Submitted:
05 May 2025
Posted:
09 May 2025
You are already at the latest version
Abstract
In controlled environment agriculture (CEA), supplementary lighting, particularly light-emitting diode (LED) technology, plays an important role in optimizing plant growth and development. Blue light has emerged as a critical wavelength significantly affecting plant morphogenesis, photosynthesis, and essential physiological processes. However, species-specific guidelines for optimizing blue light parameters such as intensity, duration, and spectral ratios remain insufficiently developed. Furthermore, the spectral requirements of plants change over different developmental stages, highlighting distinct responses to blue light at each growth phase. This review synthesizes existing knowledge on the influence of blue light on morphological adaptation, photosynthetic efficiency, flowering, and secondary metabolism, with an emphasis on differential responses across diverse plant species. We emphasize the need for growth-stage-specific lighting frameworks and scalable strategies tailored to real-world CEA systems. Interdisciplinary collaboration, integrating molecular biology, genomics, and horticultural engineering, is necessary to enhance understanding of blue light-driven regulatory networks, optimize photoreceptor responses, and facilitate systematic validation of adaptive lighting approaches, ultimately advancing sustainable horticulture and next-generation CEA innovations.
Keywords:
blue light
; light quality
; wavelength
; photomorphogenesis
; photosynthetic efficiency
; flowering
; secondary metabolism
; LED lighting
; controlled environment agriculture
1. Introduction
Controlled environment agriculture (CEA) represents a significant technological advancement in modern farming and is characterized by the precise management of environmental conditions to maximize crop productivity [1-3]. A key element of CEA is the use of supplemental lighting, since plant development and physiology are highly dependent on the availability of light spectra, intensity, and duration in controlled environmental growth conditions [4,5]. In CEA systems, supplemental lighting, particularly light-emitting diodes (LEDs), offers significant advantages over traditional lighting systems, including enhanced photosynthetic efficiency and reduced heat loss [6,7].
Light quality, defined as the spectral composition or color spectrum, shows distinct responses in plants. Red light (600–700 nm) is crucial for the development of the photosynthetic apparatus and starch accumulation [8]. Blue light (400–500 nm) plays a significant role in chlorophyll formation, stomatal opening, and photomorphogenesis [9-12]. Far-red light (700–800 nm) modulates flowering in photoperiod-sensitive species, and green light (500–600 nm) can counteract the effects of blue light on stomatal opening [13,14]. These wavelength-specific responses highlight the importance of adjusting light spectra to optimize growth conditions in CEA systems.
Despite advancements, research on optimizing blue light applications is still limited. Species-specific requirements for blue light intensity and duration remain poorly characterized, and their roles across growth phases, such as enhancing secondary metabolite biosynthesis in vegetative stages and regulating flowering timing, require further exploration [15-20]. In addition, interactions between blue light and other spectral components (e.g., red and far-red) complicate efforts to define universal ‘light recipes’ for horticultural crops. Resolving these challenges requires clarifying the effects of blue light on photosynthetic efficiency, stress resilience, and molecular pathways while addressing conflicting findings in applied studies.
This review provides a comprehensive understanding of current knowledge on the effects of blue light on plant growth and development, thereby contributing to the optimization of lighting strategies in modern horticultural practices. The specific objectives are to (1) characterize blue light-driven physiological and morphological adaptations, including stem elongation, leaf expansion, and root architecture; (2) examine its role in enhancing photosynthetic efficiency; (3) explore the role of blue light in modulating flowering and photoperiodic responses; (4) evaluate its impact on secondary metabolic pathways; and (5) evaluate its contribution to stress resilience (e.g., drought and salt stresses).
2. Blue Light
Blue light can regulate various aspects of development, including plant morphogenesis, flowering, and the synthesis of secondary metabolites (Figure 1). Unlike red light, which is primarily absorbed by phytochromes [21], blue light triggers a different set of physiological responses essential for plant adaptation and growth. Plants have evolved sophisticated mechanisms to perceive and respond to blue light, primarily through specialized photoreceptors such as cryptochromes (CRY1 and CRY2) and phytotropins (Phot1 and Phot2) [22]. These photoreceptors enable plants to decode light signals and respond physiologically, balancing energy capture with environmental adaptation.
Cryptochromes, photolyase-like receptors, regulate gene expression and photomorphogenic responses, thereby influencing plant development and growth [23]. CRY1 dominates under high-intensity blue light, suppresses hypocotyl elongation, regulates circadian rhythms, and influences light-responsive gene expression. In contrast, CRY2 is active under low- to moderate-intensity blue light, modulating flowering time via photoperiod sensing. CRY1 is stable under light conditions, but CRY2 degrades rapidly under prolonged illumination. These differences enable CRY1 and CRY2 to perform complementary roles in plant development and adaptation to varying light environments.
Phytotropins, with light‒oxygen‒voltage (LOV) domains, mediate phototropic responses, stomatal opening, chloroplast movement, and leaf positioning. Upon blue light absorption, LOV domains trigger conformational changes, activating signaling cascades that optimize light capture and gas exchange. For example, Phot1- and Phot2-driven stomatal opening increase CO2 uptake and photosystem II (Fv/Fm) efficiency, as demonstrated in tomatoes [24].
In greenhouses and vertical farms, blue light is used to manipulate crop architecture and quality. For example, blue light exposure reduces stem elongation, producing compact plants ideal for lettuce and herbs [25], while enhancing leaf pigmentation, antioxidant levels (e.g., phenolics, vitamins), and flavor compounds in leafy greens. However, the high photon energy of blue light poses risks to plants. For example, excessive blue light can induce photoprotective responses [26], such as reduced leaf/root biomass, elevated carotenoids, and altered leaf angles [27]. The adverse effects of blue light are often mitigated by combining it with red light. For example, red–blue mixtures increase the biomass and photosynthetic efficiency of basil (Ocimum basilicum), coriander (Coriandrum sativum L.), and Salvia miltiorrhiza [28,29]. Nevertheless, species- and stage-specific blue light requirements remain poorly defined. For example, seedlings may prioritize blue light for stomatal development, whereas reproductive stages might demand spectral shifts to regulate flowering. Resolving these dynamics is key to designing adaptive lighting regimes that maximize yield and nutritional value without inducing light stress.
3. Effects of Blue Light on Plant Morphology and Development
Blue light has long been recognized for its role in inhibiting stem elongation [30-32]. This phenomenon was initially observed in studies using broadband light or impurity blue light sources, such as blue-colored fluorescent lamps, which inadvertently emit residual red and far-red wavelengths [30]. These spectral impurities complicated early interpretations, as the activation of phytochromes by non-blue wavelengths potentially affected the effects of blue light.
With the advent of LED technology, studies on blue light-mediated plant elongation have reported inconsistent results, which can be attributed to the use of different plant species, genotypes (varieties), growth stages (e.g., seedlings and mature plants), LED lighting characteristics [33-35], and cultivation conditions [36]. Even within the same trial, different durations of lighting can lead to varying plant elongation responses [37-39].
3.1. Growth and Development of Young Plants
To determine whether blue light promotes growth and development more effectively than red light, we reviewed studies that utilized blue and red LED lights alone under various conditions and in different crops. Blue light has been reported to inhibit seedling growth in artichokes (Cynara cardunculus var. scolymus) [40], bamboo (Phyllostachys edulis) [41], barley (Hordeum vulgare) [42], bitter gourd (Momordica charantia) [43], coriander (Coriandrum sativum) [44], impatiens (Impatiens walleriana) [45,46], kiwi (Actinidia chinensis) [47], maize (Zea mays) [48], mulberry (Morus alba) [49], radish (Raphanus sativus) [50], rice (Oryza sativa) [51,52], salvia (Salvia splendens) [45,46], and zinnia (Zinnia elegans) [53].
Blue light has been reported to promote the growth of young plants more effectively than red light for various crops and cultivation conditions (Figure 2). In addtion, blue light was found to promote seedling growth in arugula (Brassica eruca) [54-57], calibrachoa (Calibrachoa × hybrida) [58-60], eggplant (Solanum melongena) [61,62], geranium (Pelargonium hortorum) [59], pea (Pisum sativum) [63], and sesame (Sesamum indicum) [64]. However, inconsistent results have been reported for Arabidopsis (Arabidopsis thaliana) [36,65], cabbage (Brassica oleracea var. Capitata) [54-56,66,67], cherry tomato (Solanum lycopersicum var cerasiforme) [68-70], cucumber (Cucumis sativus) [34,50,71,72], kale (Brassica napus) [54,55,58,66,73], lettuce (Lactuca sativa) [27,50,61,74,75], marigold (Tagetes erecta) [56,57,75], mustard (Brassica juncea) [54,56,57,60,76], pepper (Capsicum annuum) [50,77,78], petunia (Petunina × hydrida) [53,59,75], soybean (Glycine max) [50,79], squash (Cucurbita moschata Duch.) [38,80], and tomato (Solanum lycopersicum) [45,46,50,65,71,72,81,82].
for both treatments. Panels a-h are adapted from Kong and Zheng [36].
3.2. Stem Elongation and Leaf Expansion of Mature Plants
Similar to the growth and development of young plants, blue light can regulate the morphology of mature plants [50]. For example, blue light supplementation reduces the nitrate content in lettuce and spinach while increasing the content of phenolic compounds, ascorbic acid, and anthocyanins, thereby enhancing both yield and nutritional value [67,83-86]. However, the effects of blue light are highly dependent on the species. In marigolds and salvia, monochromatic blue light increases plant height but reduces flower bud formation and bloom duration [87]. In contrast, in strawberries [6,88] and capsicum [89], the combination of blue light with red light enhances yield and fruit quality.
The interaction between blue light and other wavelengths is critical for optimizing plant growth. Red–blue LED combinations often outperform monochromatic red or blue light in crops such as lettuce, promoting a larger leaf area, greater biomass accumulation, and improved chloroplast development, particularly when blue is optimized within the spectral composition alone [90,91]. However, species-specific responses exist [86,92]. For example, increasing the blue light percentages can decrease tomato growth, primarily through its impact on morphology and subsequent light interception [93]. Green light, although traditionally considered less efficient for photosynthesis, enhances carbon fixation by penetrating deeper into leaf layers and supporting photosynthesis in shaded tissues [94]. In lettuce, red, blue, and green LED light promotes greater fresh and dry mass than red light alone does [95,96], underscoring the benefits of spectral diversity in the regulation of plant growth.
Kong and Zheng [36] reported that increasing the proportion of blue light (0–100%) in red LED systems promoted stem elongation in petunias, calibrachoa, geraniums, and marigolds, which contrasts with the compact growth typically associated with blue light. This phenomenon highlights the role of the phytochrome photostationary state (PPS): pure blue light (e.g., LEDs) reduces PPS, promoting elongation, whereas impure blue light (e.g., fluorescent lamps with residual red/far-red light) elevates PPS, inhibiting elongation [72]. The addition of low-intensity red light to blue light counteracts elongation, but this effect is reversed by far-red light, suggesting that phytochrome B (phyB) plays a key role as a mediator. In contrast, the addition of ultraviolet (UVA/UVB) or green light has a minimal effect on elongation, emphasizing the dominance of red light in modulating blue light responses.
