
Submitted:
12 May 2025
Posted:
14 May 2025
You are already at the latest version
Abstract
Porcine reproductive and respiratory syndrome virus (PRRSV) poses a significant economic challenge to Japan's swine industry. This review synthesizes the epidemiological evolution of PRRSV in Japan by examining the available scientific literature from its initial Type 2 isolation in 1993 to recent events. Endemic Type 2 strains, initially dominated by Cluster III (Lineage 4), diversified significantly. This diversification was marked by key events including the emergence of vaccine-associated Cluster II (Lineage 5), and incursions of virulent Cluster IV/Lineage 1F (MN184A-like) strains and Type 1 virus around 2008. By 2018-2020, Clusters II and IV predominated nationwide, a trend strongly linked to widespread modified live virus (MLV) vaccination. The recent detection of the globally significant NADC34-like (Lineage 1A) strain underscores ongoing foreign incursion risks. Current MLV vaccines face challenges, including safety concerns and limited cross-protection against diverse field strains. Consequently, effective control requires integrated strategies, comprising optimized vaccination, stringent biosecurity, advanced molecular surveillance, improved diagnostics, and coordinated regional control programs guided by systematic herd classification and stakeholder partnerships. The development of next-generation vaccines and sustained multi-stakeholder collaboration are critical for mitigating the impact of PRRSV in Japan.
Keywords:
PRRSV (porcine reproductive and respiratory syndrome virus)
; epidemiology
; Japan
; molecular epidemiology
; control strategy
1. Introduction
Porcine reproductive and respiratory syndrome (PRRS), caused by the PRRS virus (PRRSV), is one of the most economically significant viral diseases affecting the global swine industry [1,2]. The disease is characterized by reproductive failure in late-gestation sows, including abortions, premature farrowing, increased stillbirths and mummified fetuses, and the birth of weak piglets, as well as respiratory distress, poor growth, and immunosuppression in pigs of all ages, particularly during the nursery and fattening stages [2]. Owing to its substantial economic and social impact, PRRS is listed by the World Organisation for Animal Health (WOAH, formerly OIE) [3], consequently affecting international animal quarantine and pork trade. Estimated annual economic losses are immense, reaching approximately USD 664 million in the United States [4] and JPY 28 billion (estimated in 2010) in Japan [5,6], thereby causing significant damage to swine production worldwide.
PRRSV, a positive-sense single-stranded RNA virus, belongs to the family Arteriviridae, order Nidovirales [1]. Its genome is approximately 15 kb long and contains at least ten open reading frames (ORFs). The virus exhibits remarkable genetic diversity and is broadly classified into two major genotypes: Type 1 (European genotype, Betaarterivirus suid 1) and Type 2 (North American genotype, Betaarterivirus suid 2), sharing only approximately 60% nucleotide identity between them [1]. Furthermore, each genotype possesses a high mutation rate (estimated at 10−2 to 10−3 substitutions/site/year [7]) and comprises numerous genetic lineages and clusters. This extensive genetic diversity and rapid evolution pose major obstacles to the development of effective control measures and broadly cross-protective vaccines [8,9]. Moreover, host genetic factors significantly influence the porcine response to PRRSV infection, adding another layer of complexity to disease control [10].
PRRSV primarily infects porcine alveolar macrophages (PAMs), monocytes, and dendritic cells [2] employing sophisticated mechanisms to evade the host immune response. These mechanisms include the suppression of type I interferon (IFN-I) production, delayed induction of neutralizing antibodies (NAbs), potential antibody-dependent enhancement (ADE) of infection, and induction of immunosuppressive cytokines such as IL-10 [11]. Such evasion contributes to persistent infection, often lasting several months, and increased susceptibility to secondary infections, frequently as part of the Porcine Respiratory Disease Complex (PRDC) wherein PRRSV interacts with other respiratory pathogens [12]. The sophisticated mechanisms by which PRRSV evades the host immune system have been comprehensively reviewed elsewhere [13]. Recent studies have revealed that specific viral proteins, such as Nsp5, interfere with key innate immune signaling pathways, including the cGAS-STING pathway [14].
In Japan, PRRS remains a primary challenge for the swine industry. Following suspected cases in the late 1980s [15] and initial clinical outbreaks around 1990 in the Kanto region [15], PRRSV was first isolated domestically in 1993 [16,17]. The initial isolates were predominantly Type 2 [16,18], which has since become endemic. Although, genetically relatively limited strains circulated initially [18], viral genetic diversity has increased over time [19]. This increase includes the incursion of novel genetic lineages from abroad, such as Type 1 [20] and virulent Type 2 strains [21]. Currently, multiple genetic clusters co-circulate within Japan, with dominant types varying by region and time (Figure 1) [22]. Furthermore, the circulation of vaccine-derived strains from widely used modified live virus (MLV) vaccines, coupled with the risk of recombination between vaccine and field strains, poses significant concerns [22,23].
This review aims to provide a comprehensive and chronological overview of the epidemiology of PRRS in Japan, from its initial discovery to the present, based on the available scientific literature. Specifically, it will cover: (1) the discovery of PRRSV and the characteristics of early genotypes (clusters) in Japan; (2) the introduction of Type 1 virus and virulent strains, leading to increased genetic diversity; (3) the impact of PRRS infection on production performance and its estimated economic losses; (4) recent trends in viral circulation (changes in cluster composition) and their association with vaccine use; (5) the latest risk posed by the incursion of foreign strains (e.g., NADC34-like virus); (6) challenges related to vaccines and preventive measures; and (7) examples of regional control efforts in Japan and future prospects. By organizing and synthesizing this information, this review seeks to clarify the current status and challenges of PRRS control in the Japanese swine industry and contribute to the development of more effective future control strategies.
2. Discovery of PRRSV and Characteristics of Early Genotypes in Japan (1993–2007)
The history of PRRS in Japan dates back to the late 1980s. Antibodies against PRRSV were detected in sera collected from domestic pigs in 1987 [15], suggesting that the virus might have been present before clinical outbreaks were recognized. The first clinical reports emerged around 1990 from multiple farms in the Kanto region [15], describing collective reproductive failures (e.g., abortions, stillbirths) and respiratory problems in piglets. These clinical manifestations closely resembled the emerging disease syndromes reported in North America ("Mystery Swine Disease") and Europe ("Blue-eared pig disease") at that time [1,2]. Subsequent investigations led to the first official isolation of PRRSV in Japan in 1993 from pigs exhibiting "Heko-Heko disease" on a farm in Chiba Prefecture (strain EDRD-1) [16,17].
From the beginning, PRRSV strains detected in Japan were predominantly of the North American type (Type 2) [16,18], potentially reflecting the origins of imported breeding stock during that period. Yoshii et al. [18] analyzed the ORF5 gene (encoding the major envelope glycoprotein GP5, a highly variable region utilized for phylogenetic analysis) of 30 PRRSV isolates collected from 23 prefectures in Japan between 1992 and 2001, confirming that all were NA-type. Their phylogenetic analysis, including global NA-type strains, revealed that NA-type viruses could be broadly classified into five major genetic clusters (Cluster I–V). This analysis further showed that a majority (20 out of 32, 63%) of the Japanese isolates belonged to a specific cluster, designated Cluster III [18]. Although Cluster III included some isolates from China and Taiwan, it was considered a distinct genetic lineage predominantly established within Japan. Furthermore, Cluster III isolates exhibited a tendency for greater geographical distribution in the eastern regions of Japan [18]. However, isolates belonging to Cluster I, which includes US isolates and the PrimePac® vaccine strain, were also present among the 1992–1993 Japanese isolates, primarily found in the western regions of Japan [18]. By the 2000–2001 period, isolates belonging to Cluster II (containing the Ingelvac® PRRS MLV vaccine strain and strains from the US and Asia) and Cluster V were detected, albeit in small numbers, along with some unclassified strains [18]. These findings indicated a gradual increase in the genetic diversity of PRRSV within Japan during the early 2000s [18,19].
