Submitted:
17 May 2025
Posted:
19 May 2025
You are already at the latest version
Abstract
Prenatal and postnatal skeletal muscle development in ruminants is coordinated by interactions between genetic, nutritional, epigenetic, and endocrine factors. This review focuses on the influence of maternal nutrition during gestation on fetal myogenesis, satellite cell dynamics, and myogenic regulatory factor (MRF) expression, including MYF5, MYOD1, and MYOG. Studies in sheep and cattle indicate that nutrient restriction or overnutrition alters muscle fiber number, cross-sectional area, and the transcriptional regulation of myogenic genes in offspring. Postnatally, muscle hypertrophy is primarily mediated by satellite cells, which are activated via Pax7, MyoD, and Myf5, and regulated through mechanisms such as Carm1-induced chromatin remodeling and miR-31-mediated mRNA expression. Hormonal signaling via the GH–IGF1 axis and thyroid hormones further modulate satellite cell proliferation and protein accretion. Genetic variants, such as myostatin mutations in Texel sheep and Belgian Blue cattle, enhance muscle mass but may compromise reproductive efficiency. Nutritional interventions, including plane of nutrition, supplementation strategies, and environmental stressors such as heat and stocking density, significantly influence muscle fiber composition and carcass traits. This review provides a comprehensive overview of skeletal muscle programming in ruminants, tracing the developmental trajectory from progenitor cell differentiation to postnatal growth and maturation. These insights underscore the need for integrated approaches combining maternal diet optimization, molecular breeding, and precision livestock management to enhance muscle growth, meat quality, and production sustainability in ruminant systems.
Keywords:
myogenesis
; maternal nutrition
; satellite cells
; myogenic regulatory factors
; postnatal hypertrophy
; prenatal programming
; carcass quality
; myostatin
; IGF-1
; meat production
1. Introduction
Muscle growth and development align with overall meat production in ruminants, and the lean-to-fat ratio measures its efficiency. However, muscle tenderness and marbling contribute to the overall quality of meat. Since all muscle fibers are formed before birth [1,2], manipulating prenatal muscle development can help manage overall beef production [3]. Both muscle fibers and intra-muscular adipocytes are the sites for intra-muscular fat accumulation or marbling, and it is affected during prenatal and neonatal stages in ruminants [4,5]. Muscles are the organs of lower priority in nutrient partitioning; their development begins during early gestation and continues across gestation [6,7]. Limited nutrition during pregnancy can affect overall muscle development, leaving a lifelong effect on the meat quality and production [8,9,10].
Both prenatal and post-natal phases of muscle development are essential for myogenesis and the marbling effect. Most prenatal perspectives involve the hyperplasia of muscle fibers and adipogenic cells; however, neonatal and postnatal stages are focused on cellular hypertrophy [6,7,11]. The dermomyotome progenitor cell, which forms the dermatome and myotome [12,13], regulated by myogenic regulatory factors, differentiates into myogenic cells [14,15,16], forming primary myofibers during embryogenesis. The non-myogenic or dermatome cells of dermomyotome differentiate into fibro-adipogenic progenitors, regulated by key transcriptional factors [17]. The maternal nutritional implications during gestation and neonatally were found to be impacting both myogenic and adipogenic cells postnatally, either in terms of hyperplasia or hypertrophy [18,19].
Skeletal muscle constitutes the primary component of meat, therefore, making its growth and development critical in the livestock industry. While prenatal muscle development establishes the number of muscle fibers, postnatal muscle growth is largely hypertrophic, relying on satellite cell proliferation, protein accretion, and hormonal regulations [20,21,22,23]. Satellite cells, the resident muscle stem cells, play a pivotal role in postnatal muscle hypertrophy by contributing additional nuclei to growing muscle fibers, thereby supporting protein synthesis and fiber enlargement [24]. Nutritional status, hormonal signals such as growth hormone and insulin-like growth factor-1, and environmental factors modulate the activity of these cells [5,24,25]. Adequate nutrition, particularly during gestation, enhances satellite cell proliferation and differentiation, promoting muscle fiber hypertrophy and improving meat yield and quality [23,24]. Conversely, nutrient restriction during critical postnatal windows can impair satellite cell function, limiting muscle growth potential and negatively affecting meat characteristics such as tenderness and marbling. Furthermore, the balance between muscle fiber growth and intramuscular fat deposition is influenced by the interaction of satellite cells with fibro-adipogenic progenitors, which are also responsive to nutritional and hormonal cues [4,5,21,23]. Understanding these complex regulatory mechanisms is essential for developing feeding and management strategies that optimize muscle development and marbling, thereby enhancing both the quantity and quality of ruminant meat production.
Ruminants exhibit variations in muscle development due to genetic, nutritional, and environmental factors. Understanding the mechanism underlying postnatal muscle growth can aid in optimizing production practices to ultimately improve meat yield and quality, while addressing the challenges related to sustainability and feed efficiency. This review provides a comprehensive overview of skeletal muscle programming in ruminants, tracing the developmental trajectory from progenitor cell differentiation to postnatal growth and maturation. We will critically evaluate the impact of maternal nutrition on fetal myogenesis, elucidating the downstream consequences for beef production traits and meat quality attributes.
2. Prenatal Muscle Development in Ruminants
Fetal Programming and Muscle Development
The concept of the fetal programming hypothesis, initially proposed by Barker and colleagues, suggests that the insults or adverse environmental conditions during gestation, such as maternal malnutrition, induce persistent structural and functional adaptations in fetal tissues [26]. Epidemiological and experimental studies in ruminants and humans demonstrate that nutritional stress during critical developmental windows (e.g., myogenesis, adipogenesis) disrupts organogenesis through gene expression regulation and altered maternal-fetal resource partitioning [6,7,18,19,27].
Muscle development is a highly regulated process. It begins during embryogenesis and continues through fetal and postnatal stages [6,7,11]. Muscle fibers, or myofibers, are the fundamental structural units of skeletal muscle, formed exclusively during prenatal development in livestock species [1,2]. Understanding the mechanisms underlying myogenesis during this critical period is essential, as events occurring at this stage profoundly impact postnatal growth, muscle composition, and meat quality [28].
Prenatal myogenesis can be divided into primary and secondary myogenesis. Primary myogenesis occurs during the embryonic stage, forming primary muscle fibers that serve as templates for secondary fibers [16]. Secondary myogenesis, during the fetal stage, generates most muscle fibers from a pool of dPC [29]. These dPC during embryogenesis initiate the primary myogenesis by expressing myogenic regulatory factors (MRF), including myogenic factor 5 (Myf5), MyoD, MYOG, and MRF4, converting dPC to differentiated muscle fibers [30,31]. Although these primary muscle fibers are limited in number, their role as templates for subsequent fiber formation highlights their importance in determining overall muscle structure. The primary myogenesis in ruminants varies in days of gestation (dG) from 31-52 to 47-85 for sheep and cattle, respectively, see Figure 1 [7,32].
