Submitted:
03 May 2025
Posted:
06 May 2025
You are already at the latest version
Abstract
Postbiotics, defined as non-viable microbial cells, their components, or metabolic byproducts, have gained recognition as promising modulators of gut health and disease, offering advantages over probiotics in terms of safety, stability and formulation. This systematic review explores the therapeutic potential of postbiotics derived from functional foods in the context of colorectal cancer (CRC), a leading cause of cancer-related mortality worldwide. Key bioactive categories of postbiotics, including short-chain fatty acids, exopolysaccharides, enzymes, cell wall fragments and cell-free supernatants are described, along with their mechanisms of action such as anti-inflammatory, immunomodulatory, and chemopreventive effects. Particular emphasis is laid on preclinical studies, in vitro, in silico, in vivo, ex vivo and advanced models such as organoids and organ-on-chip systems, that demonstrate how postbiotics may impact on tumorigenesis through pathways involving apoptosis, epigenetic regulation and gut barrier integrity. Recent advances in postbiotic delivery strategies are also summarized. Despite encouraging preclinical results, translation to clinical practice remains limited by the scarcity of human trials. This review draws attention to the need for further mechanistic research and robust clinical studies to validate the role of postbiotics as safe, effective adjuncts in CRC prevention therapy and management.
Keywords:
postbiotics
; colorectal cancer
; cancer
; functional food
; advanced preclinical models
; disease modelling
1. Introduction
In the last two decades the study and research on microbiota and its role and therapeutic potential in human health and disease have increased. The microbiota has been referred to as the “hidden organ”, with more than 100 trillion microorganisms inhabiting various body districts and contributing at a great percentage more genetic material than the entire human genome. Among the different microbiotas (e.g. respiratory, skin, oral, gut vagina …), the gut microbiota is considered particularly critical due to its profound influence on host metabolism, immunity, and disease susceptibility [1]. Increasing evidence supports the association between the gut microbiota dysbiosis, defined as an imbalance in the composition and species diversity of resident commensal microorganisms, associated with changes in metabolic activities or functions, and the pathogenesis of cardiovascular diseases, several types of cancer, respiratory diseases, diabetes and metabolic disorders, polycystic ovary syndrome, inflammatory bowel disease, neurological disorders, chronic kidney disease and liver diseases [2,3,4,5,6,7,8,9].
Food intake, nutritional status, dietary habits, and supplementation are key modulators of gut microbiota composition and activity. These factors not only determine microbial diversity and metabolic output but also critically impact host physiological functions, immune homeostasis, and disease susceptibility [10,11,12].
Recognized modulators of gut health are prebiotics (non-digestible food components, such as dietary fibers and oligosaccharides that promote the growth of beneficial commensal microorganisms) and probiotics, i.e. live microorganisms that, when administered in adequate amounts, confer a health benefit, can directly enrich and stabilize the gut microbial community. While probiotics and prebiotics have traditionally dominated this field, emerging attention is now directed toward postbiotics [13,14]. They are defined as non-viable microbial cells, their components, or metabolic products that confer health benefits to the host. The International Scientific Association for Probiotics and Prebiotics (ISAPP) recently defined postbiotics as "a preparation of inanimate microorganisms and/or their components that confers a health benefit on the host" [15,16,17]. The study on postbiotics has exponentially grown during these years as evidenced in Figure 1, in which the number of studies on postbiotics extracted from PubMed, Scopus and Web of Science in the last decade (2015-2024) were analyzed. Postbiotics have several advantages over probiotics, including greater safety, enhanced stability, and easier formulation. Their inanimate nature eliminates concerns regarding bacterial translocation, infection risk, and the transfer of antibiotic resistance genes, issues particularly critical for immunocompromised or critically ill populations. Moreover, postbiotics can exert diverse biological activities, such as anti-inflammatory, immunomodulatory, antioxidant, and anti-proliferative effects, which are increasingly being investigated in the context of chronic diseases [18,19,20].
In this complex scenario, this study aims to collect and review the role of functional foods that serve as rich sources of postbiotics, either through natural fermentation processes or via microbial biotransformation of dietary components, in a multifactorial disease like colorectal cancer (CRC) [21,22]. Despite recent advancements in the comprehension of its cellular and molecular mechanisms, CRC remains a major global health concern, ranking as the second leading cause of cancer-related deaths and the third most frequently diagnosed malignancy after lung and breast cancer. According to the Global Cancer Observatory (GCO) and the International Agency for Research on Cancer (IARC), nearly 2 million new cases of CRC were reported, with over 900,000 deaths. They account for approximately 9.3% of total cancer-related deaths. Mortality rates are notably higher in men than in women [23].
CRC is a complex disease associated with genetic predisposition and environmental risk factors. Key contributors include family history, hereditary CRC syndromes, inflammatory bowel disease, dietary habits, fiber deficiency, and obesity. Additionally, smoking, sedentary lifestyles, and increasing age have been strongly correlated with CRC incidence. The disease is often asymptomatic in its early stages, leading to late-stage diagnoses, which significantly impact treatment efficacy and patient survival [24]. Early detection through screening programs has proven to be a crucial strategy for reducing CRC incidence and mortality. Data from the European Commission (EC) CRC Statistics show that countries with long-standing, population-wide screening programs have experienced a significant decline in CRC incidence between 2000 and 2017, whereas those without widespread screening initiatives have reported either stable or rising cases. Among various screening tools, colonoscopy remains the most effective method for detecting and removing precancerous adenomas. However, due to its invasive nature, associated discomfort, and potential complications, its use as a routine screening method remains limited [25]. Pharmacological therapeutic interventions, both in a presurgical phase (neoadjuvant therapy) and after mass resection (adjuvant therapy) may have some limitations, such as severe adverse effect and chemoresistance, necessitating the exploration of novel complementary strategies [26].
For these reasons, this review aims to summarize the sources and the classes of probiotics from functional foods and then describe, using the methodologies of systematic review and metanalysis, the studies on postbiotics in CRC. Mechanisms of actions, including anti-inflammatory actions, modulation of tumor cell signaling pathways, and reinforcement of intestinal barrier integrity, through which postbiotics derived from functional foods may offer a novel, safe, and effective adjunct in CRC management are presented. This review deeply discusses properties, technological functions and health implications of postbiotics in CRC and lays a special emphasis on the research carried out using advanced preclinical study models (e.g., organoids, spheroids, organs-on-chip, ex vivo models …) as well as clinical microbiota-targeted interventions. Nevertheless, the translation of postbiotic-based interventions into clinical practice requires careful consideration of safety profiles, standardized production methods, and regulatory approval processes. While postbiotics demonstrate favorable stability and risk profiles compared to live biotherapeutics, challenges persist in the characterization of active components and in establishing robust clinical evidence of efficacy. As the field progresses, rigorous scientific validation and the establishment of clear regulatory frameworks will be pivotal for their successful integration into cancer prevention and treatment strategies.
2. Sources of Postbiotics and Classification
Although a harmonized classification has yet to be established, postbiotics are generally categorized into several types, including short-chain fatty acids, exopolysaccharides, cell wall fragments, cell-free supernatants, enzymes, and a variety of other metabolic byproducts (Figure 2) [27,28,29,30]. These bioactive compounds play a central role in regulating host health and initiating various physiological processes. Postbiotics can also be defined as structural components that include peptides, teichoic acids (TAs), and plasmalogens. These structural postbiotics contribute to maintaining microbiota integrity and functionality. They play a catalytic role in the preservation of the stability of microbial communities and the facilitation of host-microbe interactions [8,31]. Based on their chemical structure and function, postbiotics can also be categorized into carbohydrates (e.g., TAs and galactose-rich polysaccharides), proteins, lipids (including butyrate, acetate, and dimethyl acetyl-derived plasmalogens which are membrane phospholipids), vitamins (primarily B-group vitamins), organic acids (such as 3-phenyllactic acid and propionic acid), other complex biomolecules (including lipoteichoic acids, LTAs) and peptidoglycan-derived muropeptides) [32,33]. These various postbiotics exhibit a wide range of biological activities, contributing to host health and the stability of the microbiota. Their effects based on physiological functions include antioxidant, anti-inflammatory, anti-proliferative, anti-obesogenic, hypocholesterolemic, and anti-hypertensive activities [28,34].
2.1. Short Chain Fatty Acids
Short-chain fatty acids (SCFAs), which are saturated aliphatic organic acids containing between one and six carbon atoms, represent a key group of microbial metabolites. They are introduced through the diet and primarily produced by intestinal bacteria via the fermentation of non-digestible carbohydrates, such as dietary fibers [35,36]. The cecum contains the highest concentration of SCFAs, although their levels vary throughout the intestinal tract. The most physiologically significant comprise acetate, propionate, and butyrate, which account for approximately 85–95% of the total SCFAs present in the colon [37,38]. Microbial fermentation of dietary fiber in the proximal colon produces these latter compounds and can control the metabolism by increasing gut hormone secretion and suppressing appetite [39]. Interestingly, Bacteroidetes mainly produce acetate and propionate, whereas Firmicutes are major producers of butyrate [40]. Acetate is the most abundant SCFA, predominantly generated as an end-product of fermentation by enteric bacteria. Besides, it can also be produced from formate through the Wood-Ljungdahl pathway by hydrogenotrophic bacteria, such as Acetobacterium woodii [41]. Through the fermentation of nondigestible carbohydrates, Lactobacillus and Bifidobacterium could generate organic acids, like lactic acid. Under normal gut conditions, there is cross-feeding between bacteria, whereby species such as Eubacterium hallii further convert lactic acid into SCFAs [42]. Nevertheless, this affiliation is not comprehensive, since SCFA production is not severely restricted to specific bacterial phyla or genera.
KetoA (10-oxo-12(Z)-octadecenoic acid), a metabolite of linoleic acid, which is synthesized by gut lactic acid bacteria (LAB), constitutes another interesting postbiotic. This metabolite was shown to be efficient at enhancing energy expenditure and metabolic health with therapeutic potential for the management of metabolic disorders [43]. In order to produce SCFAs, prebiotics (e.g., inulin and fructooligosaccharides) are fermented by the gut microbiota and acetate, propionate, and butyrate are the principal end products [44]. These SCFAs are typically present in the colon and feces at an approximate molar ratio of 60/20/20, respectively. By comparison, glycolysis yields pyruvate, which can be further metabolized by some Lactobacilli species through fermentation to yield SCFAs. In addition, under heterofermentative conditions, these bacteria also use the phosphoketolase pathway as an alternative pathway for SCFA biosynthesis [45]. For example, Bifidobacteria have varied metabolic flexibility based on the availability of nutrients. Under carbohydrate excess, they produce largely acetate and lactate to support rapid growth. Under carbohydrate limitation, they shift metabolic strategy, however, and produce acetate and formate via committed fermentation pathways.
2. 2. Exopolysaccharides
On the basis of their chemical structure, exopolysaccharides (EPSs) are generally divided into two broad categories: (i) homopolysaccharides, including levan, cellulose, pullulan, curdlan, and dextran and composed of one type of monosaccharide unit; and (ii) heteropolysaccharides, such as gellan, xanthan, kefiran, and galactan with repeating units of 2 or more different monosaccharides [46].
These structures reflect the structural heterogeneity of EPS, which fulfill critical functions in microbial physiology and microbe–host interactions. The EPS structural complexity was affected by key microbial processes like biofilm formation, adhesion, protection against environmental stresses, and retention of nutrients. EPS also arbitrate key interactions with host organisms, counting immunomodulation, colonization, and the formation of symbiotic or pathogenic relationships [47]. Synthesized by LAB, EPS exhibited an extensive structural diversity, which is the foundation of their wide variety of biological activities. Their structural diversity enables EPS to portray a range of health-beneficial activities including antimutagenic, antitumor and immunomodulatory and activities. Besides, many LAB-derived EPS are strong antioxidant, antibacterial, antihypertensive, anti-inflammatory, and antiviral agents, and thus are excellent candidates for application in nutraceuticals and functional foods, and therapy regimens [48]. Certain EPSs yielded by Lactobacillus strains, selected from fermented durian fruit, have shown illustrious antioxidant and antimicrobial activities, corroborating their likely health-promoting effects. Aside from their biofunctional activities, the EPSs were also shown to beneficially affect lipid metabolism through the inhibition of cholesterol uptake in the gastrointestinal tract, hence supporting cardiovascular health and reducing the risk of hyperlipidemia [49]. Obtained from Lactococcus lactis subsp. Lactis, EPS showed very good antioxidant activity. This activity was supported by their ability to improve glutathione peroxidase, superoxide dismutase and catalase activities of these key antioxidant enzymes. Additionally, the EPS treatment was able to reduce the levels of lipid peroxidation in both the serum and liver mice tissues, which indicate their potential in protecting against and maintaining metabolic health and oxidative damage [50]. Originating from Lactobacillus reuteri Mh-001, EPS exhibited unprecedented immunomodulatory activities, signaling their promise to promote immune health [51]. Beyond the regulation of immune response, EPSs are also lengthily characterized for anti-inflammatory and antioxidant activities that were extremely well-studied in a wide range of biological systems. Apart from their potential bioactivity, EPSs are also of great interest in the food industry, where they find functional applications such as natural stabilizing, emulsifying, and water-binding agents contributing to the texture, shelf-life, and overall quality of various food products.
2.3. Enzymes
On the basis of their catalytic action, enzymes are classified into 6 broad categories: transferases, oxidoreductases, lyases, hydrolases, isomerases, and ligases, each catalyzing a different type of biochemical reaction [52]. These molecules perform a broad array of tasks in biological systems, extending from catalyzing biochemical reactions, and governing metabolic pathways to assist the essential physiological functions.
Industrial application of enzymes was mainly associated with a well-characterized microbial source, more exactly bacterial strains such as Bacillus subtilis and Bacillus licheniformis, and fungal strains such as Aspergillus niger and Aspergillus oryzae [53]. Their GRAS status (Generally Recognized As Safe) makes them highly suitable for applications in food, pharmaceutical, and biotechnological processes. Kim et al. reported that 2 strains of Lactobacillus fermentum produced high glutathione peroxidase concentrations with effective in vitro antioxidant activity. Antioxidative potentials of human-derived Lactobacillus strains were assessed in vitro. Lb. acidophilus KCTC 3111 showed the strongest inhibition of lipid peroxidation (50% in whole cells, 65% in lysates), strong hydroxyl radical scavenging activity, and viability in 1.0 mM hydrogen peroxide for 7 hours. L. brevis KCTC 3498 showed the highest glutathione peroxidase activity, the strain L. acidophilus 3111 demonstrated good enzymatic antioxidant activity [54]. As another example, proteases from Bacillus species are gaining increased attention as postbiotic entities due to their potent bioactivities and stability under gastrointestinal conditions. These enzymes, which are excreted via fermentation or microbial metabolism, continue to be active in the absence of viable cells, which meets the criteria of postbiotics as non-viable microbial products conferring health benefits to the host. Bacillus subtilis and Bacillus licheniformis are well recognized to produce healthy neutral and alkaline proteases that are extremely heat-resistant, pH, and digestive enzyme-resistant, thereby qualifying them as potential candidates for oral delivery in functional foods and supplements. In addition to their role in protein hydrolysis, these proteases exert health benefits through the generation of bioactive peptides with antioxidant, antimicrobial, anti-hypertensive, and immunomodulatory activities. Moreover, they can enhance protein digestibility, modulate gut microbiota, and support intestinal barrier function—main ways in which postbiotics exert beneficial effects. The use of Bacillus proteases as postbiotics is a growing field of research within microbiome-based therapies and functional nutrition, particularly since they are simple to produce, GRAS status, and their industrial scalability has already been demonstrated [30]. Catalase, which decomposes hydrogen peroxide into water and oxygen, is an enzyme crucial for lowering oxidative stress, a factor in numerous health disorders. In this regard, recent research has pinpointed the emerging potential of catalase-producing microorganisms as postbiotics with tremendous therapeutic benefits mediated by their antioxidative effects. It was demonstrated that strains that produce catalase, i.e., Lactococcus lactis, can decrease oxidative damage in the colon, thereby restricting inflammation and, in turn, the development of CRC in mice [55].
Catalase has been recently promoted as a postbiotic that can modulate oxidative stress and improve gut health. For instance, supplemental dietary administration of exogenous catalases derived from Penicillium notatum in weaned pigs was shown to enhance intestinal antioxidant defenses in the pig by alleviating lipopolysaccharide (LPS)-induced oxidative damage, where LPS is a pro-inflammatory endotoxin. The supplement not only reduced oxidative stress markers but also beneficially modulated the gut microbiota population, which cumulatively contributed to improved intestinal barrier function and overall health status [56]. Similarly, the dietary addition of catalase in broiler chickens challenged with deoxynivalenol (DON), a common mycotoxin, led to the reduction of intestinal oxidative stress by a significant amount, improvement in villus morphology, and reestablished microbiota homeostasis. Such effects suggest that catalase, either supplied through microbial fermentation or as a pure enzyme, performs effectively as a postbiotic by blocking ROS-mediated damage and supporting gut ecosystem resilience [57]. Such findings show the effect of catalase as a postbiotic molecule for promoting gut health and preventing oxidative stress disorders. Inclusion of catalase-producing microorganisms or exogenous catalase in functional foods or supplements can introduce a novel approach to enhancing health status, particularly in subjects at risk of oxidative stress-related disorders [58].
2.4. Cell Wall Fragments
Recent studies have highlighted the noticeable role played by probiotic bacterial cell wall remnants in gut health promotion and immune modulation. These remnants, e.g., peptidoglycan and lipoteichoic acid, are released during bacterial growth or lysis and can exert postbiotic activity in the absence of live cells [59]. Peptidoglycan fragments, such as muramyl dipeptide, were demonstrated to modulate immune responses by regulating cytokine secretion and activating Toll-like receptors (TLRs) as pattern recognition receptors. For example, Lactobacillus rhamnosus CRL1505 peptidoglycan could have enhanced the resistance to Streptococcus pneumoniae infection by normalizing the serum level of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6. Other than immune modulation, cell wall fragments can have anti-inflammatory and antioxidant effects. As an example, lipoteichoic acid from Lactobacillus plantarum has been reported to induce the production of TNF-α and the phosphorylation of NF-κB-p65, p-38, and JNK, thereby regulating immune responses [59]. These results highlight the promise of cell wall fragments as postbiotics in gut health promotion, immune modulation, and enhancement of the activity of other bioactive substances. Their use in the formulation of functional foods or supplements may present a new opportunity to enhance health status, especially in populations at risk of gastrointestinal diseases or chronic illnesses. Jung et al. (2022) purified LTAs from 4 Lactiplantibacillus plantarum strains named K8, K88, K5-5, and K55-5, and compared their immunostimulatory activities. The authors established those structural variations of LTAs between the strains greatly influenced their ability to modulate immune responses. More specifically, differences in glycosylation patterns, D-alanine substitutions, and molecular weights of the LTAs were found to modulate the immune cell activation, i.e., of macrophages. For instance, certain types of LTAs induced stronger expression of pro-inflammatory cytokines like TNF-α and IL-6, while others had milder or more modulatory effects. These findings highlight the strain specificity of postbiotic activity and reveal that not all LTAs of L. plantarum trigger the identical immunological responses, underlining the value of structural characterization in the development of targeted postbiotic therapies [60].
