
Submitted:
04 May 2025
Posted:
07 May 2025
You are already at the latest version
Abstract
Modern wheat breeding has largely emphasized aboveground traits, often at the expense of belowground characteristics such as root biomass, architecture, and beneficial microbial associations. This has narrowed genetic diversity, impacting traits essential for stress resilience and efficient nutrient and water acquisition—factors expected to become increasingly critical under climate change. In this study, we evaluated 36 primary synthetic (PS) hexaploid wheat lines developed by crossing Aegilops tauschii with the durum wheat cultivar Langdon (LNG) and compared them with LNG and the hexaploid variety Norin 61 (N61). We observed significant variation in root length, biomass, and associations with fungal endophytes, including arbuscular mycorrhizal fungi (AMF), Serendipita indica, and Alternaria. Clustering analysis based on these traits identified three distinct PS groups: (1) lines with greater root length and biomass, high AMF and S. indica colonization, and low Alternaria infection; (2) lines with intermediate traits; and (3) lines with reduced root traits and high Alternaria susceptibility. Notably, these phenotypic patterns corresponded closely with the soil classification of the Ae. tauschii progenitors’' origin, such as Cambisols (supportive of root growth), Gleysols and Calcisols (restrictive of root growth). This highlights the soil microenvironment as a key determinant of belowground trait expression. Our findings demonstrate the potential of wild D-genome diversity, coupled with soil–environment interactions, to restore critical root traits in wheat. Incorporating PS lines with targeted soil and microbial considerations offers a promising strategy for breeding resilient cultivars with enhanced root systems.
Keywords:
genetic diversity
; root length
; root biomass
; Arbuscular Mycorrhizal Fungi (AMF)
; Serendipita indica
; Alternaria
; soil classification
1. Introduction
The need for sustainable wheat production has become increasingly urgent as climate change—manifesting as heat stress, drought, and waterlogging—continues to threaten global wheat yields and quality [1]. Addressing these challenges requires a transition toward integrated agricultural strategies that enhance resilience to both biotic and abiotic stresses while reducing environmental impacts [2]. Wheat’s adaptability, global distribution, and high productivity have established it as a mainstay of global agriculture. However, domestication and modern breeding efforts have largely focused on aboveground traits such as grain yield and disease resistance, inadvertently narrowing genetic diversity, especially in belowground characteristics [3]. These root traits—including root length, diameter, biomass, and key plant–microbe interactions that support water and nutrient uptake—are essential for drought resilience and overall plant performance [4,5]. Every phase of root initiation, growth, and development is genetically regulated and shaped by diverse abiotic and biotic factors [6]. Additionally, root-associated microbial interactions, particularly with beneficial endophytes such as Arbuscular Mycorrhizal Fungi (AMF) and Serendipita indica (S. indica), play a pivotal role in enhancing stress tolerance, disease resistance, and plant growth [7,8]. The decline in these critical root-associated traits poses a growing concern for sustainable wheat production, especially under escalating climate pressures. Wild relatives of wheat, such as Aegilops tauschii (the D-genome donor of bread wheat), represent a valuable reservoir of genetic diversity, offering potential to improve wheat resilience and yield under both abiotic and biotic stresses. Studies have shown that the offspring of hexaploid wheat, including primary synthetic (PS) and multiple synthetic derivatives (MSD), exhibit enhanced stress tolerance and disease resistance compared to domesticated varieties [9,10,11]. However, research on root-associated traits in Ae. tauschii and its hybrids remains limited, especially relative to the extensive focus on aboveground traits. Ae. tauschii is predominantly distributed across, the Caspian Sea region, and parts of Central and South Asia—areas characterized by diverse soil types [12,13]. Soils in these regions, including Cambisols and Gleysols, exhibit distinct properties that influence both root development and microbial interactions [14]. Accessions of Ae. tauschii have demonstrated strong root systems, increased associations with AMF, and resistance to Alternaria infections [15]. Furthermore, our previous study [16] identified significant variation in root traits among Ae. tauschii accessions, with this diploid ancestor showing superior root traits (including enhanced AMF colonization) compared to cultivated tetraploid and hexaploid wheat. However, as Ae. tauschii is phenotypically distinct, these advantageous traits may not fully express in hexaploid wheat. To bridge this gap, we analyzed Primary Synthetic wheat (PS) lines derived from diverse Ae. tauschii accessions across diverse soil environmnts. These wild-derived root traits and their genetic regulation offer promising targets for improving belowground resilience and wheat performance [15,16]. By integrating root traits with microbial associations (AMF, S. indica) and aboveground selection [17,18], we aim to (1) quantify root trait variation in PS lines, (2) assess their interactions with beneficial (AMF/S. indica) and pathogenic (Alternaria) fungi relative to their Ae. tauschii soils classification, and (3) determine trait inheritance patterns to inform future breeding strategies.
2. Materials and Methods
2.1. Plant Materials and Experimental Conditions
We used 36 synthetic wheat lines (hexaploid, AABBDD) produced by crosses between a single durum wheat cultivar (Triticum turgidum var. durum cv. Langdon (LNG), tetraploid, AABB) and 36 accessions of Ae. tauschii (diploid, DD, wild species) originating from diverse regions with varied environmental conditions. The soil classification of collected Ae. tauschii accessions was determined by georeferencing coordinates against the Soil Atlas of Asia [13], with additional verification for Iranian accessions using FAO’s national fertilizer report [17]. These accessions represented three distinct lineages of Ae. tauschii (TauL1, TauL2, and TauL3). LNG and a bread wheat cultivar (Triticum aestivum cv. Norin 61 (N61), hexaploid, AABBDD) were used as check cultivars (Table S1). Information related to the PS lines and their Ae. tauschii accession, including passport data source, origin, province, soil classification, reference for soil classification, and lineage, is presented in Supplementary Table S1. The selected lines span a wide range of geographical regions, soil types, and climatic zones. Among the 36 PS lines, 14 belong to lineage 1 (TauL1), 18 to lineage 2 (TauL2), and 4 to lineage 3 (TauL3). Their diverse geographic origins and genetic backgrounds make these lines valuable material for this study.
The experiment was conducted in a greenhouse at the Arid Land Research Center (ALRC), Tottori University, Japan (35°32′04.5″N, 134°12′42.0″E), from 1 November 2023 to 6 March 2024. Temperature and relative humidity were recorded using a datalogger (SK-L754; Sato Keiryoki Mfg. Co. Ltd., Tokyo, Japan) (Figure S1). Daily temperatures ranged from 2°C to 26°C, with average daytime and nighttime temperatures of 16°C and 10°C, respectively. Relative humidity fluctuated between 40% and 90%, averaging 66% during the day and 60% at night.
2.2. Plant Growth and Experiment Setup
Seeds were sown in tall pots (60 cm high, 10 cm wide) designed for Glycyrrhiza cultivation, filled with approximately 4000 cm³ of sandy soil sourced from the ALRC wheat field. Initial soil analysis revealed low fungal levels (103 CFU/g soil) and a low AMF spore count (5.1/g). To amend these limitations, we prepared a mixture of crude inoculum extracted from the same wheat field soil (containing 10⁶ fungal CFU/ml) and a commercial AMF product, Mycogel (Agrocode BioscienceTM, Roquetas de Mar, Almeria, Spain), containing Rhizophagus irregularis strain at 5 × 10³ spores/ml [18]. Twelve days after germination, 1 ml of this mixture was inoculated into the rhizosphere to facilitate interaction and colonization of root cortical cells by AMF and root-associated endophytic fungi. The experiment was laid out in a randomized complete block design with six replications. Pots were irrigated every three days, and no fertilizers, insecticides, or fungicides were applied.
