.; Aliaya, A.; Holmberg, A.R. Somatostatin effects on the proteome of the lncap cell-line. Int J Oncol 2007, 30, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Thinnes, F.P. Neuroendocrine differentiation of lncap cells suggests: Vdac in the cell membrane is involved in the extrinsic apoptotic pathway. Mol Genet Metab 2009, 97, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Heumann, R.; Goemans, C.; Bartsch, D.; Lingenhohl, K.; Waldmeier, P.C.; Hengerer, B.; Allegrini, P.R.; Schellander, K.; Wagner, E.F.; Arendt, T.; et al. Transgenic activation of ras in neurons promotes hypertrophy and protects from lesion-induced degeneration. J Cell Biol 2000, 151, 1537–1548. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarty, K.; Serchov, T.; Mann, S.A.; Dietzel, I.D.; Heumann, R. Enhancement of dopaminergic properties and protection mediated by neuronal activation of ras in mouse ventral mesencephalic neurones. European Journal of Neuroscience 2007, 25, 1971–1981. [Google Scholar] [CrossRef]
- Felderhoff-Mueser, U.; Bittigau, P.; Sifringer, M.; Jarosz, B.; Korobowicz, E.; Mahler, L.; Piening, T.; Moysich, A.; Grune, T.; Thor, F.; et al. Oxygen causes cell death in the developing brain. Neurobiology of disease 2004, 17, 273–282. [Google Scholar] [CrossRef]
- Serdar, M.; Herz, J.; Kempe, K.; Winterhager, E.; Jastrow, H.; Heumann, R.; Felderhoff-Muser, U.; Bendix, I. Protection of oligodendrocytes through neuronal overexpression of the small gtpase ras in hyperoxia-induced neonatal brain injury. Frontiers in neurology 2018, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Hansen, H.H.; Briem, T.; Dzietko, M.; Sifringer, M.; Voss, A.; Rzeski, W.; Zdzisinska, B.; Thor, F.; Heumann, R.; Stepulak, A.; et al. Mechanisms leading to disseminated apoptosis following nmda receptor blockade in the developing rat brain. Neurobiology of disease 2004, 16, 440–453. [Google Scholar] [CrossRef]
- Makwana, M.; Serchov, T.; Hristova, M.; Bohatschek, M.; Gschwendtner, A.; Kalla, R.; Liu, Z.Q.; Heumann, R.; Raivich, G. Regulation and function of neuronal gtp-ras in facial motor nerve regeneration. Journal of Neurochemistry 2009, 108, 1453–1463. [Google Scholar] [CrossRef]
- Neumann, S.; Kuteykin-Teplyakov, K.; Heumann, R. Neuronal protection by ha-ras-gtpase signaling through selective downregulation of plasmalemmal voltage-dependent anion channel-1. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Dorszewska, J.; Prendecki, M.; Oczkowska, A.; Dezor, M.; Kozubski, W. Molecular basis of familial and sporadic alzheimer’s disease. Curr Alzheimer Res 2016, 13, 952–963. [Google Scholar] [CrossRef]
- Hauptmann, S.; Scherping, I.; Drose, S.; Brandt, U.; Schulz, K.L.; Jendrach, M.; Leuner, K.; Eckert, A.; Muller, W.E. Mitochondrial dysfunction: An early event in alzheimer pathology accumulates with age in ad transgenic mice. Neurobiol Aging 2009, 30, 1574–1586. [Google Scholar] [CrossRef] [PubMed]
- Leuner, K.; Hauptmann, S.; Abdel-Kader, R.; Scherping, I.; Keil, U.; Strosznajder, J.B.; Eckert, A.; Muller, W.E. Mitochondrial dysfunction: The first domino in brain aging and alzheimer’s disease? Antioxid Redox Signal 2007, 9, 1659–1675. [Google Scholar] [CrossRef] [PubMed]
- Manczak, M.; Park, B.S.; Jung, Y.; Reddy, P.H. Differential expression of oxidative phosphorylation genes in patients with alzheimer’s disease: Implications for early mitochondrial dysfunction and oxidative damage. Neuromolecular Med 2004, 5, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.H.; Tripathi, R.; Troung, Q.; Tirumala, K.; Reddy, T.P.; Anekonda, V.; Shirendeb, U.P.; Calkins, M.J.; Reddy, A.P.; Mao, P.; et al. Abnormal mitochondrial dynamics and synaptic degeneration as early events in alzheimer’s disease: Implications to mitochondria-targeted antioxidant therapeutics. Biochim Biophys Acta 2012, 1822, 639–649. [Google Scholar] [CrossRef]
- Gibson, G.E.; Shi, Q. A mitocentric view of alzheimer’s disease suggests multi-faceted treatments. J Alzheimers Dis 2010, 20 (Suppl. 2), S591–S607. [Google Scholar] [CrossRef]
- Rao, V.K.; Carlson, E.A.; Yan, S.S. Mitochondrial permeability transition pore is a potential drug target for neurodegeneration. Biochim Biophys Acta 2014, 1842, 1267–1272. [Google Scholar] [CrossRef]
- Moreira, P.I.; Carvalho, C.; Zhu, X.; Smith, M.A.; Perry, G. Mitochondrial dysfunction is a trigger of alzheimer’s disease pathophysiology. Biochim Biophys Acta 2010, 1802, 2–10. [Google Scholar] [CrossRef]
- Schmitt, K.; Grimm, A.; Kazmierczak, A.; Strosznajder, J.B.; Gotz, J.; Eckert, A. Insights into mitochondrial dysfunction: Aging, amyloid-beta, and tau-a deleterious trio. Antioxid Redox Signal 2012, 16, 1456–1466. [Google Scholar] [CrossRef]
- De Pinto, V.; al Jamal, J.A.; Benz, R.; Genchi, G.; Palmieri, F. Characterization of sh groups in porin of bovine heart mitochondria. Porin cysteines are localized in the channel walls. Eur J Biochem 1991, 202, 903–911. [Google Scholar] [CrossRef]
- De Pinto, V.; Reina, S.; Gupta, A.; Messina, A.; Mahalakshmi, R. Role of cysteines in mammalian vdac isoforms’ function. Biochim Biophys Acta 2016, 1857, 1219–1227. [Google Scholar] [CrossRef]
- Lemeshko, V.V. Vdac as a voltage-dependent mitochondrial gatekeeper under physiological conditions. Biochim Biophys Acta Biomembr 2023, 1865, 184175. [Google Scholar] [CrossRef] [PubMed]
- Manczak, M.; Reddy, P.H. Abnormal interaction of vdac1 with amyloid beta and phosphorylated tau causes mitochondrial dysfunction in alzheimer’s disease. Hum Mol Genet 2012, 21, 5131–5146. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado-Tejedor, M.; Vilarino, M.; Cabodevilla, F.; Del Rio, J.; Frechilla, D.; Perez-Mediavilla, A. Enhanced expression of the voltage-dependent anion channel 1 (vdac1) in alzheimer’s disease transgenic mice: An insight into the pathogenic effects of amyloid-beta. J Alzheimers Dis 2011, 23, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, C.M.; Gonzalez, M.; Diaz, M.; Alonso, R.; Ferrer, I.; Santpere, G.; Puig, B.; Meyer, G.; Marin, R. Vdac and eralpha interaction in caveolae from human cortex is altered in alzheimer’s disease. Mol Cell Neurosci 2009, 42, 172–183. [Google Scholar] [CrossRef]
- Colurso, G.J.; Nilson, J.E.; Vervoort, L.G. Quantitative assessment of DNA fragmentation and beta-amyloid deposition in insular cortex and midfrontal gyrus from patients with alzheimer’s disease. Life Sci 2003, 73, 1795–1803. [Google Scholar] [CrossRef]
- Silva, D.F.; Selfridge, J.E.; Lu, J.; E, L.; Cardoso, S.M.; Swerdlow, R.H. Mitochondrial abnormalities in alzheimer’s disease: Possible targets for therapeutic intervention. Adv Pharmacol 2012, 64, 83–126. [Google Scholar] [CrossRef]
- Geula, S.; Ben-Hail, D.; Shoshan-Barmatz, V. Structure-based analysis of vdac1: N-terminus location, translocation, channel gating and association with anti-apoptotic proteins. Biochem J 2012, 444, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Munter, L.M.; Voigt, P.; Harmeier, A.; Kaden, D.; Gottschalk, K.E.; Weise, C.; Pipkorn, R.; Schaefer, M.; Langosch, D.; Multhaup, G. Gxxxg motifs within the amyloid precursor protein transmembrane sequence are critical for the etiology of abeta42. EMBO J 2007, 26, 1702–1712. [Google Scholar] [CrossRef]
- Thinnes, F.P. Apoptogenic interactions of plasmalemmal type-1 vdac and abeta peptides via gxxxg motifs induce alzheimer’s disease - a basic model of apoptosis? Wien Med Wochenschr 2011, 161, 274–276. [Google Scholar] [CrossRef]
- Thinnes, F.P. New findings concerning vertebrate porin ii--on the relevance of glycine motifs of type-1 vdac. Mol Genet Metab 2013, 108, 212–224. [Google Scholar] [CrossRef]
- Manczak, M.; Sheiko, T.; Craigen, W.J.; Reddy, P.H. Reduced vdac1 protects against alzheimer’s disease, mitochondria, and synaptic deficiencies. J Alzheimers Dis 2013, 37, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Shteinfer-Kuzmine, A.; Kamenetsky, N.; Pittala, S.; Paul, A.; Nahon Crystal, E.; Ouro, A.; Chalifa-Caspi, V.; Pandey, S.K.; Monsonego, A.; et al. Targeting the overexpressed mitochondrial protein vdac1 in a mouse model of alzheimer’s disease protects against mitochondrial dysfunction and mitigates brain pathology. Transl Neurodegener 2022, 11, 58. [Google Scholar] [CrossRef]
- Belosludtsev, K.N.; Ilzorkina, A.I.; Matveeva, L.A.; Chulkov, A.V.; Semenova, A.A.; Dubinin, M.V.; Belosludtseva, N.V. Effect of vbit-4 on the functional activity of isolated mitochondria and cell viability. Biochim Biophys Acta Biomembr 2024, 1866, 184329. [Google Scholar] [CrossRef]
- Iwamoto, N.; Kobayashi, K.; Kosaka, K. The formation of prostaglandins in the postmortem cerebral cortex of alzheimer-type dementia patients. J Neurol 1989, 236, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Marcone, S.; Fitzgerald, D.J. Proteomic identification of the candidate target proteins of 15-deoxy-delta12,14-prostaglandin j2. Proteomics 2013, 13, 2135–2139. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the nomenclature committee on cell death 2018. Cell Death Differ 2018, 25, 486–541. [Google Scholar] [CrossRef]
