
Submitted:
16 May 2025
Posted:
16 May 2025
You are already at the latest version
Abstract
Marfan syndrome (MFS) is a connective tissue disorder caused by mutations in the fibrillin-1 gene, affecting multiple systems including the musculoskeletal, cardiovascular, and pulmonary systems. Vascular complications, particularly aortic root aneurysm, dissection, and rupture, are hallmark features of MFS. In recent decades, advances in diagnostics and treatments have significantly improved life expectancy for individuals with MFS; however, other vascular complications have become more concerning. On the other hand, the cardiovascular benefits of moderate exercise are well-documented, with evidence also suggesting that aerobic exercise improves vascular structure and cognitive function. The present study investigates the effects of mild aerobic exercise on the progression of aortic aneurysm in male and female MFS mice. Using high-resolution in vivo ultrasound imaging, the study assesses the functional properties of multiple key arteries, including the aorta, posterior cerebral, carotid, coronary, pulmonary, and renal arteries, and explore the relationship between aortic root diameters, arterial wall stiffness, and phenotypic changes across these arteries, aiming to identify key predictors that could provide insights into vascular health and the influence of exercise on arterial function and structure. Based on our findings, MFS mice exhibited significant increases in carotid PWV (wall stiffness), carotid wall thickness, coupled with reduced carotid distensibility, in both sexes compared to controls. Reduced PSV was observed in the pulmonary and posterior cerebral arteries of MFS mice, while coronary and renal arteries showed no changes. Mild exercise attenuated aortic and carotid pathology, reversing aortic root diameter growth, PWV, and carotid wall thickness, while improving carotid distensibility and restoring posterior cerebral artery blood flow, particularly in female MFS mice. In males, aortic PWV strongly correlated with posterior cerebral and pulmonary artery flow, while sinus of Valsalva diameter predicted carotid artery PWV and wall thickness in both sexes. Notably, the relationship between carotid distensibility and vascular metrics varied by sex. These findings underscore the vascular impacts of MFS, the therapeutic potential of mild exercise, and notable sex-specific differences in disease progression and arterial function.
Keywords:
Marfan syndrome
; aortic root aneurysm
; carotid artery
; cerebral artery
; pulse wave velocity
; blood flow
; exercise
1. Introduction
Marfan syndrome (MFS) is an autosomal dominant connective tissue disorder caused by mutations in the FBN1 gene, which encodes fibrillin-1, a large glycoprotein critical for extracellular matrix integrity and elastic fiber formation. The defective fibrillin-1 protein leads to dysregulation of transforming growth factor-beta (TGF-β) signaling, contributing to structural abnormalities in multiple organ systems [1,2,3]. Clinically, MFS is characterized by a wide array of manifestations, including skeletal deformities (e.g., scoliosis, pectus excavatum), ocular complications (e.g., ectopia lentis), and cardiovascular anomalies, particularly aortic root dilation and subsequent aortic dissection. Among these, cardiovascular complications represent the primary cause of morbidity and mortality in individuals with MFS. The current standard of care focuses on early diagnosis, pharmacological management, surgical intervention, and lifestyle modifications aimed at reducing hemodynamic stress on the aorta. Pharmacologic agents such as beta-blockers and angiotensin receptor blockers (ARBs) have shown efficacy in delaying the progression of aortic dilation by reducing aortic wall stress and modulating downstream TGF-β signaling. Surgical repair of the aortic root is a life-saving intervention that has significantly improved survival rates. Despite these advancements, the chronic and progressive nature of MFS, compounded by recurrent medical interventions and multisystem involvement, continues to profoundly affect patients’ quality of life [4,5,6].
Recently, attention has turned to lifestyle interventions, particularly physical activity, as a complementary approach to improve outcomes in MFS. Traditionally, individuals with MFS have been advised to avoid high-intensity physical activity due to concerns over exacerbating aortic stress and the risk of aortic rupture. However, emerging evidence suggests that mild to moderate aerobic exercise may offer protective cardiovascular benefits without compromising aortic integrity [7,8]. Preclinical studies in MFS mouse models have demonstrated that low to moderate-intensity aerobic exercise improves aortic wall structure by preserving elastin integrity and reducing aortic dilation. Mechanistically, these benefits appear to be mediated by exercise-induced improvements in endothelial function, nitric oxide (NO) production, and reduced transforming growth factor beta (TGF-β) and matrix metalloproteinase (MMP) activity [9,10].
Despite these promising findings, several critical gaps remain. For example, our knowledge of the impact of MFS’s pathology on various organ systems, particularly the vasculature beyond the aorta, remains limited [11]. Existing studies have largely focused on the aortic root and its associated complications, leaving gaps in understanding the structure, function, and interplay of other central and peripheral arteries, such as the cerebral, carotid, coronary, pulmonary, and renal arteries, in the context of MFS. Additionally, the implications of aging and common vascular comorbidities, such as atherosclerosis and hypertension, on MFS-associated arterial dysfunction are underexplored [12,13,14,15,16,17,18,19,20,21,22,23].
The interplay between physical activity and vascular health in MFS also remains poorly understood. Existing evidence highlights the potential benefits of mild aerobic exercise in enhancing aortic structure and function [24,25,26]. In this study, we aim to further explore the effects of exercise on other central and peripheral arteries in MFS mice using in vivo ultrasound imaging. The goal is to develop evidence-based recommendations for integrating safe and effective physical activity into the management of MFS, ultimately improving patient outcomes and quality of life.
2. Materials & Methods
2.1. Experimental Animals & Exercise Protocol
In this study, we utilized a transgenic mouse model carrying a missense mutation in the Fbn1 allele (C1041G), which introduces a cysteine-to-glycine substitution (Cys1041→Gly) within an epidermal growth factor–like domain of fibrillin-1 (Fbn1C1041G/+). The control group consisted of wild-type C57BL/6 littermates. At 6 weeks of age, male and female mice were randomly assigned to the following groups: Control (Ctrl), MFS, and MFS + treadmill exercise. Mice were group-housed (four per cage) under a 12-hour light/12-hour dark cycle, with unrestricted access to food and water. The exercise group were subjected to treadmill running at 55% VO2max. Acclimation to the exercise protocol was conducted over three consecutive days, starting at 5 m/min, and gradually increasing to 8 m/min for 20 minutes per session. During the training phase, mice exercised 5 days/week, and the protocol continued until the mice reached 6 months of age. All procedures and protocols involving mice were approved by the Midwestern University Institutional Animal Care and Use Committee. All animals received humane care in accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals published by the United States National Institute of Health.
2.2. Sample Size Calculation
Sample size estimation was conducted based on previous preliminary data for the diameter of the sinus of Valsalva in control and MFS mice. Assuming an expected intervention (exercise) effect of a 15–20% reduction in aortic root diameter—an effect considered clinically meaningful given that in humans, an increase from 4 to 5 cm typically warrants surgical root replacement—we estimated the required sample size to detect this difference. Using a power of 0.8 and an alpha level of 0.05, the estimated sample size for comparing two means was determined to be eight animals per group.
2.3. Echocardiography
At 7 months of age, male and female mice from both the MFS and Ctrl groups underwent in vivo high-resolution ultrasound imaging. Echocardiography was performed using the Vevo 2100 high-resolution ultrasound system, equipped with an MS550 transducer (Visualsonics, Toronto, ON, Canada). Mice were imaged under anesthesia. Initially, they were placed in an induction chamber with 3% isoflurane and 100% oxygen for 2-3 minutes, until the righting reflex was lost. Each mouse was then positioned supine on a heated platform, with their nose placed in a nosecone to maintain anesthesia with 2% isoflurane. Electrocardiogram (ECG) electrodes were attached to the limbs to monitor heart rate, and body temperature was maintained between 36-38°C using a rectal probe.
The aortic diameter at the sinus of Valsalva was measured using the B-mode aortic arch view. Pulse wave velocity (PWV) was determined from the B-mode and Doppler-mode aortic arch view, calculated using the formula: PWV (mm/s) = aortic arch distance (d2-d1) / transit time (T1-T2). The aortic arch distance (d2-d1) was measured between the two sample volume positions along the central axis of aortic arch in the B-mode image. Transit time (T1-T2) was derived from the Doppler waveform data.
Peak systolic velocity (PSV) is a parameter measured using Doppler ultrasound. On a Doppler waveform, PSV corresponds to the tallest “peaks” in the spectrum window, representing the maximum blood flow velocity during systole. The blood flow velocity can vary depending on vessel properties and pathological changes. For this study, the peak blood flow velocity of the coronary and pulmonary arteries was measured using the pulse wave (PW) Doppler-mode in the long axis view. The peak blood flow velocity of the left common carotid, posterior cerebral, and renal arteries was also measured using PW Doppler-mode. The left common carotid artery wall thickness and distensibility were assessed using M-Mode ultrasound imaging.
2.4. Statistical Analysis
Ultrasound imaging data were analyzed using Vevo LAB software (FUJIFILM, Toronto, ON, Canada), with measurements averaged over five cardiac cycles to minimize potential bias. Echocardiographic image acquisition was performed by a single investigator, while data analysis was conducted by two independent observers blinded to animal genotypes. Normality of the data for sinus of Valsalva diameter and aortic pulse wave velocity (PWV) was assessed using the Shapiro–Wilk and D’Agostino–Pearson normality tests in GraphPad Prism. All datasets passed the normality tests, allowing for parametric analysis. Statistical significance was determined using two-way ANOVA followed by Tukey’s multiple-comparison test. Data are reported as Mean ± SEM, with significance defied as P < 0.05.
The relationship between different measurements was determined using simple linear regression analysis in GraphPad Prism. The correlation coefficient (R-squared) was used to evaluate the quality of the model’s fit to the data and to quantify the proportion of variability in the dependent variable (Y) explained by the independent variable (X). R-squared values range from 0 to 1 and typically expressed as percentages, indicating the degree to which the independent variable accounts for the changes in the dependent variable (e.g., 0%-100%).
3. Results
3.1. Sex-Independent Increase of Aortic Root Diameter and Wall Stiffness in MFS Mice
Consistent with our previously published findings [9,10,27], this study also demonstrated a significant increase in the aortic root diameter at the sinus of Valsalva in MFS mice compared to Ctrl, in both female and male cohorts (Figure 1). Additionally, PWV, an index of arterial wall stiffness, was significantly increased in both male and female MFS groups compared to their respective Ctrl groups (Figure 2).
3.2. Increased Wall Stiffness and Thickness, and Reduced Distensibility in Common Carotid Artery in MFS Mice
The function of the left common carotid artery (LCCA) was assessed using in vivo ultrasound imaging by measuring pulse wave velocity (PWV), arterial wall thickness, and wall distensibility. PWV in the LCCA was significantly increased in both male and female MFS mice compared to their respective Ctrl groups, indicating elevated arterial stiffness (Figure 3). Consistently, there was also a marked increase in carotid artery wall thickness in the MFS groups compared to the Ctrl groups for both sexes (Figure 4A-B). As expected, these structural and functional changes were associated with a significant reduction in LCCA wall distensibility in both male and female MFS groups (Figure 4C).
3.3. Altered Peak Systolic Velocity in Coronary, Pulmonary, Rental, and Posterior Cerebral Arteries of MFS Mice
Blood flow reaches its maximum velocity within a vessel during the systole. Ultrasound Doppler (UD) measurements provide a reliable, non-invasive method to quantify blood flow by capturing the in vivo peak systolic velocity (PSV). PSV, which can serve as an indirect indicator of arterial wall stiffness, generally increases with progressive arterial narrowing due to elevated resistance. However, in the case of pre-occlusive lesions, PSV may paradoxically decrease as a result of critically reduced downstream flow.
In this study, no significant changes in coronary PSV were observed across experimental groups in both systole and diastole (Figure 5). A significant decrease in pulmonary artery PSV was noted in MFS mice compared to Ctrl mice, in both male and female groups. (Figure 6). PSV measurements in the right renal artery showed no statistical significance between Ctrl and MFS groups (Figure 7). In contrast, PSV in the posterior cerebral arteries was significantly reduced in both male and female MFS mice compared to their respective Ctrl groups (Figure 8).
3.4. Mild Aerobic Exercise Mitigates Arterial Pathology in MFS Mice
In this part of the study, we assessed the effects of mild aerobic exercise on vascular phenotypes in male and female MFS mice. Our findings revealed significant protective effects of exercise in attenuating aortic root pathology. Exercise training led to a reduction in the aortic root diameter at the sinus of Valsalva in both male and female MFS mice (Figure 9A-B). Notably, in exercised female MFS mice, the sinus of Valsalva diameter in exercised female MFS mice was restored to levels comparable to controls (Figure 9C). In parallel, aortic wall stiffness, as measured by pulse wave velocity (PWV), was significantly decreased in both sexes following exercise intervention (Figure 10A-C). In addition, In vivo ultrasound measurements showed that exercise improved peak blood flow velocity in the PCA of both male and female MFS mice (Figure 11A-B). Importantly, exercise training restored peak systolic velocity to control levels in both sexes MFS mice (Figure 11C).
Carotid artery measurements further underscored the positive impact of exercise training in MFS mice. In both male and female MFS mice, exercise significantly reduced PWV in LCCA, restoring it to levels observed in Ctrl groups (Figure 12A-C). Similarly, exercise led to a reduction in carotid artery wall thickness in both sexes, with measurements reaching those of healthy Ctrl mice (Figure 13A-C). Moreover, LCCA wall distensibility was markedly improved in exercised MFS mice of both sexes (Figure 14A,B), with values returning to Ctrl levels (Figure 14C).
3.5. Sex-Specific Association Between Aortic Pathology and Cerebral Blood Flow in MFs Mice
Two linear regression analyses were performed to explore the relationships between key aortic pathology parameters and cerebral blood flow. The first analysis examined the relationship between aortic PWV (wall stiffness) and the posterior cerebral artery peak blood flow. In females, the model explained 50% of the variation in posterior cerebral artery peak blood flow (R² = 0.50), indicating a moderate association between these variables (Figure 15A). In males, the relationship was stronger, with the model accounting for 74% of the variation in posterior cerebral artery peak blood flow (R² = 0.74), demonstrating a strong association (Figure 15B).
A second analysis examined the relationship between the sinus of Valsalva diameter and posterior cerebral artery peak blood flow velocity. In females, the model yielded an R² value of 0.50, consistent with the first analysis, indicating a moderate association (Figure 15C). In males, the relationship was stronger, with an R² value of 0.74, reflecting a robust predictive relationship between these variables (Figure 15D). These findings underscore sex-dependent differences in the strength of the relationships. In both analyses, male MFS mice exhibited stronger associations between the respective cardiovascular parameters, while the relationships in female MFS mice were more moderate.
