Submitted:
02 May 2025
Posted:
05 May 2025
Read the latest preprint version here
Abstract
The tumor microenvironment (TME) is a unique ecosystem that surrounds tumor tissues. The TME is composed of extracellular matrix, immune cells, blood vessels, stromal cells, and fibroblasts. These environments enhance cancer development, progression, and metastasis. Recent success in immune checkpoint blockade also supports the importance of the TME and immune cells residing in the tumor niche. Although the TME can be identified in almost all cancer types, the role of the TME may not be similar among different cancer types. Regulatory T cells (Tregs) play a pivotal role in immune homeostasis and are frequently found in the TME. Owing to their suppressive function, Tregs are often considered as unfavorable factors that allow the immune escape of cancer cells. However, the presence of Tregs is not always linked to an unfavorable phenotype, which can be explained by the heterogeneity and plasticity of Tregs. In this review, the current understanding of the role of Tregs in TME is addressed for each cancer cell type. Moreover, recently a therapeutic approach targeting the TME has been developed. Thus, understanding the role of Tregs in the TME may lead to the development of novel therapies that directly target the TME.
Keywords:
FOXP3
; Regulatory T cells
; Immune regulation
; Tolerance
; Tumor-infiltrating lymphocyte
; Cancer
1. Introduction
The tumor microenvironment (TME) is a unique ecosystem around the tumor tissue that helps escape immune recognition and elimination by the adaptive immune system [1]. The components of the TME are tumor-specific, but typically comprise extracellular matrix, immune cells, blood vessels, stromal cells, and fibroblasts [2,3]. Cancer cells utilize TME. Among the various immune cells, tumor-infiltrating lymphocytes (TIL) and tumor-associated macrophages (TAM) contribute to the generation of tumor niche [4,5,6]. Currently, immune cells residing in tissues are identified in various organs and have adapted to these tissues [7]. Recent breakthroughs in cancer immunotherapy have indicated the importance of local immune reactions that mainly occur or are influenced by the TME [8]. Tregs express inhibitory molecules, including PD-1, PD-L1, and CTLA-4; therefore, checkpoint blockade can potentially activate Tregs [9]. In addition to the adaptive immune system, the role of the innate immune system has been indicated in tumor immunity, and crosstalk between the innate and adaptive immune systems can orchestrate this process [10]. The importance of the TME in tumor metastasis has also been recognized [11,12]. Therefore, understanding the TME is critical for developing novel therapeutic approaches [13]. Checkpoint blockade is targeting TME by modulating immune cells residing in TME. Moreover, the resent research is targeting extracellular marix, blood vessels, stromal cells and fibroblasts. In correspondence with the therapeutic approaches targeting TME, the complexity and heterogenisity of TME has been described by multiple researchers [14].
In this review, the role of regulatory T cells (Tregs) in the TME is introduced together with the recent clinical and translational investigations including antidoby drug conjugate, immunotoxin and FOXP3 inhibitory peptides. Thus, understating the role of Treg in the TME may contribute to TME-directed therapy.
2. Biological Role of Tregs in the Local Immune System
Tregs are a unique cell population that contribute to immune homeostasis through their suppressive function [15]. FOXP3 is a master transcription factor that regulates Treg differentiation and phenotypic identity. The phenotype and suppressive function of Tregs are mainly regulated by FOXP3 expression [16,17]. Mutation in FOXP3 in human result in IPEX syndrome, whereas mutations in Foxp3 causes scurfy in mice, both of which are characterized by lethal autoimmunity due to the lack of functional Tregs [18]. Genetic mutations and instability or dysfunction of Tregs have been reported in multiple autoimmune diseases, allergic diseases, cancers, and neurological diseases [19]. It is commonly believed that the FOXP3 expression and suppressive function are highly correlated. Moreover, FOXP3 can drive the expression of co-inhibitory molecules including CTLA-4. Here, we address the basic phenotype and function of Tregs as well as their plasticity and heterogeneity.
2.1. Phenotype and Function of Tregs
In mice, Tregs were initially identified as CD4+ and CD25+ cells [20]. FOXP3 has been identified as the master regulator [16]. CD4+CD25+CD127low cells express FOXP3, and FOXP3 + Tregs can be isolated without FOXP3 staining [21]. In addition to “bona fide” Treg markers, Tregs express CTLA-4, GARP, GITR, and IKZF2, which are known FOXP3 biding targets [22]. Therefore, high and stable FOXP3 expressions in Tregs are important in the phenotype and suppressive function of Tregs.
Other markers (OX40, CD69, TIGIT, ICOS, Lag-3, PD-1, CD39, and CD73) were also included in the phenotypic assessments (Figure 1a) [23]. However, several other molecules (CD69, TIGIT, and ICOS) are also expressed by effector T-cells upon activation and may not be specific to Tregs. An in vitro suppression assay was conducted to assess the suppressive function [24,25]. In addition to phenotyping, the stability and function of Tregs have been assessed in vitro. Notably, majority of the Treg-related molecules including CTLA-4, GARP, GITR, TIGIT, ICOS and PD-1 has been utilized for the therapeutic targets in cancer immunotherapy. Therefore, the understanding of Treg phenotype in the cancer TME could provide information of potential targets in cancer immunotherapy.
2.2. Plasticity and Heterogeniety of Tregs
Tregs exhibit plasticity particularly under inflammation and pro-inflammatory cytokines [26,27,28]. In particular, in acquired autoimmune diseases, loss of Treg identity can contribute to disease progression [29]. Tregs express IL-1R and IL-6R, which are stimulated by IL-1 and IL-6 cytokines, and may lose FOXP3 expression [30]. To support these findings, genetic inhibition of IL-1R and IL-6R enhance Treg stability in vitro [31]. These observations provide the evidence of Treg plasticity which may be necessary to maintain immune homeostasis.
Other factors also contribute to Treg/Th17 differentiation, which may contribute to inflammation and autoimmunity [32,33,34]. In addition to their plasticity, Tregs have subpopulations similar to effector T cells [35,36]. This plasticity and heterogeneity provide Tregs with—temporal diversity (Figure 1b). Local metabolic status also contributes to Treg heterogeneity [27]. Therefore, it is speculated that the phenotype and identity of Tregs can potentially heterogenious in the each cancer tissues and affected by the presence of infection and inflammation.
2.3. Tissue-Resident Tregs
Tregs have been identified in local non-lymphoid tissues, including adipose tissues, muscles, skin, and the gastrointestinal tract [37]. Tissue-resident T cells contribute to immune reactions in local tissues [38,39,40]. Barrier tissues, including the respiratory tract and skin, have resident memory cells, primarily for a rapid response to known antigens (Figure 1c). Similar to TRM, tissue-resident Tregs have been proposed, but they are not readily identified. Recently, single-cell sequencing has allowed the identification of tissue-resident Tregs; however, the clonality of tissue-resident Tregs is similar among different tissues, and whether these Tregs are strictly tissue resident remains unclear [41,42,43]. Tissue-resident T cells and Tregs contribute to cancer initiation; however, the exact role of these newly identified T cells in carcinogenesis remains under investigation. Nevertheless, heterogeneity, plasticity and spatiotemporal diversity of Tregs could influence local immune homeostasis and may contribute to the cancer progression and metastasis. Further study is warranted to confirm the phenotype and function of tissue-resident Tregs both in physiological and pathological conditions.
3. Phenotype and Function of Tregs Isolated from TME
Tregs reside in the TME across multiple tumor types. Owing to the immunosuppressive effects of Tregs, the presence of Tregs in the TME and/or tumor tissues is considered detrimental; however, in some cancer types, the presence of Tregs in the TME could be associated with favorable outcomes. These complex clinical observations indicate the biological roles of Tregs can differe across different cancer types. Therefore, determining whether Tregs suppress tumor immunity or enhance tumor elimination is challenging. The current interpretation of tumor-infiltrating Tregs is discussed according to specific cancer type.
3.1. Colorectal Cancer
Most immune cells reside in the gastrointestinal tract; hence, the presence of immune cells in the tumor niche is presumed (Figure 2). While Tregs can be detected in the gastrointestinal tract in the physiological conditions, Tregs have also been identified in colon cancer [44]. Tregs isolated from colon cancer cells exhibit effector phenotypes [45]. Moreover, Treg depletion or inhibition can enhance antitumor activity [46,47]. Blockade of checkpoints, including CTLA-4 and PD-1, targets effector T cells and Tregs in the TME [48]. However, infiltration of Tregs is associated with better prognosis. Tregs may not be suppressive and may enhance local immune reactions [49]. Moreover, the plasticity of Tregs may be beneficial for tumor immunity. According to these data, Tregs are not fully responsible to the cancer progression and Treg ablation may not be beneficial in the colon cancer. However, enhancing tumor immunity could be beneficial to the patients with colon cancer.
3.2. Lung Cancer
Resident memory T-cells are identified in the respiratory tract [39,40]. Similaly to the gastrointestinal tract, Tregs are identified in the lung in the physiological condition. Involvement of Tregs has also been identified in other lung diseases including acute lung injury and infection (Figure 3) [50,51]. Therefore, it is speculated that Treg may control immune homeostasis in the lung both in physiological and pathological conditions.
Tregs have also been identified in lung cancer [52]. Treg infiltration in lung cancer may result in divergent phenotypes [53]. Moreover, COX-2 expression may be associated with the long-term prognosis in non-small cell lung cancer [54]. Unlike colorectal cancer, literature on Treg infiltration resulting in favorable outcomes are unavailable. Thus, Treg infiltration may result in immune escape, contributing to tumor development and relapse. To support these results, selective depletion of CCR8+ Tregs are beneficial in the lung cancer in the mice model [55]. Collectively, current clinical and translational evidence suggest beneficial effects of Treg depletion in lung cancer.
3.3. Breast Cancer
Unlike the gastrointestinal and respiratory tract which were considered as barrier tissues exposed to foreign antigen, breast tissue may not contain a numbers of immune cells in the physiological condition. However, the presence of inflammation is associated with breast cancer progression [56]. Tregs are isolated from breast cancer cells and exhibit a unique phenotype [57]. Recently, immunotherapy has been used in the management of triple-negative breast cancer, suggesting the involvement of Tregs in the TME [58]. In addition to their divergent phenotypes, Tregs in breast cancer may contribute to disease progression (Figure 4) [59]. Moreover, the interaction between Tregs in PBMC and the cancer niche may cause disease progression and relapse [60]. Indeed, Treg ablation is benefical and enhance the radiotherapy in the mice model [61]. Furthermore, the alteration of TME including the presence of Tregs are associated with breast cancer progression [62]. These results indicate the detrimental effect of Tregs in breast cancer and suggest that it may be beneficial to deplete or suppress Tregs in the TME in breast cancers.