- Rockenfeller, P. Phospholipid scramblase activity of vdac dimers: New implications for cell death, autophagy and ageing. Biomolecules 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Jope, R.S.; Yuskaitis, C.J.; Beurel, E. Glycogen synthase kinase-3 (gsk3): Inflammation, diseases, and therapeutics. Neurochem Res 2007, 32, 577–595. [Google Scholar] [CrossRef]
- Pastorino, J.G.; Hoek, J.B.; Shulga, N. Activation of glycogen synthase kinase 3beta disrupts the binding of hexokinase ii to mitochondria by phosphorylating voltage-dependent anion channel and potentiates chemotherapy-induced cytotoxicity. Cancer Res 2005, 65, 10545–10554. [Google Scholar] [CrossRef]
- Liu, M.; Sui, D.; Dexheimer, T.; Hovde, S.; Deng, X.; Wang, K.W.; Lin, H.L.; Chien, H.T.; Kweon, H.K.; Kuo, N.S.; et al. Hyperphosphorylation renders tau prone to aggregate and to cause cell death. Mol Neurobiol 2020, 57, 4704–4719. [Google Scholar] [CrossRef]
- Reddy, P.H. Abnormal tau, mitochondrial dysfunction, impaired axonal transport of mitochondria, and synaptic deprivation in alzheimer’s disease. Brain Res 2011, 1415, 136–148. [Google Scholar] [CrossRef]
- Khan, A.; Kuriachan, G.; Mahalakshmi, R. Cellular interactome of mitochondrial voltage-dependent anion channels: Oligomerization and channel (mis)regulation. ACS Chem Neurosci 2021, 12, 3497–3515. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, I. Altered mitochondria, energy metabolism, voltage-dependent anion channel, and lipid rafts converge to exhaust neurons in alzheimer’s disease. J Bioenerg Biomembr 2009, 41, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Ebneth, A.; Godemann, R.; Stamer, K.; Illenberger, S.; Trinczek, B.; Mandelkow, E. Overexpression of tau protein inhibits kinesin-dependent trafficking of vesicles, mitochondria, and endoplasmic reticulum: Implications for alzheimer’s disease. J Cell Biol 1998, 143, 777–794. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, R.; Papadopoulos, V.; Rammes, G.; Baghai, T.C.; Fan, J.; Akula, N.; Groyer, G.; Adams, D.; Schumacher, M. Translocator protein (18 kda) (tspo) as a therapeutic target for neurological and psychiatric disorders. Nat Rev Drug Discov 2010, 9, 971–988. [Google Scholar] [CrossRef]
- Gatliff, J.; East, D.; Crosby, J.; Abeti, R.; Harvey, R.; Craigen, W.; Parker, P.; Campanella, M. Tspo interacts with vdac1 and triggers a ros-mediated inhibition of mitochondrial quality control. Autophagy 2014, 10, 2279–2296. [Google Scholar] [CrossRef]
- Garland, E.F.; Dennett, O.; Lau, L.C.; Chatelet, D.S.; Bottlaender, M.; Nicoll, J.A.R.; Boche, D. The mitochondrial protein tspo in alzheimer’s disease: Relation to the severity of ad pathology and the neuroinflammatory environment. J Neuroinflammation 2023, 20, 186. [Google Scholar] [CrossRef]
- Maeda, J.; Zhang, M.R.; Okauchi, T.; Ji, B.; Ono, M.; Hattori, S.; Kumata, K.; Iwata, N.; Saido, T.C.; Trojanowski, J.Q.; et al. In vivo positron emission tomographic imaging of glial responses to amyloid-beta and tau pathologies in mouse models of alzheimer’s disease and related disorders. J Neurosci 2011, 31, 4720–4730. [Google Scholar] [CrossRef]
- James, M.L.; Belichenko, N.P.; Nguyen, T.V.; Andrews, L.E.; Ding, Z.; Liu, H.; Bodapati, D.; Arksey, N.; Shen, B.; Cheng, Z.; et al. Pet imaging of translocator protein (18 kda) in a mouse model of alzheimer’s disease using n-(2,5-dimethoxybenzyl)-2-18f-fluoro-n-(2-phenoxyphenyl)acetamide. J Nucl Med 2015, 56, 311–316. [Google Scholar] [CrossRef]
- Ji, B.; Maeda, J.; Sawada, M.; Ono, M.; Okauchi, T.; Inaji, M.; Zhang, M.R.; Suzuki, K.; Ando, K.; Staufenbiel, M.; et al. Imaging of peripheral benzodiazepine receptor expression as biomarkers of detrimental versus beneficial glial responses in mouse models of alzheimer’s and other cns pathologies. J Neurosci 2008, 28, 12255–12267. [Google Scholar] [CrossRef]
- Mirzaei, N.; Tang, S.P.; Ashworth, S.; Coello, C.; Plisson, C.; Passchier, J.; Selvaraj, V.; Tyacke, R.J.; Nutt, D.J.; Sastre, M. In vivo imaging of microglial activation by positron emission tomography with [(11)c]pbr28 in the 5xfad model of alzheimer’s disease. Glia 2016, 64, 993–1006. [Google Scholar] [CrossRef] [PubMed]
- Serriere, S.; Tauber, C.; Vercouillie, J.; Mothes, C.; Pruckner, C.; Guilloteau, D.; Kassiou, M.; Domene, A.; Garreau, L.; Page, G.; et al. Amyloid load and translocator protein 18 kda in appsweps1-de9 mice: A longitudinal study. Neurobiol Aging 2015, 36, 1639–1652. [Google Scholar] [CrossRef] [PubMed]
- Tournier, B.B.; Tsartsalis, S.; Rigaud, D.; Fossey, C.; Cailly, T.; Fabis, F.; Pham, T.; Gregoire, M.C.; Kovari, E.; Moulin-Sallanon, M.; et al. Tspo and amyloid deposits in sub-regions of the hippocampus in the 3xtgad mouse model of alzheimer’s disease. Neurobiology of disease 2019, 121, 95–105. [Google Scholar] [CrossRef]
- Cai, Y.; Yu, X.; Hu, S.; Yu, J. A brief review on the mechanisms of mirna regulation. Genomics Proteomics Bioinformatics 2009, 7, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Lennox, A.L.; Mao, H.; Silver, D.L. Rna on the brain: Emerging layers of post-transcriptional regulation in cerebral cortex development. Wiley Interdiscip Rev Dev Biol 2018, 7. [Google Scholar] [CrossRef]
- Wang, F.; Qiang, Y.; Zhu, L.; Jiang, Y.; Wang, Y.; Shao, X.; Yin, L.; Chen, J.; Chen, Z. Microrna-7 downregulates the oncogene vdac1 to influence hepatocellular carcinoma proliferation and metastasis. Tumour Biol 2016, 37, 10235–10246. [Google Scholar] [CrossRef]
- Chaudhuri, A.D.; Choi, D.C.; Kabaria, S.; Tran, A.; Junn, E. Microrna-7 regulates the function of mitochondrial permeability transition pore by targeting vdac1 expression. J Biol Chem 2016, 291, 6483–6493. [Google Scholar] [CrossRef]
- Fatima, M.; Prajapati, B.; Saleem, K.; Kumari, R.; Mohindar Singh Singal, C.; Seth, P. Novel insights into role of mir-320a-vdac1 axis in astrocyte-mediated neuronal damage in neuroaids. Glia 2017, 65, 250–263. [Google Scholar] [CrossRef]
- Bargaje, R.; Gupta, S.; Sarkeshik, A.; Park, R.; Xu, T.; Sarkar, M.; Halimani, M.; Roy, S.S.; Yates, J.; Pillai, B. Identification of novel targets for mir-29a using mirna proteomics. PLoS One 2012, 7, e43243. [Google Scholar] [CrossRef]
- Roshan, R.; Shridhar, S.; Sarangdhar, M.A.; Banik, A.; Chawla, M.; Garg, M.; Singh, V.P.; Pillai, B. Brain-specific knockdown of mir-29 results in neuronal cell death and ataxia in mice. RNA 2014, 20, 1287–1297. [Google Scholar] [CrossRef]
- Hebert, S.S.; Horre, K.; Nicolai, L.; Papadopoulou, A.S.; Mandemakers, W.; Silahtaroglu, A.N.; Kauppinen, S.; Delacourte, A.; De Strooper, B. Loss of microrna cluster mir-29a/b-1 in sporadic alzheimer’s disease correlates with increased bace1/beta-secretase expression. Proc Natl Acad Sci U S A 2008, 105, 6415–6420. [Google Scholar] [CrossRef] [PubMed]
- Stary, C.M.; Sun, X.; Ouyang, Y.; Li, L.; Giffard, R.G. Mir-29a differentially regulates cell survival in astrocytes from cornu ammonis 1 and dentate gyrus by targeting vdac1. Mitochondrion 2016, 30, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Gregg, E.W.; Yaffe, K.; Cauley, J.A.; Rolka, D.B.; Blackwell, T.L.; Narayan, K.M.; Cummings, S.R. Is diabetes associated with cognitive impairment and cognitive decline among older women? Study of osteoporotic fractures research group. Arch Intern Med 2000, 160, 174–180. [Google Scholar] [CrossRef]
- Vieira, M.N.N.; Lima-Filho, R.A.S.; De Felice, F.G. Connecting alzheimer’s disease to diabetes: Underlying mechanisms and potential therapeutic targets. Neuropharmacology 2018, 136, 160–171. [Google Scholar] [CrossRef]
- Watson, G.S.; Craft, S. The role of insulin resistance in the pathogenesis of alzheimer’s disease: Implications for treatment. CNS Drugs 2003, 17, 27–45. [Google Scholar] [CrossRef]
- Cheng, G.; Huang, C.; Deng, H.; Wang, H. Diabetes as a risk factor for dementia and mild cognitive impairment: A meta-analysis of longitudinal studies. Intern Med J 2012, 42, 484–491. [Google Scholar] [CrossRef]
- Hsu, C.C.; Wahlqvist, M.L.; Lee, M.S.; Tsai, H.N. Incidence of dementia is increased in type 2 diabetes and reduced by the use of sulfonylureas and metformin. J Alzheimers Dis 2011, 24, 485–493. [Google Scholar] [CrossRef]
- Qiu, W.Q.; Folstein, M.F. Insulin, insulin-degrading enzyme and amyloid-beta peptide in alzheimer’s disease: Review and hypothesis. Neurobiol Aging 2006, 27, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Kickstein, E.; Krauss, S.; Thornhill, P.; Rutschow, D.; Zeller, R.; Sharkey, J.; Williamson, R.; Fuchs, M.; Kohler, A.; Glossmann, H.; et al. Biguanide metformin acts on tau phosphorylation via mtor/protein phosphatase 2a (pp2a) signaling. Proc Natl Acad Sci U S A 2010, 107, 21830–21835. [Google Scholar] [CrossRef]