3.6. Sex-Specific Association Between Aortic and Carotid Artery Pathology in MFs Mice
A linear regression analysis was conducted to examine the relationship between aortic PWV and carotid artery parameters, including PWV, wall thickness, and distensibility, with the analysis stratified by sex. Using carotid PWV as a predictor, the model explained 74% of the variation in aortic PWV in females (R² = 0.74) (Figure 16A) and 73% in males (R² = 0.73) (Figure 16B), indicating a strong and comparable predictive relationship in both sexes. When carotid artery wall thickness was used as a predictor, the explanatory power remained high in females (R² = 0.74) (Figure 16C) but was slightly weaker in males (R² = 0.58) (Figure 16D), though the relationship remained significant. In contrast, carotid artery distensibility showed a weaker association with aortic PWV in females (R² = 0.39) (Figure 16E) but a stronger relationship in males (R² = 0.59) (Figure 16F). These results highlight sex-specific differences in the predictive value of carotid artery parameters, with males exhibiting stronger associations for carotid distensibility and females showing more consistent relationships across all parameters.
In addition, a linear regression analysis was performed to investigate the relationships between the sinus of Valsalva diameter and various carotid artery parameters. In female MFS mice, the results revealed a moderate relationship between the sinus of Valsalva diameter and carotid artery PWV, with an R² value of 0.55, indicating that 55% of the variation in carotid artery PWV is explained by the changes in sinus of Valsalva diameter (Figure 17A). The association between the sinus of Valsalva diameter and carotid artery wall thickness was stronger in males, with an R² value of 0.72, suggesting that 72% of the variation in carotid artery PWV can be attributed to the changes in sinus of Valsalva diameter (Figure 17B).
The association between the sinus of Valsalva diameter and carotid artery wall thickness was moderately strong in females, with an R² value of 0.73, suggesting that 73% of the variation in wall thickness can be attributed to the sinus of Valsalva diameter (Figure 17C). In males, the relationship was slightly weaker, with an R² value of 0.66, indicating a notable but less robust association compared to females (Figure 17D).
The relationship between the sinus of Valsalva diameter and carotid artery distensibility was weaker overall. In females, the model accounted for only 32% of the variation in carotid artery distensibility (R² = 0.32), reflecting a modest association (Figure 17E) Conversely, in males, the relationship was stronger, with an R² value of 0.58, demonstrating a more substantial but still moderate predictive strength (Figure 17F). These findings suggest that the diameter at the sinus of Valsalva is a strong predictor of carotid artery PWV and wall thickness, while showing a more moderate predictive value for carotid artery distensibility. Notably, sex-specific differences were observed, with females exhibiting stronger correlations between aortic root diameter and carotid wall thickness, but weaker associations with carotid distensibility compared to their male counterparts.
3.7. Sex-Specific Association Between Aortic and Pulmonary Artery Pathology in MFs Mice
In females, the model explained 46% of the variation in pulmonary artery peak blood flow (R² = 0.46), indicating a moderate association between these variables (Figure 18A). In contrast, as significantly stronger relationship was observed in males (R² = 0.80), where aortic PWV accounted for 80% of the variation in pulmonary artery peak velocity (Figure 18B). The relationship between the aortic root diameter at the sinus of Valsalva and pulmonary artery peak blood flow velocity was consistent across sexes, with an R² value of 0.50 for both females (Figure 18C) and males (Figure 18D), indicating a moderate predictive relationship in each group. These findings suggest that aortic PWV is a strong predictor of pulmonary artery hemodynamics in males, while its predictive value is more modest in females. In contrast, the sinus of Valsalva diameter exhibits a moderate and comparable association with pulmonary artery peak blood flow velocity in both sexes.
4. Discussion
This study investigated cardiovascular structural and functional changes in a mouse model of MFS and evaluated the therapeutic potential of mild exercise training. Our findings provide valuable insights into MFS pathology and demonstrate the beneficial effects of exercise on vascular health, emphasizing the importance of early intervention strategies to manage cardiovascular complications in MFS.
Our data revealed a significant increase in aortic root diameter and pulse wave velocity (PWV) in both male and female MFS mice, consistent with previous studies identifying aortic root dilation and increased arterial wall stiffness as key features of MFS [10,12,27,28,29,30,31]. Aortic root dilation, particularly at the sinus of Valsalva, is a well-established hallmark of MFS and is associated with increased risk of aortic aneurysms and dissection. Similarly, the elevated PWV observed in MFS mice indicates impaired vascular compliance, a key contributor to the elevated cardiovascular morbidity in MFS patients. These findings were corroborated by high-resolution ultrasound imaging, which revealed significant structural and functional alterations in the carotid arteries. In the left common carotid artery (LCCA), both male and female MFS mice exhibited increased PWV and wall thickness, as well as reduced distensibility. These changes reflect vascular remodeling and a reduction in elastic decreased elastic properties, consistent with early-stage atherosclerosis or generalized arterial stiffness. Such alterations may impair blood flow and contribute to the increase in cardiovascular risk in MFS.
The reduced peak systolic velocity (PSV) observed in several arteries, including the posterior cerebral and pulmonary arteries, underscores the functional consequences of the structural alterations seen in MFS mice aorta. Notably, the decrease in PSV in the posterior cerebral artery may have significant implications for cerebral perfusion, which is essential for maintaining normal brain function. The reduction in carotid and PCA blood flow velocities may have important cerebrovascular consequences, particularly with the consideration of our previous reports showing increased blood-brain barrier (BBB) permeability, heightened microglial activation, and impaired cognitive function in MFS. In our previous report, we demonstrated that 6-month-old MFS mice exhibited a significantly increased number of microglia in the hippocampus compared to age-matched control mice. This microglial profile closely resembled that observed in 12-month-old healthy control mice. Additionally, 6 -month-old MFS mice showed elevated levels of apoptosis relative to sex- and age-matched controls. Collectively, these findings suggested that MFS mice displayed features consistent with a premature brain aging phenotype [12,32].
Increased aortic PWV reflects central arterial stiffness, which can lead to enhanced transmission of pulsatile energy into cerebral microcirculation due to reduced cushioning from the elastic central arteries. When this occurs in the setting of decreased carotid artery distensibility and flow, the brain is subjected to greater hemodynamic stress, which has been shown to damage the microvasculature, disrupt the BBB, and promote microglial activation—processes linked to neurodegenerative diseases and cognitive decline [33].
Furthermore, the increased carotid wall thickness and reduced distensibility suggest early vascular remodeling, possibly driven by inflammation, altered smooth muscle cell phenotype, or fibrotic changes in response to abnormal TGF-β signaling—commonly dysregulated in MFS [34]. These vascular changes may impede blood delivery to the brain and reduce cerebral perfusion reserve, predisposing MFS mice to hypoperfusion-induced BBB breakdown, as we previously reported [12]. These results underscore the importance of evaluating cerebral hemodynamics and vascular-brain interactions in the context of MFS, particularly in light of emerging evidence that cognitive symptoms and neurovascular dysfunction may be under-recognized clinical features in this population [35].
While no significant changes were detected in the coronary or renal arteries, the reduction in pulmonary artery PSV highlights the systemic nature of vascular dysfunction in this model. Pulmonary involvement in MFS includes structural abnormalities such as emphysema and an increased risk of spontaneous pneumothorax [23,36,37,38]. These complications are believed to arise from the defective connective tissue associated with FBN1 mutations, leading to weakened alveolar walls and reduced lung elasticity. Such structural changes can elevate pulmonary vascular resistance, potentially contributing to pulmonary hypertension and altered pulmonary artery flow dynamics. Moreover, studies have reported dilation of the main pulmonary artery in a significant proportion of MFS patients. For instance, one study found that 69.4% of MFS patients exhibited pulmonary artery dilation, with 15.3% developing aneurysms [39]. Although the clinical implications of pulmonary artery dilation in MFS are not fully understood, such vascular alterations could influence pulmonary hemodynamics and contribute to the observed decrease in PSV.
The observed reduction in PSV may also reflect impaired right ventricular function. Research indicates that individuals with MFS can experience the right ventricular systolic dysfunction, which could lead to decreased blood flow velocity through the pulmonary artery [36]. Additionally, skeletal deformities common in MFS, such as scoliosis and chest wall abnormalities, can compromise respiratory mechanics and contribute to restrictive lung disease. These musculoskeletal issues may further exacerbate pulmonary complications and impact pulmonary artery hemodynamics [40,41]. These findings underscore the importance of comprehensive pulmonary assessments in MFS patients to detect and manage potential complications early.
Exercise training proved to be a promising intervention for mitigating cardiovascular pathology in MFS mice [8,10,12,27]. The results of our study clearly confirm that mild aerobic exercise significantly reduces both aortic root diameter and PWV, suggesting improved aortic compliance and decreased vascular wall stiffness. Particularly, in female MFS mice, the aortic root diameter was restored to control levels following exercise training, suggesting that exercise may fully reverse some of the pathological structural changes associated with the condition in a sex-dependent manner. These results align with previous studies in other animal models and human populations, where exercise has been shown to improve vascular function, enhance endothelial health, and reduce arterial stiffness [42,43,44,45,46,47,48,49]. The beneficial effects of exercise extended to carotid artery function as well. Exercise training reduced PWV, decreased wall thickness, and improved distensibility of left common carotid artery in both male and female MFS mice. These improvements paralleled the observed benefits in aortic function, further supporting the therapeutic potential of exercise for managing vascular health in MFS.
Furthermore, exercise training improved peak systolic velocity in the posterior cerebral artery, restoring it to control levels in female mice. This result suggests that exercise may enhance cerebral perfusion and provide protection against potential cerebrovascular complications associated with MFS. Considering the increased risk of stroke and other cerebrovascular events in individuals with MFS, this finding highlights the importance of maintaining healthy cerebral circulation as a key aspect of disease management.
Sex-specific differences were also evident in the relationships between cardiovascular parameters, with stronger associations observed between aortic PWV, aortic root diameter at the sinus of Valsalva, and blood flow velocity in males compared to females. These differences may reflect inherent biological variations in vascular function between sexes, as well as potential differences in their responses to exercise training. For instance, while the relationship between aortic PWV and cerebral blood flow was more robust in males, the improvement in carotid artery function following exercise was more pronounced in females, suggesting that the two sexes may drive distinct benefits from exercise. Studies have shown that hypertension and coronary artery disease are more prevalent in men and postmenopausal women compared to premenopausal women. This difference is partly attributed to gender-based variations in vascular tone and the potential vascular protective effects of female sex hormones, particularly estrogen and progesterone. The male sex hormone testosterone may also influence vascular function. Receptors for estrogen, progesterone, and testosterone have been identified in the blood vessels of humans and other mammals and are located in the plasmalemma, cytosol, and nuclei of various vascular cells, including endothelial and smooth muscle cells.
The interaction of sex hormones with cytosolic and nuclear receptors can trigger long-term genomic effects—such as promoting endothelial cell growth and inhibiting smooth muscle cell proliferation. In contrast, activation of membrane-bound (plasmalemmal) sex hormone receptors may lead to rapid, non-genomic effects that stimulate endothelium-dependent vascular relaxation mechanisms, including the nitric oxide-cyclic GMP, prostacyclin-cyclic AMP, and hyperpolarization pathways. Additionally, sex hormones may exert endothelium-independent effects by inhibiting signaling pathways involved in vascular smooth muscle contraction, such as intracellular calcium levels and protein kinase C activity. Together, these hormone-induced mechanisms of promoting vascular relaxation and reducing smooth muscle contraction may contribute to the observed gender differences in vascular tone and support the potential vascular benefits of hormone replacement therapy in individuals with natural or surgically induced hormone deficiencies [44,50,51]. Exercise increases blood flow and laminar shear stress, which stimulates NO production by the endothelium. NO then acts as a vasodilator, relaxing blood vessels and improving blood flow. Mild exercise improves endothelium-mediated nitric oxide (NO) production and vasorelaxation, but the magnitude of this effect can vary between sexes [46,52,53]. These findings underscore the need for personalized approaches in the management of MFS, especially in clinical settings where sex-specific factors may influence treatment outcomes.
Our study also identified significant associations between aortic parameters and carotid artery function, particularly in females. The strong correlation between the sinus of Valsalva diameter and carotid artery PWV and wall thickness, along with a moderate relationship with distensibility, suggests that changes in the aorta may serve as reliable predictors of carotid artery function in MFS. These findings are crucial for understanding how aortic pathology in MFS can extend to other vascular territories, potentially leading to widespread vascular dysfunction.
The molecular mechanisms responsible for the observed improvements in vascular function were not investigated in this study, but our previous data on MFS and exercise showed that mild aerobic exercise decreased phosphorylated Smad proteins (p-Smads), which are involved in the TGF-β signaling pathway, without necessarily decreasing the total levels of TGF-beta. This means that exercise can affect the downstream signaling of TGF-β without altering the overall amount of the signaling molecules. Further research is necessary to uncover the signaling pathways and molecular factors driving exercise-induced vascular remodeling, which could inform and enhance therapeutic strategies for MFS and other related connective tissue disorders.
5. Limitations
Hormonal fluctuations, particularly in estrogen and progesterone, during the menstrual or estrous cycle, significantly impact a wide range of physiological and behavioral outcomes, making them important considerations in biomedical research. Estrogen receptors and other factors modulate endothelial function, which can be affected by the estrous cycle. This could also explain the variability among female subjects and impact the data [54,55].
6. Conclusions
In conclusion, our study highlights the significant cardiovascular alterations observed in MFS mice, including increased aortic wall stiffness, dilation, and impaired blood flow. Importantly, these changes were shown to be reversible with mild exercise training. These findings suggest that exercise may offer a promising and non-invasive intervention to enhance vascular function and mitigate the risk of cardiovascular complications in MFS. Further research is needed to better elucidate the underlying mechanisms and to investigate the translational potential of exercise as a therapeutic strategy for individuals with MFS and related connective tissue disorders.
7. Clinical Implications & Future Directions
Moderate-intensity exercise may provide therapeutic benefits by enhancing eNOS-mediated NO production, improving endothelial function, and mitigating TGF-β/MMP-driven vascular pathology. Pharmacological Synergy: Combining exercise with treatments like angiotensin receptor blockers (ARBs), which also suppress TGF-β signaling, may offer synergistic protective effects. Targeting eNOS Pathways: Enhancing eNOS activity pharmacologically or via lifestyle interventions could serve as a novel therapeutic strategy for vascular complications in Marfan syndrome. Another consideration for future studies would be to consider the estrous cycle in female mice during sampling.
Funding
This project was funded by a National Institutes of Health (R15HL145646) to M.E.
