Submitted:
03 May 2025
Posted:
06 May 2025
You are already at the latest version
Abstract
Colorectal cancer (CRC) is a major global health concern, particularly in Western countries where there is high consumption of processed food. Gut microbiota, intestinal inflammation, and autophagy play a pivotal role in CRC initiation and progression, and probiotics have emerged as potential preventive and adjuvant therapeutics by restoring a balanced gut microbiota and dampening inflammation. Probiotics exert their anti-inflammatory effects through several mechanisms, including the modulation of immune responses, the production of anti-inflammatory molecules (such as short-chain fatty acids), the reduction of pro-inflammatory cytokines secretion, and the regulation of inflammatory signaling pathways. Moreover, probiotic metabolites have been shown to restore intestinal epithelial homeostasis and barrier integrity by modulating autophagy, a macromolecular degradation process that keeps cell proliferation, adhesion, and migration under check and that also influences gut microbiota. This narrative review discusses the current evidence supporting the anti-inflammatory effects of probiotics and explores their potential preventive and therapeutic applications in CRC management.
Keywords:
butyrate
; APC
; autophagy
; inflammation
; microbiota
; cytokines
; cancer
Introduction
Colorectal Cancer: Epidemiology, Risk Factors, Prevention and Therapy at a Glance
Worldwide statistics indicate that colorectal cancer (CRC) incidence ranks third and mortality second (after lung) in 2022, being now the first-leading and second-leading cause of cancer death in men and women, respectively [1]. The highest incidence rates are reported in high-income and well-developed regions, including Europe, Australia, North America, and Eastern Asia, suggesting a link with lifestyle and dietary factors [2]. The role of diet and lifestyle in colorectal carcinogenesis is further supported by the observation that its incidence is constantly increasing in developing countries over the last decades [3,4]. Additional factors associated with an increased risk of developing sporadic CRC include age over 50, male sex, obesity, and a sedentary lifestyle [5]. Having an impact on gut microbiota, alcohol consumption, and dietary choices constitute the most important modifiable risk factors for CRC [6,7,8]. However, the most significant risk factors for developing CRC are genetic predisposition (inherited mutations in relevant genes), a family history of CRC, and chronic inflammatory conditions of the gastrointestinal tract [9,10].
Surgery remains the gold standard treatment for CRC, although in most cases it is preceded or followed by adjuvant chemotherapy and radiotherapy to shrink or stabilize the tumor [11,12]. Standard chemotherapy protocols include monotherapy with 5-fluorouracil (5-FU) and multi-drug therapy combining 5-FU with leucovorin and oxaliplatin or with leucovorin and irinotecan [11]. More recently, novel strategies such as target therapy, gene therapy, and immunotherapy have been introduced to treat CRC [13]. Early detection of CRC through colonoscopy and modification of lifestyle risk factors are crucial for prevention and reduce the risk of developing the tumor [2]. Recently, prebiotics, probiotics, and postbiotics have emerged as potent dietary supplements for the prevention and post-surgery adjuvant therapy of CRC [14].
1. Colorectal Cancerogenesis: The Role of Genetics and Epigenetics
Most cases of CRC are sporadic and develop from polyps through the adenoma-carcinoma sequence following a characteristic sequential accumulation of genetic and epigenetic mutations in well-defined oncogenes, tumor suppressor genes, and DNA repair genes in 10 years or more [15,16]. Epigenetic regulation of oncogenes or tumor suppressor genes expression also contributes to CRC carcinogenesis [17]. CRC hereditary predisposition is linked to germline inherited monoallelic mutations in tumor suppressor or DNA repair genes [18]. Inactivation (by mutation or epigenetic silencing) of the second allele creates the conditions for the initiation step of tumorigenesis, which can be followed by additional mutations or epimutations in oncogenes and tumor suppressor genes that, eventually, will lead to frank carcinoma. The two most common forms of hereditary predisposition to CRC are hereditary non-polyposis colorectal cancer (HNPCC), with Lynch syndrome as the most common, and familial adenomatous polyposis (FAP). Lynch syndrome is characterized by mutation in DNA mismatch repair genes (MLH1, MSH2, MSH6, PMS2) leading to microsatellite instability [19]. FAP is an autosomal dominant condition caused by a germline mutation in the APC (Adenomatous Polyposis Coli) gene. It is characterized by the development of over 100 polyps at a young age. If left untreated, these polyps have a high likelihood of progressing to CRC within a few years [20]. In Lynch syndrome, the transition from adenoma to carcinoma occurs more rapidly, with an estimated progression time of 35 months [21].
Colorectal carcinogenesis involves three primary molecular pathways: chromosomal instability (CIN), microsatellite instability (MSI), and the CpG island methylator phenotype (CIMP). CIN is the most common type of molecular pathway responsible for sporadic CRC, affecting genes involved in key pathways critical for carcinogenesis, such as APC, KRAS, PI3KCA, and TP53. CIN is also involved in the development of FAP, as germline mutation in the APC gene is found in 60-80% of FAP cases [22]. Mutations in APC are often followed by mutations in KRAS, which are present in 40% of CRC cases [16]. MSI, occurring in 95% of Lynch syndrome cases and 15% of sporadic CRC, is caused by accumulated germline mutations in mismatch repair (MMR) genes, including MLH1, MSH2, MSH3, MSH6, and PMS2 [23]. These mutations lead to instability within microsatellite regions, which are repeated tandem sequences of nucleotides. As DNA polymerase struggles to bind these repetitive sequences, errors accumulate, further disrupting genome stability [16]. A small percentage (20%) of CRC cases exhibit CIMP, where the promoter of 3-5 tumor suppressor genes (e.g., p16, MGMT, TIMP3, p14, FHIT, SLC5A8) are hypermethylated due to upregulation of DNA methyltransferases (DNMT3B or DNMT1) [24]. The high-CIMP is also known as the serrated pathway, where the BRAF V600E mutation is a key event determining a fatal outcome [25].
The earliest initiating event in colorectal carcinogenesis is an inactivation mutation in the APC tumor suppressor gene in normal colonic cells, which leads to polyp formation and the development of dysplastic crypts. This is followed by an additional mutation in the oncogene KRAS and the tumor suppressor gene DCC (Deleted in Colorectal Cancer), resulting in the formation of adenomas. As the adenomas progress to carcinoma, the tumor-suppressor gene TP53 is inactivated, playing a key role in the transition to malignancy [15]. Once the adenocarcinoma becomes invasive, it can spread primarily to the liver, lungs, and bones, leading to an incurable metastatic disease [22].
The Wnt/β-Catenin pathway is a highly conserved signaling mechanism that plays a critical role in cell survival, proliferation, migration, and differentiation. Activation of the Wnt/β-Catenin pathway occurs when Wnt ligands bind to Frizzled receptors (FZD), triggering downstream signaling events that promote the nuclear translocation of β-Catenin to activate the transcription of target genes including CYCLIN D1, c-MYC, PDK, MCT-1, and MMP7 [26,27]. Additionally, hyperactivation of the Wnt/β-Catenin pathway induces epithelial to mesenchymal transition (EMT) through the transcription factors SNAIL1, SLUG, ZEB1, and TWIST [28], and also contributes to altered glucose metabolism, inducing aerobic glycolysis (so called “Warburg effect”) through hyper-expression of key glycolytic enzymes such as PDK1 and MCT-1, that shift energy metabolism towards glycolysis [29,30]. Loss-of-function mutations in the oncosuppressor APC, mutations in CTNNB1 (β-Catenin gene), and the excessive presence of Wnt ligands in the tumor microenvironment (TME) are critical drivers of colorectal carcinogenesis [31]. Wild-type APC regulates the transition from the G0/G1 to the S phase by inhibiting Wnt signaling and promoting the ubiquitin-mediated degradation of β-Catenin, a key transcription factor. Mutations in APC lead to the activation of downstream targets, including cyclin D1 and myc, which contribute to uncontrolled cell growth [32,33]. Cyclin D1, along with cyclin-dependent kinases (CDKs) like p27 and p21, is essential for regulating cell cycle progression, particularly during the G1 to S phase transition [34]. APC also plays a crucial role in processes such as chromosome segregation, migration, apoptosis, and differentiation, which become deregulated in the presence of mutated APC [32,33].
Mutated KRAS leads to the constitutive activation of the mitogen-activated protein kinase (MAPK) pathway, independent of EGFR activation, resulting in uncontrolled cell growth, proliferation, survival, migration, and invasion [35]. 10-20% of CRC cases with mutated KRAS also present mutations in PI3K during the late stages of tumor progression [36]. Activated PI3K mutations lead to the activation of the AKT-mTOR pathway, which results in downregulation of apoptosis, inhibition of autophagy, and promotion of cell survival, protein synthesis, and proliferation [37].
2. Colorectal Cancerogenesis: The Role of Autophagy
Autophagy is a catabolic process highly conserved in eukaryotic cells that maintains cell homeostasis through the lysosome-mediated degradation of damaged (unfolded, oxidized), aged, or redundant cellular components as well as the destruction of intracellular pathogens. Based on the mechanism that directs the degrading material (cargo) to the lysosome, autophagy is classified into three types: (i) macroautophagy (where the cargo is sequestered within the autophagosomes), (ii) microautophagy (small portion of the cytoplasm is internalized by invagination of the lysosome membrane), and (iii) chaperon-mediated autophagy (internalization of selected proteins is mediated by HSC73) [40]. A more in-depth description of the autophagy machinery, its biochemical regulation by extracellular signals and by genetics and epigenetics can be found in excellent reviews [40,41,42]. Here, we will only give a glance at macroautophagy (simply referred to as autophagy) since this pathway plays the major role in macromolecular degradation and turnover. This process is characterized by the sequestration of the cargo within autophagosomes (double-membrane organelles) that subsequently fuse with endosomes and lysosomes, forming the autolysosomes in which the cargo is fully degraded, and the substrates are exported in the cytosol for reutilization [40]. Autophagy is repressed by growth signals and abundancy of nutrients (glucose, amino acids) through the activation of mTORC1, while it is induced in conditions of nutrient depletion and reduced oxidative phosphorylation through the activation of AMPK [40]. Autophagy is a pro-survival pathway that opposes apoptosis, yet when hyper-induced because of overwhelming cellular toxicity, it can precipitate autophagic cell death [40]. This is reflected in cancer, where autophagy may play two opposite roles: (i) as anti-cancer it can prevent carcinogenesis by eliminating damaged subcellular structure and protecting the genome, by contrasting cell mass growth and by inhibiting cell proliferation and cell migration and by promoting cancer cell dormancy, whereas as (ii) pro-cancer it can favor survival in floating metastatic cells and can protect the cancer cell from the chemo- and radio-induced damages [43]. Autophagy also plays a role in reshaping the TME by reducing angiogenesis, inflammation, relieving immune suppression, favoring tumor dormancy, and promoting the reversal of the phenoconversion of cancer-associated fibroblasts (CAFs) into normal fibroblasts [44,45,46]. Stimulation of autophagy also re-educates M2 tumor-associated macrophages [47].
The pro- and anti-cancer role of autophagy in colorectal carcinogenesis is context-dependent [48,49] (Figure 1). In certain settings, autophagy contrasts with the induction of apoptosis in CRC cells [50,51]. On the other hand, there is evidence that supports the anti-cancer role of autophagy in inhibiting CRC growth, progression, and metastasis [52,55]. Consistent with a beneficial effect of autophagy in CRC progression, it has been reported that increased expression of the autophagy-related tumor suppressor gene BECLIN-1 associates with better overall survival in CRC patients undergoing chemotherapy [56]. Reinforcing the anti-cancer role of autophagy is the observation that autophagy may prevent the onset of intestinal inflammatory diseases that may result in CRC [57], improve the immune system functions, modulate the production of inflammatory cytokines, and improve pathogen clearance [58,59].
3. Colorectal Cancerogenesis: The Role of Inflammation
Chronic inflammation is a crucial hallmark of cancers [60]. Inflammatory immune cells and the associated cytokines in the TME are thought to play a double-faced role in the initiation and progression of CRC [61]. Inflammation starts as a defensive response of the immune system to pathogens or tissue damage. In response to intestine-localized inflammation, cytokines produced by gut immune cells (TNF-α, IL-1β, and IL-6) stimulate the crosstalk among the gut microenvironment cells [61]. The persistent activation of the inflammatory response leads to an excess release of bioactive molecules such as cytokines, chemokines, and growth factors that eventually contribute to malignant transformation, cancer growth, and progression [62]. The sequential phases of induction of inflammation, stimulation of proliferation, and tumorigenesis following microbial dysbiosis and metabolic alterations have been clearly elucidated in animal models of colitis-associated colorectal cancer (CAC) [63].
Inflammatory bowel disease (IBD) is associated with oxidative stress and DNA damage, resulting in genetic alterations [57,64], which stimulate proliferation of epithelial cells, cause dysplasia and, over time, dysplasia, eventually leading to tumor development. [65]. Accordingly, IBD patients have approximately a two-to three-fold increased risk of developing CRC compared to the general population [66,67].
