
Submitted:
15 May 2025
Posted:
16 May 2025
You are already at the latest version
Abstract
The linear polymer polyphosphate (polyP) is found across all three domains of life and fulfills diverse physiological functions, including phosphorus storage, chaperone activity and stress tolerance. In bacteria, polyP synthesis is catalyzed by polyphosphate kinase (Ppk), while its degradation is carried out by exopolyphosphatases (Ppx). The intracellular polyP level is determined by the balance between these two enzymatic activities; however, the regulatory mechanisms governing this balance remain incompletely understood. In higher eukaryotes, polyP participates in a wide variety of physiological processes. In relation to this, accumulating evidence suggests that polyP may also confer beneficial effects on human intestinal health, triggering increased interest in the production of polyP by probiotic lactic acid bacteria. This review summarizes the current knowledge on polyP metabolism in these bacteria and explores its functional properties and potential applications.
Keywords:
lactic acid bacteria
; Lactobacillaceae
; polyphosphate
; polyphosphatase
; probiotic
1. Introduction
Polyphosphate (polyP) is a linear polymer composed of phosphoryl monomers linked by high-energy phosphoanhydride bonds (Figure 1). PolyP chains range in length from three to several hundred residues. Although considered to exist almost exclusively in a linear polymer within organisms, recent evidence suggests that alternative configurations, such as branched polyP, may be more stable in aqueous media than previously thought [1,2]. PolyP can also be produced abiotically in environments such as volcanoes [3], and it is thought to have been present on prebiotic Earth, contributing to conditions for the emergence of life. It may have been incorporated into prebiotic vesicles, serving as a source of inorganic phosphate (Pi) and high-energy bonds, potentially acting as a precursor to ATP [4]. This molecule can be found in every type of cell, including bacteria, archaea, fungi, protozoa, plants, and animals [4,5,6].
In bacteria, polyP plays multiple roles, including phosphate and energy storage, survival under starvation, stress response, regulation of cell motility, biofilm formation, and virulence [4,5]. Additionally, polyP mitigates stress-induced DNA damage [7] and influences enzyme activity [2,5,8,9]. It can also form a wide variety of complexes with inorganic ions, proteins and nucleic acids [2]. PolyP was first noticed through the study of characteristic metachromatic inclusions, originally described over a century ago in Spirillum volutans and classically named as volutin granules [10]. Electron diffraction studies have shown that these structures are amorphous and elemental analysis with energy dispersive X-ray spectroscopy demonstrated that are enriched in the divalent cations Mg2+ and Ca2+ [2].
PolyP has attracted significant interest for biotechnological applications. The removal of phosphorus from wastewater by polyP-synthetizing bacteria, although still minor in wastewater treatment plants, has shown promising results and offers several advantages over chemical methods, such as eliminating the need for chemical precipitants and reducing sludge volumes [11]. Due to its highly variable properties, depending on chain length, cation composition, solvent, etc. polyP has been widely used in different industrial applications, such as fertiliser, fire retardant or additive for different materials [12]. PolyP is also attracting interest for the development of biomedical materials in regenerative medicine [13]. In the food industry, polyP is extensively used to enhance food quality, preservation, and nutritional value. It improves water-holding capacity, stabilizes emulsions, and fortifies foods with essentials minerals [14,15]. Furthermore, in recent years, a number of studies have shown that polyP produced by some bacteria can contribute to the maintenance of intestinal homeostasis. PolyP was initially proposed as the agent responsible for the probiotic activity exhibited by Levilactobacillus brevis SBC8803 [16]. This finding led to a renewed interest in polyP synthesis by lactic acid bacteria (LAB).
LAB were first defined by Orla-Jensen as a group of Gram-positive, non-sporulating, non-motile bacteria, with coccoid or rod forms and the ability to catabolize sugars mainly to lactic acid [17]. This classification encompassed a heterogeneous group, including bifidobacteria and lactobacilli, among others. Advances in molecular phylogenetic analyses have since clarified the taxonomic status of the bacterial groups included in LAB. Today, LAB refers specifically to organisms belonging to the phylum Bacillota (formerly Firmicutes), class Bacilli, order Lactobacillales. Phylogenetic studies indicate that Lactobacillales constitutes a monophyletic group [18,19], comprising the families Aerocococcaceae, Carnobacteriaceae, Enterocococcaceae, Lactobacillaceae and Streptococcaceae. Many LAB species are considered Generally Recognized as Safe (GRAS) by US FDA or QPS (Qualified Presumption of Safety), according to European legislation [20], due to their long-standing use in the production of fermented foods and their natural presence as commensals in the human gastrointestinal tract. Furthermore, some of them are acknowledged as health-promoting bacteria.
Given the functional properties of polyP, and the widespread use of LAB in the food industry and probiotic formulations, understanding how LAB regulate polyP synthesis and degradation could help to enhance their functional potential and optimize their application in fermented products and nutritional supplements. Investigating the metabolic pathways involved in polyP metabolism in LAB not only deepens our fundamental understanding of LAB physiology, but also opens new opportunities for biotechnological and health-related applications. This review aims to provide a comprehensive overview of polyP synthesis in food-associated LAB, with a focus on the key enzymes involved, physiological roles and its potential applications.
2. Synthesis of Polyphosphate in LAB
The main enzymes in bacteria involved in polyP synthesis are polyP kinases (Ppk), which, based on the similarity of their sequences and kinetic properties, are classified into two families: Ppk1 (EC 2.7.4.1) and Ppk2 (EC 2.7.4.34). The two enzymes share no significant sequence similarity and possess different structural folds [21]. Conversely, exopolyphosphatases (Ppx) hydrolyze polyP, releasing the terminal Pi. On the basis of their primary structures, they are divided into two types, Ppx1 and Ppx2, respectively. Ppx1 consists of a N-terminal domain belonging to the acetate and sugar kinase/ hsp70/actin superfamily (ASKHA) and a C-terminal domain which is responsible for the processivity of the enzyme [22]. Ppx2 enzymes share the N-terminal domain but lack the C-terminal domain of Ppx1.
The presence of polyP in LAB was first reported in the early 1960s in Lactobacillus casei (currently Lacticaseibacillus casei) [23]. Subsequently, Kakefuda et al. [24] identified polyP granules in the cytoplasm of Lactobacillus plantarum (currently Lactiplantibacillus plantarum) while studying its membrane ultrastructure. Further investigations described polyP in Lp. plantarum as a molecule complexing Mn2+ as a counterion [25]. The detection of polyP granules in 60% of Lactobacillus strains isolated from mozzarella cheese whey suggested that the ability to synthesize polyP is widespread in lactobacilli [26]. This observation has been supported by additional studies reporting polyP in various other Lactobacillaceae species [16,27,28,29]. A survey of available LAB genome sequences reveals that genes involved in polyP synthesis and degradation, namely, ppk1, ppx1 and ppx2, are uncommon in Aerococcaceae, Carnobacteriaceae, Enterococcaceae and Streptococcaceae whereas they are highly prevalent in Lactobacillaceae [29,30]. In contrast, ppk2 genes are present in far fewer Lactobacillaceae species (Table 1) and never found in the absence of ppk1. However, the capacity to synthesize polyP is not universal within this group, as several species of lactobacilli lack the enzymes required for polyP biosynthesis (Table 1).
Analysis of the genetic organization of polyP genes in Lactobacillaceae reveals that most members harbor a gene cluster with the gene order ppx1-ppk1-ppx2 [29] (Figure 2). Additional genes may be present in some genera alongside this core cluster, although there is no evidence of their involvement on polyP metabolism. Notably, the ppx1 gene is absent in some genera such as Acetilactobacillus, Companilactobacillus, Oenococcus and Xylocopilactobacillus, whereas Secundilactobacillus encode two copies of ppk1 and ppx2 (Figure 2). On the other hand, ppk2 genes are usually monocistronic and located apart from the other polyP metabolic genes, with the exception of Furfurilactobacillus, where all genes are located within the same cluster (Figure 2).
3. Polyphosphate Kinases
The Ppk1 enzyme is one of the most studied enzymes in polyP metabolism. Ppk1 was first characterized in Escherichia coli [31,32,33] and subsequently in other bacteria such as Mycobacterium smegmatis [34], Arthrobacter atrocyaneus [35], Propionibacterium shermanii [36,37] or Neisseria meningitidis [38], among others. These enzymes transfer a terminal phosphate group to the polyP chain using ATP, leading to the progressive synthesis of an elongated polyP chain [4]. The determination of the structure of the E. coli Ppk1 revealed that it forms a dimer where the active site is located in a tunnel that penetrates the center of each monomer with an ATP binding pocket in one side and conserved positively charged residues along the tunnel which possibly interact with the polyP chain during elongation [39].
In LAB, the presence of the ppk1 gene in the genome is associated with the synthesis of polyP. Alcántara et al. [29] reported that 18 out of 34 lactobacillus strains accumulated polyP, and this accumulation correlated with the presence of ppk1 genes in their genomes. A subsequent study that tested polyP accumulation in different strains of Lactobacillus and Enterococcus detected no polyP accumulation in the Enterococcus strains lacking ppk1 homologs [27]. However, in both studies, polyP accumulation widely varied even among different strains of the same species. This variability remains unexplained, as the regulation of polyP synthesis in LAB is poorly understood. In addition to differences in Ppk biosynthetic activities, variations in Pi uptake and polyP degradation may also contribute to this effect [27,29]. A dependence of polyP synthesis on Pi concentration has been observed in several LAB [29,40]. Although no LAB Ppk1 has been structurally or biochemically characterized in detail, a comparison of Lactobacillaceae Ppk1 sequences with that of E. coli reveals that critical residues for Ppk1 catalytic activity are conserved in Lactobacillaceae Ppks, suggesting that their structures possibly resemble that of E. coli Ppk1 (Figure 3). Furthermore, inactivation of ppk1 gene in Lc. paracasei BL23 abolished polyP synthesis in this organism [29] and in vitro synthesis of polyP by Lc. paracasei Ppk1 cloned and expressed in E.coli has been demonstrated [30].
The discovery of Ppk2 originated from the observation that a Pseudomonas aeruginosa mutant defective in Ppk1 still produced up to 20% of wild-type polyP levels [21,41]. This suggested that another enzyme might be responsible for additional polyphosphate kinase activity [41]. Subsequently, Ppk2 was identified as the enzyme responsible for synthesizing guanosine triphosphate (GTP) from guanosine diphosphate (GDP) and polyP in P. aeruginosa [42]. As noted above, Ppk2 proteins share no significant sequence similarity with Ppk1 and their structural analyses evidenced that they adopt different folds [21]. Biochemical analyses showed that P. aeruginosa Ppk2 exhibits a 75-fold preference for polyP degradation over synthesis [21,42]. Consequently, Ppk2 enzymes are more efficient in ATP/GTP synthesis due to their preference for nucleoside phosphorylation in the reversible reaction. Therefore, while Ppk1 acts preferentially in ATP-dependent polyP synthesis, Ppk2 is more efficient in ATP/GTP synthesis from polyP (Figure 4).
Ppk2 is a member of the P-loop-containing kinases, which are characterized by the presence of two conserved sequence motifs (Walker A and Walker B) and a lid module. Ppk2 proteins possess deviant Walker A and B motifs compared to other P-loop kinases [43]. Based on phylogenetic, biochemical and structural data, Ppk2 enzymes can be further classified into three subfamilies according to their nucleoside phosphate substrate preference: Class I phosphorylates nucleoside diphosphates, Class II phosphorylates nucleoside monophosphates, and Class III can phosphorylate both nucleoside mono or diphosphates [44]. Although no Lactobacillaceae Ppk2 protein has been biochemically characterized, phylogenetic analysis places them in Class III [44] and the alignment of Lactobacillaceae Ppk2 sequences also shows conservation of the critical catalytic domains (Figure 5).