3.4. Pancreatic Cancer
Pancreas is not immune organs, but there are lots of evidence suggesting immune cells plying central roles in inflammations and cancer development in the pancreas [63]. Tregs in the pancreas have been studied more often in patients with type I diabetes [64,65,66,67]. Moreover, pro-insulin or GAD specific Tregs that protect islet from autoimmunity and loss or disfunction of antigen specific Tregs may contribute to the development of type 1 diabetes. Moreover, adoptive Treg transfer is conducted in the multiple clinical trials based on the Treg dysfunction in T1D [68]. In contrast, pancreatic cancer called as “cold tumor” is characterized by minor immune cell infiltration [69]. TME has been identified in pancreatic cancer [70,71]. Tregs have also been identified in pancreatic cancer (Figure 5) [72]. Unlike in other cancer types, Treg depletion has shown detrimental effects [73]. Thus, the TME is controlled and maintained through different mechanisms across different cancer types. In addition, immunotherapy has been indicated for pancreatic cancer management, but its efficacy has not been fully confirmed, partially because of disease severity [74]. Collectively, the role of Tregs in pancreatic cancer has not been fully confirmed. Thus, further clincal and translational study of the Tregs in pancreatic cancer is warranted to validate the possible role of Tregs in the cancer progression.
3.5. Hepatocellular Carcinoma
While human liver is not usually considered as immune organ, the presence of various immune cells has been well documented in the past [75]. A recent study identified multiple immune cells, including Tregs, in the liver as previously suggested [76]. Either in autoimmune or viral hepatitis, both innate and adaptive immune cells are activated [77,78]. Tregs reside in liver tissue, and dysfunction of liver Tregs is associated with fibrosis [79]. Therefore, it has been speculated that Tregs reside in the liver under healthy conditions and regulate immune responses and regeneration, including fibrosis. TME has also been identified in hepatocellular carcinoma [80,81]. The association between cancer and Treg infiltration is not evident compared with other types of cancer [82]. Chronic HBV and HCV infections can contribute to the development of hepatocellular cancer; moreover, it has been suggested that HBV/HCV potentially utilizes Tregs to escape the immune system, similar to other cancer cell types [83]. Their association with chronic infection may complicate the determination of the role of Tregs in the TME independent of HBV/HCV infection. Currently, the lack of clinical and translational study in hepatocellular carcinoma focusing on TME make it difficult to consider immunotherapy and Treg ablation to enhance tumor immunity. In addition, there is few evidence on Treg ablation in hepatocellular carcinoma. The description of Tregs in hepatocellular carcinoma may provide the future insight of the therapeutic application.
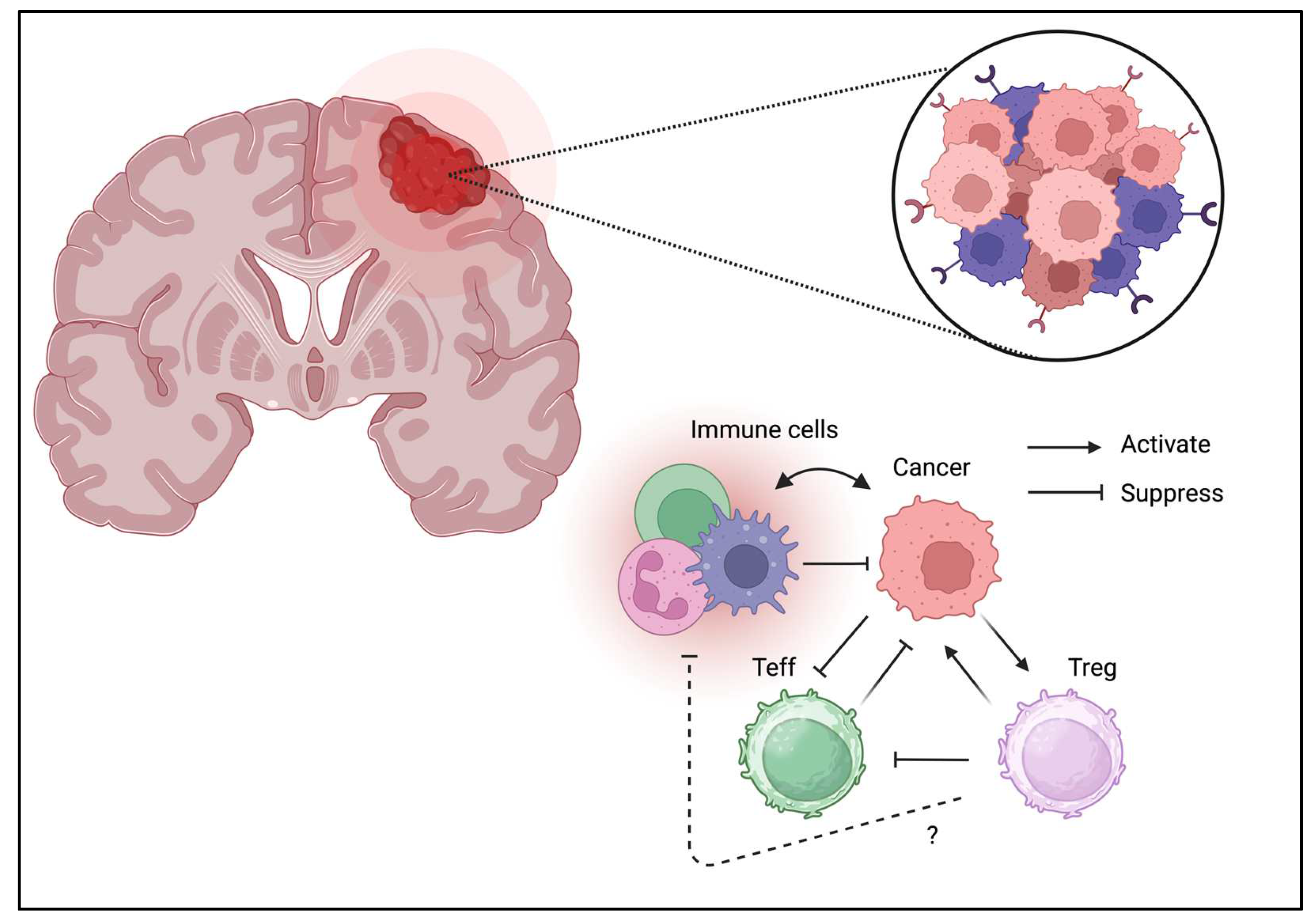
3.6. Brain Tumor
Unlike other tissues, the brain tissue is considered an “immune-privileged site; however, immune cells directly contribute to brain hemostasis [84]. Moreover, the presence of Tregs in the brain has been confirmed and may contribute to neurological function [85]. Similar to other cancer types, Treg infiltration can be a unfavorable prognostic factor for brain tumors (Figure 6) [86]. Furthermore, the brain tumor expresses various immunomodulatory molecules which are necessary to control immune response even in the “immune-privileged site.” PD-1/PD-L1 expression is also associated with clinical phenotypes [87]. Glioblastomas may suppress immune reactions owing to its strong IDO expression [88]. Moreover, GITR targeting improves therapeutic efficacy [89]. These results suggest brain tumor directly suppress immune response by similar suppressive mechanism through co-inhibitory molecules. These fundings may suggest that either immunotherapy and Treg ablation may enhance tumor immunity and potenitially improve the clinical outcome.
3.7. Melanoma
The skin tissue contains multiple immune cells, and various tissue-resident cells have been reported [90]. As expected, Tregs have been identified in the skin tissue, and their association with inflammation or allergy has been reported [91,92]. TME has been identified in melanomas, similar to other cancers [93]. Tregs have also been identified in melanomas [94]. The success of PD-1 blockade in melanoma is the benchmark for immunotherapy; therefore, immune regulation is expected to contribute to melanoma progression [95]. With regard to PD-1 blockade, the interaction between CD8+ T cells and Tregs are considered as the key effector cells for tumor rejection [96]. Although PD-1 blockade is curative in some patients, resistance to therapy needs to be investigated for better treatment outcomes. Furthermore, the role of Tregs in melanoma progression is still controversial. Similarly, the effect of Treg ablation is needed to be confirmed in the future study [94].
3.8. Kidney and Urinary Tract Cancer
Unlike other tissues, tissue-resident immune cells are not typically observed in the urinary tract, which is aseptic and may not contain immune cells. However, the urinary tract may have resident memory T cells and the presence of TRM has been confirmed after UTI [97]. The TME has been identified in renal cell carcinoma [98] and bladder cancer [99]. Tregs have also been identified in bladder cancer [100]. The roles of the TME and Tregs have not been studied much in urinary tract cancers, including renal cell carcinoma. However, immunotherapy has been used for the management of bladder and prostate cancers [101]. In addition, CCR4 blockade is shown to inhibit Treg infiltration in the bladder cancer in the canine model [102]. Moreover, CXCR4 blockade is similarly shown to inhibit Treg infiltration in the renal carcinoma [103]. While there was not enough evidence, Treg ablation might be beneficial in the patients with kidney and urinary tract cancers.
3.9. Gynecological Cancer
The female reproductive system is a barrier tissue that includes multiple immune cells [104]. Pregnancy and hormonal changes drastically alter systemic and local immunity [105,106]. There results suggest that female reproductive tract are closely associated with immune cells in physiological conditions. The presence of tissue-resident immune cells including T cells, B cells and microphage also support these results. TME has been identified in ovarian cancer [107,108]. Tregs have also been identified in ovarian cancer [109]. Immune checkpoint blockade has also been attempted in cervical cancer treatment [110]. The roles of the TME and Tregs in gynecological cancers are not fully understood, although immunotherapy has been attempted in several cancers. There are little evidence suggesting the effect of Treg ablation in gynecological cancers. Further study exploring the role of Tregs in cancer development, progression and metastasis needs to be conducted to achieve potential immunotherapy approach against female gynecological cancers.
3.10. Head and Neck Cancer
Head and neck cancers encompass multiple cancer types develop in the mouth, throat, nose, sinuses, and salivary glands. Since head and neck contain multiple lymphoid tissues includings lymphnodes and tonsil, their unique immune landscape has been described [111]. Immunotherapy approaches including check point blockade have been investigated in the various head and neck cancers [112]. Similarly to the colon cancer, infiltration of Tregs in cancer tissue is considered as a favaroble factor in head and neck cancers [113]. On the other hand, it was also reported that the increase in effector type (CD45RA-) FOXP3highTreg is negatively correlated with poor prognosis [114]. Another study indicated that CTLA-4+ Tregs may associated with the limited survival after anti-EGFR targeting by inhibiting NK cell function which is critical in the head and neck cancer. It was not possible to conclude whether Tregs are favorable factor in head and neck cancer based on the controversy in the field. Due to the heterogeneity of head and neck cancer, it may not be possible to determine the common effects of Tregs.
