Submitted:
09 May 2025
Posted:
12 May 2025
You are already at the latest version
Abstract
Male infertility is a multifactorial condition often associated with disruptions in sperm metabolism and mitochondrial function, yet traditional semen analysis provides limited insight into these molecular mechanisms. Understanding sperm bioenergetics and metabolic dysfunctions is crucial for improving diagnosis and treatment of conditions such as asthenozoospermia and azoospermia.Methods: This systematic review synthesizes recent literature, focusing on novel tools and techniques—including omics technologies, advanced imaging, spectroscopy, and functional assays—that enable comprehensive molecular assessment of sperm metabolism and development.Results: The reviewed studies highlight the effectiveness of metabolomics, proteomics, and transcriptomics in identifying metabolic biomarkers linked to male infertility. Non-invasive imaging modalities such as Raman and magnetic resonance spectroscopy offer real-time metabolic profiling, while the seminal microbiome is increasingly recognized for its role in modulating sperm metabolic health. Despite these advances, challenges remain in clinical validation and implementation of these techniques in routine infertility diagnostics.Conclusions: Integrating novel molecular metabolic assessments with conventional semen analysis promises enhanced diagnostic precision and personalized therapeutic approaches, ultimately improving reproductive outcomes. Continued research is needed to standardize biomarkers and validate clinical utility. Furthermore, these metabolic tools hold significant potential to elucidate the underlying causes of previously misunderstood and unexplained infertility cases, offering new avenues for diagnosis and treatment.
Keywords:
sperm metabolism
; mitochondrial dysfunction
; male infertility
; metabolomics
; proteomics
; molecular biomarkers
; seminal microbiome
; sperm bioenergetics
1. Introduction
Human spermatozoa are highly specialized cells responsible for delivering the paternal genome to the oocyte, a process that requires substantial energy primarily in the form of adenosine triphosphate (ATP) to maintain sperm integrity and motility [1]. Motility alone consumes the majority of sperm ATP, which is generated mainly through glycolysis and oxidative phosphorylation (OXPHOS) pathways localized in distinct sperm compartments [2,3]. These metabolic pathways are dynamically regulated to meet the energy demands during sperm development, maturation, capacitation, and transit through the male and female reproductive tracts [4]. Despite its critical role, sperm metabolism and bioenergetics remain insufficiently explored, limiting advances in diagnosing and treating male infertility conditions such as asthenozoospermia and azoospermia [5].
Infertility affects approximately 8–12% of couples worldwide, with male factors contributing to nearly half of these cases [6,7]. Among male infertility disorders, sperm motility defects have been increasingly linked to mitochondrial dysfunction and impaired energy metabolism [8]. Emerging evidence also implicates the seminal and sperm-associated microbiome in modulating sperm DNA integrity and metabolism, suggesting novel therapeutic targets [9]. The advent of omics technologies—including genomics, proteomics, and metabolomics—combined with advanced analytical tools, now enables comprehensive profiling of sperm metabolic pathways and identification of molecular biomarkers associated with fertility outcomes [10,11,12].
This systematic review synthesizes recent advances in novel tools and techniques for evaluating human sperm metabolism and development. It emphasizes methods that uncover molecular mechanisms underlying infertility and improve prediction of sperm retrieval success, particularly in azoospermia and metabolic impairments. Highlighting the central role of mitochondria and oxidative stress in idiopathic infertility, this work aims to guide future research and clinical strategies to enhance male fertility diagnostics and therapeutics.
2. Methods
2.1. Sources
Data were independently extracted by the authors. A systematic search was conducted through PubMed, Scopus, and Web of Science databases, covering records published from each database inception to 15 February 2025. The search strategy combined Medical Subject Headings (MeSH) terms and keywords tailored to capture relevant literature on sperm metabolism and development.
Specifically, key terms included: “sperm metabolism”, “sperm bioenergetics”, “glycolysis”, “oxidative phosphorylation”, “mitochondrial function”, “mitochondrial membrane potential”, “sperm motility”, “asthenozoospermia”, “azoospermia”, “male infertility”, “metabolomics”, “proteomics”, “transcriptomics””, antioxidants”, “sperm capacitation”, and “microbiome”.
Additional manual searches were performed by screening the reference lists of pertinent articles to identify further relevant studies not captured by database queries.
2.2. Study Selection
Studies that met the following inclusion criteria were included in the qualitative synthesis to focus on evaluating novel tools and techniques for assessing sperm metabolism and development in relation to male infertility:
- Design: Original research articles including observational studies, clinical trials, case reports, and technical method evaluations; reviews and small cohort studies were excluded.
- Patient inclusion criteria: Human males with infertility of various etiologies, including idiopathic infertility and metabolic-related dysfunctions affecting sperm quality.
- Patient exclusion criteria: Review articles, studies involving animal models; male infertility with well-established causes such as varicocele, cryptorchidism, infections of male accessory glands, genetic abnormalities (e.g., Y chromosome microdeletions), testicular torsion or trauma, and systemic diseases including thyroid, pituitary, adrenal disorders, kidney or liver failure.
- Study intervention: Application or evaluation of diagnostic or research tools and techniques aimed at identifying biomarkers or characterizing sperm metabolic pathways and developmental processes.
- Study outcome: Assessment of the utility, accuracy, and relevance of advanced methods and emerging technologies for evaluating sperm metabolism and their usefulness in diagnosing or understanding metabolic sperm-related infertility.
3. Results
Using the described search strategy, 2,775 records were initially identified. After removing duplicates, 2,433 records remained. Screening titles and abstracts led to the exclusion of 977 records as not pertinent, leaving 1,456 articles for further assessment. Of these, 1,150 papers were excluded with reasons during detailed screening. A total of 306 full-text articles were assessed for eligibility, with 263 excluded for not satisfying the inclusion criteria. Finally, 43 studies met the criteria and were included in this systematic review (Figure 1).
This review synthesizes the most relevant and recent tools and techniques used to evaluate human sperm metabolism and development, focusing on their application in male infertility diagnostics and research. The included studies provide a comprehensive overview of methodologies assessing metabolic pathways, mitochondrial function, and molecular biomarkers associated with various infertility causes.
Before discussing the findings on these novel techniques and their clinical relevance, subsequent sections summarize current knowledge on sperm metabolism and its critical role in male fertility, providing the foundation for future research and clinical applications.