2.3. Harvest and Data Collection
After three months of growth, all plants were carefully removed from the pots. Shoots and root systems were separated, and roots were thoroughly rinsed with tap water. Several traits were assessed, including root, shoot, and biotic characteristics.
2.3.1. Shoot Traits
The shoot systems from six replicates were weighed and then dried at 68°C for two days in a constant-temperature drying oven to determine fresh and dry weights, as well as tissue water content.
2.3.2. Root Traits
As architectural traits of roots determine the spatial configuration of the root system; root length and weight were measured. Harvested roots were digitally scanned using a desktop scanner (Perfection V500 Photo; Epson, Japan) and analyzed with WinRhizo Pro 2008a software (Regent Instruments Inc., Quebec City, QC, Canada) to determine total root length. Roots were classified into two diameter categories: thin (0–0.5 mm) and thick (0.5–1.0 mm) for subsequent analysis (Figure S2). For biotic trait analysis, 60–65 root segments of 0.5 cm each (totaling 30 cm) were randomly selected from each wheat root. The remaining roots were processed similarly to shoots to assess fresh and dry weights and tissue water content.
2.3.3. Root Biotic Traits
Biotic traits, including colonization by AMF and S. indica and Alternaria infection, were evaluated following the method of [19]. Root fragments were disinfected with 70% ethanol, soaked in 10% KOH overnight, rinsed with distilled water, acidified in 5% HCl for 3 min, and stained overnight with trypan blue (500 mL glycerol, 475 mL distilled water, 25 mL acetic acid, 0.1 g trypan blue). After rinsing with acidified water, samples were observed under a digital microscope (VHX-7000; Keyence, Osaka, Japan). Colonization levels were quantified using the gridline intersect method [20]. The three fungi were identified based on their distinct morphological structures within the root (Table S6, Figures S5–S10).
2.4. Statistical Analysis
Data were tested for normality and homogeneity of variance using the Shapiro–Wilk and Levene’s tests. Welch’s one-way analysis of variance was performed, followed by Tukey’s honestly significant difference (HSD) test for mean comparisons. Pearson’s correlation analysis was conducted to evaluate relationships among root traits. The statistical analyses were conducted using SPSS v.29 (IBM SPSS Statistics, 2024). Additionally, a hierarchical clustering heatmap and a biplot principal component analysis (PCA) were generated to classify PS lines based on trait similarities. Hierarchical clustering heatmaps and biplot PCA were created using R Statistical Software (v.4.4.1; R Core Team, 2024).
3. Results
3.1. Variation of Traits Among Wheat PS Lines
3.1.1. Root Traits
Total root length (cm), including both thin and thick root diameter classes, varied significantly across the wheat accessions studied (p < 0.05) Figure 1A. Thirty-one out of the 36 PS lines exhibited longer root lengths than LNG and N61, with PS63 showing the longest root system (6209 cm). Among the remaining lines, PS39 had a root length of 2944 cm, comparable to LNG (2906 cm), and both were significantly shorter than the first 31 PS lines. PS48 had a root length of 2679 cm, comparable to N61 (2609 cm). The shortest root lengths were recorded in PS30, PS29, and PS54 (2434 cm, 2430 cm, and 1949 cm, respectively) Figure 1A.
Root dry weight and water content also varied significantly among the studied wheat lines (p < 0.05) Figure 1B. Twenty-six PS lines exhibited significantly higher root weights than the remaining 10 lines, and LNG and N61. Among them PS47 and PS63 had the highest root dry weights of 0.498 g and 0.506 g, and water contents of 4.83 g and 4.688 g, respectively.
Regarding average root diameter, all PS lines showed significantly greater diameters than LNG, except PS 51, 36, 59, 42, and 50, which had values comparable to LNG Figure 1C. N61 displayed the smallest average root diameter among all accessions.
3.1.2. Shoot Traits
Shoot dry weight and water content were comparable across all PS lines, N61, and LNG. PS47 recorded the highest shoot dry weight (1.13 g) and water content (1.89 g). Only PS54 showed significantly lower shoot weight and water content compared to PS47, with values of 0.29 g and 0.31 g, respectively Figure 1D.
3.1.3. Biotic Traits
Biotic traits, including root colonization by beneficial fungi (AMF and S. indica) and pathogenic fungi (Alternaria), varied significantly among all wheat accessions (Figure 2). AMF colonization exhibited notable variation and allowed classification of the accessions into 11 distinct groups based on colonization levels. All PS lines demonstrated significantly higher AMF colonization than LNG and N61, which had the lowest colonization rates at 25% and 18%, respectively Figure 2A. Among the 36 PS lines, PS37, PS63, and PS42 exhibited the highest AMF colonization rates (>60%). In contrast, colonization levels gradually declined below 30% in N61 and LNG, marking them as the least colonized accessions.
S. indica colonization was lower than that of AMF but showed significant variation among all wheat accessions. The highest S. indica colonization was observed in three PS lines (63, 37, and 72), with colonization rates exceeding 25%. Similar to AMF, S. indica colonization gradually declined across the PS lines, reaching the lowest levels in PS 50 and LNG (12.5% and 11.6%, respectively). N61 exhibited an intermediate colonization rate of 16% Figure 2B.
N61 exhibited the highest infection rate of Alternaria (32%) among all 38 wheat accessions, while LNG had the lowest (11%). Among the PS lines, 19 lines (PS51, PS54, PS48, PS36, PS68, PS69, PS27, PS50, PS30, PS39, PS60, PS66, PS29, PS38, PS33, PS35, PS49, PS52, and PS59) showed higher infection rates, ranging from 24% to 17%. Infection levels gradually declined among the remaining PS lines (16%–12%), with the lowest infection among PS lines observed in PS42 (11.75%) Figure 2C.
3.1.4. Association Among Wheat Accession Traits
All wheat accession traits (root, shoot, and biotic) exhibited highly significant positive correlations with one another, except for Alternaria infection, which showed a highly significant negative correlation with all other traits, including beneficial endophytic fungi (AMF and S. indica) Figure 3. Shoot fresh and dry weights were most strongly correlated with thick root length, which plays a crucial role in water uptake for the shoot system.