Blue light also affects leaf morphology, although its effects vary depending on the stage of development. In lettuce, blue light enhances leaf thickness and cell division at later growth stages but inhibits new leaf initiation and reduces fresh weight during early growth [97,98]. In addition, blue light has been reported to alleviate ‘red light syndrome’ in cucumber plants, characterized by growth inhibition and stress symptoms, such as a low maximum quantum yield of chlorophyll fluorescence and unresponsive stomatal conductance [99].
3.3. Root Development and Architecture
While blue light is widely recognized for its role in regulating aboveground morphology, emerging evidence underscores its systemic influence on root system architecture. Roots, although shielded from direct light in natural settings, exhibit distinct architectural responses to blue light exposure. In Arabidopsis, blue light suppresses primary root elongation while stimulating lateral root formation, a response mediated by auxin redistribution and signaling [100]. This hormonal reprogramming enhances root branching, potentially improving nutrient and water uptake in compact soils. Blue light also modulates root gravitropism, i.e., the orientation of root growth in response to gravity. Under microgravity conditions, Arabidopsis roots exhibit positive phototropism toward blue light, a response typically masked by the gravitational pull of Earth [101]. These findings suggest that gravity can obscure blue light-induced phototropic responses in roots.
The effects of blue light on root development vary across species, indicating species-specific responses. In cherry rootstock (Prunus avium), blue light enhances adventitious root formation under in vitro conditions, improving transplant success rates [102]. In contrast, blue light inhibits primary root growth in cucumber (Cucumis sativus), highlighting the different regulatory mechanisms among plant species [103]. These findings suggest that while blue light can enhance root branching in certain species, it may limit root growth in others, depending on specific light perception and auxin transport mechanisms [104]. Optimizing root architecture via blue light could increase resource efficiency in hydroponic and aeroponic systems.
3.4. Factors Influencing Blue Light Responses
3.4.1. Species- and Genotype-Specific Sensitivity
The effects of blue light on plant morphology vary significantly among species and genotypes. For example, blue light promotes stem elongation in tomato [82], whereas it inhibits growth in cucumber [50] and Impatiens balsamina [53]. Such contrasting responses likely result from variations in cryptochrome expression levels or differences in auxin sensitivity among species and even cultivars within a species [97].
3.4.2. Spectral Interactions and Light Recipes
The effects of blue light on plant development are significantly influenced by its interactions with other wavelengths, resulting in varied morphological and physiological outcomes. For example, supplementing blue light with far-red light has been shown to increase stem elongation in certain species by modulating phytochrome activity, which is sensitive to red and far-red wavelengths [105]. In Cannabis sativa, blue light increased shoot dry mass but inhibited elongation, a limitation counteracted by supplemental far-red, which enhanced shoot length and yield without compromising biomass accumulation [106]. This interaction highlights the role of phytochrome-mediated signaling in balancing blue light’s growth-inhibiting effects, enabling tailored light recipes to optimize elongation and biomass simultaneously. However, in buckwheat and cucumber seedlings, optimal biomass and height were observed under blue LED treatment alone [33,34]. These findings underscore the importance of considering spectral interactions when designing light recipes for horticultural practices.
3.4.3. Night Interruption
Night interruption refers to breaking up the long dark period by briefly providing artificial lighting, thus creating modified long-day conditions for plants [107]. The quality of night interruption significantly affects morphogenesis across various plants [108-116]. Park et al. [114] reported that employing blue light during the night interruption period suppresses plant elongation. Additionally, they demonstrated that combining blue and red light promoted leaf expansion, whereas combining blue and far-red light suppressed this expansion. The enhanced relative growth rate observed under combined blue and red lights likely results from a synergistic interaction between these wavelengths. Moreover, night interruption techniques effectively simulate extended daylight hours by briefly illuminating plants during the dark phase, increasing plant growth and developmental responses associated with long-day conditions [109,117].
4. Photosynthetic Efficiency
Blue light plays a crucial role in optimizing photosynthesis by serving as an energy source for photosystems and as a regulatory signal for stomatal dynamics, chloroplast positioning, and gene expression [90,118]. Although blue light enhances photosynthetic performance, increasing blue light percentages can lead to photooxidative damage, impair chlorophyll synthesis, and reduce carbon assimilation [25]. Blue light significantly influences stomatal conductance and density, which are key factors for efficient CO₂ uptake.
In cucumber, a balanced red–blue light ratio (50:50) maximizes stomatal opening and gas exchange, surpassing monochromatic treatments [25]. As shown in Figure 3, exposure to red light alone (0% blue light) results in dysfunctional photosynthesis in plant leaves and stomatal conductance that is unresponsive to irradiance. Introducing as little as 7% blue light during growth prevents these dysfunctions, indicating that blue light is qualitatively essential for photosynthetic operation. Furthermore, the photosynthetic capacity (Aₘₐₓ) of leaves grown under 7% blue light is twice that of those grown without blue light, and Aₘₐₓ continues to increase with increasing blue light percentage up to 50% (Figure 3a). At 100% blue light, although Aₘₐₓ decreases, photosynthetic functionality remains normal. The increase in Aₘₐₓ with increasing blue light percentage (0–50%) was correlated with increases in leaf mass per unit area (LMA) (Figure 3b), chlorophyll content per area (Figure 3c), nitrogen content per area, and stomatal conductance. Notably, Aₘₐₓ per leaf chlorophyll, Aₘₐₓ per leaf dry weight, and photosynthetic nitrogen use efficiency (PNUE) increased with blue light up to 50% but remained the same between 50% and 100% (Figure 3d-f). Above 15% blue light, parameters such as Aₘₐₓ, LMA, chlorophyll content, PNUE, and the chlorophyll-to-nitrogen ratio exhibit relationships similar to those observed in leaf responses to increased irradiance intensity. These results suggest that blue light during growth is not only qualitatively required for normal photosynthetic function but also quantitatively mediates leaf responses similar to those induced by higher light intensities.
5. Flowering and Photoperiodic Responses
Blue light has a significant influence on flowering and photoperiodic responses in plants. Despite considerable advancements, inconsistencies persist in research findings due to variability among plant species and experimental parameters, such as light intensities, durations, spectral compositions, and interactions with environmental factors like temperature and irradiance [119]. Plants are broadly categorized as long-day, short-day, or day-neutral plants based on their flowering response to day length [120]. Long-day plants flower when exposed to light periods exceeding a critical threshold, whereas short-day plants require uninterrupted darkness for floral induction [121]. Day-neutral plants flower independently of photoperiod [122,123]. Long-day and Short-day plants are further categorized as qualitative (obligate), which strictly depend on photoperiodic conditions, and quantitative (facultative), where the photoperiod increases but is not essential for flowering [122].
At the molecular level, photoperiodic responses are mediated by light-activated enzymes, the circadian clock, and the FLOWERING LOCUS T (FT) gene, which produces florigen—a signal triggering flowering [120]. Blue light, perceived primarily by cryptochrome photoreceptors, stabilizes the CONSTANS (CO) protein, a key regulator of FT expression [111]. For example, in Arabidopsis, long-day conditions increase FT transcription, accelerating flowering compared with short-day environments [124]. This mechanism is conserved across species but modulated by the quality and intensity of light.
In commercial horticulture, growers strategically manipulate photoperiods via night interruption and day extension to control flowering. Night interruption disrupts darkness with low-intensity light (1–10 μmol m−2 s−1), accelerating flowering in long-day plants but delaying flowering in short-day plants [122]. Day extension extends effective daylight exposure, primarily benefiting long day plants. While broad-spectrum light at 1–2 μmol m−2 s−1 is typically sufficient for regulating flowering [125], the efficacy of blue light varies significantly across species and intensities. For example, low-intensity blue light (1–2 μmol m−2 s−1) has a minimal effect on the flowering of both long-day plants, e.g., dianthus (Dianthus chinensis) and rudbeckia (Rudbeckia hirta), and short-day plants, e.g., chrysanthemum (Chrysanthemum × morifolium), cosmos (Cosmos sulfureus), dahlia (Dahlia pinnata), and marigold (Tagetes erecta) [126]. However, long day plants such as coreopsis (Coreopsis grandiflora) and snapdragon (Antirrhinum majus) respond to intensities as low as 5 μmol m⁻² s⁻¹, whereas rudbeckia (Rudbeckia hirta) and petunia (Petunia hybrida) require 15 μmol m⁻² s⁻¹ [127].