References
- Seeburun S, Wu S, Hemani D, et al. Insights into elastic fiber fragmentation: Mechanisms and treatment of aortic aneurysm in Marfan syndrome. Vascul Pharmacol. Dec 2023;153:107215. [CrossRef]
- Bonfioli GB, Rodella L, Rosati R, Carrozza A, Metra M, Vizzardi E. Aortopathies: From Etiology to the Role of Arterial Stiffness. J Clin Med. Jun 9 2023;12(12). [CrossRef]
- Pyeritz RE. Marfan syndrome: 30 years of research equals 30 years of additional life expectancy. Heart. Mar 2009;95(3):173-5. [CrossRef]
- Coelho SG, Almeida AG. Marfan syndrome revisited: From genetics to the clinic. Rev Port Cardiol (Engl Ed). Apr 2020;39(4):215-226. Sindrome de Marfan revisitada - da genetica a clinica. [CrossRef]
- Zeigler SM, Sloan B, Jones JA. Pathophysiology and Pathogenesis of Marfan Syndrome. Adv Exp Med Biol. 2021;1348:185-206. [CrossRef]
- Marelli S, Micaglio E, Taurino J, et al. Marfan Syndrome: Enhanced Diagnostic Tools and Follow-up Management Strategies. Diagnostics (Basel). Jul 5 2023;13(13). [CrossRef]
- Selamet Tierney ES, Chung S, Stauffer KJ, et al. Can 10 000 Healthy Steps a Day Slow Aortic Root Dilation in Pediatric Patients With Marfan Syndrome? J Am Heart Assoc. Dec 6 2022;11(23):e027598. [CrossRef]
- Jouini S, Milleron O, Eliahou L, Jondeau G, Vitiello D. Is physical activity a future therapy for patients with Marfan syndrome? Orphanet J Rare Dis. Feb 10 2022;17(1):46. [CrossRef]
- Gibson C, Nielsen C, Alex R, et al. Mild aerobic exercise blocks elastin fiber fragmentation and aortic dilatation in a mouse model of Marfan syndrome associated aortic aneurysm. J Appl Physiol (1985). Jul 1 2017;123(1):147-160. [CrossRef]
- Esfandiarei M, Hoxha B, Talley NA, et al. Beneficial effects of resveratrol and exercise training on cardiac and aortic function and structure in the 3xTg mouse model of Alzheimer’s disease. Drug Des Devel Ther. 2019;13:1197-1211. [CrossRef]
- Agg B, Szilveszter B, Daradics N, et al. Increased visceral arterial tortuosity in Marfan syndrome. Orphanet J Rare Dis. Apr 15 2020;15(1):91. [CrossRef]
- Curry T, Barrameda ME, Thomas TC, Esfandiarei M. In vivo phenotypic vascular dysfunction extends beyond the aorta in a mouse model for fibrillin-1 (Fbn1) mutation. Sci Rep. Mar 9 2024;14(1):5779. [CrossRef]
- Pyeritz RE. Marfan syndrome: improved clinical history results in expanded natural history. Genet Med. Aug 2019;21(8):1683-1690. [CrossRef]
- Pyeritz RE. Marfan Syndrome: Expanding the Cardiovascular Phenotype? J Am Coll Cardiol. Jun 22 2021;77(24):3013-3015. [CrossRef]
- Spinardi L, Vornetti G, De Martino S, et al. Intracranial Arterial Tortuosity in Marfan Syndrome and Loeys-Dietz Syndrome: Tortuosity Index Evaluation Is Useful in the Differential Diagnosis. AJNR Am J Neuroradiol. Oct 2020;41(10):1916-1922. [CrossRef]
- Ekhator C, Devi M, Barker C, et al. Arterial Tortuosity Syndrome: Unraveling a Rare Vascular Disorder. Cureus. Sep 2023;15(9):e44906. [CrossRef]
- Parlapiano G, Di Lorenzo F, Salehi LB, Ruvolo G, Novelli G, Sangiuolo F. Neurovascular manifestations in connective tissue diseases: The case of Marfan Syndrome. Mech Ageing Dev. Oct 2020;191:111346. [CrossRef]
- Eberth JF, Taucer AI, Wilson E, Humphrey JD. Mechanics of carotid arteries in a mouse model of Marfan Syndrome. Ann Biomed Eng. Jun 2009;37(6):1093-104. [CrossRef]
- Latter DA, Ricci MA, Forbes RD, Graham AM. Internal carotid artery aneurysm and Marfan’s syndrome. Can J Surg. Nov 1989;32(6):463-6.
- Ito M, Kuroda S, Nakayama N, Iwasaki M, Shiiya N, Iwasaki Y. [Extracranial internal carotid artery aneurysm associated with Marfan syndrome: case report]. No Shinkei Geka. Aug 2007;35(8):793-7.
- Kim ST, Cloft H, Flemming KD, Kallmes DF, Lanzino G, Brinjikji W. Increased Prevalence of Cerebrovascular Disease in Hospitalized Patients with Marfan Syndrome. J Stroke Cerebrovasc Dis. Feb 2018;27(2):296-300. [CrossRef]
- Schoenhoff FS, Yildiz M, Langhammer B, et al. The fate of nonaortic arterial segments in Marfan patients. J Thorac Cardiovasc Surg. Jun 2019;157(6):2150-2156. [CrossRef]
- Lundby R, Rand-Hendriksen S, Hald JK, Pripp AH, Smith HJ. The pulmonary artery in patients with Marfan syndrome: a cross-sectional study. Genet Med. Nov 2012;14(11):922-7. [CrossRef]
- Tanaka H, Dinenno FA, Monahan KD, Clevenger CM, DeSouza CA, Seals DR. Aging, habitual exercise, and dynamic arterial compliance. Circulation. Sep 12 2000;102(11):1270-5. [CrossRef]
- Dyakova EY, Kapilevich LV, Shylko VG, Popov SV, Anfinogenova Y. Physical exercise associated with NO production: signaling pathways and significance in health and disease. Front Cell Dev Biol. 2015;3:19. [CrossRef]
- Higashi, Y. Exercise is a double-edged sword for endothelial function. Hypertens Res. Feb 2016;39(2):61-3. [CrossRef]
- Lee L, Cui JZ, Cua M, et al. Aortic and Cardiac Structure and Function Using High-Resolution Echocardiography and Optical Coherence Tomography in a Mouse Model of Marfan Syndrome. PLoS One. 2016;11(11):e0164778. [CrossRef]
- Sellers SL, Milad N, Chan R, et al. Inhibition of Marfan Syndrome Aortic Root Dilation by Losartan: Role of Angiotensin II Receptor Type 1-Independent Activation of Endothelial Function. Am J Pathol. Mar 2018;188(3):574-585. [CrossRef]
- Tehrani AY, White Z, Milad N, Esfandiarei M, Seidman MA, Bernatchez P. Blood pressure-independent inhibition of Marfan aortic root widening by the angiotensin II receptor blocker valsartan. Physiol Rep. May 2021;9(10):e14877. [CrossRef]
- Salvi P, Grillo A, Marelli S, et al. Aortic dilatation in Marfan syndrome: role of arterial stiffness and fibrillin-1 variants. J Hypertens. Jan 2018;36(1):77-84. [CrossRef]
- Schafer M, Browne LP, Truong U, et al. Aortic stiffness in adolescent Turner and Marfan syndrome patients. Eur J Cardiothorac Surg. Nov 1 2018;54(5):926-932. [CrossRef]
- Esfandiarei M, Anwar F, Nuthi M, et al. Apoptosis is increased in cortical neurons of female Marfan Syndrome mice. bioRxiv. Oct 17 2024. [CrossRef]
- Caughey MC, Qiao Y, Meyer ML, et al. Relationship Between Central Artery Stiffness, Brain Arterial Dilation, and White Matter Hyperintensities in Older Adults: The ARIC Study-Brief Report. Arterioscler Thromb Vasc Biol. Jun 2021;41(6):2109-2116. [CrossRef]
- Rensma SP, Stehouwer CDA, Van Boxtel MPJ, et al. Associations of Arterial Stiffness With Cognitive Performance, and the Role of Microvascular Dysfunction: The Maastricht Study. Hypertension. Jun 2020;75(6):1607-1614. [CrossRef]
- Neptune ER, Frischmeyer PA, Arking DE, et al. Dysregulation of TGF-beta activation contributes to pathogenesis in Marfan syndrome. Nat Genet. Mar 2003;33(3):407-11. [CrossRef]
- Sheikhzadeh S, De Backer J, Gorgan NR, et al. The main pulmonary artery in adults: a controlled multicenter study with assessment of echocardiographic reference values, and the frequency of dilatation and aneurysm in Marfan syndrome. Orphanet J Rare Dis. Dec 10 2014;9:203. [CrossRef]
- Mariucci E, Bonori L, Lovato L, et al. Coronary Artery Aneurysms in Patients With Marfan Syndrome: Frequent, Progressive, and Relevant. Can J Cardiol. Aug 2021;37(8):1225-1231. [CrossRef]
- Sedky K, Gaisl T, Bennett DS. Prevalence of Obstructive Sleep Apnea in Joint Hypermobility Syndrome: A Systematic Review and Meta-Analysis. J Clin Sleep Med. Feb 15 2019;15(2):293-299. [CrossRef]
- Benke K, Agg B, Polos M, et al. The effects of acute and elective cardiac surgery on the anxiety traits of patients with Marfan syndrome. BMC Psychiatry. Jul 17 2017;17(1):253. [CrossRef]
- Wang Y, Wang D, Zhang G, et al. Effects of spinal deformities on lung development in children: a review. J Orthop Surg Res. Mar 27 2023;18(1):246. [CrossRef]
- Otremski H, Widmann RF, Di Maio MF, Ovadia D. The correlation between spinal and chest wall deformities and pulmonary function in Marfan syndrome. J Child Orthop. Aug 1 2020;14(4):343-348. [CrossRef]
- Mas-Stachurska A, Siegert AM, Batlle M, et al. Cardiovascular Benefits of Moderate Exercise Training in Marfan Syndrome: Insights From an Animal Model. J Am Heart Assoc. Sep 25 2017;6(9). [CrossRef]
- Lan Y, Wu R, Feng Y, et al. Effects of Exercise on Arterial Stiffness: Mechanistic Insights into Peripheral, Central, and Systemic Vascular Health in Young Men. Metabolites. Mar 1 2025;15(3). [CrossRef]
- Green DJ, Smith KJ. Effects of Exercise on Vascular Function, Structure, and Health in Humans. Cold Spring Harb Perspect Med. Apr 2 2018;8(4). [CrossRef]
- Kresnajati S, Lin YY, Mundel T, Bernard JR, Lin HF, Liao YH. Changes in Arterial Stiffness in Response to Various Types of Exercise Modalities: A Narrative Review on Physiological and Endothelial Senescence Perspectives. Cells. Nov 9 2022;11(22). [CrossRef]
- Moreau KL, Clayton ZS, DuBose LE, Rosenberry R, Seals DR. Effects of regular exercise on vascular function with aging: Does sex matter? Am J Physiol Heart Circ Physiol. Jan 1 2024;326(1):H123-H137. [CrossRef]
- Noma K, Kihara Y, Higashi Y. Outstanding Effect of Physical Exercise on Endothelial Function Even in Children and Adolescents. Circ J. Apr 25 2017;81(5):637-639. [CrossRef]
- Di Francescomarino S, Sciartilli A, Di Valerio V, Di Baldassarre A, Gallina S. The effect of physical exercise on endothelial function. Sports Med. 2009;39(10):797-812. [CrossRef]
- Ramirez-Velez R, Aguilar AC, Mosquera M, Garcia RG, Reyes LM, Lopez-Jaramillo P. Clinical trial to assess the effect of physical exercise on endothelial function and insulin resistance in pregnant women. Trials. Nov 17 2009;10:104. [CrossRef]
- Pataky MW, Dasari S, Michie KL, et al. Impact of biological sex and sex hormones on molecular signatures of skeletal muscle at rest and in response to distinct exercise training modes. Cell Metab. Nov 7 2023;35(11):1996-2010 e6. [CrossRef]
- Orshal JM, Khalil RA. Gender, sex hormones, and vascular tone. Am J Physiol Regul Integr Comp Physiol. Feb 2004;286(2):R233-49. [CrossRef]
- Niebauer J, Cooke JP. Cardiovascular effects of exercise: role of endothelial shear stress. J Am Coll Cardiol. Dec 1996;28(7):1652-60. [CrossRef]
- Moreau KL, Ozemek C. Vascular Adaptations to Habitual Exercise in Older Adults: Time for the Sex Talk. Exerc Sport Sci Rev. Apr 2017;45(2):116-123. [CrossRef]
- Abo S, Smith D, Stadt M, Layton A. Modelling female physiology from head to Toe: Impact of sex hormones, menstrual cycle, and pregnancy. J Theor Biol. May 7 2022;540:111074. [CrossRef]
- Kehmeier MN, Bedell BR, Cullen AE, et al. In vivo arterial stiffness, but not isolated artery endothelial function, varies with the mouse estrous cycle. Am J Physiol Heart Circ Physiol. Dec 1 2022;323(6):H1057-H1067. [CrossRef]
Figure 1.
Aortic root diameter measurements at the Sinus of Valsalva. A) Representative B-mode ultrasound image showing measurements of three aortic root diameters in a 7-month-old Ctrl mouse. B) Representative B-mode ultrasound image illustrating the three aortic root diameters in a 7-month-old mouse with MFS. C) Ultrasound analysis reveals a significant increase in the diameter of the Sinus of Valsalva in both male and female MFS mice compared to age- and sex-matched Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
Figure 1.