Secondary myogenesis occurs during the fetal stage of development, accounting for most of the fibers formed in farm animals, see Figure 1. Secondary myogenesis in ruminants varies in timing; mid-gestation in cattle (ranging 119-240 days of gestation; term ~284 days) [7,18], and early-to-mid-gestation in sheep (ranging 38-110 days of gestation; term ~145 days) [32,33]. Secondary muscle fibers arise from the fusion of proliferating myogenic cells, and increasing the number of myogenic dPC will result in more fiber formation. The number of muscle fibers formed during the fetal stage is directly influenced by the availability and proliferation of myogenic dPC, and it is highly sensitive to nutrient availability [2].
Considering the timeline of muscle development in ruminants (see Figure 1), the early-to-mid gestation timeframe is important for ruminants, and limited maternal nutrition can leave life-long effects by reducing the number of myogenic dPC.
Maternal Nutrition and Muscle Development
Muscles are the organs of lower priority to nutrient partitioning during fetal programming [6]. Most studies on maternal nutrition and developmental muscle programming in ruminants have highlighted the lasting effects of both maternal restricted and overnutrition [9,34,35,36,37,38,39,40,41,42,43]. Alterations in maternal nutrition were investigated to determine their impact on overall muscle development through gene expression regulation. The restricted or limited nutrition research in ruminant models was focused on studying the consequences of escalating feed input costs, which account for up to 70% of total expenditures in livestock production [44]. The rise in feed costs is attributed to supply chain vulnerabilities and extreme weather conditions, posing significant challenges for cattle farmers and ranchers [45]. However, the overnutrition models in ruminants have been studied to enhance marbling and myogenesis during skeletal muscle development both prenatally and neonatally [34,35,36,46]. The impact of prenatal maternal nutrition on myogenesis in ruminants has been explained extensively in the literature [3,5,6,11,16,46]. However, this review will focus on the genomic aspects of prenatal myogenesis, the least explored in ruminant models.
In terms of maternal nutrient restriction, different studies have highlighted the impacts on fetal myogenesis due to alterations in fetal gene expression profiles [9,34,35,37,38,39,43,47,48,49]. During the periconceptual period, ranging 18 days before and 6 days after ovulation, the ewes that received restricted nutrition (half the maintenance requirements) showed a decrease in muscle fiber count compared to those that received 1.5 times the maintenance requirements [39]. In the periconceptual cattle model, the maternal nutrient restriction (Average Daily Gain, ADG, -0.08 kg/d) during the first 50 dG, altered the expression of MRF, MYOG and MYOD1, in fetal hind limb muscle tissues compared to fetuses from heifers provided with ADG of 0.5 kg/d [42]. In the sheep model, from early gestation till parturition (45-135 dG, term ~147 days), when both under- and over-nutrition models were studied (under, ADG = 0.047 kg/d; over, ADG = 0.226 kg/d; control, ADG = 0.135 kg/d), the increasing trend in overall cross-sectional area of fetal semitendinosus and longissimus muscles fibers was observed from 90dG till parturition in consistent with fetal growth trajectory where downregulation of MYF5 and PAX7 [37]. These transcriptomic changes were found to be negatively associated with the myogenesis of developing lambs at birth, given their metabolite profiles [35]. In the cattle model of maternal nutrient restriction (ADG = 0.59kg/d) from 147-247 dG, mid-to-late gestation, the fetuses showed an increase in expression of MYOD1 and MYOG in longissimus muscle compared to fetuses from cows receiving ADG of 1.11 kg/d [9]. All these studies from ruminant models highlighted that maternal nutrient restriction across gestation impacted the expression profiles of MRF, leaving lasting impacts on overall fetal myogenesis and offspring metabolic profiles at birth.
Like maternal nutrient restriction, overnutrition also led to perturbations in fetal myogenesis and marbling in ruminant models [34,35,36]. However, the marbling effects have not been studied extensively in ruminant models, and most of the research is focused on hepatic and muscle models of fetal development in sheep [34,37,38,50,51,52], with a few focusing on beef cattle [9,36]. In the sheep model, where ewes received maternal overnutrition, 150% of maintenance, from 60 days before gestation till parturition, showed lower leptin concentration neonatally when compared to those that received maintenance requirements only, indicating potential impacts on lambs’ marbling and myogenesis [53]. Similarly, in another study with the same nutritional sheep model during the periconceptual period, 60 days before gestation till 75 dG, the MRF (MyoD and MYOG) were downregulated in fetal semitendinosus skeletal muscle tissues from overnutrition ewes compared to fetuses from control ewes [40]. Similarly, as discussed in restricted nutritional models, the ewes received over-nutrition (under, ADG = 0.047 kg/d; over, ADG = 0.226 kg/d; control, ADG = 0.135 kg/d) from early gestation till parturition (45-135 dG, term ~147 days), also showed downregulation of MYF5 and PAX7 [37]. In the cattle model of overnutrition (ADG = 1.11 kg/d) from 147-247 dG, mid-to-late gestation, the fetuses showed a downregulation in expression of MYOD1 and MYOG in longissimus muscle compared to fetuses from cows receiving ADG of 0.59kg/d, as vice versa was observed in the nutritional restriction model [9].
3. Postnatal Muscle Development in Ruminants
Mechanisms of Postnatal Muscle Growth
Postnatal muscle growth in ruminants is predominantly a result of muscle fiber hypertrophy, where existing muscle fibers increase in size rather than number. Satellite cells, a population of muscle-resident stem cells, play a crucial role in this process by contributing additional nuclei to muscle fibers, enabling greater protein synthesis capacity [5,24,25]. These cells remain quiescent until activated by MRF, Pax7 and Pax3 [54], once activated, satellite cells proliferate, differentiate, and fuse with existing myofibers, supporting continued muscle growth and repair.