4. Energy Metabolism Pathways in Spermatozoa
Energy metabolism in spermatozoa depends on a series of tightly regulated biochemical reactions culminating in the production of adenosine triphosphate (ATP), the cell’s primary energy currency [13]. Two principal pathways generate ATP: glycolysis and oxidative phosphorylation (OXPHOS). Glycolysis occurs mainly in the sperm’s principal piece and head, providing a rapid but limited ATP supply without requiring oxygen [2] . This pathway yields only two net ATP molecules per glucose molecule.
OXPHOS, in contrast, takes place in the mitochondria densely packed within the sperm midpiece. This oxygen-dependent pathway is more efficient and produces the bulk of ATP needed to meet the high energy demands of sperm motility [3]. ATP generated in the midpiece must be transported along the flagellum to power movement. While diffusion has been proposed as a mechanism, its efficiency is limited by the narrow space within the flagellum, suggesting specialized transport systems might be involved, though these remain to be fully elucidated in humans and other mammals [14,15].
5. Energy Metabolism Pathways in Spermatozoa
5.1. Metabolic Substrates and Transport
Sperm utilize both endogenous and exogenous carbohydrates to satisfy their metabolic needs. Seminal plasma from various mammals, including humans, contains fructose as the main metabolic substrate [16,17], along with glucose and, in some species, mannose [18]. These sugars are internalized through glucose transporters (GLUTs) with varying substrate specificities [19,20]. Additionally, other sugars such as sorbitol serve as energy substrates in species including humans, rams, and mice [21,22].
Mitochondrial substrates include lactate and pyruvate, metabolized via lactate dehydrogenase (LDH) to regenerate NAD+ or enter the Krebs cycle for OXPHOS [23]. Pyruvate and lactate may originate from glycolysis or be taken up externally via monocarboxylate transporters [24,25]. Human sperm express specific LDH isoforms that enhance mitochondrial utilization of lactate, permitting efficient respiratory substrate use [26,27]. Fatty acid β-oxidation also contributes to ATP production. Boar and bull sperm metabolize long-chain fatty acids such as oleic, palmitic, and myristic acids via mitochondrial β-oxidation [28,29]. Human sperm possess enzymatic machinery for both saturated and unsaturated fatty acid β-oxidation; inhibition of this pathway impairs motility [30]. Glycogen reserves, once thought absent in mature sperm, have been found in some species such as dogs, where sperm can synthesize and utilize glycogen as an energy source [15,31,32].
5.2. Metabolic Adaptations During Maturation and Transit
Many studies use epididymal sperm to assess basal metabolic activity; however, epididymal sperm lack exposure to seminal plasma, which provides a fluid rich in energetic substrates and metabolic modulators [33]. Differences in metabolic activity between epididymal and ejaculated sperm have been observed. For example, ejaculated bull sperm rely heavily on OXPHOS for motility, whereas epididymal sperm maintain ATP levels primarily through glycolysis [33,34,35]. In mice, epididymal sperm depend mainly on glucose, but ejaculated sperm utilize a broader range of substrates, including citrate and pyruvate, through nonglycolytic pathways [36].
Sperm acquire fertilizing capacity through capacitation, a process involving extensive biochemical and physiological changes such as plasma membrane remodeling, ion fluxes, and activation of signaling cascades including cAMP-dependent pathways. Metabolic activity increases substantially during capacitation, reflecting heightened energy demands [37]. During transit through the female reproductive tract, sperm encounter variable environments with differing oxygen and substrate availability, necessitating metabolic adaptability to maintain motility and viability until fertilization [36,38,39].
6. Molecular Biomarkers and Oxidative Stress
Disruptions in sperm metabolism are frequently characterized by mitochondrial dysfunction and heightened oxidative stress, both of which play pivotal roles in the pathophysiology of male infertility. Mitochondria, being the powerhouse of sperm cells, are essential for ATP production necessary for motility and overall sperm function. However, excessive generation of reactive oxygen species (ROS) — a byproduct of impaired mitochondrial electron transport chains — leads to oxidative damage targeting mitochondrial membranes, proteins, and mitochondrial DNA [40]. This oxidative insult not only diminishes sperm motility but also compromises the integrity of nuclear DNA, increasing the risk of mutations and fragmentation that can adversely affect fertilization and embryo development [4].
Advances in high-throughput omics technologies, including proteomics, metabolomics, and transcriptomics, have enabled the identification of specific molecular biomarkers that serve as indicators of these metabolic disturbances. Proteomic analyses have revealed aberrant expression of key mitochondrial proteins and antioxidant enzymes, while metabolomic profiling has uncovered altered concentrations of metabolites involved in energy metabolism and redox balance. Transcriptomic studies further highlight dysregulation in gene expression pathways related to oxidative stress response and mitochondrial function [11]. Together, these molecular signatures provide valuable insight into the biochemical underpinnings of sperm dysfunction and hold promise for developing diagnostic tools and targeted therapies aimed at mitigating oxidative damage in infertile men.
7. Influence of the Seminal Microbiome
Recent research has begun to elucidate the significant role of the seminal microbiome—the diverse microbial communities residing within seminal fluid and associated with spermatozoa—in modulating sperm metabolic health and fertility outcomes. These microbial populations can influence the biochemical milieu of the seminal plasma, impacting factors such as pH, nutrient availability, and oxidative stress levels. Certain bacterial species have been implicated in either protective or detrimental effects on sperm function, with some microbes producing metabolites that exacerbate oxidative stress or trigger inflammatory responses, thereby compromising sperm DNA integrity and metabolic efficiency [9]. The interaction between the seminal microbiome and sperm cells represents a complex and dynamic system that may contribute to idiopathic cases of male infertility. Understanding how microbial dysbiosis alters sperm metabolism and function opens new avenues for therapeutic intervention, such as the use of probiotics, antibiotics, or microbiome modulation strategies to restore a healthy seminal environment. This emerging field holds promise for enhancing fertility treatments by addressing not only the host’s cellular dysfunction but also the microbial factors influencing reproductive health.