3.2. Wheat Accession Grouping and Traits Association
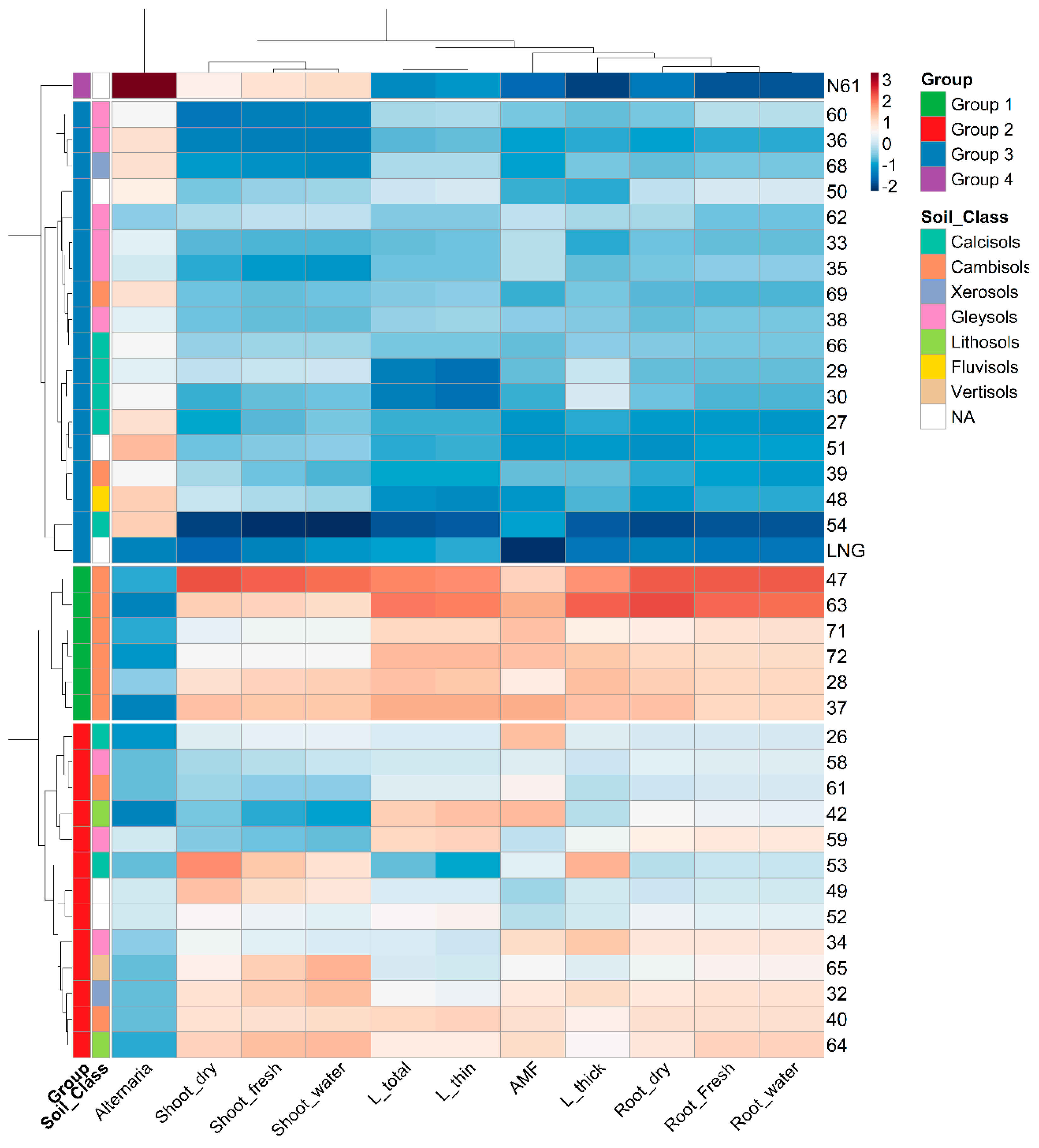
Hierarchical clustering heatmap of root traits revealed four distinct groups among the wheat accessions Figure 4. (group 1) comprised six PS lines exhibiting the highest root and shoot traits, the strongest AMF colonization, and the lowest Alternaria infection. Notably, all group 1 PS lines were derived from Ae. tauschii accessions originating exclusively from Cambisols. (group 2) included 13 PS lines with intermediate trait values. Their Ae. tauschii progenitor accessions were derived from diverse soil classes: three from Gleysols; two each from Cambisols, Calcisols, and Lithosols; and one each from Xerosols and Vertisols, along with two accessions from unclassified (NA) soils. (group 3) consisted of 17 PS lines along with their parent LNG. This group was characterized by lower root and shoot trait values and the highest Alternaria infection among PS lines. Their Ae. tauschii progenitors soil classification showed six accessions from Gleysols, five from Calcisols, two from Cambisols, and one each from Xerosols and Fluvisols, with three unclassified (NA) accessions. (group 4) included only accession N61, which displayed the most extreme phenotypic divergence. It showed minimal root development, sustained shoot growth, and the highest level of Alternaria infection observed in the study Figure 4.
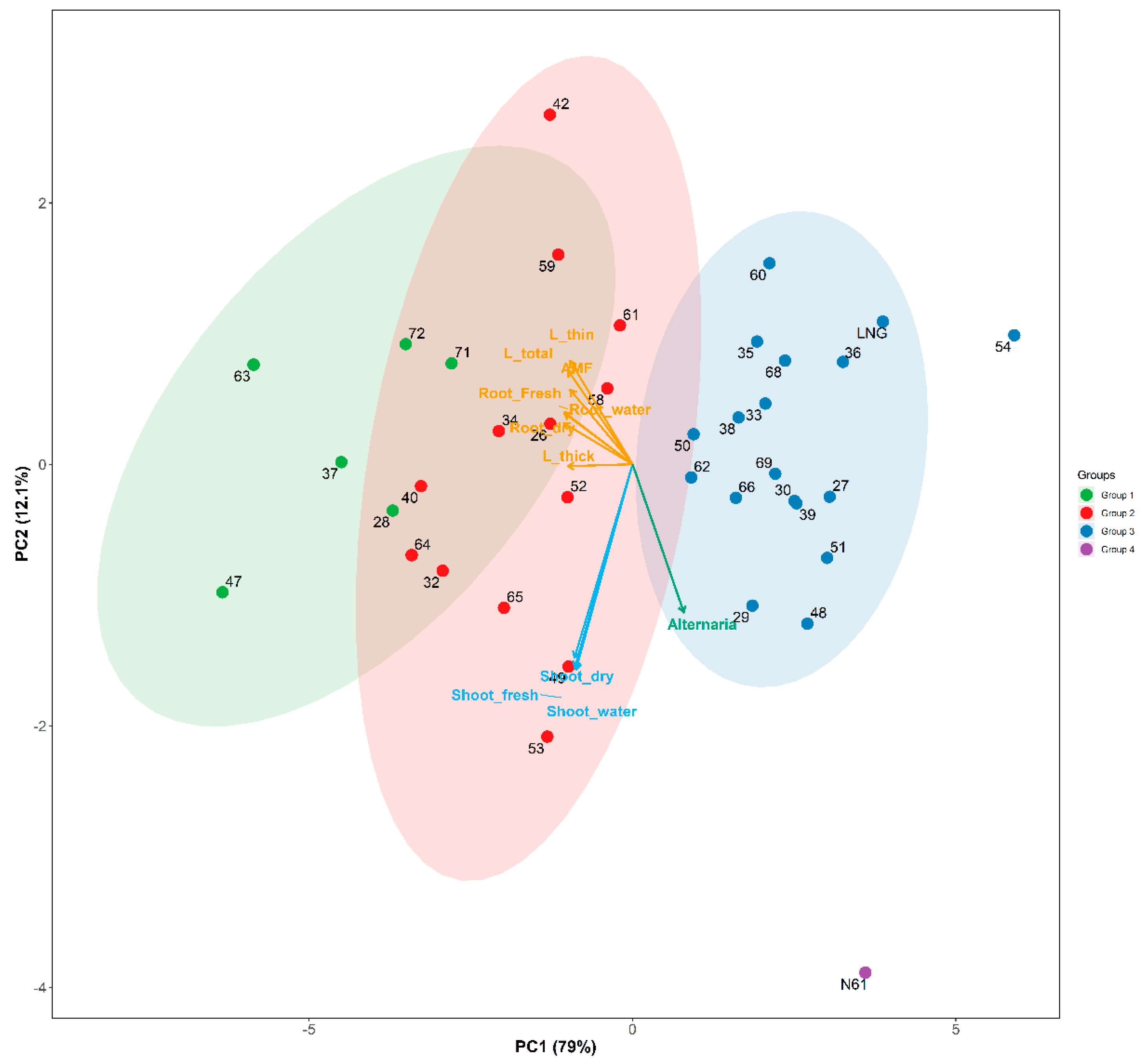
PCA biplot explained 91% of the total variance (PC1: 78.97%, PC2: 12.1%) Figure 5. The result showed similar classification into groups with hierarchical clustering heatmap shown in Figure 4. PS lines were separated into three groups, with their progenitor LNG. N61 formed the fourth group. Notably, some overlap was observed between Groups 1 and 2, primarily due to similarities in root traits. PS54, although classified within Group 3 in Figure 4, deviated even more than its LNG progenitor, from group 3, driven by its exceptionally low root and shoot traits and high Alternaria infection Figure 5.