3.11. Soft Tissue Sarcoma
Soft tissue sarcoma is usually considered as less immunologic malignancy compared to the other tumor types. However, the presence of Tregs in soft tissue sarcoma may be associated with poor survival [115]. The limited effecacy of immunotherapy such as PD-1 and PD-L1 has been reported [116]. Immune landscape of soft tissue sarcoma is not yet fully investigated but the importance of immunomonitoring in soft tissue sarcoma was suggested in the past [117]. There was few literatures describing the biological function of Tregs in soft tissue sarcoma. Due to the reduced immunogenicity, the association to the Treg infiltration and clinical outcome is not anticipated, nervertheless needs to be confirmed in the future clinical and translational study.
3.12. Leukemia/Lymphoma
Similar to other solid tumors, lymphoma has a TME [118,119]. Leukemia does not have a tumor mass, and the TME has not been fully identified; however, several studies have indicated the involvement of Tregs in disease progression [120,121]. The functions of Tregs have been studied with respect to acute and chronic GvHD [122]. Moreover, the bone marrow niche is not considered the TME; however, leukemic stem cells may utilize the bone marrow as the TME, similar to other cancers [123]. Thus, bone marrow niche may be a potential therapeutic target for leukemia [124]. Interestingly, Tregs are shown to support hematopoietic stem cells engraftment and survival by harnessing TME [125]. It is hypothesized that leukemic stem cell [126]. Collectively, the future study investigating tumor/leukemic niche will provide supporting evidence of Treg ablation in hematologic cancers.
4. Therapeutic Approach Targeting Treg in TME
Tregs are therapeutic targets in cancer immunotherapy [127]. Several therapeutic approaches have been proposed to target Tregs in the TME [128]. Among these, antibody-drug conjugates, immunotoxins, and peptides have frequently been investigated to target Tregs in the TME [129]. However, as discussed in the previous section, the role of Tregs in the TME may be tumor dependent, and its presence may be detrimental in most cancer types. However, it is important to study the beneficial effects of Tregs on TME, as observed in specific cancer types, including colon and pancreatic cancers. The presence of Tregs in the TME has been extensively studied in cancers of the digestive, respiratory, and breast tissues compared to gynecologic and urinary tract cancers. Moreover, understanding normal tissues with respect to tissue-resident cells may contribute to the evaluation and interpretation of immune cells isolated from the TME. Current therapeutic approaches targeting Treg in TME were discussed in the following sections.
4.1. Antibody Drug Conjugate
Among therapeutic candidates targeting TME, antibody durg conjugate is the most promising therapeutic option. Currently, more than 13 antibody durg conjugate products were approved by FDA [130,131]. In preclinical study, CD25 antibody conjugated with pyrrolobenzodiazepine has been shown to eliminate Tregs and enhance tumor immunity against lymphoma [132]. Currently, the safety and efficacy was tested in the phase-1 clinical trial. It may possible to utilize CD25 antibody conjugated with pyrrolobenzodiazepine for intratumoral Treg depletion.
4.2. Immunotoxin
Before the emergence of antibody drug conjugate, immunotoxin targeting IL-2 and CD25 has been studied to delplete Tregs in vivo [133,134]. Similarly to CD25 antibody drug conjugate, both IL-2 and CD25 immunotoxin successfully reduced Tregs in vivo and enhance T cell immune response suppressed by Tregs. Since mechanism actions were shared by antibody durg conjugate and immunotoxin, CD25 targeting is currently most efficiently reduced Tregs.
4.3. Peptide
Aside from antibody durg conjugate and immunotoxin, peptide durg conjugate is investigated to target Tregs. CD28 targeting aptamer was combined with P60, synthetic 15-mer FOXP3 inhibiting peptide, and shown to inhibit Tregs [135]. Similarly, P60 conjugated with CD25 targeting nanoliposome is shown to reduce intratumoral Tregs [136]. While antibody drug conjugate and immunotoxin directly eliminate Tregs, P60 inhibit FOXP3 and potentially reduce suppressive function. Therefore, inhibited Tregs were not elimitated from the tissues, however, peptide based approach may have superior safety compared to the other Treg targeting strategies.
5. Future Investigations for Targeting Treg in TME
Currently, several therapeutic applications were investigated in the early phase clinical trial such as CD25antibody drug conjugate. Immunotoxin and peptide showed similar efficacy in the preclinical models. Therefore, it may become possible to deplete Tregs in vivo. Howerver, it would be important to monitor immune response after Treg depletion to avoid unwanted immune responses such as autoimmunity. Unlike the mice, it is not readily possible to ablate Tregs in human without adverse events. Especially, it must be even more challenging to ablate tumor-infiltrating Tregs than peripheral Tregs. As suggested in the mice, the blockade of homing receptor (CXCR4) and chemokine receptor (CCR4/CCR8) may inhibit Treg infiltration and tumor-homing. More precise understanding Tregs in TME may provide more direct evidence of Treg ablation/depletion in the context of cancer immunotherapy.
In addition Tregs, the other immune cells including effector T cells, NK cells and macrophage may play a role in TME, however the biological role is not fully determined. Moreover, the role of other non-immune cells such as stromal cells, fibroblasts and blood vessels are not often studied compared to the immune cells. The interaction among immune cells, non-immune cells and cancer/tumor cells needs to be studied, nevertheless, it may not be readily possible due to the limitation of the patient sample and ethical issues. The discovery of the crosstalk between cells in TME may lead to novel therapeutic interventions.
6. Conclusions
To date, the biological role of TME and the functional properties of tumor infiltrating Tregs were vigorously investigated across various cancer types. Recent advances in tumor immunology, single cell sequencing and spatial transcriptomic analysis identified tumor infiltrating Tregs and propsed possible therapeutic candidates. Diversity of clinical background, TME, Treg heterogeneity needs to be considered and the importance of clinical and translationa study are appreciated in the field. The precise understandings of TME and tumor infiltrating Tregs may shed new light on the cancer immunotherapy.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author thank Editage for the language editing of the manuscript. The author thanks Akihito Tsubota (The Jikei University School of Medicine) for the encouragement.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Anderson, N.M.; Simon, M.C. The tumor microenvironment. Curr. Biol. 2020, 30, R921–R925. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, N.A.; Sanchez-Salas, R.; Peske, J.D.; Vano, Y.; Becht, E.; Petitprez, F.; Validire, P.; Ingels, A.; Cathelineau, X.; Fridman, W.H.; et al. The clinical role of the TME in solid cancer. Br. J. Cancer 2019, 120, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Mayer, S.; Milo, T.; Isaacson, A.; Halperin, C.; Miyara, S.; Stein, Y.; Lior, C.; Pevsner-Fischer, M.; Tzahor, E.; Mayo, A.; et al. The tumor microenvironment shows a hierarchy of cell-cell interactions dominated by fibroblasts. Nat. Commun. 2023, 14, 5810. [Google Scholar] [CrossRef]
- Sarnaik, A.A.; Hwu, P.; Mule, J.J.; Pilon-Thomas, S. Tumor-infiltrating lymphocytes: A new hope. Cancer Cell 2024, 42, 1315–1318. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Z.; Yang, Y.; Cui, J.; Sun, J.; Liu, Y. The prognostic and biology of tumour-infiltrating lymphocytes in the immunotherapy of cancer. Br. J. Cancer 2023, 129, 1041–1049. [Google Scholar] [CrossRef]
- Bied, M.; Ho, W.W.; Ginhoux, F.; Bleriot, C. Roles of macrophages in tumor development: a spatiotemporal perspective. Cell Mol. Immunol. 2023, 20, 983–992. [Google Scholar] [CrossRef]
- Gray, J.I.; Farber, D.L. Tissue-Resident Immune Cells in Humans. Annu. Rev. Immunol. 2022, 40, 195–220. [Google Scholar] [CrossRef]
- Tay, C.; Tanaka, A.; Sakaguchi, S. Tumor-infiltrating regulatory T cells as targets of cancer immunotherapy. Cancer Cell 2023, 41, 450–465. [Google Scholar] [CrossRef]
- van der Leun, A.M.; Traets, J.J.H.; Vos, J.L.; Elbers, J.B.W.; Patiwael, S.; Qiao, X.; Machuca-Ostos, M.; Thommen, D.S.; Haanen, J.; Schumacher, T.N.M.; et al. Dual Immune Checkpoint Blockade Induces Analogous Alterations in the Dysfunctional CD8+ T-cell and Activated Treg Compartment. Cancer Discov. 2023, 13, 2212–2227. [Google Scholar] [CrossRef]
- Gajewski, T.F.; Schreiber, H.; Fu, Y.X. Innate and adaptive immune cells in the tumor microenvironment. Nat. Immunol. 2013, 14, 1014–1022. [Google Scholar] [CrossRef]
- Neophytou, C.M.; Panagi, M.; Stylianopoulos, T.; Papageorgis, P. The Role of Tumor Microenvironment in Cancer Metastasis: Molecular Mechanisms and Therapeutic Opportunities. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Huppert, L.A.; Green, M.D.; Kim, L.; Chow, C.; Leyfman, Y.; Daud, A.I.; Lee, J.C. Tissue-specific Tregs in cancer metastasis: opportunities for precision immunotherapy. Cell Mol. Immunol. 2022, 19, 33–45. [Google Scholar] [CrossRef]