2.5. Cell Free Supernatants
Cell free supernatants (CFSs) from LAB characterize a heterogeneous postbiotics class with multilayered roles in promoting health and posing sustainable substitutes to conventional preservers and antibiotics. Current investigations have stressed the efficacy of LAB-derived CFS against a range of pathogenic microorganisms. For example, CFSs from LAB strains like Lb. aacidophilus, Lb. rhamnosus, and Lactiplantibacillus plantarum have established potent antibacterial effects against foodborne pathogens such as Salmonella and E. coli. These CFSs displayed antimicrobial activity even after lyophilization or neutralization, signifying their robustness and potential as natural preservatives in animal feed and food products [61]. Furthermore, CFSs have shown potential in combating biofilm formation, a common challenge in long-lasting contaminations and food spoilage. LAB-derived CFSs can disturb biofilm integrity and constrain the expression of virulence factors in pathogens like P. aeruginosa, S. aureus, and Candida species, indicating their potential in therapeutic and food safety applications. Beyond antimicrobial activities, LAB-derived CFSs possess antioxidant properties, which can be beneficial in preventing oxidative stress-related diseases. Additionally, studies have demonstrated that CFS from probiotic strains such as Lactobacillus casei and Lactobacillus rhamnosus GG can inhibit colon cancer cell invasion by modulating matrix metalloproteinase-9 (MMP-9) activity and enhancing tight junction protein levels, thereby contributing to cancer metastasis prevention [62].
2.6. Postbiotic Functional Food Sources
2.6.1. Sauerkraut (Fermented Cabbage)
Cabbage fermentations resulted in consistent metabolome profiles enriched in bioactive compounds known to be important postbiotics. They could also serve as metabolites of human gut microbiome, including D-phenyl-lactate (D-PLA) and indole-3-lactate (ILA) [63]. A popular fermented vegetable that appears with different names (e.g., sauerkraut, suan cai, curtido, etc.) is a fermented cabbage [64]. Preparation includes mixing of shredded cabbage with 2%–3% (wt/wt) sodium chloride and incubation at room temperature for 2–3 weeks with minimal exposure to ambient oxygen. Selection for LAB arises from the lack of oxygen and the presence of salt during cabbage fermentation. Phylogenetically related bacteria in the Bacillota (formerly Firmicutes) phylum are encompassed by LAB and due to their saccharolytic fermentation energy conservation metabolism they result in lactic acid as the only or the main (> 50%) end-product. There is a change of the dominant LAB in cabbage fermentations over time. It starts with heterofermentative species, such as Leuconostoc mesenteroides and Weissella spp., followed by Lactiplantibacillus plantarum and Levilactobacillus brevis after a couple of days. These heterofermentative bacteria metabolize glucose to generate lactic acid, acetic acid, carbon dioxide, and ethanol, which initiates the fermentation process [65]. As the pH decreases, the conditions become suitable for homofermentative LAB such as Lactiplantibacillus plantarum and Lactobacillus brevis, which primarily create lactic acid during fermentation. This process further decreases acidity and enhances the sauerkraut stability [66]. By extending the shelf life of sauerkraut, it becomes a preserved and safe fermented food item [67].
Lactic and acetic acids, mannitol, and CO2, are the primary secreted metabolic end-products produced by LAB. The latter break down sugars and other compounds and sometimes reach 2% (vol/wt) or more in the ferments [68]. Fermentation enhances the bioavailability of vitamins and minerals in cabbage, and generates antioxidants and probiotics, which contribute to the health advantages of sauerkraut [69].
Besides their antimicrobial effects and sensorial properties of fermented foods, lactic acid and acetic acid compounds are associated with improvement of intestinal barrier function [70,71,72], immune function [73], and metabolic health (e.g., fasting blood glucose and insulin sensitivity) [72,74,75].
Cabbage fermentations guided by LAB show higher concentrations of phenolic compounds (polyphenols, phenolic acids) [76], carotenoids [77], glucosinolate breakdown products (ascorbigen, indole-3-carbinol, and isothiocyanates) [78,79], and other bioactive metabolites [80]. Intestinal barrier-protective properties following production of microbiota-associated D-PLA and ILA are the main advantages of the amino acid derivatives, D-PLA [81] and ILA [82], [83,84,85]. The activation of signaling pathways involving PPAR-γ (D-PLA) and AHR (ILA) exert these effects. PLA and ILA along with lactic acid are significant for the immunomodulatory, antioxidant, and anti-carcinogenic properties of fermented cabbage [79,80].
2.6.2. Kefir
Kefir is a fermented drink with low alcohol content, acidic and bubbly from the fermentation carbonation of kefir grains with milk or water [86,87]. The starter is the kefir grains. Kefir grains range in size from 1 to 4 cm in length and look like small cauliflower florets in shape (irregular and lobed-shaped) and color (from white to light yellow). This gelatinous and slimy structure is comprised of kefiran, a natural matrix of EPSs and proteins. In this matrix LAB, yeasts, and acetic acid bacteria (AAB) co-exist in symbiotic connection [86].
The most pre-dominantly found bacterial species in kefir grains are Lactobacillus kefiranofaciens, Lacticaseibacillus paracasei, Lactiplantibacillus plantarum, Lactobacillus acidophilus, and Lactobacillus delbrueckii subsp. bulgaricus. The predominant yeast species present in kefir are Saccharomyces cerevisiae, S. unisporus, Candida kefyr, and Kluyveromyces marxianus ssp. marxianus [88].
Kefiran is the postbiotic from kefir that has shown potential benefits in alleviating food allergies by modulating both the intestinal microbiome and the immune system [89]. This water-soluble branched glucogalactan postbiotic showed also antimicrobial and healing activity [90]. Other postbiotics from kefir LAB include surface layer proteins (SLP) and various EPS that may have a beneficial role in gut dysbiosis and obesity management [91].
This microbial diversity is responsible for the physicochemical features and biological activities of each kefir, also comprising some major Lactobacillus species due to their probiotic strain-specific properties. Some of the most significant health benefits from kefir beverage consumption include anti-microbial, anti-tumor, anti-carcinogenic, hypocholesterolemic effects. In addition, anti-hypertensive, anti-diabetic, immunomodulatory activities along with improvement of lactose digestion are reported. Kefir exhibits excellent protein content and appears in two distinct forms: intact protein and partially digested protein [92]. These forms facilitate its utilization as prebiotic, probiotic [93].
2.6.3. Kimchi
Kimchi, a typical fermented Korean vegetable food, is considered as one of the most important postbiotic foods, since it contains Lactiplantibacillus plantarum, a homofermentative LAB type producing organic acids, and Leuconostoc mesenteroides, a heterofermentative type producing carbon dioxide, acids and weak alcohols [94]. The flavor (taste and aroma) of the product are controlled by these bacteria. Sugars are transformed into lactic acid, and this process characterizes yogurt, kimchi, and fermented cereals. This fermentation is primarily driven by LAB [95].
3. Mechanisms of Action of Postbiotics in Colorectal Cancer
It is well known that dietary intervention may have a great role in modulating human health and disease. In literature many bioactive compounds have been described to have strong benefits on CRC and modulation of gut microbiota [20]. As an example, Ou J et al. explored the impact of diet on colon cancer risk. They analyzed gut microbiota metabolites in individuals at high risk (African Americans) and low risk (rural native Africans) for colon cancer. Their research revealed significant links between a decreased production of SCFAs, elevated levels of secondary bile acid metabolites, and an increased risk of colon cancer [96]. Postbiotics may exert anti-carcinogenic and chemopreventive effects in CRC through a range of interrelated mechanisms involving modulation of epithelial cell signaling, immune and inflammatory responses, and microbial-host interactions. Although several mechanisms are similar to that of prebiotics, postbiotics offer a higher safety profile, better generalizability, longer shelf life, and more rapid biological activity. However, further larger studies are needed to fully elucidate their mechanisms and optimize their therapeutic potential.
3.1. Anti-Inflammatory and Immunomodulatory Effects
Chronic inflammation is a hallmark of CRC pathogenesis [97]. Several postbiotic compounds, in particular SCFAs, downregulate pro-inflammatory signaling cascades, acting as chemopreventive agents. As an example, butyrate suppresses nuclear factor kappa B (NF-κB) activation, reducing the expression of pro-inflammatory cytokines like TNF-α, IL-6, and IL-1β. This downregulation of inflammation mitigates a tumor-promoting microenvironment. Similarly, SCFAs may modulate the mitogen-activated protein kinase (MAPK) pathway, leading to decreased proliferation and enhanced apoptosis of transformed colonocytes [3,98]. In IEC models, in tissue samples of the patients with CRC and in mouse colon, butyrate suppresses LPS-induced NF-κB activation via GPR109A [99]. One study explored the role of EPSs in modulating inflammation. EPSs bind to the TLR2 receptor of intestinal epithelial cells, thus leading to the inhibition of IL-17 production, and promotion of IL-10 expression [100].
Moreover, butyrate and SCFAs maintain intestinal immune homeostasis, mainly through the modulation of regulatory T cells (Tregs). The proliferation of CD4+ T lymphocytes is limited by Tregs, and are therefore essential for suppressing inflammatory and allergic responses [101].
A variety of gastrointestinal disorders and diseases is associated with intestinal barrier dysfunction induced by toxins and pathogens. SCFAs, in particular butyrate, has a protective action of the barrier. In several studies, measurement of transepithelial electrical resistance (TER) and inulin permeability in Caco-2 and T-84 cells determined the effect of butyrate on the intestinal barrier. Results show that activation of AMP-activated protein kinase (AMPK) is carried out by promotion of the formation of the intestinal barrier by butyrate [102,103].
The aryl hydrocarbon receptor (AhR), activated by indole derivatives, promotes the secretion of interleukin-22 (IL-22) and interleukin-10 (IL-10) while reducing the expression of pro-inflammatory cytokines such as TNF-α and IL-6 [104,105]. Another study highlighted a novel mechanism in which Lactobacillus reuteri and dietary tryptophan interact to regulate gut immunity via AhR activation. L. reuteri produces indole derivatives from tryptophan, which activate AhR in CD4⁺ T cells. AhR activation leads to the downregulation of ThPOK, a transcription factor that enables CD4⁺ T cells to transition into DP IELs [106].
Anti-inflammatory and immunomodulatory functions are exerted from SLPs derived from various Lactobacillus species. SLPs isolated from Propionibacterium fischeri reduced TNF-α and IL-8, inversely induced a significant increase of expression TFG-β levels in HT-29 cells [107]. In a similar study SLPs from Lactobacillus helveticus MMLh5 exerts anti-inflammatory effects by reducing levels of both basal and induced NF-κ B in the presence of proinflammatory stimulus IL-1 B in the cell lines of Caco-2 [108]. Moreover, SLPs from Lactobacillus plantarum reversed intestinal epithelial cell damage induced by pathogenic E. coli [109].
TAs play a role in immune modulation for simultaneous decrease in IL-12 and production of IL-10 in animal models. The effect of these TAs on IL-10 production was mediated by TLR2-dependent ERK activation [110]. Additionally, regulation of Treg T cell function is carried out along with suppression of intestinal inflammation, maintenance of homeostasis in the intestine, and exertion of antitumor and antioxidant effects [111]. Conversely, another study has reported that lipoteichoic acid induces intestinal immune activation, resulting in the production of TNF-α and IL-12, which contribute to intestinal mucosal damage [112].
Furthermore, postbiotic CFSs downregulate pro-inflammatory pathways by reducing pro-inflammatory factors and chemokines in dendritic cells targeted by Salmonella typhi. This occurs through TLR activation, which enhances protection against highly infectious pathogens [62].
In an interesting study, it was shown that ILA, metabolite of Lactobacillus plantarum, improved intestinal inflammation, dysbiosis and slowed down tumor growth. Experiments were carried out on adenocarcinoma cell lines and animal models. The production of IL12a in dendritic cells was promoted by ILA by facilitating the increase of H3K27ac binding at IL12a enhancer regions, thereby aiding in the activation of CD8⁺ T cell (cytotoxic T lymphocyte, CTL) responses against tumor development. Additionally, suppression of Saa3 expression, linked to cholesterol metabolism in CD8⁺ T cells, was observed, likely due to alterations in chromatin accessibility, which in turn enhanced the functionality of CD8⁺ T cells infiltrating tumors i.e. killing cancer cells [113].
Yan et al. isolated and purified proteins secreted by the probiotic Lactobacillus rhamnosus. Two key proteins, p75 (75kDa) and p40 (40kDa), were used to treat human (HT-29) and mouse (YAMC) intestinal cells and murine ex vivo colon samples. The p75 and p40 proteins initiate the PI3K/Akt pathway, inhibit TNF-α induced apoptosis and pro-inflammatory cytokines, stimulate the proliferation of intestinal epithelial cells, and reduce epithelial damage induced by TNF. These results suggest that these proteins might prevent gastrointestinal diseases mediated by cytokines [114]. Bäuerl et al. demonstrated that the extracellular vesicles isolated from L. casei have on their surface the proteins P40 and P75. T84 cells were treated with P40 and P75 vesicles, which induced EGFR phosphorylation in a dose-dependent manner, showing their anti-inflammatory and antiapoptotic properties [115]. Five LAB strains from Korean kimchi were reported to have a protective role in inflammatory responses. The expression of VDR and autophagy was measured by treating human cell lines HCT-116 and intestinal organoids with conditional medium (CM) from these strains. The LAB-CM-treated groups showed higher mRNA expression of VDR and its target genes cathelicidin compared with the control group [116].
3.2. Apoptosis Induction and Tumor Suppression
Postbiotics represent a promising frontier in cancer research due to their ability to induce apoptosis and suppress tumor growth through multiple biological pathways [117]. The mechanisms behind their anticancer effects include activation of pro-apoptotic pathways, such as Bax and Bcl-2, regulation of p53 and activation of Caspase 3 and 9 and suppression of tumor migration and invasion. As an example, the cytotoxic effects on HT-29 cells of two heat killed probiotic bacteria were evaluated. Lactobacillus Brevis and Lactobacillus paracasei were isolated from a novel food from the Iranian diet 'terxine'. Both bacteria inhibited the proliferation and induced apoptosis of HT-29 cells, increasing the expression of m-RNA of caspase-3 and 9 and reduced BCL-2 were evaluated [118]. Konishi et al. demonstrated in animal models that ferrichrome suppresses tumor growth both in vivo and in vitro by triggering apoptosis (activation of caspases) and inducing mitochondrial dysfunction [119].
In several studies, it was investigated the effect and mechanism of butyrate on apoptosis, in CRC models. Butyrate was shown to activate the intrinsic apoptotic pathway by upregulating pro-apoptotic proteins like Bax and downregulating anti-apoptotic proteins like Bcl-2 [120] and via the deactivation of mTOR/S6K1 signaling [121]. Similarly, Ryu et al. demonstrated that another SCFA, propionate, downregulated of a histone arginine methyltransferase, PRMT1 in the HCT-116 cell line. This downregulation induced apoptosis by inhibiting phospho-p70 S6 kinase. To further validate the anticancer effect of propionate, cell growth assays using crystal violet staining were conducted (0 and 5 mM) in both normal enterocytes and the HCT-116 cell line. Notably, no significant effect of propionate treatment was observed in normal enterocytes [122].
3.3. Other Effects
Some chemopreventive and therapeutic effects were described in literature for several postbiotics. Butyrate is a well-characterized HDAC inhibitor (epigenetic modulation) [123]. In CRC cells, it induces histone hyperacetylation, leading to the transcriptional activation of tumor suppressor genes, thereby promoting cell cycle arrest and apoptosis. The context-specific “butyrate paradox” illustrates this: while butyrate supports proliferation of healthy colonocytes, it inhibits proliferation and induces apoptosis in CRC cells due to altered metabolic states (Warburg effect) [124,125].
4. Therapeutic Potential and Application in Biomedical System: Current Evidence from Preclinical Studies
4.1. Methodologies and Software
This In this section the selection of the studies to be analyzed was carried out using the methodologies of Extensive literature search (ELS) and the principles of the PRISMA (The Preferred Reporting Items for Systematic reviews and Meta-Analyses) guideline - 2020 version were used [128,129,130]. The initial searches were performed on three databases, PubMed, Scopus and Web of Science. All keywords and terms were selected using the MeSH (Medical Subject Headings) vocabulary. The primary strings included “postbiotic AND colorectal cancer”, “postbiotic AND colorectal cancer OR tumor”, followed by secondary keywords in order to retrieve all studies, such as “butyrate”, “p40”, “p70”, “cell free supernatant”, “Lactobacillus”, “bacteriocins”, “exopolysaccharide”, “colon organoids”.
The literature review was conducted following the methodology outlined in the referenced guidelines. Following removal of duplicate records, screening of titles and abstracts was carried out based on the inclusion and exclusion criteria detailed in Table 1. The search results from the ELS were imported into EndNoteWeb (https://web.endnote.com/), where initial selection was carried out using title and abstract screening, followed by full-text assessment. Only studies that satisfied the eligibility criteria proceeded to the next phase of the review, while those unrelated to the research focus or not meeting the inclusion standards were excluded.
The results obtained are summarized in Figure 4. The records were classified on the basis of the models used to conduct the study (in vitro, in silico, in vivo, ex vivo, advanced preclinical patient-derived models), postbiotics and microorganisms of derivation. Key findings and pathways and genes studied were reported. No clinical studies were found.
4.2. Studies on Cell Lines
A wide range of in vitro studies investigated mechanisms of action and future possible benefits for CRC patients of postbiotics, including the production of organic acids, enzymes, peptides and polysaccharides during the fermentation process. These studies provide mechanistic insights into how postbiotics may exert cytotoxic, pro-apoptotic, anti-proliferative, and immunomodulatory effects on CRC cell lines.