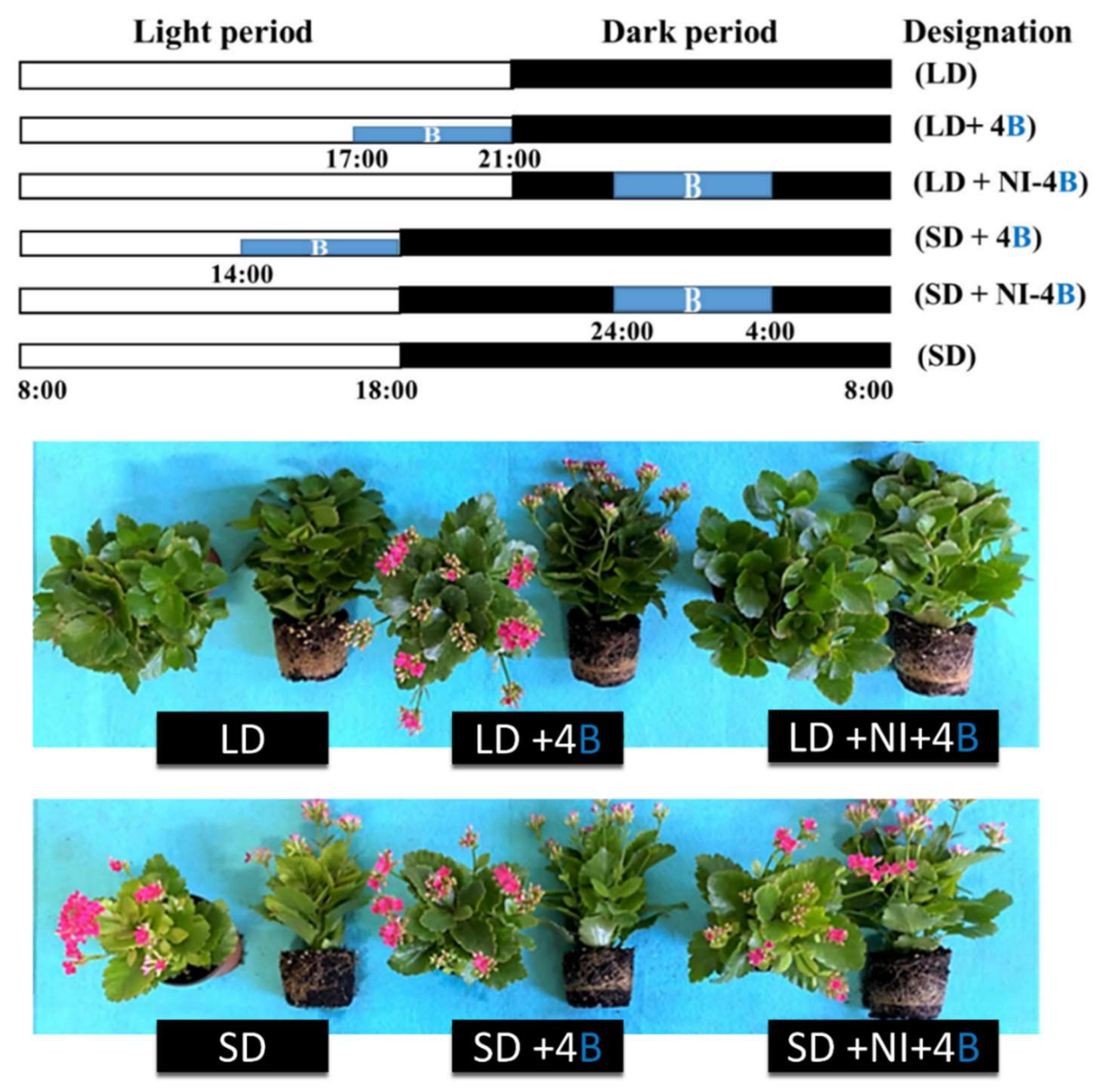
In contrast to earlier findings, low-intensity blue light (10 µmol m⁻² s⁻¹) supplementation during photoperiods promotes flowering in qualitative short-day plants such as chrysanthemum (Dendranthema grandiflorum) [109,110] and kalanchoe [16], even under noninductive long-day conditions. Shoot tip-targeted blue light night interruption reduces stem elongation by 30% and advances flowering by 14 days via CRY-mediated FT upregulation [111]. As shown in Figure 4, supplying low-intensity blue light under short-day or long-day conditions effectively promoted flowering in short-day plants, challenging traditional findings that prioritize uninterrupted darkness for floral induction. These results highlight the potential of blue light as a tool for commercial floriculture. However, the responses remain cultivar specific. For example, Kalanchoe ‘Spain’ flowers under short-day and blue light NI, but Kalanchoe ‘Lipstick’ remains unaffected [128]. The positioning of light used for night interruption further refines outcomes. Compared with leaf-targeted treatments, shoot tip-targeted blue light night interruption in chrysanthemum enhances FT expression by 50%, optimizing flowering induction [110]. Despite its advantages, challenges persist. High blue light intensity (>20 µmol m⁻² s⁻¹) delays flowering in some SDPs (e.g., chrysanthemum morifolium), necessitating dynamic spectral adjustments [129].
The influence of blue light extends to diverse crops. In everbearing strawberry plants (Fragaria × ananassa), 24-hour exposure to blue light accelerates flowering and increases yield, whereas supplemental blue light enhances flower cluster formation in greenhouse production [130]. The blue-dominant spectrum of blueberries (Vaccinium corymbosum) leads to improved fruit quality, although cultivar-specific responses are observed [131]. Vegetables such as tomatoes and peppers benefit from blue-red (RB) LED combinations that balance compact growth and early flowering [132]. Specialty crops, such as saffron (Crocus sativus) and cannabis (Cannabis sativa), underscore the economic potential of blue light. High-intensity blue light increases saffron stigma yield and elevates cannabinoid concentrations in cannabis [133].
Mechanistic gaps, such as cryptochrome‒phytochrome interactions under varying spectral ratios, require further research. Innovations such as smart lighting systems that adapt spectra in real-time could optimize energy use and crop yields. Additionally, leveraging blue light for speed breeding, i.e., accelerating generational cycles in short-day crops, is promising for sustainable agriculture. By harmonizing spectral precision with plant physiology, horticulture can achieve year-round control of flowering, aligning production with global agricultural demands.
6. Plant Secondary Metabolism
Blue light is a critical regulator of plant secondary metabolism, driving the synthesis of bioactive compounds critical for defense, stress adaptation, and ecological interactions [7]. These metabolites, including flavonoids, anthocyanins, phenolics, carotenoids, glucosinolates, alkaloids, and terpenoids, enhance plant resilience against herbivores, pathogens, and oxidative stress, while also contributing to their nutritional and medicinal value [134]. The molecular regulation of these compounds involves photoreceptors, such as CRYs and PHYs, which activate transcription factors, including hypocotyl 5 (HY5), phytochrome interacting factor 4 (PIF4), and MYC2, to modulate biosynthetic pathways [135,136]. For example, blue light upregulates functional metabolites such as rutin and catechins in longan embryogenic calli through these regulatory pathways [135,136]. Similarly, citrus fruits exposed to blue light exhibit accelerated carotenoid accumulation and chlorophyll degradation, enhancing fruit coloration via the activation of pigment-related structural genes [137-139].
Cryptochromes, particularly CRY1 and CRY2, are important to blue light signaling and regulate stomatal dynamics, biomass allocation, and metabolite biosynthesis [7,140]. These processes often synergize with phytochromes (PHYA/PHYB) for full pathway activation. In tomato, blue light induces anthocyanin accumulation through CRY1 but requires the participation of phytochrome, even at low levels [141]. The effects of blue light are further modulated by interactions with other wavelengths. Green light counteracts blue light by inactivating CRY1 signaling, reducing anthocyanin levels in Arabidopsis and lettuce [142,143], whereas far-red light synergizes with blue light, increasing anthocyanin levels in microgreens (e.g., kale, broccoli, and beet) as the light intensity increases [144].
The effects of blue light are highly species-dependent and environmentally modulated, as evidenced by divergent responses across plant varieties and growth conditions. Lettuce (Lactuca sativa), for example, exhibits a wavelength-dependent anthocyanin hierarchy (505 nm > 455 nm > 470 nm > 535 nm) [145]. Short-term exposure reduces the nitrate content and increases the antioxidant content, whereas prolonged exposure triggers adaptive oxidative defenses [83]. In Brassica varieties, red light elevates anthocyanin in ‘Red Rookie’ cabbage but does not affect "Kinshun" [67]. Herbs such as coriander (Coriandrum sativum) exhibited a 40% increase in phenolic content under blue light, which improved their radical scavenging activity [29]. In contrast, basil and sweet peppers showed increased pigment and antioxidant levels [28]. In addition to pigmentation, blue light alters lipid metabolism, optimizing fatty acid composition in leaves and maximizing lipid content in fresh water microalgae Chlorella at 200 μmol m⁻² s⁻¹[146]. In ornamental species such as Rosa hybrida, blue light enhances the production of phenolic acids and flavonoids, thereby increasing antioxidant capacity [147]. In contrast, pea sprouts (Pisum sativum) exhibit increased phenolic and flavonoid contents under blue light, which is correlated with enhanced antioxidant activity [147].
Practical applications highlight the agricultural potential of blue light. In tea plants (Camellia sinensis), blue light during dark periods induces the CRY2/3, HY5, and MYB transcription factors, promoting anthocyanin and catechin accumulation [148]. Tailored spectral regimes, such as combining blue with far-red light, increase pigmentation in microgreens, whereas green light fine-tunes anthocyanin levels [144,145]. These findings highlight the complexity of light-regulated anthocyanin biosynthesis, underscoring the need for tailored spectral regimes in agriculture to optimize crop color, nutritional quality, and stress resilience, while accounting for photoreceptor crosstalk and species-specific traits.
7. Blue Light-Mediated Stress Resilience
Blue light plays a multifaceted role in enhancing plant resilience to environmental stresses, from abiotic challenges such as drought and salinity to biotic threats such as pathogen attacks. Its ability to modulate physiological, molecular, and biochemical pathways underscores its importance in sustainable agriculture, particularly in enhancing stress tolerance and crop productivity.
7.1. Drought Stress
Blue light can increase plant resilience to drought stress by modulating stomatal dynamics, enhancing antioxidant defenses, and activating stress-responsive pathways. Phytotropins regulate stomatal closure during water scarcity, minimizing water loss while maintaining CO₂ uptake for photosynthesis [149]. For example, in Arabidopsis, blue light upregulates CYCLIN H;1, a gene that suppresses reactive oxygen species (ROS) accumulation under drought, thereby enhancing osmotic balance and antioxidant capacity [150]. Cryptochromes further fine-tune these responses, as demonstrated in tomato (Solanum lycopersicum), where blue light fluence rates influence osmotic stress tolerance through cryptochrome-mediated signaling [151]
Pre-treatment strategies leveraging blue light have proven effective across species, such as lemon balm, lettuce, and rocket. Lemon balm (Melissa officinalis) exposed to combined blue light and red light presented increased root hydraulic conductivity and proline accumulation, bolstering water retention [152], whereas lettuce (Lactuca sativa) and rocket (Eruca sativa) pretreated with blue light presented improved water-use efficiency and reduced drought-induced wilting [153].
Blue light drives secondary metabolic adaptations under drought, such as the accumulation of flavonoids and hydroxycinnamates in wild privet (Ligustrum vulgare), which mitigates oxidative damage during combined excess light and drought stress [154]. Similarly, alfalfa (Medicago sativa) subjected to blue light-enriched regimes under drought conditions displays improved forage quality and physiological performance, which are linked to elevated antioxidant activity [155]. Chinese cabbage (Brassica rapa) exhibits enhanced carotenoid biosynthesis and ROS scavenging under prolonged blue light exposure [156]. Species-specific responses underscore the need for tailored approaches: sweet pepper (Capsicum annuum) benefits from blue light-enriched spectra through improved leaf physiology and reduced water-deficit vulnerability [157], whereas rose (Rosa × hybrida) leaves that develop under high humidity show improved stomatal function when exposed to blue light [158]. Festuca arundinacea adapts to drought through blue light-mediated adjustments in leaf elongation and stomatal transpiration [159]. These findings highlight the complex role of blue light in enhancing water conservation and stress resilience while emphasizing the necessity of calibrated application.
7.2. Salt Stress
Salinity stress, caused by elevated soil salinity or saline irrigation water, poses a significant threat to global crop productivity by triggering oxidative stress, osmotic imbalance, and ionic toxicity [160]. These disruptions lead to reduced photosynthetic activity, premature senescence, and programmed cell death, which threaten food security [161]. Recent studies highlight blue light as a critical regulator of plant salt tolerance, acting as a signaling cue that activates photomorphogenic and physiological adaptations to mitigate stress [162].
Blue light enhances salt tolerance by optimizing stomatal dynamics through phytotropin photoreceptors. These photoreceptors activate guard cell signaling pathways, promoting stomatal opening via K⁺ accumulation and water potential reduction, which sustains CO₂ uptake and photosynthesis under salt stress conditions [163]. For example, supplementing red light with blue light significantly improved stomatal function and CO₂ uptake in salt-stressed plants [164].