The MyoD and Myf5 are the two major MRF involved in differentiating satellite cells to myogenic proliferation [55,56]. The regulation of Myf5 activation postnatally is intricately linked to arginine methylation of Pax7 by the enzyme Carm1, which catalyzes the recruitment of the Wdr5-Ash2L-MLL2 histone methyltransferase complex to the Myf5 locus, thereby promoting chromatin remodeling conducive to gene transcription [57,58]. The Myf5 transcripts are present even in quiescent satellite cells, but were sequestered in mRNP granules through miR-31-mediated mechanisms, preventing premature translation. Upon satellite cell activation, these granules disassemble, facilitating the rapid onset of Myf5 translation and subsequent myogenic activation [20]. Myotubule formation under the influence of MyoD induces MYOG expression, resulted in downregulation of Myf5 [59]. This coordinated activity of MyoD and MYOG leads to the expression of MRF4, regulating terminal differentiation and fusion with muscle fibers [5]. Differences in MRF expression levels among ruminant species and breeds can contribute to variations in muscle growth potential and efficiency, influencing overall meat production.
Hormones play a pivotal role in regulating postnatal muscle development. The IGF-1/mTOR signaling pathway is a key driver of muscle protein synthesis, responding to anabolic stimuli such as, growth hormone (GH) and nutrient availability [21,22]. The pituitary gland secretes GH, stimulates hepatic IFG-1 production, which subsequently enhances muscle growth by activating satellite cells and promoting protein accretion [21,22]. Additionally, thyroid hormones, insulin, and androgenic steroids contribute to muscle growth by modulating metabolic and anabolic processes in muscle tissue.
Genetic differences among breeds can influence muscle growth potential by influencing gene expression regulation. Myostatin, a negative regulator of muscle growth, plays a significant role in controlling muscle mass [60,61]. Mutations in the myostatin gene result in increased muscle fiber hypertrophy, as observed in Belgian Blue cattle and Texel sheep, leading to the “double muscling” phenotype [62]. While these genetic traits enhance muscle yield, they can also present challenges related to calving difficulties, reduced fertility, and altered meat texture.
Different ruminant breeds exhibit muscle fiber composition variations, ultimately impacting meat quality. Slow-twitch oxidative (Type I) fibers contribute to endurance and higher myoglobin content, producing darker, more flavorful meat. Whereas, fast twitch glycolytic (Type II) fibers support rapid growth and increased carcass weight, but can reduce meat tenderness [5]. Dairy cattle breeds, in contrast to beef cattle, seem to have higher oxidative metabolism and greater oxidase activities [63]. The Piemontese cattle breed is found to have higher glycolytic fibers than oxidative ones when compared to Belgian blue breed bulls [64]. Similarly, in sheep breeds, the Scottish Blackface breed presented type I muscle fibers compared to the Texel breed, which had type II muscle fibers [65]. Selective breeding programs to target optimal fiber type proportions and other growth traits to balance meat quality and production efficiency also impact the structure and quality of muscle and meat production in ruminants.
Nutritional and Environmental Influences on Muscle Growth
Nutritional management plays a crucial role in optimizing postnatal muscle growth. Adequate protein intake provides essential amino acids necessary for muscle protein synthesis, while energy availability influences the efficiency of protein accretion [23]. High-energy diets can enhance muscle growth, but may also increase fat deposition, affecting carcass composition and meat quality [66,67]. In contrast, restricted nutrition during early development can lead to long-term reductions in muscle growth potential [23]. Restricting energy intake to 65% of NRC recommendations in heifers and 50% in cows for 100 days prepartum was found to reduce calf birth and weaning weights and increase calf mortality, thereby impacting herd productivity [68]. In another study, cows were managed to either maintain a body condition score (BCS) of 5.0 or lose one BCS via energy restriction during mid-gestation, followed by re-alimentation through late gestation [69]. Although birth weights were unaffected, the calves born to restricted cows were lighter from the receiving phase till day 57 post-weaning [69]. However, when carcasses were harvested and evaluated on day 208 after birth, and found higher marbling to backfat in calves from restricted heifers, although ribeye area, hot carcass weight, and meat color and tenderness were not different [70]. The results were further supported by another study on the same cattle model, where improved marbling-to-backfat ratios and greater tenderness in steaks were found from calves of restricted dams [71]. In sheep model, the lambs born to ewes, subjected to nutrient restriction (50% of NRC requirements) from early-to-mid-gestation (day 28 to 78 of gestation), were found heavier and had more backfat when slaughtered at day 280 of age compared to lambs from control fed ewes (100% of NRC requirements) [72]. Together, these studies emphasize that mid-gestational maternal nutrient intake programs progeny outcomes, holds potential for optimizing carcass characteristics, production efficiency, and resilience in overall ruminant production systems, see Figure 1 and Table 1. Future research should aim to integrate gene expression profiling of satellite cells in offspring with maternal diet records to identify key nutritional signals that modify the myogenic epigenome during fetal life.
Other environmental factors, such as temperature stress, stocking density, and handling practices, can also impact muscle growth in ruminants. Chronic stress leads to elevated cortisol levels, which can promote muscle protein degradation and inhibit growth [73]. Proper livestock management, including adequate space, controlled feeding regimens, and minimized stress during transport and harvest, is essential for maintaining optimal growth rates and meat quality. These environmental factors, along with genetic selection and precision nutritional management, can optimize muscle development, production efficiency, and consumer acceptance of ruminant meat products.
4. Final Considerations
Muscle development in ruminants is a complex-multifactorial process beginning in utero and extending postnatally, intricately regulated by maternal nutrition, genetic programming, satellite cell activity, and environmental conditions. During gestation, especially early-to-mid gestational stages, maternal nutrition exerts profound effects on fetal myogenesis by altering the expression of MRF such as MyoD, Myf5, MYOG, and MRF4, as well as key transcription factors like Pax3 and Pax7. These molecular changes influence muscle fiber number, fiber type composition, and the myogenic potential of satellite cells, with long-term consequences for offspring muscle development, carcass traits, and overall production efficiency. Postnatally, muscle growth predominantly occurs through hypertrophy, facilitated by satellite cells that contribute additional nuclei to existing fibers. This process is tightly regulated by the same core MRF network and further modulated by epigenetic factors (e.g., Carm1-mediated chromatin remodeling), nutrient availability, and endocrine signals such as growth hormone (GH), insulin-like growth factor-1 (IGF-1), and thyroid hormones. Additionally, genetic factors, including myostatin mutations and breed-specific muscle fiber distributions, play critical roles in determining muscle growth potential, meat quality, and productivity. Evidence suggests that targeted maternal nutritional interventions during gestation can “program” satellite cell activity and muscle characteristics in offspring, highlighting the importance of developmental plasticity in livestock systems. To meet the increasing global demand for high-quality, sustainable meat, future research should adopt a systems biology approach integrating genomic and epigenomic profiling of both fetal and postnatal muscle tissues, and precision feeding to stress management strategies tailored to gestational stage and breed-specific requirements. By combining these strategies with genetic selection tools and emerging nutritional technologies, the livestock industry can enhance both the quantity and quality of ruminant meat production, ensuring resilience, sustainability, and profitability in future food systems.