8. Advanced Techniques and Tools for Evaluating Sperm Metabolism
The evaluation of sperm metabolism and its role in male infertility has advanced considerably through diverse methodologies encompassing proteomics, metabolomics, transcriptomics, imaging, and functional assays. Agarwal et al. conducted proteomic analyses on seminal plasma from 42 infertile men and 17 fertile donors, identifying a 35-protein pathway linked to sperm dysfunction, including overexpression of membrane metallo-endopeptidase (MME), which was significantly elevated in infertile patients [41]. Similarly, Amaral et al. characterized the sperm tail proteome in normozoospermic men, identifying 1,049 proteins with a prominent presence of lipid metabolism enzymes and active mitochondrial and peroxisomal pathways, highlighting the metabolic complexity essential for sperm motility [8].
Metabolomic studies have revealed significant biochemical alterations in infertile populations, Boguenet et al. applied targeted metabolomics to seminal plasma from severe oligoasthenospermia patients, documenting decreased metabolites associated with sperm membrane deterioration and energy defects [42]. Complementing this, Li et al. identified nine metabolites differing significantly in asthenozoospermic men, with nicotinamide levels correlating positively with sperm count and motility [43]. Zhao et al. detected 33 disturbed metabolites in idiopathic asthenozoospermia, implicating disrupted energy and amino acid metabolism [44], while in another study he reported 72 to 89 metabolites altered in oligospermic patients, including key amino acid and ketone body pathways [44]. Mitochondrial dysfunction emerges as a central theme linking these molecular changes to impaired sperm function. Cassina et al. demonstrated that mitochondrial respiratory deficits and nitro-oxidative protein modifications in sperm midpiece and head regions correlated with reduced motility in their cohort [40]. This functional impairment aligns with fluorescence-based assessments by Paoli et al. who showed a positive correlation between mitochondrial membrane potential—measured by JC-1 staining—and sperm motility, reinforcing mitochondrial health as a fertility marker [45].
Transcriptomic and epigenetic analyses have further enriched biomarker discovery. Larriba et al. profiled small RNAs in seminal extracellular vesicles from azoospermic men, identifying canonical and isomiR microRNAs that effectively discriminate azoospermia origin with high specificity [46]. Li et al. reported 238 differentially methylated regions in asthenozoospermic spermatozoa, affecting 114 genes essential for spermatogenesis and motility, suggesting epigenetic regulation as a contributor to metabolic dysfunction [43]. Imaging and spectroscopic techniques provide complementary functional insights. Raman microspectroscopy has been utilized to detect sperm DNA damage, enabling sensitive biochemical characterization suitable for live sperm in a study by Costa et al. [47]. Additionally, 13C-magnetic resonance spectroscopy revealed that motile sperm preferentially metabolize glucose, fructose, and pyruvate into lactate, highlighting increased glycolytic activity crucial for motility [48]. These observations align with metabolic profiling studies that differentiated metabolite peaks between sperm populations using 1H magnetic resonance spectroscopy [49].
Oxidative stress remains a critical factor in sperm dysfunction, with Aziz et al. and Irigoyen et al. quantifying reactive oxygen species (ROS) production in sperm cells [50,51]. Their studies demonstrated that increased hydrogen peroxide and peroxynitrite levels inversely correlate with mitochondrial respiratory control ratios and sperm motility, highlighting oxidative damage as a major disruptor of bioenergetics. Proteomic analyses have identified 15 proteins that are differentially expressed in spermatozoa exhibiting elevated reactive oxygen species, especially those related to energy metabolism and antioxidant defense [11]. Emerging research further implicates the seminal microbiome in sperm health; combining 16S rRNA sequencing with untargeted metabolomics revealed an enrichment of Lactobacillus iners in men with a high sperm DNA fragmentation index, associating microbial metabolites such as butanoate fermentation products with DNA damage and metabolic disruptions [9].
The integration of these multi-dimensional data sets has been facilitated by bioinformatics approaches. Milardi et al. and Sharma et al. utilized pathway enrichment and network modeling to identify disrupted metabolic nodes in infertile men, accelerating biomarker discovery and therapeutic targeting [10,11]. Such systems biology approaches are crucial for translating complex molecular insights into clinical practice.
Collectively, these studies underscore that sperm metabolism—and its perturbations involving mitochondrial dysfunction, oxidative stress, epigenetic alterations, and microbial influences—plays a pivotal role in male infertility. The high sensitivity and specificity of these advanced methodologies promise improved diagnostic precision and personalized treatment strategies. Table 1 summarizes the key methods and their contributions, illustrating the strides made toward comprehensive sperm metabolism assessment beyond traditional semen analysis.
9. Clinical Implications: Metabolic Dysfunction in Male Infertility
The reviewed studies underscore the critical role of metabolic dysfunction in prevalent infertility conditions such as asthenozoospermia and azoospermia. Alterations in energy metabolism pathways, mitochondrial dysfunction, and oxidative stress are recurrent themes linked to impaired sperm motility and viability [40,41,58]. In asthenozoospermia, metabolomic and proteomic studies have identified disrupted glycolysis and fatty acid oxidation pathways, correlating with reduced motility [8,43]. Similarly, azoospermic patients exhibit distinct proteomic and microRNA profiles predictive of spermatogenic status and sperm retrieval success [53,56,62]. These biomarkers provide valuable prognostic information, guiding clinical decisions and personalized treatment strategies.
Emerging evidence also highlights the influence of the seminal microbiome on sperm metabolic health and DNA integrity, adding complexity to infertility diagnostics [9]. Despite advances, challenges remain in standardizing assays and integrating multi-omics data into routine clinical practice [65,71].
The integration of advanced metabolic assessment tools into clinical workflows promises improved diagnosis, prognosis, and management of male infertility.
Future research should focus on validating these biomarkers in larger cohorts and developing accessible platforms for widespread clinical application.
10. Conclusions
The relationship between sperm metabolism and male fertility is increasingly recognized, yet its clinical application remains limited. Traditional semen analysis often fails to explain idiopathic infertility, highlighting the need for metabolic profiling and molecular biomarkers to improve diagnostic accuracy. These advanced tools provide deeper insights into mitochondrial function, energy metabolism, and oxidative stress—key factors for sperm quality and fertility. Challenges remain in identifying which patients will benefit most, determining optimal testing timing, and selecting the most informative biomarkers. Additionally, randomized clinical trials are needed to validate these approaches, especially for idiopathic infertility cases. Future research should focus on clinical validation and exploring targeted therapies that enhance sperm function.