Blue light also regulates the transcriptional networks critical for stress adaptation. In wheat, blue light induces G-BOX BINDING FACTOR 1 (TaGBF1), a transcription factor that enhances abscisic acid sensitivity and ionic homeostasis under salinity [165]. Combining blue light with other wavelengths optimizes the stress response. While red light alone induces stomatal opening via guard cell chloroplast responses to intercellular CO₂ changes, blue light supplementation enhances this process under salt stress [164,166]. These results highlight the potential of spectral manipulation to counteract salt stress. To translate these insights into practice, future work should prioritize (1) optimizing the light spectrum for crop-specific needs, (2) elucidating stress-response gene regulatory networks, and (3) validating field applications under realistic agronomic conditions.
8. Conclusions
The use of blue light in plant science and controlled environment agriculture represents a significant advancement in optimizing crop production and resilience. Blue light regulates essential developmental and physiological processes, including morphogenesis, photosynthesis, flowering time, secondary metabolism, and stress adaptation, through its interaction with photoreceptors such as cryptochromes and phytotropins. These interactions improve functional traits and increase the biosynthesis of health-promoting phytochemicals. However, optimizing blue light application requires careful calibration specific to plant species and their growth stages to avoid potential trade-offs.
In controlled environment agriculture systems, blue light-enriched LED technologies have demonstrated remarkable potential in increasing crop yield, nutritional quality, and resilience to environmental stresses, which are key priorities for sustainable food systems. By precisely adjusting the intensity of blue light and exposure duration, growers can effectively manage flowering, optimize phytochemical profiles, and enhance growth efficiency. Future research should focus on interdisciplinary collaboration, integrating molecular biology, genomics, and horticultural engineering to better understand blue light driven regulatory networks, improve photoreceptor efficacy, and rigorously validate practical applications across diverse crop species. As global food demands continue to rise and climate challenges intensify, blue light has emerged as a critical tool in modern agriculture, facilitating the development of resource-efficient and nutrient-dense crop production systems.
Declaration of Competing Interest: The authors declare no conflict of interest.
Acknowledgments
This work was partially supported by USDA NIFA-SAS Project Award number: 2023-69012-39038 and Charles Carter Newman Endowment funds, Clemson University, Clemson, SC.
References
- Wilkinson, A.; Gerlach, C.; Karlsson, M.; Penn, H. Controlled environment agriculture and containerized food production in northern north america. J Agric Food Syst Co 2021, 10, 127-142. [CrossRef]
- Nelkin, J.; Caplow, T. Sustainable controlled environment agriculture for urban areas. Proceedings of the International Symposium on High Technology for Greenhouse System Management, Vols 1 and 2 2008, 449-455, . [CrossRef]
- Takakura, T. Controlled environment agriculture. Xi International Congress - the Use of Plastics in Agriculture, Vol 1 1990, F3-F10.
- Kotilainen, T.; Robson, T.M.; Hernández, R. Light quality characterization under climate screens and shade nets for controlled-environment agriculture. Plos One 2018, 13. [CrossRef]
- Pocock, T. Advanced lighting technology in controlled environment agriculture. Lighting Res Technol 2016, 48, 83-94. [CrossRef]
- Choi, H.G.; Moon, B.Y.; Kang, N.J. Effects of led light on the production of strawberry during cultivation in a plastic greenhouse and in a growth chamber. Sci Hortic-Amsterdam 2015, 189, 22-31. [CrossRef]
- Al Murad, M.; Razi, K.; Jeong, B.R.; Samy, P.M.A.; Muneer, S. Light emitting diodes (leds) as agricultural lighting: Impact and its potential on improving physiology, flowering, and secondary metabolites of crops. Sustainability-Basel 2021, 13. [CrossRef]
- Saebo, A.; Krekling, T.; Appelgren, M. Light quality affects photosynthesis and leaf anatomy of birch plantlets in-vitro. Plant Cell Tiss Org 1995, 41, 177-185. [CrossRef]
- Sumida, S.; Ehara, T.; Osafune, T.; Ohkuro, I.; Hase, E. Effects of blue-light on chloroplast development in dark-grown euglena-gracilis z. J. Electron Microsc. 1984, 33, 304-305.
- Zeiger, E. Light perception in guard-cells. Plant Cell Environ 1990, 13, 739-744. [CrossRef]
- Senger, H. The effect of blue-light on plants and microorganisms. Photochemistry and Photobiology 1982, 35, 911-920. [CrossRef]
- Cosgrove, D.J. Rapid suppression of growth by blue-light - occurrence, time course, and general-characteristics. Plant Physiol. 1981, 67, 584-590. [CrossRef]
- Talbott, L.D.; Hammad, J.W.; Harn, L.C.; Nguyen, V.H.; Patel, J.; Zeiger, E. Reversal by green light of blue light-stimulated stomatal opening in intact, attached leaves of arabidopsis operates only in the potassium-dependent, morning phase of movement. PCPhy 2006, 47, 332-339. [CrossRef]
- Frechilla, S.; Talbott, L.D.; Bogomolni, R.A.; Zeiger, E. Reversal of blue light-stimulated stomatal opening by green light. PCPhy 2000, 41, 171-176.
- Yang, J.; Song, J.N.; Jeong, B.R. Blue light supplemented at intervals in long-day conditions intervenes in photoperiodic flowering, photosynthesis, and antioxidant properties in chrysanthemums. Antioxidants-Basel 2022, 11. [CrossRef]
- Yang, J.; Song, J.; Jeong, B.R. Low-intensity blue light supplemented during photoperiod in controlled environment induces flowering and antioxidant production in kalanchoe. Antioxidants-Basel 2022, 11. [CrossRef]
- Yang, J.; Song, J.; Jeong, B.R. Lighting from top and side enhances photosynthesis and plant performance by improving light usage efficiency. Int J Mol Sci 2022, 23. [CrossRef]
- Yang, J.; Jeong, B.R. Side lighting enhances morphophysiology by inducing more branching and flowering in chrysanthemum grown in controlled environment. Int J Mol Sci 2021, 22. [CrossRef]
- Yang, J.L.; Song, J.N.; Jeong, B.R. Flowering and runnering of seasonal strawberry under different photoperiods are affected by intensity of supplemental or night-interrupting blue light. Plants-Basel 2024, 13. [CrossRef]
- Jones, M.A. Using light to improve commercial value. Hortic Res-England 2018, 5. [CrossRef]
- Mccree, K.J. Action spectrum, absorptance and quantum yield of photosynthesis in crop plants. Agr. Meteorol. 1972, 9, 191-&. [CrossRef]
- Li, J.G.; Terzaghi, W.; Deng, X.W. Genomic basis for light control of plant development. Protein Cell 2012, 3, 106-116. [CrossRef]
- Cashmore, A.R.; Jarillo, J.A.; Wu, Y.J.; Liu, D.M. Cryptochromes: Blue light receptors for plants and animals. Science 1999, 284, 760-765. [CrossRef]
- Huché-Thélier, L.; Crespel, L.; Le Gourrierec, J.; Morel, P.; Sakr, S.; Leduc, N. Light signaling and plant responses to blue and uv radiations-perspectives for applications in horticulture. Environ. Exp. Bot. 2016, 121, 22-38. [CrossRef]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of grown under different combinations of red and blue light. Journal of Experimental Botany 2010, 61, 3107-3117. [CrossRef]
- Zheng, Y.J.; Zhang, Y.T.; Liu, H.C.; Li, Y.M.; Liu, Y.L.; Hao, Y.W.; Lei, B.F. Supplemental blue light increases growth and quality of greenhouse pak choi depending on cultivar and supplemental light intensity. J Integr Agr 2018, 17, 2245-2256. [CrossRef]
- Yanagi, T.; Okamoto, K.; Takita, S. Effects of blue, red, and blue/red lights of two different ppf levels on growth and morphogenesis of lettuce plants. International Symposium on Plant Production in Closed Ecosystems - Automation, Culture, and Environment 1997, 117-122.
- Naznin, M.T.; Lefsrud, M.; Gravel, V.; Azad, M.O.K. Blue light added with red leds enhance growth characteristics, pigments content, and antioxidant capacity in lettuce, spinach, kale, basil, and sweet pepper in a controlled environment. Plants-Basel 2019, 8. [CrossRef]
- Nguyen, D.T.P.; Kitayama, M.; Lu, N.; Takagaki, M. Improving secondary metabolite accumulation, mineral content, and growth of coriander (coriandrum sativum l.) by regulating light quality in a plant factory. J. Horticult. Sci. Biotechnol. 2020, 95, 356-363. [CrossRef]
- Appelgren, M. Effects of light quality on stem elongation of pelargonium invitro. Sci Hortic-Amsterdam 1991, 45, 345-351. [CrossRef]
- Wheeler, R.M.; Mackowiak, C.L.; Sager, J.C. Soybean stem growth under high-pressure sodium with supplemental blue lighting. Agron. J. 1991, 83, 903-906. [CrossRef]
- Brown, C.S.; Schuerger, A.C.; Sager, J.C. Growth and photomorphogenesis of pepper plants under red light-emitting-diodes with supplemental blue or far-red lighting. J. Am. Soc. Hort. Sci. 1995, 120, 808-813. [CrossRef]
- Lee, S.W.; Seo, J.M.; Lee, M.K.; Chun, J.H.; Antonisamy, P.; Arasu, M.V.; Suzuki, T.; Al-Dhabi, N.A.; Kim, S.J. Influence of different led lamps on the production of phenolic compounds in common and tartary buckwheat sprouts. Ind Crop Prod 2014, 54, 320-326. [CrossRef]
- Hernández, R.; Kubota, C. Physiological responses of cucumber seedlings under different blue and red photon flux ratios using leds. Environ. Exp. Bot. 2016, 121, 66-74. [CrossRef]
- Tymoszuk, A.; Kulus, D.; Błażejewska, A.; Nadolna, K.; Kulpińska, A.; Pietrzykowski, K. Application of wide-spectrum light-emitting diodes in the indoor production of cucumber and tomato seedlings. Acta Agro 2023, 76. [CrossRef]
- Kong, Y.; Zheng, Y.B. Magic blue light: A versatile mediator of plant elongation. Plants-Basel 2024, 13. [CrossRef]
- Li, C.X.; Xu, Z.G.; Dong, R.Q.; Chang, S.X.; Wang, L.Z.; Khalil-Ur-Rehman, M.; Tao, J.M. An rna-seq analysis of grape plantlets grown in vitro reveals different responses to blue, green, red led light, and white fluorescent light. Front Plant Sci 2017, 8. [CrossRef]
- Lim, Y.J.; Kwon, S.J.; Eom, S.H. Red and blue light-specific metabolic changes in soybean seedlings. Front Plant Sci 2023, 14. [CrossRef]
- Hahn, E.J.; Kozai, T.; Paek, K.Y. Combinations of blue and red leds increase the morphophysiological performance and furanocoumarin production of brosimum gaudichaudii trécul in vitro. Front. Plant Sci. 2021, 680545.