4. Discussion
Root phenomics is crucial in crop breeding; among key root traits, root length and biomass are vital for water and nutrient uptake [21]. Root traits such as increased root length, variation in thin and thick root diameters, and greater biomass enhance wheat performance, particularly under stress, by improving water uptake, boosting nutrient acquisition, and fostering beneficial microbial interactions [4,5,22]. Breeding strategies that target these traits can enhance drought resilience and nutrient efficiency, ultimately increasing yields in challenging environments [6,23,24]. This study identified significant variation in root traits-length, diameter, and biomass—as well as in biotic interactions with endophytic fungi (AMF, S. indica, and Alternaria) among 36 PS wheat lines derived from 36 Ae. tauschii accessions and LNG. The considerable variability in wheat root traits observed in this study, especially among the 36 PS lines (Figure 1A, 1B–and 1C), underscores the influential role of the wild D-genome from Ae. tauschii in shaping root architecture. The improved root traits in certain PS lines are likely inherited from Ae. tauschii [16]. Conversely, reduced root traits in some PS lines may reflect the influence of the AB genome from LNG, which has undergone domestication-related selection focused primarily on aboveground traits [25]. Previous research has highlighted the critical role of the D-genome in root trait development. For instance, [15] identified TaLBD16-D as a key gene regulating lateral root number, with the D-genome making notable contributions in both diploid and hexaploid wheat compared to the tetraploid AB genome. In our earlier study [16], Ae. tauschii accessions KU-2109, KU-2074, KU-2136, IG 126387, AT76, and IG 47259 consistently demonstrated superior root traits compared to LNG and N61. In the current study, their corresponding PS lines PS64, PS33, PS66, PS29, PS51, and PS54 retained many of these advantages, though they did not include the top-performing PS lines overall. Notably, all lines except PS29 and PS54 showed greater root length than LNG and N61, and only line PS54 exhibited a root dry weight similar to N61 and lower than LNG. Average root diameter was consistently higher across all six lines relative to LNG and N61. These findings illustrate the inheritance of beneficial root traits from Ae. tauschii progenitors and highlight their valuable contribution to root trait enhancement in synthetic hexaploid wheat.
Biotic traits variability among PS lines: AMF, S. indica, and Alternaria resistance
Beneficial AMF and S. indica have been shown to enhance wheat performance by improving nutrient uptake, promoting root growth, increasing drought tolerance, and strengthening resistance to biotic stresses such as fungal pathogens [26,27,28]. In contrast, Alternaria represents a major biotic stressor in wheat, contributing to root rot and post-harvest spoilage [7]. Investigating these root endophytic fungi holds significant promise for advancing sustainable wheat production, particularly under biotic and abiotic stress conditions. In this study, as with root traits, the biotic traits, including both beneficial and pathogenic root endophytic fungi, varied significantly among wheat accessions and across the 36 PS lines. All PS lines exhibited significantly higher colonization by AMF compared to LNG and N61, supporting the same hypothesis as for root traits: that the wild D-genome of Ae. tauschii may contribute to promoting beneficial fungal associations. [29] proposed that AMF symbiosis-related genes in wild ancestors can be transmitted without major disruption from AB genome introgression.
Resistance to Alternaria colonization also varied across the PS lines. While some PS lines retained strong resistance, others exhibited increased susceptibility; nevertheless, all PS lines showed lower Alternaria colonization than the domesticated hexaploid N61, closely resembling their progenitor Ae. Tauschii, as reported in our previous study [16]. At the same time, all PS lines exhibited significantly higher Alternaria colonization than their domesticated progenitor LNG, which appeared to suppress both beneficial and pathogenic fungal associations, a finding consistent with our earlier observations in Ae. tauschii [16]. In our prior study, Ae. tauschii accessions KU-2109, KU-2074, KU-2136, IG 126387, AT76, and IG 47259 consistently demonstrated higher AMF colonization and lower Alternaria infection compared to LNG and N61. In the current study, we assessed their derived PS lines-PS64, PS33, PS66, PS29, PS51, and PS54—which displayed intermediate biotic traits. Although not the top-performing PS lines overall, they still outperformed LNG and N61 by exhibiting higher AMF colonization and reduced Alternaria infection, reflecting the trends seen in their wild progenitors.
Root-biotic trait associations
The strong positive correlation observed between root traits and beneficial biotic traits (AMF and S. indica) (Figure 3) underscores the interconnected nature of plant–microbe interactions. Enhanced root length and biomass likely facilitated the recruitment of beneficial fungi, reinforcing symbiotic associations [30] and contributing to the suppression of Alternaria. This is further supported by the negative correlation between Alternaria and both root traits and beneficial fungi, aligning with our previous study [16], which focused exclusively on the two progenitors of the PS lines (Ae. tauschii and LNG).
Soil classification affects wheat root traits grouping
Significant variability in root traits, AMF colonization, and Alternaria susceptibility among wheat accessions and PS lines was captured through hierarchical clustering and PCA, consistently resolving four distinct groups (Figure 4 and Figure 5). Group 1, comprising six PS lines, exhibited superior root and shoot traits, stronger AMF colonization, and reduced Alternaria infection, marking them as valuable germplasm for improving belowground traits. PS47 and PS63, derived from Ae. tauschii accessions KU-2159 and KU-2103 (Iran), were among the top performers.
Remarkably, Group 1 PS lines originated from all three Ae. tauschii lineages and diverse geographic regions but shared association with Cambisols—well-drained, moderately weathered soils favorable for root growth [13,17,31]. This convergence suggests that soil type, rather than lineage or climatic region, exerts a dominant influence on belowground phenotypes. Unlike climate, soil acts as the direct interface for belowground development, which helps explain the variation observed among PS lines originating from similar regions but differing soil types, contrasting to [12] who found aboveground traits more influenced by climatic region and lineages.
In contrast, PS lines in Groups 2 and 3—despite similar lineage and regional origins—were primarily linked to Gleysols and Calcisols. These soils are known to restrict root development due to waterlogging and high CaCO₃ content, respectively. Gleysols reduce root elongation [14], while Calcisols form cemented pans and chemically suppress root hair and AMF development [32,33,34]. These edaphic limitations were reflected in reduced root biomass and higher Alternaria susceptibility.
Altogether, these findings highlight soil type—particularly Cambisols—as a critical and underappreciated factor influencing root performance. Incorporating soil origin into Ae. tauschii donor selection may enhance breeding strategies targeting belowground resilience.
Impact of domesticated LNG on PS lines
Beyond soil and genetic diversity within Ae. tauschii lineages, the hybridization effect from the second progenitor, the domesticated LNG, likely contributes to trait variability among PS lines. This is supported by the consistently lower root traits and weaker fungal associations observed in both the current and previous study [16], highlighting the presence of admixed PS lines.