However, despite significant advances, several limitations constrain the full understanding and practical application of muscle development programming in ruminants. Firstly, much of the previous studies derive knowledge from model organisms or in vitro studies, which may not fully replicate the complexity of in vivo ruminant muscle development, particularly under variable field conditions. The translation of molecular findings to on-farm nutritional strategies remain challenging due to the multifaceted interactions between genetics, nutrition, and environment, which can vary widely across breeds, management systems, and geographic regions. Secondly, the timing, dosage, and composition of maternal nutritional interventions necessary to elicit consistent and beneficial programming effects are not yet fully defined, with some studies reporting conflicting outcomes. This variability is partly due to differences in experimental design, nutrient sources, and the difficulty of controlling all confounding factors in large animal studies. Thirdly, the long-term effects of prenatal and early postnatal nutritional manipulations on animal health, welfare, and reproductive performance require further investigation to avoid unintended consequences. Moreover, epigenetic modifications implicated in muscle programming are dynamic and influenced by multiple external stimuli, raising questions about the stability and heritability of such changes across generations. Lastly, economic and practical constraints limit the widespread adoption of precision feeding and genomic technologies, especially in resource-limited production systems. To overcome these challenges, interdisciplinary research combining animal science, molecular genetics, nutrition, and bioinformatics is essential, alongside the development of cost-effective biomarkers and decision-support tools for producers. Addressing these limitations will be critical to harnessing the full potential of muscle developmental programming for sustainable and efficient ruminant meat production.
Author Contributions
K. G. and M.A. conceptualized the study, drafted the manuscript, and contributed to its editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Russell, R.G.; Oteruelo, F.T. An Ultrastructural Study of the Differentiation of Skeletal Muscle in the Bovine Fetus. Anatomy and Embryology 1981, 162, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.-J.; Ford, S.P.; Nathanielsz, P.W.; Du, M. Effect of Maternal Nutrient Restriction in Sheep on the Development of Fetal Skeletal Muscle1. Biol Reprod 2004, 71, 1968–1973. [Google Scholar] [CrossRef] [PubMed]
- Costa, T.C.; Gionbelli, M.P.; Duarte, M. de S. Fetal Programming in Ruminant Animals: Understanding the Skeletal Muscle Development to Improve Meat Quality. Animal Frontiers 2021, 11, 66–73. [Google Scholar] [CrossRef]
- Tong, J.; Zhu, M.J.; Underwood, K.R.; Hess, B.W.; Ford, S.P.; Du, M. AMP-Activated Protein Kinase and Adipogenesis in Sheep Fetal Skeletal Muscle and 3T3-L1 Cells. J Anim Sci 2008, 86, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Du, M. Prenatal Development of Muscle and Adipose and Connective Tissues and Its Impact on Meat Quality. Meat and Muscle Biology 2023, 7, 16230–16231. [Google Scholar] [CrossRef]
- Du, M.; Tong, J.; Zhao, J.; Underwood, K.R.; Zhu, M.; Ford, S.P.; Nathanielsz, P.W. Fetal Programming of Skeletal Muscle Development in Ruminant Animals1. J Anim Sci 2010, 88, E51–E60. [Google Scholar] [CrossRef]
- Anas, M.; Diniz, W.J.S.; Menezes, A.C.B.; Reynolds, L.P.; Caton, J.S.; Dahlen, C.R.; Ward, A.K. Maternal Mineral Nutrition Regulates Fetal Genomic Programming in Cattle: A Review. Metabolites 2023, 13, 593. [Google Scholar] [CrossRef]
- Marquez, D.C.; Paulino, M.F.; Rennó, L.N.; Villadiego, F.C.; Ortega, R.M.; Moreno, D.S.; Martins, L.S.; De Almeida, D.M.; Gionbelli, M.P.; Manso, M.R.; et al. Supplementation of Grazing Beef Cows during Gestation as a Strategy to Improve Skeletal Muscle Development of the Offspring. animal 2017, 11, 2184–2192. [Google Scholar] [CrossRef]
- Paradis, F.; Wood, K.M.; Swanson, K.C.; Miller, S.P.; McBride, B.W.; Fitzsimmons, C. Maternal Nutrient Restriction in Mid-to-Late Gestation Influences Fetal MRNA Expression in Muscle Tissues in Beef Cattle. BMC Genomics 2017, 18, 632. [Google Scholar] [CrossRef]
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How Muscle Structure and Composition Influence Meat and Flesh Quality. The Scientific World Journal 2016, 2016. [Google Scholar] [CrossRef]
- Summers, A.F.; Funston, R.N. FETAL PROGRAMMING: IMPLICATIONS FOR BEEF CATTLE PRODUCTION. Proceedings Range Beef Cow Symposium 2013, 5. [Google Scholar]
- Ben-Yair, R.; Kalcheim, C. Lineage Analysis of the Avian Dermomyotome Sheet Reveals the Existence of Single Cells with Both Dermal and Muscle Progenitor Fates. Development 2005, 132, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Tani, S.; Chung, U. il; Ohba, S.; Hojo, H. Understanding Paraxial Mesoderm Development and Sclerotome Specification for Skeletal Repair. Exp Mol Med 2020, 52, 1166. [Google Scholar] [CrossRef]