- Rabara, R.C.; Behrman, G.; Timbol, T.; Rushton, P.J. Effect of spectral quality of monochromatic led lights on the growth of artichoke seedlings. Front Plant Sci 2017, 8. [CrossRef]
- Li, K.; Ji, L.Y.; Xing, Y.Y.; Zuo, Z.C.; Zhang, L. Data-independent acquisition proteomics reveals the effects of red and blue light on the growth and development of moso bamboo (phyllostachys edulis) seedlings. Int J Mol Sci 2023, 24. [CrossRef]
- Kochetova, G.V.; Avercheva, O.V.; Bassarskaya, E.M.; Kushunina, M.A.; Zhigalova, T.V. Effects of red and blue led light on the growth and photosynthesis of barley (hordeum vulgare l.) seedlings. J. Plant Growth Regul. 2023, 42, 1804-1820. [CrossRef]
- Wan, Y.L.; Wu, Y.; Zhang, M.; Hong, A.Y.; Liu, Y. Effects of photoperiod extension via red-blue light-emitting diodes and high-pressure sodium lamps on the growth and photosynthetic characteristics in. Acta Physiol Plant 2020, 42. [CrossRef]
- Gao, Q.; Liao, Q.H.; Li, Q.M.; Yang, Q.C.; Wang, F.; Li, J.M. Effects of led red and blue light component on growth and photosynthetic characteristics of coriander in plant factory. Horticulturae 2022, 8. [CrossRef]
- Wollaeger, H.M.; Runkle, E.S. Growth of impatiens, petunia, salvia, and tomato seedlings under blue, green, and red light-emitting diodes. HortScience 2014, 49, 734-740. [CrossRef]
- Wollaeger, H.M.; Runkle, E.S. Growth and acclimation of impatiens, salvia, petunia, and tomato seedlings to blue and red light. HortScience 2015, 50, 522-529. [CrossRef]
- Correia, C.; Magnani, F.; Pastore, C.; Cellini, A.; Donati, I.; Pennisi, G.; Paucek, I.; Orsini, F.; Vandelle, E.; Santos, C.; et al. Red and blue light differently influence actinidia chinensis performance and its interaction with pseudomonas syringae pv. Actinidiae. Int J Mol Sci 2022, 23. [CrossRef]
- Zhao, X.Q.; Niu, Y.N.; Hossain, Z.; Zhao, B.Y.; Bai, X.D.; Mao, T.T. New insights into light spectral quality inhibits the plasticity elongation of maize mesocotyl and coleoptile during seed germination. Front Plant Sci 2023, 14. [CrossRef]
- Hu, J.W.; Dai, X.; Sun, G.Y. Morphological and physiological responses of morus alba seedlings under different light qualities. Not Bot Horti Agrobo 2016, 44, 382-392. [CrossRef]
- Snowden, M.C.; Cope, K.R.; Bugbee, B. Sensitivity of seven diverse species to blue and green light: Interactions with photon flux. Plos One 2016, 11. [CrossRef]
- Chen, C.C.; Huang, M.Y.; Lin, K.H.; Wong, S.L.; Huang, W.D.; Yang, C.M. Effects of light quality on the growth, development and metabolism of rice seedlings (oryza sativa l.). Res J Biotechnol 2014, 9, 15-24.
- Ren, M.F.; Liu, S.Z.; Tang, C.Z.; Mao, G.L.; Gai, P.P.; Guo, X.L.; Zheng, H.B.; Tang, Q.Y. Photomorphogenesis and photosynthetic traits changes in rice seedlings responding to red and blue light. Int J Mol Sci 2023, 24. [CrossRef]
- Akbarian, B.; Matloobi, M.; Mahna, N. Effects of led light on seed emergence and seedling quality of four bedding flowers. J. Ornam. Hortic. Plants 2016, 6, 115-123.
- Kong, Y.; Schiestel, K.; Zheng, Y.B. Pure blue light effects on growth and morphology are slightly changed by adding low-level uva or far-red light: A comparison with red light in four microgreen species. Environ. Exp. Bot. 2019, 157, 58-68. [CrossRef]
- Kong, Y.; Schiestel, K.; Zheng, Y.B. Maximum elongation growth promoted as a shade-avoidance response by blue light is related to deactivated phytochrome: A comparison with red light in four microgreen species. Can J Plant Sci 2020, 100, 314-326. [CrossRef]
- Kong, Y.; Kamath, D.; Zheng, Y.B. Blue versus red light can promote elongation growth independent of photoperiod: A study in four brassica microgreens species. HortScience 2019, 54, 1955-1961. [CrossRef]
- Johnson, R.E.; Kong, Y.; Zheng, Y.B. Elongation growth mediated by blue light varies with light intensities and plant species: A comparison with red light in arugula and mustard seedlings. Environ. Exp. Bot. 2020, 169. [CrossRef]
- Kong, Y.; Schiestel, K.; Zheng, Y. Blue light associated with low phytochrome activity can promote flowering: A comparison with red light in four bedding plant specie. Acta Hortic. 2020, 1296, 621-628.
- Kong, Y.; Kamath, D.; Zheng, Y. Blue-light-promoted elongation and flowering are not artifacts from 24-h lighting: A comparison with red light in four bedding plant specie. Acta Hortic. 2020, 1295, 659-666.
- Kong, Y.; Masabni, J.; Niu, G. Effect of temperature variation and blue and red leds on the elongation of arugula and mustard microgreens. Horticulturae 2023. [CrossRef]
- Hirai, T.; Amaki, W.; Watanabe, H. Action of blue or red monochromatic light on stem internodal growth depends on plant species. Sci. Hortic. 2006, 109995.
- Di, Q.H.; Li, J.; Du, Y.F.; Wei, M.; Shi, Q.H.; Li, Y.; Yang, F.J. Combination of red and blue lights improved the growth and development of eggplant (solanum melongena l.) seedlings by regulating photosynthesis. J. Plant Growth Regul. 2021, 40, 1477-1492. [CrossRef]
- Awasthi, K. Effect of different light on the growth and development of pea plant. Int. J. Res. Eng. Sci. 2023, 94-98.
- Hata, N.; Hayashi, Y.; Ono, E.; Satake, H.; Kobayashi, A.; Muranaka, T.; Okazawa, A. Differences in plant growth and leaf sesamin content of the lignan-rich sesame variety 'gomazou' under continuous light of different wavelengths. Plant Biotechnol. 2013, 30, 1-U104. [CrossRef]
- Spaninks, K.; Lamers, G.; van Lieshout, J.; Offringa, R. Light quality regulates apical and primary radial growth of arabidopsis thaliana and solanum lycopersicum. Sci Hortic-Amsterdam 2023, 317. [CrossRef]
- Kong, Y.; Zheng, Y.B. Growth and morphology responses to narrow-band blue light and its co-action with low-level uvb or green light: A comparison with red light in four microgreen species. Environ. Exp. Bot. 2020, 178. [CrossRef]
- Mizuno, T.; Amaki, W.; Watanabe, H. Effects of monochromatic light irradiation by led on the growth and anthocyanin contents in leaves of cabbage seedlings. Vi International Symposium on Light in Horticulture 2011, 907, 179-184. [CrossRef]
- Liu, X.Y.; Chang, T.T.; Guo, S.R.; Xu, Z.G.; Li, J. Effect of different light quality of led on growth and photosynthetic character in cherry tomato seedling. Vi International Symposium on Light in Horticulture 2011, 907, 325-330.
- Liu, X.; Guo, S.; Chang, T.; Xu, Z.; Tezuka, T. Regulation of the growth and photosynthesis of cherry tomato seedlings by different light irradiations of light emitting diodes (led). Afr. J. Biotechnol. 2012, 11, 6169–6177.
- Kim, E.Y.; Park, S.A.; Park, B.J.; Lee, Y.; Oh, M.M. Growth and antioxidant phenolic compounds in cherry tomato seedlings grown under monochromatic light-emitting diodes. Hortic Environ Biote 2014, 55, 506-513. [CrossRef]
- Hernández, R.; Eguchi, T.; Kubota, C. Growth and morphology of vegetable seedlings under different blue and red photon flux ratios using light-emitting diodes as sole-source lighting. Viii International Symposium on Light in Horticulture 2016, 1134, 195-200. [CrossRef]
- Liang, Y.; Kang, C.Q.; Kaiser, E.; Kuang, Y.; Yang, Q.C.; Li, T. Red/blue light ratios induce morphology and physiology alterations differently in cucumber and tomato. Sci Hortic-Amsterdam 2021, 281. [CrossRef]
- Wojciechowska, R.; Dabrowa, A.; Kolton, A. How monochromatic and composed light affect the kale 'scarlet' in its initial growth stage. Acta Sci Pol-Hortoru 2023, 22, 93-100. [CrossRef]
- Meng, Q.W.; Runkle, E.S. Growth responses of red-leaf lettuce to temporal spectral changes. Front Plant Sci 2020, 11. [CrossRef]
- Kong, Y.; Schiestel, K.; Zheng, Y. Does “blue” light invariably cause plant compactness? Not really: A comparison with red light in four bedding plant species during the transplant stage. Acta Hortic. 2020, 621-628.
- Brazaityte, A.; Miliauskiene, J.; Vastakaite-Kairiene, V.; Sutuliene, R.; Lauzike, K.; Duchovskis, P.; Malek, S. Effect of different ratios of blue and red led light on brassicaceae microgreens under a controlled environment. Plants-Basel 2021, 10. [CrossRef]
- Nie, W.F.; Li, Y.; Chen, Y.; Zhou, Y.; Yu, T.; Zhou, Y.H.; Yang, Y.X. Spectral light quality regulates the morphogenesis, architecture, and flowering in pepper (capsicum annuum l.). J Photoch Photobio B 2023, 241. [CrossRef]
- Li, Y.; Xin, G.F.; Shi, Q.H.; Yang, F.J.; Wei, M. Response of photomorphogenesis and photosynthetic properties of sweet pepper seedlings exposed to mixed red and blue light. Front Plant Sci 2023, 13. [CrossRef]
- Lim, M.J.; Murthy, H.N.; Song, H.Y.; Lee, S.Y.; Park, S.Y. Influence of white, red, blue, and combination of led lights on in vitro multiplication of shoots, rooting, and acclimatization of gerbera jamesonii cv. ‘Shy pink’ plants. Agronomy-Basel 2023, 13. [CrossRef]
- Lin, K.H.; Chen, Y.C.; Wu, Q.E.; Lin, H.H. Effects of red and blue light ratio on the morphological traits and flower sex expression in cucurbita moschata duch. Not Bot Horti Agrobo 2023, 51. [CrossRef]
- Izzo, L.G.; Mele, B.H.; Vitale, L.; Vitale, E.; Arena, C. The role of monochromatic red and blue light in tomato early photomorphogenesis and photosynthetic traits. Environ. Exp. Bot. 2020, 179. [CrossRef]
- Zhang, X.; Bisbis, M.; Heuvelink, E.; Jiang, W.J.; Marcelis, L.F.M. Green light reduces elongation when partially replacing sole blue light independently from cryptochrome 1a. Physiol. Plant. 2021, 173, 1946-1955. [CrossRef]
- Bian, Z.H.; Cheng, R.F.; Yang, Q.C.; Wang, J.; Lu, C.G. Continuous light from red, blue, and green light-emitting diodes reduces nitrate content and enhances phytochemical concentrations and antioxidant capacity in lettuce. J. Am. Soc. Hort. Sci. 2016, 141, 186-U216. [CrossRef]
- Taulavuori, K.; Hyöky, V.; Oksanen, J.; Taulavuori, E.; Julkunen-Tiitto, R. Species-specific differences in synthesis of flavonoids and phenolic acids under increasing periods of enhanced blue light. Environ. Exp. Bot. 2016, 121, 145-150. [CrossRef]
- Son, K.H.; Oh, M.M. Growth, photosynthetic and antioxidant parameters of two lettuce cultivars as affected by red, green, and blue light-emitting diodes. Hortic Environ Biote 2015, 56, 639-653. [CrossRef]
- Johkan, M.; Shoji, K.; Goto, F.; Hashida, S.; Yoshihara, T. Blue light-emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. HortScience 2010, 45, 1809-1814. [CrossRef]
- Heo, J.; Lee, C.; Chakrabarty, D.; Paek, K. Growth responses of marigold and salvia bedding plants as affected by monochromic or mixture radiation provided by a light-emitting diode (led). Plant Growth Regul 2002, 38, 225-230. [CrossRef]
- Samuoliene, G.; Brazaityte, A.; Urbonaviciute, A.; Sabajeviene, G.; Duchovskis, P. The effect of red and blue light component on the growth and development of frigo strawberries. Zemdirbyste 2010, 97, 99-104.