- Bomfim, T.R.; Forny-Germano, L.; Sathler, L.B.; Brito-Moreira, J.; Houzel, J.C.; Decker, H.; Silverman, M.A.; Kazi, H.; Melo, H.M.; McClean, P.L.; et al. An anti-diabetes agent protects the mouse brain from defective insulin signaling caused by alzheimer’s disease- associated abeta oligomers. J Clin Invest 2012, 122, 1339–1353. [Google Scholar] [CrossRef]
- Talbot, K.; Wang, H.Y.; Kazi, H.; Han, L.Y.; Bakshi, K.P.; Stucky, A.; Fuino, R.L.; Kawaguchi, K.R.; Samoyedny, A.J.; Wilson, R.S.; et al. Demonstrated brain insulin resistance in alzheimer’s disease patients is associated with igf-1 resistance, irs-1 dysregulation, and cognitive decline. J Clin Invest 2012, 122, 1316–1338. [Google Scholar] [CrossRef] [PubMed]
- Kandimalla, R.; Thirumala, V.; Reddy, P.H. Is alzheimer’s disease a type 3 diabetes? A critical appraisal. Biochim Biophys Acta Mol Basis Dis 2017, 1863, 1078–1089. [Google Scholar] [CrossRef] [PubMed]
- Kciuk, M.; Kruczkowska, W.; Galeziewska, J.; Wanke, K.; Kaluzinska-Kolat, Z.; Aleksandrowicz, M.; Kontek, R. Alzheimer’s disease as type 3 diabetes: Understanding the link and implications. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Sandhir, R.; Gupta, S. Molecular and biochemical trajectories from diabetes to alzheimer’s disease: A critical appraisal. World J Diabetes 2015, 6, 1223–1242. [Google Scholar] [CrossRef]
- Baglietto-Vargas, D.; Shi, J.; Yaeger, D.M.; Ager, R.; LaFerla, F.M. Diabetes and alzheimer’s disease crosstalk. Neurosci Biobehav Rev 2016, 64, 272–287. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, M.; Sato, N. Bidirectional interactions between diabetes and alzheimer’s disease. Neurochem Int 2017, 108, 296–302. [Google Scholar] [CrossRef]
- Perez-Gracia, E.; Torrejon-Escribano, B.; Ferrer, I. Dystrophic neurites of senile plaques in alzheimer’s disease are deficient in cytochrome c oxidase. Acta Neuropathol 2008, 116, 261–268. [Google Scholar] [CrossRef]
- Ahmed, M.; Muhammed, S.J.; Kessler, B.; Salehi, A. Mitochondrial proteome analysis reveals altered expression of voltage dependent anion channels in pancreatic beta-cells exposed to high glucose. Islets 2010, 2, 283–292. [Google Scholar] [CrossRef]
- Sasaki, K.; Donthamsetty, R.; Heldak, M.; Cho, Y.E.; Scott, B.T.; Makino, A. Vdac: Old protein with new roles in diabetes. Am J Physiol Cell Physiol 2012, 303, C1055–C1060. [Google Scholar] [CrossRef]
- Picone, P.; Vilasi, S.; Librizzi, F.; Contardi, M.; Nuzzo, D.; Caruana, L.; Baldassano, S.; Amato, A.; Mule, F.; San Biagio, P.L.; et al. Biological and biophysics aspects of metformin-induced effects: Cortex mitochondrial dysfunction and promotion of toxic amyloid pre-fibrillar aggregates. Aging (Albany NY) 2016, 8, 1718–1734. [Google Scholar] [CrossRef]
- Tysnes, O.B.; Storstein, A. Epidemiology of parkinson’s disease. J Neural Transm (Vienna) 2017, 124, 901–905. [Google Scholar] [CrossRef] [PubMed]
- Lees, A.; Ferreira, J.J.; Rocha, J.F.; Rascol, O.; Poewe, W.; Gama, H.; Soares-da-Silva, P. Safety profile of opicapone in the management of parkinson’s disease. J Parkinsons Dis 2019, 9, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Stefani, A.; Hogl, B. Sleep in parkinson’s disease. Neuropsychopharmacology 2020, 45, 121–128. [Google Scholar] [CrossRef]
- Sveinbjornsdottir, S. The clinical symptoms of parkinson’s disease. J Neurochem 2016, 139 (Suppl. 1), 318–324. [Google Scholar] [CrossRef]
- Berg, D.; Postuma, R.B.; Adler, C.H.; Bloem, B.R.; Chan, P.; Dubois, B.; Gasser, T.; Goetz, C.G.; Halliday, G.; Joseph, L.; et al. Mds research criteria for prodromal parkinson’s disease. Mov Disord 2015, 30, 1600–1611. [Google Scholar] [CrossRef] [PubMed]
- Balestrino, R.; Schapira, A.H.V. Parkinson disease. Eur J Neurol 2020, 27, 27–42. [Google Scholar] [CrossRef]
- Raza, C.; Anjum, R.; Shakeel, N.U.A. Parkinson’s disease: Mechanisms, translational models and management strategies. Life Sci 2019, 226, 77–90. [Google Scholar] [CrossRef]
- Damier, P.; Hirsch, E.C.; Agid, Y.; Graybiel, A.M. The substantia nigra of the human brain. Ii. Patterns of loss of dopamine-containing neurons in parkinson’s disease. Brain 1999, 122 Pt 8, 1437–1448. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K. Cortico-basal ganglia-cortical circuitry in parkinson’s disease reconsidered. Exp Neurol 2008, 212, 226–229. [Google Scholar] [CrossRef]
- Oertel, W.; Schulz, J.B. Current and experimental treatments of parkinson disease: A guide for neuroscientists. J Neurochem 2016, 139 (Suppl. 1), 325–337. [Google Scholar] [CrossRef]
- Heumann, R.; Moratalla, R.; Herrero, M.T.; Chakrabarty, K.; Drucker-Colin, R.; Garcia-Montes, J.R.; Simola, N.; Morelli, M. Dyskinesia in parkinson’s disease: Mechanisms and current non-pharmacological interventions. J Neurochem 2014, 130, 472–489. [Google Scholar] [CrossRef] [PubMed]
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. Alpha-synuclein in lewy bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef] [PubMed]
- Esteves, A.R.; Arduino, D.M.; Silva, D.F.; Oliveira, C.R.; Cardoso, S.M. Mitochondrial dysfunction: The road to alpha-synuclein oligomerization in pd. Parkinsons Dis 2011, 2011, 693761. [Google Scholar] [CrossRef]
- Iadanza, M.G.; Jackson, M.P.; Hewitt, E.W.; Ranson, N.A.; Radford, S.E. A new era for understanding amyloid structures and disease. Nature reviews. Molecular cell biology 2018, 19, 755–773. [Google Scholar] [CrossRef]
- Bartels, T.; Ahlstrom, L.S.; Leftin, A.; Kamp, F.; Haass, C.; Brown, M.F.; Beyer, K. The n-terminus of the intrinsically disordered protein alpha-synuclein triggers membrane binding and helix folding. Biophys J 2010, 99, 2116–2124. [Google Scholar] [CrossRef]
- Waxman, E.A.; Mazzulli, J.R.; Giasson, B.I. Characterization of hydrophobic residue requirements for alpha-synuclein fibrillization. Biochemistry 2009, 48, 9427–9436. [Google Scholar] [CrossRef]
- Anderson, J.P.; Walker, D.E.; Goldstein, J.M.; de Laat, R.; Banducci, K.; Caccavello, R.J.; Barbour, R.; Huang, J.; Kling, K.; Lee, M.; et al. Phosphorylation of ser-129 is the dominant pathological modification of alpha-synuclein in familial and sporadic lewy body disease. J Biol Chem 2006, 281, 29739–29752. [Google Scholar] [CrossRef] [PubMed]
- Bartels, T.; Choi, J.G.; Selkoe, D.J. Alpha-synuclein occurs physiologically as a helically folded tetramer that resists aggregation. Nature 2011, 477, 107–110. [Google Scholar] [CrossRef]
- Bousset, L.; Pieri, L.; Ruiz-Arlandis, G.; Gath, J.; Jensen, P.H.; Habenstein, B.; Madiona, K.; Olieric, V.; Bockmann, A.; Meier, B.H.; et al. Structural and functional characterization of two alpha-synuclein strains. Nat Commun 2013, 4, 2575. [Google Scholar] [CrossRef]
- Angot, E.; Steiner, J.A.; Hansen, C.; Li, J.Y.; Brundin, P. Are synucleinopathies prion-like disorders? Lancet Neurol 2010, 9, 1128–1138. [Google Scholar] [CrossRef]
- Masuda-Suzukake, M.; Nonaka, T.; Hosokawa, M.; Oikawa, T.; Arai, T.; Akiyama, H.; Mann, D.M.; Hasegawa, M. Prion-like spreading of pathological alpha-synuclein in brain. Brain 2013, 136, 1128–1138. [Google Scholar] [CrossRef] [PubMed]
- Fusco, G.; Chen, S.W.; Williamson, P.T.F.; Cascella, R.; Perni, M.; Jarvis, J.A.; Cecchi, C.; Vendruscolo, M.; Chiti, F.; Cremades, N.; et al. Structural basis of membrane disruption and cellular toxicity by alpha-synuclein oligomers. Science 2017, 358, 1440–1443. [Google Scholar] [CrossRef] [PubMed]
- Deas, E.; Cremades, N.; Angelova, P.R.; Ludtmann, M.H.; Yao, Z.; Chen, S.; Horrocks, M.H.; Banushi, B.; Little, D.; Devine, M.J.; et al. Alpha-synuclein oligomers interact with metal ions to induce oxidative stress and neuronal death in parkinson’s disease. Antioxid Redox Signal 2016, 24, 376–391. [Google Scholar] [CrossRef] [PubMed]
- Danzer, K.M.; Haasen, D.; Karow, A.R.; Moussaud, S.; Habeck, M.; Giese, A.; Kretzschmar, H.; Hengerer, B.; Kostka, M. Different species of alpha-synuclein oligomers induce calcium influx and seeding. J Neurosci 2007, 27, 9220–9232. [Google Scholar] [CrossRef]
- Luth, E.S.; Stavrovskaya, I.G.; Bartels, T.; Kristal, B.S.; Selkoe, D.J. Soluble, prefibrillar alpha-synuclein oligomers promote complex i-dependent, ca2+-induced mitochondrial dysfunction. J Biol Chem 2014, 289, 21490–21507. [Google Scholar] [CrossRef]
- Rostovtseva, T.K.; Gurnev, P.A.; Protchenko, O.; Hoogerheide, D.P.; Yap, T.L.; Philpott, C.C.; Lee, J.C.; Bezrukov, S.M. Alpha-synuclein shows high affinity interaction with voltage-dependent anion channel, suggesting mechanisms of mitochondrial regulation and toxicity in parkinson disease. J Biol Chem 2015, 290, 18467–18477. [Google Scholar] [CrossRef]