Aortic root diameter measurements at the Sinus of Valsalva. A) Representative B-mode ultrasound image showing measurements of three aortic root diameters in a 7-month-old Ctrl mouse. B) Representative B-mode ultrasound image illustrating the three aortic root diameters in a 7-month-old mouse with MFS. C) Ultrasound analysis reveals a significant increase in the diameter of the Sinus of Valsalva in both male and female MFS mice compared to age- and sex-matched Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
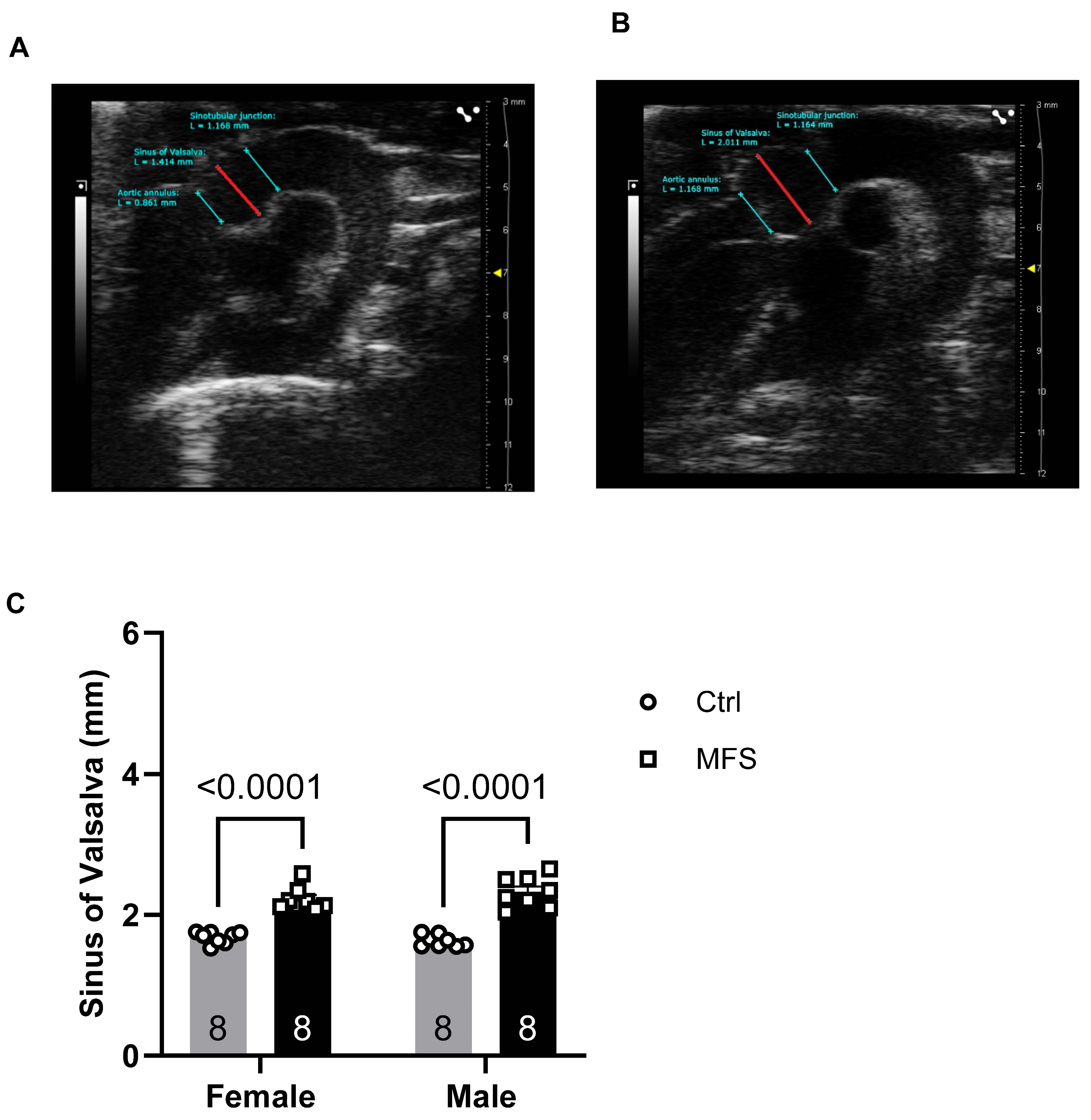
Figure 2.
Pulse wave velocity (PWV) measurements of thoracic aorta. A) PW Doppler tracing of the ascending (left panel) and descending aorta (right panel). B) Pulse wave Doppler tracing of the ascending and descending aorta. C) Thoracic aorta PWV, an indicator of aortic wall stiffness, exhibited a significant increase in 7-month-old mice with MFS compared to age- and sex-matched Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
Figure 2.
Pulse wave velocity (PWV) measurements of thoracic aorta. A) PW Doppler tracing of the ascending (left panel) and descending aorta (right panel). B) Pulse wave Doppler tracing of the ascending and descending aorta. C) Thoracic aorta PWV, an indicator of aortic wall stiffness, exhibited a significant increase in 7-month-old mice with MFS compared to age- and sex-matched Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
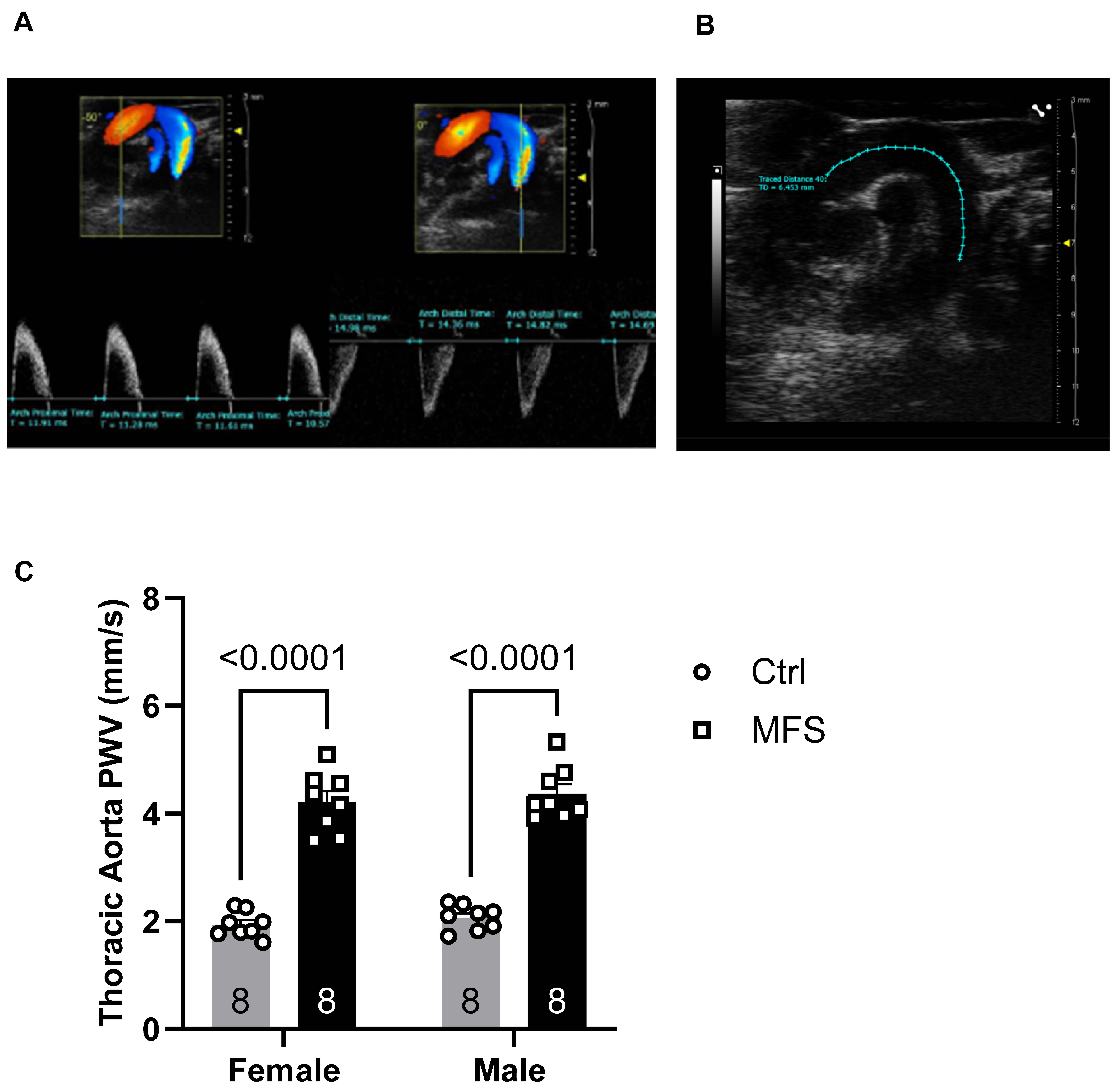
Figure 3.
Measurements of PWV of left common carotid artery (LCCA). A) Doppler mode, ultrasound image of the left common carotid artery. B) PWV was significantly increased in both male and female MFS mice compared to the age- and sex-matched Ctrl groups. (Mean ± SEM, p < 0.05 is considered significant).
Figure 3.
Measurements of PWV of left common carotid artery (LCCA). A) Doppler mode, ultrasound image of the left common carotid artery. B) PWV was significantly increased in both male and female MFS mice compared to the age- and sex-matched Ctrl groups. (Mean ± SEM, p < 0.05 is considered significant).
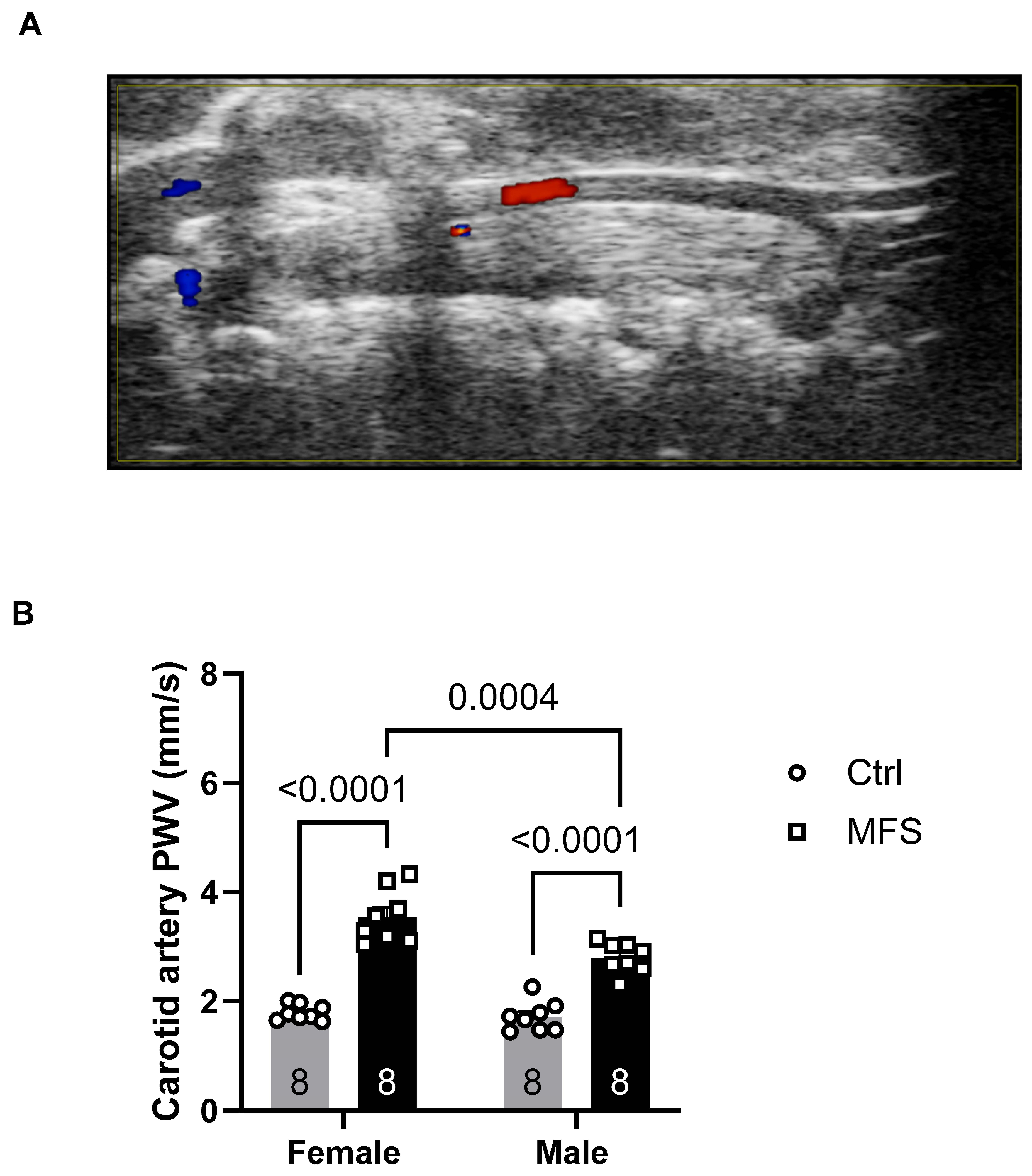
Figure 4.
Ultrasound measurements of the LCCA wall thickness and distensibility. A) M-Mode ultrasound view of the LCCA in a 7-month-old Ctrl mouse. B) Data showed a significant increase in the wall thickness and C) a significant decrease in the distensibility in LCCA in MFS mice compared to age- and sex-matched Ctrl. (Mean ± SEM, p < 0.05 is considered significant).
Figure 4.
Ultrasound measurements of the LCCA wall thickness and distensibility. A) M-Mode ultrasound view of the LCCA in a 7-month-old Ctrl mouse. B) Data showed a significant increase in the wall thickness and C) a significant decrease in the distensibility in LCCA in MFS mice compared to age- and sex-matched Ctrl. (Mean ± SEM, p < 0.05 is considered significant).

Figure 5.
Measurements of the carotid artery peak systolic velocity (PSV). A) Ultrasound PW Color Doppler mode view of the coronary artery in a 7-month-old Ctrl mouse. B) Data showed no significant changes in coronary artery peak systolic velocity between MFS and age- and sex-matched Ctrl groups. (Mean ± SEM, p < 0.05 is considered significant).
Figure 5.
Measurements of the carotid artery peak systolic velocity (PSV). A) Ultrasound PW Color Doppler mode view of the coronary artery in a 7-month-old Ctrl mouse. B) Data showed no significant changes in coronary artery peak systolic velocity between MFS and age- and sex-matched Ctrl groups. (Mean ± SEM, p < 0.05 is considered significant).
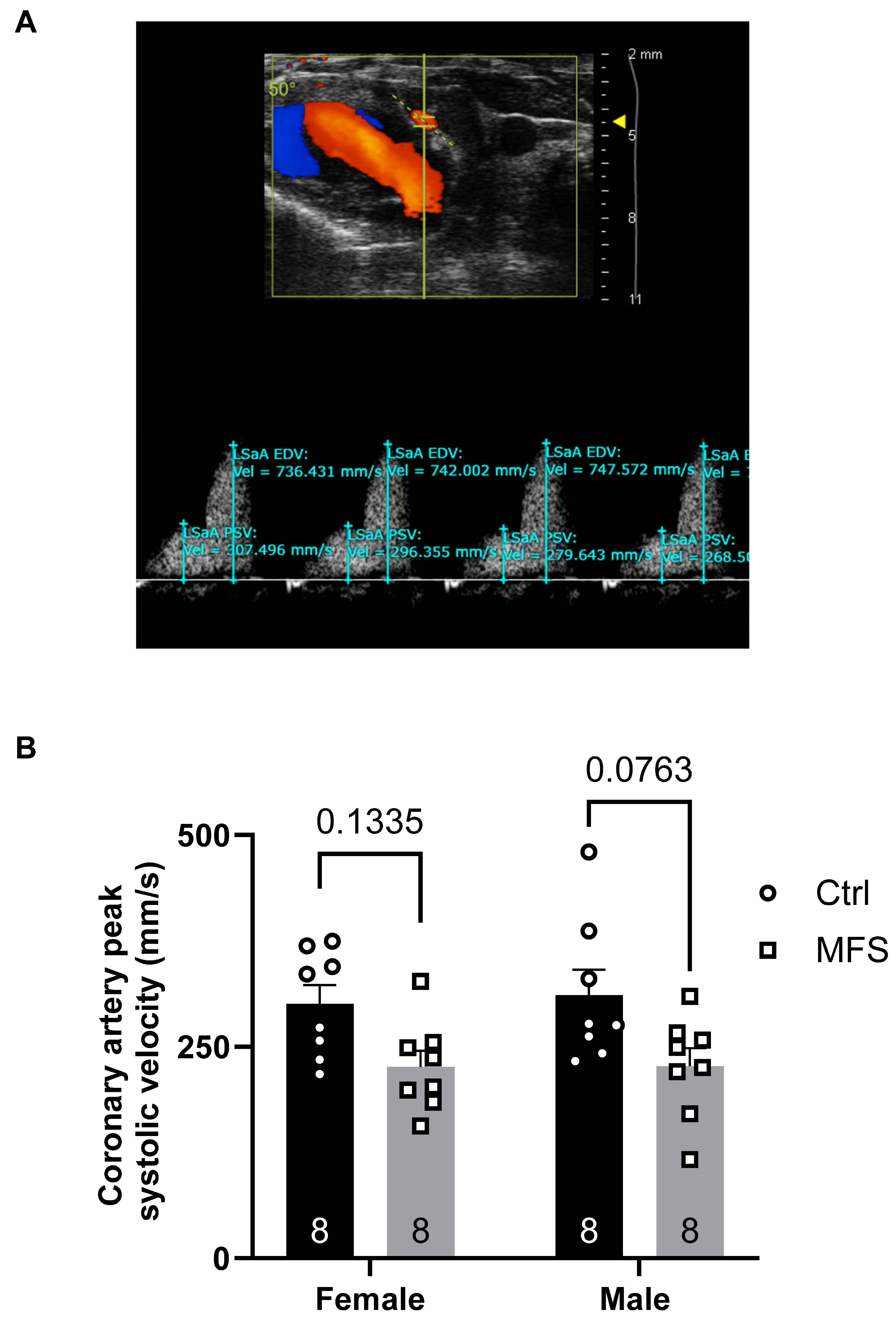
Figure 6.