Cancer cells, CAFs, tumor-associated macrophages (TAMs), and immune cells release a plethora of inflammatory cytokines that ultimately affect the microbiota, further contributing to the creation of a permissive TME for the growth and metastasization of CRC [68]. CRC stromal compartment includes fibroblasts, macrophages, and immune cells secreting a variety of cytokines, among other immune mediators. IL-6 is a master inflammatory cytokine promoting the development of CRC through multiple pathways [69,70]. The release of IL-6 by CRC-associated fibroblasts stimulates VEGF-mediated angiogenesis, inhibits CRC cell apoptosis, and induces CRC proliferation and motility [71,72]. Further, IL-6 induces the release of TGF-β, thus promoting EMT, activates the proliferation pathways (such as ERK/MAPK, PI3K, and Wnt/β-Catenin), promotes the M2 TAMs polarization, and suppresses the immune response [73,74,75,76]. The serum levels of IL-6 increase during the progression from colorectal adenoma to carcinoma, and, consistently, CAC and CRC patients with high serum levels of IL-6 present with tumors of large size, relapse, and shorter overall survival [77].
Macrophages are key players in the TME, where they can dynamically change their phenotype from M1-like to M2-like depending on the extracellular stimuli. M1-like macrophages, identified by the expression of CD80 and CD86, show a glycolytic metabolism and exert a pro-inflammatory and anti-tumor function, while M2-like macrophages (and their subtypes), identified by the expression of CD206 and CD163, show a phosphorylation oxidative metabolism and exert an anti-inflammatory and pro-tumorigenic function [78,79,80]. CRC cells co-cultured with CD68, CD204, and CD206-positive M2-like macrophages showed an increased rate of proliferation and colony formation, and this effect was abrogated when the M2-like macrophages were pre-exposed to rapamycin to induce autophagy [47].
Finally, dysbiotic microbiota may stimulate a chronic inflammatory status that may evolve into a high grade of dysplasia and then cancer due to the release of toxins, reduced production of beneficial metabolites, and disruption of epithelial integrity [81].
4. Dysbiosis and Colorectal Cancerogenesis: The Role of Diet and Microbiota
CRC incidence is strongly associated with dietary habits [82]. The western diet, characterized by a high intake of red and processed meats, saturated fats, alcohol, and sugar, is associated with an increased risk of CRC [83,84]. This is linked to the oxidative metabolism of foods that typically make up the western diet [85]. A dietary pattern rich in fats associated with inflammatory adipose tissue contributes to colorectal carcinogenesis [86]. In contrast, diets rich in low-fat dairy products such as fruits, vegetables, legumes, fiber, whole grains, and fish, combined with moderate alcohol intake, have been shown to have a protective effect against CRC. This protective effect is attributed to their content in prebiotics and specific micronutrients, such as calcium and magnesium, as well as vitamins like vitamin D and vitamin B6, and polyphenols [7,87].
Dietary patterns significantly influence the gut microbiota [8,88]. Changes in its composition, particularly a reduction in microbial diversity, can result in dysbiosis, a condition that plays a critical role in CRC by triggering malignant transformations in intestinal cells [8,89].
The gut microbiota is a dynamic community of several types of microbes, including archaea, eukaryotes, bacteria, viruses, and parasites, that have evolved to grow and survive in the gastrointestinal tract. Within the microbial community, bacteria are the most abundant group, comprising approximately 39 trillion individuals [57]. These bacteria belong to several phyla, including Firmicutes, Bacteroidetes, Actinobacteria, Fusobacteria, Proteobacteria, Verrucomicrobia, and Cyanobacteria [90], with Firmicutes and Bacteroidetes constituting 90% of the total bacterial population [91]. Throughout life, the composition of the gut microbiota can change, influenced by age, sex, lifestyle factors such as physical exercise, sedentary behavior, smoking, diet, and the use of antibiotics, creating a unique microbial profile for each individual [8,92]. Gut microbiota plays a key role in maintaining the integrity of the mucosal barrier, regulating tissue development, generating short-chain fatty acids (SCFAs) essential for cellular energy balance, providing nutrients and vitamins, and modulating the host immune response, inflammatory cascade, and protection against pathogens [57,93,94]. The bacteria in the gastrointestinal tract can ferment non-digestible carbohydrates, generating SCFAs. The main SCFAs are acetate, propionate, and butyrate. SCFAs exhibit various beneficial effects, such as maintaining intestinal barrier integrity, regulating immune function, and promoting anti-inflammatory responses. SCFAs also display anti-tumor properties by inducing apoptosis, reducing the proliferation of neoplastic cells, and mitigating oxidative stress [95]. The effects of SCFAs on the human gut are mediated by the presence of SCFAs transporters on the colonic epithelium. These transporters include MCT1, MCT4, SMCT1, and SMCT2. MCT1 is located in both the apical and basolateral membranes of the colonic epithelium, MCT4 is found in the basolateral membrane, and SMCT1 and SMCT2 are located in the apical region [96].
Environmental factors, including diet, infections, and drugs, can disrupt the intestinal bacterial flora, which consequently may affect SCFAs and vitamin synthesis, increase stress responses, cause immune dysregulation, and increase susceptibility to DNA alterations [97]. Over time, these changes will lead to chronic inflammation, immune dysfunction, and metabolic alterations that can contribute to the development of conditions such as allergies, obesity, irritable bowel syndrome, IBD, and CRC. The resulting imbalance in the composition and function of the microbiota is referred to as dysbiosis, a condition linked to the development and progression of CRC [98].
Patients with CRC exhibit low microbial biodiversity, with specific bacteria, such as Fusobacterium nucleatum, Escherichia coli, Enterococcus faecalis, Streptococcus gallolyticus, and Bacteroides fragilis present in higher abundance in their gut. Additionally, Salmonella, Clostridium difficile, and Prevotella are also linked to CRC pathogenesis [99,100].
Fusobacterium nucleatum invades the intestinal epithelium through the expression of the adhesion protein FadA. This protein forms a complex with E-cadherin, increasing intestinal barrier permeability [101] and enhancing β-Catenin activity [102], which promotes cancer cell growth. Additionally, F. nucleatum suppresses immune cell activity by binding specific inhibitory receptors on natural killer (NK) cells and T cells [103]. An increase in tumor-promoting cytokines, including IL-17A and TNF-α, in CRC is associated with the activation of the NF-κB pathway by F. nucleatum [104].
A similar role is attributed to Bacteroides fragilis, which produces a metalloprotease that cleaves E-cadherin, disrupting the intestinal barrier [105]. It also induces the nuclear translocation of β-Catenin [106] and stimulates the production of IL-8 and IL-17, triggering an aberrant immune response and promoting tumorigenesis [107].
Activation of the β-Catenin pathway is also induced by Salmonella [108] and Clostridium difficile [109] infection, along with the activation of the STAT3 signaling pathway [110] and the production of reactive oxygen species (ROS) and IL-17, respectively [109].
The virulent strain of E. coli contributes to CRC carcinogenesis by releasing toxins that induce double-stranded DNA damage [111,112].
Enterococcus faecalis plays a primary role in CRC progression through the production of oxidants, which induce chromosomal instability [113]. The hydrogen peroxide produced by E. faecalis promotes cell proliferation by activating the EGFR signaling pathway [114].
Streptococcus gallolyticus drives CRC carcinogenesis by inducing inflammation through IL-1, COX-2, and IL-8 pathways [115]. Additionally, cells infected with S. gallolyticus exhibit increased levels of β-Catenin and proliferative proteins such as c-myc and PCNA [116].
The intestinal dysbiosis is also accompanied by a reduction in butyrate-producing bacteria, such as Eubacterium, Roseburia, and Faecalibacterium, which are essential for maintaining intestinal homeostasis and exerting anti-inflammatory and anti-tumorigenic effects, thus further promoting CRC progression [117] (Figure 2).
5. Probiotics and Their Metabolites for Prevention and as Adjuvant Therapeutics in Colorectal Cancer: Impact on Inflammation and Autophagy
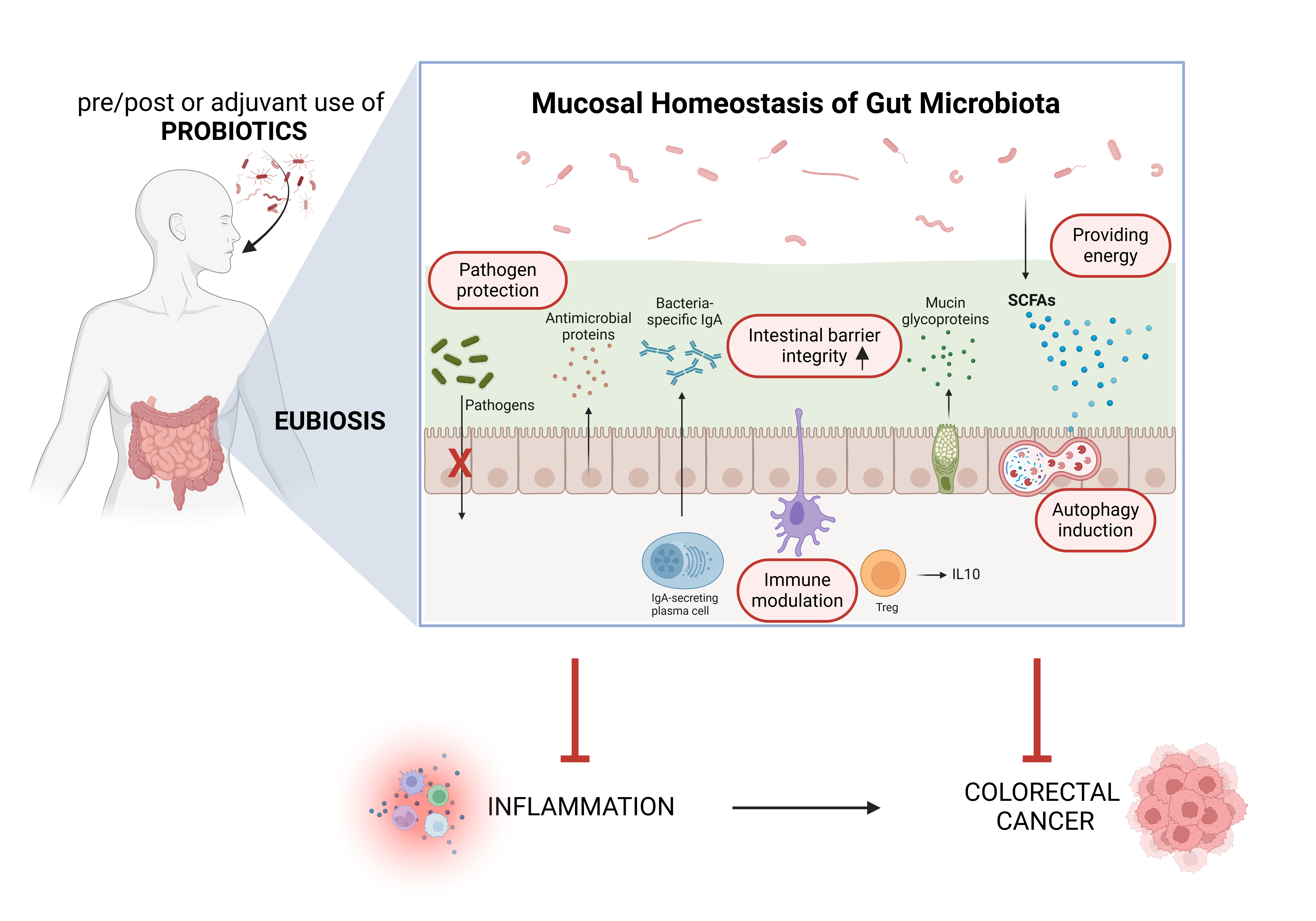
Dysbiosis plays a significant role in the development and progression of CRC. This has led to increasing interest in using probiotics as a therapeutic option, as they can modulate the gut microbiota, restore microbial balance, and potentially counteract dysbiosis, offering a promising strategy to influence CRC outcomes. Probiotics are living bacteria that elicit beneficial health effects by restoring eubiosis. Probiotics exhibit anti-cancer properties through various mechanisms, such as altering gut microbiota composition, reducing intestinal pH levels, neutralizing mutagens or carcinogens, enhancing immune responses while dampening inflammation, and promoting autophagy, apoptosis, and cell differentiation processes [118,119,120].
The release of bacterial products, such as flagellin and lipopolysaccharide (LPS), plays a significant role in activating the host immune system and contributes to the anti-inflammatory properties of probiotics. One of the ways probiotics combat inflammation is through the interaction of bacterial products with Toll-like receptors (TLRs) expressed on dendritic cells or macrophages. This interaction triggers the differentiation of naïve T cells into Treg cells, which produce IL-10 and reduce the production of TNF-α. Furthermore, probiotics inhibit the activation of NF-κB in macrophages, thereby reducing the production of pro-inflammatory cytokines such as TNF-α and IL-12. Additionally, probiotics promote the differentiation of B cells into IgA-producing plasma cells, enhancing mucus production and preventing the adhesion of microorganisms to intestinal epithelial cells, thereby reducing the risk of pathogen penetration [121,122,123].
In vitro experiments demonstrated the anti-proliferative and cytotoxic properties of probiotic bacteria administered alone or in strain combination on human CRC cells but not on normal colonic cells [124].