- Devi, L.; Raghavendran, V.; Prabhu, B.M.; Avadhani, N.G.; Anandatheerthavarada, H.K. Mitochondrial import and accumulation of alpha-synuclein impair complex i in human dopaminergic neuronal cultures and parkinson disease brain. J Biol Chem 2008, 283, 9089–9100. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, C.; Yin, J.; Li, X.; Cheng, F.; Li, Y.; Yang, H.; Ueda, K.; Chan, P.; Yu, S. Alpha-synuclein is differentially expressed in mitochondria from different rat brain regions and dose-dependently down-regulates complex i activity. Neurosci Lett 2009, 454, 187–192. [Google Scholar] [CrossRef]
- Ellis, C.E.; Murphy, E.J.; Mitchell, D.C.; Golovko, M.Y.; Scaglia, F.; Barcelo-Coblijn, G.C.; Nussbaum, R.L. Mitochondrial lipid abnormality and electron transport chain impairment in mice lacking alpha-synuclein. Mol Cell Biol 2005, 25, 10190–10201. [Google Scholar] [CrossRef]
- Elkon, H.; Don, J.; Melamed, E.; Ziv, I.; Shirvan, A.; Offen, D. Mutant and wild-type alpha-synuclein interact with mitochondrial cytochrome c oxidase. J Mol Neurosci 2002, 18, 229–238. [Google Scholar] [CrossRef]
- Nakamura, K. Alpha-synuclein and mitochondria: Partners in crime? Neurotherapeutics 2013, 10, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Hoogerheide, D.P.; Rostovtseva, T.K.; Bezrukov, S.M. Exploring lipid-dependent conformations of membrane-bound alpha-synuclein with the vdac nanopore. Biochim Biophys Acta Biomembr 2021, 1863, 183643. [Google Scholar] [CrossRef] [PubMed]
- Rosencrans, W.M.; Aguilella, V.M.; Rostovtseva, T.K.; Bezrukov, S.M. Alpha-synuclein emerges as a potent regulator of vdac-facilitated calcium transport. Cell Calcium 2021, 95, 102355. [Google Scholar] [CrossRef]
- Queralt-Martin, M.; Bergdoll, L.; Teijido, O.; Munshi, N.; Jacobs, D.; Kuszak, A.J.; Protchenko, O.; Reina, S.; Magri, A.; De Pinto, V.; et al. A lower affinity to cytosolic proteins reveals vdac3 isoform-specific role in mitochondrial biology. J Gen Physiol 2020, 152. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Zhang, C.; Cai, Q.; Lu, Q.; Duan, C.; Zhu, Y.; Yang, H. Voltage-dependent anion channel involved in the alpha-synuclein-induced dopaminergic neuron toxicity in rats. Acta Biochim Biophys Sin (Shanghai) 2013, 45, 170–178. [Google Scholar] [CrossRef]
- Martin, L.J.; Semenkow, S.; Hanaford, A.; Wong, M. Mitochondrial permeability transition pore regulates parkinson’s disease development in mutant alpha-synuclein transgenic mice. Neurobiol Aging 2014, 35, 1132–1152. [Google Scholar] [CrossRef]
- Chu, Y.; Goldman, J.G.; Kelly, L.; He, Y.; Waliczek, T.; Kordower, J.H. Abnormal alpha-synuclein reduces nigral voltage-dependent anion channel 1 in sporadic and experimental parkinson’s disease. Neurobiology of disease 2014, 69, 1–14. [Google Scholar] [CrossRef]
- Alberio, T.; Mammucari, C.; D’Agostino, G.; Rizzuto, R.; Fasano, M. Altered dopamine homeostasis differentially affects mitochondrial voltage-dependent anion channels turnover. Biochim Biophys Acta 2014, 1842, 1816–1822. [Google Scholar] [CrossRef]
- Premkumar, A.; Simantov, R. Mitochondrial voltage-dependent anion channel is involved in dopamine-induced apoptosis. J Neurochem 2002, 82, 345–352. [Google Scholar] [CrossRef]
- Xiong, Y.; Ding, H.; Xu, M.; Gao, J. Protective effects of asiatic acid on rotenone- or h2o2-induced injury in sh-sy5y cells. Neurochem Res 2009, 34, 746–754. [Google Scholar] [CrossRef]
- Burte, F.; De Girolamo, L.A.; Hargreaves, A.J.; Billett, E.E. Alterations in the mitochondrial proteome of neuroblastoma cells in response to complex 1 inhibition. J Proteome Res 2011, 10, 1974–1986. [Google Scholar] [CrossRef] [PubMed]
- Magalingam, K.B.; Somanath, S.D.; Ramdas, P.; Haleagrahara, N.; Radhakrishnan, A.K. 6-hydroxydopamine induces neurodegeneration in terminally differentiated sh-sy5y neuroblastoma cells via enrichment of the nucleosomal degradation pathway: A global proteomics approach. J Mol Neurosci 2022, 72, 1026–1046. [Google Scholar] [CrossRef]
- Araujo de Lima, L.; Oliveira Cunha, P.L.; Felicio Calou, I.B.; Tavares Neves, K.R.; Facundo, H.T.; Socorro de Barros Viana, G. Effects of vitamin d (vd3) supplementation on the brain mitochondrial function of male rats, in the 6-ohda-induced model of parkinson’s disease. Neurochem Int 2022, 154, 105280. [Google Scholar] [CrossRef] [PubMed]
- Junn, E.; Lee, K.W.; Jeong, B.S.; Chan, T.W.; Im, J.Y.; Mouradian, M.M. Repression of alpha-synuclein expression and toxicity by microrna-7. Proc Natl Acad Sci U S A 2009, 106, 13052–13057. [Google Scholar] [CrossRef] [PubMed]
- Leggio, L.; Vivarelli, S.; L’Episcopo, F.; Tirolo, C.; Caniglia, S.; Testa, N.; Marchetti, B.; Iraci, N. Micrornas in parkinson’s disease: From pathogenesis to novel diagnostic and therapeutic approaches. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef]
- McMillan, K.J.; Murray, T.K.; Bengoa-Vergniory, N.; Cordero-Llana, O.; Cooper, J.; Buckley, A.; Wade-Martins, R.; Uney, J.B.; O’Neill, M.J.; Wong, L.F.; et al. Loss of microrna-7 regulation leads to alpha-synuclein accumulation and dopaminergic neuronal loss in vivo. Mol Ther 2017, 25, 2404–2414. [Google Scholar] [CrossRef]
- Rovini, A.; Gurnev, P.A.; Beilina, A.; Queralt-Martin, M.; Rosencrans, W.; Cookson, M.R.; Bezrukov, S.M.; Rostovtseva, T.K. Molecular mechanism of olesoxime-mediated neuroprotection through targeting alpha-synuclein interaction with mitochondrial vdac. Cell Mol Life Sci 2020, 77, 3611–3626. [Google Scholar] [CrossRef]
- Bordet, T.; Buisson, B.; Michaud, M.; Drouot, C.; Galea, P.; Delaage, P.; Akentieva, N.P.; Evers, A.S.; Covey, D.F.; Ostuni, M.A.; et al. Identification and characterization of cholest-4-en-3-one, oxime (tro19622), a novel drug candidate for amyotrophic lateral sclerosis. J Pharmacol Exp Ther 2007, 322, 709–720. [Google Scholar] [CrossRef]
- Rajendran, M.; Queralt-Martin, M.; Gurnev, P.A.; Rosencrans, W.M.; Rovini, A.; Jacobs, D.; Abrantes, K.; Hoogerheide, D.P.; Bezrukov, S.M.; Rostovtseva, T.K. Restricting alpha-synuclein transport into mitochondria by inhibition of alpha-synuclein-vdac complexation as a potential therapeutic target for parkinson’s disease treatment. Cell Mol Life Sci 2022, 79, 368. [Google Scholar] [CrossRef]
- Feng, S.; Gui, J.; Qin, B.; Ye, J.; Zhao, Q.; Guo, A.; Sang, M.; Sun, X. Resveratrol inhibits vdac1-mediated mitochondrial dysfunction to mitigate pathological progression in parkinson’s disease model. Mol Neurobiol 2024. [Google Scholar] [CrossRef]
- Iaccarino, C.; Crosio, C.; Vitale, C.; Sanna, G.; Carri, M.T.; Barone, P. Apoptotic mechanisms in mutant lrrk2-mediated cell death. Hum Mol Genet 2007, 16, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Yu, M.; Niu, J.; Yue, Z.; Xu, Z. Expression of leucine-rich repeat kinase 2 (lrrk2) inhibits the processing of umtck to induce cell death in a cell culture model system. Biosci Rep 2011, 31, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Kitada, T.; Asakawa, S.; Hattori, N.; Matsumine, H.; Yamamura, Y.; Minoshima, S.; Yokochi, M.; Mizuno, Y.; Shimizu, N. Mutations in the parkin gene cause autosomal recessive juvenile parkinsonism. Nature 1998, 392, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Shimura, H.; Hattori, N.; Kubo, S.; Mizuno, Y.; Asakawa, S.; Minoshima, S.; Shimizu, N.; Iwai, K.; Chiba, T.; Tanaka, K.; et al. Familial parkinson disease gene product, parkin, is a ubiquitin-protein ligase. Nat Genet 2000, 25, 302–305. [Google Scholar] [CrossRef]
- Rogaeva, E.; Johnson, J.; Lang, A.E.; Gulick, C.; Gwinn-Hardy, K.; Kawarai, T.; Sato, C.; Morgan, A.; Werner, J.; Nussbaum, R.; et al. Analysis of the pink1 gene in a large cohort of cases with parkinson disease. Arch Neurol 2004, 61, 1898–1904. [Google Scholar] [CrossRef]
- Valente, E.M.; Abou-Sleiman, P.M.; Caputo, V.; Muqit, M.M.; Harvey, K.; Gispert, S.; Ali, Z.; Del Turco, D.; Bentivoglio, A.R.; Healy, D.G.; et al. Hereditary early-onset parkinson’s disease caused by mutations in pink1. Science 2004, 304, 1158–1160. [Google Scholar] [CrossRef]
- Pickrell, A.M.; Youle, R.J. The roles of pink1, parkin, and mitochondrial fidelity in parkinson’s disease. Neuron 2015, 85, 257–273. [Google Scholar] [CrossRef]
- Sun, K.; Jing, X.; Guo, J.; Yao, X.; Guo, F. Mitophagy in degenerative joint diseases. Autophagy 2021, 17, 2082–2092. [Google Scholar] [CrossRef]
- Geisler, S.; Holmstrom, K.M.; Skujat, D.; Fiesel, F.C.; Rothfuss, O.C.; Kahle, P.J.; Springer, W. Pink1/parkin-mediated mitophagy is dependent on vdac1 and p62/sqstm1. Nat Cell Biol 2010, 12, 119–131. [Google Scholar] [CrossRef]
- Callegari, S.; Kirk, N.S.; Gan, Z.Y.; Dite, T.; Cobbold, S.A.; Leis, A.; Dagley, L.F.; Glukhova, A.; Komander, D. Structure of human pink1 at a mitochondrial tom-vdac array. Science 2025, eadu6445. [Google Scholar] [CrossRef]