CFSs from Lactobacillus casei and Lactobacillus rhamnosus GG strains were reported to induce apoptosis and inhibit proliferation in HCT-116 CRC cells by decreasing matrix metalloproteinase-9 (MMP-9) and increasing the tight junction protein zona occludens-1 (ZO-1) levels [131]. The CFSs were subsequently fractioned, and the most active fractions were identified (>100 kDa and 50–100 kDa). Similar results were obtained by Elham et al. on Caco-2 cells [132].
Chen et al. demonstrated that damage in HT-29 cell membranes was caused by supernatants of seven strains of Lactobacillus at high concentrations. The PM177 strain showed the most potent inhibitory effect followed by PM153 and BCRC14625. Among the strains studied, BCRC17010 stood out for the best antitumoral potential, thanks to induction of apoptosis, releasing lactate dehydrogenase (LDH) and producing nitric oxide (NO) [133].
Similarly, Jastrząb et al. identified Lactococcus lactis subsp. lactis Lc4 as a promising therapeutic strain capable of releasing arginine deiminase into its supernatant, which exerts strong anti-proliferative effects on CRC cells by depleting arginine and inducing cell cycle arrest [134].
Luo et al. investigated the mechanism of sodium butyrate (NaB)-induced autophagy in CRC cell lines HCT-116 and HT-29, and their findings suggested that NaB treatment increased the formation of autolysosome and expression of phosphorylated liver kinase B1 (LKB1), AMP-activated protein kinase (AMPK) and acetyl-CoA carboxylase (ACC). In particular, LKB1 and AMPK are critical for NaB-mediated autophagy [135].
Recent studies reported on bacteriocins as postbiotic metabolites from Lactobacillus plantarum strains showing potent selective cytotoxicity effects, via anti-proliferative mechanisms and induction of apoptosis in HT-29 cells, without affecting healthy cells [136]. Selective inhibition of store-operated Ca²⁺ entry (SOCE) in CRC cells by downregulation of Orai1 and STIM1, thereby impairing cell migration might also be evident by postbiotics from Lacticaseibacillus paracasei and Lactiplantibacillus plantarum [137].
Similarly, EPSs derived from Lactobacillus acidophilus was found to inhibit the growth of Caco-2 colon cancer cell line in a dose-dependent manner and both under normoxia and hypoxia [138].
Cousin et al. showed that induction of intrinsic apoptosis of HT-29 and HCT-116 cells, alone and enhancement of the activity of the apoptosis inducer, TNF-Related Apoptosis-Inducing Ligand (TRAIL) could occur via propionate and acetate from Propionibacterium freudenreichii ITG-P9 [139].
In another study, LS174T human colorectal cells were treated with butyrate related to beneficial probiotics (Lactobacillus and Bifidobacterium) species at various concentrations. It positively influences mucin secretion with increased protein content (peak effects at 6 or 9 mM), which enhanced adherence of probiotic strains, and stimulates the MAPK signaling pathway in intestinal cells, increasing gut defense [140]. CFSs from Bifidobacterium longum were also tested for anti-cancer properties in a study using Fn-secreted extracellular vesicles (Fnev)-infected CRC cells with controversial results [141].
A preliminary in silico analysis guided the in vitro studies of Erfanian et al., which reported that Lactobacillus acidophilus postbiotics may exert their anti-proliferation and anti-migration activities via Wnt signaling pathway (RSPO2, NGF, MMP7 and SFRP1, SFRP2, SFRP4, MMP7) [142]. Similar results were obtained in another study of these authors on postbiotics from Bifidobacterium breve and Lactobacillus rhamnosus [143].
Another study evaluated the safety, antimicrobial, and anticancer effects of cell-free metabolites from Gluconobacter oxydans strains, isolated from Kombucha, as potential postbiotics. Using five AAB strains and three human cell lines, including HT-29, the results confirmed the safety and functional potential of selected AAB strains. The KNS30 strain showed the strongest anticancer activity on gastric cancer [144].
4.3. In Vivo Studies
The effectiveness of postbiotics in CRC prevention and therapy in vivo has been evaluated by several animal studies. Although limited, in vivo studies provide complementary evidence of postbiotic efficacy in CRC models. As an example, early-life supplementation with p40, a protein derived from Lactobacillus rhamnosus GG, was shown to enhance intestinal development and immune function in mice. Delivered via hydrogels, p40 promotes epithelial growth, tight junction formation, and IgA production through EGFR activation. These effects are absent without EGFR, highlighting its key role. Early p40 treatment also improves resistance to gut injury and inflammation in adulthood, suggesting long-term health benefits from probiotic-derived factors [145].
Sharma and Shukla reported the mitigation of early-stage colon cancer development in Sprague–Dawley rats by the CFS of Lactobacillus rhamnosus MD 14 MH656799, which includes acetamide, acetate, propionate, butyrate, thiocyanic acid, and oxalic acid compounds,. The protective effects were associated with a reduction in fecal procarcinogenic enzyme activity, oxidative stress, and aberrant crypt foci, alongside the suppression of oncogenes like β-catenin, K-ras, Cox-2, and NF-κB, and the upregulation of the tumor suppressor gene p53, resulting in near-normal colon tissue structure [146].
Lactobacillus casei ATCC334 supernatant, where ferrichrome is subsequently identified as the responsible molecule that induced apoptosis in CRC cells, exhibited minimal effect on normal intestinal epithelial while having stronger antitumor activity than conventional CRC drugs (5-FU and cisplatin). The findings were confirmed using xenograft models obtained by injecting SW620 cells into male BALB/c nude mice [119].
EPS application in postbiotics therapy was investigated by Ma et al., suggesting that Lactiplantibacillus plantarum-12 EPS oral administration in mice can reduce colon cancer symptoms through gut microbiota and metabolites modulation [147].
Furthermore, a new type of therapeutic strategies could lead to the use of bacterial lysates, given that L.acidophilus cell lysates, combined with immunotherapeutic antibodies, anti- CTLA-4, may help in forming the anticancer immune response in CRC-induced murine models; as resulted, the combined administration leads to meaningful inhibition of increased amounts of proteobacteria and somewhat modulates the CRC-induced dysbiosis [148].
In a promising and advanced preclinical study, Lee et al. characterized the synergic effect of MS-20 Symbiota®, a mixture of microbial metabolites generated from fermentation of a soybean-based medium with multiple strains of probiotics and yeast, with anti-PD1 antibody therapy in xenograft mouse models, showing an enhanced efficacy on tumor growth. This study also used fecal samples from CRC patient for an ex vivo treatment to confirm the modulation of immune checkpoint inhibitor (ICI)-responsive bacteria [149]. A similar study on AOM/DSS mouse model of CRC investigated the role of potential postbiotic putrescine that reduced the number and size of colonic tumors, and downmodulated the release of inflammatory cytokines in the colonic lumen [150].
Other studies tested the anti-proliferative/migration effect modulated by apoptosis, cell cycle arrest and autophagy. As an example, Zhong et al. evaluated MZY531 postbiotic and its tumor growth inhibition effect, in xenograft mouse model, through Bax/Bcl2/caspase-3 & JAK2/STAT3 mediated apoptosis and PI3K/AKT/mTOR & TGF-β/SMAD4 mediated autophagy with interesting results [151].
4.4. Investigating Postbiotic Safety and Effects Using Advanced Preclinical Models
Conventional in vitro 2D cell cultures and co-culture systems were extensively used during the last decades to predict cell behavior, morphology, physiology and molecular responses. However, these system lack of tissues complexity, cell-to-cell and cell-to-ECM interactions, and physical stimuli to be used as model for studying unique events involved in cancer development and progression or even related treatments and [152]. For this reason, studies based on animal models continue to be the gold standard even though their notable drawbacks including interspecies differences and ethical concerns. To address the gap between 2D in vitro systems and vivo animal studies, advanced technologies such as 3D organoids and organ-on-chip (OoC) platforms have been developed, offering more physiologically relevant alternatives. They showed great potential and broad applicability in drug development, safety assessment as well as in personalized medicine and advanced preclinical/clinical research. Organoids, self-organizing 3D structures derived from pluripotent stem cells or adult progenitor cells, recapitulate many key functions of the original tissue, including cellular heterogeneity and spatial organization [153,154]. Organoids derived from cancerous tissue are particularly valuable for modeling tumor biology and drug response [155,156,157,158]. However, cellular self-organization alone is insufficient to fully mimic native tissue architecture, which is why biomaterials such as hydrogels or decellularized scaffolds are employed to simulate ECM characteristics and support cell–cell communication [159,160,161]. Thanks to the development of the lithography technique, and subsequently the 3D printing once, has been possible the production of the organ-on-chip system. It consists of a silicon-based organic polymer, polydimethylsiloxane (PDMS), microfluidic device, designed for maintaining cell culture in a closed environment and for analyzing cell biological characteristics. These technologies introduce the possibility to regulate fluidic parameters like flow, pressure, oxygen, pH in real time during the entire experimental procedure [162]. Colon organoids and tumoroids were successfully developed, expanded and a long-term cultured in many studies [163,164]. Given the importance of gut microbiota, some models involve microinjecting bacteria into the organoid lumen to replicate host–microbe interactions more accurately [165]. This approach has also been used to study infections in other tissues, such as Cryptosporidium in the small intestine [166] and endometrial infections [167]. Combining organoids with microfluidic chips, organoid-on-chip systems, add greater experimental control, allowing real-time regulation of flow, oxygen, and mechanical forces. These platforms better simulate the intestinal environment, including peristalsis and crypt architecture [168].
In this context, the studies on postbiotics carried out using these advanced models are characterized by higher human relevance. Table 2 reports relevant results from SR and ELS research, focusing on strings that include primary and secondary research keywords.
Although the majority of preclinical studies still rely on 2D cultures and mouse models, some combine organoids, animals and microfluidics to investigate inflammation, epithelial regeneration, and tumor growth. Only a few studies utilize microfluidic chip systems. The main application was the production of hydrogel-based microsphere for postbiotic delivery. Several studies combined early use of cell lines and following integration of in vivo and organoid models. As an example, Sugimura et al. found that Lactobacillus gallinarum, through production of bioactive metabolites, like ILA, significantly reduced tumor number and size compared to controls in mouse models. Its culture supernatant suppressed CRC cell proliferation and induced apoptosis in CRC cells and patient-derived organoids (but not normal cells). It also altered gut microbiota towards a more beneficial composition. The study was limited to two patients for organoid derivation [169]. However, the lack of standardized, interconnected systems limits data integration and translation across platforms. Harmonized models could reduce experimental time and cost, while offering closer alignment with human biology. These studies investigated the inflammatory modulation properties of several postbiotics. Cho Y. et al focused on the COX-2 gene pathway, while Lee H. et al focused on (IFNγ)/TNFα, IL-1β, IL-6, IL-8, IL-10, and TGF-β pathway [170,171].
Fewer than others are the studies about the maintenance/recovery of intestinal epithelium integrity and functionality, following an inflammatory stress. Furone et al. analyzed Lactobacillus rhamnosus postbiotic and its protective role on the alteration induced by gliadin in Celiac disease patient-derived organoids. They found that the postbiotic acted on mTOR, inflammation and autophagy pathways, all essential for the maintenance of the epithelium integrity and functionality. Although the model was not developed for CRC, the mTOR pathway, in particular the PI3K/Akt/mTOR pathway, was extensively described as a potential target in this type of cancer [172].
Finally, an advanced preclinical model based on human iPSC-derived intestinal epithelial cells (IECs) was used to assess the anti-inflammatory properties of heat-killed Lactiplantibacillus plantarum WCFS1 against the conventional Caco-2 cell model, offering improved physiological relevance [173].
5. Clinal Evidences, Formulation and Delivery of Postbiotics
5.1. Clinical Evidences
While some large clinical trials were described for probiotics in post-surgical CRC, no clinical trials are reported for postbiotics application in CRC patients.
As an example, Zaharuddin et al. showed that probiotics containing Lactobacillus and Bifidobacteria strains (Lactobacillus acidophilus BCMC® 12,130, Lactobacillus lactis BCMC® 12,451, Lactobacillus casei subsp BCMC® 12,313, Bifidobacterium longum BCMC® 02120, Bifidobacterium bifidum BCMC® 02290,Bifidobacterium infantis BCMC® 02129) are safe to be consumed at four weeks after surgery in CRC patients and have reduced pro-inflammatory cytokines, probably by intestinal microenvironment modification [174]. In another randomized controlled prospective, a probiotic with eight bacterial cultures (Lactobacillus acidophilus, Lactobacillus casei, Lactobacillus plantarum, Lactobacillus rhamnosus, Bifidobacterium lactis, Bifidobacterium bifidum, Bifidobacterium breve, Streptococcus thermophilus) was used to treat CRC patients, and its use led to a statistically significant reduction in postoperative complication [175]. Those studies, as already mentioned, represent clear examples that underline the existence of this data gap, indeed few clinical studies regarding postbiotics application in CRC patients can be considered.
In another interesting clinical study different levels of SCFA and HSP 70 expression in CRC were compared with non-CRC patients. Fauzi et al. found that CRC patients had lower level of acetate, propionate and butyrate acids than non-CRC and that short chain fatty acids were indirectly correlated to CRC pathogenesis [176]. In placebo-controlled RIBOGUT trial, Liu et al. found that oral supplementation with 100 mg/day riboflavin for 2 weeks increased the number of F. prausnitzii in feces, promoting butyrate production in the absence of major shifts in gut microbiota composition, while the complexity and stability of the bacterial network were enhanced [177].
In another study, evaluation of the potential roles of taking probiotics postoperatively in attenuating the gastrointestinal complications and disturbed gut microbiota in CRC patients undergoing chemotherapy researchers was carried out [178]. Protection of B. infants, L. acidophilus, E. faecalis, and B. cereus against this chemotherapy-induced gut microbiota dysbiosis and also promoted the production of SCFA.
Although no clinical studies focusing on administration of postbiotics were found, these evidences show that the supplementations of these metabolites may have a beneficial effect on CRC. Surely, large clinical trials are needed to fill this research data gap.
5.2. Postbiotic Formulation and Delivery
Although a preventive, palliative and therapeutic role of several postbiotics was extensively discussed, their clinical potential may be limited by instability, rapid degradation and poor bioavailability. To overcome these challenges, innovative drug delivery systems led to obtaining postbiotics targeted delivery to their sites of action, resulting in improved therapeutic efficacy and reduced side effects. This kind of pharmaceutical approach is useful also to mask unpleasant organoleptic properties and increase patients’ compliance [179].
In particular, several novel pharmaceutical formulations, to enhance delivery of butyrate and the other SCFAs to tumor cells, were studied and described in literature [180]. Advanced drug delivery strategies, such as enteric coating, conjugation with dietary fibers, prodrug design, and nanoformulations, enhance the stability of metabolites during bodily transit, ensuring targeted delivery to specific sites. These systems also improve release profiles, helping to prolong the duration of therapeutic effectiveness. As an example, sodium butyrate, well known postbiotic with therapeutic potential, was found to similarly inhibit the growth of a variety of other cancer cell lines in vitro. For this purpose, a combination of poly (lactic-co-glycolic acid) (PLGA) and poly (N-isopropylacrylamide) (PNIPAM) was used to create sodium butyrate-loaded microspheres, which showed slow degradation, prolong retention, and enable controlled release, making them effective for treating different clinical conditions [181]. Enteric encapsulations, prodrugs and esterification to dietary fibers have been employed to ensure postbiotics reach the distal gut, where they exert their beneficial effects. As microbial metabolites are normally produced in the distal gut, targeting their delivery to this site enables them to mimic their production by a eubiotic microbiome. Enteric coatings protect drugs from gastric degradation and also can impart delayed-, modified-, and/or controlled-release characteristics by virtue of their selective disintegration at specific pH values or upon exposure to intestinal enzymes [182]. Usually, prodrugs are designed to delay metabolite release until they reach systemic circulation, improving stability, bioavailability, and duration of action of small molecule drugs [183]. Postbiotic-prodrug formulations are currently only related to SCFA prodrugs, which are, in turn, predominantly comprised of butyrate prodrugs. In this context, formulations like Tributyrin (TB), Pivaloyloxymethyl butyrate (AN-9, Pivanex), Butyroyloxymethyl diethylphosphate (AN-7) and N-(1-carbamoyl-2-phenyl-ethyl) butyramide (FBA) led to obtain improvements in pharmacokinetics characteristics [123,184,185,186]. Additionally, esterification to dietary fibers represents another pharmaceutical approach to achieve colon-targeted delivery, prolonging the release of short-chain fatty acids (SCFAs), eliminating their unpleasant organoleptic properties and supporting gut microbiota interactions [187]. As an example, SCFA-esterified HAMS (High amylose maize starch) selectively increase SCFA levels in the distal GI tract and alters gut microbiota, expanding SCFA-utilizing bacteria and shifting microbial metabolism toward SCFA utilization [188,189,190]. Offering a promising approach to extend drug exposure profiles, nanoformulations act as targeted-delivery carriers, concentrating drugs at specific sites, or as controlled-release systems, maintaining therapeutic drug levels over an extended period. Nano-scale carriers reduce the need for frequent dosing and improve stability against gastric degradation, offering another strategy for controlled postbiotics release in the lower intestine. Cholesteryl butyrate-loaded solid lipid nanoparticles (Chol-But SLNs) provide stability in acidic gastric conditions, enable sustained drug release, and support targeted delivery to inflamed tissues through preferential uptake by immune cells [191]. Polyvinyl butyrate nanoparticles (PV-But NPs) were designed for controlled butyrate delivery to the lower intestine, resisting pancreatic lipase hydrolysis and enabling slow butyrate release [192]. Liposome-encapsulated sodium butyrate (NaB-Lip), originally developed to address F. nucleatum-induced chemotherapy resistance, accumulate in F. nucleatum-infected colorectal tumors and, when combined with oxaliplatin, significantly reduced tumor growth and improved survival [193].
Certain postbiotics are believed to help alleviate colonic inflammation and support the restoration of gut barrier function. For instance, p40, a protein produced by Lactobacillus rhamnosus GG, has been shown to counteract cytokine-induced epithelial cell apoptosis and prevent disruption of the intestinal barrier. Targeted delivery of hydrogel-coating p40 (to protect p40 from degradation) is effective in preventing and treating intestinal injury and inflammation, as well as promoting protective immune response [194,195]. HM0539, a secreted protein identified by the means of liquid chromatography-tandem mass spectrometry analysis from the culture of L. rhamnosus GG, plays a protective role in maintaining the integrity of the intestinal barrier by increasing the expression of intestinal mucin and preventing intestinal barrier injury. A pectin/zein beads delivery system was used to deliver HM0539 to the colon as they protect HM0539 from protease attack. In this study its potential bioactivity was tested in vitro and in vivo [196].