In animal models, the treatment with probiotics of the Lactobacillus genus decreased the formation of tumors, reduced the growth of aerobic bacteria in the intestine, and restored the correct host immune functions [125,126,127]. In CAC models, the administration of VSL#3, a probiotic mixture containing Bifidobacterium breve, B. infantis, B. longum, Lactobacillus acidophilus, L. bulgaricus, L. casei, L. plantarum, and Streptococcus thermophilus, reduced the incidence of high-grade dysplasia and prevented the development of CRC. The probiotic strains contrasted colorectal carcinogenesis in mouse models of Dextran Sulfate Sodium (DSS)- and Azoxymethane (AOM)-induced colitis, reducing the activation of STAT3, the expression of BCL-2 and IL-6 [128,129]. L. acidophilus reduced the formation of aberrant crypt foci, the earliest neoplastic lesions in the colon epithelium [130].
In CRC patients, probiotics were shown to synergize with chemotherapy by improving their therapeutic efficacy and reducing the side effects. The co-administration of L. acidophilus and L. casei improves the efficacy of the chemotherapeutic drug 5-FU increasing its pro-apoptotic efficiency [131]. L. paracasei and L. rhamnosus lead to a decreased production of MMP-9, which reflects in reduced invasion ability of CRC cell lines [132].
CRC patients co-treated with L. rhamnosus could be treated with lower doses of 5-FU and showed reduced episodes of diarrhea and abdominal pain, and required shorter hospitalization compared to patients treated with only the chemotherapeutics [133]. Lactobacillus bacteria administered before and after surgery reduced the proliferation, the growth, and the invasion of CRC cells, decreased enteropathogenic bacteria in blood and improved diarrhea, restored the integrity of gut mucosa, and stimulated the systemic immune system [134,135]. Post-operative complications were reduced in CRC patients treated with Bifidobacterium and Lactobacillus [136,137]. Patients treated with chemotherapy supplemented with Bifidobacterium breve demonstrated increased microbiota diversity, a lower frequency of fever, and a reduced risk of infections [136]. The administration of specific Lactobacillus and Bifidobacterium strains reduces radiotherapy-related gut toxicity and diarrhea [138,139].
L. casei, L. acidophilus, L. lactis, B. bifidum, B. longum, and B. infantis ameliorated the inflammatory reactions, reducing the levels of IL-6, and improved the quality of life for CRC patients [140]. B. longum, L. acidophilus, and Enterococcus faecalis decreased the population of Fusobacterium species in CRC patients [141]. As previously mentioned, probiotics play a role in reducing harmful gut bacteria by competing for essential resources such as nutrients and adhesion sites, while also producing antimicrobial compounds like bacteriocins, hydrogen peroxide, and lactic acid [142]. Probiotics have been shown to counteract inflammation, a key factor in CRC promotion, and modulate the immune response [57]. Additionally, probiotics stimulate goblet cells to increase the production of mucus and defensins, enhance tight junction integrity, and reduce intestinal permeability, thus preventing the infiltration of pathogenic bacteria [142]. These processes help reshape the gut microbiota, promote the growth of beneficial bacteria, and reduce the risk of CRC development.
Recently, we have shown that soluble metabolites of Lactiplantibacillus plantarum OC01 could reduce the cell proliferation and migration of CRC cells cultivated as 2D or 3D-spheroids even in the presence of the inflammatory cytokine IL-6 [143]. Additionally, the metabolites from Lactiplantibacillus plantarum OC01 changed the secretome of CRC cells, reducing the release of cytokines so that M2-like macrophages were reverted into an M1-like phenotype [144].
In vitro studies have shown that certain Lactobacillus strains possess anti-genotoxic and anti-mutagenic properties, effectively reducing mutagen levels and thereby lowering the risk of CRC initiation [145,146]. Furthermore, probiotics enhance the activity of antioxidant enzymes such as superoxide dismutase, catalase, and glutathione peroxidase, which counteract oxidative stress and provide additional protection against CRC development [121]. Lactobacillus casei DG’s metabolites were shown to reduce the expression of inflammatory cytokines IL-1α, IL-6, and IL-8 and of TLR4 while increasing the expression of anti-inflammatory cytokine IL-10 [147].
The anti-carcinogenic effects of probiotics are also attributed to the production of SCFAs [148]. A decrease in the concentration of SCFAs has been linked to an increased risk of CRC. Butyrate is the primary SCFA absorbed by the intestinal epithelium and serves as the main energy source for colonocytes, while the acetate and propionate mainly reach the liver [149]. Additionally, butyrate promotes the integrity of the gut barrier by upregulating the genes that encode for the tight junction proteins, such as claudin-1, zonula occludens-1, and occludin [150]. Furthermore, by enhancing the expression of Mucin 2, butyrate reinforces the mucus layer of the gut epithelium, which is essential for protection against pathogens and immune response regulation [151]. SCFAs, particularly butyrate, negatively regulate the inflammatory signaling pathway mediated by the inflammasome complex NLRP3, thereby inhibiting macrophage activation [152]. Butyrate also reduces the production of inflammatory cytokines, including TNF-α, MCP-1, and IL-6, through the activation of GPR41 in macrophages [153], and decreases the proliferation and the cytokine production of Th1, Th17, and Th22 cells [154]. In patients with CRC, however, there is a marked reduction in butyrate levels [155,156]. This reduction contributes to increased cell proliferation and decreased apoptosis, creating an environment that promotes cancer cell growth and tumor progression. Moreover, the lower levels of butyrate may impair the intestinal barrier, increase inflammation and promote dysbiosis, all of which are risk factors for CRC. The consumption of probiotics, such as Lactobacillus and Bifidobacterium, has been shown to increase the production of butyrate in the colon. Elevated butyrate concentrations help restore the balance between cell proliferation and apoptosis, promoting cancer cell death while inhibiting abnormal cell proliferation. Additionally, butyrate enhances the integrity of the intestinal barrier, reduces inflammation, and promotes the production of anti-inflammatory cytokines like IL-10. Furthermore, increased butyrate production can help reduce the formation of harmful secondary bile acids, contributing to a more favorable gut environment that inhibits CRC progression [122,142,157].
Butyrate treatment stimulated autophagy in CRC cells through the activation of the LKB1-AMPK pathway [158]. Dysregulation of autophagy may lead to impaired bacteria clearance and pathogens invasion and overall gut microbiota dysbiosis and exacerbated chronic inflammation that increases the risk of CRC [159]. Of relevance, in cancer cells there is a crucial interplay between the Wnt pathway and autophagy. Activation of the Wnt/β-Catenin-signaling pathway has been shown to inhibit autophagy while promoting CRC growth [160]. We have shown that butyrate influences the Wnt/β-Catenin pathway, a key player in the early stages of CRC development. By inducing autophagy as an alternative mechanism to the proteasomal degradation, which is ineffective in APC and β-Catenin mutated CRC, butyrate promotes the autophagic degradation of β-Catenin, and this results in reduced CRC cell proliferation and migration [161]. Probiotics could also affect TAMs polarization through modulation of autophagy [152] (Figure 3).
Conclusions
Autophagy plays a crucial role in maintaining cellular homeostasis through the turnover of damaged proteins and organelles [162,163]. Dysregulation of autophagy can influence the composition and function of the gut microbiota, contributing to CRC development [159,164]. This bidirectional relationship, in which autophagy impacts microbial balance and dysbiosis affects autophagic regulation, plays a significant role in the development of CRC. This relationship between genetic alterations, microbial dysbiosis, and autophagy represents the complex nature of CRC pathogenesis and could provide new therapeutic strategies targeting autophagy and microbiota to prevent or treat CRC.
The gut microbiota plays a crucial role in maintaining intestinal homeostasis through numerous functions, including the breakdown of non-digestible dietary components, the renewal of epithelial cells, the maintenance of intestinal mucosal integrity, immune system modulation, and the secretion of antimicrobial substances. It also protects against infections by competing with pathogenic organisms for nutrients and receptors, while promoting overall health through the synthesis of vitamins, regulation of fat reserves, and energy storage [94,165]. However, gut microbiota is highly sensitive to environmental factors such as dietary macro- and micronutrients, infections, and lifestyle, which can lead to alterations in its composition and function, a condition known as dysbiosis. Gut dysbiosis is often associated with intestinal inflammation, which may evolve to IBD and CRC [166]. Specifically, changes in the abundance of beneficial bacteria or an overgrowth of pathogenic organisms such as Bacteroides, Prevotella, Proteobacteria, Eubacterium, Fusobacterium, Proteobacteria, Escherichia coli, Clostriudium, and Salmonella spp. can disrupt the balance of the microbiota, creating an environment that promotes inflammation and tumorigenesis [167].
In addition to the influence of gut microbiota, genetic factors also play a significant role in the development of CRC. Around 80% of CRCs are associated with mutations in the Wnt/β-Catenin signaling pathway [168], which is involved in key processes such as adult tissue renewal and homeostasis, hematopoietic stem cell maintenance, and regulation of cell proliferation, migration, and differentiation [169]. Mutations in this pathway, notably in the APC and β-Catenin genes, lead to disruption of these regulatory mechanisms, resulting in uncontrolled tumor growth [27,170]. The APC mutation specifically initiates the formation of adenomas, or intestinal polyps, some of which progress to adenocarcinomas due to the accumulation of additional mutations, including those in KRAS, DCC, TP53, and SMAD4 [171]. These genetic mutations are observed in both hereditary and sporadic CRC cases [27] and interact with environmental factors, such as diet, lifestyle, and gut microbiota, thereby accelerating the development and progression of the disease [172].
The state of eubiosis, which refers to the balance between the host and its microbiota, is essential for proper gut function and overall health [173]. The most important metabolites released by the intestinal microbiota, which have a beneficial effect on human health, are SCFAs, including acetate, butyrate, and propionate. These metabolites are produced during the fermentation of non-digestible plant fibers in the diet [60]. Among them, butyrate is the most abundant and serves as the primary energy source for intestinal cells [149]. Butyrate has a wide range of beneficial effects on health. By interacting with its receptors, it modulates immune system activity, promotes the release of anti-inflammatory cytokines, and exhibits various anti-tumor properties, including inducing autophagy and apoptosis and reducing the proliferation of neoplastic cells. Additionally, butyrate plays a crucial role in reducing oxidative stress and reinforcing the intestinal barrier, preserving gut integrity [174,175].
Probiotics are live microorganisms that when supplemented in equilibrate formulas provide beneficial effects to the host. They are primarily lactic acid bacteria from genera such as Lactobacillus, Streptococcus, Enterococcus, Lactococcus, Leuconostoc, Bifidobacterium, and Saccharomyces [176]. The therapeutic and preventive effects of probiotics are largely due to their ability to maintain a state of eubiosis, ensuring intestinal health [177]. Probiotics help maintain gut health by protecting the intestinal mucosa, reducing the release of carcinogenic and oxidative molecules, and modulating immune responses, inflammation, autophagy, apoptosis, and cell differentiation. They also compete with harmful bacteria for nutrients and space, producing antimicrobial substances that limit the proliferation of pathogenic microbes, thereby preventing dysbiosis [178,179]. Moreover, probiotics produce SCFAs, which are important for maintaining gut barrier integrity and regulating inflammatory processes, further supporting gut health [119].
By modulating key signaling pathways such as Wnt/β-Catenin and reshaping the TME, microbial metabolites offer a promising avenue for therapeutic intervention in CRC. Collectively, these insights pave the way for future translational research and therapeutic development aimed at introducing microbiota-derived metabolites for improved CRC management, offering new hope for more personalized and targeted treatments. The implementation of probiotics in CRC therapy is not only related to their ability to reduce the risk of CRC development and progression through the mechanisms described above, but also to their potential to synergize with while decreasing the side effects of current therapies [177]. Moreover, the supplementation of probiotics pre- and/or post-surgery may improve prognosis, quality of life, and reduce the recurrence of CRC [135,180,181].
Taken together, in vitro and in vivo evidence demonstrated that probiotics could counteract the malignant phenotype of CRC cancer and support the employment of probiotic supplementation for CRC management. Bacterial strains improve the immune system and intestinal integrity, increase the antimicrobial defense, reduce the adverse effects related to chemotherapy treatment, the postoperative complications, as well as the symptoms associated with the gastrointestinal environment. In CRC patients, the adjuvant therapy with probiotics decreases the occurrence of septicemia, the incidence of postoperative infections and diarrhea, and the rate of postoperative antibiotic use. It is to be stressed that, despite the promising and encouraging results, not all probiotics showed positive health effects in CRC patients, emphasizing the need for further investigations to reveal the potential of probiotics for personalized CRC treatment.
Author Contributions
Conceptualization, B.G., L.V., A.F., and C.I.; formal analysis, A.V. and C.M.; data curation, A.V. and C.M.; writing—original draft preparation, B.G., L.V., and A.F.; writing—review and editing, C.I.; visualization, B.G., L.V., and A.F.; supervision, C.I.; project administration, A.A. and M.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article. This review article is part of the PhD thesis discussed by Dr. Beatrice Garavaglia on February 26, 2025. We mention this to anticipate that the overlapping text does not count as self-plagiarism.