Iseki et al. [19] compared ORF5 sequences from Japanese PRRSV isolates collected during three periods (1992–1993, 2000–2001, and 2007–2008) and confirmed a trend of increasing genetic heterogeneity over time. In particular, the nucleotide identity among Cluster III isolates decreased from an average of 95.7% (relative to reference strain EDRD-1) in 1992–1993 to 91.2% in 2007–2008, indicating ongoing genetic divergence within this cluster. While Cluster III remained the most prevalent cluster in 2007–2008 (accounting for 12 out of 17 isolates, 71%), Cluster II isolates were detected more frequently (3 out of 17, 17%) [19]. Many of these Cluster II isolates were genetically very close to the Ingelvac® PRRS MLV vaccine strain which was widely used in Japan at that time, suggesting the potential circulation and establishment of vaccine-derived strains in the field [19]. These findings highlight the establishment of the NA-type virus in Japan, the presence of a unique predominant domestic lineage (Cluster III), and increasing genetic diversity, including the emergence of Cluster II potentially linked to vaccine use, by the late 2000s.
3. Incursion of European Type and Expansion of Genetic Diversity (2008–2013)
Until 2007, all PRRSV strains identified in Japan were Type 2. However, the genetic landscape of PRRSV in the country became significantly more complex in the subsequent years. Iseki et al., in their survey conducted during 2007–2008, identified a novel genetic lineage distinct from Clusters I, II, III, and V [19]. This new lineage, represented by the isolate Jpn5-37, was designated Cluster IV [19]. The ORF5 gene of Jpn5-37 shared a high nucleotide identity (97.1%) with MN184A, a virulent strain isolated in Minnesota, USA, in 2001 [21]. Further analysis by Iseki et al. [21] revealed that the full genome of Jpn5-37 shared 93.6% identity with MN184A, exhibiting particularly high homology in ORF1b, ORF6, and ORF7 (91.8–97.3%). Crucially, the non-structural protein 2 (NSP2) gene region of Jpn5-37 exhibited three discontinuous deletions totaling 131 amino acids when compared to the VR-2332 prototype strain (111 aa + 1 aa + 19 aa deletions), a pattern identical to that found in MN184A [21]. The NSP2 gene is known to be the most variable region in the PRRSV genome, and specific deletions, particularly large ones such as the 111-aa deletions in MN184A/Jpn5-37, have been associated with increased virulence [21,24]. Yoshii et al. [25] had previously characterized the NSP2 of the endemic Japanese Cluster III strain EDRD-1, noting a unique 117-bp (39-aa) deletion and a 108-bp (36-aa) insertion (defined as nsp2-type E) relative to VR-2332. In contrast, Jpn5-37 and MN184A lacked this deletion/insertion pattern (belonging to nsp2-type V) but possessed the distinct large deletions within NSP2 [21,25]. Animal experiments confirmed the higher pathogenicity of Jpn5-37 compared to EDRD-1, with Jpn5-37-infected animals exhibiting significantly higher viremia levels and more pronounced fever [21]. These genetic and pathogenic characteristics strongly suggested that Cluster IV represented a newly introduced Virulent-PRRSV lineage, likely originating from the US.
Furthermore, in 2008, the first isolation of European type (Type 1) PRRSV in Japan was officially reported [20]. The isolate, Jpn EU 4-37, was found to be genetically related to Type 1 strains based on ORF5 sequence analysis. Animal experiments conducted by Iseki et al. [26] showed that while Jpn EU 4-37 alone caused only mild pathology [27], prior immunity to Type 2 PRRSV (EDRD-1) did not prevent infection with or viremia by Jpn EU 4-37. Conversely, prior Type 1 immunity did partially reduce viremia following Type 2 challenge [26]. These findings indicated that the introduction of the Type 1 virus posed a new epidemiological risk, even in herds endemic for the Type 2 virus.
Thus, from 2008 onwards, Japan faced a situation characterized by the co-circulation of diverse PRRSV strains: the endemic Japanese lineage (Cluster III / nsp2-type E) [18,25], vaccine-related lineages (Cluster II / nsp2-type V) [19], a virulent North American lineage (Cluster IV / nsp2-type V with deletions) [19,21], and the European type (Type 1) [20]. This rapid expansion of genetic diversity significantly complicated PRRS diagnosis, vaccine selection, and control strategies, heralding a new and more challenging phase for PRRS management in the Japanese swine industry.
4. Impact on Production Performance and Economic Losses (2010 Survey and National Estimates)
The spread of PRRS has directly impacted the productivity of Japanese pig farms, posing a major economic burden. Several studies have quantitatively assessed this impact. Ishizeki et al. [28] conducted a cross-sectional study of 92 farrow-to-finish farms in 2010, analyzing the association between PRRSV status and production performance. Farm PRRSV status was determined based on routine serological (ELISA) and/or virological (PCR) testing under veterinary supervision. The study revealed a high prevalence of PRRSV-positive farms (76 out of 92, 82.6%) [28]. Comparison of production parameters revealed that PRRSV-positive farms had significantly higher post-weaning mortality (6.1% vs. 3.9% in negative farms; p<0.01) and significantly lower average daily gain (ADG) (585.1 g vs. 638.8 g; p<0.01), resulting in considerably longer days to market (196.9 days vs. 180.3 days; p<0.01) [28]. No significant differences were observed in carcass weight, feed conversion ratio, or reproductive parameters such as farrowing rate and litter size. These findings suggesst that PRRS infection primarily affects the performance of growing-finishing pigs, potentially by reducing feed intake and growth rate, thereby necessitating extended feeding periods to reach market weight.
The decline in production performance inevitably translates into economic losses. Yamane et al. conducted a detailed study to estimate these losses. Based on data from six PRRS-affected farms [6], they calculated loss units per sow per day for specific clinical manifestations: JPY 22 for increased pre-weaning mortality, JPY 57 for increased post-weaning mortality, JPY 35 for increased finishing mortality, JPY 84 for abortions, JPY 30 for stillbirths, and JPY 32 for reduced growth rate. Abortions and post-weaning mortality were identified as having the largest economic impact [6]. Furthermore, Yamane et al. utilized these loss units, combined with data from a nationwide questionnaire survey of 121 farms (of which 65% reported experiencing PRRS outbreaks in the previous two years), to simulate annual economic losses. The simulation estimated a total annual loss of approximately JPY 3.525 billion for the 121 surveyed farms. Extrapolating this figure to the national sow population at that time (approximately 900,000 sows) resulted in an estimated nationwide annual economic loss of approximately JPY 28 billion [5]. This study was the first in Japan to quantify the substantial economic impact of PRRS, clearly demonstrating the severity of the damage [5,6,29]. This estimate provided a strong rationale for prioritizing PRRS control measures and investments in research and development by both governmental bodies and the swine industry. The economic burden of PRRS encompasses not only direct losses from mortality and reduced growth but also increased costs for medication, vaccination, and disinfection as well as losses due to secondary infections.
5. Shift in Cluster Composition and Association with Vaccines (2018–2020)
The genetic landscape of PRRSV in Japan continued to evolve into the late 2010s. Historically, PRRSV genotypes in Japan have been classified into Clusters I to V based on ORF5 sequences [18]. While this classification is widely utilized in domestic research and will be primarily employed in this review, the international standard is increasingly shifting towards a more detailed Lineage classification system, such as that proposed by Yim-im et al. [30], which comprises 11 Lineages (L1–L11) and 21 Sub-lineages. The correspondence between these systems, according to Yonezawa et al. [31], is approximately as follows: Cluster I correspond to Lineage 8, Cluster II to Lineage 5, Cluster III to Lineage 4, and Cluster IV to Lineage 1. Cluster V is also considered part of Lineage 4. Efforts are ongoing to refine PRRSV genetic classification systems globally to better track viral evolution and diversity, including proposals for dynamic nomenclature systems [32]. Bearing this correspondence in mind, this section discusses the recent shift in cluster composition in Japan and its potential association with vaccine use, focusing on the study by Kyutoku et al. [22].