A key distinction between the hexaploid PS lines and the domesticated hexaploid N61 lies in the origin of their D-genome. While N61’s domesticated DD genome has undergone selection pressures that may have reduced investment in belowground traits, the PS lines retain a wild DD genome, preserving the genetic potential for enhanced root development and beneficial biotic interactions [29,35,36]. This highlights the critical importance of conserving wild germplasm to improve wheat resilience to both abiotic and biotic stresses.
Limitations and future directions
A primary limitation of this study is the restricted number of PS wheat lines derived from Ae. tauschii. Although these lines represent various geographical regions and taxonomical lineages, they cover only a small fraction of the extensive Ae. tauschii germplasm available. Additionally, all PS lines were developed using the AB genome background from LNG, which may not fully capture the diversity of the A and B genomes found in wheat. Future directions include utilizing Multiple synthetic derivatives (MSDs), created by crossing PS lines with elite hexaploid wheat, as a powerful platform for tracking trait inheritance in breeding populations. The application of Genome-Wide Association Study could further identify genes linked to beneficial root traits. Lastly, exploring how soil types and microbial communities influence root development can inform the strategic deployment of cultivars across diverse global environments.
5. Conclusions
This study highlights substantial variation in root traits and biotic interactions among 36 PS wheat lines, shaped by the genomic interplay between their two progenitors: the wild Ae. tauschii and the domesticated LNG. The presence of enhanced root traits and beneficial fungal associations in several PS lines indicates a strong influence from the Ae. tauschii genome, whereas reduced traits in other lines reflect domestication-related effects inherited from LNG. Trait expression was further modulated by soil type, with Cambisols supporting superior root development and microbial colonization compared to Gleysols. These findings underscore the complex genetic and environmental interactions that govern belowground traits in wheat. The clustering of PS lines into three distinct phenotypic groups offers a practical framework for selecting genotypes based on root traits and endophytic fungal associations. From Group 1, six PS lines—47, 63, 71, 72, 28, and 37—are recommended for future exploration in wheat breeding programs due to their superior performance. Leveraging wild genetic diversity through the targeted use of PS lines, while accounting for soil environments and microbial interactions, presents a promising path toward developing resilient wheat cultivars optimized for root performance and stress tolerance.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org; 1- Mohammedali et al., Supplementary tables excel sheet containing: Table S1: Information of Aegilops tauschii accessions used for production of 36 primary syntetic (PS) wheat lines. Table S2: Root , shoot and biotic traits; and their statistical significance of differences among the 38 wheat accessions (36 P.S lines, LNG and N61). Table S3: Eigenvalues for the screeplot Fig S2 and Biplot PCA Fig 5. Table S4: Wheat accessions scores for Biplot PCA Fig 5. Table S5: Traits loading for Biplot PCA Fig 5. 2- Mohammedali et al., Supplementary materials PDF containing: Figure S1: Daily average temperature and relative humidity in greenhouse during the experiment (March 30th to June 30th, 2023). Figure S2: Screeplot for the percentage of explained variance for the dimensions of the biplot PCA in figure 5. Figure S3: scanned Images with Epson scanner for WinRhizo analysis of wheat accessions root system from both studies demonstrates the variation of the root systems among P.S accessions, and both their parents Aegilops tauschii and Langdon compared to the control Norin61. Figure S4: Image of scanned root fragment illustrating the subcategory classes of the root according to the diameter: Thin root 0 > 0.5 mm and thick root 0.5-1.0 mm. Figure S5: Arbuscules of Mycorrhizal Fungi structure inside wheat root cortical cells: Visualized by Keyence VHX digital microscope. Figure S6: Spores of Arbuscular Mycorrhizal Fungi structure inside wheat root cortical cells: Visualized by Keyence VHX digital microscope. Figure S7: Vesicles of Arbuscular Mycorrhizal Fungi structure inside wheat root cortical cells: Visualized by Keyence VHX digital microscope. Figure S8: Inter and intracellular Hyphae and Vesicles of Serendipita indica Fungi structure inside wheat root cortical cells: Visualized by Keyence VHX digital microscope. Figure S9: Spores of Serendipita indica Fungi structure inside wheat root cortical cells: Visualized by Keyence VHX digital microscope. Figure S10: Alternaria spp. Fungi structure inside wheat root cortical cells: Visualized by Keyence VHX digital microscope. Table S6: features and structures of root endophytic fungi: AMF (Rhizophagus irregularis), Serendipita indica and Alternaria sp. inside wheat root cells.
Author Contributions
Conceptualization, Ahmed Mohammedali, Nasrein Kamal, Yasir Gorafi, Izzat Tahir, Hisashi Tsujimoto and Takeshi Taniguchi; Data curation, Ahmed Mohammedali, Nasrein Kamal and Takeshi Taniguchi; Formal analysis, Ahmed Mohammedali and Nasrein Kamal; Funding acquisition, Takeshi Taniguchi; Investigation, Ahmed Mohammedali, Yasir Gorafi, Izzat Tahir and Takeshi Taniguchi; Methodology, Ahmed Mohammedali, Yasir Gorafi, Izzat Tahir and Takeshi Taniguchi; Project administration, Takeshi Taniguchi; Resources, Hisashi Tsujimoto and Takeshi Taniguchi; Software, Ahmed Mohammedali and Nasrein Kamal; Supervision, Takeshi Taniguchi; Validation, Ahmed Mohammedali, Yasir Gorafi, Izzat Tahir and Takeshi Taniguchi; Visualization, Ahmed Mohammedali, Nasrein Kamal and Takeshi Taniguchi; Writing – original draft, Ahmed Mohammedali; Writing – review & editing, Nasrein Kamal, Yasir Gorafi, Izzat Tahir, Hisashi Tsujimoto and Takeshi Taniguchi.
Funding
“This research received no external funding”
Data Availability Statement
The data supporting the findings of this study are available within the article and its supplementary materials. Restrictions may apply to data derived from third-party sources or due to privacy/ethical considerations. For further details, refer to MDPI’s research data policies at https://www.mdpi.com/ethics.
Acknowledgments
The authors express their gratitude to Researcher Maha F. A. Hassan for her valuable contributions to formal Analysis. They also extend their appreciation to the members of the Laboratory of Microbial Ecology and Molecular Breeding and the technical staff of the Arid Land Research Center, Tottori University, for their invaluable laboratory assistance.
Conflicts of Interest
The authors declare no conflict of interest.
Ethical Compliance
The authors confirm that all applicable ethical standards were followed. No human or animal subjects were involved in this study.
References
- Calvin, K.; Dasgupta, D.; Krinner, G.; Mukherji, A.; Thorne, P.W.; Trisos, C.; Romero, J.; Aldunce, P.; Barrett, K.; Blanco, G.; et al. IPCC, 2023: Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, H. Lee and J. Romero (Eds.)]. IPCC, Geneva, Switzerland.; First.; Intergovernmental Panel on Climate Change (IPCC), 2023;
- Constas, M.A.; d’Errico, M.; Hoddinott, J.F.; Pietrelli, R. Resilient Food Systems – A Proposed Analytical Strategy for Empirical Applications Available online: https://ageconsearch.umn.edu/record/319840 (accessed on 28 March 2025).