Measurement of the pulmonary artery peak blood flow velocity. A) Ultrasound PW Color Doppler mode view of the coronary artery in a 7-month-old Ctrl mouse. B) A significant decrease was observed in the pulmonary artery peak systolic velocity in MFS mice compared to age- and sex-matched Ctrl in groups. (Mean ± SEM, p < 0.05 is considered as significant).
Figure 6.
Measurement of the pulmonary artery peak blood flow velocity. A) Ultrasound PW Color Doppler mode view of the coronary artery in a 7-month-old Ctrl mouse. B) A significant decrease was observed in the pulmonary artery peak systolic velocity in MFS mice compared to age- and sex-matched Ctrl in groups. (Mean ± SEM, p < 0.05 is considered as significant).
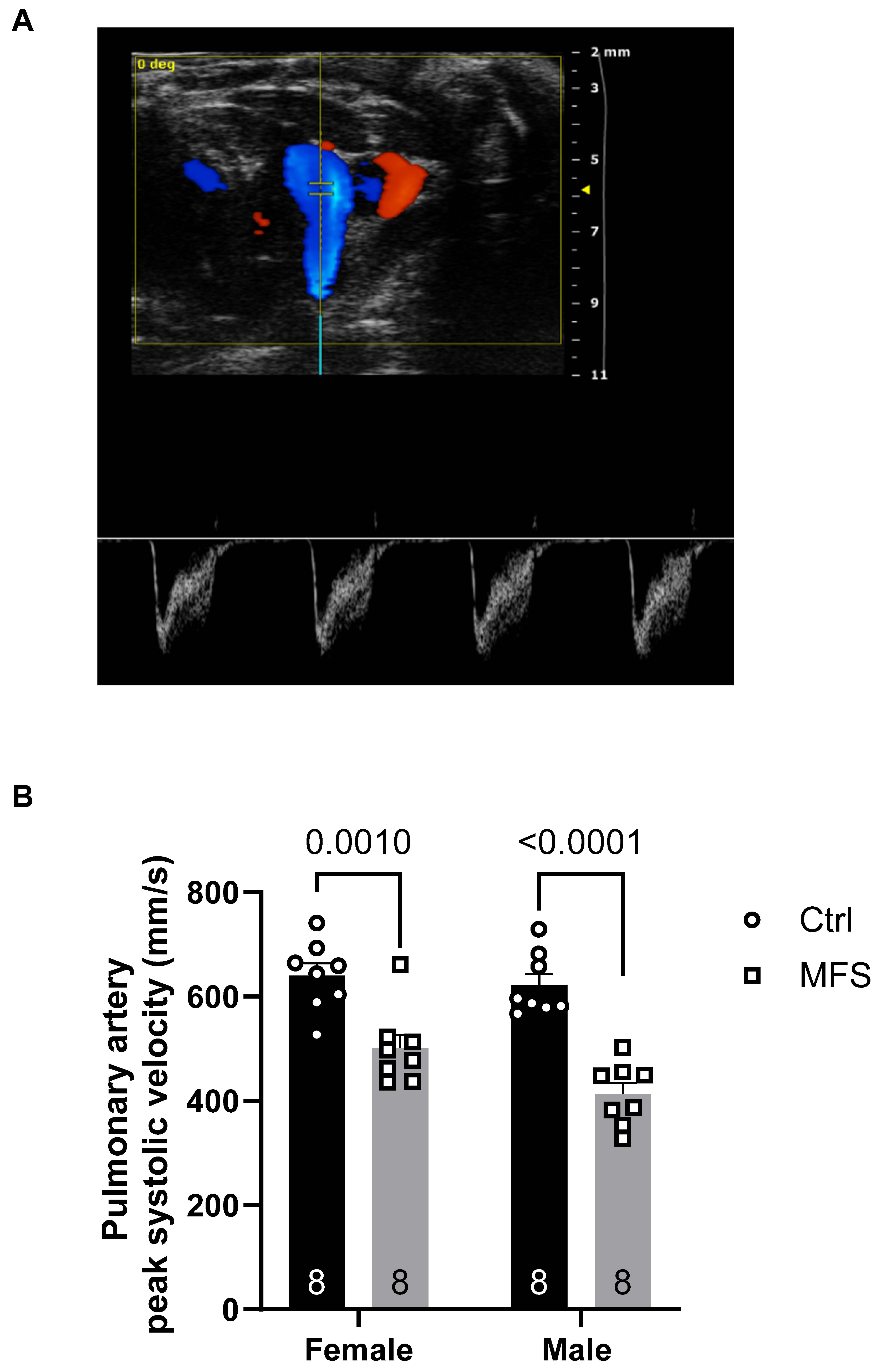
Figure 7.
Ultrasound measurements of the right renal artery peak systolic velocity. A) Ultrasound PW Color Doppler mode view of the right renal artery in a 7-month-old Ctrl mouse. B) There was no significant difference in the peak systolic velocity in the right renal artery between MFS and age- and sex-matched groups. (Mean ± SEM, p < 0.05 is considered as significant).
Figure 7.
Ultrasound measurements of the right renal artery peak systolic velocity. A) Ultrasound PW Color Doppler mode view of the right renal artery in a 7-month-old Ctrl mouse. B) There was no significant difference in the peak systolic velocity in the right renal artery between MFS and age- and sex-matched groups. (Mean ± SEM, p < 0.05 is considered as significant).
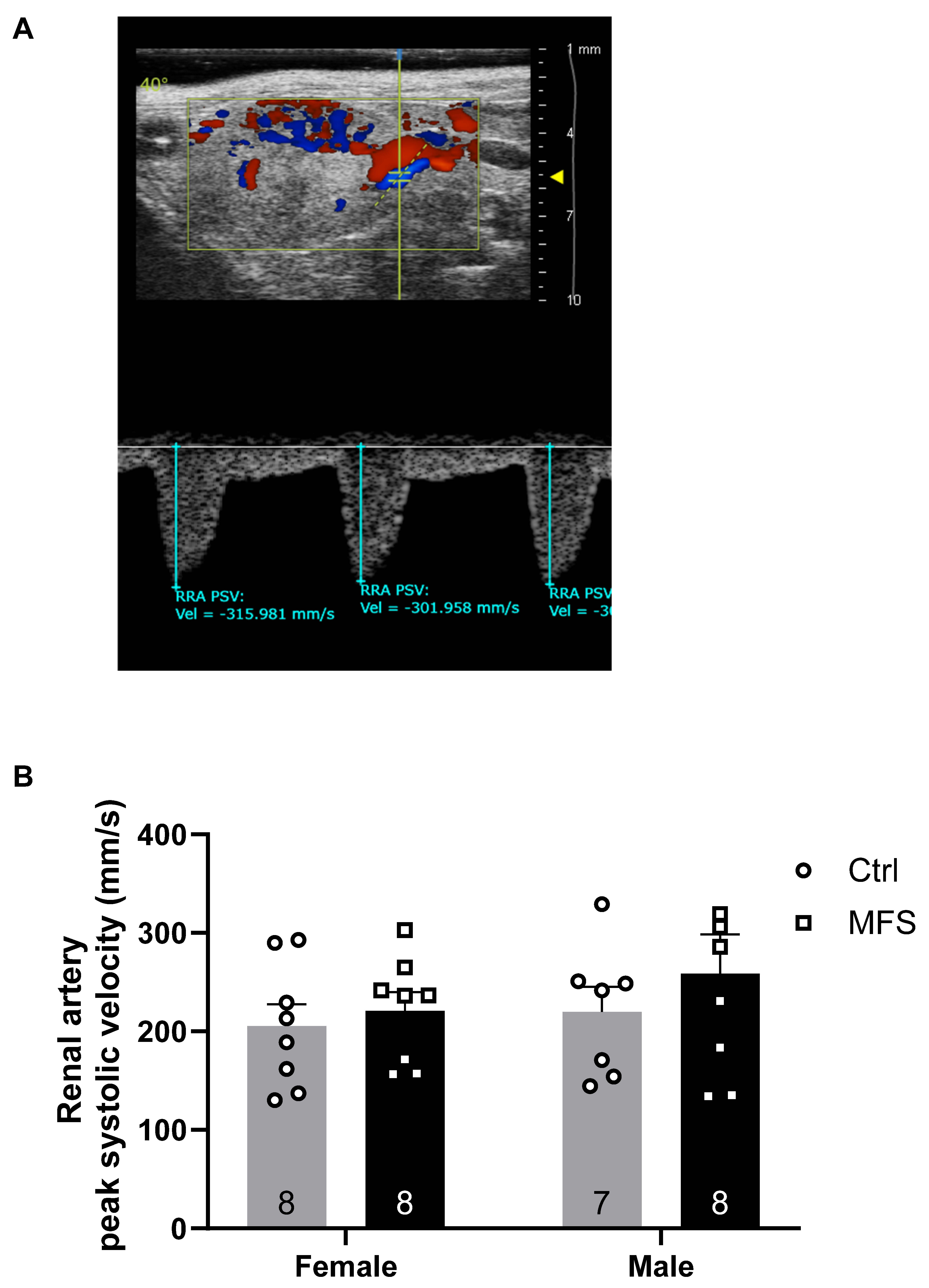
Figure 8.
Measurements of the posterior cerebral artery peak systolic velocity. A) Ultrasound, B-Mode color Doppler view of the left and right posterior cerebral arteries in a 7-month-old Ctrl mouse. B) Peak systolic velocity of the posterior cerebral artery was significantly decreased in male and female MFS mice compared to age- and sex-matched Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
Figure 8.
Measurements of the posterior cerebral artery peak systolic velocity. A) Ultrasound, B-Mode color Doppler view of the left and right posterior cerebral arteries in a 7-month-old Ctrl mouse. B) Peak systolic velocity of the posterior cerebral artery was significantly decreased in male and female MFS mice compared to age- and sex-matched Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
Figure 9.
Effects of mild aerobic exercise on the aortic diameter. A) Exercise training significantly improved the aortic diameter in female MFS mice, bringing the values back to the levels observed in age- and sex-matched healthy Ctrl mice. B) Exercise training partially but significantly improved the aortic diameter in mice MFS mice. C) There were no significant differences between male and female sexes. (Mean ± SEM, p < 0.05 is considered as significant).
Figure 9.
Effects of mild aerobic exercise on the aortic diameter. A) Exercise training significantly improved the aortic diameter in female MFS mice, bringing the values back to the levels observed in age- and sex-matched healthy Ctrl mice. B) Exercise training partially but significantly improved the aortic diameter in mice MFS mice. C) There were no significant differences between male and female sexes. (Mean ± SEM, p < 0.05 is considered as significant).
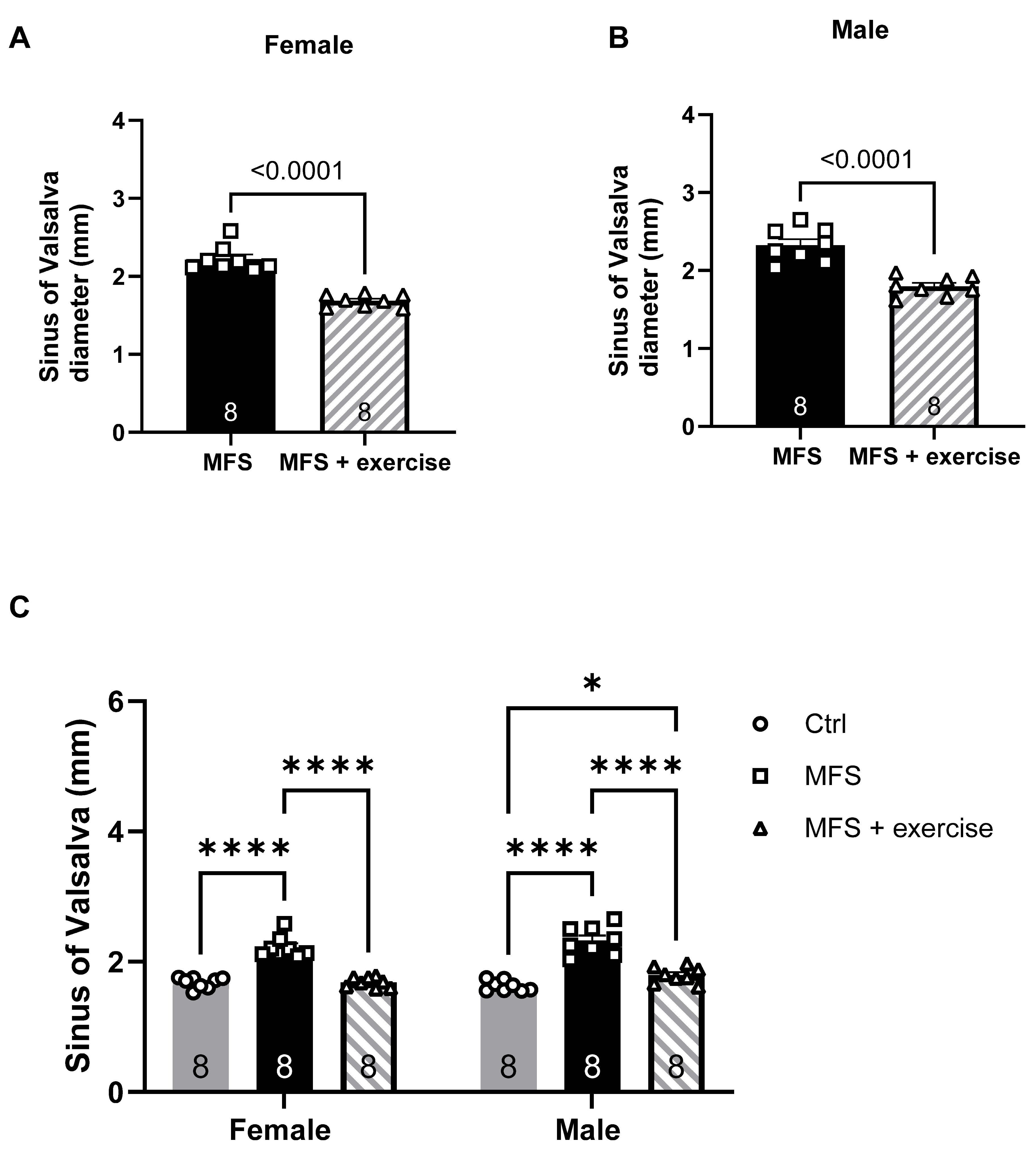
Figure 10.