- Bejarano, L.; Jordao, M.J.C.; Joyce, J.A. Therapeutic Targeting of the Tumor Microenvironment. Cancer Discov. 2021, 11, 933–959. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Wang, A.; Yuan, Y.; Zhu, B.; Long, H. Heterogeneity of the tumor immune microenvironment and its clinical relevance. Exp. Hematol. Oncol. 2022, 11, 24. [Google Scholar] [CrossRef]
- Rudensky, A.Y. Regulatory T cells and Foxp3. Immunol. Rev. 2011, 241, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef]
- Fontenot, J.D.; Gavin, M.A.; Rudensky, A.Y. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat. Immunol. 2003, 4, 330–336. [Google Scholar] [CrossRef]
- Ziegler, S.F. FOXP3: of mice and men. Annu. Rev. Immunol. 2006, 24, 209–226. [Google Scholar] [CrossRef]
- Sato, Y. Forkhead Box Protein P3 in the Immune System. Allergies 2025, 5, 6. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Sakaguchi, N.; Asano, M.; Itoh, M.; Toda, M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol. 1995, 155, 1151–1164. [Google Scholar] [CrossRef]
- Liu, W.; Putnam, A.L.; Xu-Yu, Z.; Szot, G.L.; Lee, M.R.; Zhu, S.; Gottlieb, P.A.; Kapranov, P.; Gingeras, T.R.; Fazekas de St Groth, B.; et al. CD127 expression inversely correlates with FoxP3 and suppressive function of human CD4+ T reg cells. J. Exp. Med. 2006, 203, 1701–1711. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Perea, A.L.; Arcia, E.D.; Rueda, C.M.; Velilla, P.A. Phenotypical characterization of regulatory T cells in humans and rodents. Clin. Exp. Immunol. 2016, 185, 281–291. [Google Scholar] [CrossRef]
- Benamar, M.; Chen, Q.; Wang, M.; Chan, T.M.F.; Chatila, T.A. CPHEN-016: Comprehensive phenotyping of human regulatory T cells. Cytom. A 2022, 101, 1006–1011. [Google Scholar] [CrossRef]
- Collison, L.W.; Vignali, D.A. In vitro Treg suppression assays. Methods Mol. Biol. 2011, 707, 21–37. [Google Scholar] [CrossRef] [PubMed]
- McMurchy, A.N.; Levings, M.K. Suppression assays with human T regulatory cells: a technical guide. Eur. J. Immunol. 2012, 42, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Hoeppli, R.E.; Wu, D.; Cook, L.; Levings, M.K. The environment of regulatory T cell biology: cytokines, metabolites, and the microbiome. Front. Immunol. 2015, 6, 61. [Google Scholar] [CrossRef]
- Carbone, F.; Colamatteo, A.; La Rocca, C.; Lepore, M.T.; Russo, C.; De Rosa, G.; Matarese, A.; Procaccini, C.; Matarese, G. Metabolic Plasticity of Regulatory T Cells in Health and Autoimmunity. J. Immunol. 2024, 212, 1859–1866. [Google Scholar] [CrossRef] [PubMed]
- Colamatteo, A.; Carbone, F.; Bruzzaniti, S.; Galgani, M.; Fusco, C.; Maniscalco, G.T.; Di Rella, F.; de Candia, P.; De Rosa, V. Molecular Mechanisms Controlling Foxp3 Expression in Health and Autoimmunity: From Epigenetic to Post-translational Regulation. Front. Immunol. 2019, 10, 3136. [Google Scholar] [CrossRef] [PubMed]
- Qiu, R.; Zhou, L.; Ma, Y.; Zhou, L.; Liang, T.; Shi, L.; Long, J.; Yuan, D. Regulatory T Cell Plasticity and Stability and Autoimmune Diseases. Clin. Rev. Allergy Immunol. 2020, 58, 52–70. [Google Scholar] [CrossRef]
- Doganci, A.; Eigenbrod, T.; Krug, N.; De Sanctis, G.T.; Hausding, M.; Erpenbeck, V.J.; Haddad el, B.; Lehr, H.A.; Schmitt, E.; Bopp, T.; et al. The IL-6R alpha chain controls lung CD4+CD25+ Treg development and function during allergic airway inflammation in vivo. J. Clin. Invest. 2005, 115, 313–325. [Google Scholar] [CrossRef]
- Van Zeebroeck, L.; Arroyo Hornero, R.; Corte-Real, B.F.; Hamad, I.; Meissner, T.B.; Kleinewietfeld, M. Fast and Efficient Genome Editing of Human FOXP3(+) Regulatory T Cells. Front. Immunol. 2021, 12, 655122. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Mukasa, R.; Hatton, R.D.; Weaver, C.T. Developmental plasticity of Th17 and Treg cells. Curr. Opin. Immunol. 2009, 21, 274–280. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, X.; Wan, Y.Y. Intricacies of TGF-beta signaling in Treg and Th17 cell biology. Cell Mol. Immunol. 2023, 20, 1002–1022. [Google Scholar] [CrossRef] [PubMed]
- Kleinewietfeld, M.; Hafler, D.A. The plasticity of human Treg and Th17 cells and its role in autoimmunity. Semin. Immunol. 2013, 25, 305–312. [Google Scholar] [CrossRef]
- Zemmour, D.; Zilionis, R.; Kiner, E.; Klein, A.M.; Mathis, D.; Benoist, C. Single-cell gene expression reveals a landscape of regulatory T cell phenotypes shaped by the TCR. Nat. Immunol. 2018, 19, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Xu, C.; Wang, B.; Niu, Q.; Su, X.; Bai, Y.; Zhu, S.; Zhao, C.; Sun, Y.; Wang, J.; et al. Single-cell transcriptomic analysis reveals disparate effector differentiation pathways in human T(reg) compartment. Nat. Commun. 2021, 12, 3913. [Google Scholar] [CrossRef]
- Lui, P.P.; Cho, I.; Ali, N. Tissue regulatory T cells. Immunology 2020, 161, 4–17. [Google Scholar] [CrossRef]
- Kumar, B.V.; Ma, W.; Miron, M.; Granot, T.; Guyer, R.S.; Carpenter, D.J.; Senda, T.; Sun, X.; Ho, S.H.; Lerner, H.; et al. Human Tissue-Resident Memory T Cells Are Defined by Core Transcriptional and Functional Signatures in Lymphoid and Mucosal Sites. Cell Rep. 2017, 20, 2921–2934. [Google Scholar] [CrossRef]
- Pizzolla, A.; Nguyen, T.H.O.; Smith, J.M.; Brooks, A.G.; Kedzieska, K.; Heath, W.R.; Reading, P.C.; Wakim, L.M. Resident memory CD8(+) T cells in the upper respiratory tract prevent pulmonary influenza virus infection. Sci. Immunol. 2017, 2. [Google Scholar] [CrossRef]
- Zheng, M.Z.M.; Wakim, L.M. Tissue resident memory T cells in the respiratory tract. Mucosal Immunol. 2022, 15, 379–388. [Google Scholar] [CrossRef]
- Delacher, M.; Simon, M.; Sanderink, L.; Hotz-Wagenblatt, A.; Wuttke, M.; Schambeck, K.; Schmidleithner, L.; Bittner, S.; Pant, A.; Ritter, U.; et al. Single-cell chromatin accessibility landscape identifies tissue repair program in human regulatory T cells. Immunity 2021, 54, 702–720. [Google Scholar] [CrossRef] [PubMed]
- Hewavisenti, R.V.; Ferguson, A.L.; Gasparini, G.; Ohashi, T.; Braun, A.; Watkins, T.S.; Miles, J.J.; Elliott, M.; Sierro, F.; Feng, C.G.; et al. Tissue-resident regulatory T cells accumulate at human barrier lymphoid organs. Immunol. Cell Biol. 2021, 99, 894–906. [Google Scholar] [CrossRef]
- Burton, O.T.; Bricard, O.; Tareen, S.; Gergelits, V.; Andrews, S.; Biggins, L.; Roca, C.P.; Whyte, C.; Junius, S.; Brajic, A.; et al. The tissue-resident regulatory T cell pool is shaped by transient multi-tissue migration and a conserved residency program. Immunity 2024, 57, 1586–1602. [Google Scholar] [CrossRef] [PubMed]
- Olguin, J.E.; Medina-Andrade, I.; Rodriguez, T.; Rodriguez-Sosa, M.; Terrazas, L.I. Relevance of Regulatory T Cells during Colorectal Cancer Development. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef]
- Szeponik, L.; Ahlmanner, F.; Sundstrom, P.; Rodin, W.; Gustavsson, B.; Bexe Lindskog, E.; Wettergren, Y.; Quiding-Jarbrink, M. Intratumoral regulatory T cells from colon cancer patients comprise several activated effector populations. BMC Immunol. 2021, 22, 58. [Google Scholar] [CrossRef]
- Nie, Y.; He, J.; Shirota, H.; Trivett, A.L.; Yang, D.; Klinman, D.M.; Oppenheim, J.J.; Chen, X. Blockade of TNFR2 signaling enhances the immunotherapeutic effect of CpG ODN in a mouse model of colon cancer. Sci. Signal 2018, 11. [Google Scholar] [CrossRef]
- Pastille, E.; Bardini, K.; Fleissner, D.; Adamczyk, A.; Frede, A.; Wadwa, M.; von Smolinski, D.; Kasper, S.; Sparwasser, T.; Gruber, A.D.; et al. Transient ablation of regulatory T cells improves antitumor immunity in colitis-associated colon cancer. Cancer Res. 2014, 74, 4258–4269. [Google Scholar] [CrossRef] [PubMed]
- Aristin Revilla, S.; Kranenburg, O.; Coffer, P.J. Colorectal Cancer-Infiltrating Regulatory T Cells: Functional Heterogeneity, Metabolic Adaptation, and Therapeutic Targeting. Front. Immunol. 2022, 13, 903564. [Google Scholar] [CrossRef]
- Saito, T.; Nishikawa, H.; Wada, H.; Nagano, Y.; Sugiyama, D.; Atarashi, K.; Maeda, Y.; Hamaguchi, M.; Ohkura, N.; Sato, E.; et al. Two FOXP3(+)CD4(+) T cell subpopulations distinctly control the prognosis of colorectal cancers. Nat. Med. 2016, 22, 679–684. [Google Scholar] [CrossRef]
- Guan, T.; Zhou, X.; Zhou, W.; Lin, H. Regulatory T cell and macrophage crosstalk in acute lung injury: future perspectives. Cell Death Discov. 2023, 9, 9. [Google Scholar] [CrossRef]
- Jovisic, M.; Mambetsariev, N.; Singer, B.D.; Morales-Nebreda, L. Differential roles of regulatory T cells in acute respiratory infections. J. Clin. Invest. 2023, 133. [Google Scholar] [CrossRef]
- Liang, J.; Bi, G.; Shan, G.; Jin, X.; Bian, Y.; Wang, Q. Tumor-Associated Regulatory T Cells in Non-Small-Cell Lung Cancer: Current Advances and Future Perspectives. J. Immunol. Res. 2022, 2022, 4355386. [Google Scholar] [CrossRef]
- Dykema, A.G.; Zhang, J.; Cheung, L.S.; Connor, S.; Zhang, B.; Zeng, Z.; Cherry, C.M.; Li, T.; Caushi, J.X.; Nishimoto, M.; et al. Lung tumor-infiltrating T(reg) have divergent transcriptional profiles and function linked to checkpoint blockade response. Sci. Immunol. 2023, 8, eadg1487. [Google Scholar] [CrossRef]
- Shimizu, K.; Nakata, M.; Hirami, Y.; Yukawa, T.; Maeda, A.; Tanemoto, K. Tumor-infiltrating Foxp3+ regulatory T cells are correlated with cyclooxygenase-2 expression and are associated with recurrence in resected non-small cell lung cancer. J. Thorac. Oncol. 2010, 5, 585–590. [Google Scholar] [CrossRef]
- Chen, P.; Wang, H.; Tang, Z.; Shi, J.; Cheng, L.; Zhao, C.; Li, X.; Zhou, C. Selective depletion of CCR8+Treg cells enhances anti-tumor immunity of cytotoxic T cells in lung cancer via dendritic cells. J. Thorac. Oncol. 2025. [Google Scholar] [CrossRef] [PubMed]