Kyutoku et al. [22] analyzed ORF5 sequences from 2,482 PRRSV-positive samples collected nationwide (32 prefectures, approximately 260 farms/year) between January 2018 and December 2020. It should be noted that these samples were collected under a diagnostic program initiated by a specific vaccine manufacturer and thus may not be fully representative of the entire Japanese pig population. Their extensive analysis revealed that Cluster II (accounting for 44.9–50.6% of total isolates) and Cluster IV (accounting for 34.0–40.8%) were the predominantly detected clusters during this period, collectively accounting for over 80% of the isolates [22]. In contrast, the proportion of Cluster III, previously the dominant endemic lineage in 2007–2008 [19], had significantly decreased to 7.8–12.1% (p<0.01) [22]. Cluster I accounted for 3.1–6.7% of isolates, and Cluster V for only 0.1–0.2% [22]. These finding indicates a major shift in the dominant PRRSV genotypes in Japan during the 2010s.
Significant regional variations in cluster distribution were also observed. In 2018, Cluster II was dominant across all regions. However, in 2019, Cluster II remained dominant only in the Hokkaido and Tohoku regions, while Cluster IV became dominant in other regions (Kanto/Tosan, Tokai, Chugoku/Shikoku, and Kyushu/Okinawa). In 2020, Cluster IV was dominant in Kanto/Tosan and Kyushu/Okinawa, whereas Cluster II predominated elsewhere, illustrating a complex and dynamic epidemiological situation [22].
The study strongly suggested that the widespread use of MLV played a significant role in this shift [22]. Many farms included in the study utilized MLV vaccines, primarily Ingelvac® PRRS MLV or Fostera® PRRS. A large majority of the detected Cluster II isolates exhibited extremely high ORF5 sequence identity (average >98%) to the Ingelvac® PRRS MLV strain (derived from US strain VR-2332, belonging to Lineage 5A) [22]. This finding strongly suggests circulation of the vaccine strain itself, or slightly mutated derivatives thereof, or detection from vaccinated animals. Similarly, some Cluster I isolates showed high identity (97–100%) to the Fostera® PRRS vaccine strain (derived from US strain P129, Lineage 8C) [22]. Fostera® PRRS was commercially introduced in Japan in 2018, potentially linking its use to the detection of these Cluster I strains. Conversely, no isolates showed high homology to the PrimePac® vaccine strain (Cluster I/Lineage 7), which is not approved for use in Japan [22]. These findings suggest that the long-term, widespread use of Ingelvac® PRRS MLV and the more recent introduction of Fostera® PRRS may be exerting selection pressure (i.e., vaccine pressure) on the domestic PRRSV population, favoring the spread of specific clusters (Clusters II and I) highly homologous to the vaccine strains [22]. While the increase in Cluster IV (Lineage 1) might be attributed to its high infectivity [21], its association with virulent strains such as MN184A (L1F) [21] and the NADC-like strains (L1A, L1C) warrants particular attention [31].
6. Incursion of Foreign Strains and New Risks (2024: Emergence of NADC34-like strain)
The genetic diversity of PRRSV in Japan is influenced not only by domestic evolution and vaccine pressure but also by the constant risk of the incursion of novel viral lineages from abroad. Examples include the first confirmation of Type 1 virus in 2008 [20] and the emergence of Cluster IV (MN184A-like/Lineage 1F) [19,21]. In 2024, this risk materialized anew with the detection of another significant foreign lineage.
Yonezawa et al. [31] reported the first detection in Japan of PRRSV-2 strains belonging to Lineage 1, Sub-lineage 1.5 (L1A), from serum samples obtained from 18 piglets (42–56 days old) on a farm in Okinawa Prefecture. Phylogenetic analysis placed these Okinawan strains in a cluster closely related to the US NADC34 strain and NADC34-like strains isolated in Peru and China [31]. The NADC34 strain, first identified in the US in 2014, has rapidly spread globally in recent years, particularly across Asia (e.g., China, Korea, Thailand, and Taiwan), often exhibiting high pathogenicity (especially causing abortion storms in sows) and inflicting severe economic losses [24,31]. Studies in China have confirmed the prevalence and high pathogenicity of NADC34-like strains in sow herds [33].
Analysis by Yonezawa et al. [31] showed that the ORF5 gene of the Okinawan L1A isolates shared 90.9–92.9% (average 91.9%) nucleotide identity with the US NADC34 reference strain. This identity was significantly higher than that with other Lineage 1 reference strains such as NADC30 (L1C), MN184 (L1F), or Jpn5-37 (L1F). Restriction Fragment Length Polymorphism (RFLP) analysis using the MluI-HincII-SacII enzyme combination [34] revealed that 10 of the 18 Okinawan isolates displayed the "1-7-4" pattern characteristic of NADC34, while the remaining 8 exhibited the "1-4-4" pattern often observed in NADC30 strains [31]. This RFLP pattern distribution is consistent with reports for L1A strains (where 59.4% show the 1-7-4 and 11.4% show the 1-4-4 pattern), further supporting their classification as NADC34-like, although RFLP patterns alone are not definitive for genotyping [31].
Prediction of N-linked glycosylation sites in the GP5 protein sequence identified glycosylation at asparagine residue 57 (N57) in 12 of the 18 Okinawan isolates (67%) [31]. This N57 glycosylation, along with the conserved N44 and N51 sites, is a characteristic feature that has been re-emerging in US L1A strains since 2015 [24,31]. The presence of strains lacking N57 glycosylation (6 isolates) on the same farm suggests the co-circulation or evolution of multiple variants [31]. Based on the combined evidence from phylogenetic analysis, RFLP patterns, and glycosylation site characteristics, Yonezawa et al. [31] concluded that the isolates detected in Okinawa were NADC34-like strains (L1A), marking the first official report of this lineage in Japan.
NADC34-like strains pose a significant threat owing to their potential high pathogenicity and antigenic divergence from existing vaccine strains, raising concerns about reduced cross-protection afforded by current vaccines [24,31]. Indeed, development of new vaccines targeting NADC34-like strains is underway in countries such as China. Interestingly, the farm in Okinawa where these strains were detected reportedly experienced only a transient increase in piglet mortality and mild respiratory signs, without severe clinical manifestations like abortion storms typically associated with NADC34 [31]. This discrepancy could be attributed to lower pathogenicity of the specific isolates, farm-specific environmental factors, co-infections, or the herd's immune status, all of which require further investigation. Regardless of the observed pathogenicity in this specific case, the confirmed incursion of the NADC34-like lineage into Japan represents a serious warning for the nation's PRRS control efforts. The potential spread of this virus to other regions could lead to significant damage, possibly overwhelming existing control measures. This finding underscores the urgent need to strengthen national PRRSV surveillance, particularly by monitoring for incursions of foreign strains, and to develop and update contingency plans for responding to outbreaks caused by novel, high-consequence lineages.
7. Challenges in Vaccines and Preventive Measures
Vaccination is a cornerstone of PRRS control worldwide. In Japan, while the primary vaccines currently approved and utilized are modified live virus (MLV) vaccines based on North American type (Type 2) strains, such as Ingelvac® PRRS MLV (Lineage 5A) and Fostera® PRRS (Lineage 8C), inactivated vaccines (KV) (Nisseiken PRRS Vaccine ME) are also available as an option (Table 1) [22].