- Maroto, M.; Reshef, R.; Münsterberg, A.E.; Koester, S.; Goulding, M.; Lassar, A.B. Ectopic Pax-3 Activates MyoD and Myf-5 Expression in Embryonic Mesoderm and Neural Tissue. Cell 1997, 89, 139–148. [Google Scholar] [CrossRef]
- Hyatt, J.P.K.; McCall, G.E.; Kander, E.M.; Zhong, H.; Roy, R.R.; Huey, K.A. PAX3/7 Expression Coincides with Myod during Chronic Skeletal Muscle Overload. Muscle Nerve 2008, 38, 861–866. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhu, M.-J.; Dodson, M. V.; Du, M. Developmental Programming of Fetal Skeletal Muscle and Adipose Tissue Development. J Genomics 2013, 1, 29. [Google Scholar] [CrossRef]
- Avram, M.M.; Avram, A.S.; James, W.D. Subcutaneous Fat in Normal and Diseased States: 3. Adipogenesis: From Stem Cell to Fat Cell. J Am Acad Dermatol 2007, 56, 472–492. [Google Scholar] [CrossRef]
- Bonnet, M.; Cassar-Malek, I.; Chilliard, Y.; Picard, B. Ontogenesis of Muscle and Adipose Tissues and Their Interactions in Ruminants and Other Species. Animal 2010, 4, 1093–1109. [Google Scholar] [CrossRef]
- Gonzalez, J.M.; Camacho, L.E.; Ebarb, S.M.; Swanson, K.C.; Vonnahme, K.A.; Stelzleni, A.M.; Johnson, S.E. Realimentation of Nutrient Restricted Pregnant Beef Cows Supports Compensatory Fetal Muscle Growth. J Anim Sci 2013, 91, 4797–4806. [Google Scholar] [CrossRef]
- Crist, C.G.; Montarras, D.; Buckingham, M. Muscle Satellite Cells Are Primed for Myogenesis but Maintain Quiescence with Sequestration of Myf5 MRNA Targeted by MicroRNA-31 in MRNP Granules. Cell Stem Cell 2012, 11, 118–126. [Google Scholar] [CrossRef]
- Schiaffino, S.; Mammucari, C. Regulation of Skeletal Muscle Growth by the IGF1-Akt/PKB Pathway: Insights from Genetic Models. Skelet Muscle 2011, 1, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Zhao, J.X.; Yan, X.; Huang, Y.; Nicodemus, L. V.; Yue, W.; Mccormick, R.J.; Zhu, M.J. Fetal Muscle Development, Mesenchymal Multipotent Cell Differentiation, and Associated Signaling Pathways. J Anim Sci 2011, 89, 583–590. [Google Scholar] [CrossRef]
- Costa, T.C.; Du, M.; Nascimento, K.B.; Galvão, M.C.; Meneses, J.A.M.; Schultz, E.B.; Gionbelli, M.P.; Duarte, M. de S. Skeletal Muscle Development in Postnatal Beef Cattle Resulting from Maternal Protein Restriction during Mid-Gestation. Animals 2021, 11, 860. [Google Scholar] [CrossRef]
- Pallafacchina, G.; Blaauw, B.; Schiaffino, S. Role of Satellite Cells in Muscle Growth and Maintenance of Muscle Mass. Nutrition, Metabolism and Cardiovascular Diseases 2013, 23, S12–S18. [Google Scholar] [CrossRef]
- Aiello, D.; Patel, K.; Lasagna, E. The Myostatin Gene: An Overview of Mechanisms of Action and Its Relevance to Livestock Animals. Anim Genet 2018, 49, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.P. The Fetal and Infant Origins of Adult Disease. BMJ : British Medical Journal 1990, 301. [Google Scholar] [CrossRef] [PubMed]
- Costa, T.C.; Gionbelli, M.P.; Duarte, M. de S. Fetal Programming in Ruminant Animals: Understanding the Skeletal Muscle Development to Improve Meat Quality. Anim Front 2021, 11, 66. [Google Scholar] [CrossRef]
- Dauncey, M.J.; Gilmour, R.S. Regulatory Factors in the Control of Muscle Development. Proceedings of the Nutrition Society 1996, 55, 543–559. [Google Scholar] [CrossRef]
- Bailey, P.; Holowacz, T.; Lassar, A.B. The Origin of Skeletal Muscle Stem Cells in the Embryo and the Adult. Curr Opin Cell Biol 2001, 13, 679–689. [Google Scholar] [CrossRef]
- Moncaut, N.; Rigby, P.W.J.; Carvajal, J.J. Dial M(RF) for Myogenesis. FEBS J 2013, 280, 3980–3990. [Google Scholar] [CrossRef]
- Buckingham, M.; Rigby, P.W.J. Gene Regulatory Networks and Transcriptional Mechanisms That Control Myogenesis. Dev Cell 2014, 28, 225–238. [Google Scholar] [CrossRef]
- Wilson, S.J.; McEwan, J.C.; Sheard, P.W.; Harris, A.J. Early Stages of Myogenesis in a Large Mammal: Formation of Successive Generations of Myotubes in Sheep Tibialis Cranialis Muscle. J Muscle Res Cell Motil 1992, 13, 534–550. [Google Scholar] [CrossRef]
- Maier, A.; McEwan, J.C.; Dodds, K.G.; Fischman, D.A.; Fitzsimons, R.B.; Harris, A.J. Myosin Heavy Chain Composition of Single Fibres and Their Origins and Distribution in Developing Fascicles of Sheep Tibialis Cranialis Muscles. J Muscle Res Cell Motil 1992, 13, 551–572. [Google Scholar] [CrossRef]
- Reed, S.A.; Raja, J.S.; Hoffman, M.L.; Zinn, S.A.; Govoni, K.E. Poor Maternal Nutrition Inhibits Muscle Development in Ovine Offspring. J Anim Sci Biotechnol 2014, 5, 43. [Google Scholar] [CrossRef]
- Martin, D.E.; Jones, A.K.; Pillai, S.M.; Hoffman, M.L.; McFadden, K.K.; Zinn, S.A.; Govoni, K.E.; Reed, S.A. Maternal Restricted- and Over-Feeding During Gestation Result in Distinct Lipid and Amino Acid Metabolite Profiles in the Longissimus Muscle of the Offspring. Front Physiol 2019, 10, 448206. [Google Scholar] [CrossRef]
- Duarte, M.S.; Gionbelli, M.P.; Paulino, P.V.R.; Serão, N.V.L.; Nascimento, C.S.; Botelho, M.E.; Martins, T.S.; Filho, S.C. V.; Dodson, M. V.; Guimarães, S.E.F.; et al. Maternal Overnutrition Enhances MRNA Expression of Adipogenic Markers and Collagen Deposition in Skeletal Muscle of Beef Cattle Fetuses1. J Anim Sci 2014, 92, 3846–3854. [Google Scholar] [CrossRef]