- DeNardo, D.G.; Coussens, L.M. Inflammation and breast cancer. Balancing immune response: crosstalk between adaptive and innate immune cells during breast cancer progression. Breast Cancer Res. 2007, 9, 212. [Google Scholar] [CrossRef] [PubMed]
- Plitas, G.; Konopacki, C.; Wu, K.; Bos, P.D.; Morrow, M.; Putintseva, E.V.; Chudakov, D.M.; Rudensky, A.Y. Regulatory T Cells Exhibit Distinct Features in Human Breast Cancer. Immunity 2016, 45, 1122–1134. [Google Scholar] [CrossRef]
- Debien, V.; De Caluwe, A.; Wang, X.; Piccart-Gebhart, M.; Tuohy, V.K.; Romano, E.; Buisseret, L. Immunotherapy in breast cancer: an overview of current strategies and perspectives. NPJ Breast Cancer 2023, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Kos, K.; de Visser, K.E. The Multifaceted Role of Regulatory T Cells in Breast Cancer. Annu. Rev. Cancer Biol. 2021, 5, 291–310. [Google Scholar] [CrossRef]
- Wang, L.; Simons, D.L.; Lu, X.; Tu, T.Y.; Solomon, S.; Wang, R.; Rosario, A.; Avalos, C.; Schmolze, D.; Yim, J.; et al. Connecting blood and intratumoral T(reg) cell activity in predicting future relapse in breast cancer. Nat. Immunol. 2019, 20, 1220–1230. [Google Scholar] [CrossRef]
- Bos, P.D.; Plitas, G.; Rudra, D.; Lee, S.Y.; Rudensky, A.Y. Transient regulatory T cell ablation deters oncogene-driven breast cancer and enhances radiotherapy. J. Exp. Med. 2013, 210, 2435–2466. [Google Scholar] [CrossRef]
- Clark, N.M.; Martinez, L.M.; Murdock, S.; deLigio, J.T.; Olex, A.L.; Effi, C.; Dozmorov, M.G.; Bos, P.D. Regulatory T Cells Support Breast Cancer Progression by Opposing IFN-gamma-Dependent Functional Reprogramming of Myeloid Cells. Cell Rep. 2020, 33, 108482. [Google Scholar] [CrossRef]
- Zheng, L.; Xue, J.; Jaffee, E.M.; Habtezion, A. Role of immune cells and immune-based therapies in pancreatitis and pancreatic ductal adenocarcinoma. Gastroenterology 2013, 144, 1230–1240. [Google Scholar] [CrossRef]
- Pugliese, A. Autoreactive T cells in type 1 diabetes. J. Clin. Invest. 2017, 127, 2881–2891. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, A.; Socci, C.; Stabilini, A.; Valle, A.; Monti, P.; Piemonti, L.; Nano, R.; Olek, S.; Maffi, P.; Scavini, M.; et al. Expansion of Th17 cells and functional defects in T regulatory cells are key features of the pancreatic lymph nodes in patients with type 1 diabetes. Diabetes 2011, 60, 2903–2913. [Google Scholar] [CrossRef]
- Hull, C.M.; Peakman, M.; Tree, T.I.M. Regulatory T cell dysfunction in type 1 diabetes: what’s broken and how can we fix it? Diabetologia 2017, 60, 1839–1850. [Google Scholar] [CrossRef] [PubMed]
- Tonkin, D.R.; Haskins, K. Regulatory T cells enter the pancreas during suppression of type 1 diabetes and inhibit effector T cells and macrophages in a TGF-beta-dependent manner. Eur. J. Immunol. 2009, 39, 1313–1322. [Google Scholar] [CrossRef] [PubMed]
- Bluestone, J.A.; Buckner, J.H.; Fitch, M.; Gitelman, S.E.; Gupta, S.; Hellerstein, M.K.; Herold, K.C.; Lares, A.; Lee, M.R.; Li, K.; et al. Type 1 diabetes immunotherapy using polyclonal regulatory T cells. Sci. Transl. Med. 2015, 7, 315ra189. [Google Scholar] [CrossRef]
- Ullman, N.A.; Burchard, P.R.; Dunne, R.F.; Linehan, D.C. Immunologic Strategies in Pancreatic Cancer: Making Cold Tumors Hot. J. Clin. Oncol. 2022, 40, 2789–2805. [Google Scholar] [CrossRef]
- Truong, L.H.; Pauklin, S. Pancreatic Cancer Microenvironment and Cellular Composition: Current Understandings and Therapeutic Approaches. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Ho, W.J.; Jaffee, E.M.; Zheng, L. The tumour microenvironment in pancreatic cancer—clinical challenges and opportunities. Nat. Rev. Clin. Oncol. 2020, 17, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Mota Reyes, C.; Demir, E.; Cifcibasi, K.; Istvanffy, R.; Friess, H.; Demir, I.E. Regulatory T Cells in Pancreatic Cancer: Of Mice and Men. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Zhang, Y.; Lazarus, J.; Steele, N.G.; Yan, W.; Lee, H.J.; Nwosu, Z.C.; Halbrook, C.J.; Menjivar, R.E.; Kemp, S.B.; Sirihorachai, V.R.; et al. Regulatory T-cell Depletion Alters the Tumor Microenvironment and Accelerates Pancreatic Carcinogenesis. Cancer Discov. 2020, 10, 422–439. [Google Scholar] [CrossRef] [PubMed]
- Farhangnia, P.; Khorramdelazad, H.; Nickho, H.; Delbandi, A.A. Current and future immunotherapeutic approaches in pancreatic cancer treatment. J. Hematol. Oncol. 2024, 17, 40. [Google Scholar] [CrossRef]
- Robinson, M.W.; Harmon, C.; O’Farrelly, C. Liver immunology and its role in inflammation and homeostasis. Cell Mol. Immunol. 2016, 13, 267–276. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, S.; Liu, Y.; He, X.; Qu, M.; Xu, G.; Wang, H.; Huang, M.; Pan, J.; Liu, Z.; et al. Single-cell RNA sequencing reveals the heterogeneity of liver-resident immune cells in human. Cell Discov. 2020, 6, 22. [Google Scholar] [CrossRef]
- Terziroli Beretta-Piccoli, B.; Mieli-Vergani, G.; Vergani, D. Autoimmmune hepatitis. Cell Mol. Immunol. 2022, 19, 158–176. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.C.; Sung, P.S.; Park, S.H. Immune responses and immunopathology in acute and chronic viral hepatitis. Nat. Rev. Immunol. 2016, 16, 509–523. [Google Scholar] [CrossRef]
- Wu, K.J.; Qian, Q.F.; Zhou, J.R.; Sun, D.L.; Duan, Y.F.; Zhu, X.; Sartorius, K.; Lu, Y.J. Regulatory T cells (Tregs) in liver fibrosis. Cell Death Discov. 2023, 9, 53. [Google Scholar] [CrossRef]
- Sas, Z.; Cendrowicz, E.; Weinhauser, I.; Rygiel, T.P. Tumor Microenvironment of Hepatocellular Carcinoma: Challenges and Opportunities for New Treatment Options. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Yin, Y.; Feng, W.; Chen, J.; Chen, X.; Wang, G.; Wang, S.; Xu, X.; Nie, Y.; Fan, D.; Wu, K.; et al. Immunosuppressive tumor microenvironment in the progression, metastasis, and therapy of hepatocellular carcinoma: from bench to bedside. Exp. Hematol. Oncol. 2024, 13, 72. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Li, Q.; Wang, Y. Impact of regulatory T cells on the prognosis of hepatocellular carcinoma: A protocol for systematic review and meta analysis. Med. (Baltim. ) 2021, 100, e23957. [Google Scholar] [CrossRef]
- Jung, M.K.; Shin, E.C. Regulatory T Cells in Hepatitis B and C Virus Infections. Immune Netw. 2016, 16, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Castellani, G.; Croese, T.; Peralta Ramos, J.M.; Schwartz, M. Transforming the understanding of brain immunity. Science 2023, 380, eabo7649. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Komai, K.; Mise-Omata, S.; Iizuka-Koga, M.; Noguchi, Y.; Kondo, T.; Sakai, R.; Matsuo, K.; Nakayama, T.; Yoshie, O.; et al. Brain regulatory T cells suppress astrogliosis and potentiate neurological recovery. Nature 2019, 565, 246–250. [Google Scholar] [CrossRef]
- Jacobs, J.F.; Idema, A.J.; Bol, K.F.; Grotenhuis, J.A.; de Vries, I.J.; Wesseling, P.; Adema, G.J. Prognostic significance and mechanism of Treg infiltration in human brain tumors. J. Neuroimmunol. 2010, 225, 195–199. [Google Scholar] [CrossRef]
- Jacobs, J.F.; Idema, A.J.; Bol, K.F.; Nierkens, S.; Grauer, O.M.; Wesseling, P.; Grotenhuis, J.A.; Hoogerbrugge, P.M.; de Vries, I.J.; Adema, G.J. Regulatory T cells and the PD-L1/PD-1 pathway mediate immune suppression in malignant human brain tumors. Neuro Oncol. 2009, 11, 394–402. [Google Scholar] [CrossRef]
- Wainwright, D.A.; Dey, M.; Chang, A.; Lesniak, M.S. Targeting Tregs in Malignant Brain Cancer: Overcoming IDO. Front. Immunol. 2013, 4, 116. [Google Scholar] [CrossRef]
- Amoozgar, Z.; Kloepper, J.; Ren, J.; Tay, R.E.; Kazer, S.W.; Kiner, E.; Krishnan, S.; Posada, J.M.; Ghosh, M.; Mamessier, E.; et al. Targeting Treg cells with GITR activation alleviates resistance to immunotherapy in murine glioblastomas. Nat. Commun. 2021, 12, 2582. [Google Scholar] [CrossRef]
- Park, S.; Matte-Martone, C.; Gonzalez, D.G.; Lathrop, E.A.; May, D.P.; Pineda, C.M.; Moore, J.L.; Boucher, J.D.; Marsh, E.; Schmitter-Sanchez, A.; et al. Skin-resident immune cells actively coordinate their distribution with epidermal cells during homeostasis. Nat. Cell Biol. 2021, 23, 476–484. [Google Scholar] [CrossRef]
- Ali, N.; Rosenblum, M.D. Regulatory T cells in skin. Immunology 2017, 152, 372–381. [Google Scholar] [CrossRef]
- Tong, X.; Kim, S.H.; Che, L.; Park, J.; Lee, J.; Kim, T.G. Foxp3(+) Treg control allergic skin inflammation by restricting IFN-gamma-driven neutrophilic infiltration and NETosis. J. Dermatol. Sci. 2024, 115, 2–12. [Google Scholar] [CrossRef]
- Falcone, I.; Conciatori, F.; Bazzichetto, C.; Ferretti, G.; Cognetti, F.; Ciuffreda, L.; Milella, M. Tumor Microenvironment: Implications in Melanoma Resistance to Targeted Therapy and Immunotherapy. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Guo, Y.; Liu, S.; Wang, H.; Zhu, J.; Ou, L.; Xu, X. Targeting regulatory T cells for immunotherapy in melanoma. Mol. Biomed. 2021, 2, 11. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.C.; Zappasodi, R. A decade of checkpoint blockade immunotherapy in melanoma: understanding the molecular basis for immune sensitivity and resistance. Nat. Immunol. 2022, 23, 660–670. [Google Scholar] [CrossRef] [PubMed]
- Geels, S.N.; Moshensky, A.; Sousa, R.S.; Murat, C.; Bustos, M.A.; Walker, B.L.; Singh, R.; Harbour, S.N.; Gutierrez, G.; Hwang, M.; et al. Interruption of the intratumor CD8(+) T cell:Treg crosstalk improves the efficacy of PD-1 immunotherapy. Cancer Cell 2024, 42, 1051–1066. [Google Scholar] [CrossRef]
- Rousseau, M.; Lacerda Mariano, L.; Canton, T.; Ingersoll, M.A. Tissue-resident memory T cells mediate mucosal immunity to recurrent urinary tract infection. Sci. Immunol. 2023, 8, eabn4332. [Google Scholar] [CrossRef]