- Ham, S.J.; Lee, D.; Yoo, H.; Jun, K.; Shin, H.; Chung, J. Decision between mitophagy and apoptosis by parkin via vdac1 ubiquitination. Proc Natl Acad Sci U S A 2020, 117, 4281–4291. [Google Scholar] [CrossRef] [PubMed]
- Cui, T.; Fan, C.; Gu, L.; Gao, H.; Liu, Q.; Zhang, T.; Qi, Z.; Zhao, C.; Zhao, H.; Cai, Q.; et al. Silencing of pink1 induces mitophagy via mitochondrial permeability transition in dopaminergic mn9d cells. Brain Res 2011, 1394, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Sun, W.; Shen, M.; Zhang, Y.; Jiang, M.; Liu, A.; Ma, H.; Lai, X.; Wu, J. Idebenone improves motor dysfunction, learning and memory by regulating mitophagy in mptp-treated mice. Cell Death Discov 2022, 8, 28. [Google Scholar] [CrossRef]
- van der Merwe, C.; Jalali Sefid Dashti, Z.; Christoffels, A.; Loos, B.; Bardien, S. Evidence for a common biological pathway linking three parkinson’s disease-causing genes: Parkin, pink1 and dj-1. Eur J Neurosci 2015, 41, 1113–1125. [Google Scholar] [CrossRef] [PubMed]
- Ottolini, D.; Cali, T.; Negro, A.; Brini, M. The parkinson disease-related protein dj-1 counteracts mitochondrial impairment induced by the tumour suppressor protein p53 by enhancing endoplasmic reticulum-mitochondria tethering. Hum Mol Genet 2013, 22, 2152–2168. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, X.; Fujioka, H.; Liu, J.; Chen, S.; Zhu, X. Dj-1 regulates the integrity and function of er-mitochondria association through interaction with ip3r3-grp75-vdac1. Proc Natl Acad Sci U S A 2019, 116, 25322–25328. [Google Scholar] [CrossRef]
- Basso, V.; Marchesan, E.; Ziviani, E. A trio has turned into a quartet: Dj-1 interacts with the ip3r-grp75-vdac complex to control er-mitochondria interaction. Cell Calcium 2020, 87, 102186. [Google Scholar] [CrossRef]
- Cleveland, D.W.; Rothstein, J.D. From charcot to lou gehrig: Deciphering selective motor neuron death in als. Nat Rev Neurosci 2001, 2, 806–819. [Google Scholar] [CrossRef]
- Mulder, D.W.; Kurland, L.T.; Offord, K.P.; Beard, C.M. Familial adult motor neuron disease: Amyotrophic lateral sclerosis. Neurology 1986, 36, 511–517. [Google Scholar] [CrossRef]
- Gibbons, C.; Pagnini, F.; Friede, T.; Young, C.A. Treatment of fatigue in amyotrophic lateral sclerosis/motor neuron disease. Cochrane Database Syst Rev 2018, 1, CD011005. [Google Scholar] [CrossRef]
- Wang, H.; Guan, L.; Deng, M. Recent progress of the genetics of amyotrophic lateral sclerosis and challenges of gene therapy. Front Neurosci 2023, 17, 1170996. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, L.; Anakor, E.; Connolly, O.; Vijayakumar, U.G.; Duddy, W.J.; Duguez, S. Molecular and cellular mechanisms affected in als. J Pers Med 2020, 10. [Google Scholar] [CrossRef]
- Rosen, D.R.; Siddique, T.; Patterson, D.; Figlewicz, D.A.; Sapp, P.; Hentati, A.; Donaldson, D.; Goto, J.; O’Regan, J.P.; Deng, H.X.; et al. Mutations in cu/zn superoxide dismutase gene are associated with familial amyotrophic lateral sclerosis. Nature 1993, 362, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Sanghai, N.; Tranmer, G.K. Hydrogen peroxide and amyotrophic lateral sclerosis: From biochemistry to pathophysiology. Antioxidants (Basel) 2021, 11. [Google Scholar] [CrossRef]
- Abu-Hamad, S.; Kahn, J.; Leyton-Jaimes, M.F.; Rosenblatt, J.; Israelson, A. Misfolded sod1 accumulation and mitochondrial association contribute to the selective vulnerability of motor neurons in familial als: Correlation to human disease. ACS Chem Neurosci 2017, 8, 2225–2234. [Google Scholar] [CrossRef] [PubMed]
- Israelson, A.; Arbel, N.; Da Cruz, S.; Ilieva, H.; Yamanaka, K.; Shoshan-Barmatz, V.; Cleveland, D.W. Misfolded mutant sod1 directly inhibits vdac1 conductance in a mouse model of inherited als. Neuron 2010, 67, 575–587. [Google Scholar] [CrossRef]
- Li, Q.; Vande Velde, C.; Israelson, A.; Xie, J.; Bailey, A.O.; Dong, M.Q.; Chun, S.J.; Roy, T.; Winer, L.; Yates, J.R.; et al. Als-linked mutant superoxide dismutase 1 (sod1) alters mitochondrial protein composition and decreases protein import. Proc Natl Acad Sci U S A 2010, 107, 21146–21151. [Google Scholar] [CrossRef]
- Shteinfer-Kuzmine, A.; Argueti-Ostrovsky, S.; Leyton-Jaimes, M.F.; Anand, U.; Abu-Hamad, S.; Zalk, R.; Shoshan-Barmatz, V.; Israelson, A. Targeting the mitochondrial protein vdac1 as a potential therapeutic strategy in als. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Tan, W.; Naniche, N.; Bogush, A.; Pedrini, S.; Trotti, D.; Pasinelli, P. Small peptides against the mutant sod1/bcl-2 toxic mitochondrial complex restore mitochondrial function and cell viability in mutant sod1-mediated als. J Neurosci 2013, 33, 11588–11598. [Google Scholar] [CrossRef]
- Pedrini, S.; Sau, D.; Guareschi, S.; Bogush, M.; Brown, R.H., Jr.; Naniche, N.; Kia, A.; Trotti, D.; Pasinelli, P. Als-linked mutant sod1 damages mitochondria by promoting conformational changes in bcl-2. Hum Mol Genet 2010, 19, 2974–2986. [Google Scholar] [CrossRef]
- Shteinfer-Kuzmine, A.; Argueti, S.; Gupta, R.; Shvil, N.; Abu-Hamad, S.; Gropper, Y.; Hoeber, J.; Magri, A.; Messina, A.; Kozlova, E.N.; et al. A vdac1-derived n-terminal peptide inhibits mutant sod1-vdac1 interactions and toxicity in the sod1 model of als. Front Cell Neurosci 2019, 13, 346. [Google Scholar] [CrossRef] [PubMed]
- Magri, A.; Belfiore, R.; Reina, S.; Tomasello, M.F.; Di Rosa, M.C.; Guarino, F.; Leggio, L.; De Pinto, V.; Messina, A. Hexokinase i n-terminal based peptide prevents the vdac1-sod1 g93a interaction and re-establishes als cell viability. Sci Rep 2016, 6, 34802. [Google Scholar] [CrossRef] [PubMed]
- Magri, A.; Risiglione, P.; Caccamo, A.; Formicola, B.; Tomasello, M.F.; Arrigoni, C.; Zimbone, S.; Guarino, F.; Re, F.; Messina, A. Small hexokinase 1 peptide against toxic sod1 g93a mitochondrial accumulation in als rescues the atp-related respiration. Biomedicines 2021, 9. [Google Scholar] [CrossRef]
- Fukada, K.; Zhang, F.; Vien, A.; Cashman, N.R.; Zhu, H. Mitochondrial proteomic analysis of a cell line model of familial amyotrophic lateral sclerosis. Mol Cell Proteomics 2004, 3, 1211–1223. [Google Scholar] [CrossRef]
- Pittala, M.G.G.; Reina, S.; Cubisino, S.A.M.; Cucina, A.; Formicola, B.; Cunsolo, V.; Foti, S.; Saletti, R.; Messina, A. Post-translational modification analysis of vdac1 in als-sod1 model cells reveals specific asparagine and glutamine deamidation. Antioxidants (Basel) 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Pittala, M.G.G.; Reina, S.; Nibali, S.C.; Cucina, A.; Cubisino, S.A.M.; Cunsolo, V.; Amodeo, G.F.; Foti, S.; De Pinto, V.; Saletti, R.; et al. Specific post-translational modifications of vdac3 in als-sod1 model cells identified by high-resolution mass spectrometry. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Sunyach, C.; Michaud, M.; Arnoux, T.; Bernard-Marissal, N.; Aebischer, J.; Latyszenok, V.; Gouarne, C.; Raoul, C.; Pruss, R.M.; Bordet, T.; et al. Olesoxime delays muscle denervation, astrogliosis, microglial activation and motoneuron death in an als mouse model. Neuropharmacology 2012, 62, 2346–2352. [Google Scholar] [CrossRef]
- Lenglet, T.; Lacomblez, L.; Abitbol, J.L.; Ludolph, A.; Mora, J.S.; Robberecht, W.; Shaw, P.J.; Pruss, R.M.; Cuvier, V.; Meininger, V.; et al. A phase ii-iii trial of olesoxime in subjects with amyotrophic lateral sclerosis. Eur J Neurol 2014, 21, 529–536. [Google Scholar] [CrossRef]
- Magri, A.; Lipari, C.L.R.; Caccamo, A.; Battiato, G.; Conti Nibali, S.; De Pinto, V.; Guarino, F.; Messina, A. Aav-mediated upregulation of vdac1 rescues the mitochondrial respiration and sirtuins expression in a sod1 mouse model of inherited als. Cell Death Discov 2024, 10, 178. [Google Scholar] [CrossRef]
- Pappalardo, X.G.; Jansen, G.; Amaradio, M.; Costanza, J.; Umeton, R.; Guarino, F.; De Pinto, V.; Oliver, S.G.; Messina, A.; Nicosia, G. Inferring gene regulatory networks of als from blood transcriptome profiles. Heliyon 2024, 10, e40696. [Google Scholar] [CrossRef]
- Yu, C.H.; Davidson, S.; Harapas, C.R.; Hilton, J.B.; Mlodzianoski, M.J.; Laohamonthonkul, P.; Louis, C.; Low, R.R.J.; Moecking, J.; De Nardo, D.; et al. Tdp-43 triggers mitochondrial DNA release via mptp to activate cgas/sting in als. Cell 2020, 183, 636–649.e618. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.A.; Itaman, S.; Khalid-Janney, C.M.; Sherard, J.A.; Dowell, J.A.; Cairns, N.J.; Gitcho, M.A. Tdp-43 interacts with mitochondrial proteins critical for mitophagy and mitochondrial dynamics. Neurosci Lett 2018, 678, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Pilotto, F.; Schmitz, A.; Maharjan, N.; Diab, R.; Odriozola, A.; Tripathi, P.; Yamoah, A.; Scheidegger, O.; Oestmann, A.; Dennys, C.N.; et al. Polyga targets the er stress-adaptive response by impairing grp75 function at the mam in c9orf72-als/ftd. Acta Neuropathol 2022, 144, 939–966. [Google Scholar] [CrossRef]