Effects of mild aerobic exercise on the aortic PWV. A) Exercise training significantly improved aortic wall elasticity in female MFS mice. B) Exercise training significantly improved aortic wall elasticity in male MFS mice. C) In both male and female MFS groups, exercise training did not fully reverse aortic wall functional integrity to levels observed in age- and sex-matched healthy Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
Figure 10.
Effects of mild aerobic exercise on the aortic PWV. A) Exercise training significantly improved aortic wall elasticity in female MFS mice. B) Exercise training significantly improved aortic wall elasticity in male MFS mice. C) In both male and female MFS groups, exercise training did not fully reverse aortic wall functional integrity to levels observed in age- and sex-matched healthy Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
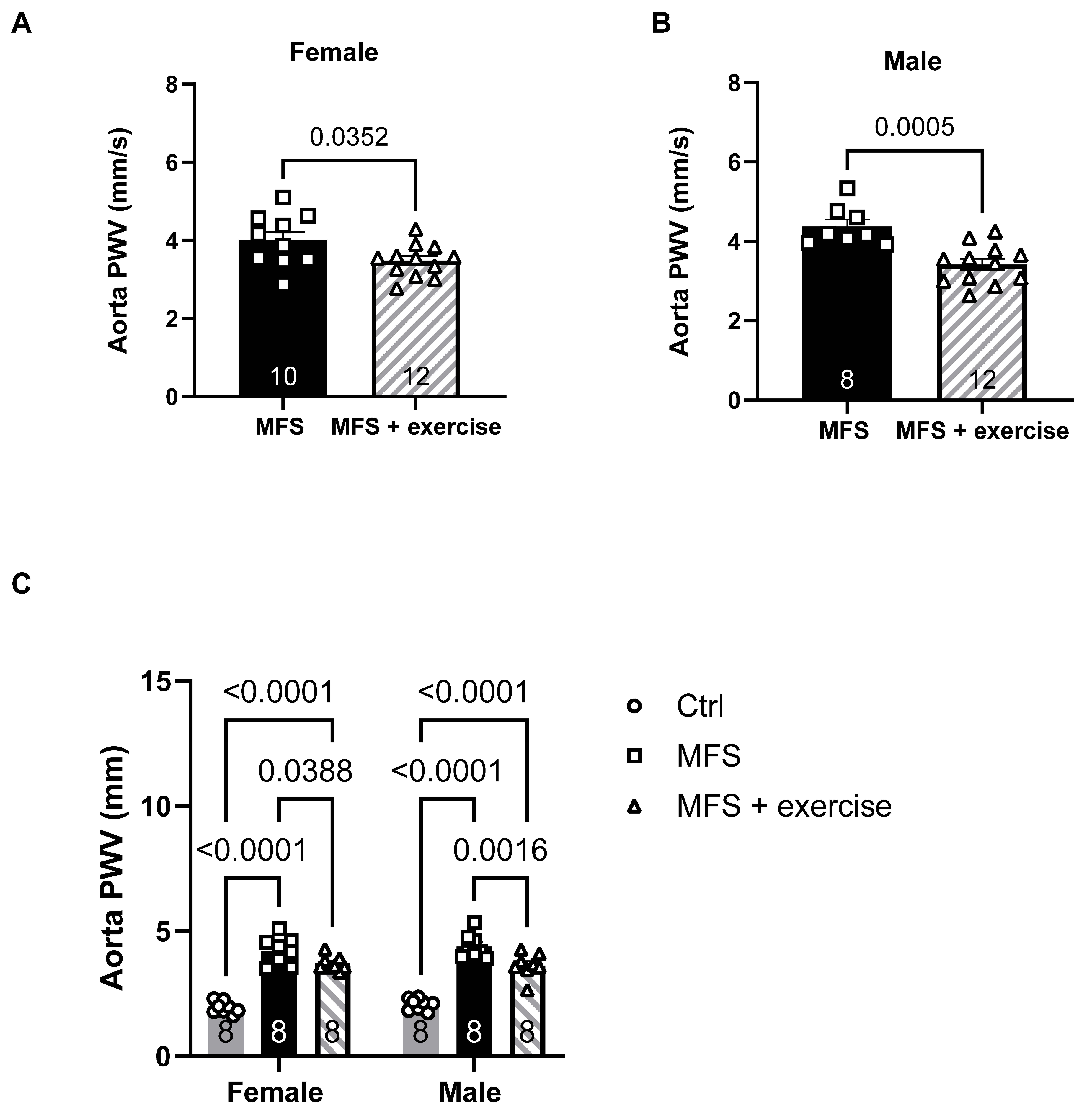
Figure 11.
Effects of mild aerobic exercise on the PCA peak blood flow velocity. In vivo ultrasound measurements showed that exercise improved the peak blood flow velocity in the PCA of both female (A) and male (B) MFS mice. C) In both female and male MFS groups, exercise training effectively restored peak systolic velocity to levels observed in age- and sex-matched healthy Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
Figure 11.
Effects of mild aerobic exercise on the PCA peak blood flow velocity. In vivo ultrasound measurements showed that exercise improved the peak blood flow velocity in the PCA of both female (A) and male (B) MFS mice. C) In both female and male MFS groups, exercise training effectively restored peak systolic velocity to levels observed in age- and sex-matched healthy Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
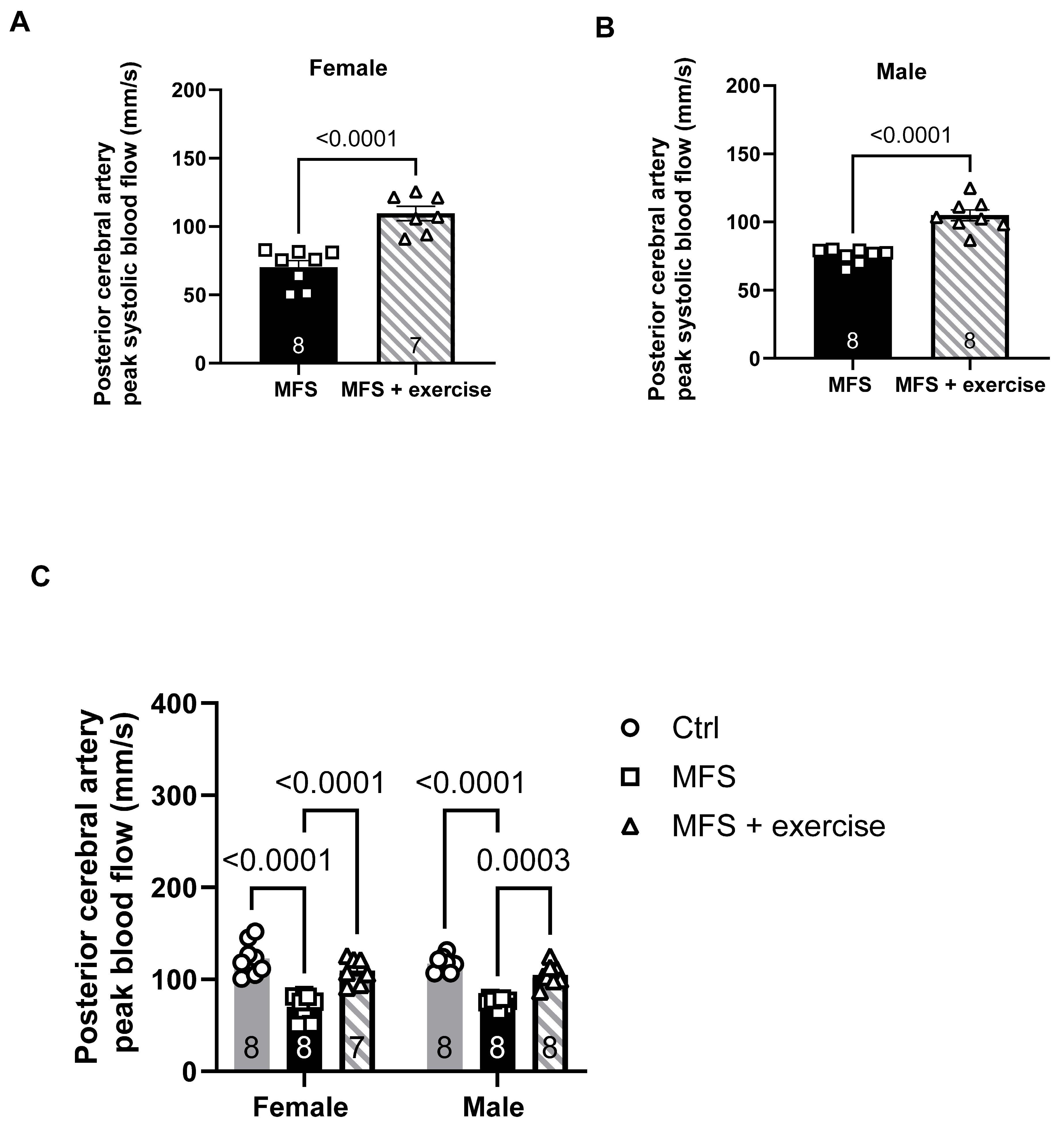
Figure 12.
Effect of mild aerobic exercise on the carotid artery PWV. Exercise significantly improved the carotid artery PWV in female (A) and male (B) MFS mice. C) Mild exercise fully restored carotid artery wall elasticity in both male and female MFS groups to the levels observed in healthy Ctrl groups. In addition, when compared to age-matched male groups, female MFS mice showed a higher carotid artery stiffness, indicating a sex-dependent effect. (Mean ± SEM, p < 0.05 is considered as significant).
Figure 12.
Effect of mild aerobic exercise on the carotid artery PWV. Exercise significantly improved the carotid artery PWV in female (A) and male (B) MFS mice. C) Mild exercise fully restored carotid artery wall elasticity in both male and female MFS groups to the levels observed in healthy Ctrl groups. In addition, when compared to age-matched male groups, female MFS mice showed a higher carotid artery stiffness, indicating a sex-dependent effect. (Mean ± SEM, p < 0.05 is considered as significant).
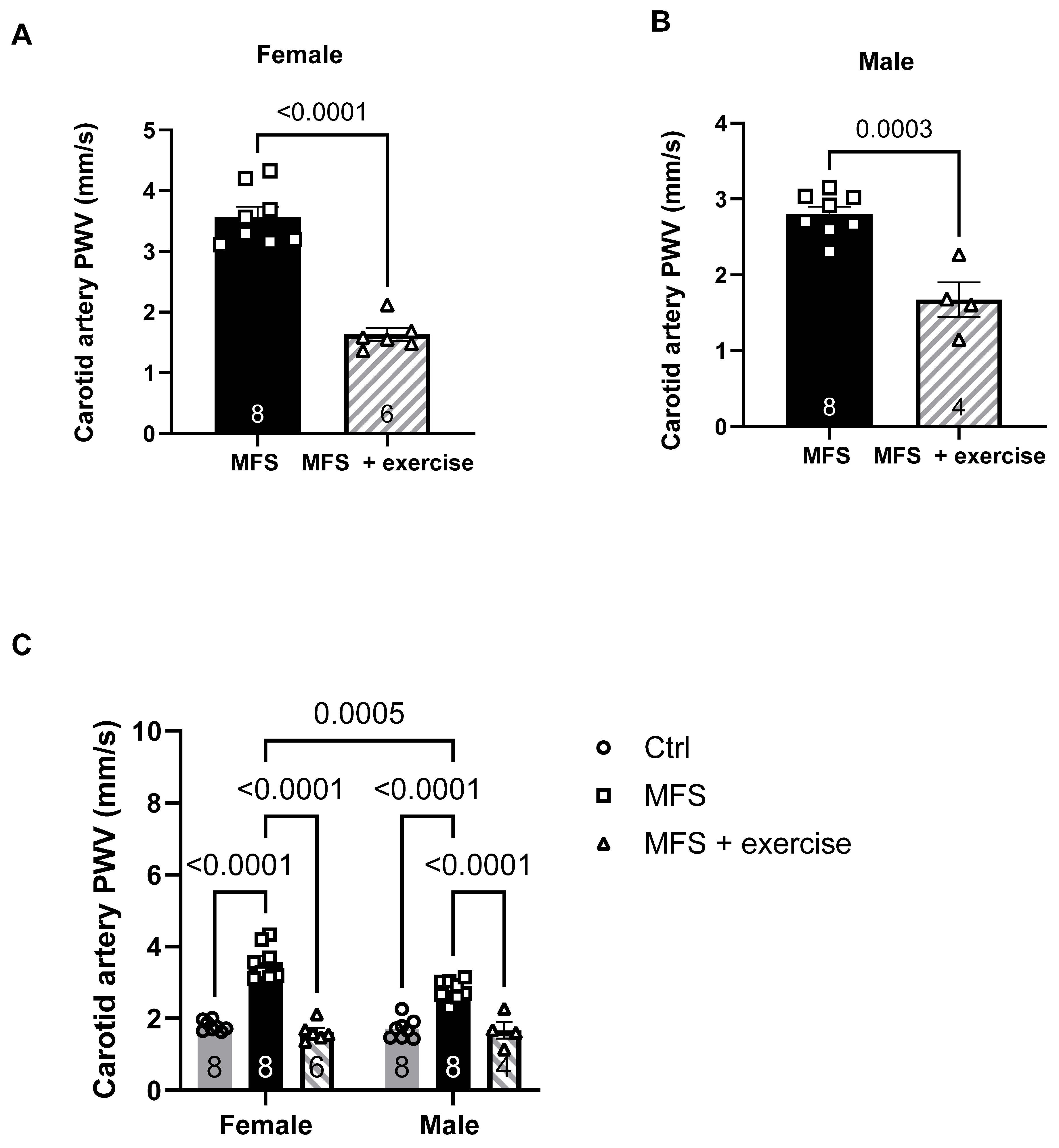
Figure 13.