- Heidegger, I.; Pircher, A.; Pichler, R. Targeting the Tumor Microenvironment in Renal Cell Cancer Biology and Therapy. Front. Oncol. 2019, 9, 490. [Google Scholar] [CrossRef]
- Hatogai, K.; Sweis, R.F. The Tumor Microenvironment of Bladder Cancer. Adv. Exp. Med. Biol. 2020, 1296, 275–290. [Google Scholar] [CrossRef]
- Koll, F.J.; Banek, S.; Kluth, L.; Kollermann, J.; Bankov, K.; Chun, F.K.; Wild, P.J.; Weigert, A.; Reis, H. Tumor-associated macrophages and Tregs influence and represent immune cell infiltration of muscle-invasive bladder cancer and predict prognosis. J. Transl. Med. 2023, 21, 124. [Google Scholar] [CrossRef]
- Nair, S.S.; Weil, R.; Dovey, Z.; Davis, A.; Tewari, A.K. The Tumor Microenvironment and Immunotherapy in Prostate and Bladder Cancer. Urol. Clin. North. Am. 2020, 47, e17–e54. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Murakami, K.; Inoue, A.; Yonezawa, T.; Matsuki, N. CCR4 Blockade Depletes Regulatory T Cells and Prolongs Survival in a Canine Model of Bladder Cancer. Cancer Immunol. Res. 2019, 7, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Santagata, S.; Rea, G.; Bello, A.M.; Capiluongo, A.; Napolitano, M.; Desicato, S.; Fragale, A.; D’Alterio, C.; Trotta, A.M.; Ierano, C.; et al. Targeting CXCR4 impaired T regulatory function through PTEN in renal cancer patients. Br. J. Cancer 2024, 130, 2016–2026. [Google Scholar] [CrossRef]
- Yuzen, D.; Arck, P.C.; Thiele, K. Tissue-resident immunity in the female and male reproductive tract. Semin. Immunopathol. 2022, 44, 785–799. [Google Scholar] [CrossRef]
- Mor, G.; Cardenas, I. The immune system in pregnancy: a unique complexity. Am. J. Reprod. Immunol. 2010, 63, 425–433. [Google Scholar] [CrossRef]
- Taneja, V. Sex Hormones Determine Immune Response. Front. Immunol. 2018, 9, 1931. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, Y.; Yang, J.; Zhao, X.; Wei, X. Tumor Microenvironment in Ovarian Cancer: Function and Therapeutic Strategy. Front. Cell Dev. Biol. 2020, 8, 758. [Google Scholar] [CrossRef]
- Garg, P.; Ramisetty, S.K.; Raghu Subbalakshmi, A.; Krishna, B.M.; Pareek, S.; Mohanty, A.; Kulkarni, P.; Horne, D.; Salgia, R.; Singhal, S.S. Gynecological cancer tumor Microenvironment: Unveiling cellular complexity and therapeutic potential. Biochem. Pharmacol. 2024, 229, 116498. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ke, X.; Zeng, S.; Wu, M.; Lou, J.; Wu, L.; Huang, P.; Huang, L.; Wang, F.; Pan, S. Analysis of CD8+ Treg cells in patients with ovarian cancer: a possible mechanism for immune impairment. Cell Mol. Immunol. 2015, 12, 580–591. [Google Scholar] [CrossRef]
- Lynam, S.; Lugade, A.A.; Odunsi, K. Immunotherapy for Gynecologic Cancer: Current Applications and Future Directions. Clin. Obs. Obstet. Gynecol. 2020, 63, 48–63. [Google Scholar] [CrossRef]
- Mandal, R.; Senbabaoglu, Y.; Desrichard, A.; Havel, J.J.; Dalin, M.G.; Riaz, N.; Lee, K.W.; Ganly, I.; Hakimi, A.A.; Chan, T.A.; et al. The head and neck cancer immune landscape and its immunotherapeutic implications. JCI Insight 2016, 1, e89829. [Google Scholar] [CrossRef]
- Fasano, M.; Corte, C.M.D.; Liello, R.D.; Viscardi, G.; Sparano, F.; Iacovino, M.L.; Paragliola, F.; Piccolo, A.; Napolitano, S.; Martini, G.; et al. Immunotherapy for head and neck cancer: Present and future. Crit. Rev. Oncol. Hematol. 2022, 174, 103679. [Google Scholar] [CrossRef]
- Seminerio, I.; Descamps, G.; Dupont, S.; de Marrez, L.; Laigle, J.A.; Lechien, J.R.; Kindt, N.; Journe, F.; Saussez, S. Infiltration of FoxP3+ Regulatory T Cells is a Strong and Independent Prognostic Factor in Head and Neck Squamous Cell Carcinoma. Cancers (Basel) 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Ihara, F.; Sakurai, D.; Horinaka, A.; Makita, Y.; Fujikawa, A.; Sakurai, T.; Yamasaki, K.; Kunii, N.; Motohashi, S.; Nakayama, T.; et al. CD45RA(-)Foxp3(high) regulatory T cells have a negative impact on the clinical outcome of head and neck squamous cell carcinoma. Cancer Immunol. Immunother. 2017, 66, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Smolle, M.A.; Herbsthofer, L.; Granegger, B.; Goda, M.; Brcic, I.; Bergovec, M.; Scheipl, S.; Prietl, B.; Pichler, M.; Gerger, A.; et al. T-regulatory cells predict clinical outcome in soft tissue sarcoma patients: a clinico-pathological study. Br. J. Cancer 2021, 125, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Fazel, M.; Dufresne, A.; Vanacker, H.; Waissi, W.; Blay, J.Y.; Brahmi, M. Immunotherapy for Soft Tissue Sarcomas: Anti-PD1/PDL1 and Beyond. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef]
- Almeida, J.S.; Sousa, L.M.; Couceiro, P.; Andrade, T.F.; Alves, V.; Martinho, A.; Rodrigues, J.; Fonseca, R.; Freitas-Tavares, P.; Santos-Rosa, M.; et al. Peripheral immune profiling of soft tissue sarcoma: perspectives for disease monitoring. Front. Immunol. 2024, 15, 1391840. [Google Scholar] [CrossRef]
- Garcia-Dominguez, D.J.; Hontecillas-Prieto, L.; Palazon-Carrion, N.; Jimenez-Cortegana, C.; Sanchez-Margalet, V.; de la Cruz-Merino, L. Tumor Immune Microenvironment in Lymphoma: Focus on Epigenetics. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Scott, D.W.; Gascoyne, R.D. The tumour microenvironment in B cell lymphomas. Nat. Rev. Cancer 2014, 14, 517–534. [Google Scholar] [CrossRef]
- Ustun, C.; Miller, J.S.; Munn, D.H.; Weisdorf, D.J.; Blazar, B.R. Regulatory T cells in acute myelogenous leukemia: is it time for immunomodulation? Blood 2011, 118, 5084–5095. [Google Scholar] [CrossRef]
- Xu, Y.; Mou, J.; Wang, Y.; Zhou, W.; Rao, Q.; Xing, H.; Tian, Z.; Tang, K.; Wang, M.; Wang, J. Regulatory T cells promote the stemness of leukemia stem cells through IL10 cytokine-related signaling pathway. Leukemia 2022, 36, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Mutis, T.; van Rijn, R.S.; Simonetti, E.R.; Aarts-Riemens, T.; Emmelot, M.E.; van Bloois, L.; Martens, A.; Verdonck, L.F.; Ebeling, S.B. Human regulatory T cells control xenogeneic graft-versus-host disease induced by autologous T cells in RAG2-/-gammac-/- immunodeficient mice. Clin. Cancer Res. 2006, 12, 5520–5525. [Google Scholar] [CrossRef]
- Vinado, A.C.; Calvo, I.A.; Cenzano, I.; Olaverri, D.; Cocera, M.; San Martin-Uriz, P.; Romero, J.P.; Vilas-Zornoza, A.; Vera, L.; Gomez-Cebrian, N.; et al. The bone marrow niche regulates redox and energy balance in MLL::AF9 leukemia stem cells. Leukemia 2022, 36, 1969–1979. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, K.; Zhang, H. Targeting Leukemia-Initiating Cells and Leukemic Niches: The Next Therapy Station for T-Cell Acute Lymphoblastic Leukemia? Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Liao, W.; Liu, C.; Yang, K.; Chen, J.; Wu, Y.; Zhang, S.; Yu, K.; Wang, L.; Ran, L.; Chen, M.; et al. Aged hematopoietic stem cells entrap regulatory T cells to create a prosurvival microenvironment. Cell Mol. Immunol. 2023, 20, 1216–1231. [Google Scholar] [CrossRef] [PubMed]
- Riether, C. Regulation of hematopoietic and leukemia stem cells by regulatory T cells. Front. Immunol. 2022, 13, 1049301. [Google Scholar] [CrossRef]
- Shan, F.; Somasundaram, A.; Bruno, T.C.; Workman, C.J.; Vignali, D.A.A. Therapeutic targeting of regulatory T cells in cancer. Trends Cancer 2022, 8, 944–961. [Google Scholar] [CrossRef]
- Nishikawa, H.; Koyama, S. Mechanisms of regulatory T cell infiltration in tumors: implications for innovative immune precision therapies. J. Immunother. Cancer 2021, 9. [Google Scholar] [CrossRef]
- Yang, J.; Bae, H. Drug conjugates for targeting regulatory T cells in the tumor microenvironment: guided missiles for cancer treatment. Exp. Mol. Med. 2023, 55, 1996–2004. [Google Scholar] [CrossRef]
- Fu, Z.; Li, S.; Han, S.; Shi, C.; Zhang, Y. Antibody drug conjugate: the “biological missile” for targeted cancer therapy. Signal Transduct. Target. Ther. 2022, 7, 93. [Google Scholar] [CrossRef]
- Veneziani, A.C.; Sneha, S.; Oza, A.M. Antibody-Drug Conjugates: Advancing from Magic Bullet to Biological Missile. Clin. Cancer Res. 2024, 30, 1434–1437. [Google Scholar] [CrossRef] [PubMed]
- Zammarchi, F.; Havenith, K.; Bertelli, F.; Vijayakrishnan, B.; Chivers, S.; van Berkel, P.H. CD25-targeted antibody-drug conjugate depletes regulatory T cells and eliminates established syngeneic tumors via antitumor immunity. J. Immunother. Cancer 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Litzinger, M.T.; Fernando, R.; Curiel, T.J.; Grosenbach, D.W.; Schlom, J.; Palena, C. IL-2 immunotoxin denileukin diftitox reduces regulatory T cells and enhances vaccine-mediated T-cell immunity. Blood 2007, 110, 3192–3201. [Google Scholar] [CrossRef] [PubMed]
- Onda, M.; Kobayashi, K.; Pastan, I. Depletion of regulatory T cells in tumors with an anti-CD25 immunotoxin induces CD8 T cell-mediated systemic antitumor immunity. Proc. Natl. Acad. Sci. U S A 2019, 116, 4575–4582. [Google Scholar] [CrossRef]
- Lozano, T.; Soldevilla, M.M.; Casares, N.; Villanueva, H.; Bendandi, M.; Lasarte, J.J.; Pastor, F. Targeting inhibition of Foxp3 by a CD28 2′-Fluro oligonucleotide aptamer conjugated to P60-peptide enhances active cancer immunotherapy. Biomaterials 2016, 91, 73–80. [Google Scholar] [CrossRef]
- Serrano, A.; Casares, N.; Troconiz, I.F.; Lozano, T.; Lasarte, J.J.; Zalba, S.; Garrido, M.J. Foxp3 inhibitory peptide encapsulated in a novel CD25-targeted nanoliposome promotes efficient tumor regression in mice. Acta Pharmacol. Sin. 2025, 46, 171–183. [Google Scholar] [CrossRef]
Figure 1.