Integrating metabolic assessments with conventional analysis promises more personalized treatments and better outcomes. By advancing biomarker-driven diagnostics, we move closer to resolving male infertility’s complexities and improving reproductive success through precision medicine.
Author Contributions
“Conceptualization, A.Z. and I.D.; methodology, A.Z. and I.D..; software,A.Z..; validation, A.Z.,., I.D., A.B. and N.L..; formal analysis, A.Z.; investigation, X.X.; resources, X.X.; data curation, I.D., A.B. and N.L.; writing—original draft preparation, A.Z.; writing—review and editing, A.Z., I.D., A.B. and N.L.;; visualization, A.Z., I.D., A.B. and N.L .; supervision, I.D., A.B. and N.L .; project administration, A.B.; All authors have read and agreed to the published version of the manuscript.”.
Funding
This study was conducted without the support of any external funding sources.
Conflicts of Interest
The authors report no conflicts of interest.
References
- R. Bohnensack and W. Halangk, “Control of respiration and of motility in ejaculated bull spermatozoa,” Biochim Biophys Acta, vol. 850, no. 1, pp. 72–79, Jun. 1986. [CrossRef]
- S. S. Du Plessis, A. Agarwal, J. Halabi, and E. Tvrda, “Contemporary evidence on the physiological role of reactive oxygen species in human sperm function,” J Assist Reprod Genet, vol. 32, no. 4, pp. 509–520, Apr. 2015. [CrossRef]
- P. E. Visconti, “Sperm bioenergetics in a nutshell,” Biol Reprod, vol. 87, no. 3, p. 72, Sep. 2012. [CrossRef]
- R. Sciorio and S. D. Fleming, “Intracytoplasmic sperm injection vs. in-vitro fertilization in couples in whom the male partners had a semen analysis within normal reference ranges: An open debate,” Andrology, vol. 12, no. 1, pp. 20–29, 2024. [CrossRef]
- W. Ombelet et al., “Now is the time to introduce new innovative assisted reproduction methods to implement accessible, affordable, and demonstrably successful advanced infertility services in resource-poor countries,” Human Reproduction Open, vol. 2025, no. 1, p. hoaf001, Jan. 2025. [CrossRef]
- “Infertility.” Accessed: Apr. 17, 2025. [Online]. Available: https://www.who.int/news-room/factsheets/detail/infertility.
- Agarwal, A. Mulgund, A. Hamada, and M. R. Chyatte, “A unique view on male infertility around the globe,” Reprod Biol Endocrinol, vol. 13, p. 37, Apr. 2015. [CrossRef]
- A. Amaral, J. Castillo, J. M. Estanyol, J. L. Ballesca, J. Ramalho-Santos, and R. Oliva, “Human sperm tail proteome suggests new endogenous metabolic pathways,” Molecular and Cellular Proteomics, vol. 12, no. 2, pp. 330–342, 2013. [CrossRef]
- J. He et al., “Association between semen microbiome disorder and sperm DNA damage.,” Microbiol Spectr, vol. 12, no. 8, p. e0075924, Aug. 2024. [CrossRef]
- D. Milardi et al., “Novel Biomarkers of Androgen Deficiency From Seminal Plasma Profiling Using High-Resolution Mass Spectrometry,” JOURNAL OF CLINICAL ENDOCRINOLOGY & METABOLISM, vol. 99, no. 8, pp. 2813–2820, Aug. 2014. [CrossRef]
- R. Sharma et al., “Functional proteomic analysis of seminal plasma proteins in men with various semen parameters,” REPRODUCTIVE BIOLOGY AND ENDOCRINOLOGY, vol. 11, May 2013. [CrossRef]
- R. Sharma et al., “Proteomic analysis of human spermatozoa proteins with oxidative stress,” REPRODUCTIVE BIOLOGY AND ENDOCRINOLOGY, vol. 11, May 2013. [CrossRef]
- Quijano, M. Trujillo, L. Castro, and A. Trostchansky, “Interplay between oxidant species and energy metabolism,” Redox Biology, vol. 8, pp. 28–42, Aug. 2016. [CrossRef]
- R. M. Tombes and B. M. Shapiro, “Metabolite channeling: a phosphorylcreatine shuttle to mediate high energy phosphate transport between sperm mitochondrion and tail,” Cell, vol. 41, no. 1, pp. 325–334, May 1985. [CrossRef]
- W. C. L. Ford, “Glycolysis and sperm motility: does a spoonful of sugar help the flagellum go round?,” Hum Reprod Update, vol. 12, no. 3, pp. 269–274, 2006. [CrossRef]
- T. Mann, “Fructose, a Constituent of Semen,” Nature, vol. 157, no. 3977, pp. 79–79, Jan. 1946. [CrossRef]
- G. Frenette, C. G. Frenette, C. Lessard, E. Madore, M. A. Fortier, and R. Sullivan, “Aldose Reductase and Macrophage Migration Inhibitory Factor Are Associated with Epididymosomes and Spermatozoa in the Bovine Epididymis1,” Biology of Reproduction, vol. 69, no. 5, pp. 1586–1592, Nov. 2003. [CrossRef]
- T. Rigau et al., “Differential effects of glucose and fructose on hexose metabolism in dog spermatozoa,” Apr. 2002. [CrossRef]
- F. Burant, J. F. Burant, J. Takeda, E. Brot-Laroche, G. I. Bell, and N. O. Davidson, “Fructose transporter in human spermatozoa and small intestine is GLUT5.,” Journal of Biological Chemistry, vol. 267, no. 21, pp. 14523–14526, Jul. 1992. [CrossRef]
- Bucci, J. E. Rodriguez-Gil, C. Vallorani, M. Spinaci, G. Galeati, and C. Tamanini, “GLUTs and Mammalian Sperm Metabolism,” Journal of Andrology, vol. 32, no. 4, pp. 348–355, 2011. [CrossRef]
- T. E. King and T. R. R. Mann, “Sorbitol metabolism in spermatozoa,” Proceedings of the Royal Society of London. Series B. Biological Sciences, vol. 151, no. 943, pp. 226–243, Jan. 1997. [CrossRef]
- W. Cao, H. K. W. Cao, H. K. Aghajanian, L. A. Haig-Ladewig, and G. L. Gerton, “Sorbitol Can Fuel Mouse Sperm Motility and Protein Tyrosine Phosphorylation via Sorbitol Dehydrogenase1,” Biology of Reproduction, vol. 80, no. 1, pp. 124–133, Jan. 2009. [CrossRef]
- C. R. Darr, D. D. C. R. Darr, D. D. Varner, S. Teague, G. A. Cortopassi, S. Datta, and S. A. Meyers, “Lactate and Pyruvate Are Major Sources of Energy for Stallion Sperm with Dose Effects on Mitochondrial Function, Motility, and ROS Production1,” Biology of Reproduction, vol. 95, no. 2, pp. 34, 1–11, Aug. 2016. [CrossRef]
- P. Halestrap and N. T. Price, “The proton-linked monocarboxylate transporter (MCT) family: structure, function and regulation,” Biochem J, vol. 343 Pt 2, no. Pt 2, pp. 281–299, Oct. 1999.