- Reif, J.C.; Zhang, P.; Dreisigacker, S.; Warburton, M.L.; van Ginkel, M.; Hoisington, D.; Bohn, M.; Melchinger, A.E. Wheat Genetic Diversity Trends during Domestication and Breeding. Theor Appl Genet 2005, 110, 859–864. [CrossRef]
- Maeght, J.-L.; Rewald, B.; Pierret, A. How to Study Deep Roots—and Why It Matters. Front Plant Sci 2013, 4, 299. [CrossRef]
- Pierret, A.; Maeght, J.-L.; Clément, C.; Montoroi, J.-P.; Hartmann, C.; Gonkhamdee, S. Understanding Deep Roots and Their Functions in Ecosystems: An Advocacy for More Unconventional Research. Annals of Botany 2016, 118, 621–635. [CrossRef]
- Ober, E.S.; Alahmad, S.; Cockram, J.; Forestan, C.; Hickey, L.T.; Kant, J.; Maccaferri, M.; Marr, E.; Milner, M.; Pinto, F.; et al. Wheat Root Systems as a Breeding Target for Climate Resilience. Theor Appl Genet 2021, 134, 1645–1662. [CrossRef]
- Jaster-Keller, J.; Müller, M.E.H.; El-Khatib, A.H.; Lorenz, N.; Bahlmann, A.; Mülow-Stollin, U.; Bunzel, M.; Scheibenzuber, S.; Rychlik, M.; von der Waydbrink, G.; et al. Root Uptake and Metabolization of Alternaria Toxins by Winter Wheat Plants Using a Hydroponic System. Mycotoxin Res 2023, 39, 109–126. [CrossRef]
- Qiao, X.; Sun, T.; Lei, J.; Xiao, L.; Xue, L.; Zhang, H.; Jia, J.; Bei, S. Frontiers | Arbuscular Mycorrhizal Fungi Contribute to Wheat Yield in an Agroforestry System with Different Tree Ages.. [CrossRef]
- Crespo-Herrera, L.A.; Govindan, V.; Stangoulis, J.; Hao, Y.; Singh, R.P. QTL Mapping of Grain Zn and Fe Concentrations in Two Hexaploid Wheat RIL Populations with Ample Transgressive Segregation. Front. Plant Sci. 2017, 8. [CrossRef]
- Elbashir, A.A.E.; Gorafi, Y.S.A.; Tahir, I.S.A.; Kim, J.-S.; Tsujimoto, H. Wheat Multiple Synthetic Derivatives: A New Source for Heat Stress Tolerance Adaptive Traits. Breed. Sci. 2017, 67, 248–256. [CrossRef]
- Itam, M.O.; Mega, R.; Gorafi, Y.S.A.; Yamasaki, Y.; Tahir, I.S.A.; Akashi, K.; Tsujimoto, H. Genomic Analysis for Heat and Combined Heat–Drought Resilience in Bread Wheat under Field Conditions. Theor Appl Genet 2022, 135, 337–350. [CrossRef]
- Mahjoob, M.M.M.; Chen, T.-S.; Gorafi, Y.S.A.; Yamasaki, Y.; Kamal, N.M.; Abdelrahman, M.; Iwata, H.; Matsuoka, Y.; Tahir, I.S.A.; Tsujimoto, H. Traits to Differentiate Lineages and Subspecies of Aegilops Tauschii, the D Genome Progenitor Species of Bread Wheat. Diversity 2021, 13, 217. [CrossRef]
- Soil Atlas of Asia; Jones, A., Caon, L., Yigini, Y., Verbeke, I., Konyushkova, M., Vargas, R., Viatkin, K., Michéli, E., European Commission, Eds.; Publications Office: Luxembourg, 2024; ISBN 978-92-68-03116-2.
- Mauer, O.; Palátová, E. Mountain Ash (Sorbus Aucuparia L.) Root System Morphogenesis. J. For. Sci. 2002, 48, 342–350. [CrossRef]
- Wang, H.; Hu, Z.; Huang, K.; Han, Y.; Zhao, A.; Han, H.; Song, L.; Fan, C.; Li, R.; Xin, M.; et al. Three Genomes Differentially Contribute to the Seedling Lateral Root Number in Allohexaploid Wheat: Evidence from Phenotype Evolution and Gene Expression. The Plant Journal 2018, 95, 976–987. [CrossRef]
- Mohammedali, A.Kh.H.; Gorafi, Y.S.A.; Kamal, N.M.; Tahir, I.S.A.; Tsujimoto, H.; Taniguchi, T. Wheat Root Traits and Endophytic Fungal Association Differ with Ploidy. under_review 2025.
- FAO Fertilizer Use by Crop in the Islamic Republic of Iran Available online: https://openknowledge.fao.org/server/api/core/bitstreams/a59738f1-92fc-4bac-bbd9-00e6eadb0483/content (accessed on 30 April 2025).
- Martín, M.; Rubio, A.; Remesal, E.; Cano, C.; Bago, A. Application of the Ultimate Arbuscular Mycorrhizal Inoculant MYCOGEL® in Japan: Results and Prospects.
- Phillips, J.M.; Hayman, D.S. Improved Procedures for Clearing Roots and Staining Parasitic and Vesicular-Arbuscular Mycorrhizal Fungi for Rapid Assessment of Infection. Transactions of the British Mycological Society 1970, 55, 158-IN18. [CrossRef]
- McGONIGLE, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A New Method Which Gives an Objective Measure of Colonization of Roots by Vesicular—Arbuscular Mycorrhizal Fungi. New Phytologist 1990, 115, 495–501. [CrossRef]
- Smith, F.A. Plant Roots. Growth, Activity and Interaction with Soils. Annals of Botany 2007, 100, 151–152. [CrossRef]
- Lynch, J.P. Root Phenotypes for Improved Nutrient Capture: An Underexploited Opportunity for Global Agriculture. New Phytologist 2019, 223, 548–564. [CrossRef]
- Hafeez, A.; Ali, S.; Javed, M.A.; Iqbal, R.; Khan, M.N.; ÇIĞ, F.; Sabagh, A.E.; Abujamel, T.; Harakeh, S.; Ercisli, S.; et al. Breeding for Water-Use Efficiency in Wheat: Progress, Challenges and Prospects. Mol Biol Rep 2024, 51, 429. [CrossRef]
- Chen, S.; Long, L.; Sun, X.; Parsons, D.; Zhou, Z. Responsive Root Traits and Mitigating Strategies for Wheat Production under Single or Combined Abiotic Stress. European Journal of Agronomy 2025, 162, 127393. [CrossRef]
- Lynch, J.P. Harnessing Root Architecture to Address Global Challenges. The Plant Journal 2022, 109, 415–431. [CrossRef]
- Cheng, S.; Zou, Y.-N.; Kuča, K.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.-S. Elucidating the Mechanisms Underlying Enhanced Drought Tolerance in Plants Mediated by Arbuscular Mycorrhizal Fungi. Front. Microbiol. 2021, 12. [CrossRef]
- Hosseini, F.; Mosaddeghi, M.R.; Zarebanadkouki, M. Enhancing Rhizosphere Soil Water Retention in Wheat through Colonization with Endophytic Fungus Serendipita Indica. Rhizosphere 2023, 26, 100709. [CrossRef]
- Duan, H.-X.; Luo, C.-L.; Zhu, Y.; Zhao, L.; Wang, J.; Wang, W.; Xiong, Y.-C. Arbuscular Mycorrhizal Fungus Activates Wheat Physiology for Higher Reproductive Allocation under Drought Stress in Primitive and Modern Wheat. European Journal of Agronomy 2024, 161, 127376. [CrossRef]
- Tkacz, A.; Pini, F.; Turner, T.R.; Bestion, E.; Simmonds, J.; Howell, P.; Greenland, A.; Cheema, J.; Emms, D.M.; Uauy, C.; et al. Agricultural Selection of Wheat Has Been Shaped by Plant-Microbe Interactions. Front. Microbiol. 2020, 11, 132. [CrossRef]
- Lattacher, A.; Le Gall, S.; Rothfuss, Y.; Gao, C.; Harings, M.; Pagel, H.; Giraud, M.; Alahmad, S.; Hickey, L.T.; Kandeler, E.; et al. Rooting for Microbes: Impact of Root Architecture on the Microbial Community and Function in Top- and Subsoil. Plant and Soil 2025, 1–19. [CrossRef]
- Klimek-Kopyra, A.; Kulig, B.; Głąb, T.; Zając, T.; Skowera, B.; Kopcińska, J. EFFECT OF PLANT INTERCROPPING AND SOIL TYPE ON SPECIFIC ROOT LENGTH. ROMANIAN AGRICULTURAL RESEARCH 2015.