Treg phenotype, plasticity, heterogeneity and tissue residency.
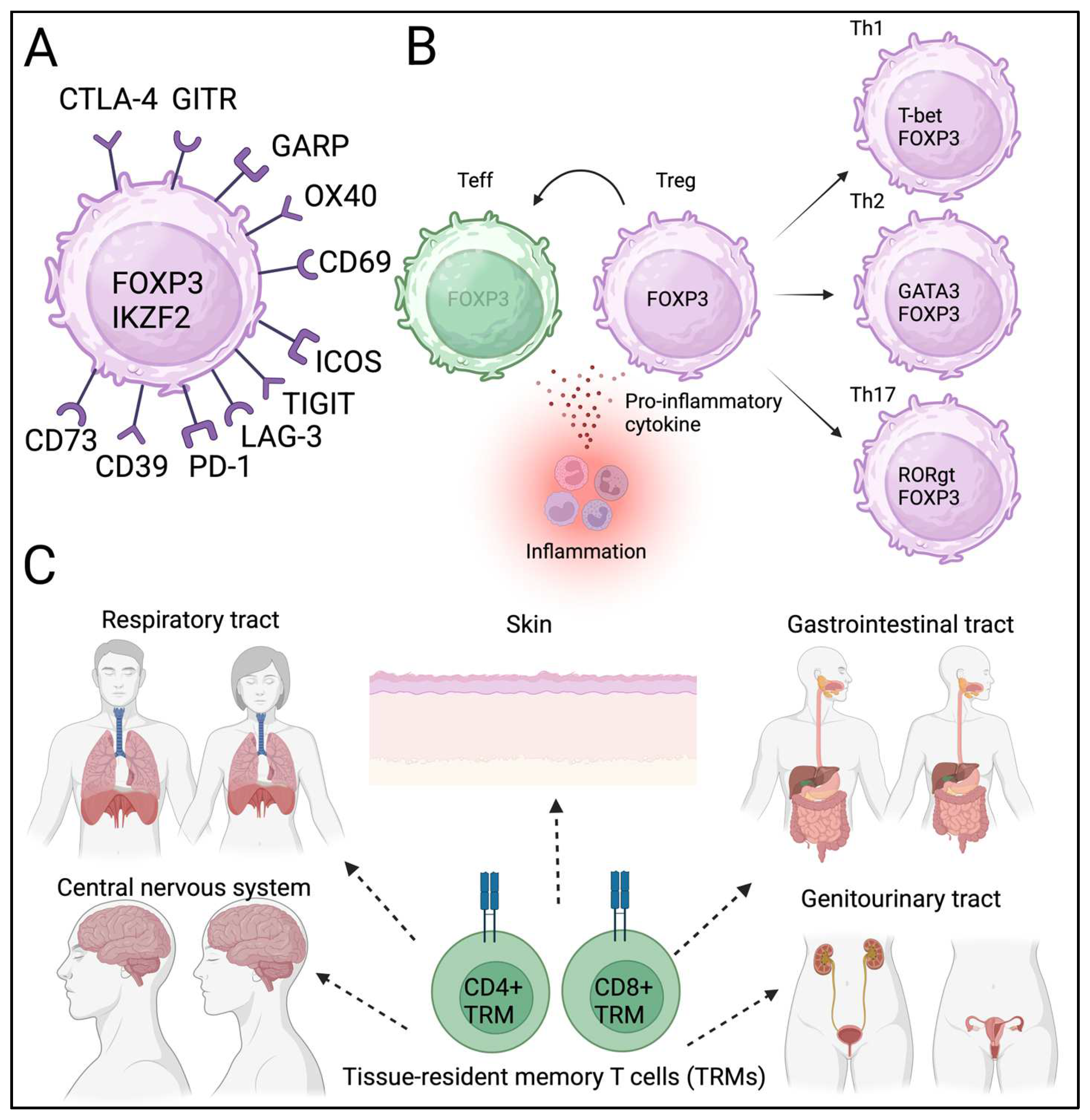
Figure 2.
Tregs can be detected in TME of colon cancer.
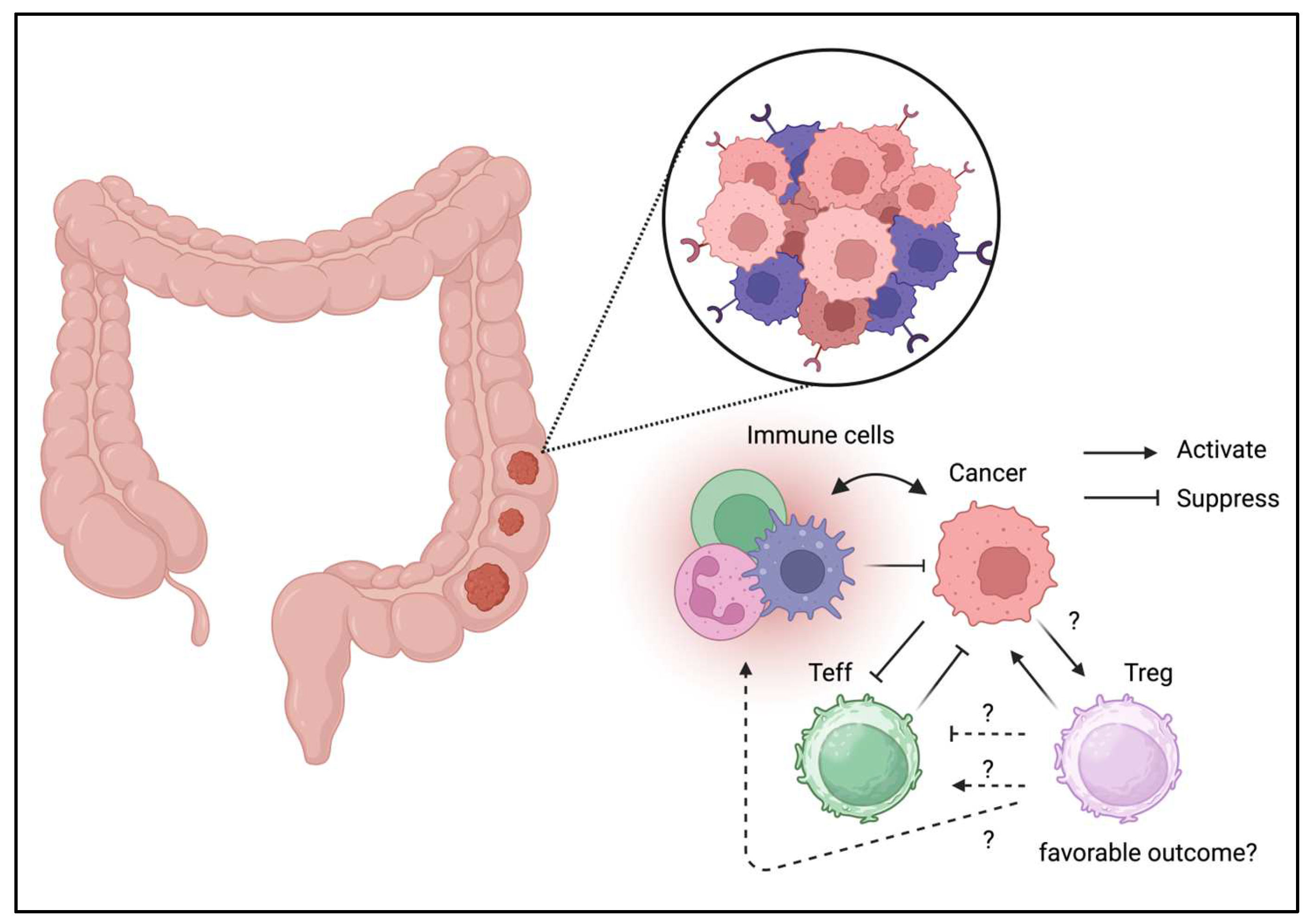
Figure 3.
Tregs can be detected in TME of lung cancer.