MLV vaccines have demonstrated efficacy in reducing clinical signs (e.g., reproductive failure, respiratory disease), decreasing viral shedding, and shortening the duration of infection in numerous studies [8,9]. However, current MLV vaccines face several significant challenges and limitations: 1. Incomplete Protection: MLV vaccines do not provide sterilizing immunity; consequently, they cannot completely prevent infection, viral replication, or transmission. Vaccinated pigs can still become infected with field strains, exhibit mild symptoms, and shed virus, thereby acting as potential sources of infection [8,9]. 2. Safety Concerns: MLV vaccine viruses can persist in vaccinated animals for weeks to months, posing a risk of shedding and transmission to unvaccinated animals. Transmission via semen from vaccinated boars is a notable concern. Furthermore, vaccination of pregnant sows, especially in late gestation, carries a risk of transplacental fetal infection of fetuses, potentially causing reproductive losses [8,9]. 3. Risk of Reversion and Recombination: As live viruses, MLVs can mutate during replication, presenting a theoretical risk of reverting to a more virulent form. A more significant concern is the potential for genetic recombination between MLV vaccine strains and circulating field strains (or even different MLV strains) when co-infection occurs [8,23]. Such recombination events have been reported globally [23] and can lead to the emergence of novel viruses with altered antigenicity or increased virulence, sometimes resulting in severe outbreaks. 4. Limited Heterologous Protection: Owing to the vast genetic diversity of PRRSV [18,19], immunity induced by one specific vaccine strain often fails to provide effective protection against genetically distant field strains [8,9,50]. This lack of broad cross-protection is a major limitation. For example, protection between Type 1 and Type 2 viruses is generally poor [26]. Even within Type 2, protection against different lineages (e.g., L1 vs. L5 vs. L8) can be insufficient [22,30]. This is particularly relevant with the emergence of novel strains such as NADC34-like (L1A) [31], where existing vaccine efficacy may be compromised. Indeed, studies evaluating commercial MLV vaccines (including VR2332-derived strains like Ingelvac®) against recent NADC34-like isolates have shown only partial effectiveness, highlighting this challenge [35]. Interestingly, reduced dosage strategies for some MLV vaccines are being explored for potential benefits against NADC34-like strains, although further investigation is required [36]. Experimental studies in Japan comparing Japanese Type 1 (Jpn EU 4-37) and Type 2 (EDRD-1) isolates revealed asymmetric cross-protection: prior Type 1 infection partially reduced Type 2 viremia, whereas prior Type 2 infection offered no protection against Type 1 challenge [26]. Conversely, prior infection with the endemic Japanese strain EDRD-1 (Lineage 4) did significantly reduce viremia and clinical signs upon challenge with a heterologous highly pathogenic PRRSV strain from Vietnam (Lineage 8) [37], suggesting some cross-protection is possible depending on the specific strains involved. 5. Delayed and Skewed Immune Response: PRRSV's inherent immune evasion mechanisms can lead to delayed induction of neutralizing antibodies and cell-mediated immunity, even following vaccination [8,11]. The induced response may also be skewed towards less effective Th2 immunity [11].
To address these challenges, global research efforts are focused on developing next-generation vaccines [8,9,38]. Inactivated vaccines, while generally safer, require improved immunogenicity, often pursued through advanced adjuvants (e.g., nanoparticles) and antigen processing techniques. Vector vaccines, which use other viruses or bacteria to deliver PRRSV antigens, offer the potential for enhanced safety and immunogenicity. Subunit vaccines, utilizing specific viral proteins or epitopes, provide high safety and design flexibility but necessitate potent adjuvants and delivery systems (e.g., baculovirus expression system, plant-based expression, nanoparticles, multi-epitope constructs). Nucleic acid vaccines (DNA and mRNA) represent a rapidly evolving platform, offering the potential for rapid development and the induction of both humoral and cellular immunity, although delivery and stability remain key areas for improvement [38]. Promising results have been observed with novel platforms like ferritin nanoparticle-based vaccines, which demonstrated the induction of protective immune responses [39].
In the Japanese context, MLVs remain the primary tool, but their application requires careful consideration of each farm's specific situation. Kawabata [40] demonstrated that on a PRRS-endemic farm in a high-density area in Kagoshima that a two-dose MLV vaccination strategy (0.5 dose at 1 and 3 weeks of age) was more effective in reducing mortality and improving growth compared to a single dose administrated at 3 weeks. These findings suggested that early infection (pre-weaning) was occurring, and the two-dose regimen likely provided earlier immunity while potentially mitigating interference from maternal antibodies. This underscores the importance of understanding the infection dynamics on each farm (e.g., prevalent strain type, timing of infection) for optimizing vaccine selection and administration timing. Combination vaccination strategies, such as an MLV prime followed by inactivated or DNA boost, are also being explored internationally to enhance immunity [38].
However, reliance solely on vaccination is insufficient for effective PRRS control. Stringent biosecurity is crucial for both preventing virus introduction and controlling its spread within the farm. Comprehensive, science-based biosecurity programs (often termed "Next Generation Biosecurity") have demonstrated significant reductions in PRRSV incidence risk in large production systems [41]. Specific interventions such as air filtration and the use of feed mitigants have also shown protective effects against outbreaks [42]. These measures include external biosecurity (e.g., quarantine and testing of incoming animals, control of vehicle/personnel entry, disinfection, pest control) and internal measures (all-in/all-out pig flow, thorough cleaning and disinfection between batches, proper needle usage, movement control from contaminated to clean areas).
Accurate diagnosis and monitoring are also vital, yet the genetic diversity of PRRSV poses challenges to these efforts. Fukunaga et al. [43] highlighted that some conventional PCR primers designed before 2010 may fail to detect recent Japanese PRRSV strains owing to mutations in primer binding sites. This emphasizes the need for updated diagnostic tools, such as their newly designed primer pair (PRRSV-M-2F/4R), to ensure reliable detection.
8. Future Actions and Prospects
Experience in Japan has demonstrated that controlling PRRS solely through individual farm efforts is often insufficient, particularly in densely populated pig production areas where the risk of virus re-introduction from neighboring infected farms via aerosols, personnel, vehicles, or other fomites is high [40,44]. Consequently, regional control and elimination programs, defined as deliberate efforts to reduce disease incidence to locally acceptable levels or to achieve eradication within a defined area, are increasingly recognized as crucial strategies [45]. This understanding has led to a growing emphasis on regional control programs (RCPs), which involve coordinated efforts among multiple stakeholders within a defined geographical area.
A pioneering example in Japan is the regional eradication initiative in Tahara City, Aichi Prefecture [44]. Faced with high farm density and a farm-level PRRSV prevalence of 98% in 2011, local stakeholders established the "Tahara City PRRS Eradication Committee," comprising producers (led by young farmer representatives), municipal agricultural authorities, and local veterinarians. This committee functions as the central coordinating body, holding regular meetings for information exchange and strategic planning. Key activities include disseminating PRRS information to producers, collecting farm-level data on hygiene and control measures via questionnaires, and systematically assessing and sharing the PRRS status of participating farms. A crucial tool adopted is the "PRRS Stage Classification System" developed by P-JET (PRRS JAPAN ELIMINATION TEAM) (Figure 2) [46]. This system categorizes farms into five stages (Stage I: Positive-Unstable to Stage V: Negative-Naive/Clean) based on regular serological (ELISA) and virological (PCR) testing results. By applying this classification, the Tahara committee objectively monitors the status of each farm (e.g., as of 2013: 7 farms in Stage I, 2 in Stage II, 12 in Stage III, 1 in Stage V, and 49 undetermined) and utilizes mapping to visualize regional risk, thereby facilitating targeted interventions and progress tracking [44]. Importantly, written consent is obtained from producers for data sharing, ensuring trust and participation. Planned future activities include genetic analysis of regional strains to understand transmission pathways and the implementation of stage-specific interventions (e.g., enhanced biosecurity, revised vaccination strategies for Stage I/II; surveillance for Stage III/IV). While the initiative has strengthened collaboration and awareness, challenges include coordinating diverse opinions, maintaining motivation, and addressing knowledge gaps among stakeholders [44]. International experiences, such as those in the US, highlight the evolution of voluntary regional control programs and provide valuable insights into structuring effective collaborations and addressing associated challenges [47].