Effect of mild aerobic exercise on the carotid artery wall thickness. Exercise significantly improved the carotid artery wall thickness in female (A) and male (B) MFS mice. C) Mild exercise fully normalized carotid artery wall thickness in both male and female MFS groups to the levels observed in healthy Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
Figure 13.
Effect of mild aerobic exercise on the carotid artery wall thickness. Exercise significantly improved the carotid artery wall thickness in female (A) and male (B) MFS mice. C) Mild exercise fully normalized carotid artery wall thickness in both male and female MFS groups to the levels observed in healthy Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
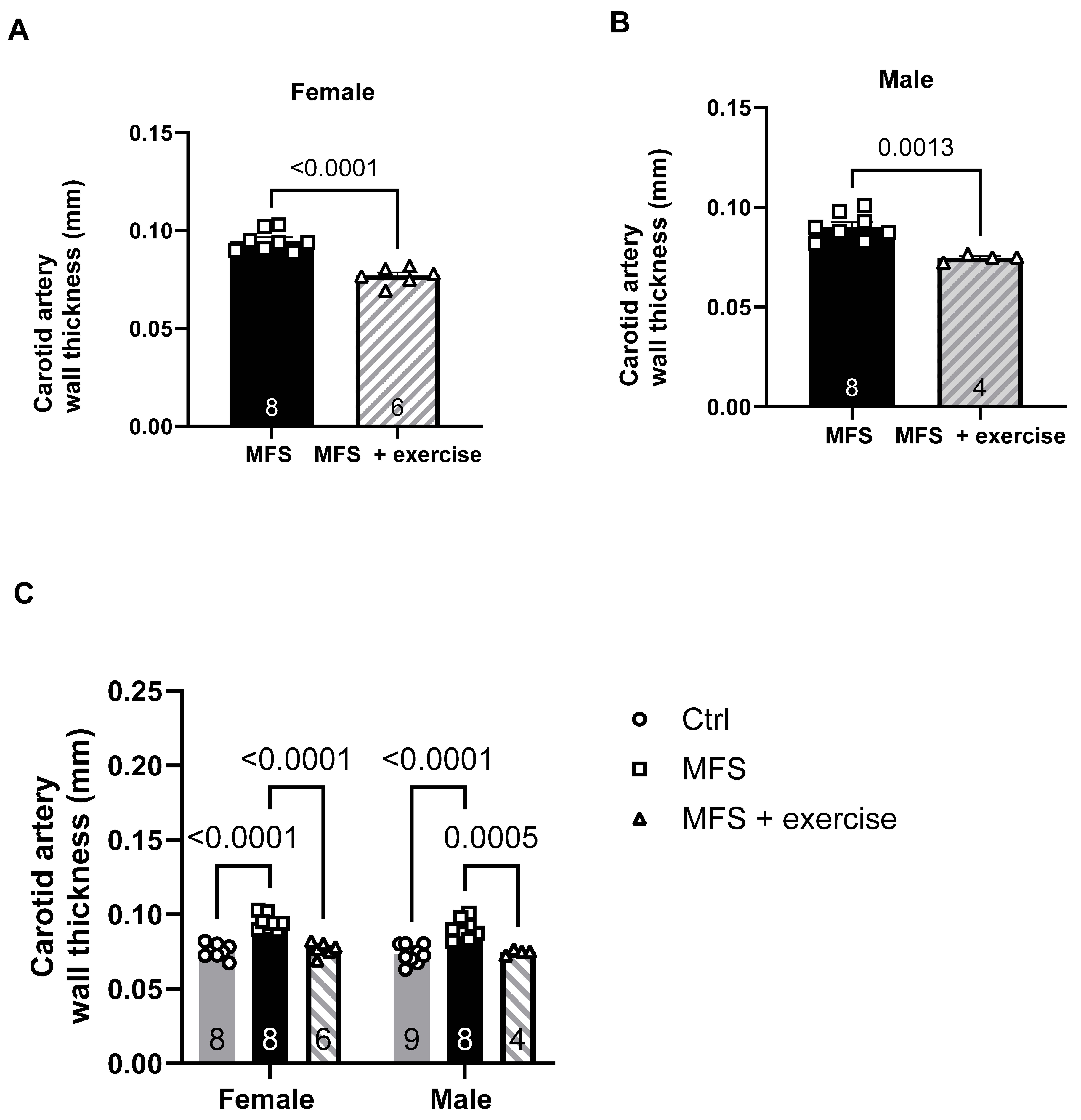
Figure 14.
Effect of mild aerobic exercise on the carotid artery wall distensibility. Exercise significantly improved the carotid artery wall distensibility in female (A) and male (B) MFS mice. C) Mild exercise fully normalized carotid artery wall distensibility in both male and female MFS groups to the levels observed in age- and sex-matched healthy Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
Figure 14.
Effect of mild aerobic exercise on the carotid artery wall distensibility. Exercise significantly improved the carotid artery wall distensibility in female (A) and male (B) MFS mice. C) Mild exercise fully normalized carotid artery wall distensibility in both male and female MFS groups to the levels observed in age- and sex-matched healthy Ctrl groups. (Mean ± SEM, p < 0.05 is considered as significant).
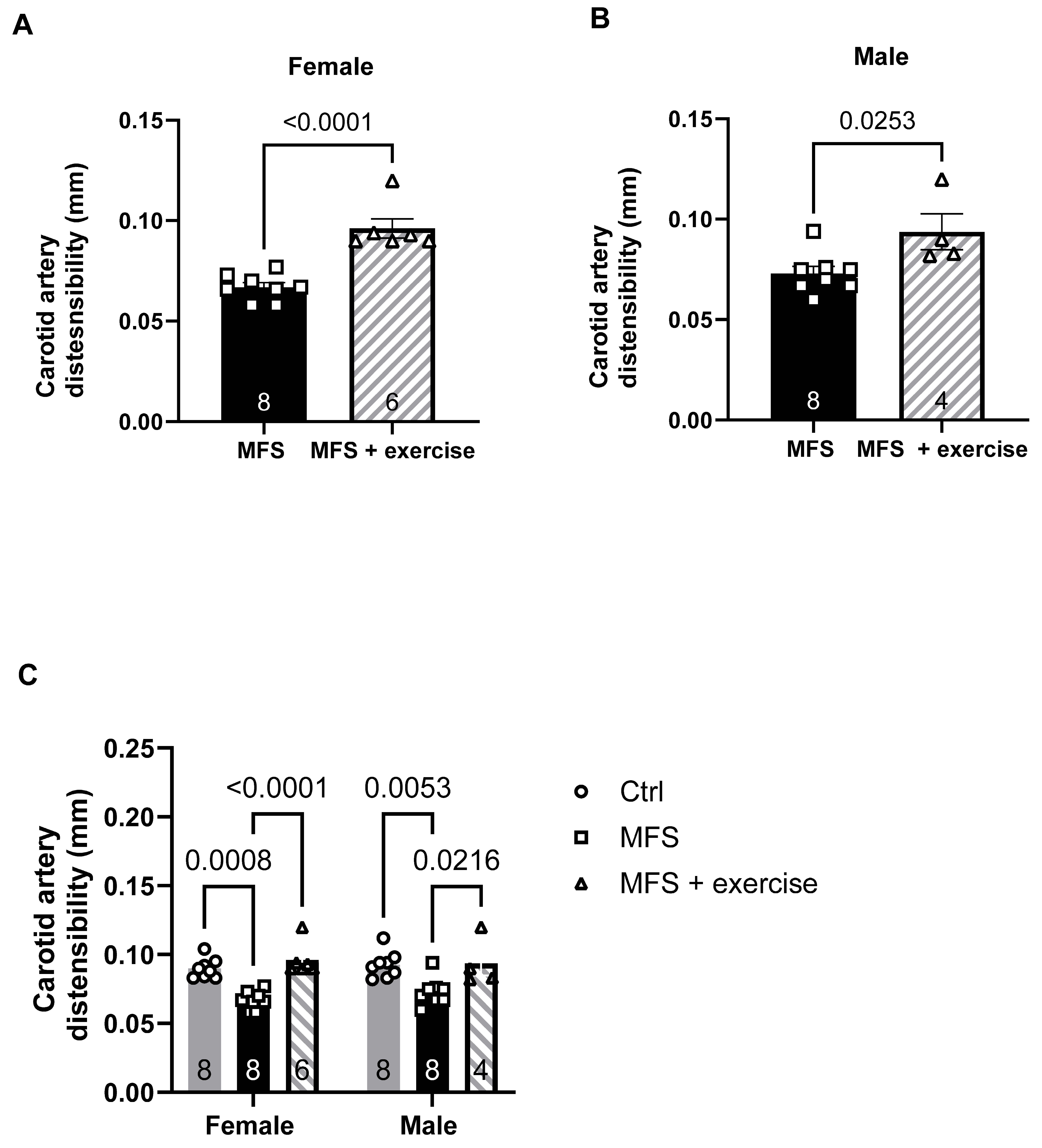
Figure 15.
Inverse correlation between PCA peak blood flow and aortic root dilation and wall stiffness in male and female mice. A) Linear regression analysis showed a moderate inverse relationship between the aortic PWV and the PCA peak blood flow in the female group (R2 = 0.5054). B) There is a stronger inverse relationship between the PCA peak blood flow and aortic PWV in the male group (R2 = 0.7422). C) Linear regression analysis showed a moderate inverse relationship between the PCA peak blood flow and aortic root diameters in the female group (R2 = 0.5054). D) There is a stronger inverse relationship between the PCA peak blood flow and aortic root diameters in the male group (R2 = 0.7422).
Figure 15.
Inverse correlation between PCA peak blood flow and aortic root dilation and wall stiffness in male and female mice. A) Linear regression analysis showed a moderate inverse relationship between the aortic PWV and the PCA peak blood flow in the female group (R2 = 0.5054). B) There is a stronger inverse relationship between the PCA peak blood flow and aortic PWV in the male group (R2 = 0.7422). C) Linear regression analysis showed a moderate inverse relationship between the PCA peak blood flow and aortic root diameters in the female group (R2 = 0.5054). D) There is a stronger inverse relationship between the PCA peak blood flow and aortic root diameters in the male group (R2 = 0.7422).
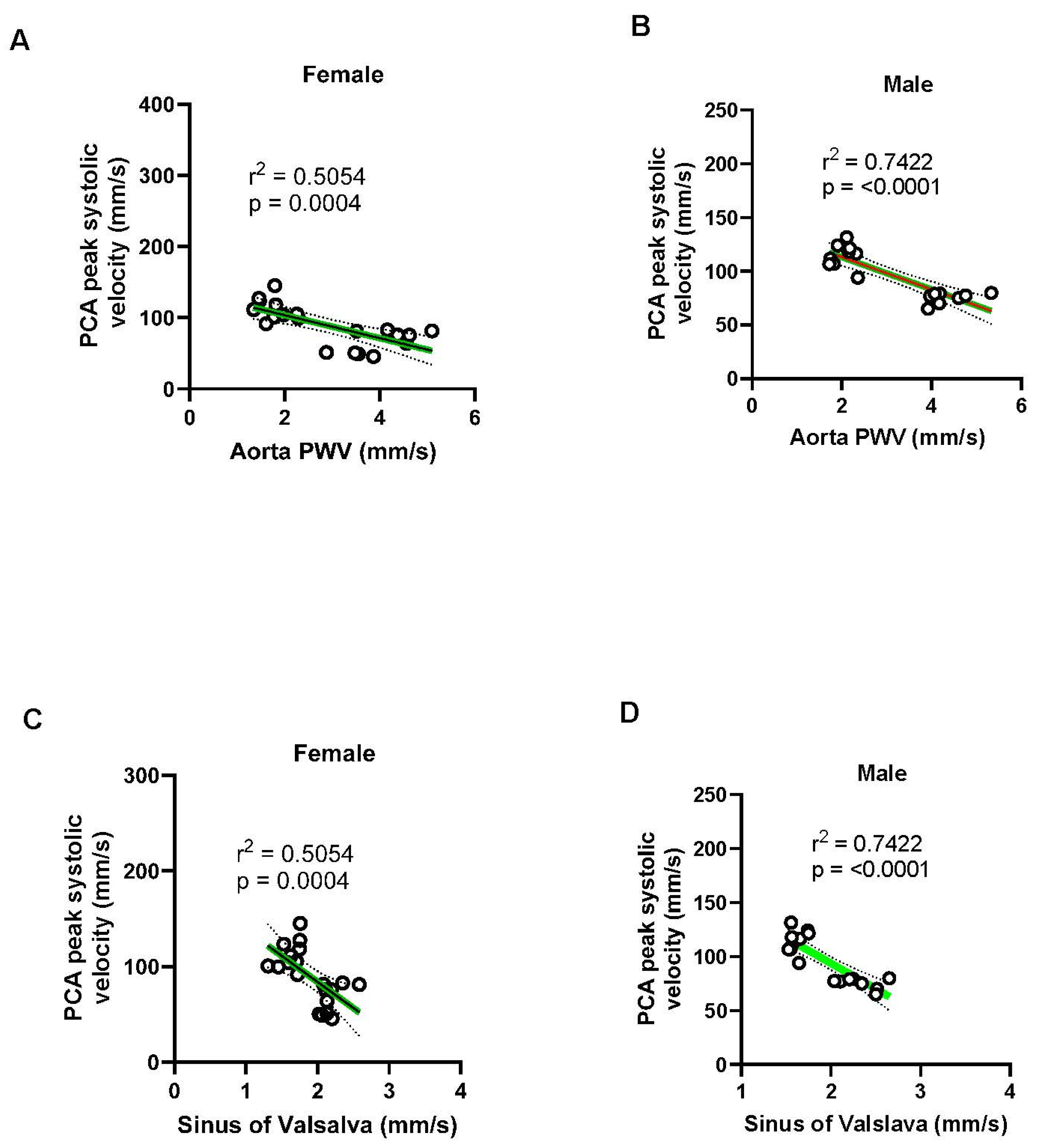
Figure 16.
Linear regression plots between the aortic PWV carotid artery parameters. (A-B) Carotid PWV showed a strong positive correlation with aortic PWV in both sexes (females: R² = 0.7427; males: R² = 0.7324), indicating that systemic arterial stiffening is associated with local carotid stiffening. (C-D) Carotid wall thickness also positively correlated with aortic PWV (females: R² = 0.7427; males: R² = 0.5816), with stronger associations observed in females. (E-F) Carotid artery distensibility index negatively correlated with aortic PWV in both sexes, suggesting reduced carotid compliance with increasing aortic stiffness (females: R² = 0.3943; males: R² = 0.5972, with the relationship being more pronounced in males. These data highlight sex-dependent differences in the impact of aortic stiffness on carotid artery function and structure.
Figure 16.