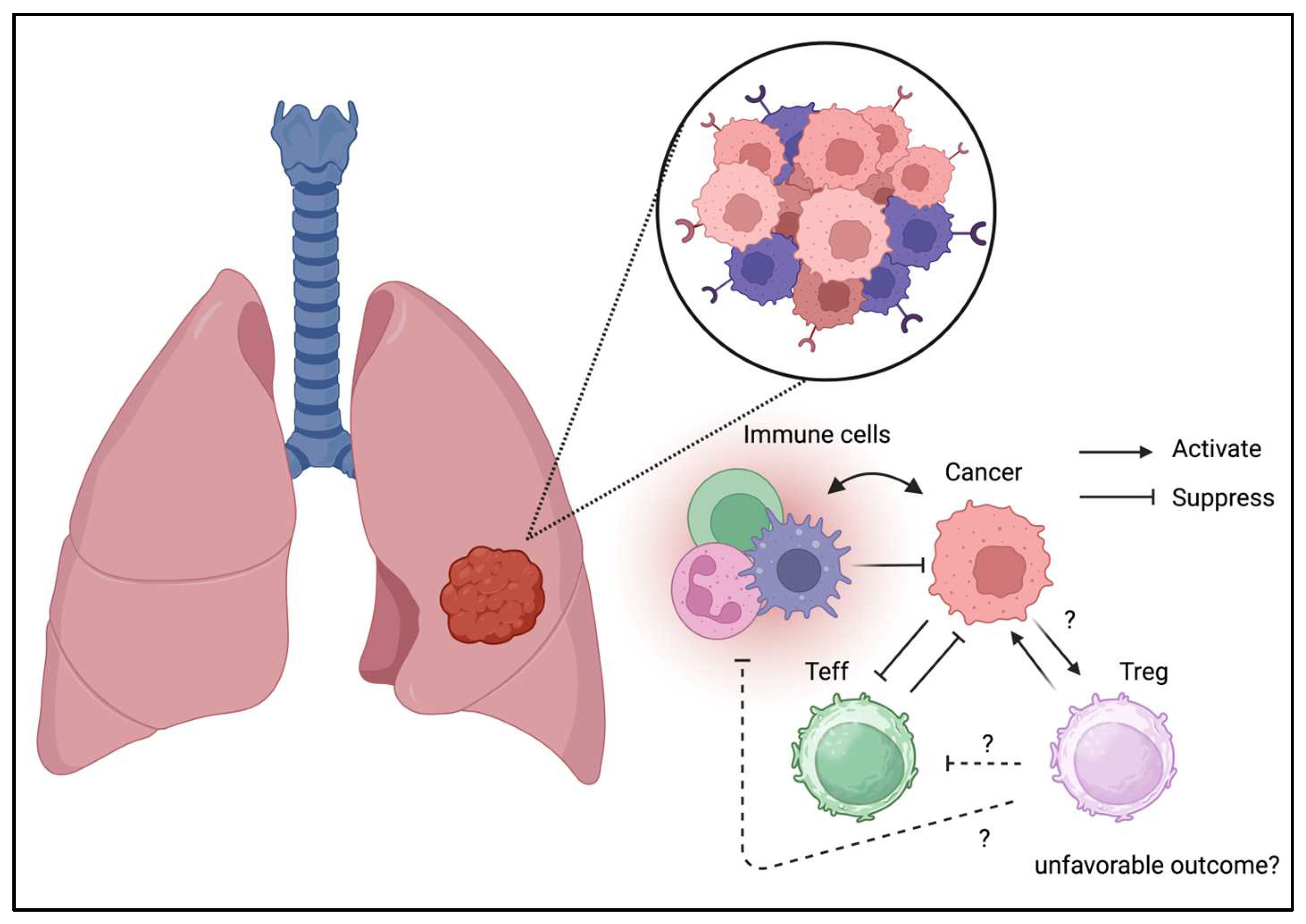
Figure 4.
Tregs can be detected in TME of breast cancer.
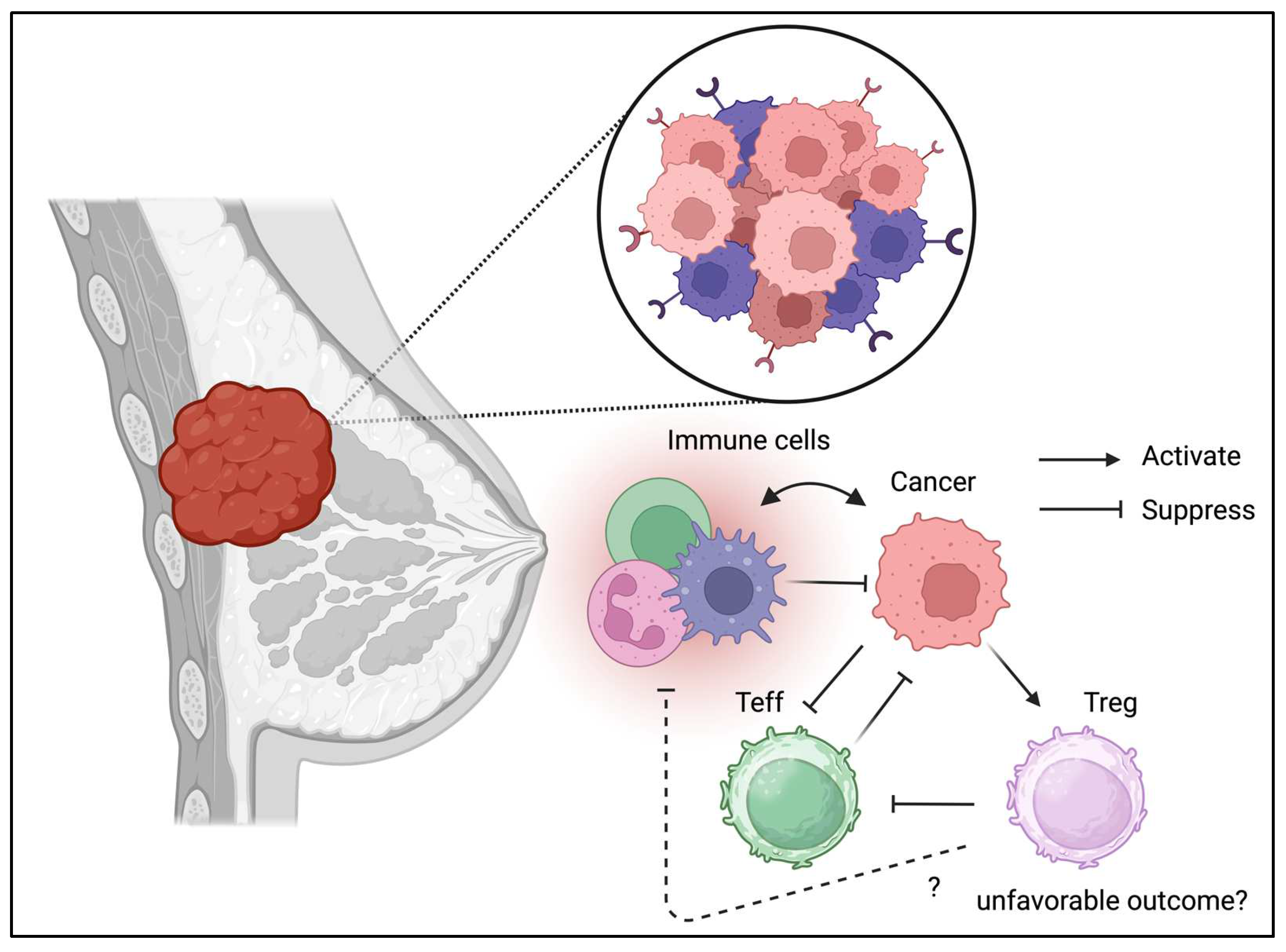
Figure 5.
Tregs can be detected in TME of pancreatic cancer.
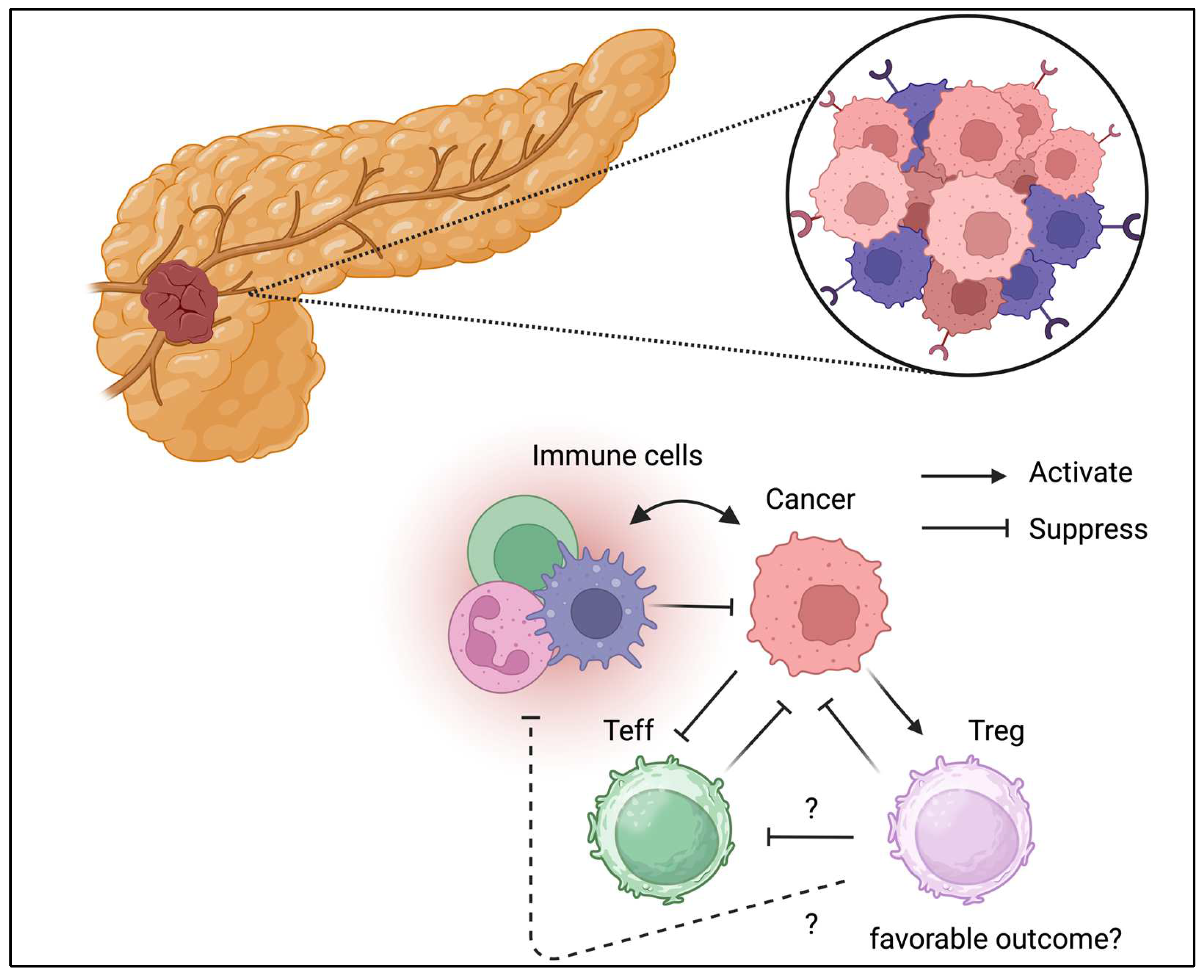
Figure 6.
Tregs can be detected in TME of brain tumor.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