6. Conclusions
The growing body of evidence reviewed in this work underscores the promising role of postbiotics as co-adjuvant agents in the therapy and management of CRC. Postbiotics, including short-chain fatty acids, exopolysaccharides, enzymes, cell wall fragments, and other microbial metabolites, demonstrate multiple health-promoting activities, such as anti-inflammatory, immunomodulatory, antioxidant, and antitumor effects. These mechanisms collectively contribute to intestinal barrier protection, immune homeostasis, and inhibition of CRC progression through apoptosis induction, modulation of oncogenic signaling pathways, and epigenetic regulation. Preclinical studies, especially those employing advanced models such as organoids and organ-on-chip systems, reveal the potential of postbiotics to emulate host–microbe interactions with higher physiological relevance than traditional 2D cell cultures. These models offer crucial insights into the multifaceted bioactivities of postbiotics, though their broader adoption is still limited by the lack of standardization and translational frameworks. Despite these encouraging findings, the clinical application of postbiotics in CRC remains at a preliminary stage. Critical challenges persist, including the need for standardized definitions, reproducible manufacturing processes, comprehensive safety profiling, and robust clinical validation. Additionally, regulatory pathways for the approval and integration of postbiotic-based interventions into therapeutic protocols require clearer delineation.
Future research should aim at translating preclinical efficacy into clinical settings through well-designed human trials, while also leveraging precision medicine approaches to tailor postbiotic therapies to individual microbiome profiles and genetic backgrounds. With continued interdisciplinary effort, postbiotics hold strong potential to emerge as effective, safe, and sustainable adjuncts in CRC prevention and treatment strategies.
Author Contributions
Conceptualization and methodology, T.D. and T.V.; investigation T.D., C.Z., V.L., A.D.C.; data curation, T.D.; writing—original draft preparation, T.D., C.Z., V.L., A.D.C., S.S., T.V.; writing—review and editing, T.D., A.V., T.V., S.S; visualization, A.V.; funding acquisition, T.D., T.V. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
Dr. D’Amore (Principal Investigator), Dr. Di Ciancia, Dr. Lauciello, Dr. Vagliasindi and Dr. Zolfanelli (Collaborators) acknowledge the support by Italian Ministry of Health - Ricerca Corrente 2025, for the project “Relevance assessment of human-based next generation in vitro models for Colorectal Cancer: new perspectives in Cancer Research” (ID 2791643).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hou, K.; Wu, Z.-X.; Chen, X.-Y.; Wang, J.-Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in Health and Diseases. Signal Transduct Target Ther 2022, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.-Y.; Mei, J.-X.; Yu, G.; Lei, L.; Zhang, W.-H.; Liu, K.; Chen, X.-L.; Kołat, D.; Yang, K.; Hu, J.-K. Role of the Gut Microbiota in Anticancer Therapy: From Molecular Mechanisms to Clinical Applications. Sig Transduct Target Ther 2023, 8, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Fong, W.; Li, Q.; Yu, J. Gut Microbiota Modulation: A Novel Strategy for Prevention and Treatment of Colorectal Cancer. Oncogene 2020, 39, 4925–4943. [Google Scholar] [CrossRef] [PubMed]
- Vivarelli, S.; Salemi, R.; Candido, S.; Falzone, L.; Santagati, M.; Stefani, S.; Torino, F.; Banna, G.L.; Tonini, G.; Libra, M. Gut Microbiota and Cancer: From Pathogenesis to Therapy. Cancers (Basel) 2019, 11, 38. [Google Scholar] [CrossRef]
- Murgiano, M.; Bartocci, B.; Puca, P.; di Vincenzo, F.; Del Gaudio, A.; Papa, A.; Cammarota, G.; Gasbarrini, A.; Scaldaferri, F.; Lopetuso, L.R. Gut Microbiota Modulation in IBD: From the Old Paradigm to Revolutionary Tools. Int J Mol Sci 2025, 26, 3059. [Google Scholar] [CrossRef]
- Senthilkumar, H.; Arumugam, M. Gut Microbiota: A Hidden Player in Polycystic Ovary Syndrome. J Transl Med 2025, 23, 443. [Google Scholar] [CrossRef]
- Xie, Q.; Liu, J.; Yu, P.; Qiu, T.; Jiang, S.; Yu, R. Unlocking the Power of Probiotics, Postbiotics: Targeting Apoptosis for the Treatment and Prevention of Digestive Diseases. Front Nutr 2025, 12, 1570268. [Google Scholar] [CrossRef]
- Mishra, S.; Jain, S.; Agadzi, B.; Yadav, H. A Cascade of Microbiota-Leaky Gut-Inflammation- Is It a Key Player in Metabolic Disorders? Curr Obes Rep 2025, 14, 32. [Google Scholar] [CrossRef]
- Golden, A.; Williams, C.; Yadav, H.; Masternak, M.M.; Labyak, C.; Holland, P.J.; Arikawa, A.Y.; Jain, S. The Selection of Participants for Interventional Microbiota Trials Involving Cognitively Impaired Older Adults. Geroscience 2025. [Google Scholar] [CrossRef]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Lopetuso, L.R.; Scaldaferri, F.; Pulcini, G.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. Food Components and Dietary Habits: Keys for a Healthy Gut Microbiota Composition. Nutrients 2019, 11, 2393. [Google Scholar] [CrossRef]
- Del Chierico, F.; Vernocchi, P.; Dallapiccola, B.; Putignani, L. Mediterranean Diet and Health: Food Effects on Gut Microbiota and Disease Control. International Journal of Molecular Sciences 2014, 15, 11678–11699. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Ju, Z.; Zuo, T. Time for Food: The Impact of Diet on Gut Microbiota and Human Health. Nutrition 2018, 51–52, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Green, K.M.; Rawat, M. A Comprehensive Overview of Postbiotics with a Special Focus on Discovery Techniques and Clinical Applications. Foods 2024, 13, 2937. [Google Scholar] [CrossRef] [PubMed]
- Prajapati, N.; Patel, J.; Singh, S.; Yadav, V.K.; Joshi, C.; Patani, A.; Prajapati, D.; Sahoo, D.K.; Patel, A. Postbiotic Production: Harnessing the Power of Microbial Metabolites for Health Applications. Front. Microbiol. 2023, 14. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Postbiotics. Nat Rev Gastroenterol Hepatol 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Vinderola, G.; Sanders, M.E.; Salminen, S. The Concept of Postbiotics. Foods 2022, 11, 1077. [Google Scholar] [CrossRef]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics-A Step Beyond Pre- and Probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef]
- Liang, B.; Xing, D. The Current and Future Perspectives of Postbiotics. Probiotics & Antimicro. Prot. 2023, 15, 1626–1643. [Google Scholar] [CrossRef]
- Rad, A.H.; Aghebati-Maleki, L.; Kafil, H.S.; Abbasi, A. Molecular Mechanisms of Postbiotics in Colorectal Cancer Prevention and Treatment. Critical Reviews in Food Science and Nutrition 2021, 61, 1787–1803. [Google Scholar] [CrossRef]
- Feizi, H.; Plotnikov,Andrey; Rezaee,Mohammad Ahangarzadeh; Ganbarov,Khudaverdi; Kamounah,Fadhil S. ; Nikitin,Sergei; Kadkhoda,Hiva; Gholizadeh,Pourya; Pagliano,Pasquale; and Kafil, H.S. Postbiotics versus Probiotics in Early-Onset Colorectal Cancer. Critical Reviews in Food Science and Nutrition 2024, 64, 3573–3582. [Google Scholar] [CrossRef]
- Zhong, Y.; Liu,Zhijia; Wang,Yanfei; Cai,Shengbao; Qiao,Zhu; Hu,Xiaosong; Wang,Tao; and Yi, J. Preventive Methods for Colorectal Cancer Through Dietary Interventions: A Focus on Gut Microbiota Modulation. Food Reviews International 2025, 41, 720–748. [Google Scholar] [CrossRef]
- Xu, Y.; Wu, X.; Li, Y.; Liu, X.; Fang, L.; Jiang, Z. Probiotics and the Role of Dietary Substrates in Maintaining the Gut Health: Use of Live Microbes and Their Products for Anticancer Effects against Colorectal Cancer. J Microbiol Biotechnol 2024, 34, 1933–1946. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global Cancer Statistics 2022: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA: A Cancer Journal for Clinicians 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.W.; Harpaz, N.; Itzkowitz, S.H.; Parsons, R.E. Molecular Mechanisms in Colitis-Associated Colorectal Cancer. Oncogenesis 2023, 12, 1–11. [Google Scholar] [CrossRef]
- Cardoso, R.; Guo, F.; Heisser, T.; Hackl, M.; Ihle, P.; De Schutter, H.; Van Damme, N.; Valerianova, Z.; Atanasov, T.; Májek, O.; et al. Colorectal Cancer Incidence, Mortality, and Stage Distribution in European Countries in the Colorectal Cancer Screening Era: An International Population-Based Study. Lancet Oncol 2021, 22, 1002–1013. [Google Scholar] [CrossRef]
- van de Wetering, M.; Francies, H.E.; Francis, J.M.; Bounova, G.; Iorio, F.; Pronk, A.; van Houdt, W.; van Gorp, J.; Taylor-Weiner, A.; Kester, L.; et al. Prospective Derivation of a Living Organoid Biobank of Colorectal Cancer Patients. Cell 2015, 161, 933–945. [Google Scholar] [CrossRef]
- Asefa, Z.; Belay, A.; Welelaw, E.; Haile, M. Postbiotics and Their Biotherapeutic Potential for Chronic Disease and Their Feature Perspective: A Review. Front. Microbiomes 2025, 4, 1489339. [Google Scholar] [CrossRef]
- Hijová, E. Postbiotics as Metabolites and Their Biotherapeutic Potential. International Journal of Molecular Sciences 2024, 25, 5441. [Google Scholar] [CrossRef]
- Suthar, P.; Kumar, S.; Kumar, V.; Sharma, V.; Dhiman, A. Postbiotics: An Exposition on next Generation Functional Food Compounds- Opportunities and Challenges. Crit Rev Food Sci Nutr 2025, 65, 1163–1182. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An Evolving Term within the Functional Foods Field. Trends in Food Science & Technology 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Saedi, S.; Derakhshan, S.; Hasani, A.; Khoshbaten, M.; Poortahmasebi, V.; Milani, P.G.; Sadeghi, J. Recent Advances in Gut Microbiome Modulation: Effect of Probiotics, Prebiotics, Synbiotics, and Postbiotics in Inflammatory Bowel Disease Prevention and Treatment. Curr Microbiol 2024, 82, 12. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Martino, L.D.; Li, J. Natural Polysaccharides-Based Postbiotics and Their Potential Applications. Explor Med. 2024, 5, 444–458. [Google Scholar] [CrossRef]
- Jin, X.; Nguyen, T.T.M.; Yi, E.-J.; Zheng, Q.; Park, S.-J.; Yi, G.-S.; Yang, S.-J.; Kim, M.-J.; Yi, T.-H. Emerging Trends in Skin Anti-Photoaging by Lactic Acid Bacteria: A Focus on Postbiotics. Chemistry 2024, 6, 1495–1508. [Google Scholar] [CrossRef]
- Harat, S.G.; Pourjafar, H. Health Benefits and Safety of Postbiotics Derived from Different Probiotic Species. Curr Pharm Des 2025, 31, 116–127. [Google Scholar] [CrossRef]
- Rauf, A.; Khalil, A.A.; Rahman, U.-U.-; Khalid, A.; Naz, S.; Shariati, M.A.; Rebezov, M.; Urtecho, E.Z.; de Albuquerque, R.D.D.G.; Anwar, S.; et al. Recent Advances in the Therapeutic Application of Short-Chain Fatty Acids (SCFAs): An Updated Review. Crit Rev Food Sci Nutr 2022, 62, 6034–6054. [Google Scholar] [CrossRef]
- Ramos Meyers, G.; Samouda, H.; Bohn, T. Short Chain Fatty Acid Metabolism in Relation to Gut Microbiota and Genetic Variability. Nutrients 2022, 14, 5361. [Google Scholar] [CrossRef]
- van der Hee, B.; Wells, J.M. Microbial Regulation of Host Physiology by Short-Chain Fatty Acids. Trends Microbiol 2021, 29, 700–712. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, J.; Xie, F.; He, H.; Johnston, L.J.; Dai, X.; Wu, C.; Ma, X. Dietary Fiber-Derived Short-Chain Fatty Acids: A Potential Therapeutic Target to Alleviate Obesity-Related Nonalcoholic Fatty Liver Disease. Obes Rev 2021, 22, e13316. [Google Scholar] [CrossRef]
- Bourebaba, Y.; Marycz, K.; Mularczyk, M.; Bourebaba, L. Postbiotics as Potential New Therapeutic Agents for Metabolic Disorders Management. Biomed Pharmacother 2022, 153, 113138. [Google Scholar] [CrossRef]
- Houtman, T.A.; Eckermann, H.A.; Smidt, H.; de Weerth, C. Gut Microbiota and BMI throughout Childhood: The Role of Firmicutes, Bacteroidetes, and Short-Chain Fatty Acid Producers. Sci Rep 2022, 12, 3140. [Google Scholar] [CrossRef]
- Bongiovanni, T.; Yin, M.O.L.; Heaney, L.M. The Athlete and Gut Microbiome: Short-Chain Fatty Acids as Potential Ergogenic Aids for Exercise and Training. Int J Sports Med 2021, 42, 1143–1158. [Google Scholar] [CrossRef] [PubMed]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Reynés, B.; Palou, M.; Rodríguez, A.M.; Palou, A. Regulation of Adaptive Thermogenesis and Browning by Prebiotics and Postbiotics. Front Physiol 2018, 9, 1908. [Google Scholar] [CrossRef] [PubMed]
- Ashaolu, T.J.; Ashaolu, J.O.; Adeyeye, S. a. O. Fermentation of Prebiotics by Human Colonic Microbiota in Vitro and Short-Chain Fatty Acids Production: A Critical Review. J Appl Microbiol 2021, 130, 677–687. [Google Scholar] [CrossRef]
- Fang, H.; Rodrigues E-Lacerda, R.; Barra, N.G.; Kukje Zada, D.; Robin, N.; Mehra, A.; Schertzer, J.D. Postbiotic Impact on Host Metabolism and Immunity Provides Therapeutic Potential in Metabolic Disease. Endocr Rev 2025, 46, 60–79. [Google Scholar] [CrossRef]
- Wu, J.; Li, Z.; Zhang, Z.; Zhang, J.; Hu, H.; Lan, H.; Hong, W.; Yang, Z. Characterization of a Postbiotic Exopolysaccharide Produced by Lacticaseibacillus Paracasei ET-22 with Antioxidant and Anti-Inflammatory Efficacy. Int J Biol Macromol 2025, 306, 141608. [Google Scholar] [CrossRef]
- Goh, J.X.H.; Tan, L.T.H.; Law, J.W.F.; Ser, H.L.; Khaw, K.Y.; Letchumanan, V.; Lee, L.H.; Goh, B.H. Harnessing the Potentialities of Probiotics, Prebiotics, Synbiotics, Paraprobiotics, and Postbiotics for Shrimp Farming. Reviews in Aquaculture 2022, 14, 1478–1557. [Google Scholar] [CrossRef]
- Kango, N.; Nath, S. Prebiotics, Probiotics and Postbiotics: The Changing Paradigm of Functional Foods. J Diet Suppl 2024, 21, 709–735. [Google Scholar] [CrossRef]
- Vijayaganapathi, A. ; K.a., A.; Shree Kumari, G.R.; Subathra Devi; Vaithilingam, M. Chapter 28 - Antiatherosclerotic Effects of Postbiotics. In Postbiotics; Dharumadurai, D., Halami, P.M., Eds.; Developments in Applied Microbiology and Biotechnology; Academic Press, 2025; pp. 513–528 ISBN 978-0-443-22188-0.