- Schneider, F.; Don, A. Root-Restricting Layers in German Agricultural Soils. Part I: Extent and Cause. Plant and Soil 2019, 442, 433–451. [CrossRef]
- Labidi, S.; Calonne, M.; Ben Jeddi, F.; Debiane, D.; Rezgui, S.; Laruelle, F.; Tisserant, B.; Grandmougin-Ferjani, A.; Lounès-Hadj Sahraoui, A. Calcareous Impact on Arbuscular Mycorrhizal Fungus Development and on Lipid Peroxidation in Monoxenic Roots. Phytochemistry 2011, 72, 2335–2341. [CrossRef]
- Sediqui, N.; Amin, M.W.; Dawlatzai, N.; Gulab, G.; Poyesh, D.S.; Terada, N.; Sanada, A.; Kamata, A.; Koshio, K. Elucidation of Shoot and Root Growth, Physiological Responses, and Quality Traits of Tomato (Solanum Lycopersicon L.) Exposed to Elevated Calcium Carbonate Concentrations. Horticulturae 2024, 10, 573. [CrossRef]
- Bektas, H.; Hohn, C.E.; Waines, J.G. Characteristics of the Root System in the Diploid Genome Donors of Hexaploid Wheat (Triticum Aestivum L.). Genet Resour Crop Evol 2017, 64, 1641–1650. [CrossRef]
- Li, A.; Liu, D.; Yang, W.; Kishii, M.; Mao, L. Synthetic Hexaploid Wheat: Yesterday, Today, and Tomorrow. Engineering 2018, 4, 552–558. [CrossRef]
Figure 1.
Root and shoot traits of 38 wheat accessions, including 36 primary synthetics (PS), Langdon, and Norin 61. (A) Total root length (cm), subdivided into thin and thick root diameter classes. (B) Root fresh weight (g), with subcategories of dry weight and water content. (C) Average root diameter (mm). (D) Shoot fresh weight (g), with subcategories of dry weight and water content (g). L_total, total root length (cm); L_thin, thin roots (0 > 0.5 mm diameter); L_thick, thick roots (0.5-1 mm diameter). Bars sharing the same letter are not significantly different (Tukey’s Honestly Significant Difference [HSD] test, p< 0.01).
Figure 1.
Root and shoot traits of 38 wheat accessions, including 36 primary synthetics (PS), Langdon, and Norin 61. (A) Total root length (cm), subdivided into thin and thick root diameter classes. (B) Root fresh weight (g), with subcategories of dry weight and water content. (C) Average root diameter (mm). (D) Shoot fresh weight (g), with subcategories of dry weight and water content (g). L_total, total root length (cm); L_thin, thin roots (0 > 0.5 mm diameter); L_thick, thick roots (0.5-1 mm diameter). Bars sharing the same letter are not significantly different (Tukey’s Honestly Significant Difference [HSD] test, p< 0.01).
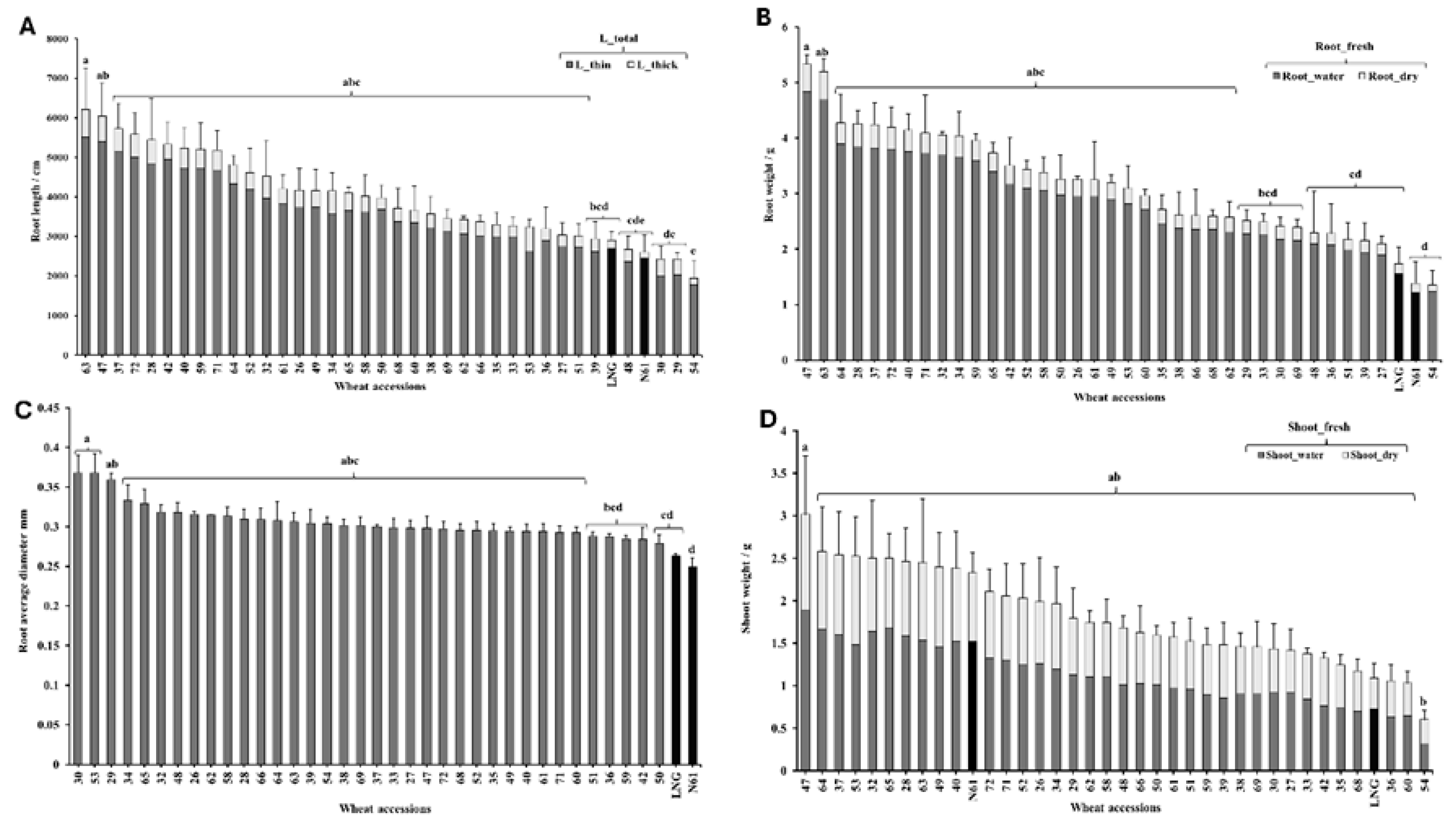
Figure 2.