4. Exopolyphosphatases
Exopolyphosphatases (Ppx) can progressively hydrolyze and release the terminal Pi from linear polyP chains containing three or more phosphoanhydride bonds. In initial investigations in E. coli, a gene encoding the exopolyphosphatase enzyme (ppx) was identified adjacent to the gene encoding Ppk1 (ppk). These two genes constitute a polycistronic operon, in which transcription of the ppx gene is regulated by the ppk promoters [45]. Elucidation of the structure of E. coli Ppx revealed that the protein is composed of four domains and forms a dimer with a deep canyon at the dimer interface [46]. This canyon, lined with numerous basic residues, opens to the active site and is postulated as the polyP binding site [46]. The active site region contains glycine-rich phosphate-binding loops named as P-loops (distinct from the P-loops previously described in kinases). Domains I and II, which form the putative Ppx active site at their interface, share structural similarity with proteins possessing a ribonuclease-H-like fold. Domain III shows structural similarity to the N-terminal, HD domain of the (p)ppGpp synthetase SpoT whereas domain IV has structural counterparts in cold-shock associated RNA-binding proteins [46]. E. coli Ppx is homologous to GppA and shares the same structural arrangement [47]. GppA regulates the pppGpp/ppGpp ratio in the cell by converting pppGpp into ppGpp [48,49]. Despite their structural similarity, Ppx preferentially acts on polyP whereas GppA targets pppGpp. This substrate specificity is attributed to conformational differences in their active sites, whereby Ppx adopts a closed conformation that hinders pppGpp binding whereas GppA adopts an open conformation that facilitates it [50]. Other Ppx proteins, such as that encoded by Aquifex aeolicus, possibly act on both substrates, as it possess an active site with an open conformation [46,50]. A. aeolicus Ppx is monomeric and lacks the C-terminal domains III and IV [51].
As noted above (see Figure 2), Lactobacillaceae typically encode two Ppx proteins: Ppx1, which possesses the four domains present in E. coli Ppx, and Ppx2, which contains only the two N-terminal catalytic domains, similar to A. aeolicus Ppx. However, available evidence suggests that Ppx1 in Lactobacillaceae is not involved in polyP hydrolysis, in contrast to all other characterized Ppx1 homologs. It was observed that Lc. paracasei BL23, harboring an in-frame deletion of ppx1 was unable to accumulate polyP. Complementation of ppx1 in trans restored the ability to synthetize polyP [30]. These results indicate not only that Ppx1 does not participate in polyP hydrolysis, but also that it is essential for polyP synthesis in Lc. paracasei. Inspection of the amino-acid sequence of Ppx1 proteins from Lactobacillaceae revealed amino-acid substitutions in key residues postulated as critical for polyphosphatase catalytic activity (Figure 6). In particular, changes were identified in the conserved P-loops and catalytic residues [30].
A phylogenetic analysis of Ppx1 homologs from taxa within the phylum Bacillota revealed two major clusters (Figure 7). Cluster I encompassed Ppx1 proteins from taxa in the orders Bacillales and Lactobacillales, excluding those from the family Lactobacillaceae. Cluster II primarily comprised proteins from Lactobacillaceae, as well as from the classes Clostridia and Negativicutes. Inspection of the sequences revealed that Cluster I Ppx1 retain conserved motifs and residues involved in polyphosphatase activity, whereas Cluster II proteins exhibit changes in part or most of those residues [30]. These findings suggested that Cluster II Ppx1 proteins lack polyphosphatase activity and might instead function as activators of their Ppk1 counterparts, as described in Lc. paracasei BL23. The mechanism by which Ppx1 regulates Ppk1 activity remains to be elucidated. Notwithstanding, it is worth noting that some species such as Oenococcus oeni and Xylocopilactobacillus apicola encode only the ppx2 gene (Figure 2). Unfortunately, evidence regarding their ability to synthesize polyP is lacking.
On the other hand, the Lc. paracasei BL23 purified Ppx2 enzyme was shown to exhibit in vitro exopolyphosphatase activity and displayed full conservation of P-loops 1 and 2, along with other key catalytic amino acid residues (Figure 6) [30]. These results suggested that Ppx2 is the main polyphosphatase in Lactobacillaceae. However, conclusive evidence is still lacking, as inactivation of ppx2 in Lc. paracasei BL23 did not affect polyP accumulation in this organism [30].
5. Physiological Roles of Polyphosphate in Lactic Acid Bacteria
PolyP is involved in a wide range of physiological processes across all domains of life [2,52,53,54], with its specific functions depending on the organism and cellular context. In bacteria, polyP participates in several processes, including: (i) serving as a structural component of lipid membranes [55] and contributing to the formation of non-protein Ca²⁺ channels [56]; (ii) supporting resistance to various stress conditions [57,58]; (iii) acting as an ATP substitute, an energy source, and a Pi reservoir [59,60,61]; (iv) contributing to nucleoid and chromosomal organization as well as gene regulation [2]; and (v) enhancing the virulence of certain pathogens [62,63]. Despite extensive research on these roles in many bacterial taxa, the involvement of polyP in the physiology of LAB remains comparatively underexplored.
The relationship between polyP and the stress response is the most extensively studied aspect of polyP metabolism in LAB and has emerged as a key factor in their microbial physiology. As a polyanion, PolyP is a strong chelator of metal cations [60], a property proposed to increase the tolerance of polyP-producing microorganisms to toxic metals. Several studies have shown that both polyP synthesis and degradation are required for its detoxifying function. A proposed model suggests that toxic metals are initially sequestered by polyP which is then degraded by polyphosphatases. The resulting metal-Pi complexes are subsequently exported via Pi transporters [64]. Whether LAB use a similar mechanism remains to be elucidated.
In LAB, heavy metals interaction has been suggested to occur primarily at the bacterial surface through binding to cell wall components [65]. Intracellular interactions between polyP and metals in LAB has only been described for Mn2+. Lp. plantarum has been shown to accumulate over 20 mM of intracellular Mn2+ [25,66] as a mechanism of defense against oxidative stress. While the exact protective mechanisms of polyP under oxidative conditions are not fully understood, in some cases mechanisms of action have been proposed. For example, its activity against superoxide ion (O2-) can be explained by its ability to coordinate cations such as Fe3+ or Mn2+, forming complexes that facilitate O2- dismutation [60]. In this model, Mn2+ is chelated by polyP, forming a Mn2+-polyP complex, which is then hydrolyzed by exopolyphosphatases to generate MnHPO4, a compound capable of detoxifying O2-.
PolyP may also inhibit the Fenton reaction, which generates highly reactive hydroxyl radicals, by chelating cations such as Fe2+ or Cu2+ and stabilizing the Fe3+ intermediates [57,60]. In, Lp. plantarum, polyP–complexed Mn²⁺ also appears to contribute directly to O2− scavenging [67]. Notably, Pi limitation reduces Mn accumulation in Lp. plantarum ATCC 14917 [68] and conversely, Mn-depleted conditions inhibit polyP synthesis in both Lp. plantarum [68] and Lacticaseibacillus rhamnosus ATCC 7469 [69]. These findings suggest a regulatory mechanism coordinating intracellular Mn and Pi accumulation.
Mutants of Lc. paracasei unable to produce polyP, due to disruption of the ppk1 gene, exhibited increased sensitivity to osmotic (NaCl), acidic (pH 4), and oxidative (plumbagin-induced) stresses compared to the wild-type strain [29]. In contrast, inactivation of ppk1 in Lp. plantarum did not affect sensitivity to osmotic or acidic stress. However, ppk1 mutants of both Lp. plantarum and Lc. paracasei were more susceptible to both inorganic and organic mercury [65].
Heat stress is a relevant factor in industrial applications of LAB. In Lc. rhamnosus CRL1505, high content of polyP in the presence of inorganic salts (MnSO₄, MgSO₄, and Pi) was associated with increased survival to heat stress [28]. Similarly, Lc. paracasei BL23 grown in hyper-concentrated (30%) sweet whey formed intracellular polyP granules, which were absent under isotonic (5% sweet whey) conditions. This high-polyP condition also correlated with enhanced survival during spray drying, suggesting a role for polyP in thermotolerance [70]. Although the precise mechanisms behind polyP-mediated stress resistance in LAB remain unclear, several pathways may be involved, including links to oxidative stress resistance and the chaperone activity of polyP [58]. Notably, the potential role of polyP in transcriptional regulation during stress response is entirely unexplored in LAB. In E. coli, the general stress response is controlled by the sigma factor σ38 (encoded by the rpoS) whose transcription require both the Ppk enzyme and polyP. Moreover, rpoS-deficient mutants fail to accumulate polyP under osmotic stress or nitrogen limitation [57,71,72]. However, LAB generally lack alternative sigma factors, implying that different, yet unidentified, regulatory mechanisms likely mediate the connection between polyP and stress responses in these organisms.
Studies using E. coli strains deficient in the ppk1 gene revealed increased sensitivity to hypochlorous acid (HOCl) treatment compared to the wild-type strain. HOCl induces severe protein damage by oxidizing amino acids such as cysteine, methionine, and histidine, leading to protein unfolding and aggregation. PolyP has been shown to interact directly with unfolded proteins, preventing their aggregation and thus mitigating HOCl-induced stress [58]. In contrast, inactivation of ppk1 in Limosilactobacillus reuteri does not significantly impair resistance to HOCl-induced protein damage. Notably, a ppk1 mutation in this species only reduces polyP levels by approximately 50%, suggesting that the Ppk2 enzyme also contributes to polyP synthesis [73]. However, mutation of ppk2 in this bacterium leads to a strong defect in HOCl resistance [74], suggesting that ATP/GTP generation via Ppk2 activity may be critical for stress survival in this species.
6. Regulation of Polyphosphate Synthesis in Lactic Acid Bacteria
Despite intensive research into bacterial polyP synthesis in recent years, the mechanisms underlying the regulation of its synthesis and degradation remain largely unknown. Early models based on E. coli proposed that the alarmone (p)ppGpp inhibits Ppx exopolyphosphatase activity, thereby promoting polyP accumulation. However, these models have been recently questioned [75], and no evidence currently supports a role for (p)ppGpp in polyP accumulation in LAB. Similar to other bacteria, polyP levels in LAB rise during exponential growth and sharply increase at the onset of the stationary phase [27], a phenomenon likely triggered by signals related to nutritional downshift. PolyP concentrations in LAB growth supernatants are much lower than intracellular levels, but they follow a similar temporal pattern. The role of polyP degradation during the stationary phase, whether mediated by Ppx or Ppk2, remains unknown. Additionally, growth under high Pi conditions significantly enhances polyP accumulation in LAB compared to growth under Pi-limited conditions [29,30]. Regulation of the transcription of polyP metabolism genes has been little studied in lactobacilli. Transcription of ppk1, ppx1 and ppx2 has been evidenced in Lc. rhamnosus CRL1505 concomitant with polyP synthesis [76] and growth under high- or low-Pi conditions had only a moderate effect on ppx/ppk expression in Lc. paracasei BL23 [30]. Furthermore, no effect of the PhoP regulator, that controls transcription of Pi-regulated genes, has been evidenced for ppk, ppx1 or ppx2 expression in this species [77]. The fact that the genes for polyP synthesizing and degrading enzymes are clustered in an operon in LAB and are jointly transcribed, probably excludes transcriptional regulation as a main control mechanism. However, a different regulation may exist for ppk2, which is not located in ppx1-ppk1-ppx2 gene clusters. In Lactiplantibacillus paraplantarum CRL1905, the enzyme Ppk2 was not detected in proteomic analyses when cells were cultivated in medium with high Pi, while it was present under low Pi conditions. This correlated with the increased accumulation and persistence of polyP at the stationary phase under high Pi in this strain [40].