Acknowledgments
B.G. was supported with PhD fellowship granted by Comoli, Ferrari & SpA (Novara, Italy). L.V. was supported with post-doctoral fellowship granted by Probiotical S.p.A (Novara, Italy). A.F. is recipient of a post-doctoral fellowship granted by Fondazione Veronesi (FUV 2025) (Milan, Italy). Thanks are due to Associazione per la Ricerca Medica “Ippocrate-Rhazi” (Novara, Italy) and to Consorzio Interuniversitario per le Biotecnologie (CIB, Trieste, Italy) for support.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: A Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA: A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Hossain, S.; Karuniawati, H.; Jairoun, A.A.; Urbi, Z.; Ooi, D.J.; John, A.; Lim, Y.C.; Kibria, K.M.K.; Mohiuddin, A.M.; Ming, L.C.; et al. Colorectal Cancer: A Review of Carcinogenesis, Global Epidemiology, Current Challenges, Risk Factors, Preventive and Treatment Strategies. Cancers 2022, 14, 1732. [Google Scholar] [CrossRef]
- Morgan, E.; Arnold, M.; Gini, A.; Lorenzoni, V.; Cabasag, C.J.; Laversanne, M.; Vignat, J.; Ferlay, J.; Murphy, N.; Bray, F. Global burden of colorectal cancer in 2020 and 2040: incidence and mortality estimates from GLOBOCAN. Gut 2023, 72, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Roshandel, G.; Ghasemi-Kebria, F.; Malekzadeh, R. Colorectal Cancer: Epidemiology, Risk Factors, and Prevention. Cancers 2024, 16, 1530. [Google Scholar] [CrossRef]
- Tsuruya, A.; Kuwahara, A.; Saito, Y.; Yamaguchi, H.; Tsubo, T.; Suga, S.; Inai, M.; Aoki, Y.; Takahashi, S.; Tsutsumi, E.; et al. Ecophysiological consequences of alcoholism on human gut microbiota: implications for ethanol-related pathogenesis of colon cancer. Sci. Rep. 2016, 6, 27923. [Google Scholar] [CrossRef]
- Munteanu, C.; Schwartz, B. Interactions between Dietary Antioxidants, Dietary Fiber and the Gut Microbiome: Their Putative Role in Inflammation and Cancer. Int. J. Mol. Sci. 2024, 25, 8250. [Google Scholar] [CrossRef]
- Ionescu, V.A.; Gheorghe, G.; Georgescu, T.F.; Buica, V.; Catanescu, M.-S.; Cercel, I.-A.; Budeanu, B.; Budan, M.; Bacalbasa, N.; Diaconu, C. Exploring the Role of the Gut Microbiota in Colorectal Cancer Development. Gastrointest. Disord. 2024, 6, 526–537. [Google Scholar] [CrossRef]
- Valle, L.; Vilar, E.; Tavtigian, S.V.; Stoffel, E.M. Genetic predisposition to colorectal cancer: syndromes, genes, classification of genetic variants and implications for precision medicine. J. Pathol. 2018, 247, 574–588. [Google Scholar] [CrossRef]
- Fantini, M.C.; Guadagni, I. From inflammation to colitis-associated colorectal cancer in inflammatory bowel disease: Pathogenesis and impact of current therapies. Dig. Liver Dis. 2021, 53, 558–565. [Google Scholar] [CrossRef]
- Fadlallah, H.; El Masri, J.; Fakhereddine, H.; Youssef, J.; Chemaly, C.; Doughan, S.; Abou-Kheir, W. Colorectal cancer: Recent advances in management and treatment. World J. Clin. Oncol. 2024, 15, 1136–1156. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Gautam, V.; Sandhu, A.; Rawat, K.; Sharma, A.; Saha, L. Current and emerging therapeutic approaches for colorectal cancer: A comprehensive review. World J. Gastrointest. Surg. 2023, 15, 495–519. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.-H.; Chen, Y.-X.; Fang, J.-Y. Comprehensive review of targeted therapy for colorectal cancer. Signal Transduct. Target. Ther. 2020, 5, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Moreira, M.M.; Carriço, M.; Capelas, M.L.; Pimenta, N.; Santos, T.; Ganhão-Arranhado, S.; Mäkitie, A.; Ravasco, P.; M, A. The impact of pre-, pro- and synbiotics supplementation in colorectal cancer treatment: a systematic review. Front. Oncol. 2024, 14, 1395966. [Google Scholar] [CrossRef]
- Fearon, E.R.; Vogelstein, B. A genetic model for colorectal tumorigenesis. Cell 1990, 61, 759–767. [Google Scholar] [CrossRef]
- Nguyen, L.H.; Goel, A.; Chung, D.C. Pathways of Colorectal Carcinogenesis. Gastroenterology 2020, 158, 291–302. [Google Scholar] [CrossRef]
- Cao, Q.; Tian, Y.; Deng, Z.; Yang, F.; Chen, E. Epigenetic Alteration in Colorectal Cancer: Potential Diagnostic and Prognostic Implications. Int. J. Mol. Sci. 2024, 25, 3358. [Google Scholar] [CrossRef]
- Mármol, I.; Sánchez-De-Diego, C.; Pradilla Dieste, A.; Cerrada, E.; Rodriguez Yoldi, M. Colorectal Carcinoma: A General Overview and Future Perspectives in Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 197. [Google Scholar] [CrossRef]
- Jansen, M.; Menko, F.H.; Brosens, L.A.; Giardiello, F.M.; Offerhaus, G.J. Establishing a clinical and molecular diagnosis for hereditary colorectal cancer syndromes: Present tense, future perfect? Gastrointest. Endosc. 2014, 80, 1145–1155. [Google Scholar] [CrossRef]
- Trimbath, J.D.; Giardiello, F.M. Genetic testing and counselling for hereditary colorectal cancer. Aliment. Pharmacol. Ther. 2002, 16, 1843–1857. [Google Scholar] [CrossRef]
- Edelstein, D.L.; Axilbund, J.; Baxter, M.; Hylind, L.M.; Romans, K.; Griffin, C.A.; Cruz–Correa, M.; Giardiello, F.M. Rapid Development of Colorectal Neoplasia in Patients With Lynch Syndrome. Clin. Gastroenterol. Hepatol. 2010, 9, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Bonk, T.; Humeny, A.; Sutter, C.; Gebert, J.; Doeberitz, M.v.K.; Becker, C.-M. Molecular diagnosis of familial adenomatous polyposis (FAP): genotyping of adenomatous polyposis coli (APC) alleles by MALDI-TOF mass spectrometry. Clin. Biochem. 2002, 35, 87–92. [Google Scholar] [CrossRef]
- Geiersbach, K.B.; Samowitz, W.S. Microsatellite Instability and Colorectal Cancer. Arch. Pathol. Lab. Med. 2011, 135, 1269–1277. [Google Scholar] [CrossRef]
- Magzoub, M.M.; Prunello, M.; Brennan, K.; Gevaert, O. The impact of DNA methylation on the cancer proteome. PLOS Comput. Biol. 2019, 15, e1007245. [Google Scholar] [CrossRef] [PubMed]
- Colussi, D.; Brandi, G.; Bazzoli, F.; Ricciardiello, L. Molecular Pathways Involved in Colorectal Cancer: Implications for Disease Behavior and Prevention. Int. J. Mol. Sci. 2013, 14, 16365–16385. [Google Scholar] [CrossRef]
- Lecarpentier, Y.; Schussler, O.; Hébert, J.-L.; Vallée, A. Multiple Targets of the Canonical WNT/β-Catenin Signaling in Cancers. Front. Oncol. 2019, 9, 1248. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Gan, W.-J. Wnt/β-Catenin Signaling Pathway in the Development and Progression of Colorectal Cancer. Cancer Manag. Res. 2023, ume 15, 435–448. [Google Scholar] [CrossRef]
- Farahmand, L.; Darvishi, B.; Majidzadeh-A, K.; Ansari, A.M. Naturally occurring compounds acting as potent anti-metastatic agents and their suppressing effects on Hedgehog and WNT/β-catenin signalling pathways. Cell Prolif. 2016, 50. [Google Scholar] [CrossRef]
- Pate, K.T.; Stringari, C.; Sprowl-Tanio, S.; Wang, K.; TeSlaa, T.; Hoverter, N.P.; McQuade, M.M.; Garner, C.; A Digman, M.; A Teitell, M.; et al. Wnt signaling directs a metabolic program of glycolysis and angiogenesis in colon cancer. EMBO J. 2014, 33, 1454–1473. [Google Scholar] [CrossRef]
- Sprowl-Tanio, S.; Habowski, A.N.; Pate, K.T.; McQuade, M.M.; Wang, K.; Edwards, R.A.; Grun, F.; Lyou, Y.; Waterman, M.L. Lactate/pyruvate transporter MCT-1 is a direct Wnt target that confers sensitivity to 3-bromopyruvate in colon cancer. Cancer Metab. 2016, 4, 1–18. [Google Scholar] [CrossRef]
- Yaeger, R.; Chatila, W.K.; Lipsyc, M.D.; Hechtman, J.F.; Cercek, A.; Sanchez-Vega, F.; Jayakumaran, G.; Middha, S.; Zehir, A.; Donoghue, M.T.A.; et al. Clinical Sequencing Defines the Genomic Landscape of Metastatic Colorectal Cancer. Cancer Cell 2018, 33, 125–136.e3. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Shay, J.W. Multiple Roles of APC and its Therapeutic Implications in Colorectal Cancer. J. Natl. Cancer Inst. 2017, 109, djw332. [Google Scholar] [CrossRef] [PubMed]
- Aghabozorgi, A.S.; Bahreyni, A.; Soleimani, A.; Bahrami, A.; Khazaei, M.; Ferns, G.A.; Avan, A.; Hassanian, S.M. Role of adenomatous polyposis coli (APC) gene mutations in the pathogenesis of colorectal cancer; current status and perspectives. Biochimie 2019, 157, 64–71. [Google Scholar] [CrossRef]
- Heinen, C.D.; Goss, K.H.; Cornelius, J.R.; Babcock, G.F.; Knudsen, E.S.; Kowalik, T.; Groden, J. The APC tumor suppressor controls entry into S-phase through its ability to regulate the cyclin D/RB pathway. Gastroenterology 2002, 123, 751–763. [Google Scholar] [CrossRef]
- Zhu, G.; Pei, L.; Xia, H.; Tang, Q.; Bi, F. Role of oncogenic KRAS in the prognosis, diagnosis and treatment of colorectal cancer. Mol. Cancer 2021, 20, 143. [Google Scholar] [CrossRef]
- Liao, X.; Morikawa, T.; Lochhead, P.; Imamura, Y.; Kuchiba, A.; Yamauchi, M.; Nosho, K.; Qian, Z.R.; Nishihara, R.; Meyerhardt, J.A.; et al. Prognostic Role of PIK3CA Mutation in Colorectal Cancer: Cohort Study and Literature Review. Clin. Cancer Res. 2012, 18, 2257–2268. [Google Scholar] [CrossRef]
- Nosho, K.; Kawasaki, T.; Longtine, J.A.; Fuchs, C.S.; Ohnishi, M.; Suemoto, Y.; Kirkner, G.J.; Zepf, D.; Yan, L.; Ogino, S. PIK3CA Mutation in Colorectal Cancer: Relationship with Genetic and Epigenetic Alterations. Neoplasia 2008, 10, 534–541. [Google Scholar] [CrossRef]
- Smith, G.; Carey, F.A.; Beattie, J.; Wilkie, M.J.V.; Lightfoot, T.J.; Coxhead, J.; Garner, R.C.; Steele, R.J.; Wolf, C.R. Mutations in APC, Kirsten-ras, and p53--alternative genetic pathways to colorectal cancer. Proc. Natl. Acad. Sci. USA 2002, 99, 9433–9438. [Google Scholar] [CrossRef]
- Russo, A.; Bazan, V.; Iacopetta, B.; Kerr, D.; Soussi, T.; Gebbia, N. The TP53 Colorectal Cancer International Collaborative Study on the Prognostic and Predictive Significance of p53 Mutation: Influence of Tumor Site, Type of Mutation, and Adjuvant Treatment. J. Clin. Oncol. 2005, 23, 7518–7528. [Google Scholar] [CrossRef]
- Yu, G.; Klionsky, D.J. Life and Death Decisions—The Many Faces of Autophagy in Cell Survival and Cell Death. Biomolecules 2022, 12, 866. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Abdel-Aziz, A.K.; Abdelfatah, S.; Abdellatif, M.; Abdoli, A.; Abel, S.; Abeliovich, H.; Abildgaard, M.H.; Abudu, Y.P.; Acevedo-Arozena, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (4th edition). Autophagy 2021, 17, 1–382. [Google Scholar] [CrossRef]
- Vidoni, C.; Ferraresi, A.; Secomandi, E.; Vallino, L.; Dhanasekaran, D.N.; Isidoro, C. Epigenetic targeting of autophagy for cancer prevention and treatment by natural compounds. Semin. Cancer Biol. 2020, 66, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Vidoni, C.; Vallino, L.; Ferraresi, A.; Secomandi, E.; Salwa, A.; Chinthakindi, M.; Galetto, A.; Dhanasekaran, D.N.; Isidoro, C. Epigenetic control of autophagy in women’s tumors: role of non-coding RNAs. J. Cancer Metastasis Treat. 2021, 2021. [Google Scholar] [CrossRef]