- J. Lee, D. R. Lee, and S. Lee, “The genetic variation in Monocarboxylic acid transporter 2 (MCT2) has functional and clinical relevance with male infertility,” Asian Journal of Andrology, vol. 16, no. 5, p. 694, Oct. 2014. [CrossRef]
- A. Blanco and W., H. Zinkham, “Lactate Dehydrogenases in Human Testes,” Science, vol. 139, no. 3555, pp. 601–602, Feb. 1963. [CrossRef]
- Swegen, B. J. Curry, Z. Gibb, S. R. Lambourne, N. D. Smith, and R. J. Aitken, “Investigation of the stallion sperm proteome by mass spectrometry,” Mar. 2015. [CrossRef]
- Z. Zhu et al., “Exogenous Oleic Acid and Palmitic Acid Improve Boar Sperm Motility via Enhancing Mitochondrial Β-Oxidation for ATP Generation,” Animals, vol. 10, no. 4, Art. no. 4, Apr. 2020. [CrossRef]
- Md. M. Islam, T. Umehara, N. Tsujita, and M. Shimada, “Saturated fatty acids accelerate linear motility through mitochondrial ATP production in bull sperm,” Reproductive Medicine and Biology, vol. 20, no. 3, pp. 289–298, 2021. [CrossRef]
- A. Amaral, B. Lourenço, M. Marques, and J. Ramalho-Santos, “Mitochondria functionality and sperm quality,” Reproduction, vol. 146, no. 5, pp. R163-174, 2013. [CrossRef]
- J. Ballester et al., “Evidence for a functional glycogen metabolism in mature mammalian spermatozoa,” Molecular Reproduction and Development, vol. 56, no. 2, pp. 207–219, 2000. [CrossRef]
- M. J. Palomo, J. M. FernÁndez-Novell, A. Peña, J. J. Guinovart, T. Rigau, and J. E. Rodríguez-Gil, “Glucose- and fructose-induced dog-sperm glycogen synthesis shows specific changes in the location of the sperm glycogen deposition,” Molecular Reproduction and Development, vol. 64, no. 3, pp. 349–359, 2003. [CrossRef]
- J. Losano et al., “Effect of mitochondrial uncoupling and glycolysis inhibition on ram sperm functionality,” Reproduction in Domestic Animals, vol. 52, no. 2, pp. 289–297, 2017. [CrossRef]
- C. R. Moraes et al., “Effect of glucose concentration and cryopreservation on mitochondrial functions of bull spermatozoa and relationship with sire conception rate,” Animal Reproduction Science, vol. 230, p. 106779, Jul. 2021. [CrossRef]
- E. A. Bulkeley et al., “Effects from disruption of mitochondrial electron transport chain function on bull sperm motility,” Theriogenology, vol. 176, pp. 63–72, Dec. 2021. [CrossRef]
- M. Balbach et al., “Capacitation induces changes in metabolic pathways supporting motility of epididymal and ejaculated sperm,” Front. Cell Dev. Biol., vol. 11, Jun. 2023. [CrossRef]
- S.-K. Jin and W.-X. Yang, “Factors and pathways involved in capacitation: how are they regulated?,” Oncotarget, vol. 8, no. 2, pp. 3600–3627, Sep. 2016. [CrossRef]
- M. Tourmente, E. M. Tourmente, E. Sansegundo, E. Rial, and E. R. S. Roldan, “Capacitation promotes a shift in energy metabolism in murine sperm,” Front. Cell Dev. Biol., vol. 10, Aug. 2022. [CrossRef]
- Y. Mateo-Otero et al., “Sperm physiology and in vitro fertilising ability rely on basal metabolic activity: insights from the pig model,” Commun Biol, vol. 6, no. 1, pp. 1–9, Mar. 2023. [CrossRef]
- A. Cassina, P. Silveira, L. Cantu, J. Maria Montes, R. Radi, and R. Sapiro, “Defective Human Sperm Cells Are Associated with Mitochondrial Dysfunction and Oxidant Production,” BIOLOGY OF REPRODUCTION, vol. 93, no. 5, Nov. 2015. [CrossRef]
- A. Agarwal et al., “Comparative proteomic network signatures in seminal plasma of infertile men as a function of reactive oxygen species,” Clinical Proteomics, vol. 12, no. 1, 2015. [CrossRef]
- M. Boguenet et al., “Metabolomic signature of the seminal plasma in men with severe oligoasthenospermia,” ANDROLOGY, vol. 8, no. 6, pp. 1859–1866, Nov. 2020. [CrossRef]
- L. Li et al., “Metabolomic characterization of semen from asthenozoospermic patients using ultra-high-performance liquid chromatography-tandem quadrupole time-of-flight mass spectrometry,” BIOMEDICAL CHROMATOGRAPHY, vol. 34, no. 9, Sep. 2020. [CrossRef]
- K. Zhao et al., “Metabolomic Profiling of Human Spermatozoa in Idiopathic Asthenozoospermia Patients Using Gas Chromatography-Mass Spectrometry.,” Biomed Res Int, vol. 2018, p. 8327506, 2018. [CrossRef]