- Illarioshkin, S.N.; Klyushnikov, S.A.; Vigont, V.A.; Seliverstov, Y.A.; Kaznacheyeva, E.V. Molecular pathogenesis in huntington’s disease. Biochemistry (Mosc) 2018, 83, 1030–1039. [Google Scholar] [CrossRef]
- Tabrizi, S.J.; Estevez-Fraga, C.; van Roon-Mom, W.M.C.; Flower, M.D.; Scahill, R.I.; Wild, E.J.; Munoz-Sanjuan, I.; Sampaio, C.; Rosser, A.E.; Leavitt, B.R. Potential disease-modifying therapies for huntington’s disease: Lessons learned and future opportunities. Lancet Neurol 2022, 21, 645–658. [Google Scholar] [CrossRef]
- Zielonka, D. Factors contributing to clinical picture and progression of huntington’s disease. Neural Regen Res 2018, 13, 1364–1365. [Google Scholar] [CrossRef]
- Jurcau, A. Molecular pathophysiological mechanisms in huntington’s disease. Biomedicines 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Clemens, L.E.; Weber, J.J.; Wlodkowski, T.T.; Yu-Taeger, L.; Michaud, M.; Calaminus, C.; Eckert, S.H.; Gaca, J.; Weiss, A.; Magg, J.C.; et al. Olesoxime suppresses calpain activation and mutant huntingtin fragmentation in the bachd rat. Brain 2015, 138, 3632–3653. [Google Scholar] [CrossRef]
- Lou, S.; Lepak, V.C.; Eberly, L.E.; Roth, B.; Cui, W.; Zhu, X.H.; Oz, G.; Dubinsky, J.M. Oxygen consumption deficit in huntington disease mouse brain under metabolic stress. Hum Mol Genet 2016, 25, 2813–2826. [Google Scholar] [CrossRef]
- Napoli, E.; Wong, S.; Hung, C.; Ross-Inta, C.; Bomdica, P.; Giulivi, C. Defective mitochondrial disulfide relay system, altered mitochondrial morphology and function in huntington’s disease. Hum Mol Genet 2013, 22, 989–1004. [Google Scholar] [CrossRef]
- Perluigi, M.; Poon, H.F.; Maragos, W.; Pierce, W.M.; Klein, J.B.; Calabrese, V.; Cini, C.; De Marco, C.; Butterfield, D.A. Proteomic analysis of protein expression and oxidative modification in r6/2 transgenic mice: A model of huntington disease. Mol Cell Proteomics 2005, 4, 1849–1861. [Google Scholar] [CrossRef] [PubMed]
- Karachitos, A.; Grobys, D.; Kulczynska, K.; Sobusiak, A.; Kmita, H. The association of vdac with cell viability of pc12 model of huntington’s disease. Front Oncol 2016, 6, 238. [Google Scholar] [CrossRef] [PubMed]
- Beatriz, M.; Vilaca, R.; Anjo, S.I.; Manadas, B.; Januario, C.; Rego, A.C.; Lopes, C. Defective mitochondria-lysosomal axis enhances the release of extracellular vesicles containing mitochondrial DNA and proteins in huntington’s disease. J Extracell Biol 2022, 1, e65. [Google Scholar] [CrossRef] [PubMed]
- Brondani, M.; Roginski, A.C.; Ribeiro, R.T.; de Medeiros, M.P.; Hoffmann, C.I.H.; Wajner, M.; Leipnitz, G.; Seminotti, B. Mitochondrial dysfunction, oxidative stress, er stress and mitochondria-er crosstalk alterations in a chemical rat model of huntington’s disease: Potential benefits of bezafibrate. Toxicol Lett 2023, 381, 48–59. [Google Scholar] [CrossRef]
- El-Emam, M.A.; Sheta, E.; El-Abhar, H.S.; Abdallah, D.M.; El Kerdawy, A.M.; Eldehna, W.M.; Gowayed, M.A. Morin suppresses mtorc1/ire-1alpha/jnk and ip3r-vdac-1 pathways: Crucial mechanisms in apoptosis and mitophagy inhibition in experimental huntington’s disease, supported by in silico molecular docking simulations. Life Sci 2024, 338, 122362. [Google Scholar] [CrossRef]
- Zhang, Y.; Du, B.; Zou, M.; Peng, B.; Rao, Y. Neuronal ceroid lipofuscinosis-concepts, classification, and avenues for therapy. CNS Neurosci Ther 2025, 31, e70261. [Google Scholar] [CrossRef]
- Kielar, C.; Wishart, T.M.; Palmer, A.; Dihanich, S.; Wong, A.M.; Macauley, S.L.; Chan, C.H.; Sands, M.S.; Pearce, D.A.; Cooper, J.D.; et al. Molecular correlates of axonal and synaptic pathology in mouse models of batten disease. Hum Mol Genet 2009, 18, 4066–4080. [Google Scholar] [CrossRef]
- Homewood, J.; Bond, N.W. Thiamin deficiency and korsakoff’s syndrome: Failure to find memory impairments following nonalcoholic wernicke’s encephalopathy. Alcohol 1999, 19, 75–84. [Google Scholar] [CrossRef]
- Hazell, A.S.; Butterworth, R.F. Update of cell damage mechanisms in thiamine deficiency: Focus on oxidative stress, excitotoxicity and inflammation. Alcohol Alcohol 2009, 44, 141–147. [Google Scholar] [CrossRef]
- Bueno, K.O.; de Souza Resende, L.; Ribeiro, A.F.; Dos Santos, D.M.; Goncalves, E.C.; Vigil, F.A.; de Oliveira Silva, I.F.; Ferreira, L.F.; de Castro Pimenta, A.M.; Ribeiro, A.M. Spatial cognitive deficits in an animal model of wernicke-korsakoff syndrome are related to changes in thalamic vdac protein concentrations. Neuroscience 2015, 294, 29–37. [Google Scholar] [CrossRef]
- Mishra, E.; Thakur, M.K. Alterations in hippocampal mitochondrial dynamics are associated with neurodegeneration and recognition memory decline in old male mice. Biogerontology 2022, 23, 251–271. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic outline for the involvement of pl-VDAC-1 in the neuroprotection in a mouse model expressing human V12-Ha-RAS in postmitotic neurons. Ha-RAS is anchored to the cytoplasmic membrane by two palmitoyl (black) and one farnesyl (gray) anchors. Guanine–nucleotide exchange factors (GEF) activate RAS by the exchange of GDP to GTP. V12-Ha-RAS is locked in its GTP bound state because it cannot be inactivated by GTPase-activating proteins (GAP) due to the V12 mutation. V12-Ha-RAS then permanently activates the mitogen activated protein kinase (MAPK) signal transduction via rapidly activated fibrosarcoma (RAF), MAPK/ERK kinase (MEK) and extracellular-signal-regulated-kinase (ERK). MEK promotes BRAF activation through allosteric control of KSR proteins, a kinase suppressor of RAS (KSR) serving as molecular scaffold for effective signalling. Phosphorylations are indicated by yellow circles with Pi. MEK can be inhibited by U0126 resulting in a blockage of the downstream signalling. In the nucleus, a dimer of activated ERK stimulates the transcription of certain genes. The VDAC-1 mRNA becomes alternatively spliced into pl-VDAC-1 mRNA and mt-VDAC-1 mRNA. The level of pl-VDAC-1 mRNA is selectively decreased by the activation of the RAS/MAPK pathway in comparison to wild type mice. In mouse, the pl-VDAC-1 requires a signal peptide guiding the VDAC-1 to the plasmalemmal membrane, this sequence is schematically outlined in red in the pl-VDAC-1 mRNA. Upon translation, the mt-VDAC-1 is inserted into the OMM while the pl-VDAC-1 is passed by the endoplasmic reticulum and the Golgi apparatus and directly inserted into the cell membrane. In this mouse model, a reduced expression of pl-VDAC-1 causes neuroprotection which is phenocopied by the extracellular application of an anti-VDAC-1 antibody resulting in channel blocking. Mt-VDAC-1 and pl-VDAC-1 are shown in their three-dimensional protein structure (3EMN; [17]). This figure was created in https://BioRender.com (accessed 15th May 2025) in modification from [109].
Figure 1.
Schematic outline for the involvement of pl-VDAC-1 in the neuroprotection in a mouse model expressing human V12-Ha-RAS in postmitotic neurons. Ha-RAS is anchored to the cytoplasmic membrane by two palmitoyl (black) and one farnesyl (gray) anchors. Guanine–nucleotide exchange factors (GEF) activate RAS by the exchange of GDP to GTP. V12-Ha-RAS is locked in its GTP bound state because it cannot be inactivated by GTPase-activating proteins (GAP) due to the V12 mutation. V12-Ha-RAS then permanently activates the mitogen activated protein kinase (MAPK) signal transduction via rapidly activated fibrosarcoma (RAF), MAPK/ERK kinase (MEK) and extracellular-signal-regulated-kinase (ERK). MEK promotes BRAF activation through allosteric control of KSR proteins, a kinase suppressor of RAS (KSR) serving as molecular scaffold for effective signalling. Phosphorylations are indicated by yellow circles with Pi. MEK can be inhibited by U0126 resulting in a blockage of the downstream signalling. In the nucleus, a dimer of activated ERK stimulates the transcription of certain genes. The VDAC-1 mRNA becomes alternatively spliced into pl-VDAC-1 mRNA and mt-VDAC-1 mRNA. The level of pl-VDAC-1 mRNA is selectively decreased by the activation of the RAS/MAPK pathway in comparison to wild type mice. In mouse, the pl-VDAC-1 requires a signal peptide guiding the VDAC-1 to the plasmalemmal membrane, this sequence is schematically outlined in red in the pl-VDAC-1 mRNA. Upon translation, the mt-VDAC-1 is inserted into the OMM while the pl-VDAC-1 is passed by the endoplasmic reticulum and the Golgi apparatus and directly inserted into the cell membrane. In this mouse model, a reduced expression of pl-VDAC-1 causes neuroprotection which is phenocopied by the extracellular application of an anti-VDAC-1 antibody resulting in channel blocking. Mt-VDAC-1 and pl-VDAC-1 are shown in their three-dimensional protein structure (3EMN; [17]). This figure was created in https://BioRender.com (accessed 15th May 2025) in modification from [109].