- Thuwajit, C.; Ferraresi, A.; Titone, R.; Thuwajit, P.; Isidoro, C. The metabolic cross-talk between epithelial cancer cells and stromal fibroblasts in ovarian cancer progression: Autophagy plays a role. Med. Res. Rev. 2017, 38, 1235–1254. [Google Scholar] [CrossRef] [PubMed]
- Ferraresi, A.; Girone, C.; Esposito, A.; Vidoni, C.; Vallino, L.; Secomandi, E.; Dhanasekaran, D.N.; Isidoro, C. How Autophagy Shapes the Tumor Microenvironment in Ovarian Cancer. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef]
- Ferraresi, A.; Girone, C.; Maheshwari, C.; Vallino, L.; Dhanasekaran, D.N.; Isidoro, C. Ovarian Cancer Cell-Conditioning Medium Induces Cancer-Associated Fibroblast Phenoconversion through Glucose-Dependent Inhibition of Autophagy. Int. J. Mol. Sci. 2024, 25, 5691. [Google Scholar] [CrossRef]
- Shao, L.-N.; Zhu, B.-S.; Xing, C.-G.; Yang, X.-D.; Young, W.; Cao, J.-P. Effects of autophagy regulation of tumor-associated macrophages on radiosensitivity of colorectal cancer cells. Mol. Med. Rep. 2016, 13, 2661–2670. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, H.; Lv, L.; Lu, K.; Li, H.; Zhang, W.; Cui, T. Autophagy: Dual roles and perspective for clinical treatment of colorectal cancer. Biochimie 2022, 206, 49–60. [Google Scholar] [CrossRef]
- Manzoor, S.; Muhammad, J.S.; Maghazachi, A.A.; Hamid, Q. Autophagy: A Versatile Player in the Progression of Colorectal Cancer and Drug Resistance. Front. Oncol. 2022, 12, 924290. [Google Scholar] [CrossRef]
- Tian, X.; Han, Z.; Zhu, Q.; Tan, J.; Liu, W.; Wang, Y.; Chen, W.; Zou, Y.; Cai, Y.; Huang, S.; et al. Silencing of cadherin-17 enhances apoptosis and inhibits autophagy in colorectal cancer cells. Biomed. Pharmacother. 2018, 108, 331–337. [Google Scholar] [CrossRef]
- Qureshi-Baig, K.; Kuhn, D.; Viry, E.; Pozdeev, V.I.; Schmitz, M.; Rodriguez, F.; Ullmann, P.; Koncina, E.; Nurmik, M.; Frasquilho, S.; et al. Hypoxia-induced autophagy drives colorectal cancer initiation and progression by activating the PRKC/PKC-EZR (ezrin) pathway. Autophagy 2019, 16, 1436–1452. [Google Scholar] [CrossRef] [PubMed]
- Trincheri, N.F.; Follo, C.; Nicotra, G.; Peracchio, C.; Castino, R.; Isidoro, C. Resveratrol-induced apoptosis depends on the lipid kinase activity of Vps34 and on the formation of autophagolysosomes. Carcinog. 2007, 29, 381–389. [Google Scholar] [CrossRef]
- Che, N.; Yang, Z.; Liu, X.; Li, M.; Feng, Y.; Zhang, C.; Li, C.; Cui, Y.; Xuan, Y. Suppression of LETM1 inhibits the proliferation and stemness of colorectal cancer cells through reactive oxygen species–induced autophagy. J. Cell. Mol. Med. 2020, 25, 2110–2120. [Google Scholar] [CrossRef]
- Wang, H.; Wang, Y.; Qian, L.; Wang, X.; Dong, X.; Huang, S.; Jin, M.; Ge, H.; Xu, C.; Zhang, Y. RNF216 contributes to proliferation and migration of colorectal cancer via suppressing BECN1-dependent autophagy. Oncotarget 2016, 7, 51174–51183. [Google Scholar] [CrossRef]
- Shen, T.; Cai, L.-D.; Liu, Y.-H.; Li, S.; Gan, W.-J.; Li, X.-M.; Wang, J.-R.; Guo, P.-D.; Zhou, Q.; Lu, X.-X.; et al. Ube2v1-mediated ubiquitination and degradation of Sirt1 promotes metastasis of colorectal cancer by epigenetically suppressing autophagy. J. Hematol. Oncol. 2018, 11, 1–16. [Google Scholar] [CrossRef]
- Li, B.-X.; Li, C.-Y.; Peng, R.-Q.; Wu, X.-J.; Wang, H.-Y.; Wan, D.-S.; Zhu, X.-F.; Zhang, X.-S. The expression ofbeclin 1is associated with favorable prognosis in stage IIIB colon cancers. Autophagy 2009, 5, 303–306. [Google Scholar] [CrossRef]
- Garavaglia, B.; Vallino, L.; Amoruso, A.; Pane, M.; Ferraresi, A.; Isidoro, C. The role of gut microbiota, immune system, and autophagy in the pathogenesis of inflammatory bowel disease: Molecular mechanisms and therapeutic approaches. Asp. Mol. Med. 2024, 4. [Google Scholar] [CrossRef]
- Hooper, K.M.; Barlow, P.G.; Henderson, P.; Stevens, C. Interactions Between Autophagy and the Unfolded Protein Response: Implications for Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2018, 25, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Iida, T.; Onodera, K.; Nakase, H. Role of autophagy in the pathogenesis of inflammatory bowel disease. World J. Gastroenterol. 2017, 23, 1944–1953. [Google Scholar] [CrossRef]
- Liu, X.; Yin, L.; Shen, S.; Hou, Y. Inflammation and cancer: paradoxical roles in tumorigenesis and implications in immunotherapies. Genes Dis. 2023, 10, 151–164. [Google Scholar] [CrossRef]
- Borowczak, J.; Szczerbowski, K.; Maniewski, M.; Kowalewski, A.; Janiczek-Polewska, M.; Szylberg, A.; Marszałek, A.; Szylberg, Ł. The Role of Inflammatory Cytokines in the Pathogenesis of Colorectal Carcinoma—Recent Findings and Review. Biomedicines 2022, 10, 1670. [Google Scholar] [CrossRef] [PubMed]
- Muthusami, S.; Ramachandran, I.K.; Babu, K.N.; Krishnamoorthy, S.; Guruswamy, A.; Queimado, L.; Chaudhuri, G. Role of Inflammation in the Development of Colorectal Cancer. Endocrine, Metab. Immune Disord. - Drug Targets 2021, 21, 77–90. [Google Scholar] [CrossRef]
- Wang, D.; Zhu, L.; Liu, H.; Feng, X.; Zhang, C.; Liu, B.; Li, T.; Liu, L.; Chang, H.; Sun, J.; et al. Altered gut metabolites and metabolic reprogramming involved in the pathogenesis of colitis-associated colorectal cancer and the transition of colon "inflammation to cancer". J. Pharm. Biomed. Anal. 2024, 253, 116553. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Z.; Li, Y.-Y. Inflammatory bowel disease: Pathogenesis. World J. Gastroenterol. 2014, 20, 91–99. [Google Scholar] [CrossRef]
- Shah, S.C.; Itzkowitz, S.H. Colorectal Cancer in Inflammatory Bowel Disease: Mechanisms and Management. Gastroenterology 2021, 162, 715–730.e3. [Google Scholar] [CrossRef]
- Sato, Y.; Tsujinaka, S.; Miura, T.; Kitamura, Y.; Suzuki, H.; Shibata, C. Inflammatory Bowel Disease and Colorectal Cancer: Epidemiology, Etiology, Surveillance, and Management. Cancers 2023, 15, 4154. [Google Scholar] [CrossRef]
- Fanizza, J.; Bencardino, S.; Allocca, M.; Furfaro, F.; Zilli, A.; Parigi, T.L.; Fiorino, G.; Peyrin-Biroulet, L.; Danese, S.; D’amico, F. Inflammatory Bowel Disease and Colorectal Cancer. Cancers 2024, 16, 2943. [Google Scholar] [CrossRef]
- Heo, G.; Lee, Y.; Im, E. Interplay between the Gut Microbiota and Inflammatory Mediators in the Development of Colorectal Cancer. Cancers 2021, 13, 734. [Google Scholar] [CrossRef]
- Turano, M.; Cammarota, F.; Duraturo, F.; Izzo, P.; De Rosa, M. A Potential Role of IL-6/IL-6R in the Development and Management of Colon Cancer. Membranes 2021, 11, 312. [Google Scholar] [CrossRef]
- West, N.R.; McCuaig, S.; Franchini, F.; Powrie, F. Emerging cytokine networks in colorectal cancer. Nat. Rev. Immunol. 2015, 15, 615–629. [Google Scholar] [CrossRef]
- Nagasaki, T.; Hara, M.; Nakanishi, H.; Takahashi, H.; Sato, M.; Takeyama, H. Interleukin-6 released by colon cancer-associated fibroblasts is critical for tumour angiogenesis: anti-interleukin-6 receptor antibody suppressed angiogenesis and inhibited tumour–stroma interaction. Br. J. Cancer 2014, 110, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T. IL-6 in inflammation, autoimmunity and cancer. Int. Immunol. 2020, 33, 127–148. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Shang, Y.; Sun, F.; Dong, X.; Niu, J.; Li, F. Interleukin-6 Promotes Epithelial-Mesenchymal Transition and Cell Invasion through Integrin β6 Upregulation in Colorectal Cancer. Oxidative Med. Cell. Longev. 2020, 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Zou, X.; Xia, W.; Gao, H.; Li, Z.; Liu, N.; Xu, Z.; Gao, C.; He, Z.; Niu, W.; et al. Integrin αvβ6 plays a bi-directional regulation role between colon cancer cells and cancer-associated fibroblasts. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, L.; Lai, W.; Zeng, Y.; Xu, H.; Lan, Q.; Su, P.; Chu, Z. Interaction with tumor-associated macrophages promotes PRL-3-induced invasion of colorectal cancer cells via MAPK pathway-induced EMT and NF-κB signaling-induced angiogenesis. Oncol. Rep. 2019, 41, 2790–2802. [Google Scholar] [CrossRef]
- Zhang, D.; Bi, J.; Liang, Q.; Wang, S.; Zhang, L.; Han, F.; Li, S.; Qiu, B.; Fan, X.; Chen, W.; et al. VCAM1 Promotes Tumor Cell Invasion and Metastasis by Inducing EMT and Transendothelial Migration in Colorectal Cancer. Front. Oncol. 2020, 10, 1066. [Google Scholar] [CrossRef]
- Knüpfer, H.; Preiss, R. Serum interleukin-6 levels in colorectal cancer patients—a summary of published results. Int. J. Color. Dis. 2009, 25, 135–140. [Google Scholar] [CrossRef]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS–) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef]
- Wang, S.; Liu, G.; Li, Y.; Pan, Y. Metabolic Reprogramming Induces Macrophage Polarization in the Tumor Microenvironment. Front. Immunol. 2022, 13, 840029. [Google Scholar] [CrossRef]
- Sun, X.; Li, Y.; Deng, Q.; Hu, Y.; Dong, J.; Wang, W.; Wang, Y.; Li, C. Macrophage Polarization, Metabolic Reprogramming, and Inflammatory Effects in Ischemic Heart Disease. Front. Immunol. 2022, 13, 934040. [Google Scholar] [CrossRef]
- Keller, D.S.; Windsor, A.; Cohen, R.; Chand, M. Colorectal cancer in inflammatory bowel disease: review of the evidence. Tech. Coloproctol. 2019, 23, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Veettil, S.K.; Wong, T.Y.; Loo, Y.S.; Playdon, M.C.; Lai, N.M.; Giovannucci, E.L.; Chaiyakunapruk, N. Role of Diet in Colorectal Cancer Incidence. JAMA Netw. Open 2021, 4, e2037341–e2037341. [Google Scholar] [CrossRef]
- Lewandowska, A.; Rudzki, G.; Lewandowski, T.; Stryjkowska-Góra, A.; Rudzki, S. Risk Factors for the Diagnosis of Colorectal Cancer. Cancer Control. 2022, 29. [Google Scholar] [CrossRef] [PubMed]
- Vernia, F.; Longo, S.; Stefanelli, G.; Viscido, A.; Latella, G. Dietary Factors Modulating Colorectal Carcinogenesis. Nutrients 2021, 13, 143. [Google Scholar] [CrossRef]
- Rubio, K.; Hernández-Cruz, E.Y.; Rogel-Ayala, D.G.; Sarvari, P.; Isidoro, C.; Barreto, G.; Pedraza-Chaverri, J. Nutriepigenomics in Environmental-Associated Oxidative Stress. Antioxidants 2023, 12, 771. [Google Scholar] [CrossRef]
- Conti, L. , Corn, M.D., Scazzocchio, B., Var, R., D’Archivio, M., et al. (2019). Dietary fatty acids and adipose tissue inflammation at the crossroad between obesity and colorectal cancer. Journal of Cancer Metastasis and Treatment.