A case study from a high-density swine production area in Kagoshima Prefecture [40] further illustrates the challenges and necessity of regional approaches. Detection of identical RFLP patterns among neighboring farms strongly suggested inter-farm transmission, hindering individual farm control efforts. Although an optimized MLV vaccination strategy (two 0.5 doses at 1 and 3 weeks of age) improved piglet mortality and growth on one farm, performance remained suboptimal compared to that of clean herds [40]. This situation prompted the implementation of more drastic regional measures, including partial depopulation of nursery units and off-site gilt acclimatization, aimed at reduce the overall viral load in the area [40].
These Japanese examples underscore the limitations of farm-level control and the potential of coordinated regional strategies. P-JET [46] serves as a national platform facilitating such efforts by promoting science-based guidelines (including the Stage Classification), and supporting regional projects. Moving forward, advancing PRRS control in Japan will require scaling up these regional models and strengthening strategies in several key areas:
1. Advanced Molecular Surveillance: Enhance nationwide surveillance using not only ORF5 sequencing but also whole genome sequence (WGS) to rapidly detect genetic shifts, recombination events [23], virulence markers (e.g., NSP2 deletions) [21,25], antigenic changes, and transmission pathways [48]. The systematic analysis of large-scale sequencing data, as demonstrated in the US, is vital for tracking genetic evolution and identifying emerging strains [49]; dynamic classification systems can aid in monitoring these changes effectively [32]. Continuous monitoring for emerging domestic variants and incursions of foreign strains, such as the NADC34-like virus [31], is critical.
2. Strengthened Foreign Strain Risk Management: Maintain up-to-date knowledge of global PRRSV epidemiology, especially concerning Lineage 1 variants and HP-PRRSV. Bolster border biosecurity and quarantine measures. Improve early detection surveillance for foreign strains and refine contingency plans for rapid containment and eradication should incursions occur.
3. Next-Generation Vaccine Development and Evaluation: Accelerate research and development of novel vaccines (e.g., improved MLVs, nanoparticle-adjuvanted inactivated vaccines, vector vaccines, subunit vaccines, mRNA vaccines) [8,9,38] that overcome the limitations of current MLVs by offering better safety profiles and broader cross-protection against diverse strains. Promising results have been observed with novel platforms such as ferritin nanoparticle-based vaccines, which have demonstrated the induction of protective immune responses [39]. Establish robust systems for evaluating the efficacy of new vaccines against relevant domestic circulating strains.
4. Promotion and Support of Regional Control Programs (RCPs): Expand successful RCP models, such as Tahara's [44], nationwide, adapting them to local conditions. Robust RCP establishment and sustainability recommend a dual support approach: strengthening governmental financial and technical aid (e.g., for testing, culling, initial setup, emergency response), potentially via frameworks like the "Regional Chronic Disease Eradication Support Measures", complemented by producer-led funding initiatives such as check-off systems. This dual approach is expected to bolster industry ownership and RCP resilience. Load-Close-Homogenize (LCH) strategies combined with vaccination also warrant consideration.
5. Data-Driven Disease Control: Develop and implement integrated data management systems that combine genomic, farm management, host genetic [10], and production data. This will enable real-time risk assessment, outbreak prediction, and evaluation of control measure effectiveness.
6. Continued Stakeholder Collaboration: Foster and maintain strong collaborative networks involving producers, veterinarians, government agencies, researchers, and industry partners (e.g., through P-JET [46]). Such networks are essential for continuous information sharing, technology transfer, capacity building, and maintaining high biosecurity awareness.
9. Conclusions
This review has outlined the epidemiological trajectory of Porcine Reproductive and Respiratory Syndrome (PRRS) in Japan over approximately three decades, since the first domestic isolation of PRRSV in 1993 [16,17]. It has traced the dynamic evolution of the virus, from the establishment of NA-type virus [16,18] and a unique endemic lineage (Cluster III) [18,25]. This evolution continued through a subsequent increase in genetic diversity (manifested by the rise of Clusters II and IV, and the decline of Cluster III) [19,22], the incursion of the European type (Type 1) [20] and virulent NA-type lineages (Cluster IV/L1F) [19,21], and culminated in the most recent emergence of a significant foreign lineage (NADC34-like/L1A) [31]. Furthermore, this review has examined the tangible impact of PRRS on production performance [28], its substantial economic burden (estimated at JPY 28 billion annually) [5], the challenges associated with widely used MLV vaccines (e.g., limited efficacy, safety concerns) [8,9,23,26], the complexities introduced by host genetic factors [10] and co-infections [12], and the growing importance of regional control initiatives [40,44] coupled with robust biosecurity measures [41,42].
PRRSV presents a formidable challenge owing to its high mutation rate [7], propensity for recombination [23], and sophisticated immune evasion strategies [11], rendering its control exceptionally difficult. The genetic pool of PRRSV in Japan has become increasingly complex, shaped by domestic evolution [18,19], vaccine-induced selection pressure [22], and incursions from abroad [20,21,31]. However, mitigating the impact of PRRS and progressing towards improved control is achievable through a multifaceted approach. This approach should combine advanced molecular epidemiology (e.g., phylogenetics, WGS) for understanding circulating strains, objective diagnostic methods [43], appropriate farm-specific strategies that integrate vaccination [40] (considering limitations against emerging strains [35,36]) and stringent biosecurity [41,42], and, crucially, coordinated regional efforts [40,44]. The recognition that farm-level interventions alone are insufficient, particularly in high-density areas, has spurred initiatives such as those in Aichi [44] and Kagoshima [40] prefectures. These initiatives demonstrate the potential of collaborative, science-based regional programs (e.g., utilizing Stage Classification [46]). Although eradication remains a long-term aspiration, effective PRRS control is an urgent national priority for the sustainability of Japan's swine industry. Continued progress necessitates leveraging the latest scientific knowledge, as outlined in this review, alongside practical field applications driven by strong partnerships (including platforms such as P-JET [46]). Strengthening surveillance, developing next-generation vaccines [38], enhancing biosecurity practices, exploring host genetic resistance [10], and fostering robust collaboration among all stakeholders are essential steps to effectively combat this persistent and costly disease.
Author Contributions
O.T. was responsible for the conceptualization (ideation), literature search, analysis (synthesis of information), drafting, and revision of this review article. A.K., N.T., and K.S. contributed to the discussion and reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work did not receive support from any organization except the regular institutional budget.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable since this study does not include any works with human nor animal samples that require permission to use.
Data Availability Statement
The datasets analyzed during the current study are available from the corresponding author upon reasonable request.
Conflicts of Interest
All authors declare no conflict of interest. However, N.T. is an employee of Nisseiken Co., Ltd.
References
- Snijder, E.J.; Kikkert, M.; Fang, Y. Arterivirus molecular biology and pathogenesis. J. Gen. Virol. 2013, 94, 2141–2163. [CrossRef]
- Rossow, K.D. Porcine Reproductive and Respiratory Syndrome. Vet. Pathol. 1998, 35, 1–20.
- World Organisation for Animal Health (WOAH). Infection with porcine reproductive and respiratory syndrome virus. In Terrestrial Animal Health Code. Available online: https://www.google.com/search?q=https://www.woah.org/en/what-we-do/standards/codes-and-manuals/terrestrial-code-online-access/%3Fid%3D169%26L%3D1%26htmfile%3Dchapitre_prrs.htm (accessed on 22 April 2025).
- Holtkamp, D.J.; Kliebenstein, J.B.; Neumann, E.J.; Zimmerman, J.J.; Rotto, H.F.; Yoder, T.K.; Wang, C.; Yeske, P.E.; Mowrer, C.L.; Haley, C.A. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J. Swine Health Prod. 2013, 21, 72–84. [CrossRef]
- Yamane, I.; Kure, K.; Ishikawa, H.; Takagi, M.; Miyazaki, A.; Suzuki, T.; et al. Evaluation of the economical losses due to the outbreaks of porcine reproductive and respiratory syndrome. (A questionnaire-based epidemiological survey and estimation of the total economical losses in Japan). Proc. Jpn. Pig Vet. Soc.2009, 55, 33–37. (In Japanese).