- Gangadhar, B.H.; Mishra, R.K.; Pandian, G.; Park, S.W. Comparative study of color, pungency, and biochemical composition in chili pepper (capsicum annuum) under different light-emitting diode treatments. HortScience 2012, 47, 1729-1735. [CrossRef]
- Matsuda, R.; Ohashi-Kaneko, K.; Fujiwara, K.; Kurata, K. Analysis of the relationship between blue-light photon flux density and the photosynthetic properties of spinach (l.) leaves with regard to the acclimation of photosynthesis to growth irradiance. Soil Sci. Plant Nutr. 2007, 53, 459-465. [CrossRef]
- Yorio, N.C.; Goins, G.D.; Kagie, H.R.; Wheeler, R.M.; Sager, J.C. Improving spinach, radish, and lettuce growth under red light-emitting diodes (leds) with blue light supplementation. HortScience 2001, 36, 380-383. [CrossRef]
- Heo, J.W.; Kang, D.H.; Bang, H.S.; Hong, S.G.; Chun, C.; Kang, K.K. Early growth, pigmentation, protein content, and phenylalanine ammonia-lyase activity of red curled lettuces grown under different lighting conditions. Korean J Hortic Sci 2012, 30, 6-12. [CrossRef]
- Kalaitzoglou, P.; Taylor, C.; Calders, K.; Hogervorst, M.; van Ieperen, W.; Harbinson, J.; de Visser, P.; Nicole, C.C.S.; Marcelis, L.F.M. Unraveling the effects of blue light in an artificial solar background light on growth of tomato plants. Environ. Exp. Bot. 2021, 184. [CrossRef]
- Terashima, I.; Fujita, T.; Inoue, T.; Chow, W.S.; Oguchi, R. Green light drives leaf photosynthesis more efficiently than red light in strong white light: Revisiting the enigmatic question of why leaves are green. PCPhy 2009, 50, 684-697. [CrossRef]
- Kim, H.H.; Wheeler, R.M.; Sager, J.C.; Goins, G.D. A comparison of growth and photosynthetic characteristics of lettuce grown under red and blue light-emitting diodes (leds) with and without supplemental green leds. Proceedings of the Viith International Symposium on Protected Cultivation in Mild Winter Climates: Production, Pest Management and Global Competition, Vols 1 and 2 2004, 467-475. [CrossRef]
- Kim, H.H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Green-light supplementation for enhanced lettuce growth under red- and blue-light-emitting diodes. HortScience 2004, 39, 1617-1622. [CrossRef]
- Ouzounis, T.; Heuvelink, E.; Ji, Y.; Schouten, H.J.; Visser, R.G.F.; Marcelis, L.F.M. Blue and red led lighting effects on plant biomass, stomata! Conductance, and metabolite content in nine tomato genotypes. Viii International Symposium on Light in Horticulture 2016, 1134, 251-258. [CrossRef]
- Pashkovskiy, P.P.; Kartashov, A.V.; Zlobin, I.E.; Pogosyan, S.I.; Kuznetsov, V.V. Blue light alters mir167 expression and microrna-targeted auxin response factor genes in arabidopsis thaliana plants. Plant Physiol. Biochem. 2016, 104, 146-154. [CrossRef]
- Miao, Y.X.; Chen, Q.Y.; Qu, M.; Gao, L.H.; Hou, L.P. Blue light alleviates 'red light syndrome' by regulating chloroplast ultrastructure, photosynthetic traits and nutrient accumulation in cucumber plants. Sci Hortic-Amsterdam 2019, 257. [CrossRef]
- Canamero, R.C.; Bakrim, N.; Bouly, J.P.; Garay, A.; Dudkin, E.E.; Habricot, Y.; Ahmad, M. Cryptochrome photoreceptors cry1 and cry2 antagonistically regulate primary root elongation in. Planta 2006, 224, 995-1003. [CrossRef]
- Vandenbrink, J.P.; Herranz, R.; Medina, F.J.; Edelmann, R.E.; Kiss, J.Z. A novel blue-light phototropic response is revealed in roots of arabidopsis thalianain microgravity. Planta 2016, 244, 1201-1215. [CrossRef]
- Iacona, C.; Muleo, R. Light quality affects adventitious rooting and performance of cherry rootstock colt. Sci Hortic-Amsterdam 2010, 125, 630-636. [CrossRef]
- Jin, D.Z.; Su, X.F.; Li, Y.F.; Shi, M.M.; Yang, B.B.; Wan, W.C.; Wen, X.; Yang, S.J.; Ding, X.T.; Zou, J. Effect of red and blue light on cucumber seedlings grown in a plant factory. Horticulturae 2023, 9. [CrossRef]
- Zhai, S.; Cai, W.; Xiang, Z.X.; Chen, C.Y.; Lu, Y.T.; Yuan, T.T. Pin3-mediated auxin transport contributes to blue light-induced adventitious root formation in arabidopsis. Plant Sci. 2021, 312. [CrossRef]
- Heo, J.W.; Lee, C.W.; Paek, K.Y. Influence of mixed led radiation on the growth of annual plants. J Plant Biol 2006, 49, 286-290. [CrossRef]
- Mckay, M.; Faust, J.E.; Taylor, M.; Adelberg, J. The effects of blue light and supplemental far-red on an in vitro multiple harvest system for the production of cannabis sativa. Plants-Basel 2025, 14. [CrossRef]
- Vince-Prue, D.; Canham, A.E. Horticultural significance of photomorphogenesis. Encyclopedia of Plant Physiol 1983, NS 16B, 518-544.
- Park, Y.G.; Jeong, B.R. Night interruption light quality changes morphogenesis, flowering, and gene expression in dendranthema grandiflorum. Hortic Environ Biote 2019, 60, 167-173. [CrossRef]
- Park, Y.G.; Jeong, B.R. How supplementary or night-interrupting low-intensity blue light affects the flower induction in chrysanthemum, a qualitative short-day plant. Plants-Basel 2020, 9. [CrossRef]
- Park, Y.G.; Jeong, B.R. Both the quality and positioning of the night interruption light are important for flowering and plant extension growth. J. Plant Growth Regul. 2020, 39, 583-593. [CrossRef]
- Park, Y.G.; Jeong, B.R. The quality and quality shifting of the night interruption light 2022.
- Park, Y.G.; Jeong, B.R. Shift in the light quality of night interruption affects flowering and morphogenesis of petunia hybrida. Plants-Basel 2023, 12. [CrossRef]
- Park, Y.G.; Jeong, B.R. Photoreceptors modulate the flowering and morphogenesis responses of pelargonium × hortorum to night-interruption light quality shifting. Agronomy-Basel 2023, 13. [CrossRef]
- Park, Y.G.; Muneer, S.; Jeong, B.R. Morphogenesis, flowering, and gene expression of dendranthema grandiflorum in response to shift in light quality of night interruption. Int J Mol Sci 2015, 16, 16497-16513. [CrossRef]
- Park, Y.G.; Muneer, S.; Soundararajan, P.; Manivnnan, A.; Jeong, B.R. Light quality during night interruption affects morphogenesis and flowering in petunia hybrida , a qualitative long-day plant. Hortic Environ Biote 2016, 57, 371-377. [CrossRef]
- Park, Y.G.; Muneer, S.; Soundararajan, P.; Manivnnan, A.; Jeong, B.R. Light quality during night interruption affects morphogenesis and flowering in geranium. Hortic Environ Biote 2017, 58, 212-217. [CrossRef]
- Yamada, A.; Tanigawa, T.; Suyama, T.; Matsuno, T.; Kunitake, T. Night break treatment using different light sources promotes or delays growth and flowering of (raf.) shinn. J Jpn Soc Hortic Sci 2008, 77, 69-74. [CrossRef]
- Pérez, C.P.; Ulrichs, C.; Huyskens-Keil, S.; Schreiner, M.; Krumbein, A.; Schwarz, D.; Kläring, H.P. Composition of carotenoids in tomato fruits as affected by moderate uv-b radiation before harvest. International Symposium on Tomato in the Tropics 2009, 821, 217-221. [CrossRef]
- Kong, Y.; Zheng, Y.B. Diverse flowering response to blue light manipulation: Application of electric lighting in controlled-environment plant production. Horticulturae 2024, 10. [CrossRef]
- Jackson, S.D. Plant responses to photoperiod. New Phytol. 2009, 181, 517-531. [CrossRef]
- Walters, K.J.; Hurt, A.A.; Lopez, R.G. Flowering, stem extension growth, and cutting yield of foliage annuals in response to photoperiod. HortScience 2019, 54, 661-666. [CrossRef]
- Chandel, A.; Thakur, M.; Singh, G.; Dogra, R.; Bajad, A.; Soni, V.; Bhargava, B. Flower regulation in floriculture: An agronomic concept and commercial use. J. Plant Growth Regul. 2023, 42, 2136-2161. [CrossRef]
- Izawa, T. Daylength measurements by rice plants in photoperiodic short-day flowering. International Review of Cytology - a Survey of Cell Biology, Vol 256 2007, 256, 191-+. [CrossRef]
- Kobayashi, Y.; Kaya, H.; Goto, K.; Iwabuchi, M.; Araki, T. A pair of related genes with antagonistic roles in mediating flowering signals. Science 1999, 286, 1960-1962. [CrossRef]
- Whitman, C.M.; Heins, R.D.; Cameron, A.C.; Carlson, W.H. Lamp type and irradiance level for daylength extensions influence flowering of 'blue clips', 'early sunrise', and 'moonbeam'. J. Am. Soc. Hort. Sci. 1998, 123, 802-807. [CrossRef]
- Meng, Q.; Runkle, E.S. Low-intensity blue light in night-interruption lighting does not influence flowering of herbaceous ornamentals. Sci Hortic-Amsterdam 2015, 186, 230-238.