- Gauvin, M.C.; Pillai, S.M.; Reed, S.A.; Stevens, J.R.; Hoffman, M.L.; Jones, A.K.; Zinn, S.A.; Govoni, K.E. Poor Maternal Nutrition during Gestation in Sheep Alters Prenatal Muscle Growth and Development in Offspring. J Anim Sci 2020, 98, 1–15. [Google Scholar] [CrossRef]
- Hoffman, M.L.; Peck, K.N.; Wegrzyn, J.L.; Reed, S.A.; Zinn, S.A.; Govoni, K.E. Poor Maternal Nutrition during Gestation Alters the Expression of Genes Involved in Muscle Development and Metabolism in Lambs1. J Anim Sci 2016, 94, 3093–3099. [Google Scholar] [CrossRef]
- Quigley, S.P.; Kleemann, D.O.; Kakar, M.A.; Owens, J.A.; Nattrass, G.S.; Maddocks, S.; Walker, S.K. Myogenesis in Sheep Is Altered by Maternal Feed Intake during the Peri-Conception Period. Anim Reprod Sci 2005, 87, 241–251. [Google Scholar] [CrossRef]
- Tong, J.F.; Yan, X.; Zhu, M.J.; Ford, S.P.; Nathanielsz, P.W.; Du, M. Maternal Obesity Downregulates Myogenesis and β-Catenin Signaling in Fetal Skeletal Muscle. American Journal of Physiology-Endocrinology and Metabolism 2009, 296, E917–E924. [Google Scholar] [CrossRef]
- Thorn, S.R.; Regnault, T.R.H.; Brown, L.D.; Rozance, P.J.; Keng, J.; Roper, M.; Wilkening, R.B.; Hay, W.W.; Friedman, J.E. Intrauterine Growth Restriction Increases Fetal Hepatic Gluconeogenic Capacity and Reduces Messenger Ribonucleic Acid Translation Initiation and Nutrient Sensing in Fetal Liver and Skeletal Muscle. Endocrinology 2009, 150, 3021–3030. [Google Scholar] [CrossRef]
- Crouse, M.S.; Caton, J.S.; Cushman, R.A.; McLean, K.J.; Dahlen, C.R.; Borowicz, P.P.; Reynolds, L.P.; Ward, A.K. Moderate Nutrient Restriction of Beef Heifers Alters Expression of Genes Associated with Tissue Metabolism, Accretion, and Function in Fetal Liver, Muscle, and Cerebrum by Day 50 of Gestation. Transl Anim Sci 2019, 3, 855–866. [Google Scholar] [CrossRef]
- Diniz, W.J.S.; Crouse, M.S.; Cushman, R.A.; McLean, K.J.; Caton, J.S.; Dahlen, C.R.; Reynolds, L.P.; Ward, A.K. Cerebrum, Liver, and Muscle Regulatory Networks Uncover Maternal Nutrition Effects in Developmental Programming of Beef Cattle during Early Pregnancy. Scientific Reports 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Review: Feed Demand Landscape and Implications of Food-Not Feed Strategy for Food Security and Climate Change. animal 2018, 12, 1744–1754. [Google Scholar] [CrossRef]
- Cowley, C.; Cowley; Cortney Long-Term Pressures and Prospects for the U.S. Cattle Industry. Economic Review 2021, vol.107. [CrossRef]
- Du, M.; Ford, S.P.; Zhu, M.-J. Optimizing Livestock Production Efficiency through Maternal Nutritional Management and Fetal Developmental Programming. Animal Frontiers 2017, 7, 5–11. [Google Scholar] [CrossRef]
- Yan, X.; Huang, Y.; Zhao, J.X.; Rogers, C.J.; Zhu, M.J.; Ford, S.P.; Nathanielsz, P.W.; Du, M. Maternal Obesity Downregulates MicroRNA Let-7g Expression, a Possible Mechanism for Enhanced Adipogenesis during Ovine Fetal Skeletal Muscle Development. International Journal of Obesity 2012, 37, 568–575. [Google Scholar] [CrossRef]
- Crouse, M.S.; Caton, J.S.; Cushman, R.A.; McLean, K.J.; Dahlen, C.R.; Borowicz, P.P.; Reynolds, L.P.; Ward, A.K. Moderate Nutrient Restriction of Beef Heifers Alters Expression of Genes Associated with Tissue Metabolism, Accretion, and Function in Fetal Liver, Muscle, and Cerebrum by Day 50 of Gestation. Transl Anim Sci 2019, 3, 855–866. [Google Scholar] [CrossRef]
- Thorn, S.R.; Regnault, T.R.H.; Brown, L.D.; Rozance, P.J.; Keng, J.; Roper, M.; Wilkening, R.B.; Hay, W.W.; Friedman, J.E. Intrauterine Growth Restriction Increases Fetal Hepatic Gluconeogenic Capacity and Reduces Messenger Ribonucleic Acid Translation Initiation and Nutrient Sensing in Fetal Liver and Skeletal Muscle. Endocrinology 2009, 150, 3021–3030. [Google Scholar] [CrossRef]
- Fernández-Foren, A.; Meikle, A.; de Brun, V.; Graña-Baumgartner, A.; Abecia, J.A.; Sosa, C. Metabolic Memory Determines Gene Expression in Liver and Adipose Tissue of Undernourished Ewes. Livest Sci 2022, 260, 104949. [Google Scholar] [CrossRef]
- Hoffman, M.L.; Rokosa, M.A.; Zinn, S.A.; Hoagland, T.A.; Govoni, K.E. Poor Maternal Nutrition during Gestation in Sheep Reduces Circulating Concentrations of Insulin-like Growth Factor-I and Insulin-like Growth Factor Binding Protein-3 in Offspring. Domest Anim Endocrinol 2014, 49, 39–48. [Google Scholar] [CrossRef]
- Jones, A.K.; Hoffman, M.L.; Pillai, S.M.; McFadden, K.K.; Govoni, K.E.; Zinn, S.A.; Reed, S.A. Gestational Restricted- and over-Feeding Promote Maternal and Offspring Inflammatory Responses That Are Distinct and Dependent on Diet in Sheep†. Biol Reprod 2018, 98, 184–196. [Google Scholar] [CrossRef]
- Long, N.M.; Ford, S.P.; Nathanielsz, P.W. Maternal Obesity Eliminates the Neonatal Lamb Plasma Leptin Peak. J Physiol 2011, 589, 1455–1462. [Google Scholar] [CrossRef] [PubMed]
- Kassar-Duchossoy, L.; Giacone, E.; Gayraud-Morel, B.; Jory, A.; Gomès, D.; Tajbakhsh, S. Pax3/Pax7 Mark a Novel Population of Primitive Myogenic Cells during Development. Genes Dev 2005, 19, 1426. [Google Scholar] [CrossRef]