- Yamane, I.; Kure, K.; Ishikawa, H.; Takagi, M.; Yoshii, M.; Okinaga, T.; et al. Evaluation of the economical losses due to the outbreaks of porcine reproductive and respiratory syndrome. (Results from 6 targeted farms being investigated). Proc. Jpn. Pig Vet. Soc. 2009, 54, 8–13. (In Japanese).
- Hanada, K.; Suzuki, Y.; Nakane, T.; Hirose, O.; Gojobori, T. The origin and evolution of porcine reproductive and respiratory syndrome viruses. Mol. Biol. Evol. 2005, 22, 1024–1031. [CrossRef]
- Kimman, T.G.; Cornelissen, L.A.; Moormann, R.J.; Rebel, J.M.; Stockhofe-Zurwieden, N. Challenges for porcine reproductive and respiratory syndrome virus (PRRSV) vaccinology. Vaccine 2009, 27, 3704–3718. [CrossRef]
- Charerntantanakul, W. Porcine reproductive and respiratory syndrome virus vaccines: Immunogenicity, efficacy and safety aspects. World J. Virol. 2012, 1, 23–30. [CrossRef]
- Peng, N.; Zhou, P.; Pei, Y.; Lin, C.; Li, H.; Feng, Z.; Liu, X.; Chen, Y.; Wu, Z.; Ma, J.; Huang, L. Genetic background influences pig responses to porcine reproductive and respiratory syndrome virus. Front. Vet. Sci. 2023, 10, 1289570.
- Mateu, E.; Diaz, I. The challenge of PRRS immunology. Vet. J. 2008, 177, 345–351. [CrossRef]
- Opriessnig, T.; Giménez-Lirola, L.G.; Halbur, P.G. PRRSV: Interaction with other respiratory pathogens. Pig333, 29 July 2013. Available online: https://www.pig333.com/articles/prrsv-interaction-with-other-respiratory-pathogens_7443/ (accessed on 23 April 2025).
- Fiers, J.; Cay, A.B.; Maes, D.; Tignon, M. A Comprehensive Review on Porcine Reproductive and Respiratory Syndrome Virus with Emphasis on Immunity. Vaccines (Basel) 2024, 12, 935. [CrossRef]
- Wang, H.; Cao, D.; Zhang, T.; Tang, Y.-D.; Liu, W.-M.; Cao, Y.-C.; Wei, J.-C.; Zhang, L.-C.; Zhang, G.-H. PRRSV evades innate immune cGAS-STING antiviral function via its Nsp5 to deter STING translocation and activation. bioRxiv 2025, 2025.02.24.639812 (Preprint).
- Ito, H.; Ouchi, S.; Kato, K.; Komoda, M.; Higuchi, A.; Yamada, T.; et al. The first outbreak of predicted PRRS in Japan. (1) The case in Gunma Prefecture. Proc. Jpn. Pig Vet. Soc. 1995, 26, 6–9. (In Japanese).
- Murakami, Y.; Kato, A.; Tsuda, T.; Morozumi, T.; Miura, Y.; Sugimura, T. Isolation and Serological Characterization of Porcine Reproductive and Respiratory Syndrome (PRRS) Viruses from Pigs with Reproductive and Respiratory Disorders in Japan. J. Vet. Med. Sci. 1994, 56, 891–894. [CrossRef]
- Shimizu, M.; Yamada, S.; Murakami, Y.; Tsuda, T.; Morozumi, T.; Kokuho, T.; et al. Isolation of porcine reproductive and respiratory syndrome (PRRS) virus from Heko-Heko disease of pigs. J. Vet. Med. Sci.1994, 56, 389–391. [CrossRef]
- Yoshii, M.; Kaku, Y.; Murakami, Y.; Shimizu, M.; Kato, K.; Ikeda, H. Genetic variation and geographic distribution of porcine reproductive and respiratory syndrome virus in Japan. Arch. Virol. 2005, 150, 2313–2324. [CrossRef]
- Iseki, H.; Takagi, M.; Miyazaki, A.; Katsuda, K.; Mikami, O.; Tsunemitsu, H. Genetic analysis of ORF5 in porcine reproductive and respiratory syndrome virus in Japan. Microbiol. Immunol. 2011, 55, 211–216. [CrossRef]
- Iseki, H.; Takagi, M.; Kawashima, K.; Shibahara, T.; Kuroda, Y.; Tsunemitsu, H. Type 1 porcine reproductive and respiratory syndrome virus emerged in Japan. In Proceedings of the 22nd International Pig Veterinary Society Congress, Jeju, Korea, 10–13 June 2012; p. 978.
- Iseki, H.; Morozumi, T.; Takagi, M.; Kawashima, K.; Shibahara, T.; Uenishi, H.; et al. Genomic sequence and virulence evaluation of MN184A-like porcine reproductive and respiratory syndrome virus in Japan. Microbiol. Immunol. 2016, 60, 824–834. [CrossRef]
- Kyutoku, F.; Yokoyama, T.; Sugiura, K. Genetic Diversity and Epidemic Types of Porcine Reproductive and Respiratory Syndrome (PRRS) Virus in Japan from 2018 to 2020. Epidemiologia (Basel) 2022, 3, 285–296. [CrossRef]
- Trevisan, G.; Magstadt, D.; Woods, A.; Sparks, J.; Zeller, M.; Li, G.; Krueger, K.M.; Saxena, A.; Zhang, J.; Gauger, P.C.; Linhares, D.C.L. A recombinant porcine reproductive and respiratory syndrome virus type 2 field strain derived from two PRRSV-2-modified live virus vaccines. Front. Vet. Sci. 2023, 10, 1149293. [CrossRef]
- Ruedas-Torres, I.; Rodríguez-Gómez, I.M.; Sánchez-Carvajal, J.M.; Larenas-Muñoz, F.; Gómez-Laguna, J.; Pallarés, F.J.; Carrasco, L.; Oleksiewicz, M.B.; Risalde, M.Á.; Botti, S.; Fraile, L.; Barasona, J.A.; Mínguez, O.; Sánchez-Vizcaíno, J.M.; Mateu, E.; Martín-Acebes, M.A. The jigsaw of PRRSV virulence. Vet. Microbiol. 2021, 260, 109168.
- Yoshii, M.; Okinaga, T.; Miyazaki, A.; Kato, K.; Ikeda, H.; Tsunemitsu, H. Genetic polymorphism of the nsp2 gene in North American type-Porcine reproductive and respiratory syndrome virus. Arch. Virol. 2008, 153, 1173–1178. [CrossRef]
- Iseki, H.; Kawashima, K.; Takagi, M.; Shibahara, T.; Mase, M. Studies on heterologous protection between Japanese type 1 and type 2 porcine reproductive and respiratory syndrome virus isolates. J. Vet. Med. Sci. 2020, 82, 935–942. [CrossRef]
- Iseki, H.; Takagi, M.; Kawashima, K.; Shibahara, T.; Kuroda, Y.; Tsunemitsu, H.; et al. Pathogenicity of emerging Japanese type 1 porcine reproductive and respiratory syndrome virus in experimentally infected pigs. J. Vet. Med. Sci. 2015, 77, 1507–1510. [CrossRef]
- Ishizeki, S.; Ishikawa, H.; Adachi, Y.; Yamazaki, H.; Yamane, I. Study on association of productivity and farm level status of porcine reproductive and respiratory syndrome in pig farms in Japan. Nihon Chikusan Gakkaiho 2014, 85, 171–177. (In Japanese). [CrossRef]
- Yamane, I. Evaluation of the economical losses due to the porcine reproductive and respiratory syndrome. Proc. Jpn. Pig Vet. Soc. 2013, 61, 1–4. (In Japanese).
- Yim-im, W.; Anderson, T.K.; Paploski, I.A.D.; VanderWaal, K.; Gauger, P.; Krueger, K.; Shi, M.; Main, R.; Zhang, J.Q.; Chao, D.Y. Refining PRRSV-2 genetic classification based on global ORF5 sequences and investigation of their geographic distributions and temporal changes. Microbiol. Spectr. 2023, 11, e02916-23.