An Lc. paracasei mutant strain has been isolated that displayed an enhanced polyP production (100-fold increase compared to the parental strain) [27]. In this mutant strain, extracellularly located polyP also followed a similar trend. Unfortunately, the nature of the mutation(s) producing such enhanced production is not known. A mutant strain has also been characterized from Lp. paraplantarum KCCM11826P which consumed 76 % more Pi from culture medium than its parental strain and produced more polyP. In this case, mutations in tuf, dnaK, and groL genes were found in the genome of this high polyP accumulating strain [78]. This probably links polyP accumulation to a mechanism triggered by some type of stress that resulted from defects in protein folding and/or translation elongation.
As already mentioned, in contrast to Ppx2, Ppx1 from LAB does not possess the characteristics of a true exopolyphosphatase [30]. Combined with the observation that deletion of the gene encoding this enzyme in Lc. paracasei BL23 completely abolishes polyP synthesis, this strongly suggests that polyP accumulation in LAB is controlled through signal integration affecting Ppk1 and/or Ppx2 enzymatic activity, with Ppx1 playing a regulatory role. However, the specific nature of this putative regulatory mechanism remains unknown.
7. Functional Properties of Probiotic-Derived Polyphosphate: Current Evidence
In prokaryotes, polyP is primarily found intracellularly. In contrast, higher eukaryotes, including humans, produce polyP both intra- and extracellularly, where it is involved in a wide range of physiological processes. In humans, polyP plays roles in diverse processes across different tissues, such as blood coagulation, osteogenesis and chondrogenesis, neurotransmission, regulation of mitochondrial energetics, inflammation and the response to infections, among others [52,53,79]. As previously noted, polyP produced by LAB has been identified as a functional molecule in the supernatants of probiotic cultures. This extracellular polyP contributes to the maintenance of intestinal homeostasis and has recently attracted interest for its potential therapeutic applications. The capacity of certain LAB to accumulate polyP, and excrete it to a certain level, may influence host inflammatory pathways and Pi balance. Understanding the underlying mechanisms of these effects could lead to novel strategies to manage inflammatory diseases and disorders associated with Pi imbalance, among others.
Early research showed that Lv. brevis SBC8803 induces cytoprotective heat-shock proteins in the small intestine and protects against intestinal injury in a mouse model of acute colitis [80]. The responsible factor, identified in the bacterial culture supernatant, was polyP [16]. This probiotic-derived polyP replicated the beneficial effects of the SBC8803 strain in Caco-2/BBE cells, inducing HSP27 and protecting cells against the oxidative damage caused by H2O2 or NH2Cl. In a murine colitis model, polyP suppressed the induction of the proinflammatory cytokines IL-1β and IL-6 and improved survival [16]. Mechanistic analyses indicated that polyP protects intestinal barrier function via the integrin-p38 MAPK signaling pathway with no apparent action on other MAPK pathways such as ERK, JNK or Akt. It was also evidenced that polyP interacted with integrin β1, leading to the activation of p38 MAPK [16]. Unlike other host–microbe interaction factors, polyP signaling was not mediated by pattern recognition receptors (PRR) pathways. Instead, it is internalized through the integrin–caveolin-mediated endocytic pathway and induces TNFAIP3 (tumor necrosis factor alpha-induced protein 3) expression in Caco-2/BBE cells. This provides a mechanistic basis for its anti-inflammatory properties through inhibition of TNF-α/NF-κB signaling and enhancement of intestinal barrier function [81].
Further studies in mouse models of disease support the anti-inflammatory and barrier-protective properties of LAB-derived polyP. In chronic colitis models (TNBS- and DSS-induced), polyP prevented fibrosis by suppressing expression of collagen types I and IV, along with proinflammatory cytokines IL-1β, TNF-α, and IFN-γ [82]. This effect was attributed to the downregulation of TGF-β1 expression in Caco-2/BBE epithelial cells, but not in THP-1 macrophages. Notably, polyP did not reduce collagen expression in TGF-β1-stimulated CDD-18 fibroblasts, suggesting that epithelial cells are the primary targets of polyP action. [82]. In addition, polyP isolated from Lp. plantarum (average length of 250 Pi residues) inhibited lipopolysaccharide-stimulated M1 polarization of RAW264.7 macrophages, providing another alternative anti-inflammatory mechanism [83]. Moreover, incubation of IEC-18 cells with platelets pretreated with probiotic-derived polyP led to activation of the ERK, Akt, JNK, and p38 pathways. These platelets released unidentified factors that activated the Raf–MEK signaling cascade upstream of ERK, promoting epithelial wound healing in vitro [84]. This response was linked to the beneficial effects of polyP on DSS-induced colitis in mice, where platelet accumulation at the intestinal epithelium was observed [84].
The effects of LAB-derived polyP have also been explored beyond the intestinal tract. Oral administration of polyP from Lv. brevis (1 mg/kg body weight) to mice alleviated acute pancreatitis induced by intraperitoneal injection of cerulein. Treatment with polyP suppressed histological signs of inflammation in the pancreas, diminished expression of IL-6 and the monocyte chemoattractant protein-1 (MCP-1/CCL2) in the pancreas, and lowered serum levels of amylase and lipase [85]. Additionally, polyP administration modified the composition of the intestinal microbiota, decreasing the abundance of potentially virulent bacteria such as Desulfovibrio, while increasing levels of potentially beneficial bacteria, such as Alistipes and Candidatus Saccharimonas. It remains unclear whether these microbiota changes result from a direct effect of polyP or are secondary to polyP-induced modifications of the intestinal epithelium.
Although most studies on polyP functionality have utilized polyP isolated from LAB cultures or enzymatically synthesized polyP (via Ppk), other investigations have tested functionality directly using LAB culture supernatants or whole cells. Growth supernatants from Lp. plantarum WCFS1 and Lp. plantarum Lpp+ induced HSP27 expression in Caco-2 cultures, whereas supernatants from their respective ppk1 mutants failed to elicit this response [65]. Similarly, supernatants from Lm. reuteri SBC8803 cultures induced apoptosis in the human colon carcinoma SW620 cell line, but had no such effect on primary epithelial cells derived from normal small intestine. This antitumor effect was abolished when polyP present in these supernatants was hydrolyzed by Ppx treatment [86]. In this case, the antitumor effect of polyP was associated with activation of the ERK pathway [86]. The polyP-accumulating strain Lc. rhamnosus CRL1505 was also studied in a murine model of acute respiratory inflammation induced by lipopolysaccharide [76]. Intracellular extracts rich in polyP from this strain normalized serum levels of several pro-inflammatory cytokines (IL-17, IL-6, IL-2, IL-4, IFN-γ) in nasally-treated mice, suggesting a protective effect against respiratory tract inflammation [76]. These findings support the potential of LAB-derived polyP as an innovative biotherapeutic agent for inflammatory diseases beyond the gastrointestinal tract. However, some studies have pointed to possible negative effects for polyP-accumulating LAB. For instance, certain LAB species found in the oral cavity able to accumulate polyP may have negative health impacts by facilitating tooth demineralization. Thus, experiments with Lc. rhamnosus, which inhabits caries lesions, showed that Pi draining for polyP synthesis may create physicochemical conditions that favors tooth dissolution [69].
In addition to their proposed role in maintaining host homeostasis via modulation of signaling pathways, recent studies have highlighted further biotechnological and health-related applications of polyP-producing LAB. One promising avenue is the treatment of hyperphosphatemia in patients with chronic kidney disease (CKD) by reducing intestinal phosphorus absorption. In a rat model of CKD, supplementation with Lp. paraplantarum KCCM 11826P significantly lowered serum Pi levels [87]. This attenuation is attributed to the remarkable capacity of this strain to absorb Pi and store it as polyP, suggesting a potential alternative to conventional Pi binders for phosphorus control in CKD patients [87]. Additional beneficial properties have also been ascribed to lactobacilli-derived polyP. Due to its high negative charge density, polyP is hypothesized to chelate toxic metal cations such as Hg²⁺ and Cd²⁺. Probiotic strains generally demonstrate a high capacity for binding such cations, potentially decreasing their bioavailability and toxicity in the gastrointestinal tract [88,89]. However, studies using ppk1 mutants deficient in polyP synthesis of Lc. paracasei and Lp. plantarum revealed no reduction in their metal-binding capacity despite increased sensitivity to mercury exposure [65]. These findings challenge the presumed direct role of polyP in heavy metal sequestration in LAB.
7. Future Prospects
Overall, polyP has emerged as a key factor in the functionality of probiotic LAB. Its roles in microbial physiology, particularly in stress resistance, cellular homeostasis, and adaptation to adverse environments, are increasingly recognized as central to LAB survival in the gastrointestinal tract and their ability to confer health benefits. Notwithstanding, further research is essential to optimize polyP metabolism for industrial and probiotic applications. Notably, the pathways governing polyP synthesis and degradation in LAB appear to diverge from those described in classical bacterial models, underscoring the need for targeted studies in this group. A deeper understanding of the regulatory mechanisms controlling polyP accumulation, particularly under stress conditions, is necessary to enhance its production and functional availability. Specifically, elucidating whether polyP reaches the extracellular environment through active secretion or as a result of cell lysis is essential, as this determines its availability to interact with host target cells. Finally, since polyP chain length critically influences its biological activity [79], identifying LAB strains that preferentially synthesize polyP of specific average lengths represents a promising area for strain selection and engineering. Advancing our knowledge in these areas will be instrumental for the rational development of LAB strains with enhanced functionalities tailored for a range of biotechnological and therapeutic applications.
Author Contributions
Conceptualization, M.Z.; validation, V.M. and M.Z.; investigation, D.C., C.A., V.M. and M.Z.; resources, V.M. and M.Z.; writing—original draft preparation, D.C.; writing—review and editing, V.M. and M.Z.; visualization, D.C., C.A., V.M. and M.Z.; supervision, V.M. and M.Z.; project administration, V.M. and M.Z.; funding acquisition, V.M. and M.Z.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministerio de Ciencia, Innovación y Universidades/Agencia Estatal de Investigación (MCIU/AEI/10.13039/501100011033) from Spain, grant number PID2022-137342OB-I00, and it was co-funded by the European Commission through the European Regional Development Fund (Multiregional operative program for Spain 2014–2020).