- Baena, R.; Salinas, P. Diet and colorectal cancer. Maturitas 2015, 80, 258–264. [Google Scholar] [CrossRef]
- O'Keefe, S.J.D. Diet, microorganisms and their metabolites, and colon cancer. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.H.; Yu, J. Gut microbiota in colorectal cancer: mechanisms of action and clinical applications. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 690–704. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-Bacterial Mutualism in the Human Intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Al Bander, Z.; Nitert, M.D.; Mousa, A.; Naderpoor, N. The Gut Microbiota and Inflammation: An Overview. Int. J. Environ. Res. Public Heal. 2020, 17, 7618. [Google Scholar] [CrossRef] [PubMed]
- Natividad, J.M.M.; Verdu, E.F. Modulation of intestinal barrier by intestinal microbiota: Pathological and therapeutic implications. Pharmacol. Res. 2013, 69, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Liu, P.; Wang, Y.; Yang, G.; Zhang, Q.; Meng, L.; Xin, Y.; Jiang, X. The role of short-chain fatty acids in intestinal barrier function, inflammation, oxidative stress, and colonic carcinogenesis. Pharmacol. Res. 2021, 165, 105420. [Google Scholar] [CrossRef]
- Sivaprakasam, S.; Bhutia, Y.D.; Yang, S.; Ganapathy, V. Short-Chain Fatty Acid Transporters: Role in Colonic Homeostasis. In Comprehensive Physiology; Wiley: New York, NY, USA, 2017; Volume 8, pp. 299–314. [Google Scholar] [CrossRef]
- Lan, Y.; Kriete, A.; Rosen, G.L. Selecting age-related functional characteristics in the human gut microbiome. Microbiome 2013, 1, 2–2. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.-X.; Chen, X.-Y.; Wang, J.-Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in health and diseases. Signal Transduct. Target. Ther. 2022, 7, 1–28. [Google Scholar] [CrossRef]
- Sobhani, I.; Tap, J.; Roudot-Thoraval, F.; Roperch, J.P.; Letulle, S.; Langella, P.; Corthier, G.; Van Nhieu, J.T.; Furet, J.P. Microbial Dysbiosis in Colorectal Cancer (CRC) Patients. PLOS ONE 2011, 6, e16393. [Google Scholar] [CrossRef] [PubMed]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012, 22, 299–306. [Google Scholar] [CrossRef]
- Fardini, Y.; Wang, X.; Témoin, S.; Nithianantham, S.; Lee, D.; Shoham, M.; Han, Y.W. Fusobacterium nucleatumadhesin FadA binds vascular endothelial cadherin and alters endothelial integrity. Mol. Microbiol. 2011, 82, 1468–1480. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum Promotes Colorectal Carcinogenesis by Modulating E-Cadherin/β-Catenin Signaling via its FadA Adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef]
- Gur, C.; Ibrahim, Y.; Isaacson, B.; Yamin, R.; Abed, J.; Gamliel, M.; Enk, J.; Bar-On, Y.; Stanietsky-Kaynan, N.; Coppenhagen-Glazer, S.; et al. Binding of the Fap2 Protein of Fusobacterium nucleatum to Human Inhibitory Receptor TIGIT Protects Tumors from Immune Cell Attack. Immunity 2015, 42, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Wang, R.; Bhattacharya, R.; Boulbes, D.R.; Fan, F.; Xia, L.; Adoni, H.; Ajami, N.J.; Wong, M.C.; Smith, D.P.; et al. Fusobacterium Nucleatum Subspecies Animalis Influences Proinflammatory Cytokine Expression and Monocyte Activation in Human Colorectal Tumors. Cancer Prev. Res. 2017, 10, 398–409. [Google Scholar] [CrossRef]
- Wu, S.; Lim, K.-C.; Huang, J.; Saidi, R.F.; Sears, C.L. Bacteroides fragilis enterotoxin cleaves the zonula adherens protein, E-cadherin. Proc. Natl. Acad. Sci. 1998, 95, 14979–14984. [Google Scholar] [CrossRef]
- Wu, S.; Morin, P.J.; Maouyo, D.; Sears, C.L. Bacteroides fragilis enterotoxin induces c-Myc expression and cellular proliferation. Gastroenterology 2003, 124, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Chung, L.; Orberg, E.T.; Geis, A.L.; Chan, J.L.; Fu, K.; Shields, C.E.D.; Dejea, C.M.; Fathi, P.; Chen, J.; Finard, B.B.; et al. Bacteroides fragilis Toxin Coordinates a Pro-carcinogenic Inflammatory Cascade via Targeting of Colonic Epithelial Cells. Cell Host Microbe 2018, 23, 203–214.e5. [Google Scholar] [CrossRef]
- Lu, R.; Wu, S.; Zhang, Y.-G.; Xia, Y.; Liu, X.; Zheng, Y.; Chen, H.; Schaefer, K.L.; Zhou, Z.; Bissonnette, M.; et al. Enteric bacterial protein AvrA promotes colonic tumorigenesis and activates colonic beta-catenin signaling pathway. Oncogenesis 2014, 3, e105–e105. [Google Scholar] [CrossRef]
- Drewes, J.L.; Chen, J.; Markham, N.O.; Knippel, R.J.; Domingue, J.C.; Tam, A.J.; Chan, J.L.; Kim, L.; McMann, M.; Stevens, C.; et al. Human Colon Cancer–Derived Clostridioides difficile Strains Drive Colonic Tumorigenesis in Mice. Cancer Discov. 2022, 12, 1873–1885. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Wu, S.; Zhang, Y.-G.; Xia, Y.; Zhou, Z.; Kato, I.; Dong, H.; Bissonnette, M.; Sun, J. Salmonella Protein AvrA Activates the STAT3 Signaling Pathway in Colon Cancer. Neoplasia 2016, 18, 307–316. [Google Scholar] [CrossRef]
- Nowrouzian, F.L.; Oswald, E. Escherichia coli strains with the capacity for long-term persistence in the bowel microbiota carry the potentially genotoxic pks island. Microb. Pathog. 2012, 53, 180–182. [Google Scholar] [CrossRef]
- Dziubańska-Kusibab, P.J.; Berger, H.; Battistini, F.; Bouwman, B.A.M.; Iftekhar, A.; Katainen, R.; Cajuso, T.; Crosetto, N.; Orozco, M.; Aaltonen, L.A.; et al. Colibactin DNA-damage signature indicates mutational impact in colorectal cancer. Nat. Med. 2020, 26, 1063–1069. [Google Scholar] [CrossRef]
- Wang, X.; Allen, T.D.; May, R.J.; Lightfoot, S.; Houchen, C.W.; Huycke, M.M. Enterococcus faecalis Induces Aneuploidy and Tetraploidy in Colonic Epithelial Cells through a Bystander Effect. Cancer Res. 2008, 68, 9909–9917. [Google Scholar] [CrossRef] [PubMed]
- Boonanantanasarn, K.; Gill, A.L.; Yap, Y.; Jayaprakash, V.; Sullivan, M.A.; Gill, S.R. Enterococcus faecalis Enhances Cell Proliferation through Hydrogen Peroxide-Mediated Epidermal Growth Factor Receptor Activation. Infect. Immun. 2012, 80, 3545–3558. [Google Scholar] [CrossRef] [PubMed]
- Abdulamir, A.S.; Hafidh, R.R.; Bakar, F. Molecular detection, quantification, and isolation of Streptococcus gallolyticus bacteria colonizing colorectal tumors: inflammation-driven potential of carcinogenesis via IL-1, COX-2, and IL-8. Mol. Cancer 2010, 9, 249–249. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Herold, J.L.; Schady, D.; Davis, J.; Kopetz, S.; Martinez-Moczygemba, M.; Murray, B.E.; Han, F.; Li, Y.; Callaway, E.; et al. Streptococcus gallolyticus subsp. gallolyticus promotes colorectal tumor development. PLOS Pathog. 2017, 13, e1006440. [Google Scholar] [CrossRef]
- Balamurugan, R.; Rajendiran, E.; George, S.; Samuel, G.V.; Ramakrishna, B.S. Real-time polymerase chain reaction quantification of specific butyrate-producing bacteria, Desulfovibrio and Enterococcus faecalis in the feces of patients with colorectal cancer. J. Gastroenterol. Hepatol. 2008, 23, 1298–1303. [Google Scholar] [CrossRef]
- Nemati, M.; Omrani, G.R.; Ebrahimi, B.; Montazeri-Najafabady, N. The Beneficial Effects of Probiotics via Autophagy: A Systematic Review. BioMed Res. Int. 2021, 2021. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of Action of Probiotics. Adv. Nutr. Int. Rev. J. 2019, 10, S49–S66. [Google Scholar] [CrossRef]
- Lu, K.; Dong, S.; Wu, X.; Jin, R.; Chen, H. Probiotics in Cancer. Front. Oncol. 2021, 11. [Google Scholar] [CrossRef]
- Kahouli, I.; Tomaro-Duchesneau, C.; Prakash, S. Probiotics in colorectal cancer (CRC) with emphasis on mechanisms of action and current perspectives. J. Med Microbiol. 2013, 62, 1107–1123. [Google Scholar] [CrossRef]
- Molska, M.; Reguła, J. Potential Mechanisms of Probiotics Action in the Prevention and Treatment of Colorectal Cancer. Nutrients 2019, 11, 2453. [Google Scholar] [CrossRef]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: A Door to the Body. Front. Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef] [PubMed]
- Tiptiri-Kourpeti, A.; Spyridopoulou, K.; Santarmaki, V.; Aindelis, G.; Tompoulidou, E.; Lamprianidou, E.E.; Saxami, G.; Ypsilantis, P.; Lampri, E.S.; Simopoulos, C.; et al. Lactobacillus casei Exerts Anti-Proliferative Effects Accompanied by Apoptotic Cell Death and Up-Regulation of TRAIL in Colon Carcinoma Cells. PLOS ONE 2016, 11, e0147960. [Google Scholar] [CrossRef] [PubMed]
- Bertkova, I.; Hijova, E.; Chmelarova, A.; Mojzisova, G.; Petrasova, D.; Strojny, L.; Bomba, A.; Zitnan, R. The effect of probiotic microorganisms and bioactive compounds on chemically induced carcinogenesis in rats. Neoplasma 2010, 57, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, N.K.; Sinha, P.R. Inhibition of 1,2-dimethylhydrazine induced colon genotoxicity in rats by the administration of probiotic curd. Mol. Biol. Rep. 2009, 37, 1373–1376. [Google Scholar] [CrossRef]
- Chang, J.-H.; Shim, Y.Y.; Cha, S.-K.; Reaney, M.J.T.; Chee, K.M. Effect of Lactobacillus acidophilus KFRI342 on the development of chemically induced precancerous growths in the rat colon. J. Med Microbiol. 2012, 61, 361–368. [Google Scholar] [CrossRef]
- Appleyard, C.B.; Cruz, M.L.; Isidro, A.A.; Arthur, J.C.; Jobin, C.; De Simone, C. Pretreatment with the probiotic VSL#3 delays transition from inflammation to dysplasia in a rat model of colitis-associated cancer. Am. J. Physiol. Liver Physiol. 2011, 301, G1004–G1013. [Google Scholar] [CrossRef]
- Do, E.; Hwang, S.W.; Kim, S.; Ryu, Y.; Cho, E.A.; Chung, E.; Park, S.; Lee, H.J.; Byeon, J.; Ye, B.D.; et al. Suppression of colitis-associated carcinogenesis through modulation of IL-6/STAT3 pathway by balsalazide and VSL#3. J. Gastroenterol. Hepatol. 2016, 31, 1453–1461. [Google Scholar] [CrossRef]
- Verma, A.; Shukla, G. ProbioticsLactobacillus rhamnosus GG,Lactobacillus acidophilusSuppresses DMH-Induced Procarcinogenic Fecal Enzymes and Preneoplastic Aberrant Crypt Foci in Early Colon Carcinogenesis in Sprague Dawley Rats. Nutr. Cancer 2013, 65, 84–91. [Google Scholar] [CrossRef]
- Baldwin∗, C.; Millette∗, M.; Oth, D.; Ruiz, M.T.; Luquet, F.-M.; Lacroix, M. ProbioticLactobacillus AcidophilusandL. CaseiMix Sensitize Colorectal Tumoral Cells to 5-Fluorouracil-Induced Apoptosis. Nutr. Cancer 2010, 62, 371–378. [Google Scholar] [CrossRef]
- Escamilla, J.; Lane, M.A.; Maitin, V. Cell-Free Supernatants from ProbioticLactobacillus caseiandLactobacillus rhamnosusGG Decrease Colon Cancer Cell Invasion In Vitro. Nutr. Cancer 2012, 64, 871–878. [Google Scholar] [CrossRef]
- Österlund, P.; Ruotsalainen, T.; Korpela, R.; Saxelin, M.; Ollus, A.; Valta, P.; Kouri, M.; Elomaa, I.; Joensuu, H. Lactobacillus supplementation for diarrhoea related to chemotherapy of colorectal cancer: a randomised study. Br. J. Cancer 2007, 97, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Akedo, I.; Otani, T.; Suzuki, T.; Nakamura, T.; Takeyama, I.; Ishiguro, S.; Miyaoka, E.; Sobue, T.; Kakizoe, T. Randomized trial of dietary fiber andLactobacillus casei administration for prevention of colorectal tumors. Int. J. Cancer 2005, 116, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Qin, H.; Yang, Z.; Xia, Y.; Liu, W.; Yang, J.; Jiang, Y.; Zhang, H.; Yang, Z.; Wang, Y.; Zheng, Q. Randomised clinical trial: the effects of perioperative probiotic treatment on barrier function and post-operative infectious complications in colorectal cancer surgery-A double-blind study. Aliment. Pharmacol. Ther. 2011, 33, 50–63. [Google Scholar] [CrossRef]