- Yonezawa, Y.; Taira, O.; Omori, T.; Tsutsumi, N.; Sugiura, K. First detection of NADC34-like porcine reproductive and respiratory syndrome virus strains in Japan. J. Vet. Med. Sci. 2025, 87, 110–114. [CrossRef]
- VanderWaal, K.; Pamornchainavakul, N.; Kikuti, M.; Zhang, J.; Zeller, M.; Trevisan, G.; Rossow, S.; Schwartz, M.; Linhares, D.C.L.; Holtkamp, D.J.; Herrera da Silva, J.P.; Corzo, C.A.; Baker, J.P.; Anderson, T.K.; Makau, D.N.; Paploski, I.A.D. PRRSV-2 variant classification: a dynamic nomenclature for enhanced monitoring and surveillance. mSphere 2024, 9, e00709-24. [CrossRef]
- Mei, Y.; Chen, J.; Chen, Y.; Huang, C.; Chen, X.; Zhang, Z.; Li, Y.; Zhou, L.; Wu, J.; Liu, X.; Zhang, L.; He, Q.; Chen, H.; Qian, P. Porcine Reproductive and Respiratory Syndrome Virus Prevalence and Pathogenicity of One NADC34-like Virus Isolate Circulating in China. Microorganisms 2025, 13, 796.
- Wesley, R.D.; Mengeling, W.L.; Lager, K.M.; Clouser, D.F.; Landgraf, J.G.; Frey, M.L. Differentiation of a Porcine Reproductive and Respiratory Syndrome Virus Vaccine Strain from North American Field Strains by Restriction Fragment Length Polymorphism Analysis of ORF 5. J. Vet. Diagn. Invest. 1998, 10, 140–144. [CrossRef]
- Zhang, R.; Zhu, L.; Li, Y.; Yan, Y.; Wang, Z.; Wang, X.; Zhang, J.; Ji, H.; Zhang, Y.; Zhou, E.-M.; Chen, H.-B. Protective evaluation of the commercialized porcine reproductive and respiratory syndrome virus vaccines in piglets challenged by NADC34-like strain. Front. Microbiol. 2024, 15, 1422335.
- Luo, Y.; Xu, J.; Xia, W.; Hou, G.; Wang, Z.; Wu, Y.; Cui, L.; Wang, X.; Xiao, Y.; Zhang, G.; Li, J.; Liu, Y. Efficacy of a reduced-dosage PRRS MLV vaccine against a NADC34-like strain of porcine reproductive and respiratory syndrome virus. Front. Vet. Sci. 2024, 11, 1493384.
- Iseki, H.; Kawashima, K.; Shibahara, T.; Mase, M. Immunity against a Japanese local strain of porcine reproductive and respiratory syndrome virus decreases viremia and symptoms of a highly pathogenic strain. BMC Vet. Res. 2021, 17, 156. [CrossRef]
- Li, J.; Miller, L.C.; Sang, Y. Current Status of Vaccines for Porcine Reproductive and Respiratory Syndrome: Interferon Response, Immunological Overview, and Future Prospects. Vaccines (Basel) 2024, 12, 606. [CrossRef]
- Sun, Y., Gao, Y., Su, T., Zhang, L., Zhou, H., Zhang, J., Sun, H., Bai, J., & Jiang, P. Nanoparticle Vaccine Triggers Interferon-Gamma Production and Confers Protective Immunity against Porcine Reproductive and Respiratory Syndrome Virus. ACS Nano. 2025, 9 (1), 852-870. [CrossRef]
- Kawabata, T. Control measures against porcine reproductive and respiratory syndrome in a high-density swine production area. Proc. Jpn. Pig Vet. Soc. 2014, 63, 22–26. (In Japanese).
- Dee, S.; Brands, L.; Edler, R.; Schelkopf, A.; Nerem, J.; Spronk, G.; Kikuti, M.; Corzo, C.A. Further Evidence That Science-Based Biosecurity Provides Sustainable Prevention of Porcine Reproductive and Respiratory Syndrome Virus Infection and Improved Productivity in Swine Breeding Herds. Animals (Basel) 2024, 14, 2530. [CrossRef]
- Niederwerder, M.C.; Ressenig, K.R.; Gebhardt, J.T.; Bryer, C.L.; Pouggouras, K.C.; Woodworth, J.C.; DeRouchey, J.M.; Dritz, S.S.; Goodband, R.D. An assessment of enhanced biosecurity interventions and their impact on porcine reproductive and respiratory syndrome virus outbreaks within a managed group of farrow-to-wean farms, 2020–2021. J. Am. Vet. Med. Assoc. 2023, 261 (S1), S57–S66.
- Fukunaga, W.; Hayakawa-Sugaya, Y.; Koike, F.; Diep, N.V.; Kojima, I.; Yoshida, Y.; et al. Newly-designed primer pairs for the detection of type 2 porcine reproductive and respiratory syndrome virus genes. J. Virol. Methods 2021, 291, 114071. [CrossRef]
- Mizukami, Y. Construction of regional network for the PRRS free project. Proc. Jpn. Pig Vet. Soc. 2013, 61, 11–13. (In Japanese).
- Dee, S. How to control and eliminate PRRS from swine herds on farm and regional level. CAB Rev. 2012, 7, 1–9.
- Farm Stage Definitions for Promoting Eradication of Porcine Reproductive and Respiratory Syndrome (PRRS). Available online: https://site-pjet.com/wp-content/uploads/2017/07/document2.pdf (accessed on 22 April 2025). (In Japanese).
- Magalhães, E.S.; Zimmerman, J.J.; Holtkamp, D.J.; Classen, D.M.; Groth, D.D.; Glowzenski, L.; Philips, R.; Silva, G.S.; Linhares, D.C.L. Next Generation of Voluntary PRRS Virus Regional Control Programs. Front. Vet. Sci. 2021, 8, 769312. [CrossRef]
- Baker, J.P.; Rovira, A.; VanderWaal, K. Repeat offenders: PRRSV-2 clinical re-breaks from a whole genome perspective. Vet. Microbiol. 2025, 302, 110411. [CrossRef]
- Chandra, A.M.; Linhares, D.C.L.; Trevisan, G.; Crim, B.; Burrough, E.; Gauger, P.; Madson, D.; Thomas, J.; Zeller, M.; Zhang, J.; Main, R.; Rovira, A.; Thurn, M.; Lages, P.; Corzo, C.A.; Sturos, M.; VanderWaal, K.; Naikare, H.; Matias-Ferreyra, F.; McGaughey, R.; Retallick, J.; McReynolds, S.; Arruda, L.C.S.P.; Schwartz, M.; Yeske, P.; Murray, D.; Mason, B.; Schneider, P.; Copeland, S.; Dufresne, L.; Boykin, D.; Fruge, C.; Hollis, W.; Robbins, R.; Petznick, T.; Kuecker, K.; Glowzenski, L.; Niederwerder, M.; Huang, X. Harnessing sequencing data for porcine reproductive and respiratory syndrome virus (PRRSV): tracking genetic evolution dynamics and emerging sequences in US swine industry. Front. Vet. Sci. 2025, Mar 6:12:1571020. [CrossRef]
- Zuckermann, F.A.; Garcia, E.A.; Luque, I.D.; Christopher-Hennings, J.; Doster, A.R.; Murtaugh, M.P.; Nelson, E.A.; Schwartz, K.-J.; Torrison, J.L.; Wasilk, A.M. Assessment of the efficacy of commercial porcine reproductive and respiratory syndrome virus (PRRSV) vaccines based on evaluation of virologic protection following challenge with a virulent strain of type 2 PRRSV. Vaccine 2007, 25, 5098–5108.
Figure 1.
Genetic evolution of PRRSV in Japan (1993-2024). Timeline illustrating the genetic evolution of PRRSV in Japan since 1993, highlighting the emergence and shifts of major lineages and key incursions of novel strains.
Figure 1.