- Beauchamp, J.R.; Heslop, L.; Yu, D.S.W.; Tajbakhsh, S.; Kelly, R.G.; Wernig, A.; Buckingham, M.E.; Partridge, T.A.; Zammit, P.S. Expression of Cd34 and Myf5 Defines the Majority of Quiescent Adult Skeletal Muscle Satellite Cells. J Cell Biol 2000, 151, 1221. [Google Scholar] [CrossRef] [PubMed]
- Kuang, S.; Kuroda, K.; Le Grand, F.; Rudnicki, M.A. Asymmetric Self-Renewal and Commitment of Satellite Stem Cells in Muscle. Cell 2007, 129, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- McKinnell, I.W.; Ishibashi, J.; Le Grand, F.; Punch, V.G.J.; Addicks, G.C.; Greenblatt, J.F.; Dilworth, F.J.; Rudnicki, M.A. Pax7 Activates Myogenic Genes by Recruitment of a Histone Methyltransferase Complex. Nat Cell Biol 2007, 10, 77. [Google Scholar] [CrossRef]
- Kawabe, Y.I.; Wang, Y.X.; McKinnell, I.W.; Bedford, M.T.; Rudnicki, M.A. Carm1 Regulates Pax7 Transcriptional Activity Through MLL1/2 Recruitment During Asymmetric Satellite Stem Cell Divisions. Cell Stem Cell 2012, 11, 333. [Google Scholar] [CrossRef]
- Deato, M.D.E.; Marr, M.T.; Sottero, T.; Inouye, C.; Hu, P.; Tjian, R. MyoD Targets TAF3 /TRF3 to Activate Myogenin Transcription. Mol Cell 2008, 32, 96. [Google Scholar] [CrossRef]
- Lee, S.J. Myostatin: A Skeletal Muscle Chalone. Annu Rev Physiol 2023, 85, 269–291. [Google Scholar] [CrossRef]
- Chen, P.R.; Lee, K. INVITED REVIEW: Inhibitors of Myostatin as Methods of Enhancing Muscle Growth and Development. J Anim Sci 2016, 94, 3125–3134. [Google Scholar] [CrossRef]
- Bellinge, R.H.S.; Liberles, D.A.; Iaschi, S.P.A.; O’Brien, P.A.; Tay, G.K. Myostatin and Its Implications on Animal Breeding: A Review. Anim Genet 2005, 36, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Schreurs, N.M.; Garcia, F.; Jurie, C.; Agabriel, J.; Micol, D.; Bauchart, D.; Listrat, A.; Picard, B. Meta-Analysis of the Effect of Animal Maturity on Muscle Characteristics in Different Muscles, Breeds, and Sexes of Cattle. J Anim Sci 2008, 86, 2872–2887. [Google Scholar] [CrossRef] [PubMed]
- Bittante, G.; Cecchinato, A.; Tagliapietra, F.; Verdiglione, R.; Simonetto, A.; Schiavon, S. Crossbred Young Bulls and Heifers Sired by Double-Muscled Piemontese or Belgian Blue Bulls Exhibit Different Effects of Sexual Dimorphism on Fattening Performance and Muscularity but Not on Meat Quality Traits. Meat Sci 2018, 137, 24–33. [Google Scholar] [CrossRef]
- Bünger, L.; Navajas, E.A.; Stevenson, L.; Lambe, N.R.; Maltin, C.A.; Simm, G.; Fisher, A.V.; Chang, K.C. Muscle Fibre Characteristics of Two Contrasting Sheep Breeds: Scottish Blackface and Texel. Meat Sci 2009, 81, 372–381. [Google Scholar] [CrossRef]
- Ozawa, S.; Mitsuhashi, T.; Mitsumoto, M.; Matsumoto, S.; Itoh, N.; Itagaki, K.; Kohno, Y.; Dohgo, T. The Characteristics of Muscle Fiber Types of Longissimus Thoracis Muscle and Their Influences on the Quantity and Quality of Meat from Japanese Black Steers. Meat Sci 2000, 54, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Hudson, N.J.; Reverter, A.; Greenwood, P.L.; Guo, B.; Cafe, L.M.; Dalrymple, B.P. Longitudinal Muscle Gene Expression Patterns Associated with Differential Intramuscular Fat in Cattle. Animal 2015, 9, 650–659. [Google Scholar] [CrossRef]
- Corah, L.R.; Dunn, T.G.; Kaltenbach, C.C. Influence of Prepartum Nutrition on the Reproductive Performance of Beef Females and the Performance of Their Progeny. J Anim Sci 1975, 41, 819–824. [Google Scholar] [CrossRef]
- Taylor, A.R.; Mohrhauser, D.A.; Pritchard, R.H.; Underwood, K.R.; Wertz-Lutz, A.E.; Blair, A.D. The Influence of Maternal Energy Status during Mid-Gestation on Growth, Cattle Performance, and the Immune Response in the Resultant Beef Progeny. Professional Animal Scientist 2016, 32, 389–399. [Google Scholar] [CrossRef]
- Mohrhauser, D.A.; Taylor, A.R.; Underwood, K.R.; Pritchard, R.H.; Wertz-Lutz, A.E.; Blair, A.D. The Influence of Maternal Energy Status during Midgestation on Beef Offspring Carcass Characteristics and Meat Quality. J Anim Sci 2015, 93, 786–793. [Google Scholar] [CrossRef]
- Gardner, J.M.; Ineck, N.E.; Quarnberg, S.M.; Legako, J.F.; Carpenter, C.E.; Rood, K.A.; Thornton-Kurth, K.J.; Gardner, J.M.; Ineck, N.E.; Quarnberg, S.M.; et al. The Influence of Maternal Dietary Intake During Mid-Gestation on Growth, Feedlot Performance, MiRNA and MRNA Expression, and Carcass and Meat Quality of Resultant Offspring. Meat and Muscle Biology 2021, 5, 3–4. [Google Scholar] [CrossRef]
- Ford, S.P.; Long, N.M.; Ford, S.P.; Long, N.M. Evidence for Similar Changes in Offspring Phenotype Following Either Maternal Undernutrition or Overnutrition: Potential Impact on Fetal Epigenetic Mechanisms. Reprod Fertil Dev 2011, 24, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Rehfeldt, C.; Te Pas, M.F.W.; Wimmers, K.; Brameld, J.M.; Nissen, P.M.; Berri, C.; Valente, L.M.P.; Power, D.M.; Picard, B.; Stickland, N.C.; et al. Advances in Research on the Prenatal Development of Skeletal Muscle in Animals in Relation to the Quality of Muscle-Based Food. I. Regulation of Myogenesis and Environmental Impact. Animal 2011, 5, 703–717. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Timeline indicating myogenesis in ruminant models based on days in gestation (dG) [7,18,32,33].