- Yang, Y.; Fan, G.; Lan, J.; Li, X.; Li, X.; Liu, R. Polysaccharide-Mediated Modulation of Gut Microbiota in the Treatment of Liver Diseases: Promising Approach with Significant Challenges. Int J Biol Macromol 2024, 280, 135566. [Google Scholar] [CrossRef]
- Gezginç, Y.; Karabekmez-erdem, T.; Tatar, H.D.; Ayman, S.; Ganiyusufoğlu, E.; Dayısoylu, K.S. Health Promoting Benefits of Postbiotics Produced by Lactic Acid Bacteria: Exopolysaccharide. Biotech Studies 2022, 31, 61–70. [Google Scholar] [CrossRef]
- Moradi, M.; Kousheh, S.A.; Almasi, H.; Alizadeh, A.; Guimarães, J.T.; Yılmaz, N.; Lotfi, A. Postbiotics Produced by Lactic Acid Bacteria: The next Frontier in Food Safety. Compr Rev Food Sci Food Saf 2020, 19, 3390–3415. [Google Scholar] [CrossRef] [PubMed]
- Razzaq, A.; Shamsi, S.; Ali, A.; Ali, Q.; Sajjad, M.; Malik, A.; Ashraf, M. Microbial Proteases Applications. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Chae, H.S.; Jeong, S.G.; Ham, J.S.; Im, S.K.; Ahn, C.N.; Lee, J.M. In Vitro Antioxidative Properties of Lactobacilli. Asian-Australasian Journal of Animal Sciences 2005, 19, 262–265. [Google Scholar] [CrossRef]
- de Moreno de LeBlanc, A.; LeBlanc, J.G.; Perdigón, G.; Miyoshi, A.; Langella, P.; Azevedo, V.; Sesma, F. Oral Administration of a Catalase-Producing Lactococcus Lactis Can Prevent a Chemically Induced Colon Cancer in Mice. J Med Microbiol 2008, 57, 100–105. [Google Scholar] [CrossRef]
- Wang, W.; Zhu, J.; Cao, Q.; Zhang, C.; Dong, Z.; Feng, D.; Ye, H.; Zuo, J. Dietary Catalase Supplementation Alleviates Deoxynivalenol-Induced Oxidative Stress and Gut Microbiota Dysbiosis in Broiler Chickens. Toxins 2022, 14, 830. [Google Scholar] [CrossRef]
- Peng, M.; Tabashsum, Z.; Anderson, M.; Truong, A.; Houser, A.K.; Padilla, J.; Akmel, A.; Bhatti, J.; Rahaman, S.O.; Biswas, D. Effectiveness of Probiotics, Prebiotics, and Prebiotic-like Components in Common Functional Foods. Compr Rev Food Sci Food Saf 2020, 19, 1908–1933. [Google Scholar] [CrossRef]
- Chen, J.; Li, F.; Yang, W.; Jiang, S.; Li, Y. Supplementation with Exogenous Catalase from Penicillium Notatum in the Diet Ameliorates Lipopolysaccharide-Induced Intestinal Oxidative Damage through Affecting Intestinal Antioxidant Capacity and Microbiota in Weaned Pigs. Microbiol Spectr 9, e00654-21. [CrossRef]
- Wei, L.; Wang, B.; Bai, J.; Zhang, Y.; Liu, C.; Suo, H.; Wang, C. Postbiotics Are a Candidate for New Functional Foods. Food Chem X 2024, 23, 101650. [Google Scholar] [CrossRef]
- Jung, B.-J.; Kim, H.; Chung, D.-K. Differential Immunostimulatory Effects of Lipoteichoic Acids Isolated from Four Strains of Lactiplantibacillus Plantarum. Applied Sciences 2022, 12, 954. [Google Scholar] [CrossRef]
- Evangelista, A.G.; Corrêa, J.A.F.; Dos Santos, J.V.G.; Matté, E.H.C.; Milek, M.M.; Biauki, G.C.; Costa, L.B.; Luciano, F.B. Cell-Free Supernatants Produced by Lactic Acid Bacteria Reduce Salmonella Population in Vitro. Microbiology (Reading) 2021, 167. [Google Scholar] [CrossRef]
- Bermudez-Brito, M.; Muñoz-Quezada, S.; Gomez-Llorente, C.; Matencio, E.; Bernal, M.J.; Romero, F.; Gil, A. Cell-Free Culture Supernatant of Bifidobacterium Breve CNCM I-4035 Decreases Pro-Inflammatory Cytokines in Human Dendritic Cells Challenged with Salmonella Typhi through TLR Activation. PLOS ONE 2013, 8, e59370. [Google Scholar] [CrossRef]
- Wei, L.; Marco, M.L. The Fermented Cabbage Metabolome and Its Protection against Cytokine-Induced Intestinal Barrier Disruption of Caco-2 Monolayers. Applied and Environmental Microbiology 2025, 0, e02234–24. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; Coda, R.; De Angelis, M.; Gobbetti, M. Exploitation of Vegetables and Fruits through Lactic Acid Fermentation. Food Microbiology 2013, 33, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Plengvidhya, V.; Breidt, F.; Lu, Z.; Fleming, H.P. DNA Fingerprinting of Lactic Acid Bacteria in Sauerkraut Fermentations. Appl Environ Microbiol 2007, 73, 7697–7702. [Google Scholar] [CrossRef] [PubMed]
- Tamang, J.P.; Shin, D.-H.; Jung, S.-J.; Chae, S.-W. Functional Properties of Microorganisms in Fermented Foods. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Zabat, M.A.; Sano, W.H.; Wurster, J.I.; Cabral, D.J.; Belenky, P. Microbial Community Analysis of Sauerkraut Fermentation Reveals a Stable and Rapidly Established Community. Foods 2018, 7, 77. [Google Scholar] [CrossRef]
- Fermented Foods in Health and Disease Prevention; 2016; ISBN 978-0-12-802309-9.
- Marco, M.L.; Pavan, S.; Kleerebezem, M. Towards Understanding Molecular Modes of Probiotic Action. Current Opinion in Biotechnology 2006, 17, 204–210. [Google Scholar] [CrossRef]
- Iraporda, C.; Errea, A.; Romanin, D.E.; Cayet, D.; Pereyra, E.; Pignataro, O.; Sirard, J.C.; Garrote, G.L.; Abraham, A.G.; Rumbo, M. Lactate and Short Chain Fatty Acids Produced by Microbial Fermentation Downregulate Proinflammatory Responses in Intestinal Epithelial Cells and Myeloid Cells. Immunobiology 2015, 220, 1161–1169. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Kim, T.-Y.; Kim, Y.; Lee, S.-H.; Kim, S.; Kang, S.W.; Yang, J.-Y.; Baek, I.-J.; Sung, Y.H.; Park, Y.-Y.; et al. Microbiota-Derived Lactate Accelerates Intestinal Stem-Cell-Mediated Epithelial Development. Cell Host Microbe 2018, 24, 833–846.e6. [Google Scholar] [CrossRef]
- Pedersen, M.G.B.; Søndergaard, E.; Nielsen, C.B.; Johannsen, M.; Gormsen, L.C.; Møller, N.; Jessen, N.; Rittig, N. Oral Lactate Slows Gastric Emptying and Suppresses Appetite in Young Males. Clinical Nutrition 2022, 41, 517–525. [Google Scholar] [CrossRef]
- Yan, Y.; Xu,Ruijie; Li,Xiu; Yao,Zhijie; Zhang,Hao; Li,Haitao; and Chen, W. Unexpected Immunoregulation Effects of D-Lactate, Different from L-Lactate. Food and Agricultural Immunology 2022, 33, 286–301. [Google Scholar] [CrossRef]
- Valdes, D.S.; So, D.; Gill, P.A.; Kellow, N.J. Effect of Dietary Acetic Acid Supplementation on Plasma Glucose, Lipid Profiles, and Body Mass Index in Human Adults: A Systematic Review and Meta-Analysis. Journal of the Academy of Nutrition and Dietetics 2021, 121, 895–914. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.O.; Moraes, W.M.A.M. de; Silva, G.A.R. da; Prestes, J.; Schoenfeld, B.J. Vinegar (Acetic Acid) Intake on Glucose Metabolism: A Narrative Review. Clinical Nutrition ESPEN 2019, 32, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hunaefi, D.; Akumo, D.N.; Smetanska, I. Effect of Fermentation on Antioxidant Properties of Red Cabbages. Food Biotechnology 2013. [Google Scholar] [CrossRef]
- Kaulmann, A.; Jonville, M.-C.; Schneider, Y.-J.; Hoffmann, L.; Bohn, T. Carotenoids, Polyphenols and Micronutrient Profiles of Brassica Oleraceae and Plum Varieties and Their Contribution to Measures of Total Antioxidant Capacity. Food Chemistry 2014, 155, 240–250. [Google Scholar] [CrossRef]
- Martinez-Villaluenga, C.; Peñas, E.; Sidro, B.; Ullate, M.; Frias, J.; Vidal-Valverde, C. White Cabbage Fermentation Improves Ascorbigen Content, Antioxidant and Nitric Oxide Production Inhibitory Activity in LPS-Induced Macrophages. LWT - Food Science and Technology 2012, 46, 77–83. [Google Scholar] [CrossRef]
- Krajka-Kuźniak, V.; Szaefer, H.; Bartoszek, A.; Baer-Dubowska, W. Modulation of Rat Hepatic and Kidney Phase II Enzymes by Cabbage Juices: Comparison with the Effects of Indole-3-Carbinol and Phenethyl Isothiocyanate. British Journal of Nutrition 2011, 105, 816–826. [Google Scholar] [CrossRef]
- Siddeeg, A.; Afzaal,Muhammad; Saeed,Farhan; Ali,Rehman; Shah,Yasir Abbas; Shehzadi,Umber; Ateeq,Huda; Waris,Numra; Hussain,Muzzamal; Raza,Muhammad Ahtisham; et al. Recent Updates and Perspectives of Fermented Healthy Super Food Sauerkraut: A Review. International Journal of Food Properties 2022, 25, 2320–2331. [Google Scholar] [CrossRef]
- Peters, A.; Krumbholz, P.; Jäger, E.; Heintz-Buschart, A.; Çakir, M.V.; Rothemund, S.; Gaudl, A.; Ceglarek, U.; Schöneberg, T.; Stäubert, C. Metabolites of Lactic Acid Bacteria Present in Fermented Foods Are Highly Potent Agonists of Human Hydroxycarboxylic Acid Receptor 3. PLOS Genetics 2019, 15, e1008145. [Google Scholar] [CrossRef]
- Kasperek, M.C.; Velasquez Galeas, A.; Caetano-Silva, M.E.; Xie, Z.; Ulanov, A.; La Frano, M.; Devkota, S.; Miller, M.J.; Allen, J.M. Microbial Aromatic Amino Acid Metabolism Is Modifiable in Fermented Food Matrices to Promote Bioactivity. Food Chemistry 2024, 454, 139798. [Google Scholar] [CrossRef]
- Shelton, C.D.; Sing, E.; Mo, J.; Shealy, N.G.; Yoo, W.; Thomas, J.; Fitz, G.N.; Castro, P.R.; Hickman, T.T.; Torres, T.P.; et al. An Early-Life Microbiota Metabolite Protects against Obesity by Regulating Intestinal Lipid Metabolism. Cell Host & Microbe 2023, 31, 1604–1619.e10. [Google Scholar] [CrossRef]
- Henrick, B.M.; Rodriguez, L.; Lakshmikanth, T.; Pou, C.; Henckel, E.; Arzoomand, A.; Olin, A.; Wang, J.; Mikes, J.; Tan, Z.; et al. Bifidobacteria-Mediated Immune System Imprinting Early in Life. Cell 2021, 184, 3884–3898.e11. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Choi, K.-B.; Park, J.H.; Kim, K.H. Metabolite Profile Changes and Increased Antioxidative and Antiinflammatory Activities of Mixed Vegetables after Fermentation by Lactobacillus Plantarum. PLOS ONE 2019, 14, e0217180. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, C.; Ferrocino, I.; Reale, A.; Sabbatini, R.; Milanović, V.; Alkić-Subašić, M.; Boscaino, F.; Aquilanti, L.; Pasquini, M.; Trombetta, M.F.; et al. Study of Kefir Drinks Produced by Backslopping Method Using Kefir Grains from Bosnia and Herzegovina: Microbial Dynamics and Volatilome Profile. Food Res Int 2020, 137, 109369. [Google Scholar] [CrossRef]
- Azizi, N.F.; Kumar, M.R.; Yeap, S.K.; Abdullah, J.O.; Khalid, M.; Omar, A.R.; Osman, M.A.; Mortadza, S.A.S.; Alitheen, N.B. Kefir and Its Biological Activities. Foods 2021, 10, 1210. [Google Scholar] [CrossRef]
- Prado, M.R.; Blandón, L.M.; Vandenberghe, L.P.S.; Rodrigues, C.; Castro, G.R.; Thomaz-Soccol, V.; Soccol, C.R. Milk Kefir: Composition, Microbial Cultures, Biological Activities, and Related Products. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Barros, S.É. de L.; Rocha, C. dos S.; Moura, M.S.B. de; Barcelos, M.P.; Silva, C.H.T. de P. da; Hage-Melim, L.I. da S. Potential Beneficial Effects of Kefir and Its Postbiotic, Kefiran, on Child Food Allergy. Food Funct. 2021, 12, 3770–3786. [Google Scholar] [CrossRef]
- Rodrigues, K.L.; Caputo, L.R.G.; Carvalho, J.C.T.; Evangelista, J.; Schneedorf, J.M. Antimicrobial and Healing Activity of Kefir and Kefiran Extract. International Journal of Antimicrobial Agents 2005, 25, 404–408. [Google Scholar] [CrossRef]
- Seo, K.-H.; Gyu Lee, H.; Young Eor, J.; Jin Jeon, H.; Yokoyama, W.; Kim, H. Effects of Kefir Lactic Acid Bacteria-Derived Postbiotic Components on High Fat Diet-Induced Gut Microbiota and Obesity. Food Research International 2022, 157, 111445. [Google Scholar] [CrossRef]
- Rosa, D.D.; Dias, M.M.S.; Grześkowiak, Ł.M.; Reis, S.A.; Conceição, L.L.; Peluzio, M. do C.G. Milk Kefir: Nutritional, Microbiological and Health Benefits. Nutr Res Rev 2017, 30, 82–96. [Google Scholar] [CrossRef]
- Tingirikari, J.M.R.; Sharma, A.; Lee, H.-J. Kefir: A Fermented Plethora of Symbiotic Microbiome and Health. Journal of Ethnic Foods 2024, 11, 35. [Google Scholar] [CrossRef]
- Nugroho, D.; Thinthasit, A.; Surya, E.; Hartati; Oh, J. -S.; Jang, J.-G.; Benchawattananon, R.; Surya, R. Immunoenhancing and Antioxidant Potentials of Kimchi, an Ethnic Food from Korea, as a Probiotic and Postbiotic Food. Journal of Ethnic Foods 2024, 11, 12. [Google Scholar] [CrossRef]
- Sionek, B.; Szydłowska, A.; Küçükgöz, K.; Kołożyn-Krajewska, D. Traditional and New Microorganisms in Lactic Acid Fermentation of Food. Fermentation 2023, 9, 1019. [Google Scholar] [CrossRef]
- Ou, J.; Carbonero, F.; Zoetendal, E.G.; DeLany, J.P.; Wang, M.; Newton, K.; Gaskins, H.R.; O’Keefe, S.J.D. Diet, Microbiota, and Microbial Metabolites in Colon Cancer Risk in Rural Africans and African Americans. Am J Clin Nutr 2013, 98, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Pitmon, E.; Wang, K. Microbiome, Inflammation and Colorectal Cancer. Semin Immunol 2017, 32, 43–53. [Google Scholar] [CrossRef]
- Tan, J.K.; Macia, L.; Mackay, C.R. Dietary Fiber and SCFAs in the Regulation of Mucosal Immunity. Journal of Allergy and Clinical Immunology 2023, 151, 361–370. [Google Scholar] [CrossRef]
- Thangaraju, M.; Cresci, G.A.; Liu, K.; Ananth, S.; Gnanaprakasam, J.P.; Browning, D.D.; Mellinger, J.D.; Smith, S.B.; Digby, G.J.; Lambert, N.A.; et al. GPR109A Is a G-Protein-Coupled Receptor for the Bacterial Fermentation Product Butyrate and Functions as a Tumor Suppressor in Colon. Cancer Res 2009, 69, 2826–2832. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Round, J.L.; Kasper, D.L. A Microbial Symbiosis Factor Prevents Intestinal Inflammatory Disease. Nature 2008, 453, 620–625. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic Treg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef]
- Zheng, L.; Kelly, C.J.; Battista, K.D.; Schaefer, R.; Lanis, J.M.; Alexeev, E.E.; Wang, R.X.; Onyiah, J.C.; Kominsky, D.J.; Colgan, S.P. Microbial-Derived Butyrate Promotes Epithelial Barrier Function through IL-10 Receptor–Dependent Repression of Claudin-2. The Journal of Immunology 2017, 199, 2976–2984. [Google Scholar] [CrossRef]
- Peng, L.; Li, Z.-R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers. J Nutr 2009, 139, 1619–1625. [Google Scholar] [CrossRef]
- Lanis, J.M.; Alexeev, E.E.; Curtis, V.F.; Kitzenberg, D.A.; Kao, D.J.; Battista, K.D.; Gerich, M.E.; Glover, L.E.; Kominsky, D.J.; Colgan, S.P. Tryptophan Metabolite Activation of the Aryl Hydrocarbon Receptor Regulates IL-10 Receptor Expression on Intestinal Epithelia. Mucosal Immunology 2017, 10, 1133–1144. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Du, P.; Xie, Q.; Wang, N.; Li, H.; Smith, E.E.; Li, C.; Liu, F.; Huo, G.; Li, B. Protective Effects of Tryptophan-Catabolizing Lactobacillus Plantarum KLDS 1.0386 against Dextran Sodium Sulfate-Induced Colitis in Mice. Food Funct. 2020, 11, 10736–10747. [Google Scholar] [CrossRef] [PubMed]
- Cervantes-Barragan, L.; Chai, J.N.; Tianero, M.D.; Di Luccia, B.; Ahern, P.P.; Merriman, J.; Cortez, V.S.; Caparon, M.G.; Donia, M.S.; Gilfillan, S.; et al. Lactobacillus Reuteri Induces Gut Intraepithelial CD4+CD8αα+ T Cells. Science 2017, 357, 806–810. [Google Scholar] [CrossRef]
- Rabah, H.; Ménard, O.; Gaucher, F.; do Carmo, F.L.R.; Dupont, D.; Jan, G. Cheese Matrix Protects the Immunomodulatory Surface Protein SlpB of Propionibacterium Freudenreichii during in Vitro Digestion. Food Research International 2018, 106, 712–721. [Google Scholar] [CrossRef]
- Taverniti, V.; Stuknyte, M.; Minuzzo, M.; Arioli, S.; De Noni, I.; Scabiosi, C.; Cordova, Z.M.; Junttila, I.; Hämäläinen, S.; Turpeinen, H.; et al. S-Layer Protein Mediates the Stimulatory Effect of Lactobacillus Helveticus MIMLh5 on Innate Immunity. Applied and Environmental Microbiology 2013, 79, 1221–1231. [Google Scholar] [CrossRef]
- Liu, Z.; Shen, T.; Zhang, P.; Ma, Y.; Qin, H. Lactobacillus Plantarum Surface Layer Adhesive Protein Protects Intestinal Epithelial Cells against Tight Junction Injury Induced by Enteropathogenic Escherichia Coli. Mol Biol Rep 2011, 38, 3471–3480. [Google Scholar] [CrossRef]
- Kaji, R.; Kiyoshima-Shibata, J.; Nagaoka, M.; Nanno, M.; Shida, K. Bacterial Teichoic Acids Reverse Predominant IL-12 Production Induced by Certain Lactobacillus Strains into Predominant IL-10 Production via TLR2-Dependent ERK Activation in Macrophages. The Journal of Immunology 2010, 184, 3505–3513. [Google Scholar] [CrossRef]
- Tomkovich, S.; Jobin, C. Microbiota and Host Immune Responses: A Love–Hate Relationship. Immunology 2016, 147, 1–10. [Google Scholar] [CrossRef]
- Zadeh, M.; Khan, M.W.; Goh, Y.J.; Selle, K.; Owen, J.L.; Klaenhammer, T.; Mohamadzadeh, M. Induction of Intestinal Pro-Inflammatory Immune Responses by Lipoteichoic Acid. Journal of Inflammation 2012, 9, 7. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, Q.; Li, T.; Lu, L.; Wang, F.; Zhang, H.; Liu, Z.; Ma, H.; Zhu, Q.; Wang, J.; et al. Lactobacillus Plantarum-Derived Indole-3-Lactic Acid Ameliorates Colorectal Tumorigenesis via Epigenetic Regulation of CD8+ T Cell Immunity. Cell Metab 2023, 35, 943–960.e9. [Google Scholar] [CrossRef]
- Yan, F.; Cao, H.; Cover, T.L.; Whitehead, R.; Washington, M.K.; Polk, D.B. Soluble Proteins Produced by Probiotic Bacteria Regulate Intestinal Epithelial Cell Survival and Growth. Gastroenterology 2007, 132, 562–575. [Google Scholar] [CrossRef] [PubMed]
- Bäuerl, C.; Coll-Marqués, J.M.; Tarazona-González, C.; Pérez-Martínez, G. Lactobacillus Casei Extracellular Vesicles Stimulate EGFR Pathway Likely Due to the Presence of Proteins P40 and P75 Bound to Their Surface. Sci Rep 2020, 10, 19237. [Google Scholar] [CrossRef]
- Lu, R.; Shang, M.; Zhang, Y.-G.; Jiao, Y.; Xia, Y.; Garrett, S.; Bakke, D.; Bäuerl, C.; Martinez, G.P.; Kim, C.-H.; et al. Lactic Acid Bacteria Isolated From Korean Kimchi Activate the Vitamin D Receptor–Autophagy Signaling Pathways. Inflammatory Bowel Diseases 2020, 26, 1199–1211. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, A.; Rad, A.H.; Maleki, L.A.; Kafil, H.S.; Baghbanzadeh, A. Antigenotoxicity and Cytotoxic Potentials of Cell-Free Supernatants Derived from Saccharomyces Cerevisiae Var. Boulardii on HT-29 Human Colon Cancer Cell Lines. Probiotics & Antimicro. Prot. 2023, 15, 1583–1595. [Google Scholar] [CrossRef]
- Karimi Ardestani, S.; Tafvizi, F.; Tajabadi Ebrahimi, M. Heat-Killed Probiotic Bacteria Induce Apoptosis of HT-29 Human Colon Adenocarcinoma Cell Line via the Regulation of Bax/Bcl2 and Caspases Pathway. Hum Exp Toxicol 2019, 38, 1069–1081. [Google Scholar] [CrossRef]
- Konishi, H.; Fujiya, M.; Tanaka, H.; Ueno, N.; Moriichi, K.; Sasajima, J.; Ikuta, K.; Akutsu, H.; Tanabe, H.; Kohgo, Y. Probiotic-Derived Ferrichrome Inhibits Colon Cancer Progression via JNK-Mediated Apoptosis. Nat Commun 2016, 7, 12365. [Google Scholar] [CrossRef]
- Donohoe, D.R.; Holley, D.; Collins, L.B.; Montgomery, S.A.; Whitmore, A.C.; Hillhouse, A.; Curry, K.P.; Renner, S.W.; Greenwalt, A.; Ryan, E.P.; et al. A Gnotobiotic Mouse Model Demonstrates That Dietary Fiber Protects against Colorectal Tumorigenesis in a Microbiota- and Butyrate-Dependent Manner. Cancer Discovery 2014, 4, 1387–1397. [Google Scholar] [CrossRef]
- Cao, M.; Zhang, Z.; Han, S.; Lu, X. Butyrate Inhibits the Proliferation and Induces the Apoptosis of Colorectal Cancer HCT116 Cells via the Deactivation of MTOR/S6K1 Signaling Mediated Partly by SIRT1 Downregulation. Molecular Medicine Reports 2019, 19, 3941–3947. [Google Scholar] [CrossRef]
- Ryu, T.Y.; Kim, K.; Son, M.-Y.; Min, J.-K.; Kim, J.; Han, T.-S.; Kim, D.-S.; Cho, H.-S. Downregulation of PRMT1, a Histone Arginine Methyltransferase, by Sodium Propionate Induces Cell Apoptosis in Colon Cancer. Oncology Reports 2019, 41, 1691–1699. [Google Scholar] [CrossRef]
- Tarasenko, N.; Nudelman, A.; Tarasenko, I.; Entin-Meer, M.; Hass-Kogan, D.; Inbal, A.; Rephaeli, A. Histone Deacetylase Inhibitors: The Anticancer, Antimetastatic and Antiangiogenic Activities of AN-7 Are Superior to Those of the Clinically Tested AN-9 (Pivanex). Clin Exp Metastasis 2008, 25, 703–716. [Google Scholar] [CrossRef]
- Ali, S.R.; Humphreys, K.J.; Simpson, K.J.; McKinnon, R.A.; Meech, R.; Michael, M.Z. Functional High-Throughput Screen Identifies MicroRNAs That Promote Butyrate-Induced Death in Colorectal Cancer Cells. Molecular Therapy Nucleic Acids 2022, 30, 30–47. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Fullaondo, A.; Odriozola, I.; Odriozola, A. Chapter Eight - Microbiota and Beneficial Metabolites in Colorectal Cancer. In Advances in Genetics; Martínez, A.O., Ed.; Advances in Host Genetics and Microbiome in Colorectal Cancer-Related Phenotypes; Academic Press, 2024; Vol. 112, pp. 367–409.