- D. Paoli et al., “Mitochondrial membrane potential profile and its correlation with increasing sperm motility,” FERTILITY AND STERILITY, vol. 95, no. 7, pp. 2315–2319, Jun. 2011. [CrossRef]
- S. Larriba, J. F. Sanchez-Herrero, R. Pluvinet, O. Lopez-Rodrigo, L. Bassas, and L. Sumoy, “Seminal extracellular vesicle sncRNA sequencing reveals altered miRNA/isomiR profiles as sperm retrieval biomarkers for azoospermia,” ANDROLOGY, vol. 12, no. 1, pp. 137–156, Jan. 2024. [CrossRef]
- R. D. Costa, S. Amaral, K. Redmann, S. Kliesch, and S. Schlatt, “Spectral features of nuclear DNA in human sperm assessed by Raman Microspectroscopy: Effects of UV-irradiation and hydration,” PLoS ONE, vol. 13, no. 11, 2018. [CrossRef]
- S. J. Calvert, S. Reynolds, M. N. Paley, S. J. Walters, and A. A. Pacey, “Probing human sperm metabolism using 13C-magnetic resonance spectroscopy,” MOLECULAR HUMAN REPRODUCTION, vol. 25, no. 1, pp. 30–41, Jan. 2019. [CrossRef]
- S. Reynolds, S. J. Calvert, M. N. Paley, and A. A. Pacey, “1H Magnetic Resonance Spectroscopy of live human sperm.,” Mol Hum Reprod, vol. 23, no. 7, pp. 441–451, Jul. 2017. [CrossRef]
- N. Aziz, J. Novotny, I. Oborna, H. Fingerova, J. Brezinova, and M. Svobodova, “Comparison of chemiluminescence and flow cytometry in the estimation of reactive oxygen and nitrogen species in human semen.,” Fertil Steril, vol. 94, no. 7, pp. 2604–2608, Dec. 2010. [CrossRef]
- P. Irigoyen, P. Pintos-Polasky, L. Rosa-Villagran, M. F. Skowronek, A. Cassina, and R. Sapiro, “Mitochondrial metabolism determines the functional status of human sperm and correlates with semen parameters,” Frontiers in Cell and Developmental Biology, vol. 10, 2022. [CrossRef]
- C. Almeida, M. Cunha, L. Ferraz, J. Silva, A. Barros, and M. Sousa, “Caspase-3 detection in human testicular spermatozoa from azoospermic and non-azoospermic patients,” INTERNATIONAL JOURNAL OF ANDROLOGY, vol. 34, no. 5, pp. E407–E414, Oct. 2011. [CrossRef]
- M. Barceló, A. Mata, L. Bassas, and S. Larriba, “Exosomal microRNAs in seminal plasma are markers of the origin of azoospermia and can predict the presence of sperm in testicular tissue.,” Hum Reprod, vol. 33, no. 6, pp. 1087–1098, Jun. 2018. [CrossRef]
- V. Calle-Guisado et al., “AMP-activated kinase in human spermatozoa: identification, intracellular localization, and key function in the regulation of sperm motility,” ASIAN JOURNAL OF ANDROLOGY, vol. 19, no. 6, pp. 707–714, Dec. 2017. [CrossRef]
- T. Deng et al., “Lipidomics random forest algorithm of seminal plasma is a promising method for enhancing the diagnosis of necrozoospermia,” METABOLOMICS, vol. 20, no. 3, May 2024. [CrossRef]
- D. Fietz et al., “Proteomic biomarkers in seminal plasma as predictors of reproductive potential in azoospermic men.,” Front Endocrinol (Lausanne), vol. 15, p. 1327800, 2024. [CrossRef]
- T. Fukuda, H. Miyake, N. Enatsu, K. Matsushita, and M. Fujisawa, “Seminal level of clusterin in infertile men as a significant biomarker reflecting spermatogenesis,” ANDROLOGIA, vol. 48, no. 10, pp. 1188–1194, Dec. 2016. [CrossRef]
- M. Hashemitabar, S. Sabbagh, M. Orazizadeh, A. Ghadiri, and M. Bahmanzadeh, “A proteomic analysis on human sperm tail: comparison between normozoospermia and asthenozoospermia.,” J Assist Reprod Genet, vol. 32, no. 6, pp. 853–863, Jun. 2015. [CrossRef]
- V. Jayaraman, S. Ghosh, A. Sengupta, S. Srivastava, H. M. Sonawat, and P. K. Narayan, “Identification of biochemical differences between different forms of male infertility by nuclear magnetic resonance (NMR) spectroscopy,” JOURNAL OF ASSISTED REPRODUCTION AND GENETICS, vol. 31, no. 9, pp. 1195–1204, Sep. 2014. [CrossRef]
- M.-A. Kyrgiafini, T. Giannoulis, A. Chatziparasidou, N. Christoforidis, and Z. Mamuris, “Unveiling the Genetic Complexity of Teratozoospermia: Integrated Genomic Analysis Reveals Novel Insights into lncRNAs’ Role in Male Infertility,” International Journal of Molecular Sciences, vol. 24, no. 19, 2023. [CrossRef]