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
D. Corrales is grateful for her doctoral fellowship from the Ministerio de Ciencia, Tecnología e Innovación of Colombia (Convocatoria 906 de 2021). The Accreditation as Center of Excellence Severo Ochoa CEX2021–001189-S funded by MCIU/AEI is also fully acknowledged.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| DSS | Dextran Sodium Sulfate |
| ERK | Extracellular signal-Regulated Kinase |
| LAB | Lactic Acid Bacteria |
| MAPK | Mitogen-Activated Protein Kinase |
| PolyP | Polyphosphate |
| TNBS | 2,4,6-trinitrobenzenesulfonic acid |
References
- Dürr-Mayer, T.; Qiu, D.; Eisenbeis, V. B.; Steck, N.; Häner, M.; Hofer, A.; Mayer, A.; Siegel, J. S.; Baldridge, K. K.; Jessen, H. J. The chemistry of branched condensed phosphates. Nat. Commun. 2021, 12, 5368. [Google Scholar] [CrossRef] [PubMed]
- Racki, L. R.; Freddolino, L. Polyphosphate: The “Dark Matter” of Bacterial Chromatin Structure. Mol. Microbiol. 2025, 123, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, Y.; Watanabe, H.; Saitoh, M.; Namba, T. Volcanic production of polyphosphates and its relevance to prebiotic evolution. Nature 1991, 352, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Rao, N. N.; Gómez-García, M. R.; Kornberg, A. Inorganic Polyphosphate: Essential for Growth and Survival. Annu. Rev. Biochem. 2009, 78, 605–647. [Google Scholar] [CrossRef]
- Bowlin, M. Q.; Gray, M. J. Inorganic polyphosphate in host and microbe biology. Trends Microbiol. 2021, 29, 1013–1023. [Google Scholar] [CrossRef]
- Kulaev, I. S.; Vagabov, V. M. Polyphosphate Metabolism in Micro-Organisms. In Adv. Microb. Physiol., Rose, A. H.; Morris, J. G.; Tempest, D. W., Eds. Academic Press: 1983; Vol. 24, pp 83-171. [CrossRef]
- Beaufay, F.; Quarles, E.; Franz, A.; Katamanin, O.; Wholey, W.-Y.; Jakob, U. Polyphosphate Functions In Vivo as an Iron Chelator and Fenton Reaction Inhibitor. mBio, 2020; 11. [Google Scholar] [CrossRef]
- Kuroda, A.; Nomura, K.; Ohtomo, R.; Kato, J.; Ikeda, T.; Takiguchi, N.; Ohtake, H.; Kornberg, A. Role of Inorganic Polyphosphate in Promoting Ribosomal Protein Degradation by the Lon Protease in E. coli. Science 2001, 293, 705–708. [Google Scholar] [CrossRef]
- Gross, M. H.; Konieczny, I. Polyphosphate induces the proteolysis of ADP-bound fraction of initiator to inhibit DNA replication initiation upon stress in Escherichia coli. Nucleic Acids Res. 2020, 48, 5457–5466. [Google Scholar] [CrossRef]
- Beck, R. W. A chronology of microbiology in historical context. ASM Press: Washington, D.C., 2000.
- McGrath, J. W.; Quinn, J. P. Microbial Phosphate Removal and Polyphosphate Production from Wastewaters. In Adv. Appl. Microbiol., Academic Press: 2003; Vol. 52, pp 75-100. [CrossRef]
- Hansen–Bruhn, I.; Laura Craig, J.; Hinge, M.; Hull, T. R. Ammonium polyphosphates: Correlating structure to application. Eur. Polym. J. 2025, 223, 113644. [Google Scholar] [CrossRef]
- Müller, W. E. G.; Neufurth, M.; Wang, S.; Schröder, H. C.; Wang, X. The Physiological Inorganic Polymers Biosilica and Polyphosphate as Key Drivers for Biomedical Materials in Regenerative Nanomedicine. Int. J. Nanomedicine 2024, 19, 1303–1337. [Google Scholar] [CrossRef]
- Demling, P.; Baier, M.; Deitert, A.; Fees, J.; Blank, L. M. Biotechnological polyphosphate as an opportunity to contribute to the circularization of the phosphate economy. Curr. Opin. Biotechnol. 2024, 87, 103107. [Google Scholar] [CrossRef]
- Lemos Junior, W. J. F.; Santinello, D.; Mohammadzadeh, S.; Treu, L.; Sant’Ana, A. S.; Campanaro, S. Polyphosphate in food systems: Their roles and applications in foods and contribution to sustainable processing practices. Trends Food Sci. Technol. 2024, 152, 104696. [Google Scholar] [CrossRef]
- Segawa, S.; Fujiya, M.; Konishi, H.; Ueno, N.; Kobayashi, N.; Shigyo, T.; Kohgo, Y. Probiotic-Derived Polyphosphate Enhances the Epithelial Barrier Function and Maintains Intestinal Homeostasis through Integrin–p38 MAPK Pathway. PLoS One 2011, 6, e23278. [Google Scholar] [CrossRef] [PubMed]
- Orla-Jensen, S. The lactic acid bacteria. Fred Host and Son: Copenhagen, Denmark, 1919.
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, N.; Shakhova, V.; Grigoriev, I.; Lou, Y.; Rohksar, D.; Lucas, S.; Huang, K.; Goodstein, D. M.; Hawkins, T.; Plengvidhya, V.; Welker, D.; Hughes, J.; Goh, Y.; Benson, A.; Baldwin, K.; Lee, J. H.; Díaz-Múñiz, I.; Dosti, B.; Smeianov, V.; Wechter, W.; Barabote, R.; Lorca, G.; Altermann, E.; Barrangou, R.; Ganesan, B.; Xie, Y.; Rawsthorne, H.; Tamir, D.; Parker, C.; Breidt, F.; Broadbent, J.; Hutkins, R.; O’Sullivan, D.; Steele, J.; Unlu, G.; Saier, M.; Klaenhammer, T.; Richardson, P.; Kozyavkin, S.; Weimer, B.; Mills, D. Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. U.S.A. 2006, 103, 15611–6. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C. M. A. P.; Harris, H. M. B.; Mattarelli, P.; O’Toole, P. W.; Pot, B.; Vandamme, P.; Walter, J.; Watanabe, K.; Wuyts, S.; Felis, G. E.; Gänzle, M. G.; Lebeer, S. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ). Update of the list of qualified presumption of safety (QPS) recommended microbiological agents intentionally added to food or feed as notified to EFSA 21: Suitability of taxonomic units notified to EFSA until September 2024. EFSA J. 2025, 23, e9169. [CrossRef]
- Neville, N.; Roberge, N.; Jia, Z. Polyphosphate Kinase 2 (PPK2) Enzymes: Structure, Function, and Roles in Bacterial Physiology and Virulence. Int. J. Mol. Sci. 2022, 23, 670. [Google Scholar] [CrossRef]
- Bolesch, D. G.; Keasling, J. D. Polyphosphate Binding and Chain Length Recognition of Escherichia coli Exopolyphosphatase. J. Biol. Chem. 2000, 275, 33814–33819. [Google Scholar] [CrossRef]
- Hugues, D. E.; Muhammed, D. The metabolism of polyphosphate in bacteria. Colloq. Int. Centre Natl. Recherche Sci. (Paris) 1962, 106, 591–602. [Google Scholar]
- Kakefuda, T.; Holden, J. T.; Utech, N. M. Ultrastructure of the Membrane System in Lactobacillus plantarum. J. Bacteriol. 1967, 93, 472–482. [Google Scholar] [CrossRef]
- Archibald, F. S.; Fridovich, I. Investigations of the state of the manganese in Lactobacillus plantarum. Arch. Biochem. Biophys. 1982, 215, 589–596. [Google Scholar] [CrossRef]
- Aprea, G.; Mullan, W. M. A.; Mullan, A.; Murru, N.; Tozzi, M.; Cortesi, M. L. Isolation of polyphosphate-accumulating lactic acid bacteria from natural whey starters. Milchwissenschaft 2005, 60, 256–258. [Google Scholar]
- Saiki, A.; Ishida, Y.; Segawa, S.; Hirota, R.; Nakamura, T.; Kuroda, A. A Lactobacillus mutant capable of accumulating long-chain polyphosphates that enhance intestinal barrier function. Biosci. Biotechnol. Biochem. 2016, 80, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Correa Deza, M. A.; Grillo-Puertas, M.; Salva, S.; Rapisarda, V. A.; Gerez, C. L.; Font de Valdez, G. Inorganic salts and intracellular polyphosphate inclusions play a role in the thermotolerance of the immunobiotic Lactobacillus rhamnosus CRL 1505. PLoS One 2017, 12, e0179242. [Google Scholar] [CrossRef] [PubMed]
- Alcántara, C.; Blasco, A.; Zúñiga, M.; Monedero, V. Accumulation of polyphosphate in Lactobacillus spp. and its involvement in stress resistance. Appl. Environ. Microbiol. 2014, 80, 1650–1659. [Google Scholar] [CrossRef]
- Corrales, D.; Alcántara, C.; Zúñiga, M.; Monedero, V. Ppx1 putative exopolyphosphatase is essential for polyphosphate accumulation in Lacticaseibacillus paracasei. Appl. Environ. Microbiol. 2024, 90, e02290–23. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K.; Kornberg, A. Polyphosphate kinase from Escherichia coli. Purification and demonstration of a phosphoenzyme intermediate. J. Biol. Chem. 1990, 265, 11734–11739. [Google Scholar] [CrossRef]
- Akiyama, M.; Crooke, E.; Kornberg, A. The polyphosphate kinase gene of Escherichia coli. Isolation and sequence of the ppk gene and membrane location of the protein. J. Biol. Chem. 1992, 267, 22556–22561. [Google Scholar] [CrossRef]
- Kornberg, A.; Kornberg, S. R.; Simms, E. S. Metaphosphate synthesis by an enzyme from Escherichia coli. Biochimica et biophysica acta 1956, 20, 215–227. [Google Scholar] [CrossRef]
- Suzuki, H.; Kaneko, T.; Ikeda, Y. Properties of polyphosphate kinase prepared from Mycobacterium smegmatis. Biochim. Biophys. Acta 1972, 268, 381–390. [Google Scholar] [CrossRef]
- Levinson, S. L.; Jacobs, L. H.; Krulwich, T. A.; Li, H.-C. Purification and characterization of a polyphosphate kinase from Arthrobacter atrocyaneus. Microbiology 1975, 88, 65–74. [Google Scholar] [CrossRef]
- Robinson, N. A.; Clark, J. E.; Wood, H. G. Polyphosphate kinase from Propionibacterium shermanii. Demonstration that polyphosphates are primers and determination of the size of the synthesized polyphosphate. J. Biol. Chem. 1987, 262, 5216–5222. [Google Scholar] [CrossRef]
- Robinson, N. A.; Wood, H. G. Polyphosphate kinase from Propionibacterium shermanii. Demonstration that the synthesis and utilization of polyphosphate is by a processive mechanism. J. Biol. Chem. 1986, 261, 4481–4485. [Google Scholar] [CrossRef]
- Tinsley, C. R.; Manjula, B. N.; Gotschlich, E. C. Purification and characterization of polyphosphate kinase from Neisseria meningitidis. Infect. Immun. 1993, 61, 3703–3710. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Huang, W.; Lee, S. S. K.; Xu, W. Crystal structure of a polyphosphate kinase and its implications for polyphosphate synthesis. EMBO Rep. 2005, 6, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Araoz, M.; Grillo-Puertas, M.; de Moreno de LeBlanc, A.; Hebert, E. M.; Villegas, J. M.; Rapisarda, V. A. Inorganic phosphate modifies stationary phase fitness and metabolic pathways in Lactiplantibacillus paraplantarum CRL 1905. Front. Microbiol. 2024, 15, 1343541. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M. H.; Rao, N. N.; Kornberg, A. Inorganic Polyphosphate Is Required for Motility of Bacterial Pathogens. J. Bacteriol. 2000, 182, 225–227. [Google Scholar] [CrossRef]
- Ishige, K.; Zhang, H.; Kornberg, A. Polyphosphate kinase (PPK2), a potent, polyphosphate-driven generator of GTP. Proc. Natl. Acad. Sci. U.S.A. 2002, 99, 16684–16688. [Google Scholar] [CrossRef]
- Leipe, D. D.; Koonin, E. V.; Aravind, L. Evolution and Classification of P-loop Kinases and Related Proteins. J. Mol. Biol. 2003, 333, 781–815. [Google Scholar] [CrossRef]
- Motomura, K.; Hirota, R.; Okada, M.; Ikeda, T.; Ishida, T.; Kuroda, A. A New Subfamily of Polyphosphate Kinase 2 (Class III PPK2) Catalyzes both Nucleoside Monophosphate Phosphorylation and Nucleoside Diphosphate Phosphorylation. Appl. Environ. Microbiol. 2014, 80, 2602–2608. [Google Scholar] [CrossRef]
- Akiyama, M.; Crooke, E.; Kornberg, A. An exopolyphosphatase of Escherichia coli. The enzyme and its ppx gene in a polyphosphate operon. J. Biol. Chem. 1993, 268, 633–639. [Google Scholar] [CrossRef]