- Wada, M.; Nagata, S.; Saito, M.; Shimizu, T.; Yamashiro, Y.; Matsuki, T.; Asahara, T.; Nomoto, K. Effects of the enteral administration of Bifidobacterium breve on patients undergoing chemotherapy for pediatric malignancies. Support. Care Cancer 2009, 18, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Kotzampassi, K.; Stavrou, G.; Damoraki, G.; Georgitsi, M.; Basdanis, G.; Tsaousi, G.; Giamarellos-Bourboulis, E.J. A Four-Probiotics Regimen Reduces Postoperative Complications After Colorectal Surgery: A Randomized, Double-Blind, Placebo-Controlled Study. World J. Surg. 2015, 39, 2776–2783. [Google Scholar] [CrossRef]
- Urbancsek, H.; Kazar, T.; Mezes, I.; Neumann, K. Results of a double-blind, randomized study to evaluate the efficacy and safety of Antibiophilus® in patients with radiation-induced diarrhoea. Eur. J. Gastroenterol. Hepatol. 2001, 13, 391–396. [Google Scholar] [CrossRef]
- Chitapanarux, I.; Chitapanarux, T.; Traisathit, P.; Kudumpee, S.; Tharavichitkul, E.; Lorvidhaya, V. Randomized controlled trial of live lactobacillus acidophilus plus bifidobacterium bifidum in prophylaxis of diarrhea during radiotherapy in cervical cancer patients. Radiat. Oncol. 2010, 5, 31–31. [Google Scholar] [CrossRef]
- Golkhalkhali, B.; Rajandram, R.; Paliany, A.S.; Ho, G.F.; Ishak, W.Z.W.; Johari, C.S.; Chin, K.F. Strain-specific probiotic (microbial cell preparation) and omega-3 fatty acid in modulating quality of life and inflammatory markers in colorectal cancer patients: a randomized controlled trial. Asia-Pacific J. Clin. Oncol. 2017, 14, 179–191. [Google Scholar] [CrossRef]
- Gao, Z.; Guo, B.; Gao, R.; Zhu, Q.; Wu, W.; Qin, H. Probiotics modify human intestinal mucosa-associated microbiota in patients with colorectal cancer. Mol. Med. Rep. 2015, 12, 6119–6127. [Google Scholar] [CrossRef]
- dos Reis, S.A.; da Conceição, L.L.; Siqueira, N.P.; Rosa, D.D.; da Silva, L.L.; Peluzio, M.D.C.G. Review of the mechanisms of probiotic actions in the prevention of colorectal cancer. Nutr. Res. 2017, 37, 1–19. [Google Scholar] [CrossRef]
- Vallino, L.; Garavaglia, B.; Visciglia, A.; Amoruso, A.; Pane, M.; Ferraresi, A.; Isidoro, C. Cell-free Lactiplantibacillus plantarum OC01 supernatant suppresses IL-6-induced proliferation and invasion of human colorectal cancer cells: Effect on β-Catenin degradation and induction of autophagy. J. Tradit. Complement. Med. 2023, 13, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Garavaglia, B.; Vallino, L.; Ferraresi, A.; Amoruso, A.; Pane, M.; Isidoro, C. Probiotic-Derived Metabolites from Lactiplantibacillus plantarum OC01 Reprogram Tumor-Associated Macrophages to an Inflammatory Anti-Tumoral Phenotype: Impact on Colorectal Cancer Cell Proliferation and Migration. Biomedicines 2025, 13, 339. [Google Scholar] [CrossRef] [PubMed]
- Ambalam, P.; Dave, J.; Nair, B.M.; Vyas, B. In vitro Mutagen binding and antimutagenic activity of human Lactobacillus rhamnosus 231. Anaerobe 2011, 17, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Pithva, S.P.; Ambalam, P.S.; Ramoliya, J.M.; Dave, J.M.; Vyas, B.R.M. Antigenotoxic and Antimutagenic Activities of ProbioticLactobacillus rhamnosusVc against N-Methyl-N′-Nitro-N-Nitrosoguanidine. Nutr. Cancer 2015, 67, 1142–1150. [Google Scholar] [CrossRef]
- Compare, D.; Rocco, A.; Coccoli, P.; Angrisani, D.; Sgamato, C.; Iovine, B.; Salvatore, U.; Nardone, G. Lactobacillus casei DG and its postbiotic reduce the inflammatory mucosal response: an ex-vivo organ culture model of post-infectious irritable bowel syndrome. BMC Gastroenterol. 2017, 17, 1–8. [Google Scholar] [CrossRef]
- Hou, H.; Chen, D.; Zhang, K.; Zhang, W.; Liu, T.; Wang, S.; Dai, X.; Wang, B.; Zhong, W.; Cao, H. Gut microbiota-derived short-chain fatty acids and colorectal cancer: Ready for clinical translation? Cancer Lett. 2022, 526, 225–235. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The Role of Short-Chain Fatty Acids in Health and Disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [CrossRef]
- Wang, H.-B.; Wang, P.-Y.; Wang, X.; Wan, Y.-L.; Liu, Y.-C. Butyrate Enhances Intestinal Epithelial Barrier Function via Up-Regulation of Tight Junction Protein Claudin-1 Transcription. Dig. Dis. Sci. 2012, 57, 3126–3135. [Google Scholar] [CrossRef]
- Burger-van Paassen, N.; Vincent, A.; Puiman, P.J.; Van Der Sluis, M.; Bouma, J.; Boehm, G.; van Goudoever, J.B.; Van Seuningen, I.; Renes, I.B. The regulation of intestinal mucin MUC2 expression by short-chain fatty acids: implications for epithelial protection. Biochem. J. 2009, 420, 211–219. [Google Scholar] [CrossRef]
- Shao, X.; Sun, S.; Zhou, Y.; Wang, H.; Yu, Y.; Hu, T.; Yao, Y.; Zhou, C. Bacteroides fragilis restricts colitis-associated cancer via negative regulation of the NLRP3 axis. Cancer Lett. 2021, 523, 170–181. [Google Scholar] [CrossRef]
- Ohira, H.; Fujioka, Y.; Katagiri, C.; Mamoto, R.; Aoyama-Ishikawa, M.; Amako, K.; Izumi, Y.; Nishiumi, S.; Yoshida, M.; Usami, M.; et al. Butyrate Attenuates Inflammation and Lipolysis Generated by the Interaction of Adipocytes and Macrophages. J. Atheroscler. Thromb. 2013, 20, 425–442. [Google Scholar] [CrossRef]
- Kibbie, J.J.; Dillon, S.M.; Thompson, T.A.; Purba, C.M.; McCarter, M.D.; Wilson, C.C. Butyrate directly decreases human gut lamina propria CD4 T cell function through histone deacetylase (HDAC) inhibition and GPR43 signaling. Immunobiology 2021, 226, 152126. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-M.; Yu, Y.-N.; Wang, J.-L.; Lin, Y.-W.; Kong, X.; Yang, C.-Q.; Yang, L.; Liu, Z.-J.; Yuan, Y.-Z.; Liu, F.; et al. Decreased dietary fiber intake and structural alteration of gut microbiota in patients with advanced colorectal adenoma. Am. J. Clin. Nutr. 2013, 97, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Weir, T.L.; Manter, D.K.; Sheflin, A.M.; Barnett, B.A.; Heuberger, A.L.; Ryan, E.P. Stool Microbiome and Metabolome Differences between Colorectal Cancer Patients and Healthy Adults. PLoS ONE 2013, 8, e70803. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Umar, S.; Rust, B.; Lazarova, D.; Bordonaro, M. Secondary Bile Acids and Short Chain Fatty Acids in the Colon: A Focus on Colonic Microbiome, Cell Proliferation, Inflammation, and Cancer. Int. J. Mol. Sci. 2019, 20, 1214. [Google Scholar] [CrossRef]
- Luo, S.; Li, Z.; Mao, L.; Chen, S.; Sun, S. Sodium butyrate induces autophagy in colorectal cancer cells through LKB1/AMPK signaling. J. Physiol. Biochem. 2018, 75, 53–63. [Google Scholar] [CrossRef]
- Yang, L.; Liu, C.; Zhao, W.; He, C.; Ding, J.; Dai, R.; Xu, K.; Xiao, L.; Luo, L.; Liu, S.; et al. Impaired Autophagy in Intestinal Epithelial Cells Alters Gut Microbiota and Host Immune Responses. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef]
- Wang, J.; Hu, J.; Wang, M.; Yuan, H.; Xing, Y.; Zhou, X.; Ding, M.; Chen, W.; Qu, B.; Zhu, L. CISD2 Promotes Proliferation of Colorectal Cancer Cells by Inhibiting Autophagy in a Wnt/β-Catenin-Signaling-Dependent Pathway. Biochem. Genet. 2022, 61, 615–627. [Google Scholar] [CrossRef]
- Garavaglia, B.; Vallino, L.; Ferraresi, A.; Esposito, A.; Salwa, A.; Vidoni, C.; Gentilli, S.; Isidoro, C. Butyrate Inhibits Colorectal Cancer Cell Proliferation through Autophagy Degradation of β-Catenin Regardless of APC and β-Catenin Mutational Status. Biomedicines 2022, 10, 1131. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Baehrecke, E.H.; Brumell, J.H.; Chu, C.T.; Codogno, P.; Cuervo, A.M.; Debnath, J.; Deretic, V.; Elazar, Z.; Eskelinen, E.-L.; et al. A comprehensive glossary of autophagy-related molecules and processes (2ndedition). Autophagy 2011, 7, 1273–1294. [Google Scholar] [CrossRef]
- Yang, Y.; Klionsky, D.J. Autophagy and disease: unanswered questions. Cell Death Differ. 2020, 27, 858–871. [Google Scholar] [CrossRef] [PubMed]
- Foerster, E.G.; Mukherjee, T.; Cabral-Fernandes, L.; Rocha, J.D.; Girardin, S.E.; Philpott, D.J. How autophagy controls the intestinal epithelial barrier. Autophagy 2021, 18, 86–103. [Google Scholar] [CrossRef]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Quaglio, A.E.V.; Grillo, T.G.; De Oliveira, E.C.S.; Di Stasi, L.C.; Sassaki, L.Y. Gut microbiota, inflammatory bowel disease and colorectal cancer. World J. Gastroenterol. 2022, 28, 4053–4060. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, H.K. Potential Role of the Gut Microbiome In Colorectal Cancer Progression. Front. Immunol. 2022, 12, 807648. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Jeong, S. Mutation Hotspots in the β-Catenin Gene: Lessons from the Human Cancer Genome Databases. Mol Cells. 2019, 42, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Wnt/β-Catenin Signaling in Development and Disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef]
- Zhao, H.; Ming, T.; Tang, S.; Ren, S.; Yang, H.; Liu, M.; Tao, Q.; Xu, H. Wnt signaling in colorectal cancer: pathogenic role and therapeutic target. Mol. Cancer 2022, 21, 1–34. [Google Scholar] [CrossRef]
- Markowitz, S.D.; Bertagnolli, M.M. Molecular origins of cancer: Molecular Basis of Colorectal Cancer. New Engl. J. Med. 2009, 361, 2449–2460. [Google Scholar] [CrossRef]
- Ionescu, V.A.; Gheorghe, G.; Bacalbasa, N.; Chiotoroiu, A.L.; Diaconu, C. Colorectal Cancer: From Risk Factors to Oncogenesis. Medicina 2023, 59, 1646. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Kaźmierczak-Siedlecka, K.; Marano, L.; Merola, E.; Roviello, F.; Połom, K. Sodium butyrate in both prevention and supportive treatment of colorectal cancer. Front. Cell. Infect. Microbiol. 2022, 12, 1023806. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Lee, G.; Son, H.; Koh, H.; Kim, E.S.; Unno, T.; Shin, J.-H. Butyrate producers, “The Sentinel of Gut”: Their intestinal significance with and beyond butyrate, and prospective use as microbial therapeutics. Front. Microbiol. 2023, 13, 1103836. [Google Scholar] [CrossRef]
- Food and Agriculture Organization and of the United Nations/World Health Organization. (2002). Guidelines for the evaluation of probiotics in foods, Available at: https://www.who.int>fs management>probiotic guidelines.
- Zhou, X.; Zhao, Y.; Zhao, R.; Shafi, S.; Yang, Y.; Liu, G.; Liu, S.-B. Research progress of intestinal microbiota in targeted therapy and immunotherapy of colorectal cancer. J. Cancer Metastasis Treat. 2024, 10. [Google Scholar] [CrossRef]
- Azad, M.A.K.; Sarker, M.; Li, T.; Yin, J. Probiotic Species in the Modulation of Gut Microbiota: An Overview. BioMed Res. Int. 2018, 2018, 9478630. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, P.; Weiskirchen, S.; Weiskirchen, R. Effects of Probiotics on Gut Microbiota: An Overview. Int. J. Mol. Sci. 2024, 25, 6022. [Google Scholar] [CrossRef]
- Zhang, J.-W.; Du, P.; Yang, B.-R.; Gao, J.; Fang, W.-J.; Ying, C.-M. Preoperative Probiotics Decrease Postoperative Infectious Complications of Colorectal Cancer. Am. J. Med Sci. 2012, 343, 199–205. [Google Scholar] [CrossRef]
- Nanno M, Kato I, Kobayashi T, Shida K. Biological effects of probiotics: what impact does Lactobacillus casei shirota have on us? Int J Immunopathol Pharmacol. 2011 Jan-Mar;24(1 Suppl):45S-50S. [PubMed]
Figure 1.
Role of autophagy in regulating intestinal homeostasis. Autophagy counteracts colorectal cancer (CRC) by limiting cell proliferation and migration, while also mitigating inflammation associated with inflammatory bowel disease (IBD), a condition that can progress to CRC. This highlights the dual protective role of autophagy in maintaining intestinal integrity and preventing tumorigenesis (created with BioRender).
Figure 1.