- Da, M.; Sun, J.; Ma, C.; Li, D.; Dong, L.; Wang, L.-S.; Chen, F. Postbiotics: Enhancing Human Health with a Novel Concept. eFood 2024, 5, e180. [Google Scholar] [CrossRef]
- Osman, A.; El-Gazzar, N.; Almanaa, T.N.; El-Hadary, A.; Sitohy, M. Lipolytic Postbiotic from Lactobacillus Paracasei Manages Metabolic Syndrome in Albino Wistar Rats. Molecules 2021, 26, 472. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. Systematic Reviews 2021, 10, 89. [Google Scholar] [CrossRef]
- Vedrine, M.L.; Hanlon, J.; Bevan, R.; Floyd, P.; Brown, T.; Matthies, F. Extensive Literature Search, Selection for Relevance and Data Extraction of Studies Related to the Toxicity of PCDD/Fs and DL-PCBs in Humans. EFSA Supporting Publications 2018, 15, 1136E. [Google Scholar] [CrossRef]
- D’Amore, T.; Chaari, M.; Falco, G.; De Gregorio, G.; Zaraî Jaouadi, N.; Ali, D.S.; Sarkar, T.; Smaoui, S. When Sustainability Meets Health and Innovation: The Case of Citrus by-Products for Cancer Chemoprevention and Applications in Functional Foods. Biocatalysis and Agricultural Biotechnology 2024, 58, 103163. [Google Scholar] [CrossRef]
- Escamilla, J.; Lane, M.A.; Maitin, V. Cell-Free Supernatants from Probiotic Lactobacillus Casei and Lactobacillus Rhamnosus GG Decrease Colon Cancer Cell Invasion In Vitro. Nutrition and Cancer 2012. [Google Scholar] [CrossRef]
- Elham, N.; Naheed, M.; Elahe, M.; Hossein, M.M.; Majid, T. Selective Cytotoxic Effect of Probiotic, Paraprobiotic and Postbiotics of L.Casei Strains against Colorectal Cancer Cells: Invitro Studies. Braz. J. Pharm. Sci. 2022, 58, e19400. [Google Scholar] [CrossRef]
- Chen, Z.-Y.; Hsieh, Y.-M.; Huang, C.-C.; Tsai, C.-C. Inhibitory Effects of Probiotic Lactobacillus on the Growth of Human Colonic Carcinoma Cell Line HT-29. Molecules 2017, 22, 107. [Google Scholar] [CrossRef]
- Jastrząb, R.; Tomecki, R.; Jurkiewicz, A.; Graczyk, D.; Szczepankowska, A.K.; Mytych, J.; Wolman, D.; Siedlecki, P. The Strain-Dependent Cytostatic Activity of Lactococcus Lactis on CRC Cell Lines Is Mediated through the Release of Arginine Deiminase. Microbial Cell Factories 2024, 23, 82. [Google Scholar] [CrossRef]
- Luo, S.; Li, Z.; Mao, L.; Chen, S.; Sun, S. Sodium Butyrate Induces Autophagy in Colorectal Cancer Cells through LKB1/AMPK Signaling. J Physiol Biochem 2019, 75, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Chuah, L.-O.; Foo, H.L.; Loh, T.C.; Mohammed Alitheen, N.B.; Yeap, S.K.; Abdul Mutalib, N.E.; Abdul Rahim, R.; Yusoff, K. Postbiotic Metabolites Produced by Lactobacillus Plantarum Strains Exert Selective Cytotoxicity Effects on Cancer Cells. BMC Complementary and Alternative Medicine 2019, 19, 114. [Google Scholar] [CrossRef] [PubMed]
- Macias-Diaz, A.; Lopez, J.J.; Bravo, M.; Jardín, I.; Garcia-Jimenez, W.L.; Blanco-Blanco, F.J.; Cerrato, R.; Rosado, J.A. Postbiotics of Lacticaseibacillus Paracasei CECT 9610 and Lactiplantibacillus Plantarum CECT 9608 Attenuates Store-Operated Calcium Entry and FAK Phosphorylation in Colorectal Cancer Cells. Molecular Oncology 2024, 18, 1123–1142. [Google Scholar] [CrossRef] [PubMed]
- Deepak, V.; Ram Kumar Pandian,Sureshbabu; Sivasubramaniam,Shiva D. ; Nellaiah,Hariharan; and Sundar, K. Optimization of Anticancer Exopolysaccharide Production from Probiotic Lactobacillus Acidophilus by Response Surface Methodology. Preparative Biochemistry & Biotechnology 2016, 46, 288–297. [Google Scholar] [CrossRef]
- Cousin, F.J.; Jouan-Lanhouet, S.; Théret, N.; Brenner, C.; Jouan, E.; Moigne-Muller, G.L.; Dimanche-Boitrel, M.-T.; Jan, G. The Probiotic Propionibacterium Freudenreichii as a New Adjuvant for TRAIL-Based Therapy in Colorectal Cancer. Oncotarget 2016, 7, 7161–7178. [Google Scholar] [CrossRef]
- Jung, T.-H.; Park, J.H.; Jeon, W.-M.; Han, K.-S. Butyrate Modulates Bacterial Adherence on LS174T Human Colorectal Cells by Stimulating Mucin Secretion and MAPK Signaling Pathway. Nutrition Research and Practice 2015, 9, 343–349. [Google Scholar] [CrossRef]
- Xu, J.; Wu, X.; Yang, L.; Xu, X. The Intervention of B. Longum Metabolites in Fnevs’ Carcinogenic Capacity: A Potential Double-Edged Sword. Experimental Cell Research 2025, 445, 114407. [Google Scholar] [CrossRef]
- Erfanian, N.; Nasseri, S.; Miraki Feriz, A.; Safarpour, H.; Namaei, M.H. Characterization of Wnt Signaling Pathway under Treatment of Lactobacillus Acidophilus Postbiotic in Colorectal Cancer Using an Integrated in Silico and in Vitro Analysis. Sci Rep 2023, 13, 22988. [Google Scholar] [CrossRef]
- Erfanian, N.; Safarpour, H.; Tavakoli, T.; Mahdiabadi, M.A.; Nasseri, S.; Namaei, M.H. Investigating the Therapeutic Potential of Bifidobacterium Breve and Lactobacillus Rhamnosus Postbiotics through Apoptosis Induction in Colorectal HT-29 Cancer Cells. Iran J Microbiol 2024, 16, 68–78. [Google Scholar] [CrossRef]
- Neffe-Skocińska, K.; Długosz, E.; Szulc-Dąbrowska, L.; Zielińska, D. Novel Gluconobacter Oxydans Strains Selected from Kombucha with Potential Postbiotic Activity. Appl Microbiol Biotechnol 2024, 108, 27. [Google Scholar] [CrossRef]
- Shen, X.; Liu, L.; Peek, R.M.; Acra, S.A.; Moore, D.J.; Wilson, K.T.; He, F.; Polk, D.B.; Yan, F. Supplementation of P40, a Lactobacillus Rhamnosus GG-Derived Protein, in Early Life Promotes Epidermal Growth Factor Receptor-Dependent Intestinal Development and Long-Term Health Outcomes. Mucosal Immunology 2018, 11, 1316–1328. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Shukla, G. Administration of Metabiotics Extracted From Probiotic Lactobacillus Rhamnosus MD 14 Inhibit Experimental Colorectal Carcinogenesis by Targeting Wnt/β-Catenin Pathway. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Song, Y.; Sun, M.; Wang, A.; Jiang, S.; Mu, G.; Tuo, Y. Exopolysaccharide Produced by Lactiplantibacillus Plantarum-12 Alleviates Intestinal Inflammation and Colon Cancer Symptoms by Modulating the Gut Microbiome and Metabolites of C57BL/6 Mice Treated by Azoxymethane/Dextran Sulfate Sodium Salt. Foods 2021, 10, 3060. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Q.; Yu, B.; Zhou, J.; Zhang, J.; Zhang, R.; Xie, J.; Wang, Q.; Zhao, S. Lysates of Lactobacillus Acidophilus Combined with CTLA-4-Blocking Antibodies Enhance Antitumor Immunity in a Mouse Colon Cancer Model. Sci Rep 2019, 9, 20128. [Google Scholar] [CrossRef]
- Lee, P.-J.; Hung,Chien-Min; Yang,Ai-Jen; Hou,Cheng-Yu; Chou,Hung-Wen; Chang,Yi-Chung; Chu,Wen-Cheng; Huang,Wen-Yen; Kuo,Wen-Chih; Yang,Chia-Chun; et al. MS-20 Enhances the Gut Microbiota-Associated Antitumor Effects of Anti-PD1 Antibody. Gut Microbes 2024, 16, 2380061. [Google Scholar] [CrossRef]
- Oliero, M.; Cuisiniere, T.; Ajayi, A.S.; Gerkins, C.; Hajjar, R.; Fragoso, G.; Calvé, A.; Vennin Rendos, H.; Mathieu-Denoncourt, A.; Dagbert, F.; et al. Putrescine Supplementation Limits the Expansion of Pks+ Escherichia Coli and Tumor Development in the Colon. Cancer Research Communications 2024, 4, 1777–1792. [Google Scholar] [CrossRef]
- Zhong, B.; Zhao, Y.; Gao, L.; Yang, G.; Gao, Y.; Li, F.; Li, S. Anticancer Effects of Weizmannia Coagulans MZY531 Postbiotics in CT26 Colorectal Tumor-Bearing Mice by Regulating Apoptosis and Autophagy. Life 2024, 14, 1334. [Google Scholar] [CrossRef]
- Ma, C.; Duan, X.; Lei, X. 3D Cell Culture Model: From Ground Experiment to Microgravity Study. Front. Bioeng. Biotechnol. 2023, 11. [Google Scholar] [CrossRef]
- Clevers, H. Modeling Development and Disease with Organoids. Cell 2016, 165, 1586–1597. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Knoblich, J.A. Organogenesis in a Dish: Modeling Development and Disease Using Organoid Technologies. Science 2014, 345, 1247125. [Google Scholar] [CrossRef]
- Drost, J.; Clevers, H. Organoids in Cancer Research. Nat Rev Cancer 2018, 18, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Yuki, K.; Cheng, N.; Nakano, M.; Kuo, C.J. Organoid Models of Tumor Immunology. Trends in Immunology 2020, 41, 652–664. [Google Scholar] [CrossRef]
- Schnalzger, T.E.; de Groot, M.H.; Zhang, C.; Mosa, M.H.; Michels, B.E.; Röder, J.; Darvishi, T.; Wels, W.S.; Farin, H.F. 3D Model for CAR-mediated Cytotoxicity Using Patient-derived Colorectal Cancer Organoids. The EMBO Journal 2019, 38, e100928. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, K.K.; Cattaneo, C.M.; Weeber, F.; Chalabi, M.; Haar, J. van de; Fanchi, L.F.; Slagter, M.; Velden, D.L. van der; Kaing, S.; Kelderman, S.; et al. Generation of Tumor-Reactive T Cells by Co-Culture of Peripheral Blood Lymphocytes and Tumor Organoids. Cell 2018, 174, 1586–1598.e12. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, I.S.; Cardoso Galante, R.S.; Dua, K.; Malipeddi, V.R.; Awasthi, R.; Ghisleni, D.D.M.; de Jesus Andreoli Pinto, T. Hydrogel Based Drug Delivery Systems: A Review with Special Emphasis on Challenges Associated with Decontamination of Hydrogels and Biomaterials. Curr Drug Deliv 2017, 14, 917–925. [Google Scholar] [CrossRef]
- Park, Y.; Huh, K.M.; Kang, S.-W. Applications of Biomaterials in 3D Cell Culture and Contributions of 3D Cell Culture to Drug Development and Basic Biomedical Research. Int J Mol Sci 2021, 22, 2491. [Google Scholar] [CrossRef]
- Fan, Z.; Cheng, P.; Liu, M.; Prakash, S.; Han, J.; Ding, Z.; Zhao, Y.; Wang, Z. Dynamic Crosslinked and Injectable Biohydrogels as Extracellular Matrix Mimics for the Delivery of Antibiotics and 3D Cell Culture. RSC Adv 2020, 10, 19587–19599. [Google Scholar] [CrossRef]
- Bhatia, S.N.; Ingber, D.E. Microfluidic Organs-on-Chips. Nat Biotechnol 2014, 32, 760–772. [Google Scholar] [CrossRef]
- Puschhof, J.; Pleguezuelos-Manzano, C.; Clevers, H. Organoids and Organs-on-Chips: Insights into Human Gut-Microbe Interactions. Cell Host Microbe 2021, 29, 867–878. [Google Scholar] [CrossRef]
- Sato, T.; Stange, D.E.; Ferrante, M.; Vries, R.G.J.; Van Es, J.H.; Van den Brink, S.; Van Houdt, W.J.; Pronk, A.; Van Gorp, J.; Siersema, P.D.; et al. Long-Term Expansion of Epithelial Organoids from Human Colon, Adenoma, Adenocarcinoma, and Barrett’s Epithelium. Gastroenterology 2011, 141, 1762–1772. [Google Scholar] [CrossRef]
- Bartfeld, S. Modeling Infectious Diseases and Host-Microbe Interactions in Gastrointestinal Organoids. Dev Biol 2016, 420, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Heo, I.; Dutta, D.; Schaefer, D.A.; Iakobachvili, N.; Artegiani, B.; Sachs, N.; Boonekamp, K.E.; Bowden, G.; Hendrickx, A.P.A.; Willems, R.J.L.; et al. Modelling Cryptosporidium Infection in Human Small Intestinal and Lung Organoids. Nat Microbiol 2018, 3, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Fan, L.; Zhang, L.; Liu, Z. Comparative Analysis of Organoid, Air-Liquid Interface, and Direct Infection Models for Studying Pathogen-Host Interactions in Endometrial Tissue. Sci Rep 2025, 15, 8531. [Google Scholar] [CrossRef]
- Workman, M.J.; Gleeson, J.P.; Troisi, E.J.; Estrada, H.Q.; Kerns, S.J.; Hinojosa, C.D.; Hamilton, G.A.; Targan, S.R.; Svendsen, C.N.; Barrett, R.J. Enhanced Utilization of Induced Pluripotent Stem Cell-Derived Human Intestinal Organoids Using Microengineered Chips. Cell Mol Gastroenterol Hepatol 2018, 5, 669–677.e2. [Google Scholar] [CrossRef]
- Sugimura, N.; Li, Q.; Chu, E.S.H.; Lau, H.C.H.; Fong, W.; Liu, W.; Liang, C.; Nakatsu, G.; Su, A.C.Y.; Coker, O.O.; et al. Lactobacillus Gallinarum Modulates the Gut Microbiota and Produces Anti-Cancer Metabolites to Protect against Colorectal Tumourigenesis. Gut 2022, 71, 2011–2021. [Google Scholar] [CrossRef]
- Cho, Y.; Sung, M.-H.; Kang, H.-T.; Lee, J.H. Establishment of an Apical-Out Organoid Model for Directly Assessing the Function of Postbiotics. J Microbiol Biotechnol 2024, 34, 2184–2191. [Google Scholar] [CrossRef]
- Lee, H.; Jung,Kwang Bo; Kwon,Ohman; Son,Ye Seul; Choi,Eunho; Yu,Won Dong; Son,Naeun; Jeon,Jun Hyoung; Jo,Hana; Yang,Haneol; et al. Limosilactobacillus Reuteri DS0384 Promotes Intestinal Epithelial Maturation via the Postbiotic Effect in Human Intestinal Organoids and Infant Mice. Gut Microbes 2022, 14, 2121580. [Google Scholar] [CrossRef]
- Furone, F.; Bellomo, C.; Carpinelli, M.; Nicoletti, M.; Hewa-Munasinghege, F.N.; Mordaa, M.; Mandile, R.; Barone, M.V.; Nanayakkara, M. The Protective Role of Lactobacillus Rhamnosus GG Postbiotic on the Alteration of Autophagy and Inflammation Pathways Induced by Gliadin in Intestinal Models. Front. Med. 2023, 10. [Google Scholar] [CrossRef]
- Janssen, A.W.F.; van der Lugt, B.; Duivenvoorde, L.P.M.; Vos, A.P.; Bastiaan-Net, S.; Tomassen, M.M.M.; Verbokkem, J.A.C.; Blok-Heimerikx, E.; Hooiveld, G.J.E.J.; van Baarlen, P.; et al. Comparison of IPSC-Derived Human Intestinal Epithelial Cells with Caco-2 Cells and Human in Vivo Data after Exposure to Lactiplantibacillus Plantarum WCFS1. Sci Rep 2024, 14, 26464. [Google Scholar] [CrossRef]
- Zaharuddin, L.; Mokhtar, N.M.; Muhammad Nawawi, K.N.; Raja Ali, R.A. A Randomized Double-Blind Placebo-Controlled Trial of Probiotics in Post-Surgical Colorectal Cancer. BMC Gastroenterology 2019, 19, 131. [Google Scholar] [CrossRef]
- Bajramagic, S.; Hodzic, E.; Mulabdic, A.; Holjan, S.; Smajlovic, S.V.; Rovcanin, A. Usage of Probiotics and Its Clinical Significance at Surgically Treated Patients Sufferig from Colorectal Carcinoma. Med Arch 2019, 73, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, F.; Adewiah, S.; Fatchiyah, F. The Level Short Chain Fatty Acids and HSP 70 in Colorectal Cancer and Non-Colorectal Cancer. Acta Inform Med 2018, 26, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Sadaghian Sadabad, M.; Gabarrini, G.; Lisotto, P.; von Martels, J.Z.H.; Wardill, H.R.; Dijkstra, G.; Steinert, R.E.; Harmsen, H.J.M. Riboflavin Supplementation Promotes Butyrate Production in the Absence of Gross Compositional Changes in the Gut Microbiota. Antioxidants & Redox Signaling 2023, 38, 282–297. [Google Scholar] [CrossRef]
- Huang, F.; Li, S.; Chen, W.; Han, Y.; Yao, Y.; Yang, L.; Li, Q.; Xiao, Q.; Wei, J.; Liu, Z.; et al. Postoperative Probiotics Administration Attenuates Gastrointestinal Complications and Gut Microbiota Dysbiosis Caused by Chemotherapy in Colorectal Cancer Patients. Nutrients 2023, 15, 356. [Google Scholar] [CrossRef]
- D’Argenio, G.; Mazzacca, G. Short-Chain Fatty Acid in the Human Colon. In Advances in Nutrition and Cancer 2; Zappia, V., Della Ragione, F., Barbarisi, A., Russo, G.L., Iacovo, R.D., Eds.; Springer US: Boston, MA, 1999; pp. 149–158 ISBN 978-1-4757-3230-6.