- Lopez, R.G.; Meng, Q.W.; Runkle, E.S. Blue radiation signals and saturates photoperiodic flowering of several long-day plants at crop-specific photon flux densities. Sci Hortic-Amsterdam 2020, 271. [CrossRef]
- Kang, D.I.; Jeong, H.K.; Park, Y.G.; Jeong, B.R. Flowering and morphogenesis of kalanchoe in response to quality and intensity of night interruption light. Plants-Basel 2019, 8. [CrossRef]
- Nissim-Levi, A.; Kitron, M.; Nishri, Y.; Ovadia, R.; Forer, I.; Oren-Shamir, M. Effects of blue and red led lights on growth and flowering of. Sci Hortic-Amsterdam 2019, 254, 77-83. [CrossRef]
- Magar, Y.G.; Ohyama, K.; Noguchi, A.; Amaki, W.; Furufuji, S. Effects of light quality during supplemental lighting on the flowering in an everbearing strawberry. Xiii International Symposium on Plant Bioregulators in Fruit Production 2018, 1206, 279-284. [CrossRef]
- Cho, H.Y.; Kadowaki, M.; Che, J.; Takahashi, S.; Horiuchi, N.; Ogiwara, I. Influence of light quality on flowering characteristics, potential for year-round fruit production and fruit quality of blueberry in a plant factory. Fruits 2019, 74, 3-10. [CrossRef]
- Javanmardi, J.; and Emami, S. Response of tomato and pepper transplants to light spectra provided by light emitting diodes. International Journal of Vegetable Science 2013, 19, 138-149. [CrossRef]
- Zhu, J.; Zhang, Y.C.; Zhou, L.; Yang, L.Y. Growth and flowering of saffron (crocus sativus l.) with three corm weights under different led light qualities. Sci Hortic-Amsterdam 2022, 303. [CrossRef]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.O. Spectral effects of artificial light on plant physiology and secondary metabolism: A review. HortScience 2015, 50, 1128-1135. [CrossRef]
- Li, H.S.; Lyu, Y.M.; Chen, X.H.; Wang, C.Q.; Yao, D.H.; Ni, S.S.; Lin, Y.L.; Chen, Y.K.; Zhang, Z.H.; Lai, Z.X. Exploration of the effect of blue light on functional metabolite accumulation in longan embryonic calli via rna sequencing. Int J Mol Sci 2019, 20. [CrossRef]
- Li, H.S.; Lin, Y.L.; Chen, X.H.; Bai, Y.; Wang, C.Q.; Xu, X.P.; Wang, Y.; Lai, Z.X. Effects of blue light on flavonoid accumulation linked to the expression of mir393, mir394 and mir395 in longan embryogenic calli. Plos One 2018, 13. [CrossRef]
- Deng, L.L.; Yuan, Z.Y.; Xie, J.; Yao, S.X.; Zeng, K.F. Sensitivity to ethephon degreening treatment is altered by blue led light irradiation in mandarin fruit. Journal of Agricultural and Food Chemistry 2017, 65, 6158-6168. [CrossRef]
- Yuan, Z.Y.; Deng, L.L.; Yin, B.F.; Yao, S.X.; Zeng, K.F. Effects of blue led light irradiation on pigment metabolism of ethephon-degreened mandarin fruit. Postharvest Biol. Technol. 2017, 134, 45-54. [CrossRef]
- Zhang, L.C.; Ma, G.; Yamawaki, K.; Ikoma, Y.; Matsumoto, H.; Yoshioka, T.; Ohta, S.; Kato, M. Effect of blue led light intensity on carotenoid accumulation in citrus juice sacs. J. Plant Physiol. 2015, 188, 58-63. [CrossRef]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59-64. [CrossRef]
- Ahmad, M.; Cashmore, A.R. The blue-light receptor cryptochrome 1 shows functional dependence on phytochrome a or phytochrome b in arabidopsis thaliana. Plant J. 1997, 11, 421-427. [CrossRef]
- Folta, K.M.; Maruhnich, S.A. Green light: A signal to slow down or stop. Journal of Experimental Botany 2007, 58, 3099-3111. [CrossRef]
- Zhang, T.T.; Folta, K.M. Green light signaling and adaptive response. Plant Signal Behav 2012, 7, 75-78. [CrossRef]
- Kopsell, D.A.; Sams, C.E.; Barickman, T.C.; Morrow, R.C. Sprouting broccoli accumulate higher concentrations of nutritionally important metabolites under narrow-band light-emitting diode lighting. J. Am. Soc. Hort. Sci. 2014, 139, 469-477. [CrossRef]
- Samuoliene, G.; Sirtautas, R.; Brazaityte, A.; Duchovskis, P. Led lighting and seasonality effects antioxidant properties of baby leaf lettuce. Food Chem. 2012, 134, 1494-1499. [CrossRef]
- Atta, M.; Idris, A.; Bukhari, A.; Wahidin, S. Intensity of blue led light: A potential stimulus for biomass and lipid content in fresh water microalgae. Bioresour. Technol. 2013, 148, 373-378. [CrossRef]
- Zhao, T.Y.; Nie, J.W.; Yan, X.Y.; Xue, W.T. Identifying the critical led light condition for optimum yield and flavonoid of pea sprouts. Sci Hortic-Amsterdam 2024, 327. [CrossRef]
- Zheng, C.; Ma, J.Q.; Ma, C.L.; Shen, S.Y.; Liu, Y.F.; Chen, L. Regulation of growth and flavonoid formation of tea plants () by blue and green light. Journal of Agricultural and Food Chemistry 2019, 67, 2408-2419. [CrossRef]
- Inoue, S.; Kinoshita, T. Blue light regulation of stomatal opening and the plasma membrane h(+)-atpase. Plant Physiol. 2017, 174, 531-538. [CrossRef]
- Zhou, X.F.; Jin, Y.H.; Yoo, C.Y.; Lin, X.L.; Kim, W.Y.; Yun, D.J.; Bressan, R.A.; Hasegawa, P.M.; Jin, J.B. Cyclin h;1 regulates drought stress responses and blue light-induced stomatal opening by inhibiting reactive oxygen species accumulation in arabidopsis. Plant Physiol. 2013, 162, 1030-1041. [CrossRef]
- D'Amico-Damiao, V.; Lúcio, J.C.B.; Oliveira, R.; Gaion, L.A.; Barreto, R.F.; Carvalho, R.F. Cryptochrome 1a depends on blue light fluence rate to mediate osmotic stress responses in tomato. J. Plant Physiol. 2021, 258. [CrossRef]
- Ahmadi, T.; Shabani, L.; Sabzalian, M.R. Improvement in drought tolerance of lemon balm, melissa officinalis l. Under the pre-treatment of led lighting. Plant Physiol. Biochem. 2019, 139, 548-557. [CrossRef]
- Ginzburg, D.N.; Klein, J.D. Led pre-exposure shines a new light on drought tolerance complexity in lettuce (lactuca sativa) and rocket (eruca sativa). Environ. Exp. Bot. 2020, 180. [CrossRef]
- Tattini, M.; Galardi, C.; Pinelli, P.; Massai, R.; Remorini, D.; Agati, G. Differential accumulation of flavonoids and hydroxycinnamates in leaves of ligustrum vulgare under excess light and drought stress. New Phytol. 2004, 163, 547-561. [CrossRef]
- He, C.X.; Zhao, Y.; Wang, Y.; Cai, J.F.; Gao, J.; Zhang, J.S. Forage quality and physiological performance of mowed alfalfa (medicago sativa l.) subjected to combined light quality and drought. Front Plant Sci 2022, 13. [CrossRef]
- Zhang, R.X.; Yang, W.J.; Pan, Q.M.; Zeng, Q.; Yan, C.T.; Bai, X.; Liu, Y.; Zhang, L.G.; Li, B.H. Effects of long-term blue light irradiation on carotenoid biosynthesis and antioxidant activities in chinese cabbage (brassica rapa l. Ssp. Pekinensis). Food Res. Int. 2023, 174. [CrossRef]
- Klein, S.; Fiebig, A.; Noga, G.; Hunsche, M. Influence of light quality on leaf physiology of sweet pepper plants grown underdrought. Theor Exp Plant Phys 2018, 30, 287-296. [CrossRef]
- Terfa, M.T.; Olsen, J.E.; Torre, S. Blue light improves stomatal function and dark-induced closure of rose leaves (rosa x hybrida) developed at high air humidity. Front Plant Sci 2020, 11. [CrossRef]
- Barillot, R.; De Swaef, T.; Combes, D.; Durand, J.L.; Escobar-Gutiérrez, A.J.; Martre, P.; Perrot, C.; Roy, E.; Frak, E. Leaf elongation response to blue light is mediated by stomatal-induced variations in transpiration in. Journal of Experimental Botany 2021, 72, 2642-2656. [CrossRef]
- Su, Q.; Kambale, R.D.; Tzeng, J.-H.; Amy, G.L.; Ladner, D.A.; Karthikeyan, R. The growing trend of saltwater intrusion and its impact on coastal agriculture: Challenges and opportunities. Sci. Tot. Environ. 2025, 966, 178701.
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. Journal of Integrative Plant Biology 2018, 60, 796-804.
- Peng, Y.X.; Zhu, H.Y.; Wang, Y.T.; Kang, J.; Hu, L.X.; Li, L.; Zhu, K.Y.; Yan, J.R.; Bu, X.; Wang, X.J.; et al. Revisiting the role of light signaling in plant responses to salt stress. Hortic Res-England 2025, 12. [CrossRef]
- Hiyama, A.; Takemiya, A.; Munemasa, S.; Okuma, E.; Sugiyama, N.; Tada, Y.; Murata, Y.; Shimazaki, K. Blue light and co2 signals converge to regulate light-induced stomatal opening. Nat Commun 2017, 8. [CrossRef]
- Schuerger, A.C.; Brown, C.S.; Stryjewski, E.C. Anatomical features of pepper plants (capsicum annuum l) grown under red light-emitting diodes supplemented with blue or far-red light. Ann. Bot. 1997, 79, 273-282. [CrossRef]
- Sun, Y.; Xu, W.; Jia, Y.B.; Wang, M.C.; Xia, G.M. The wheat gene is involved in the blue-light response and salt tolerance. Plant J. 2015, 84, 1219-1230. [CrossRef]
- Shimazaki, K.; Doi, M.; Assmann, S.M.; Kinoshita, T. Light regulation of stomatal movement. Annu. Rev. Plant Biol. 2007, 58, 219-247. [CrossRef]
Figure 1.
Schematic diagram of the effects of blue light on plant growth and development.