Linear regression plots between the aortic PWV carotid artery parameters. (A-B) Carotid PWV showed a strong positive correlation with aortic PWV in both sexes (females: R² = 0.7427; males: R² = 0.7324), indicating that systemic arterial stiffening is associated with local carotid stiffening. (C-D) Carotid wall thickness also positively correlated with aortic PWV (females: R² = 0.7427; males: R² = 0.5816), with stronger associations observed in females. (E-F) Carotid artery distensibility index negatively correlated with aortic PWV in both sexes, suggesting reduced carotid compliance with increasing aortic stiffness (females: R² = 0.3943; males: R² = 0.5972, with the relationship being more pronounced in males. These data highlight sex-dependent differences in the impact of aortic stiffness on carotid artery function and structure.
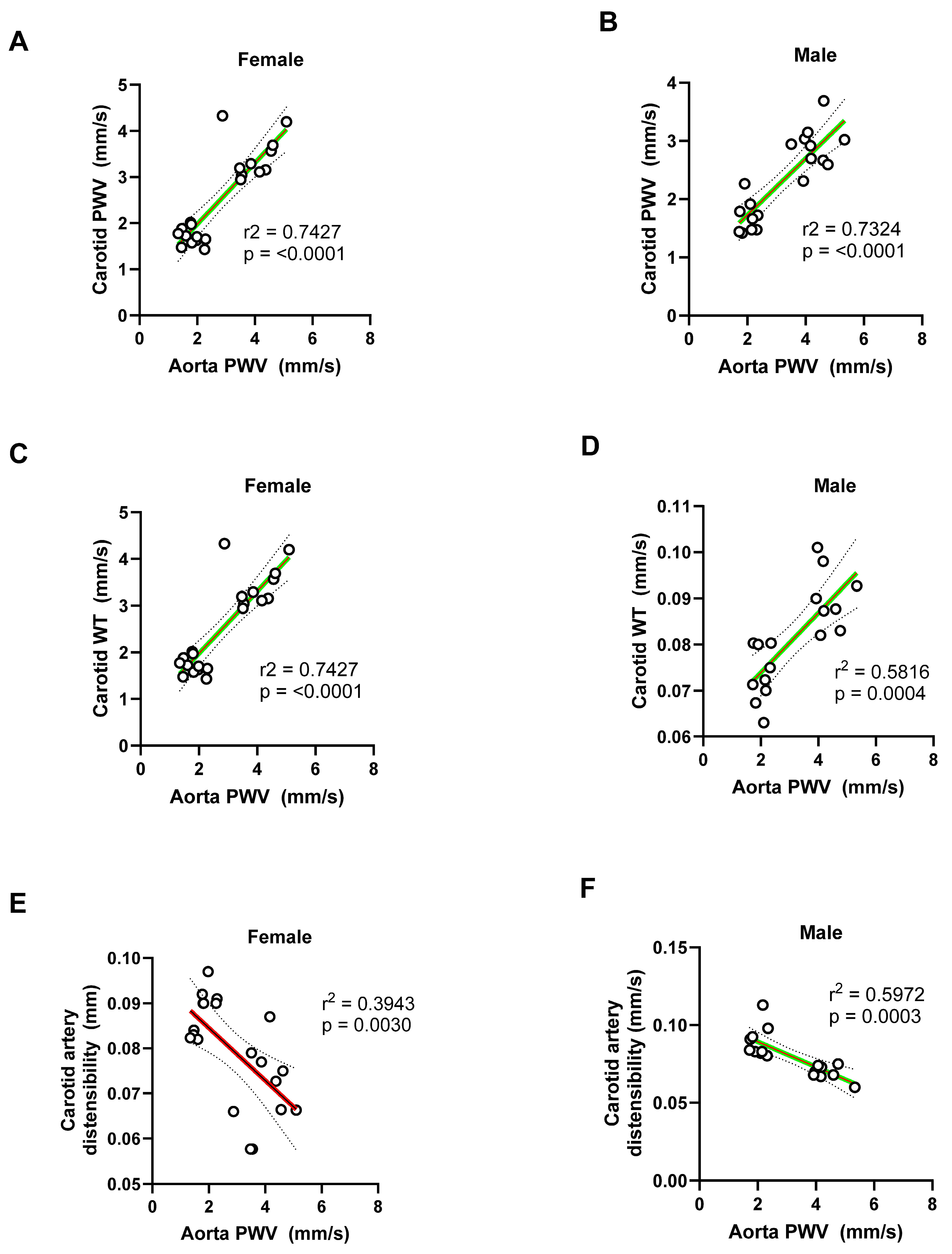
Figure 17.
Linear regression plots between the aortic root diameter and the carotid artery parameters in male and female mice. (A-B) Carotid city (PWV) was positively correlated with aortic root diameter in both females (R² = 0.5546) and males (R² = 0.7234), suggesting that dilation of the aortic root is associated with increased carotid stiffness. (C-D) Carotid artery wall thickness also positively correlated with aortic root diameter in both sexes, with stronger correlation in males (females: Rr² = 0.7335; males: R² = 0.8678, p < 0.0001), indicating vascular remodeling linked to aortic root enlargement. (E-F) Carotid artery distensibility index showed an inverse correlation with aortic diameter, indicating reduced carotid compliance with larger sinus diameters. This relationship was modest in females (R² = 0.3296) and more pronounced in males (R² = 0.5834).
Figure 17.
Linear regression plots between the aortic root diameter and the carotid artery parameters in male and female mice. (A-B) Carotid city (PWV) was positively correlated with aortic root diameter in both females (R² = 0.5546) and males (R² = 0.7234), suggesting that dilation of the aortic root is associated with increased carotid stiffness. (C-D) Carotid artery wall thickness also positively correlated with aortic root diameter in both sexes, with stronger correlation in males (females: Rr² = 0.7335; males: R² = 0.8678, p < 0.0001), indicating vascular remodeling linked to aortic root enlargement. (E-F) Carotid artery distensibility index showed an inverse correlation with aortic diameter, indicating reduced carotid compliance with larger sinus diameters. This relationship was modest in females (R² = 0.3296) and more pronounced in males (R² = 0.5834).
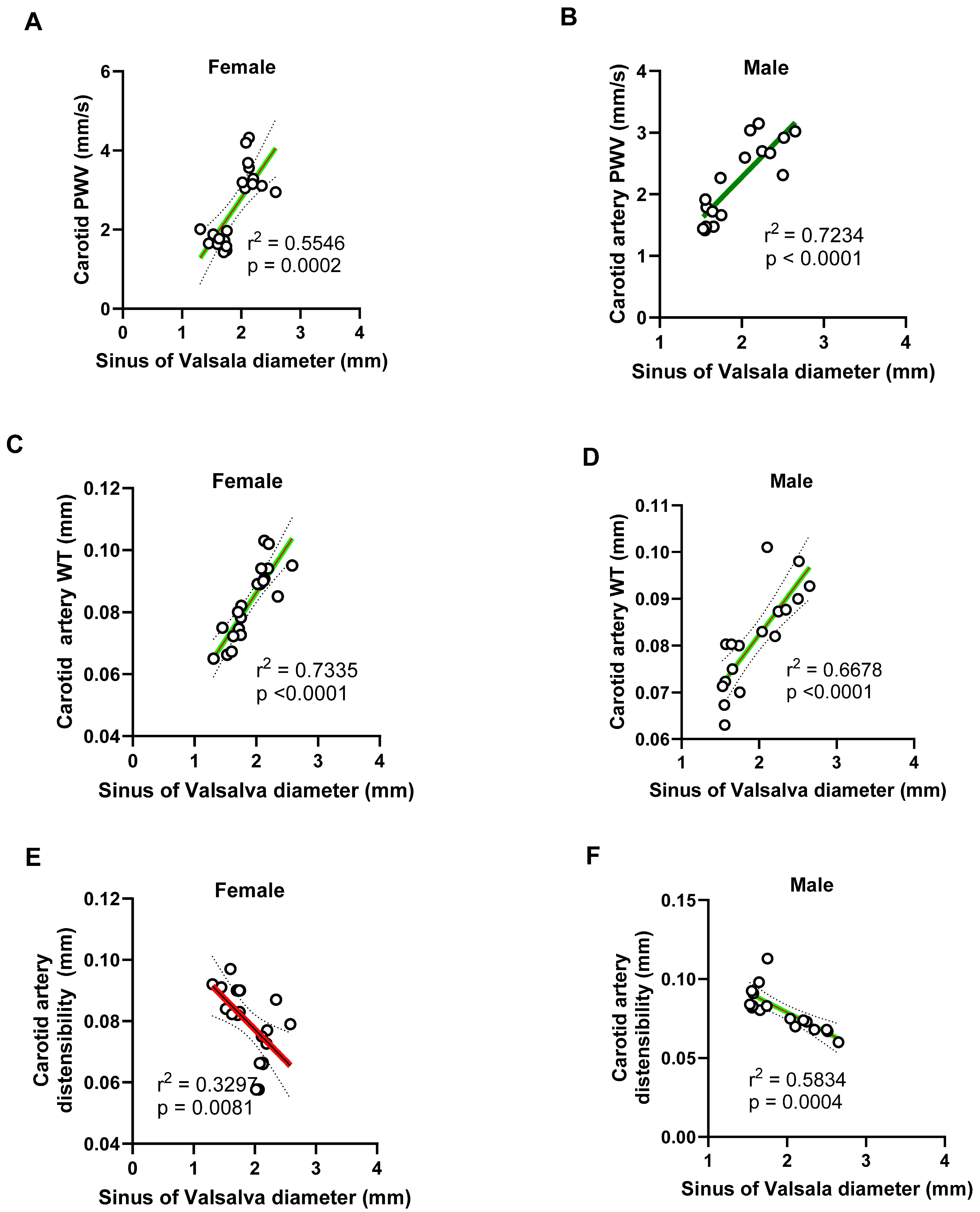
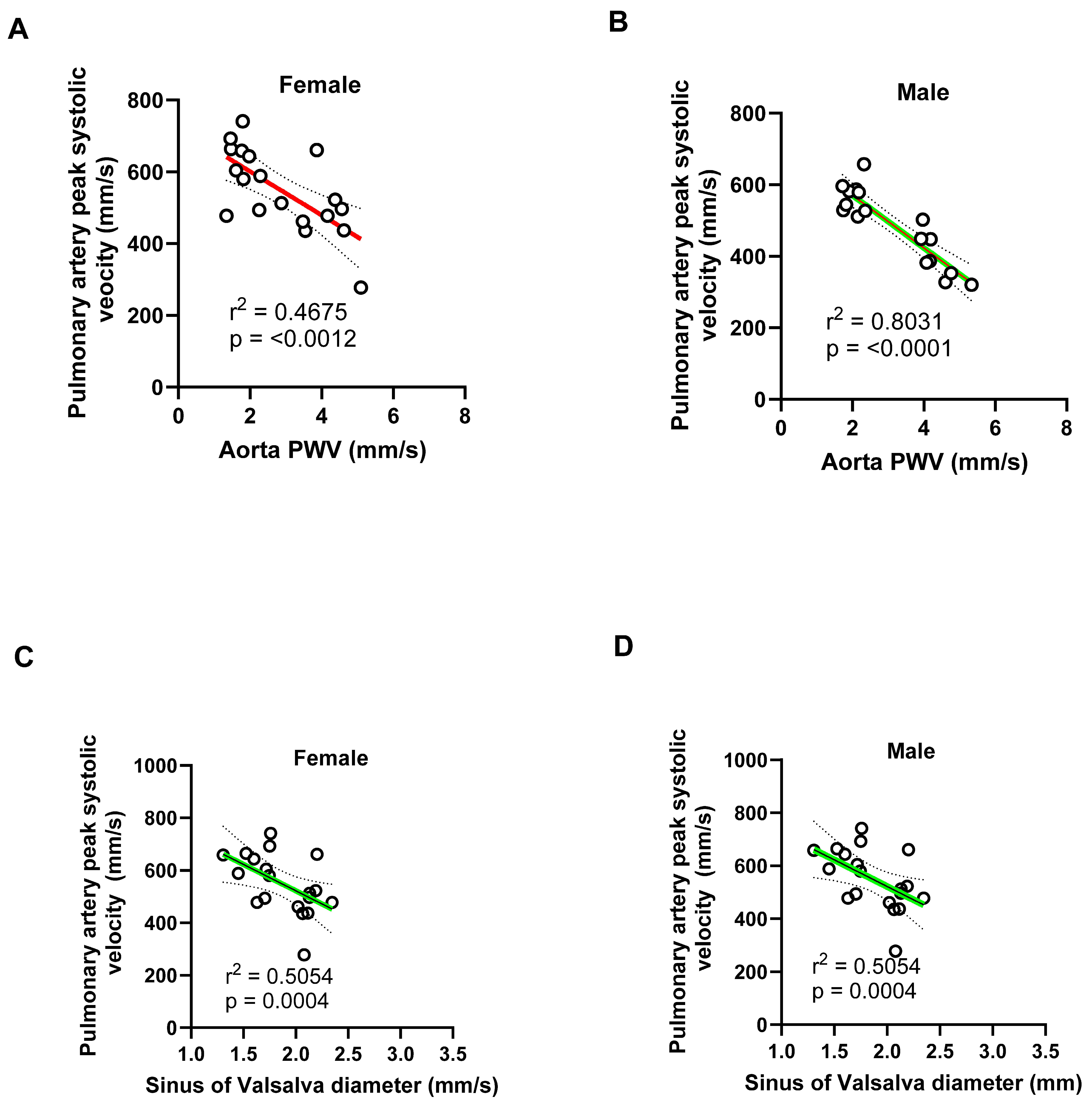
Figure 18.
Negative correlation between pulmonary artery systolic blood flow and aortic stiffness and diameter in male and female mice. (A-B) Pulmonary artery (PA) peak systolic blood velocity negatively correlated with aortic wall stiffness (PWV) in both females (R² = 0.4875) and males (R² = 0.8031), indicating that increased aortic wall stiffness is associated with reduced pulmonary flow velocity, with a stronger relationship observed in males. (C-D) Similarly, PA peak systolic blood velocity negatively correlated with aortic root diameters in both sexes (females: R² = 0.5054; males: R² = 0.5854), suggesting that progressive dilation of the aortic root impairs pulmonary hemodynamics.
Figure 18.
Negative correlation between pulmonary artery systolic blood flow and aortic stiffness and diameter in male and female mice. (A-B) Pulmonary artery (PA) peak systolic blood velocity negatively correlated with aortic wall stiffness (PWV) in both females (R² = 0.4875) and males (R² = 0.8031), indicating that increased aortic wall stiffness is associated with reduced pulmonary flow velocity, with a stronger relationship observed in males. (C-D) Similarly, PA peak systolic blood velocity negatively correlated with aortic root diameters in both sexes (females: R² = 0.5054; males: R² = 0.5854), suggesting that progressive dilation of the aortic root impairs pulmonary hemodynamics.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