- Rangarajan, E. S.; Nadeau, G.; Li, Y.; Wagner, J.; Hung, M.-N.; Schrag, J. D.; Cygler, M.; Matte, A. The Structure of the Exopolyphosphatase (PPX) from Escherichia coli O157:H7 Suggests a Binding Mode for Long Polyphosphate Chains. J. Mol. Biol. 2006, 359, 1249–1260. [Google Scholar] [CrossRef]
- Keasling, J. D.; Bertsch, L.; Kornberg, A. Guanosine pentaphosphate phosphohydrolase of Escherichia coli is a long-chain exopolyphosphatase. Proc. Natl. Acad. Sci. U.S.A. 1993, 90, 7029–7033. [Google Scholar] [CrossRef] [PubMed]
- Hara, A.; Sy, J. Guanosine 5′-triphosphate, 3′-diphosphate 5′-phosphohydrolase. Purification and substrate specificity. J. Biol. Chem. 1983, 258, 1678–1683. [Google Scholar] [CrossRef]
- Mechold, U.; Potrykus, K.; Murphy, H.; Murakami, K. S.; Cashel, M. Differential regulation by ppGpp versus pppGpp in Escherichia coli. Nucleic Acids Res. 2013, 41, 6175–6189. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Dharmasena, M. N.; Wang, C.; Shaw, G. X.; Cherry, S.; Tropea, J. E.; Jin, D. J.; Ji, X. Structure and activity of PPX/GppA homologs from Escherichia coli and Helicobacter pylori. FEBS J. 2020, 287, 1865–1885. [Google Scholar] [CrossRef]
- Kristensen, O.; Laurberg, M.; Liljas, A.; Kastrup, J. S.; Gajhede, M. Structural Characterization of the Stringent Response Related Exopolyphosphatase/Guanosine Pentaphosphate Phosphohydrolase Protein Family. Biochemistry 2004, 43, 8894–8900. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Jakob, U. Polyphosphate: a cellular Swiss army knife. Curr. Opin. Biotechnol. 2025, 93, 103303. [Google Scholar] [CrossRef]
- Schoeppe, R.; Waldmann, M.; Jessen, H. J.; Renné, T. An Update on Polyphosphate In Vivo Activities. Biomolecules 2024, 14, 937. [Google Scholar] [CrossRef]
- Rosigkeit, H.; Kneißle, L.; Obruča, S.; Jendrossek, D. The Multiple Roles of Polyphosphate in Ralstonia eutropha and Other Bacteria. Microb. Physiol. 2021, 31, 163–177. [Google Scholar] [CrossRef]
- Reusch, R. N.; Sadoff, H. L. Putative structure and functions of a poly-beta-hydroxybutyrate/calcium polyphosphate channel in bacterial plasma membranes. Proc. Natl. Acad. Sci. U.S.A. 1988, 85, 4176–4180. [Google Scholar] [CrossRef]
- Jendrossek, D.; Pfeiffer, D. New insights in the formation of polyhydroxyalkanoate granules (carbonosomes) and novel functions of poly(3-hydroxybutyrate). Environ. Microbiol. 2014, 16, 2357–2373. [Google Scholar] [CrossRef]
- Gray, M. J.; Jakob, U. Oxidative stress protection by polyphosphate—new roles for an old player. Curr. Opin. Microbiol. 2015, 24, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Gray, M. J.; Wholey, W.-Y.; Wagner, N. O.; Cremers, C. M.; Mueller-Schickert, A.; Hock, N. T.; Krieger, A. G.; Smith, E. M.; Bender, R. A.; Bardwell, J. C. A.; Jakob, U. Polyphosphate Is a Primordial Chaperone. Mol. Cell 2014, 53, 689–699. [Google Scholar] [CrossRef]
- Kornberg, A.; Rao, N. N.; Ault-Riché, D. Inorganic Polyphosphate: A Molecule of Many Functions. Annu. Rev. Biochem. 1999, 68, 89–125. [Google Scholar] [CrossRef] [PubMed]
- Albi, T.; Serrano, A. Inorganic polyphosphate in the microbial world. Emerging roles for a multifaceted biopolymer. World J. Microbiol. Biotechnol. 2016, 32, 27. [Google Scholar] [CrossRef]
- Albi, T.; Serrano, A. Two strictly polyphosphate-dependent gluco(manno)kinases from diazotrophic Cyanobacteria with potential to phosphorylate hexoses from polyphosphates. Appl. Microbiol. Biotechnol. 2015, 99, 3887–3900. [Google Scholar] [CrossRef]
- Rojas, D.; Marcoleta, A. E.; Gálvez-Silva, M.; Varas, M. A.; Díaz, M.; Hernández, M.; Vargas, C.; Nourdin-Galindo, G.; Koch, E.; Saldivia, P.; Vielma, J.; Gan, Y.-H.; Chen, Y.; Guiliani, N.; Chávez, F. P. Inorganic Polyphosphate Affects Biofilm Assembly, Capsule Formation, and Virulence of Hypervirulent ST23 Klebsiella pneumoniae. ACS Infect. Dis. 2024, 10, 606–623. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Zhou, Y.; Liu, B.; Guan, J.; Zhang, P.; Deng, X.; Li, D.; Wang, J. Polyphosphate Kinase Is Required for the Processes of Virulence and Persistence in Acinetobacter baumannii. Microbiol. Spectr. 2022, 10, e01230–22. [Google Scholar] [CrossRef]
- Keasling, J. D. Regulation of Intracellular Toxic Metals and Other Cations by Hydrolysis of Polyphosphate. Ann. N. Y. Acad. Sci. 1997, 829, 242–249. [Google Scholar] [CrossRef]
- Alcántara, C.; Coll-Marqués, J. M.; Jadán-Piedra, C.; Vélez, D.; Devesa, V.; Zúñiga, M.; Monedero, V. Polyphosphate in Lactobacillus and its link to stress tolerance and probiotic properties. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Archibald, F. S.; Fridovich, I. Manganese, superoxide dismutase, and oxygen tolerance in some lactic acid bacteria. J. Bacteriol. 1981, 146, 928–936. [Google Scholar] [CrossRef]
- Archibald, F. Manganese: Its Acquisition by and Function in the Lactic Acid Bacteria. Crit. Rev. Microbiol. 1986, 13, 63–109. [Google Scholar] [CrossRef] [PubMed]
- Archibald, F. S.; Duong, M. N. Manganese acquisition by Lactobacillus plantarum. J. Bacteriol. 1984, 158, 1–8. [Google Scholar] [CrossRef]
- Breiland, A. A.; Flood, B. E.; Nikrad, J.; Bakarich, J.; Husman, M.; Rhee, T.; Jones, R. S.; Bailey, J. V. Polyphosphate-Accumulating Bacteria: Potential Contributors to Mineral Dissolution in the Oral Cavity. Appl. Environ. Microbiol. 2018, 84, e02440–17. [Google Scholar] [CrossRef]
- Huang, S.; Gaucher, F.; Cauty, C.; Jardin, J.; Le Loir, Y.; Jeantet, R.; Chen, X. D.; Jan, G. Growth in Hyper-Concentrated Sweet Whey Triggers Multi Stress Tolerance and Spray Drying Survival in Lactobacillus casei BL23: From the Molecular Basis to New Perspectives for Sustainable Probiotic Production. Front. Microbiol. 2018, 9, 2548. [Google Scholar] [CrossRef] [PubMed]
- Ault-Riché, D.; Fraley, C. D.; Tzeng, C.-M.; Kornberg, A. Novel Assay Reveals Multiple Pathways Regulating Stress-Induced Accumulations of Inorganic Polyphosphate in Escherichia coli. J. Bacteriol. 1998, 180, 1841–1847. [Google Scholar] [CrossRef] [PubMed]
- Shiba, T.; Tsutsumi, K.; Yano, H.; Ihara, Y.; Kameda, A.; Tanaka, K.; Takahashi, H.; Munekata, M.; Rao, N. N.; Kornberg, A. Inorganic polyphosphate and the induction of rpoS expression. Proc. Natl. Acad. Sci. U.S.A. 1997, 94, 11210–11215. [Google Scholar] [CrossRef]
- Pokhrel, A.; Lingo, J. C.; Wolschendorf, F.; Gray, M. J. Assaying for Inorganic Polyphosphate in Bacteria. J. Vis. Exp. 2019, 143, e58818. [Google Scholar] [CrossRef]
- Thakur, P. B.; Long, A. R.; Nelson, B. J.; Kumar, R.; Rosenberg, A. F.; Gray, M. J. Complex Responses to Hydrogen Peroxide and Hypochlorous Acid by the Probiotic Bacterium Lactobacillus reuteri. mSystems 2019, 4. [Google Scholar] [CrossRef]
- Gray, M. J. Inorganic Polyphosphate Accumulation in Escherichia coli Is Regulated by DksA but Not by (p)ppGpp. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef]
- Correa Deza, M. A.; Rodríguez de Olmos, A.; Suárez, N. E.; Font de Valdez, G.; Salva, S.; Gerez, C. L. Inorganic polyphosphate from the immunobiotic Lactobacillus rhamnosus CRL1505 prevents inflammatory response in the respiratory tract. Saudi J. Biol. Sci. 2021, 28, 5684–5692. [Google Scholar] [CrossRef]
- Corrales, D.; Alcántara, C.; Vélez, D.; Devesa, V.; Monedero, V.; Zúñiga, M. Unveiling the role of the PhoP master regulator in arsenite resistance through ackA downregulation in Lacticaseibacillus paracasei. Curr. Res. Microb. Sci. 2025, 8, 100357. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J. P.; Moon, S. J.; Lee, M.; Kwon, M. Y.; Pyo, J.-Y.; Lee, S. C.; Kim, H. J. Engineering polyphosphate-accumulating probiotics for therapeutic applications in chronic kidney disease. Food Biosci. 2025, 68, 106360. [Google Scholar] [CrossRef]
- Kus, F.; Smolenski, R. T.; Tomczyk, M. Inorganic Polyphosphate—Regulator of Cellular Metabolism in Homeostasis and Disease. Biomedicines 2022, 10, 913. [Google Scholar] [CrossRef]
- Ueno, N.; Fujiya, M.; Segawa, S.; Nata, T.; Moriichi, K.; Tanabe, H.; Mizukami, Y.; Kobayashi, N.; Ito, K.; Kohgo, Y. Heat-killed body of Lactobacillus brevis SBC8803 ameliorates intestinal injury in a murine model of colitis by enhancing the intestinal barrier function. Inflamm. Bowel Dis. 2011, 17, 2235–2250. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Fujiya, M.; Konishi, H.; Ueno, N.; Kashima, S.; Sasajima, J.; Moriichi, K.; Ikuta, K.; Tanabe, H.; Kohgo, Y. Probiotic-derived polyphosphate improves the intestinal barrier function through the caveolin-dependent endocytic pathway. Biochem. Biophys. Res. Commun. 2015, 467, 541–548. [Google Scholar] [CrossRef]
- Kashima, S.; Fujiya, M.; Konishi, H.; Ueno, N.; Inaba, Y.; Moriichi, K.; Tanabe, H.; Ikuta, K.; Ohtake, T.; Kohgo, Y. Polyphosphate, an active molecule derived from probiotic Lactobacillus brevis, improves the fibrosis in murine colitis. Transl. Res. 2015, 166, 163–175. [Google Scholar] [CrossRef]
- Li, S.; Zheng, A.; Chen, Z.; Wang, X.; Chen, J.; Zou, Z.; Liu, G. Lactobacillus plantarum-Derived Inorganic Polyphosphate Regulates Immune Function via Inhibiting M1 Polarization and Resisting Oxidative Stress in Macrophages. Antioxidants 2025, 14, 428. [Google Scholar] [CrossRef]
- Isozaki, S.; Konishi, H.; Fujiya, M.; Tanaka, H.; Murakami, Y.; Kashima, S.; Ando, K.; Ueno, N.; Moriichi, K.; Okumura, T. Probiotic-Derived Polyphosphate Accelerates Intestinal Epithelia Wound Healing through Inducing Platelet-Derived Mediators. Mediators Inflamm. 2021, 2021, 5582943. [Google Scholar] [CrossRef]
- Takauji, S.; Konishi, H.; Fujiya, M.; Ueno, N.; Tanaka, H.; Sato, H.; Isozaki, S.; Kashima, S.; Moriichi, K.; Mizukami, Y.; Okumura, T. Polyphosphate, Derived from Lactobacillus brevis, Modulates the Intestinal Microbiome and Attenuates Acute Pancreatitis. Dig. Dis. Sci. 2021, 66, 3872–3884. [Google Scholar] [CrossRef]
- Sakatani, A.; Fujiya, M.; Ueno, N.; Kashima, S.; Sasajima, J.; Moriichi, K.; Ikuta, K.; Tanabe, H.; Kohgo, Y. Polyphosphate Derived from Lactobacillus brevis Inhibits Colon Cancer Progression Through Induction of Cell Apoptosis. Anticancer Res. 2016, 36, 591–598. [Google Scholar]
- Moon, S. J.; Hwang, J.; Kang, W. K.; Ahn, J.-P.; Kim, H. J. Administration of the probiotic Lactiplantibacillus paraplantarum is effective in controlling hyperphosphatemia in 5/6 nephrectomy rat model. Life Sci. 2022, 306, 120856. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Viso, P.; Domene, A.; Sánchez, A.; Vélez, D.; Monedero, V.; Devesa, V.; Zúñiga, M. Challenges and strategies for preventing intestinal damage associated to mercury dietary exposure. Toxicology 2023, 494, 153580. [Google Scholar] [CrossRef] [PubMed]
- Chiocchetti, G. M.; Jadán-Piedra, C.; Monedero, V.; Zúñiga, M.; Vélez, D.; Devesa, V. Use of lactic acid bacteria and yeasts to reduce exposure to chemical food contaminants and toxicity. Crit. Rev. Food Sci. Nutr. 2018, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structure of a polyphosphate (polyP) molecule. The number of phosphate residues (n) can range from tens to several hundreds.