- Compare, D.; Nardone, G. Contribution of Gut Microbiota to Colonic and Extracolonic Cancer Development. Digestive Diseases 2011, 29, 554–561. [Google Scholar] [CrossRef]
- Cheng, P.; Zeng, W.; Li, L.; Huo, D.; Zeng, L.; Tan, J.; Zhou, J.; Sun, J.; Liu, G.; Li, Y.; et al. PLGA-PNIPAM Microspheres Loaded with the Gastrointestinal Nutrient NaB Ameliorate Cardiac Dysfunction by Activating Sirt3 in Acute Myocardial Infarction. Advanced Science 2016, 3, 1600254. [Google Scholar] [CrossRef]
- Sabatino, A.D.; Morera, R.; Ciccocioppo, R.; Cazzola, P.; Gotti, S.; Tinozzi, F.P.; Tinozzi, S.; Corazza, G.R. Oral Butyrate for Mildly to Moderately Active Crohn’s Disease. Alimentary Pharmacology & Therapeutics 2005, 22, 789–794. [Google Scholar] [CrossRef]
- Chien, S.-T.; Suydam, I.T.; Woodrow, K.A. Prodrug Approaches for the Development of a Long-Acting Drug Delivery Systems. Advanced Drug Delivery Reviews 2023, 198, 114860. [Google Scholar] [CrossRef]
- Egorin, M.J.; Yuan, Z.-M.; Sentz, D.L.; Plaisance, K.; Eiseman, J.L. Plasma Pharmacokinetics of Butyrate after Intravenous Administration of Sodium Butyrate or Oral Administration of Tributyrin or Sodium Butyrate to Mice and Rats. Cancer Chemother Pharmacol 1999, 43, 445–453. [Google Scholar] [CrossRef]
- Patnaik, A.; Rowinsky, E.K.; Villalona, M.A.; Hammond, L.A.; Britten, C.D.; Siu, L.L.; Goetz, A.; Felton, S.A.; Burton, S.; Valone, F.H.; et al. A Phase I Study of Pivaloyloxymethyl Butyrate, a Prodrug of the Differentiating Agent Butyric Acid, in Patients with Advanced Solid Malignancies. Clin Cancer Res 2002, 8, 2142–2148. [Google Scholar]
- Russo, R.; Santarcangelo, C.; Badolati, N.; Sommella, E.; De Filippis, A.; Dacrema, M.; Campiglia, P.; Stornaiuolo, M.; Daglia, M. In Vivo Bioavailability and in Vitro Toxicological Evaluation of the New Butyric Acid Releaser N-(1-Carbamoyl-2-Phenyl-Ethyl) Butyramide. Biomedicine & Pharmacotherapy 2021, 137, 111385. [Google Scholar] [CrossRef]
- Annison, G.; Illman, R.J.; Topping, D.L. Acetylated, Propionylated or Butyrylated Starches Raise Large Bowel Short-Chain Fatty Acids Preferentially When Fed to Rats. The Journal of Nutrition 2003, 133, 3523–3528. [Google Scholar] [CrossRef] [PubMed]
- Bell, K.J.; Saad, S.; Tillett, B.J.; McGuire, H.M.; Bordbar, S.; Yap, Y.A.; Nguyen, L.T.; Wilkins, M.R.; Corley, S.; Brodie, S.; et al. Metabolite-Based Dietary Supplementation in Human Type 1 Diabetes Is Associated with Microbiota and Immune Modulation. Microbiome 2022, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Toden, S.; Lockett,Trevor J; Topping,David L; Scherer,Benjamin L; Watson,Emma-Jane L; Southwood,Jessica G; and Clarke, J. M. Butyrylated Starch Affects Colorectal Cancer Markers Beneficially and Dose-Dependently in Genotoxin-Treated Rats. Cancer Biology & Therapy 2014, 15, 1515–1523. [Google Scholar] [CrossRef]
- Clarke, J.M.; Young, G.P.; Topping, D.L.; Bird, A.R.; Cobiac, L.; Scherer, B.L.; Winkler, J.G.; Lockett, T.J. Butyrate Delivered by Butyrylated Starch Increases Distal Colonic Epithelial Apoptosis in Carcinogen-Treated Rats. Carcinogenesis 2012, 33, 197–202. [Google Scholar] [CrossRef]
- Dianzani, C.; Foglietta, F.; Ferrara, B.; Rosa, A.C.; Muntoni, E.; Gasco, P.; Pepa, C.D.; Canaparo, R.; Serpe, L. Solid Lipid Nanoparticles Delivering Anti-Inflammatory Drugs to Treat Inflammatory Bowel Disease: Effects in an in Vivo Model. World Journal of Gastroenterology 2017, 23, 4200–4210. [Google Scholar] [CrossRef]
- Mu, Y.; Kinashi, Y.; Li, J.; Yoshikawa, T.; Kishimura, A.; Tanaka, M.; Matsui, T.; Mori, T.; Hase, K.; Katayama, Y. Polyvinyl Butyrate Nanoparticles as Butyrate Donors for Colitis Treatment. ACS Appl. Bio Mater. 2021, 4, 2335–2341. [Google Scholar] [CrossRef]
- Chen, L.; Zhao, R.; Kang, Z.; Cao, Z.; Liu, N.; Shen, J.; Wang, C.; Pan, F.; Zhou, X.; Liu, Z.; et al. Delivery of Short Chain Fatty Acid Butyrate to Overcome Fusobacterium Nucleatum-Induced Chemoresistance. Journal of Controlled Release 2023, 363, 43–56. [Google Scholar] [CrossRef]
- Wang, L.; Cao, H.; Liu, L.; Wang, B.; Walker, W.A.; Acra, S.A.; Yan, F. Activation of Epidermal Growth Factor Receptor Mediates Mucin Production Stimulated by P40, a Lactobacillus Rhamnosus GG-Derived Protein *. Journal of Biological Chemistry 2014, 289, 20234–20244. [Google Scholar] [CrossRef]
- Yan, F.; and Polk, D.B. Characterization of a Probiotic-Derived Soluble Protein Which Reveals a Mechanism of Preventive and Treatment Effects of Probiotics on Intestinal Inflammatory Diseases. Gut Microbes 2012, 3, 25–28. [Google Scholar] [CrossRef]
- Gao, J.; Li, Y.; Wan, Y.; Hu, T.; Liu, L.; Yang, S.; Gong, Z.; Zeng, Q.; Wei, Y.; Yang, W.; et al. A Novel Postbiotic From Lactobacillus Rhamnosus GG With a Beneficial Effect on Intestinal Barrier Function. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Trend of Studies on postbiotics in the last ten years (2015-2024).
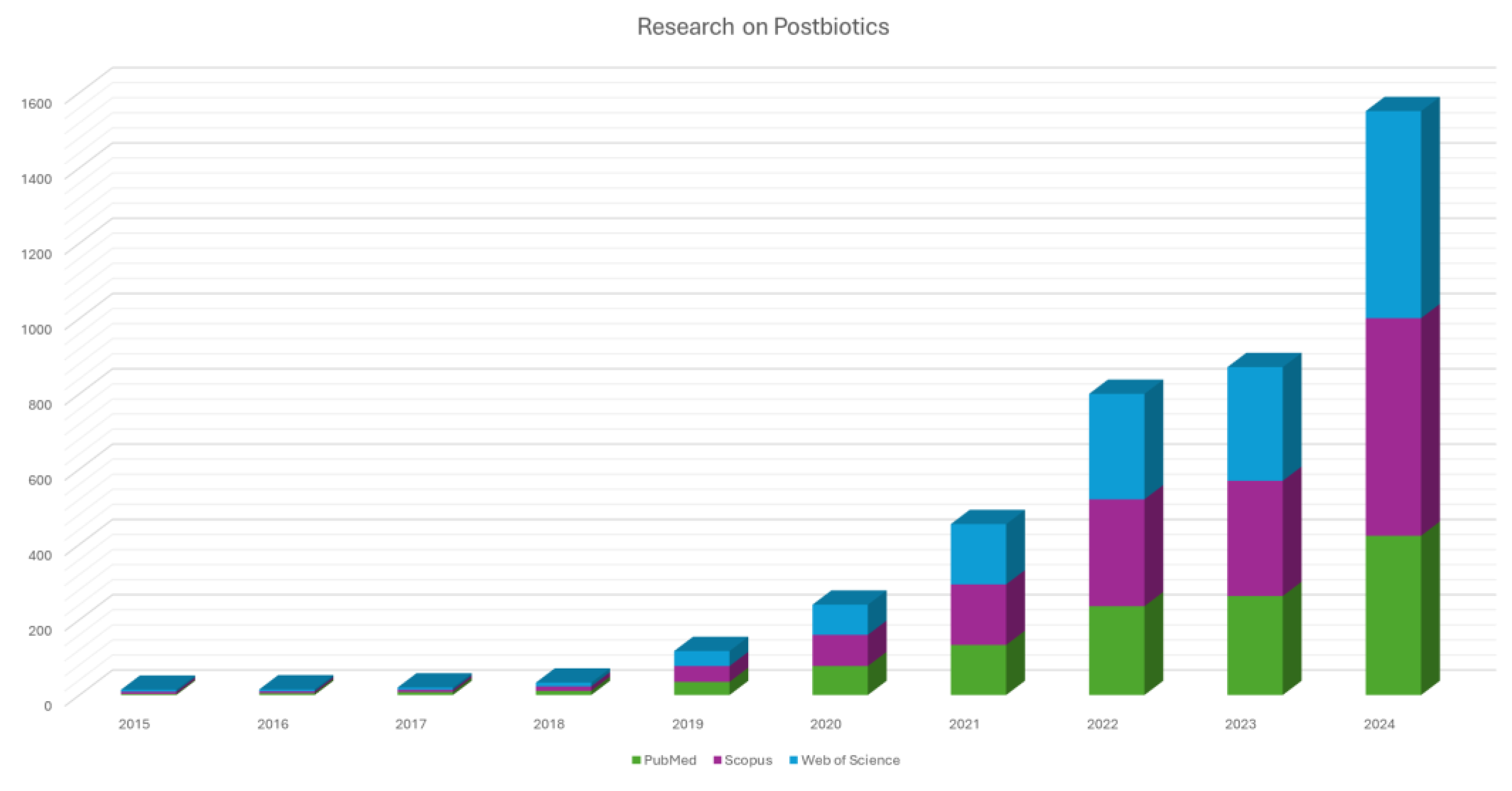
Figure 2.
Postbiotic bioactive metabolites.
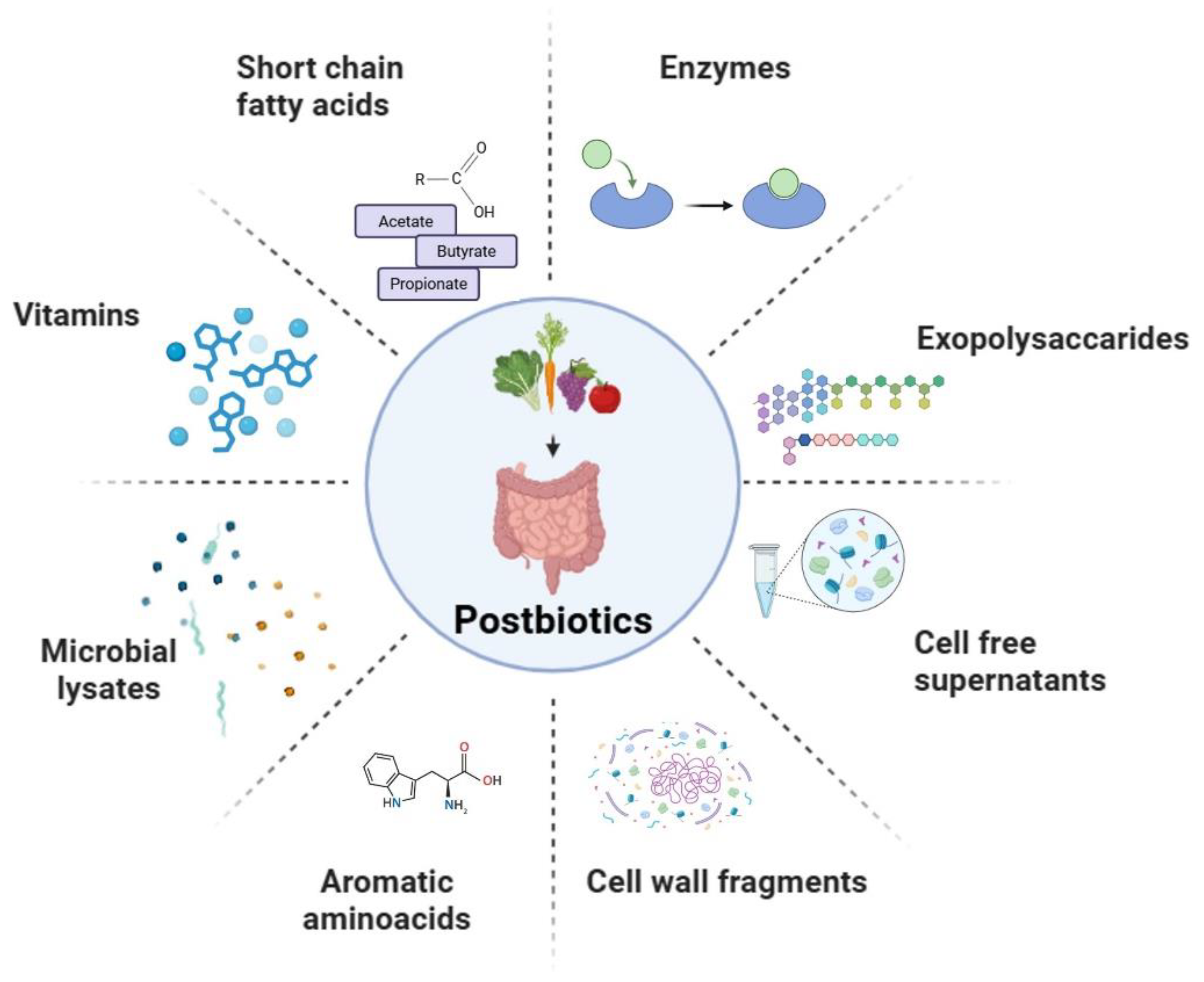
Figure 3.