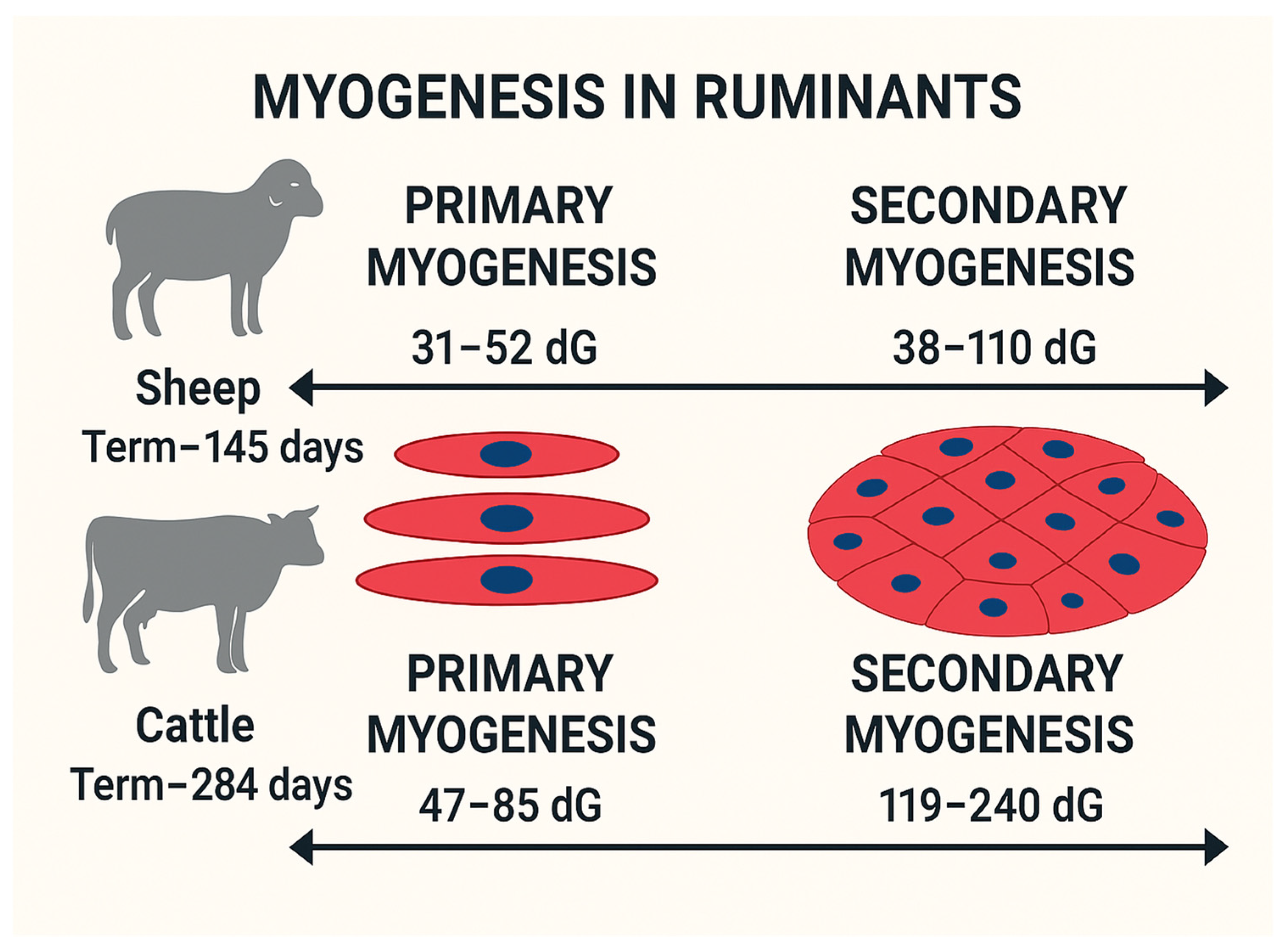

Table 1.
Studies demonstrating the maternal nutritional effects on pre- and post-natal skeletal muscle development in ruminant models.
Table 1.
Studies demonstrating the maternal nutritional effects on pre- and post-natal skeletal muscle development in ruminant models.
| Model Animal | Treatments | Gestation Stage | Effects | Reference |
| Sheep | Maternal nutrient restriction (half maintenance requirements vs. 1.5 times maintenance requirements) | Periconceptual period (18 days before and 6 days after ovulation) | Decrease in muscle fiber count in the restricted nutrition group compared to the control | [39] |
| Cattle | Maternal nutrient restriction (ADG -0.08 kg/d vs. ADG 0.5 kg/d) | Early gestation (first 50 days of gestation) | Altered expressions of MRF, MYOG, and MYOD1 in fetal hind limb muscle tissues in the restricted nutrition group. | [42] |
| Sheep | Maternal nutrient restriction and overnutrition (under: ADG = 0.047 kg/d; over: ADG = 0.226 kg/d; control: ADG = 0.135 kg/d) | Early gestation to parturition (45-135 dG) | Increased cross-sectional area of muscle fibers consistent with fetal growth trajectory; downregulation of MYF5 and PAX7 associated with negative impacts on myogenesis at birth. | [37] |
| Cattle | Maternal nutrient restriction and overnutrition (ADG = 0.59 kg/d vs. ADG = 1.11 kg/d) | Mid-to-late gestation (147-247 dG) | Increased expression of MYOD1 and MYOG in the restricted nutrition group compared to the overnutrition group. | [9] |
| Sheep | Maternal overnutrition (150% of maintenance requirements vs. maintenance requirements only) | From 60 days before gestation to parturition | Lower leptin concentration neonatally in the overnutrition group, indicating potential impacts on marbling and myogenesis. | [53] |
| Sheep | Maternal overnutrition (150% of maintenance requirements vs. control) | Periconceptual period (60 days before gestation to 75 dG) | Downregulation of MRF (MyoD and MYOG) in the overnutrition group compared to the control group. | [40] |
| Cattle | Maternal nutrient restriction leads to the loss of body condition score by 1 or maintains score of 5.0 | Mid gestational nutrient restriction (from start to end of mid-gestation) to study postnatal birth and weaning effects | Early growth suppressed; improved marbling-to-backfat ratio and tenderness in the restricted group | [69,70,71] |
| Sheep | Early to mid gestation maternal nutrient restriction (50% of NRC requirement) vs control-fed ewes (100% of NRC requirement) | Study the impacts of midgestational maternal Nutrient restriction on carcass and morphometric measures of lambs at harvesting (day 280 postnatally) | Lambs from restricted ewes were heavier and had more backfat at slaughter | [72] |
| Cattle | Prepartum nutrient restriction (heifers 65%, cows 50% NRC compared to 100% NRC as control for 100 days) | Postnatal (birth to weaning) | Lower calf birth/weaning weights; increased mortality rates | [68] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