Figure 1.
Structure of a polyphosphate (polyP) molecule. The number of phosphate residues (n) can range from tens to several hundreds.
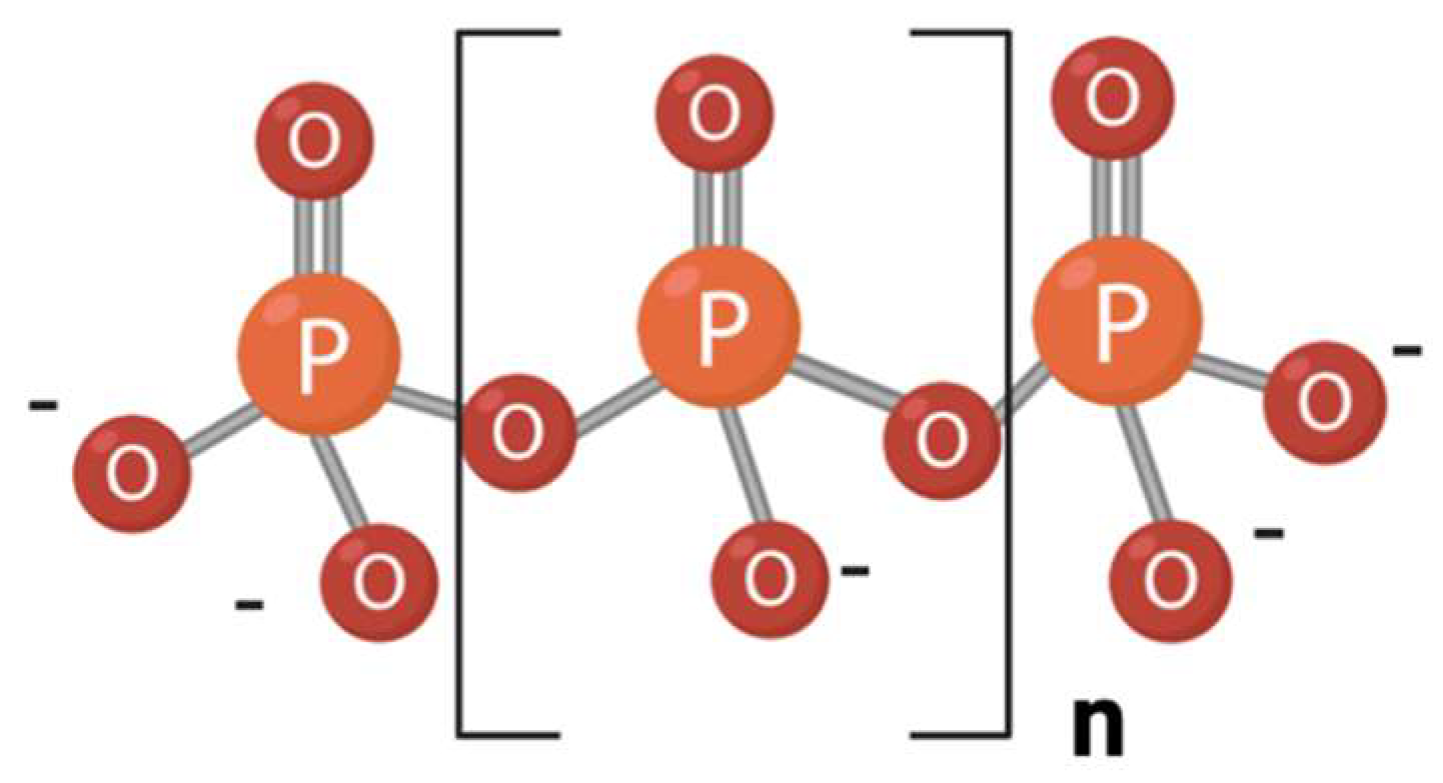
Figure 2.
PolyP metabolic gene clusters identified in representative sequenced genomes of bacterial species within the Lactobacillaceae family. The presence of ppk, ppx1, and ppx2 homologs, along with the occurrence of similar flanking genes across different strains and species, is depicted. Homologous genes are indicated by matching colors. A double black bar represents an intervening genomic region not shown to scale.
Figure 2.
PolyP metabolic gene clusters identified in representative sequenced genomes of bacterial species within the Lactobacillaceae family. The presence of ppk, ppx1, and ppx2 homologs, along with the occurrence of similar flanking genes across different strains and species, is depicted. Homologous genes are indicated by matching colors. A double black bar represents an intervening genomic region not shown to scale.
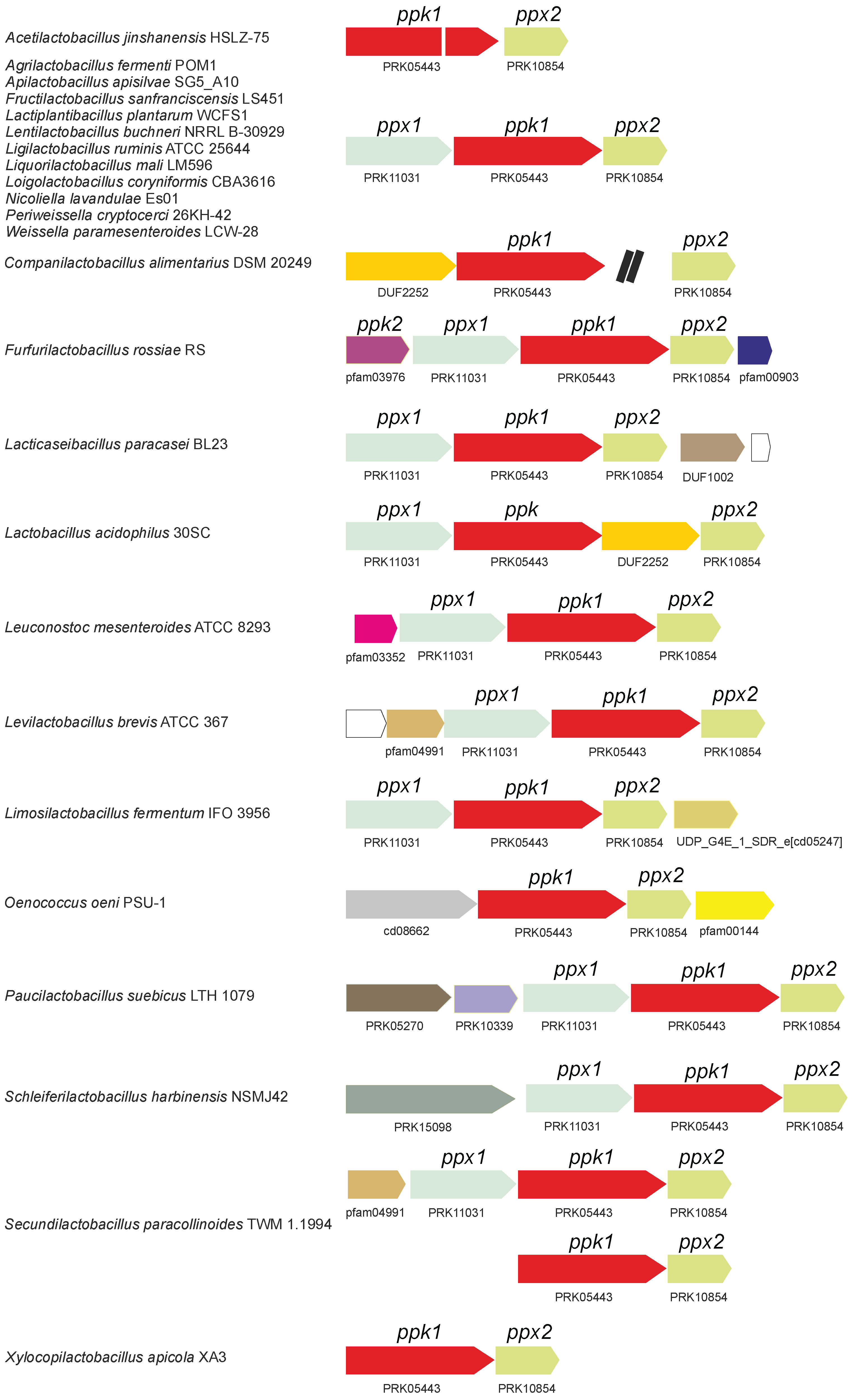
Figure 3.
Sequence alignment of polyphosphate kinases (Ppk1) from representative Lactobacillaceae species and Escherichia coli K12 (NP_416996.1). Lactobacillaceae sequences comprise Lactobacillus intestinalis (WP_304402131.1), Leuconostoc citreum (WP_349532772.1), Lc. paracasei BL23 (CAQ67833.1), Loigolactobacillus coryniformis (WP_146990929.1), Lp. plantarum WCFS1 (CCC78296.1), Lv. brevis (WP_141373826.1) and Secundilactobacillus similis (WP_057152190.1). Identical or highly conserved residues are shaded in yellow. Secondary structure elements, derived from the E. coli Ppk1 structure as described by Zhu et al. [39], are shown above the sequences: α-helices are depicted as bars and β-strands as arrows. Colors denote the four structural domains of E. coli Ppk1. The phosphorylated residue H435 and the residues involved in autophosphorylation (D470 and E623) are marked with red stars.