Postbiotics from functional foods: mechanisms of action in colorectal cancer.
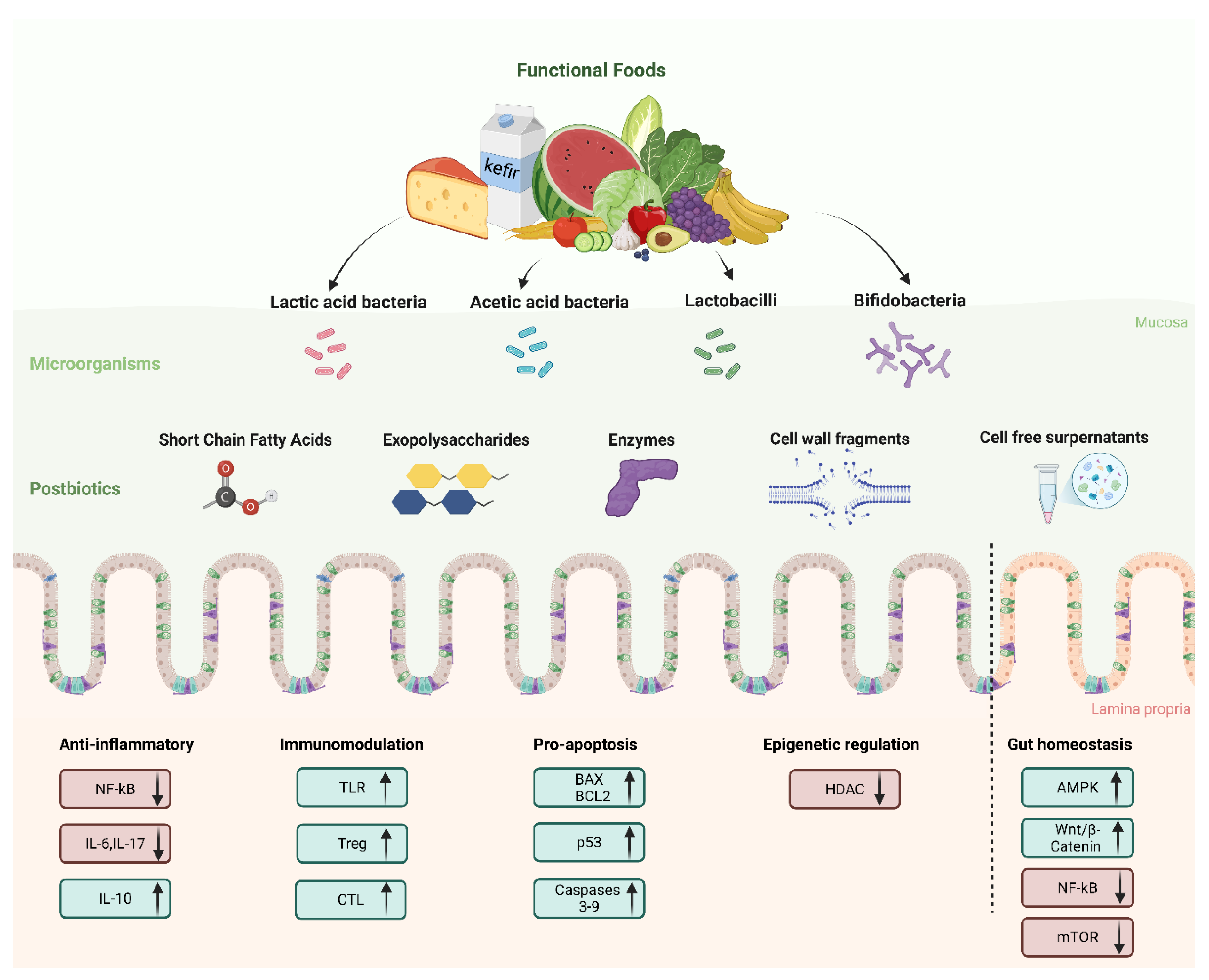
Figure 4.
Flow diagram for the extensive literature search of postbiotics and colorectal cancer.
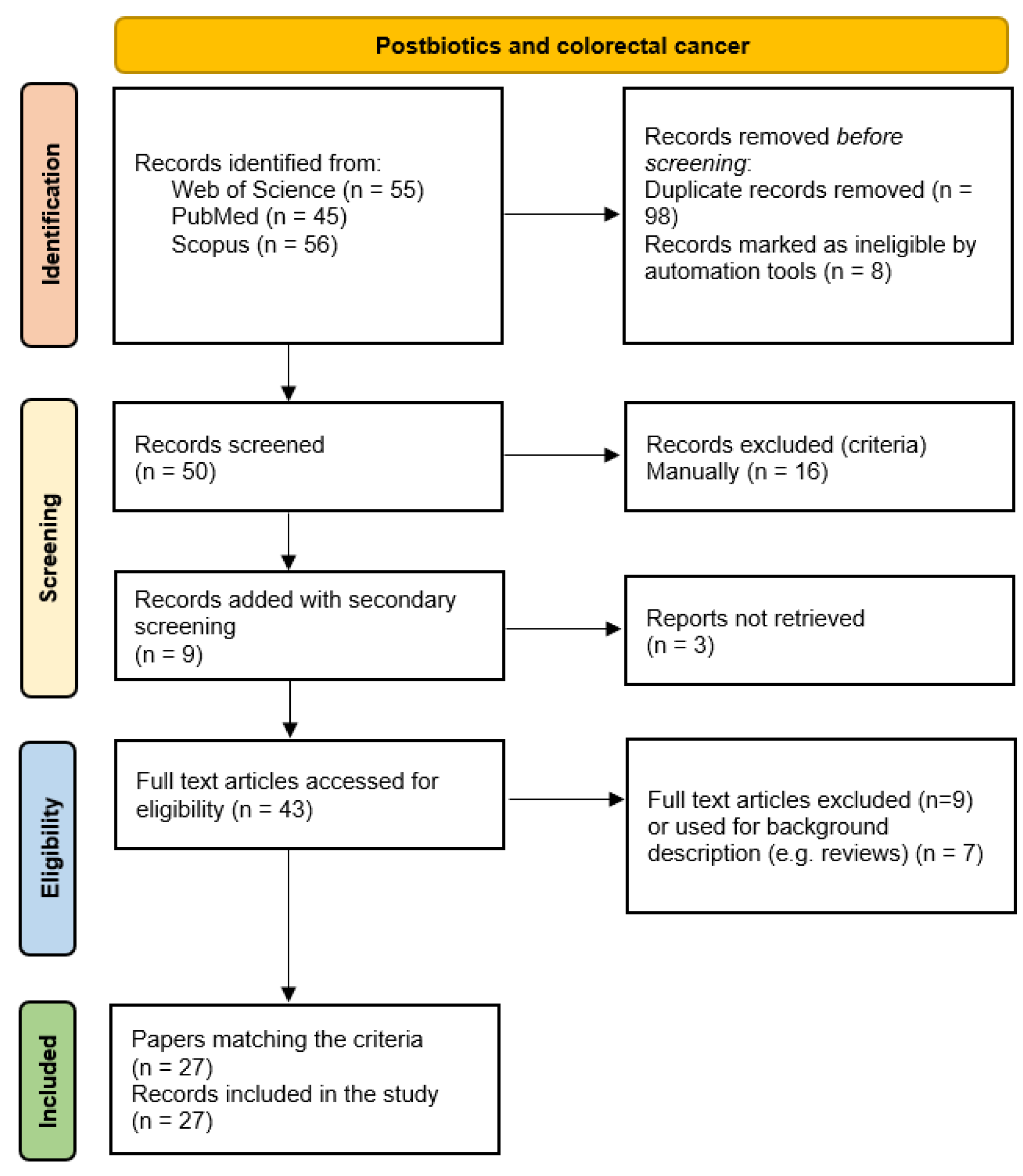

Table 1.
Inclusion and Exclusion Criteria for Systematic Review and Extensive Literature Search.
| Criterion | Decision | |
|---|---|---|
| Inclusion | Exclusion | |
| Default keywords and search terms exist as a whole or at least in the title, keywords, or abstract of the article | × | |
| The article is published in a peer-review scientific journal | × | |
| The article is written in English | × | |
| Studies where terms were referred to prebiotics, however supernatant/heat-killed cultures were used for testing/assessing | × | |
| Studies on diseases considered high risk factors for colorectal cancer and a relevant study model was developed/used | × | |
| Duplicate records | × | |
| The full text is not available | × | |
| Articles published before 2010 | × | |
| Only testing live microorganisms | × | |
| Studies on gut microbiota transplantation | × | |
Table 2.
Studies on postbiotics and colorectal cancer.
| ID | Model | Study type | Microorganism Strain/Species |
Molecules of interest | Events | Pathway/Gene involved | Notes | Ref |
|---|---|---|---|---|---|---|---|---|
| 1 | HCT-116 | in vitro | Lactobacillus casei and Lactobacillus rhamnosus GG | cell free supernatant | decreasing matrix metalloproteinase-9 (MMP-9) and increasing the tight junction protein zona occludens-1 (ZO-1) levels | cell invasion | [131] | |
| 2 | HT-29 | in vitro | 7 strains of Lactobacillus | cell free supernatant | lactate dehydrogenase regulation | apoptosis | [133] | |
| 3 | HCT-116 HT-29 |
in vitro | synthetic | sodium butyrate | autophagy | LKB1–AMPK pathway | [135] | |
| 4 | HT-29 | in vitro |
Lactobacillus plantarum |
bacteriocins | antiproliferative effect | apoptosis | study on several cancer cell lines | [136] |
| 5 | Caco-2 | in vitro |
Lactobacillus acidophilus |
exopolysaccharide | upregulation of the expression of PPAR-γ | [138] | ||
| 6 | HT-29 HCT-116 |
in vitro | Propionibacterium freudenreichii | culture supernatant, metabolites (propionate/acetate) | increased pro-apoptotic gene expression (TRAIL-R2/DR5) and decreased anti-apoptotic gene expression (FLIP, XIAP); death receptors (TRAIL-R1/DR4, TRAIL-R2/DR5) and caspases (caspase-8, -9 and -3) activation; Bcl-2 expression inhibition | extrinsic apoptotic pathway | in combination with TNF-Related Apoptosis-Inducing Ligand (TRAIL) | [139] |
| 7 | LS174T | in vitro | Lactobacillus acidophilus and Bifidobacterium longum | butyrate | dose-dependent increase in mucin protein contents; increased transcriptional levels of MUC3, MUC4, and MUC12 | MAPK signaling pathway | doses: 6 or 9 mM | [140] |
| 8 | scRNA-seq analysis and DEGs analysis HT-29 human dermal fibroblasts |
in silico and in vitro | Lactobacillus acidophilus ATCC4356 | cell free supernatant | cell cycle arrest at G1 phase, anti-proliferative and anti-migration effects, anti-proliferative activity on control fibroblasts. | Wnt signaling (SFRP1, SFRP2, SFRP4, MMP7) | [142] | |
| 9 | HT-29 human dermal fibroblast |
in vitro | Bifidobacterium breve and Lactobacillus rhamnosus | cell free supernatant | anti-proliferation, anti-migration, and apoptosis-related effects | apoptosis: Bax/Bcl2/caspase-3; Wnt signaling: RSPO2, NGF, MMP7 | [143] | |
| 10 | Caco-2 | in vitro | Lactobacillus casei | cell free supernatant | tumor cells cytotoxic effect | apoptosis | comparison of probiotic (live), paraprobiotic (heat-killed) and postbiotics (CFS) | [132] |
| 11 | HT-29 | in vitro | Gluconobacter oxydans strains isolated from Kombucha (KNS30, KNS31, KNS32, K1, and K2) | gluconic acid, glucuronic acid, acetic acid, pyruvic acid, fumaric acid, and lactic acid |
tumor cells cytotoxic effect | apoptotic/necrotic: annexin V and PI positive | study also on gastric cell line: AGS; HUVEC cell lines used as control | [144] |
| 12 | HT-29 HCT-116 |
in vitro | Lactobacillus lactis | cell free supernatant | depletion of arginine, decreased levels of c-Myc, reduced phosphorylation of p70-S6 kinase | cell cycle arrest | [134] | |
| 13 | NCM460 Caco-2 HT-29 |
in vitro | Lacticaseibacillus paracasei and Lactiplantibacillus plantarum | heat-inactivated cultures | downregulation of Orai1 and STIM1 | FAK pathway (Store-operated calcium entry) | [137] | |
| 14 | HT-29 | in vitro | Saccharomyces boulardii | cell free supernatant | increased expression of Caspase3 and PTEN genes; decreased expression of RelA and Bcl-XL genes | apoptosis | [117] | |
| 15 | HT-29 Fnevs infection model | in vitro | Bifidobacterium longum | cell free supernatant | inhibition of proliferation, migration and invasion | inhibitory effects on the expression of specific oncogenes (e.g., Myc, IL16, KCNN2, ACSBG1, Pum1, MET, NR5A2) | controversial results | [141] |
| 16 | mouse colon carcinoma CT26.WT tumor cells were injected subcutaneously - BALB/c mice | in vivo | Weizmannia coagulans MZY531 | powder of W. coagulans MZY531; oligosaccharidesuspension | inhibiting tumor growth by modulating apoptosis and autophagy in tumor cells | apoptosis: Bax/Bcl2/caspase-3 & JAK2/STAT3 Autophagy: PI3K/AKT/mTOR & TGF-β/SMAD4 | [151] | |
| 17 | Sprague–Dawley rats | in vivo | Lactobacillus rhamnosus MD 14 | metabiotic extract (acetate, butyrate, propionate, acetamide, thiocyanic acid, and oxalic acid) | downregulation of oncogenes (K-ras, β-catenin, Cox-2, nuclear factor kappa B (NF-κB)] and upregulation tumor suppressor p53 gene leading to almost normal colon histology | Wnt/β-Catenin Pathway | active components in the metabiotic extract were characterized byLC-MS | [146] |
| 18 | xenograft mousemodel CT-26 cells subcutaneously injected into BALB/c mice |
in vivo ex vivo |
multiple strains of probiotics and yeast | MS-20 “Symbiota®” in combination with anti-programmed cell death 1 (PD1) antibody | inhibited colon and lung cancer growth | CD8+ T cells and PD1 expression | fecal samples from six patients were used for ex vivo evaluation | [149] |
| 19 | C57BL/6 mouse model where cancer was induced via AOM/DSS administration | in vivo | Escherichia coli Nissle 1917 | putrescine | inhibit the growth of thepathogenic strain pks+ E. coli NC101; reduced the number and size of colonic tumors, regulation of inflammatory cytokines; shift in the composition of gut microbiota | cell proliferation; fecal Lcn-2 marker of inflammation in inflammatory bowel diseases, TNFα, IL6 and IL10; 16S rRNA amplicon sequencing | [150] | |
| 20 | xenograft models obtained by injecting SW620 cells into male BALB/c nude mice Caco-2/bbe SKCO-1 SW620 |
in vivo in vitro |
Lactobacillus casei ATCC334 | ferrichrome | activation of the JNK-DDIT3-mediated apoptotic pathway | JNK-DDIT3-mediated apoptotic pathway | effect of ferrichrome was compared with 5-FU and cisplatin | [119] |
| 21 | C57BL/6 mouse model where cancer was induced via AOM/DSS administration | in vivo | Lactiplantibacillus plantarum-12 | exopolysaccharide | activation of caspase cascade and NF-κB signaling (IκB-α, p65, p-p65, p38, and p-p38) | inflammatory signaling and apoptosis | additional untargeted fecal metabolomic analysis | [147] |
| 22 | BALB/c mice CRC models induced via AOM/DSS administration | in vivo | Lactobacillus acidophilus | lysates | increased CD8 + T cell and effector memory T cells, decreased Treg and M2 macrophages | TLR signaling pathway | combination with CTLA-4-blocking antibodies | [148] |
| 23 | C57B/6 mice model CRC cell lines organoids from CRC patients |
in vitro in vivo organoids |
Lactobacillus gallinarum | cell free supernatant (indole-3-lactictate most enriched metabolite) | antitumorigenic role: proliferation, apoptosis, cell cycle distribution, gut microbiota modulation |
cell proliferation apoptosis |
[169] | |
| 24 | Organoids derived from C57BL/6 male mice small intestines and colon |
in vitro in vivo organoids |
Lactiplantibacillus plantarum KM2 & Bacillusvelezensis KMU01 | cell free supernatant | inflammatory response LPS-induced and mitochondrial homeostasis through mitophagy and mitochondrial biogenesis | COX-2 decreased; expression of tight-junctionmarkers ZO-1, claudin, and occludin increased, and expression of mitochondrial homeostasisfactors PINK1, parkin, and PGC1a also increased. | [170] | |
| 25 | hPSC-derived intestinal organoids C57BL Mice Caco-2 |
in vitro in vivo organoids |
Limosilactobacillus reuteri DS0384 | N-carbamyl glutamic acid (NCG) | intestinal epithelial maturation; inflammatory response and intestinal epithelial barrier integrity | mature specific marker: (CDX2), (OLFM4), (DEFA5 and LYZ), (KRT20, CREB3L3, DPP4, LCT, SLC5A1, and MUC13); Inflammatory pathway: (IFNγ)/TNFα, IL-1β, IL-6, IL-8, and TNFα; localization of zonula occludens-1 | [171] | |
| 26 | Caco-2 organoids derived from celiac disease patientbiopsies |
in vitro organoids |
Lactobacillus rhamnosus GG | cell free supernatant | alteration of autophagy and inflammation pathways induced by gliadin in Celiac disease (CD) |
mTOR pathway: phosphorylation of p70S6K, p4EBP-1; inflammatory marker: NF- kb; autophagy: LC3II and p62 protein, SQSTM1 autophagosome membrane marker |
[172] | |
| 27 | Caco-2 hiPSC derived IEC monolayers |
in vitro advanced patient-derived in vitro |
Lactiplantibacillus plantarum | heat-killed | inflammatory response | IL-8, REG3α and HBD2 | [173] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