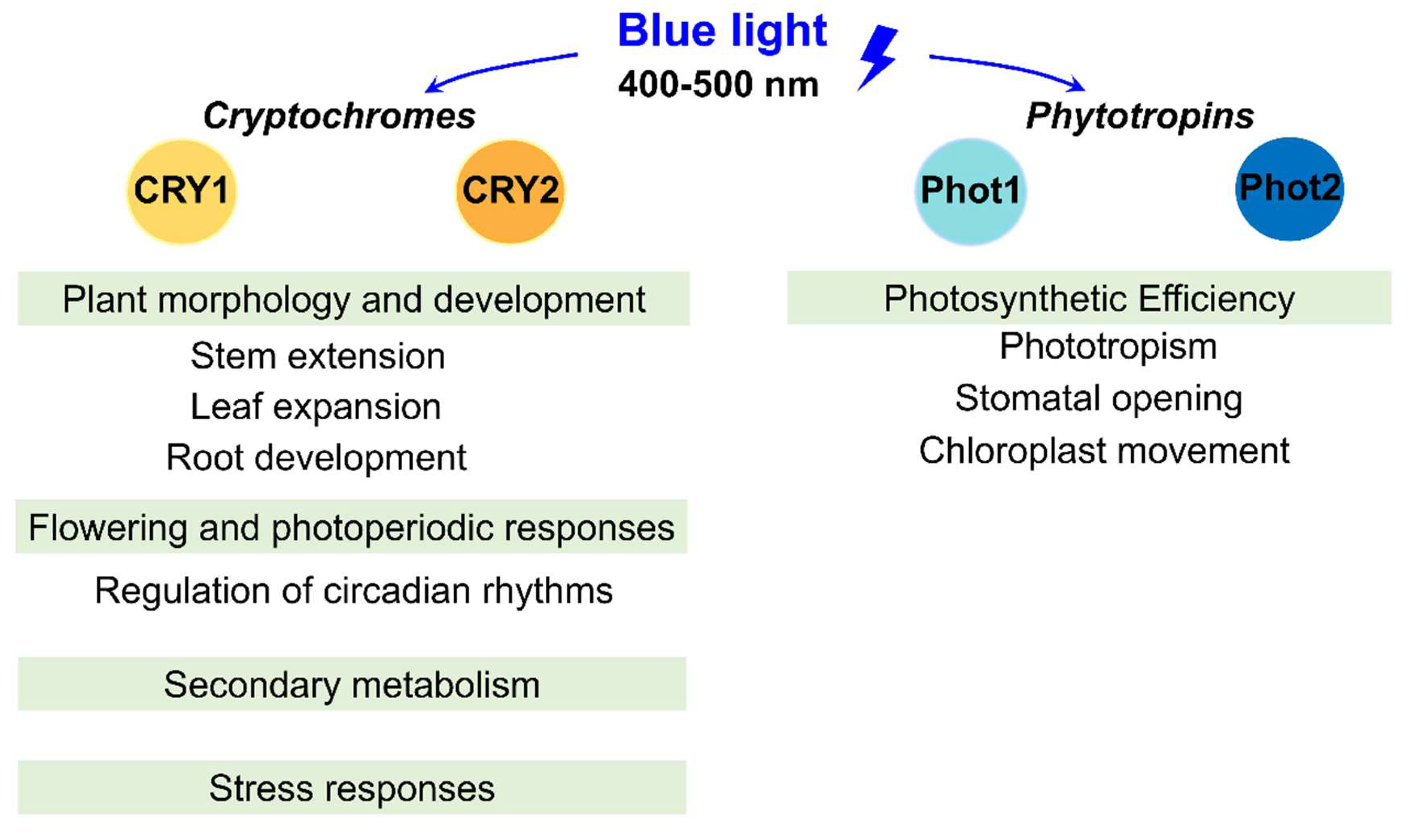
Figure 2.
Comparison of the effects of blue and red LED lights on seedling growth across different crops and cultivation conditions. (a) Cilantro, (b) green-leaf pak choi, (c) red-leaf pak choi, (d) sunflower, (e) Arugula growing at relatively high density, (f) Arugula, (g) Mustard, and (h) Arabidopsis growing in rockwool cubes. R is red LED light, and B is blue LED light. The photosynthetic photon flux density (PPFD) of the LED lights was 100
Figure 2.
Comparison of the effects of blue and red LED lights on seedling growth across different crops and cultivation conditions. (a) Cilantro, (b) green-leaf pak choi, (c) red-leaf pak choi, (d) sunflower, (e) Arugula growing at relatively high density, (f) Arugula, (g) Mustard, and (h) Arabidopsis growing in rockwool cubes. R is red LED light, and B is blue LED light. The photosynthetic photon flux density (PPFD) of the LED lights was 100

Figure 3.
Blue light dose-response curves for photosynthetic and leaf properties. The effects of light quality on (a) the photosynthetic capacity (Amax) of leaves, (b) chlorophyll content, (c) leaf mass per unit area (LMA), (d) Amax per leaf chlorophyll content, (e) Amax per leaf dry weight (DW), and (f) Amax per in the leaf on a dry weight basis, and (g) Amax per unit nitrogen during the growth of cucumber. Adapted from Hogewoning et al. [25].
Figure 3.
Blue light dose-response curves for photosynthetic and leaf properties. The effects of light quality on (a) the photosynthetic capacity (Amax) of leaves, (b) chlorophyll content, (c) leaf mass per unit area (LMA), (d) Amax per leaf chlorophyll content, (e) Amax per leaf dry weight (DW), and (f) Amax per in the leaf on a dry weight basis, and (g) Amax per unit nitrogen during the growth of cucumber. Adapted from Hogewoning et al. [25].
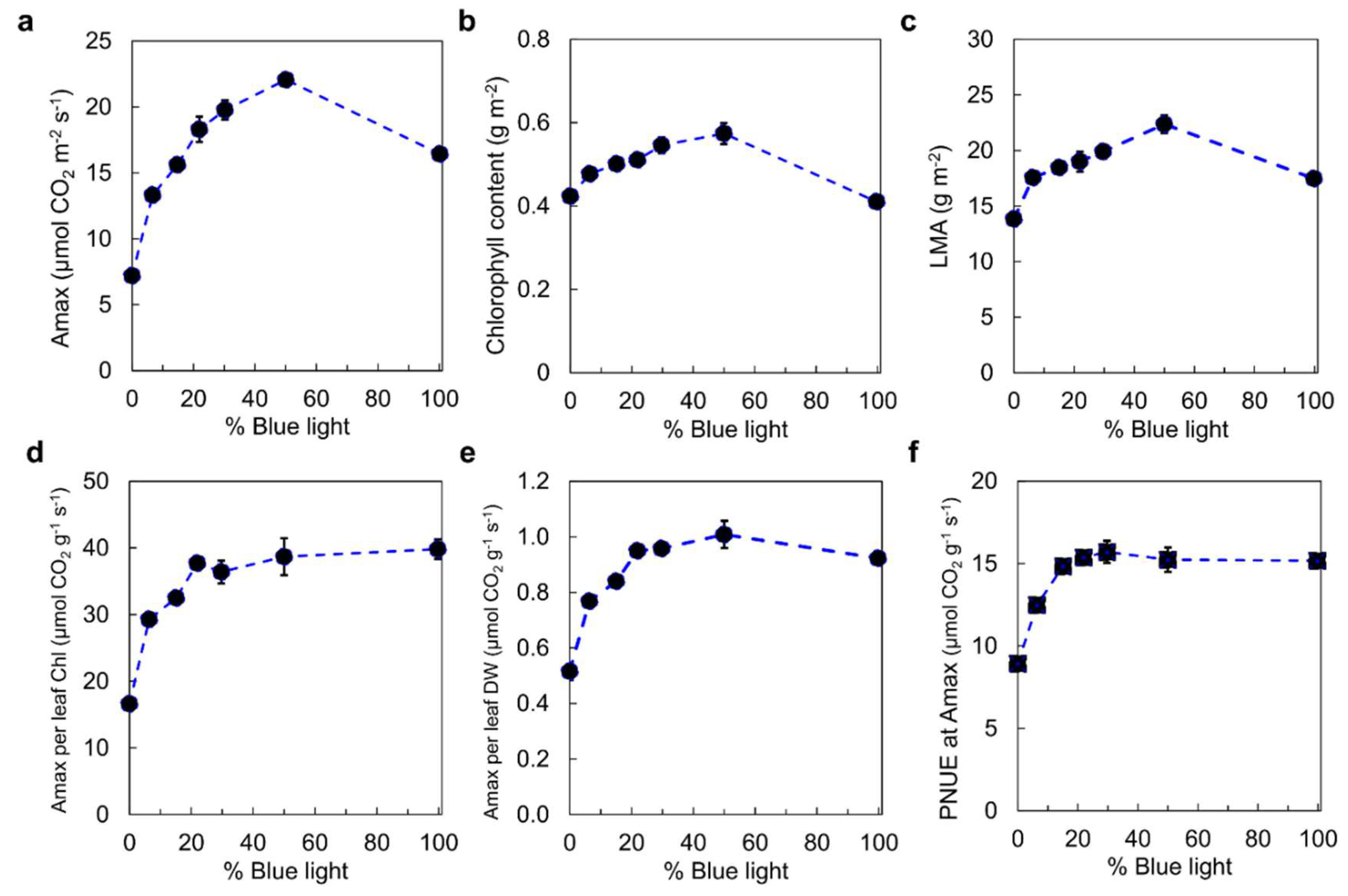
Figure 4.
Effects of supplementary and night interruption at 10 μmol m−2 s−1 PPFD on the flowering of kalanchoe (Kalanchoe blossfeldiana ‘Rudak’) 60 days after treatment. Supplementary and night-interrupting blue light schemes employed in this study: The control plants were subjected to a 10-h short-day (SD, positive control) or 13-h long-day (LD, negative control) treatment without any blue light. The blue light was used for 4 h either (1) to supplement the W LEDs at the end of the SD (SD + 4B) and LD (LD + 4B) periods or (2) to provide night interruption (NI) in the SD (SD + NI-4B) and LD (LD + NI-4B) periods. The light period began at 8:00 a.m., and the dark period ended at 8:00 a.m. Adapted from Yang et al. [16].
Figure 4.
Effects of supplementary and night interruption at 10 μmol m−2 s−1 PPFD on the flowering of kalanchoe (Kalanchoe blossfeldiana ‘Rudak’) 60 days after treatment. Supplementary and night-interrupting blue light schemes employed in this study: The control plants were subjected to a 10-h short-day (SD, positive control) or 13-h long-day (LD, negative control) treatment without any blue light. The blue light was used for 4 h either (1) to supplement the W LEDs at the end of the SD (SD + 4B) and LD (LD + 4B) periods or (2) to provide night interruption (NI) in the SD (SD + NI-4B) and LD (LD + NI-4B) periods. The light period began at 8:00 a.m., and the dark period ended at 8:00 a.m. Adapted from Yang et al. [16].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