Figure 3.
Sequence alignment of polyphosphate kinases (Ppk1) from representative Lactobacillaceae species and Escherichia coli K12 (NP_416996.1). Lactobacillaceae sequences comprise Lactobacillus intestinalis (WP_304402131.1), Leuconostoc citreum (WP_349532772.1), Lc. paracasei BL23 (CAQ67833.1), Loigolactobacillus coryniformis (WP_146990929.1), Lp. plantarum WCFS1 (CCC78296.1), Lv. brevis (WP_141373826.1) and Secundilactobacillus similis (WP_057152190.1). Identical or highly conserved residues are shaded in yellow. Secondary structure elements, derived from the E. coli Ppk1 structure as described by Zhu et al. [39], are shown above the sequences: α-helices are depicted as bars and β-strands as arrows. Colors denote the four structural domains of E. coli Ppk1. The phosphorylated residue H435 and the residues involved in autophosphorylation (D470 and E623) are marked with red stars.
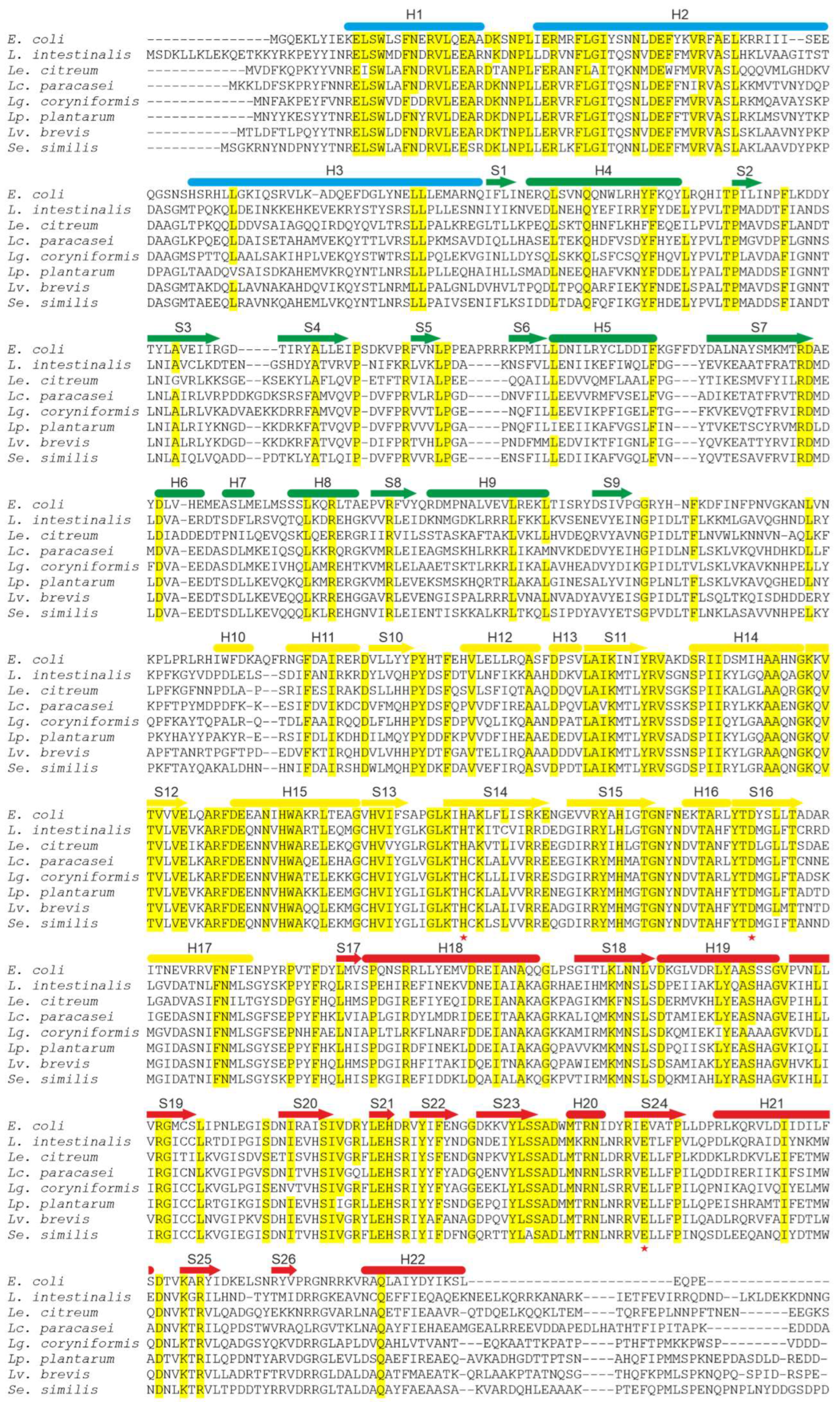
Figure 4.
Enzymatic reactions carried out by the different polyphosphate kinases.
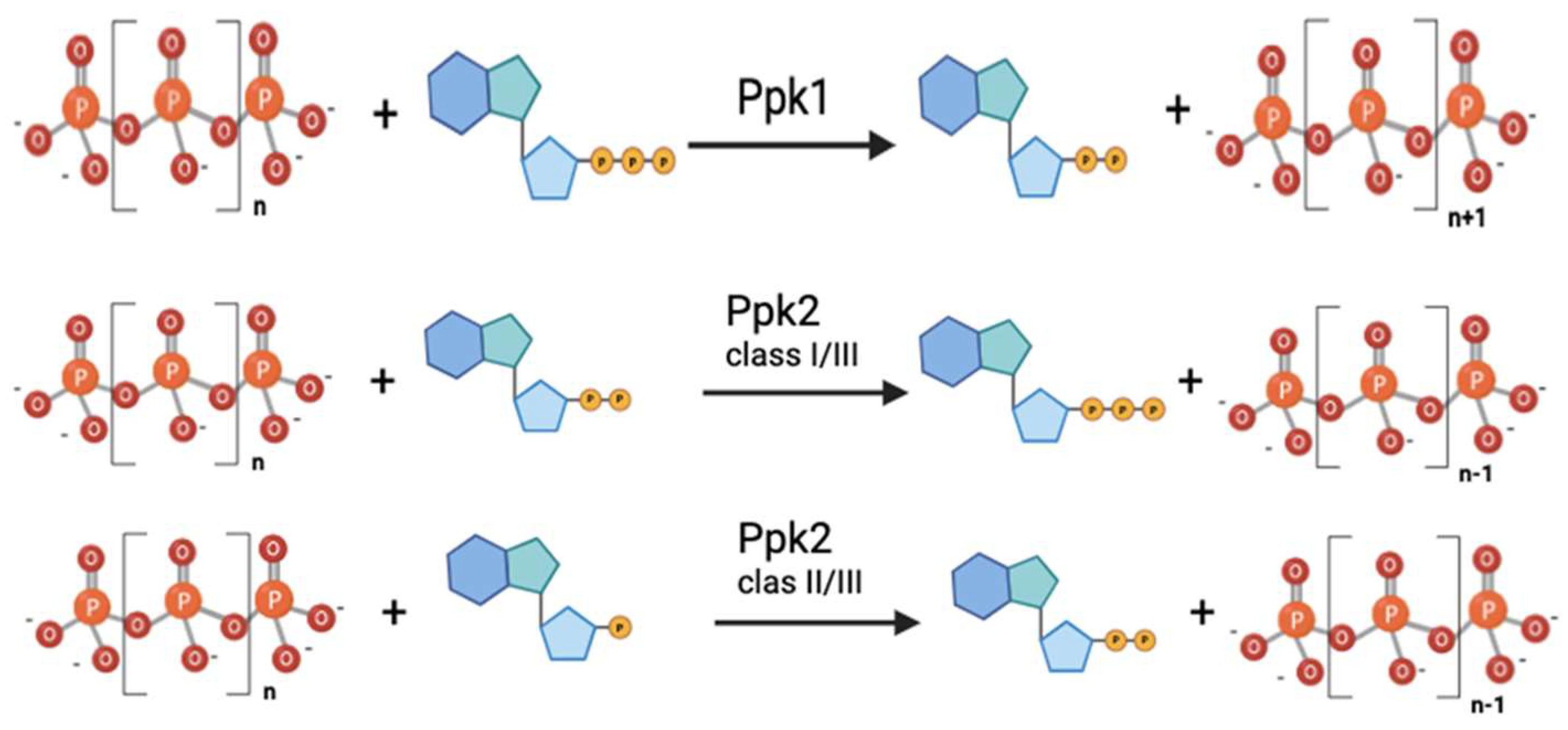
Figure 5.
Sequence alignment of selected Lactobacillaceae species and representative sequences of the three subfamilies of polyphosphate kinases 2 proteins (Ppk2). Identical or highly conserved residues are shaded in yellow. The Walker A motif is designated by a red bar, the Walker B motif is indicated by a green bar, and the lid module is indicated by a black bar, as identified by Nocek et al. (2008). Lactobacillaceae sequences comprise Lc. paracasei BL23 (CAQ66314.1), Lp. plantarum WCFS1 (WP_003642687.1), Agrilactobacillus composti DSM 18527 (KRM35117.1), Furfurilactobacillus rossiae DSM 15814 (KRL53865.1) and L. intestinalis DSM 6629 (KRM32733.1). Sinorhizobium meliloti 1021 (WP_010968631.1) represents class I Ppk2, Pseudomonas aeruginosa PAO1 (NP_252145.1) represents class II, and Paenarthrobacter aurescens TC1 (WP_261609683.1) represents class III. The two domains of P. aeruginosa class II Ppk2 are aligned separatedly and indicated as Nt and Ct, respectively.
Figure 5.
Sequence alignment of selected Lactobacillaceae species and representative sequences of the three subfamilies of polyphosphate kinases 2 proteins (Ppk2). Identical or highly conserved residues are shaded in yellow. The Walker A motif is designated by a red bar, the Walker B motif is indicated by a green bar, and the lid module is indicated by a black bar, as identified by Nocek et al. (2008). Lactobacillaceae sequences comprise Lc. paracasei BL23 (CAQ66314.1), Lp. plantarum WCFS1 (WP_003642687.1), Agrilactobacillus composti DSM 18527 (KRM35117.1), Furfurilactobacillus rossiae DSM 15814 (KRL53865.1) and L. intestinalis DSM 6629 (KRM32733.1). Sinorhizobium meliloti 1021 (WP_010968631.1) represents class I Ppk2, Pseudomonas aeruginosa PAO1 (NP_252145.1) represents class II, and Paenarthrobacter aurescens TC1 (WP_261609683.1) represents class III. The two domains of P. aeruginosa class II Ppk2 are aligned separatedly and indicated as Nt and Ct, respectively.

Figure 6.
Schematic representation of Ppx1 and Ppx2. The presence of different protein domains (ASKHA and HDc) is depicted, together with relevant portions of multiple sequence alignments of Ppx1 from Lc. paracasei (WP_003606084.1), Lv. brevis (AYM03703.1), Lp. plantarum (WP_103851489.1), Limosilactobacillus reuteri (MCH5356744.1), and Lactobacillus acidophilus (MCT3601606.1) and from Ppx2 from Lc. paracasei (CAQ67832.1), Lv. brevis (WP_135367400.1), Lp. plantarum (WP_003641094.1), Lm. reuteri (KRK50994.1), and L. acidophilus (AZN76677.1), compared with Ppx from E. coli (WP_001121363.1). The consensus sequences of the conserved P-loop 1 and P-loop 2 in the GppA/Ppx family are shown in blue above the alignments. The catalytic amino acids arginine and glutamic acid are marked with red diamonds. The amino acids characteristic of the HDc domain in C-terminal Ppx1 are marked with red circles. Reproduced with permission from Corrales et al. [30].