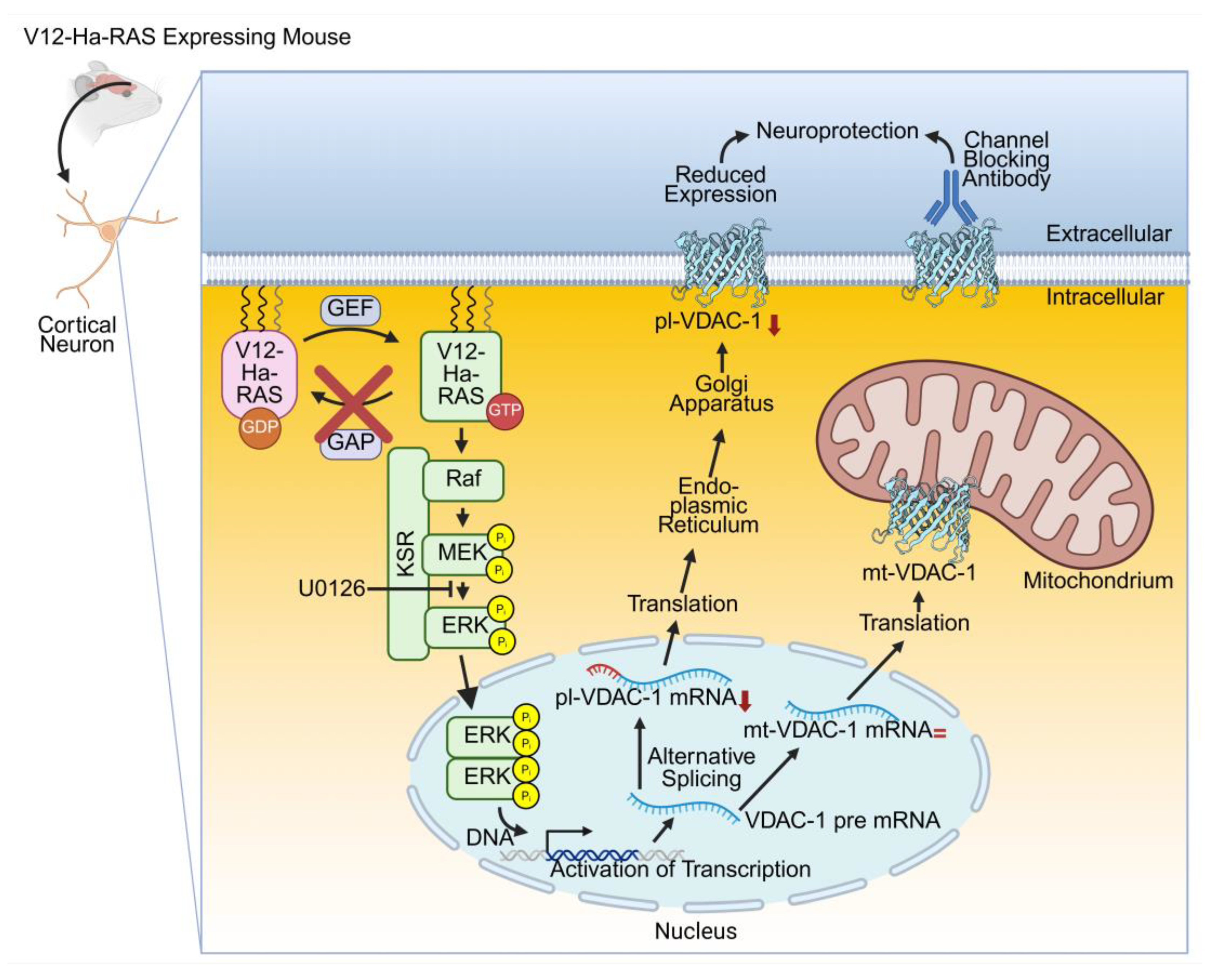
Figure 2.
Schematic outline for the role of VDAC-1 in AD. Extracellular Aβ interacts via the GXXXG motif with the N-terminus of pl-VDAC-1 by electrostatic interactions as Aβ provides negative charges and the N-terminus has positive charges. A hetero-oligomer of pl-VDAC-1 and Aβ is formed allowing the entrance of Aβ into the cytosol. Intracellularly, mt-VDAC-1 interacts with Aβ forming a hetero-oligomer of mt-VDAC-1 with Aβ enabling the release of Cytochrome c resulting in apoptosis. Aβ can induce the detachment of HK from mt-VDAC-1. Furthermore, GSK3β can phosphorylates mt-VDAC-1 and thereby inhibits the binding of HK to mt-VDAC-1. In addition, phosphorylated Tau can interact with mt-VDAC-1 leading to the closure of the channel with the consequence that there is no further transport of nucleotides, metabolites, lipids, ions etc. Additionally, mt-VDAC-1 and TSPO form a complex contributing to ROS. This figure was created in https://BioRender.com (accessed 15th May 2025).
Figure 2.
Schematic outline for the role of VDAC-1 in AD. Extracellular Aβ interacts via the GXXXG motif with the N-terminus of pl-VDAC-1 by electrostatic interactions as Aβ provides negative charges and the N-terminus has positive charges. A hetero-oligomer of pl-VDAC-1 and Aβ is formed allowing the entrance of Aβ into the cytosol. Intracellularly, mt-VDAC-1 interacts with Aβ forming a hetero-oligomer of mt-VDAC-1 with Aβ enabling the release of Cytochrome c resulting in apoptosis. Aβ can induce the detachment of HK from mt-VDAC-1. Furthermore, GSK3β can phosphorylates mt-VDAC-1 and thereby inhibits the binding of HK to mt-VDAC-1. In addition, phosphorylated Tau can interact with mt-VDAC-1 leading to the closure of the channel with the consequence that there is no further transport of nucleotides, metabolites, lipids, ions etc. Additionally, mt-VDAC-1 and TSPO form a complex contributing to ROS. This figure was created in https://BioRender.com (accessed 15th May 2025).
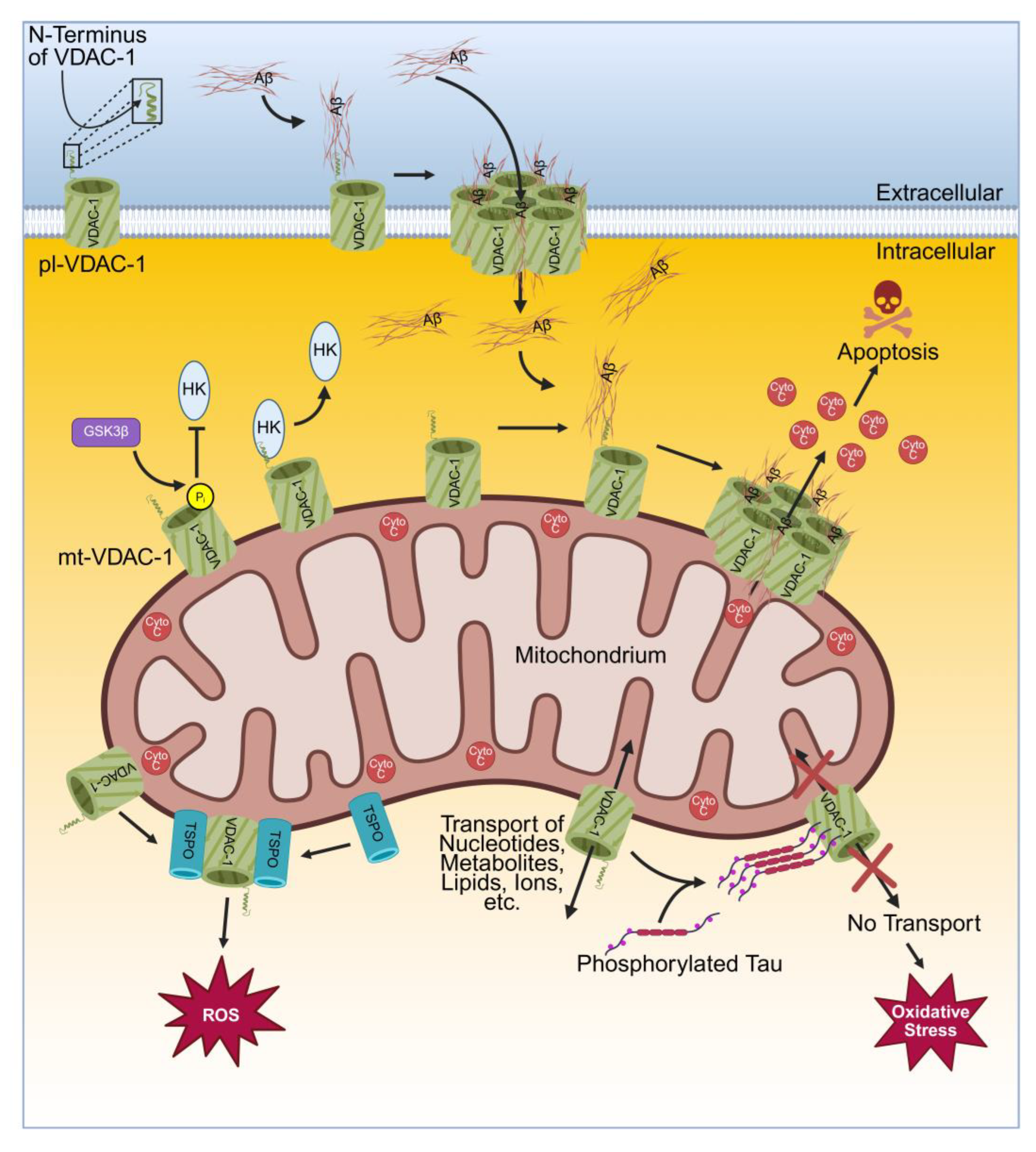
Figure 3.
Schematic layout for the role of VDAC-1 in PD. Monomeric αSyn can translocate through the mt-VDAC-1 into IMS where it directly interferes with complexes I, II, III and IV of the ETC in the IMM resulting in increased ROS production. Furthermore, monomeric αSyn can bind into the pore of mt-VDAC-1 blocking the transfer of ADP and ATP and thereby depleting the ATP synthase. The Ca2+ ion permeability of VDAC-1 changes upon binding of monomeric αSyn. Under the influence of ROS, monomeric αSyn oligomerise and these oligomers of αSyn can attach to the OMM which induces mitochondrial dysfunction leading to neuronal cell death. At the MAM, the IP3R3-Grp75-VDAC-1 requires DJ-1 and this complex enables the interorganelle transfer of Ca2+ ions between the endoplasmic reticulum and mitochondria. In PD, mutant DJ-1 L166P acquires a reduced interaction with the IP3R3-Grp75-VDAC-1 complex and in sporadic PD patients show a lower level of DJ-1 resulting in a disturbed interorganelle Ca2+ transport. LRRK2 can inhibit the mitochondrial entry of uMtCK. Normally, uMtCK enters the mitochondria, it becomes processed to CK and interacts with ANT and mt-VDAC-1. Without CK, ANT and VDAC-1 interact with each other leading to the opening of the PTP yielding to release of Cyto c inducing neuronal apoptosis. At the OMM, phosphorylated PINK1 recruits PRKN which becomes phosphorylated and subsequently ubiquitinates several proteins located in the OMM, among them mt-VDAC-1. While mono-ubiquitinated VDAC-1 inhibits apoptosis, poly-ubiquitinated VDAC-1 induces mitophagy. In PD, mutated PRKN impairs ubiquitination of mt-VDAC-1. This figure was created in https://BioRender.com (accessed 15th May 2025).