Biotic traits of 38 wheat accessions, including 36 primary synthetics (PS), Langdon, and Norin 61. (A) Arbuscular mycorrhizal fungi (AMF) colonization % (B) Serendipita indica (S.i) colonization %. (C) Alternaria infection (%). Bars sharing the same letter are not significantly different (Tukey’s honestly significant difference (HSD) test, p < 0.01).
Figure 2.
Biotic traits of 38 wheat accessions, including 36 primary synthetics (PS), Langdon, and Norin 61. (A) Arbuscular mycorrhizal fungi (AMF) colonization % (B) Serendipita indica (S.i) colonization %. (C) Alternaria infection (%). Bars sharing the same letter are not significantly different (Tukey’s honestly significant difference (HSD) test, p < 0.01).
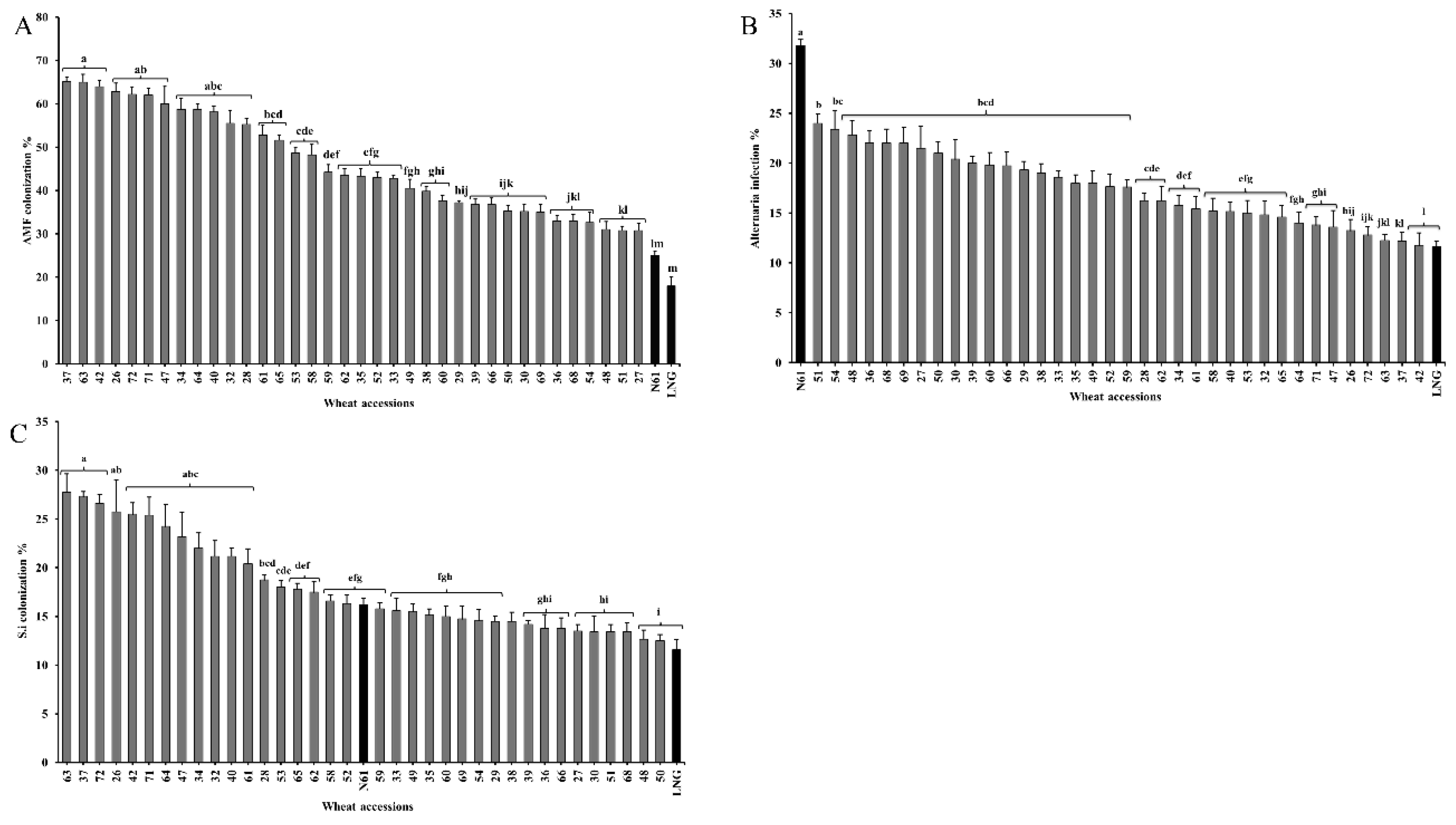
Figure 3.
Pearson’s correlation coefficients among root traits, shoot traits, and biotic traits (endophytic fungi colonization). Significance levels: *p < 0.05, **p < 0.01. AMF: arbuscular mycorrhizal fungi colonization (%); S.i: Serendipita indica colonization (%); Alternaria: Alternaria colonization (%), L_total: total root length (cm), L_thin: thin root (0 - 0.5 mm diameter) length (cm), L_thick: thick root (0.5-1.0 mm diameter) length (cm).
Figure 3.
Pearson’s correlation coefficients among root traits, shoot traits, and biotic traits (endophytic fungi colonization). Significance levels: *p < 0.05, **p < 0.01. AMF: arbuscular mycorrhizal fungi colonization (%); S.i: Serendipita indica colonization (%); Alternaria: Alternaria colonization (%), L_total: total root length (cm), L_thin: thin root (0 - 0.5 mm diameter) length (cm), L_thick: thick root (0.5-1.0 mm diameter) length (cm).

Figure 4.
Hierarchical clustering heatmap based on 11 standardized traits values across 38 wheat accessions, including 36 primary synthetics (PS), Langdon and Norin 61. Group shows the four accession groups classified by the clustering analysis. Soil Class shows the soil classes determined from GPS coordination of Ae. tauschii progenitor’s origin (NA: unknown soil class because GPS data was not found). AMF: arbuscular mycorrhizal fungi colonization (%), Alternari: Alternaria, colonization (%), L_total: total root length (cm), L_thin: thin root (0- 0.5 mm) length (cm), L_thick: thick root (0.5-1.0 mm) length (cm), Root_Fresh: root fresh weight (g), Root_Dry: root dry weight (g), Root_water: root water content (g), Shoot_fresh: shoot fresh weight (g), Shoot_dry: shoot dry weight (g), Shoot_water: shoot water content (g).
Figure 4.
Hierarchical clustering heatmap based on 11 standardized traits values across 38 wheat accessions, including 36 primary synthetics (PS), Langdon and Norin 61. Group shows the four accession groups classified by the clustering analysis. Soil Class shows the soil classes determined from GPS coordination of Ae. tauschii progenitor’s origin (NA: unknown soil class because GPS data was not found). AMF: arbuscular mycorrhizal fungi colonization (%), Alternari: Alternaria, colonization (%), L_total: total root length (cm), L_thin: thin root (0- 0.5 mm) length (cm), L_thick: thick root (0.5-1.0 mm) length (cm), Root_Fresh: root fresh weight (g), Root_Dry: root dry weight (g), Root_water: root water content (g), Shoot_fresh: shoot fresh weight (g), Shoot_dry: shoot dry weight (g), Shoot_water: shoot water content (g).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