Figure 6.
Schematic representation of Ppx1 and Ppx2. The presence of different protein domains (ASKHA and HDc) is depicted, together with relevant portions of multiple sequence alignments of Ppx1 from Lc. paracasei (WP_003606084.1), Lv. brevis (AYM03703.1), Lp. plantarum (WP_103851489.1), Limosilactobacillus reuteri (MCH5356744.1), and Lactobacillus acidophilus (MCT3601606.1) and from Ppx2 from Lc. paracasei (CAQ67832.1), Lv. brevis (WP_135367400.1), Lp. plantarum (WP_003641094.1), Lm. reuteri (KRK50994.1), and L. acidophilus (AZN76677.1), compared with Ppx from E. coli (WP_001121363.1). The consensus sequences of the conserved P-loop 1 and P-loop 2 in the GppA/Ppx family are shown in blue above the alignments. The catalytic amino acids arginine and glutamic acid are marked with red diamonds. The amino acids characteristic of the HDc domain in C-terminal Ppx1 are marked with red circles. Reproduced with permission from Corrales et al. [30].
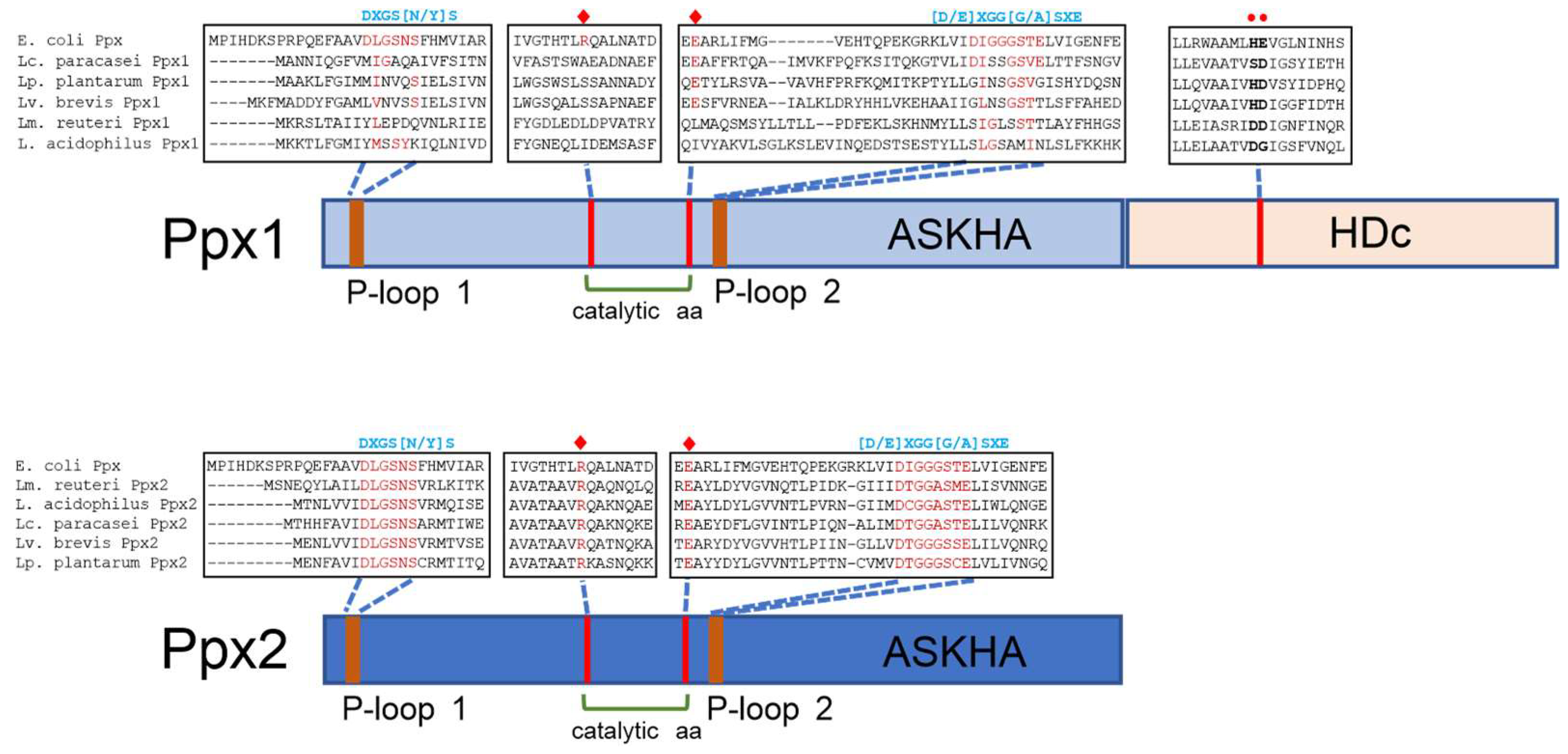
Figure 7.
Maximum likelihood phylogenetic tree of Ppx1 proteins of selected species of the phylum Bacillota. Support values are given for those nodes with support higher than 75%. The tree has been arbitrarily rooted for ease of visualization. Colors: black: class Negativicutes; brown: class Clostridia; blue: class Tissierellia; green: order Lactobacillales; and red: order Bacillales. Reproduced with permission from Corrales et al. [30].
Figure 7.
Maximum likelihood phylogenetic tree of Ppx1 proteins of selected species of the phylum Bacillota. Support values are given for those nodes with support higher than 75%. The tree has been arbitrarily rooted for ease of visualization. Colors: black: class Negativicutes; brown: class Clostridia; blue: class Tissierellia; green: order Lactobacillales; and red: order Bacillales. Reproduced with permission from Corrales et al. [30].
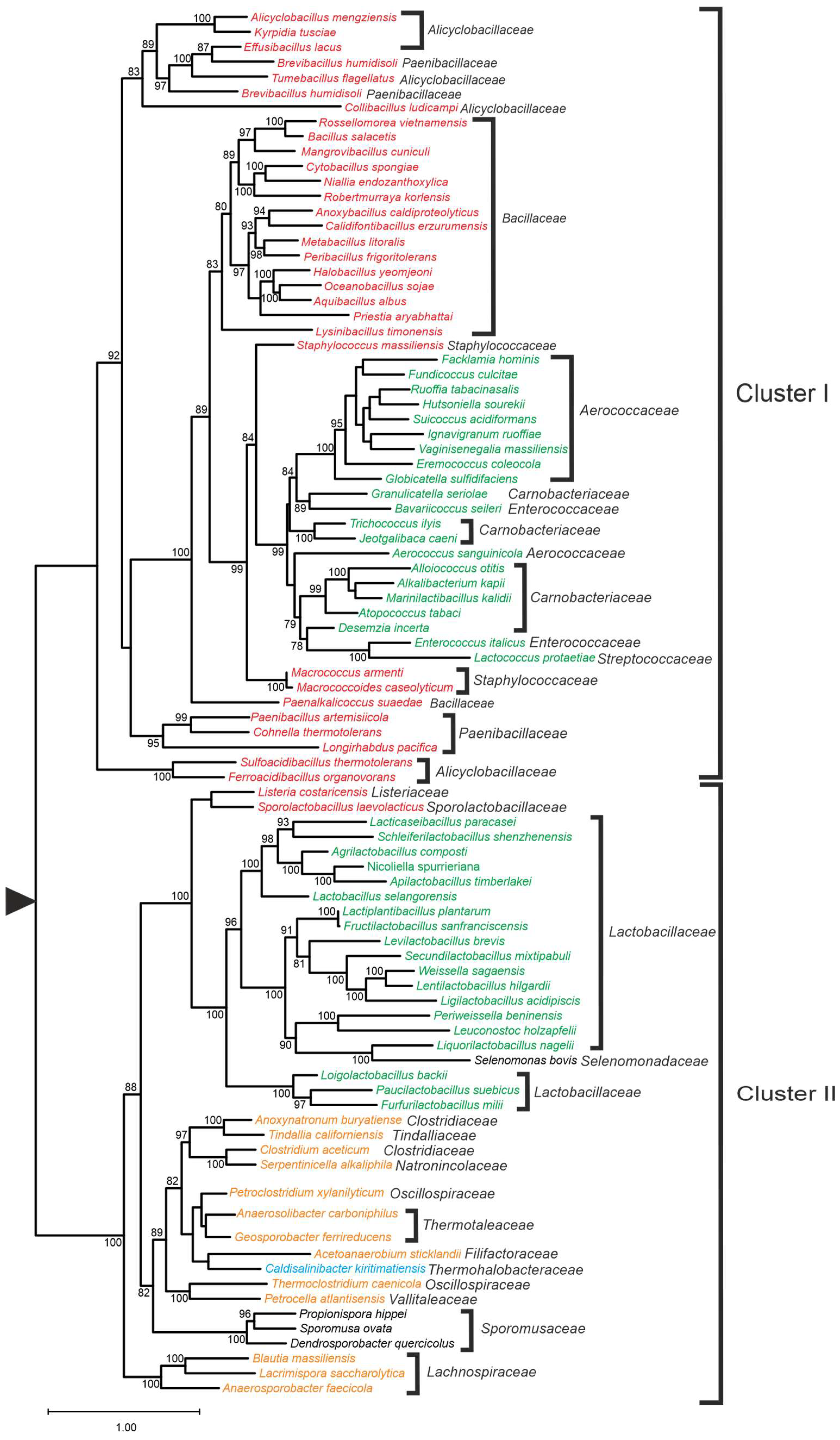

Table 1.
Presence of polyP metabolic genes in family Lactobacillaceae.
| Genus | ppk1 | ppk2 | ppx1 | ppx2 |
|---|---|---|---|---|
| Acetilactobacillus | + | - | - | + |
| Agrilactobacillus | + | ± | + | + |
| Amylolactobacillus | - | - | - | - |
| Apilactobacillus | ± | - | ± | ± |
| Bombilactobacillus | - | - | - | ± |
| Companilactobacillus | + | - | - | + |
| Convivina | - | - | - | + |
| Dellaglioa | - | - | - | + |
| Eupransor | + | - | - | + |
| Fructilactobacillus | + | - | ± | + |
| Fructobacillus | ± | - | - | + |
| Furfurilactobacillus | + | + | + | + |
| Holzapfeliella | - | - | - | - |
| Lacticaseibacillus | + | ± | + | + |
| Lactiplantibacillus | + | + | + | + |
| Lactobacillus | ± | ± | ± | ± |
| Lapidilactobacillus | - | - | - | - |
| Latilactobacillus | - | - | - | - |
| Lentilactobacillus | ± | ± | ± | + |
| Leuconostoc | + | - | + | + |
| Levilactobacillus | + | + | + | + |
| Ligilactobacillus | ± | - | ± | ± |
| Limosilactobacillus | + | + | + | + |
| Liquorilactobacillus | + | ± | + | + |
| Loigolactobacillus | ± | ± | ± | ± |
| Nicoliella | + | - | + | + |
| Oenococcus | + | - | - | + |
| Paucilactobacillus | ± | - | ± | + |
| Pediococcus | - | - | - | + |
| Periweissella | + | - | + | + |
| Philodulcilactobacillus | + | - | - | + |
| Schleiferilactobacillus | + | + | + | + |
| Secundilactobacillus | ± | - | ± | + |
| Weissella | ± | - | ± | + |
| Xylocopilactobacillus | + | - | - | + |
Note: Genes were detected by blastp using as query sequences Ppk1 (CAQ67834.1), Ppk2 (CAQ66314.1), Ppx1 (CAQ67834) and Ppx2 (CAQ67832) from Lc. paracasei BL23 against the nr database in NIH blast server. + >90% species harbour the target gene, - 0%, ± <90%.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