Genetic evolution of PRRSV in Japan (1993-2024). Timeline illustrating the genetic evolution of PRRSV in Japan since 1993, highlighting the emergence and shifts of major lineages and key incursions of novel strains.
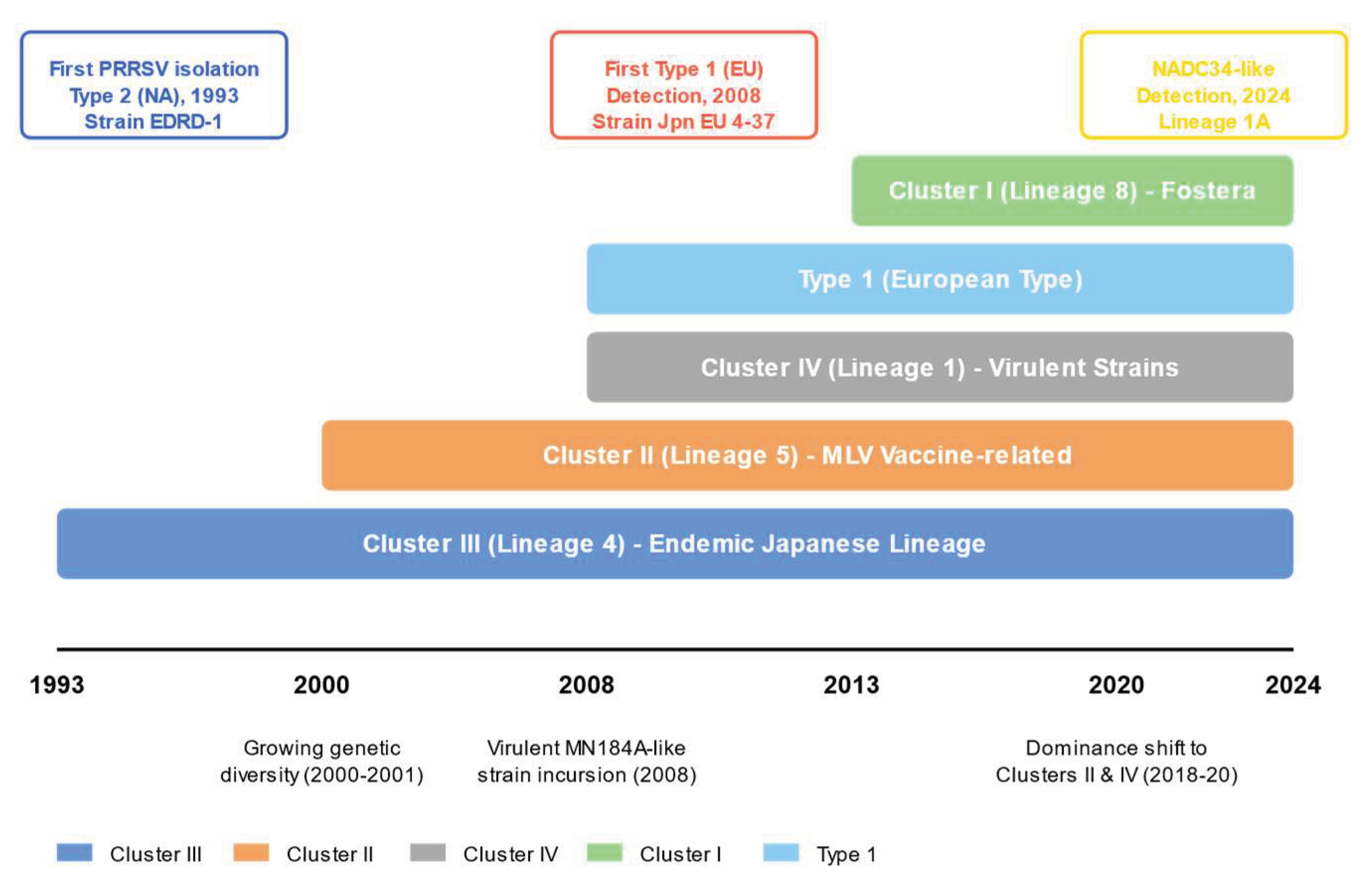
Figure 2.
PRRS stage classification system for regional control programs. A five-stage PRRS herd classification system based on diagnostic status is used to guide interventions in Japanese regional control programs.
Figure 2.
PRRS stage classification system for regional control programs. A five-stage PRRS herd classification system based on diagnostic status is used to guide interventions in Japanese regional control programs.
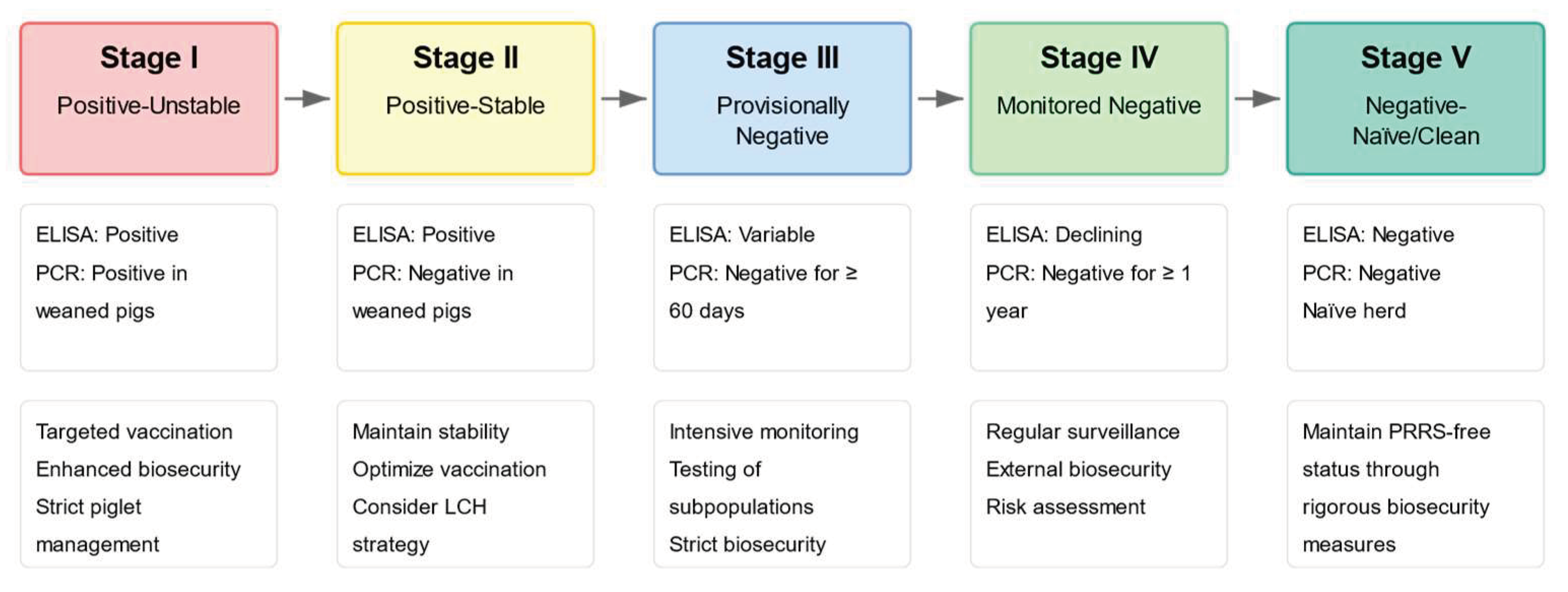

Table 1.
PRRS vaccines currently approved and used in Japan.
| Vaccine Name | Type | Lineage | Manufacturer |
|---|---|---|---|
| Ingelvac® PRRS MLV | MLV (Type 2) | Lineage 5 | Boehringer Ingelheim |
| Fostera® PRRS | MLV (Type 2) | Lineage 8 | Zoetis |
| Nisseiken PRRS Vaccine ME | KV (Type 2) | Lineage 4 | Nisseiken Co., Ltd. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