Role of autophagy in regulating intestinal homeostasis. Autophagy counteracts colorectal cancer (CRC) by limiting cell proliferation and migration, while also mitigating inflammation associated with inflammatory bowel disease (IBD), a condition that can progress to CRC. This highlights the dual protective role of autophagy in maintaining intestinal integrity and preventing tumorigenesis (created with BioRender).
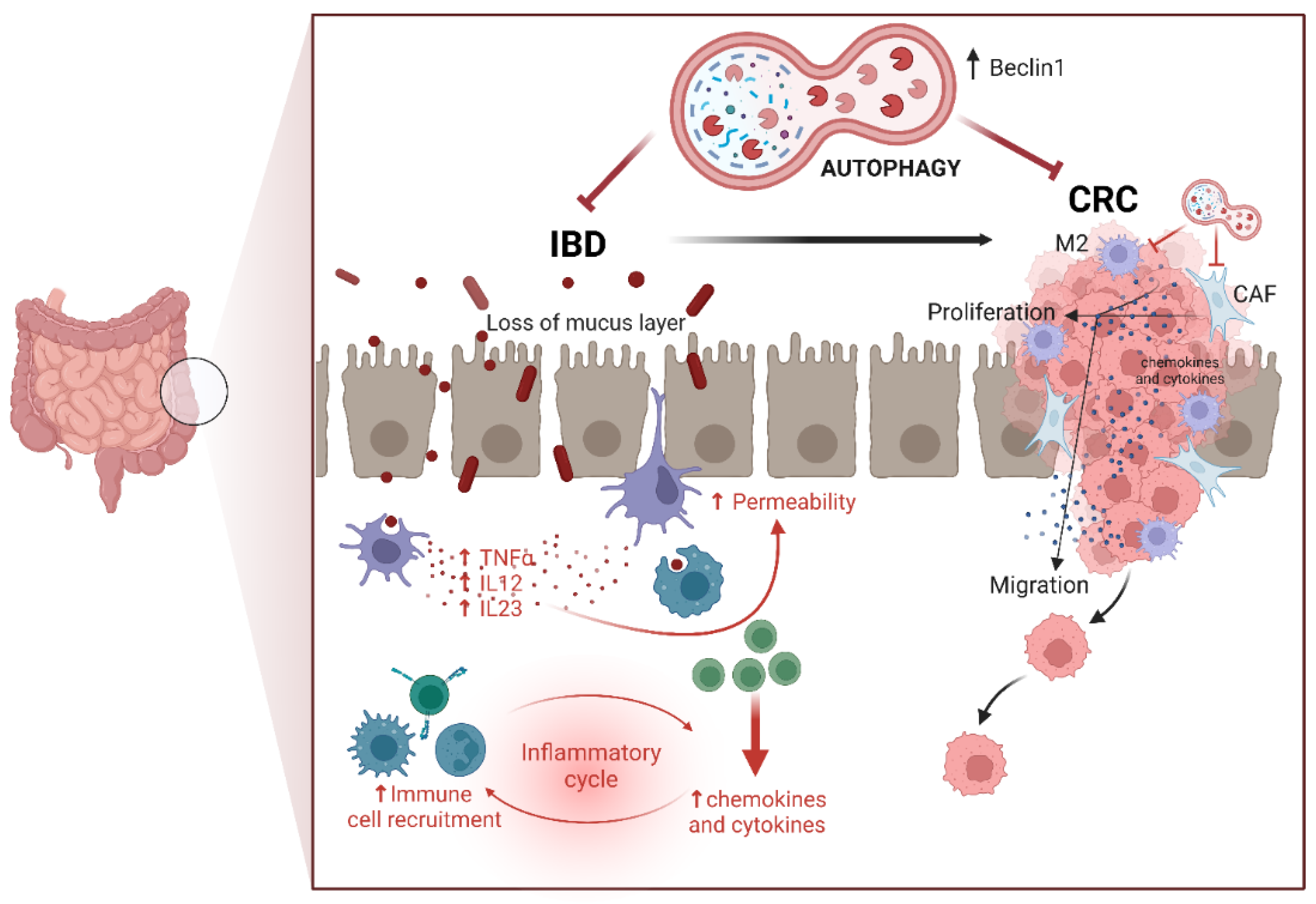
Figure 2.
Impact of gut microbiota on colorectal cancer progression. Healthy gut microbiota maintains epithelial integrity and energy balance, prevents inflammation, provides essential nutrients and vitamins, produces short-chain fatty acids (SCFAs), and protects against pathogen invasion. In contrast, dysbiotic gut microbiota increases intestinal permeability, promotes chronic inflammation and oxidative stress, induces DNA damage, activates oncogenic pathways, and reduces SCFAs production, all contributing to colorectal cancer development (created with BioRender).
Figure 2.
Impact of gut microbiota on colorectal cancer progression. Healthy gut microbiota maintains epithelial integrity and energy balance, prevents inflammation, provides essential nutrients and vitamins, produces short-chain fatty acids (SCFAs), and protects against pathogen invasion. In contrast, dysbiotic gut microbiota increases intestinal permeability, promotes chronic inflammation and oxidative stress, induces DNA damage, activates oncogenic pathways, and reduces SCFAs production, all contributing to colorectal cancer development (created with BioRender).
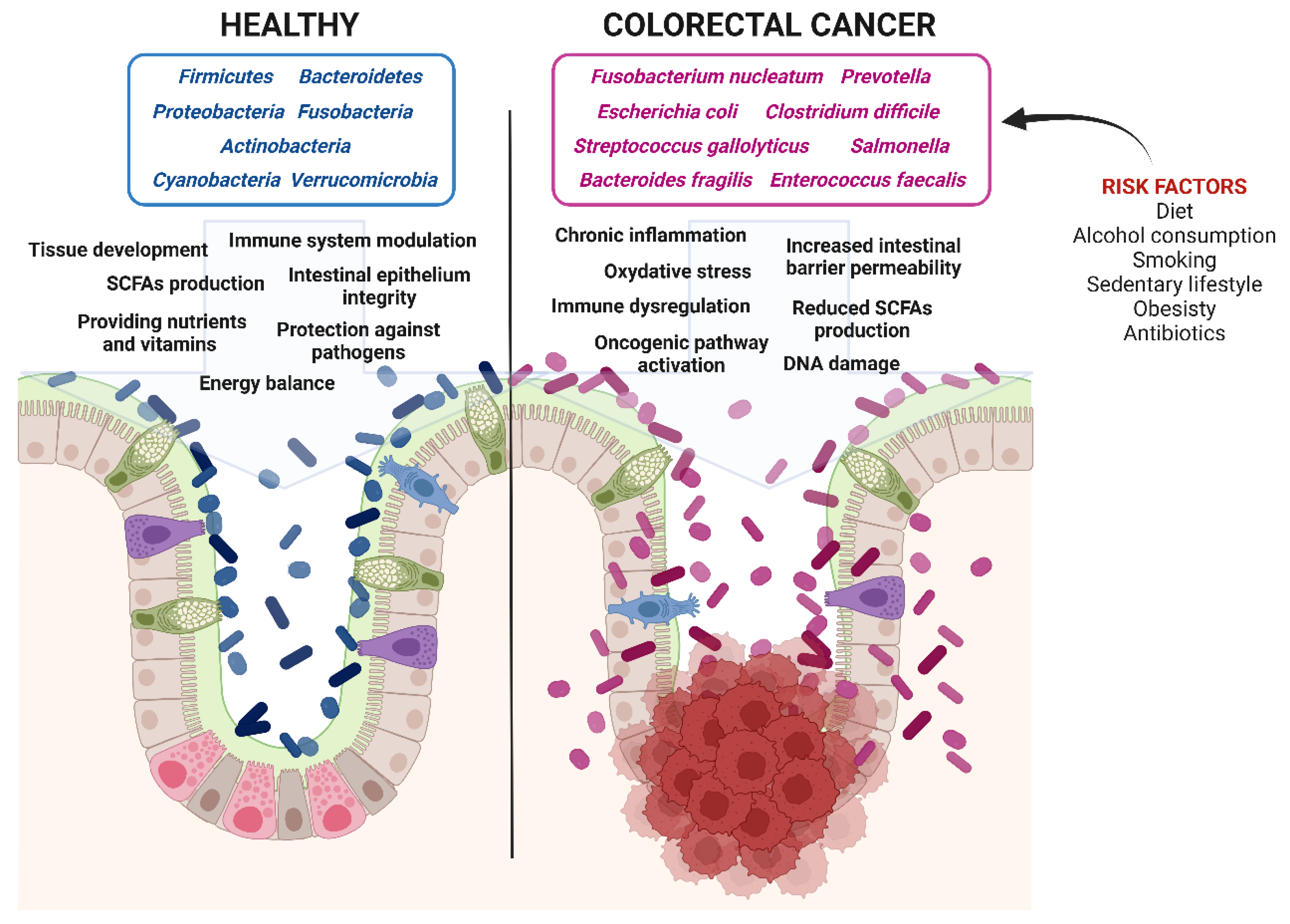
Figure 3.
Role of SCFAs in the intestinal epithelium. SCFAs, particularly butyrate, exhibit anti-carcinogenic and immunomodulatory effects, protect against pathogen invasion through mucus production, maintain intestinal integrity, and act as modulators of gene expression (created with BioRender).
Figure 3.
Role of SCFAs in the intestinal epithelium. SCFAs, particularly butyrate, exhibit anti-carcinogenic and immunomodulatory effects, protect against pathogen invasion through mucus production, maintain intestinal integrity, and act as modulators of gene expression (created with BioRender).
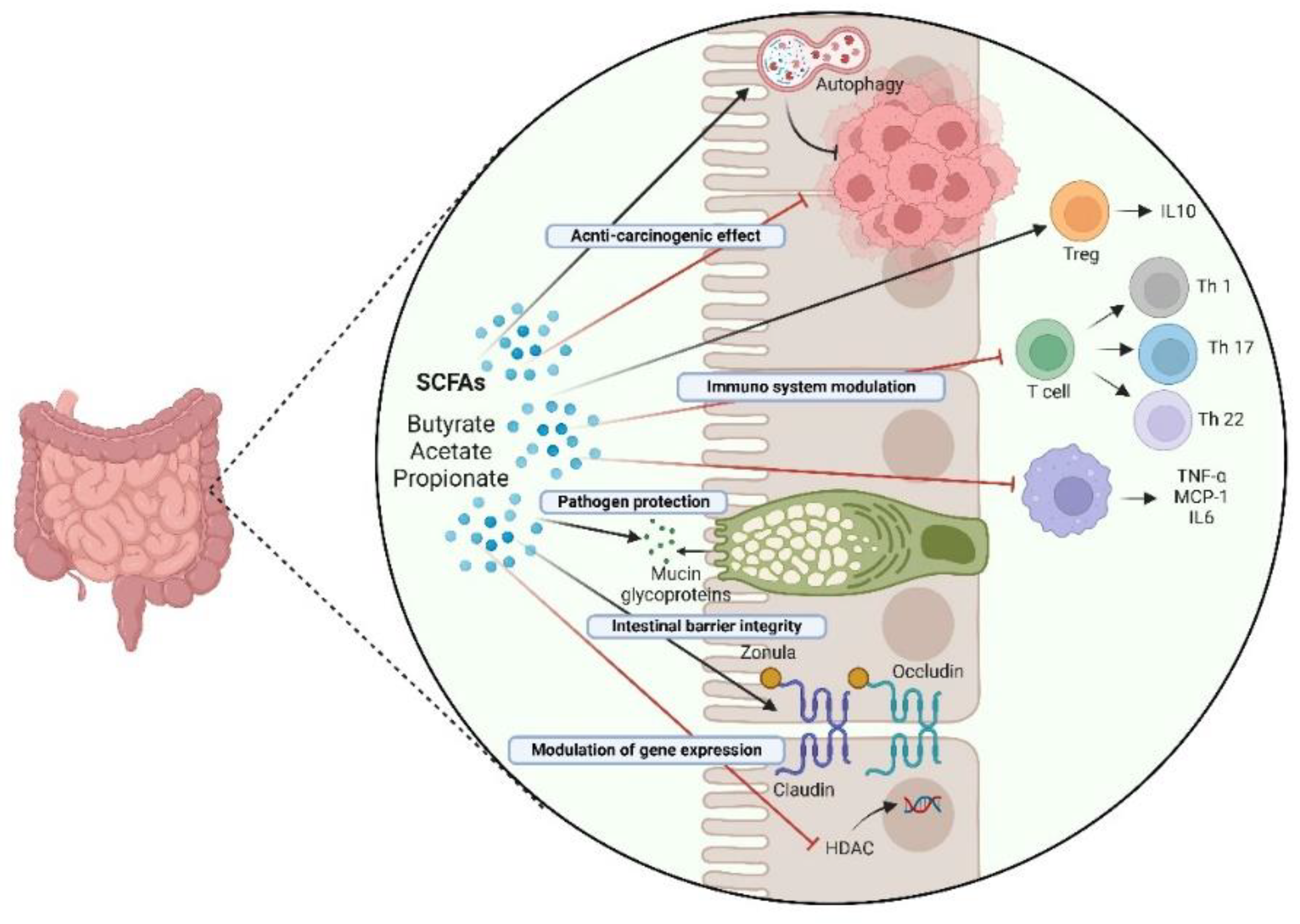
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