Figure 3.
Schematic layout for the role of VDAC-1 in PD. Monomeric αSyn can translocate through the mt-VDAC-1 into IMS where it directly interferes with complexes I, II, III and IV of the ETC in the IMM resulting in increased ROS production. Furthermore, monomeric αSyn can bind into the pore of mt-VDAC-1 blocking the transfer of ADP and ATP and thereby depleting the ATP synthase. The Ca2+ ion permeability of VDAC-1 changes upon binding of monomeric αSyn. Under the influence of ROS, monomeric αSyn oligomerise and these oligomers of αSyn can attach to the OMM which induces mitochondrial dysfunction leading to neuronal cell death. At the MAM, the IP3R3-Grp75-VDAC-1 requires DJ-1 and this complex enables the interorganelle transfer of Ca2+ ions between the endoplasmic reticulum and mitochondria. In PD, mutant DJ-1 L166P acquires a reduced interaction with the IP3R3-Grp75-VDAC-1 complex and in sporadic PD patients show a lower level of DJ-1 resulting in a disturbed interorganelle Ca2+ transport. LRRK2 can inhibit the mitochondrial entry of uMtCK. Normally, uMtCK enters the mitochondria, it becomes processed to CK and interacts with ANT and mt-VDAC-1. Without CK, ANT and VDAC-1 interact with each other leading to the opening of the PTP yielding to release of Cyto c inducing neuronal apoptosis. At the OMM, phosphorylated PINK1 recruits PRKN which becomes phosphorylated and subsequently ubiquitinates several proteins located in the OMM, among them mt-VDAC-1. While mono-ubiquitinated VDAC-1 inhibits apoptosis, poly-ubiquitinated VDAC-1 induces mitophagy. In PD, mutated PRKN impairs ubiquitination of mt-VDAC-1. This figure was created in https://BioRender.com (accessed 15th May 2025).
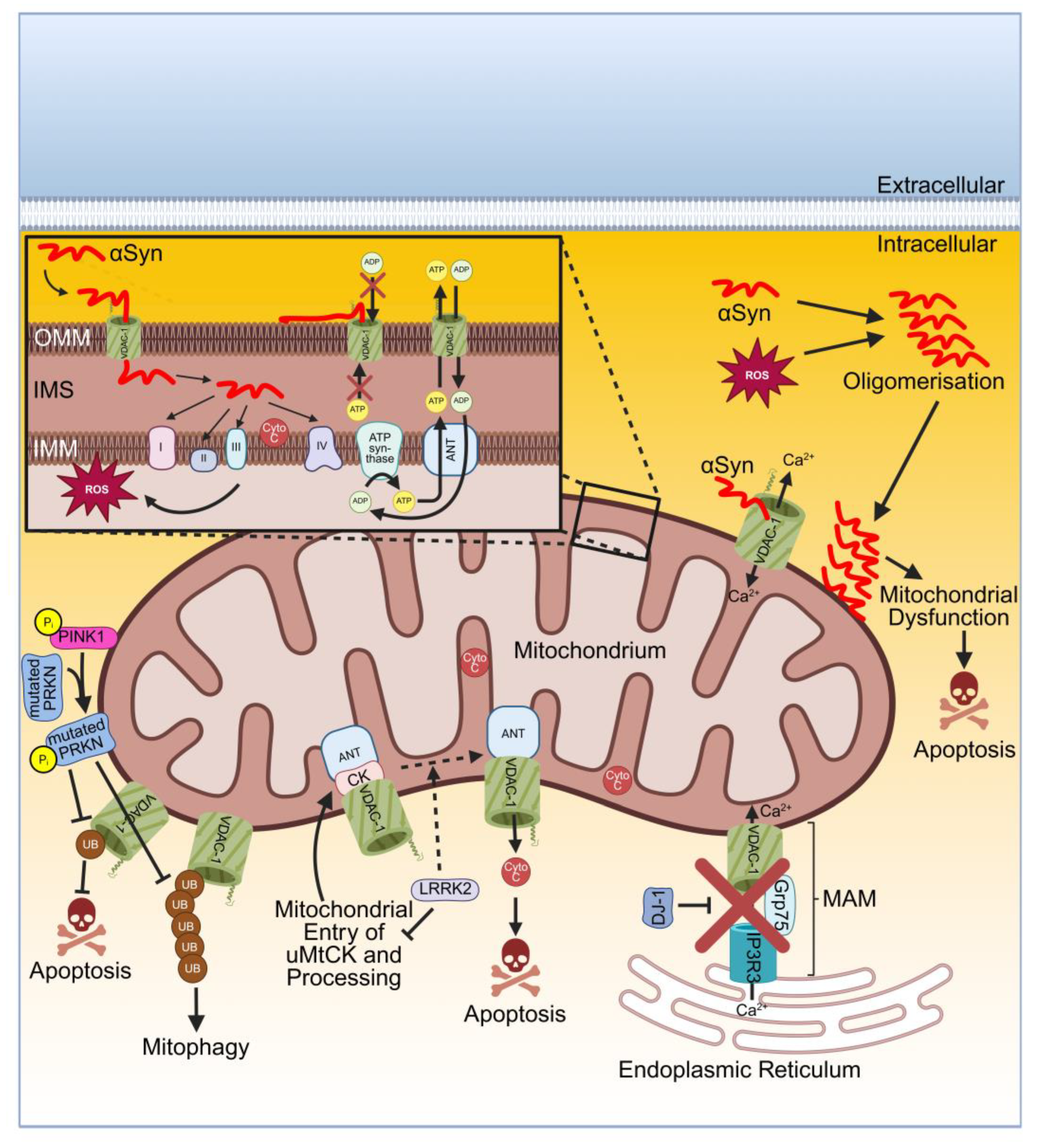
Figure 4.
Overview of disease specific proteins interacting with VDAC of the discussed neurodegenerative diseases. VDAC is symbolized by the three-dimensional structure of human VDAC-1 (Protein Data Bank entry: 2JK4; [16]). This figure was created in https://BioRender.com (accessed 15th May 2025).
Figure 4.
Overview of disease specific proteins interacting with VDAC of the discussed neurodegenerative diseases. VDAC is symbolized by the three-dimensional structure of human VDAC-1 (Protein Data Bank entry: 2JK4; [16]). This figure was created in https://BioRender.com (accessed 15th May 2025).
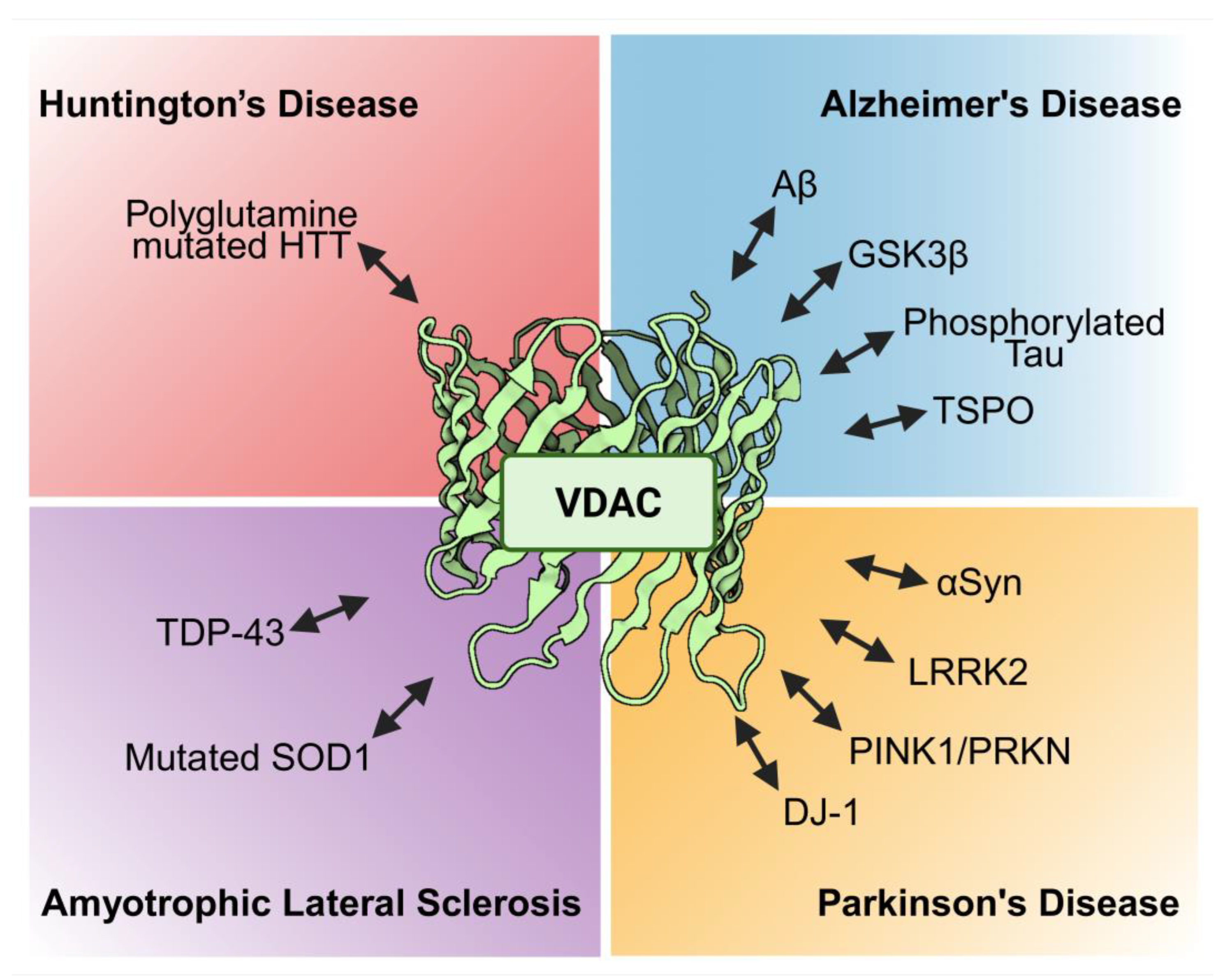
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