- J. Li et al., “Genome-wide methylation analyses of human sperm unravel novel differentially methylated regions in asthenozoospermia,” EPIGENOMICS, vol. 14, no. 16, pp. 951–964, Aug. 2022. [CrossRef]
- J. Liang et al., “Proteomic Profile of Sperm in Infertile Males Reveals Changes in Metabolic Pathways,” Protein Journal, vol. 40, no. 6, pp. 929–939, 2021. [CrossRef]
- Y. Liu et al., “Raman spectroscopy as an ex vivo noninvasive approach to distinguish complete and incomplete spermatogenesis within human seminiferous tubules,” Fertility and Sterility, vol. 102, no. 1, pp. 54-60.e2, 2014. [CrossRef]
- A. D. Martins, M. K. P. Selvam, A. Agarwal, M. G. Alves, and S. Baskaran, “Alterations in seminal plasma proteomic profile in men with primary and secondary infertility,” SCIENTIFIC REPORTS, vol. 10, no. 1, May 2020. [CrossRef]
- E. Olesti et al., “Low-polarity untargeted metabolomic profiling as a tool to gain insight into seminal fluid,” Metabolomics, vol. 19, no. 6, 2023. [CrossRef]
- E. Olesti et al., “Steroid profile analysis by LC-HRMS in human seminal fluid,” Journal of Chromatography B: Analytical Technologies in the Biomedical and Life Sciences, vol. 1136, 2020. [CrossRef]
- C. Paiva et al., “Identification of endogenous metabolites in human sperm cells using proton nuclear magnetic resonance (1H-NMR) spectroscopy and gas chromatography-mass spectrometry (GC-MS),” Andrology, vol. 3, no. 3, pp. 496–505, 2015. [CrossRef]
- S. Qiao et al., “Seminal plasma metabolomics approach for the diagnosis of unexplained male infertility,” PLOS ONE, vol. 12, no. 8, Aug. 2017. [CrossRef]
- L. Soler, V. Labas, A. Thélie, I. Grasseau, A.-P. Teixeira-Gomes, and E. Blesbois, “Intact cell MALDI-TOF MS on sperm: A molecular test for male fertility diagnosis,” Molecular and Cellular Proteomics, vol. 15, no. 6, pp. 1998–2010, 2016. [CrossRef]
- A. Sulc et al., “MALDI Imaging Mass Spectrometry Reveals Lipid Alterations in Physiological and Sertoli Cell-Only Syndrome Human Testicular Tissue Sections.,” Int J Mol Sci, vol. 25, no. 15, Jul. 2024. [CrossRef]
- M. Torra-Massana et al., “Altered mitochondrial function in spermatozoa from patients with repetitive fertilization failure after ICSI revealed by proteomics,” Andrology, vol. 9, no. 4, pp. 1192–1204, 2021. [CrossRef]
- E. Torrezan-Nitao, S. G. Brown, E. Mata-Martínez, C. L. Treviño, C. Barratt, and S. Publicover, “[Ca2+]ioscillations in human sperm are triggered in the flagellum by membrane potential-sensitive activity of CatSper,” Human Reproduction, vol. 36, no. 2, pp. 293–304, 2021. [CrossRef]
- A. Vashisht, P. K. Ahluwalia, and G. K. Gahlay, “A Comparative Analysis of the Altered Levels of Human Seminal Plasma Constituents as Contributing Factors in Different Types of Male Infertility.,” Curr Issues Mol Biol, vol. 43, no. 3, pp. 1307–1324, Sep. 2021. [CrossRef]
- A. Wainstein et al., “MicroRNAs expression in semen and testis of azoospermic men,” ANDROLOGY, vol. 11, no. 4, pp. 687–697, 23. 20 May. [CrossRef]
- C. Wang et al., “Altered Profile of Seminal Plasma MicroRNAs in the Molecular Diagnosis of Male Infertility,” CLINICAL CHEMISTRY, vol. 57, no. 12, pp. 1722–1731, Dec. 2011. [CrossRef]
- X. Wu et al., “Unraveling epigenomic abnormality in azoospermic human males by WGBS, RNA-Seq, and transcriptome profiling analyses.,” J Assist Reprod Genet, vol. 37, no. 4, pp. 789–802, Apr. 2020. [CrossRef]
- Y. Xu et al., “RNASET2 impairs the sperm motility via PKA/PI3K/calcium signal pathways,” REPRODUCTION, vol. 155, no. 4, pp. 383–392, Apr. 2018. [CrossRef]
- K. Zhao, Q. Zhang, R. Cong, Z. Xu, Y. Xu, and J. Han, “Metabolomic profiling of human semen in patients with oligospermia using high performance liquid chromatography-tandem mass spectrometry.,” Sci Rep, vol. 14, no. 1, p. 23739, Oct. 2024. [CrossRef]
Figure 1.
Flowchart detailing the selection of studies for inclusion in the review. PRISM.
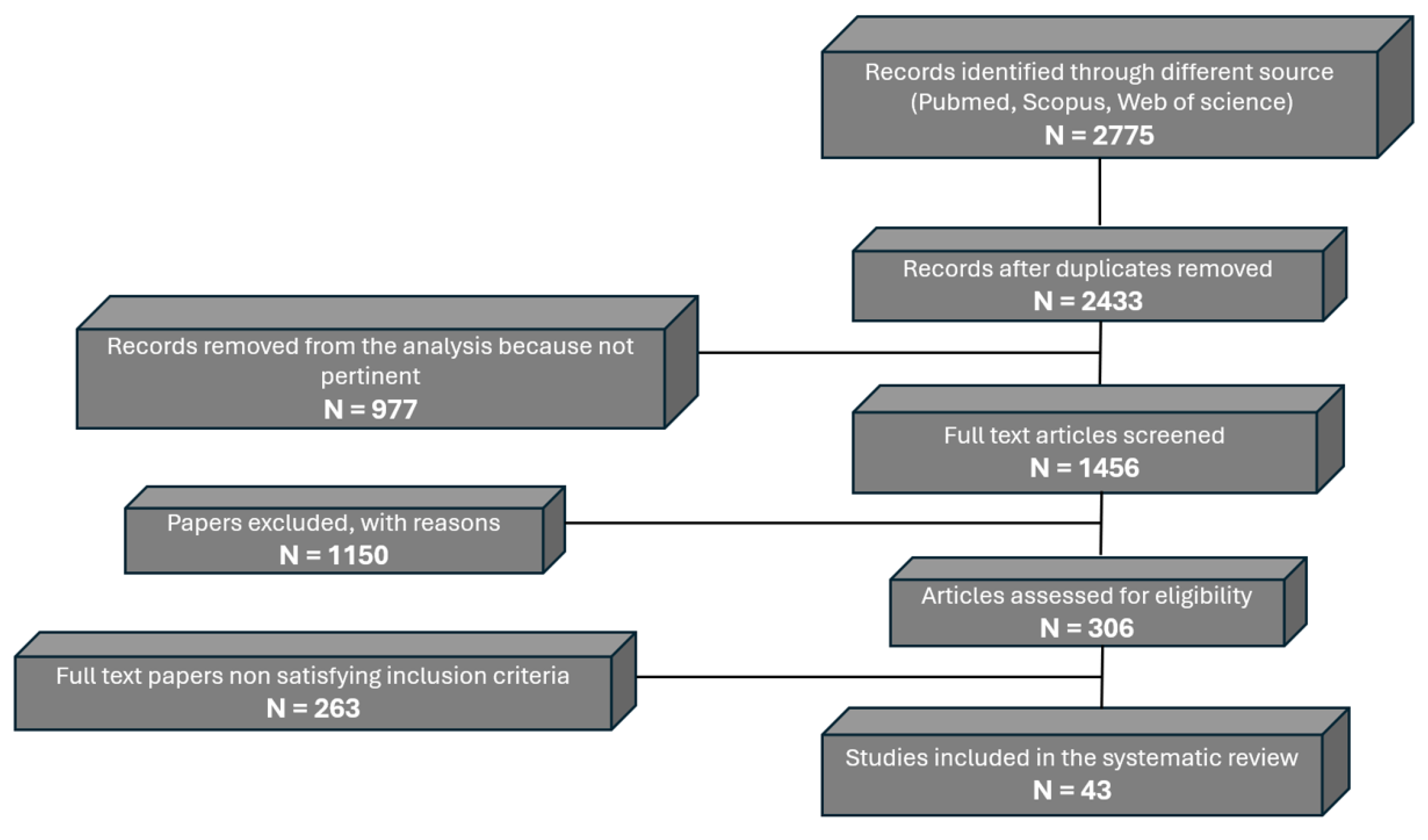

Table 1.
Studies on Innovative Techniques in Sperm Metabolism and Male Infertility.
| Authors | Study Aim | Methods/Tools | Results |
|---|---|---|---|
| Agarwal et al., 2015 [41] | Identify seminal plasma proteins involved in ROS-mediated male infertility |
|
|
| Almeida et al., 2011 [52] | Quantify testicular sperm apoptosis via active caspase-3 in normal and impaired spermatogenesis |
|
|
| Amaral et al., 2013 [8] | Characterize human sperm tail proteome, focusing on metabolism-related proteins |
|
|
| Aziz et al., 2010 [50] | Determine cell type contributions to intracellular H2O2 and peroxynitrite production in sperm |
|
|
| Barceló et al., 2018 [53] | Evaluate seminal plasma exosomal mirnas as markers for Azoospermia origin and sperm presence |
|
|
| Boguenet et al., 2020 [42] | Assess metabolomic signatures of seminal plasma in severe Oligoasthenospermia |
|
|
| Calle-Guisado et al., 2017 [54] | Identify and localize AMP- activated protein kinase (AMPK) in human sperm and evaluate its role in sperm motility |
|
|
| Calvert et al., 2019 [48] | Investigate sperm metabolism |
|
|
| Cassina et al., 2015 [40] | Analyze mitochondrial function and oxidative stress in human sperm affecting fertility |
|
|
| Costa et al., 2018 [47] | Assessement of sperm DNA damage and biochemical features |
|
|
| Deng et al., 2024 [55] | Profile seminal plasma lipid composition in necrozoospermia and evaluate lipid biomarkers |
|
|
| Fietz et al., 2024 [56] | Discover seminal plasma biomarkers for non-invasive differential diagnosis of Obstructive Azoospermia (OA) vs. Non-Obstructive Azoospermia (NOA) |
|
|
| Fukuda et al., 2016 [57] | Assess impact of seminal clusterin level on spermatogenesis and sperm retrieval in infertile men |
|
|
| Hashemitabar et al., 2015 [58] | Identify novel biomarkers for asthenozoospermia via sperm tail proteomic analysis |
|
|
| He et al., 2024 [9] | Investigate seminal microbiome and metabolome role in high sperm DNA fragmentation index (HDFI) |
|
|
| Irigoyen et al., 2022 [51] | Assess sperm mitochondrial metabolism and ROS production as tools to complement semen analysis |
|
|
| Jayaraman et al., 2014 [59] | Analyze seminal plasma metabolic profiles in idiopathic/male factor infertility |
|
|
| Kyrgiafini et al., 2023 [60] | Identify lncRNA mutations and expression in teratozoospermia |
|
|
| Larriba et al., 2024 [46] | Small RNA profiling in seminal extracellular vesicles for azoospermia classification |
|
|
| Li et al., 2022 [61] | Investigate DNA methylation patterns in asthenozoospermia |
|
|
| Liang et al., 2021 [62] | Proteomic profiling of sperm in severe oligoasthenoteratozoospermia |
|
|
| Liu et al., 2014 [63] | Identify spermatogenesis in testicular tissue |
|
|
| Martins et al., 2020 [64] | Characterize seminal plasma proteome in primary and secondary infertility |
|
|
| Milardi et al., 2014 [10] | Identify seminal biomarkers for secondary male hypogonadism (HH) |
|
|
| Olesti et al., 2023 [65] | Correlate metabolomic profiles with semen quality in young men |
|
|
| Olesti et al., 2020 [66] | Develop steroidomics strategy for human seminal fluid |
|
|
| Paiva et al., 2015 [67] | Comprehensive metabolomic characterization of human sperm cell |
|
|
| Paoli et al., 2011 [45] | Correlate sperm mitochondrial integrity with motility |
|
|
| Qiao et al., 2017 [68] | Metabolic profiling of unexplained male infertility (UMI) |
|
|
| Reynolds et al., 2017 [49] | Examine sperm molecules |
|
|
| Sharma et al., 2013 [11] | Identify seminal plasma proteins as biomarkers of sperm quality |
|
|
| Sharma et al., 2013 [12] | Proteomic profile changes in spermatozoa with elevated ROS |
|
|
| Soler et al., 2016 [69] | Fertility-predictive model profiles of spermatozoa |
|
|
| Sulc et al., 2024 [70] | Phospholipid expression in Sertoli cell-only syndrome (SCOS) testis |
|
|
| Massana et al., 2021 [71] | Proteomic analysis of sperm in fertilization failure after ICSI |
|
|
| Nitao et al., 2021 [72] | Progesterone-induced Ca2+ oscillations in human sperm |
|
|
| Vashisht et al., 2021 [73] | Evaluate seminal plasma biochemical and immunological markers in male infertility |
|
|
| Wainstein et al., 2023 [74] | Study microRNA profiles in semen and testicular tissue of azoospermic men |
|
|
| Wang et al., 2011 [75] | Seminal plasma miRNAs in infertile men and diagnostic value |
|
|
| Wu et al., 2020 [76] | DNA methylation in testicular cells of azoospermia patients |
|
|
| Xu et al., 2018 [77] | Ribonuclease (RNASET2) levels in sperm and relation to motility |
|
|
| Zhao et al., 2018 [44] | Metabolic profiling of idiopathic asthenozoospermia sperm cells |
|
|
| Zhao et al., 2024 [78] | Semen metabolic profiling in oligospermia patients |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



