Submitted:
30 April 2025
Posted:
02 May 2025
You are already at the latest version
Abstract
The Irish potato (Solanum tuberosum) is an important food and cash crop in many countries worldwide. However, it is susceptible to infection by many diseases, including Fusarium wilt and root knot nematodes, leading to severe yield losses. The use of synthetic chemicals to control these diseases is potentially harmful to consumers and the environment. Studies to evaluate the innovative and sustainable use of three Trichoderma isolates (T28, T77 and Dam 3) in controlling the Fusarium wilt – root knot nematode disease complex and promoting potato plant growth were carried out under laboratory and field conditions. After sequencing, the Trichoderma isolates were identified as Trichoderma asperellum, Trichoderma spp and Trichoderma asperolloides for T28, T77 and Dam 3, respectively. In the laboratory studies, the T77 isolate was most effective in inhibiting F. oxysporum growth (61.33%), suppressing nematode egg hatching and inducing juvenile mortality (96%) while the Dam 3 isolate was least effective. In the field studies, the T28+T77 isolate significantly increased potato leaf area index by 74.92%, and caused the least Fusarium wilt disease incidence and galling at both trial sites. The T77 isolate significantly increased plant height (33%), chlorophyll content (10.1%), and potato yield (>100%) when compared to the control treatment. The isolates can either be seed or furrow-applied. The study showed that T77 and T28+T77 isolates are effective in suppressing the development of Fusarium wilt and root knot nematodes and they also promote potato plant growth. Being biocontrol agents, it is recommended that the two isolates be incorporated into integrated disease management programmes for the sustainable control of the Fusarium wilt - root knot nematode disease complex in potato production.
Keywords:
Trichoderma
; Fusarium wilt
; root knot nematodes
; disease complex
; disease control
; disease incidence
; disease severity
; plant growth promotion
1. Introduction
The Irish potato, Solanum tuberosum L., is the fourth most-produced crop in the world and an important staple in many countries [1]. Its tubers are an excellent source of vitamins, proteins and carbohydrates [2]. The potato is prone to infection by pathogens during all stages of its growth [3,4,5], resulting in considerable losses in yield and quality. Amongst the pathogens are the ascomycetous fungi belonging to the genus Fusarium and root knot nematodes (Meloidogyne spp). Potato yield losses due to Fusarium spp infection are estimated at 10 - 53% [6] while Meloidogyne spp infection cause losses of up to 100%, depending on the season and the level of field infestation [7].
Fusarium spp and Meloidogyne spp are capable of co-infecting crops resulting in the development of a Fusarium wilt-root knot nematode (FW-RKN) disease complex [8,9]. This concomitant infection usually results in greater damage to the host plant and more disease severity than when the pathogens infect a host singly [10]. Infected plants display a wide range of symptoms that include leaf chlorosis, wilting, retarded growth and ultimately, plant death. The FW-RKN disease complex has been reported infecting potatoes in Zimbabwe [11] but its impact on yield has not yet been quantified.
The management of Fusarium wilt and root knot nematodes can be achieved by employing cultural, biological, host resistance and chemical control tactics. However, these control tactics have some limitations. For instance, the use of fumigants like metam sodium, methyl bromide and ethylene dibromide, is detrimental to the environment and human health [12]. Also, both root knot nematodes and Fusarium spp can develop resistance to synthetic chemicals [13,14]. Host resistance, though environmentally-friendly, is neither durable nor absolute as it tends to break down when disease pressure is high or environmental conditions are unfavourable for crop growth. Cultural practices such as crop rotation and fallowing are proving difficult to implement due to limited land availability, the polyphagous natures of both pathogens and their abilities to survive for long periods in the absence of their hosts [15]. Thus, there is a need to develop alternative management strategies that are economical, environmentally-friendly and sustainable in those cropping systems where the disease complex is present.
Biological control (or biocontrol) is a safer alternative disease management strategy to the use of synthetic chemicals [16]. It involves the harnessing and utilization of such microbes as bacteria, fungi, viruses, nematodes and actinomycetes to control or suppress pathogenic microbes. The biocontrol agents employ a diverse range of mechanisms to protect plants against pathogenic invasion. For example, they compete for space and resources with the pathogens [17], disrupt the pathogens’ quorum sensing by inhibiting the production of signal molecules that launch infections [18] and/or trigger a defence response in the host. Some biocontrol agents can promote plant growth by enhancing water and nutrient uptake [19,20]. Several fungal species have biocontrol properties including Chaetomium globosum [21], Glomus spp [22], Paecilomyces lilacinus [23], Pochonia chlamydosporia [24], and Trichoderma spp [25,26].
Trichoderma spp are common soilborne fungi that are naturally present in many soil types including cultivated, forest, fallow and pastureland soils in temperate and tropical environments [27]. They are characterised by rapid growth that produces large amounts of conidia whose pigmentation can vary from dark to light green [28]. As biocontrol agents, Trichoderma spp induce antibiosis, mycoparasitism and also stimulate defences against diseases [29]. They also compete with other microbes for nutrients, space and key exudates from seeds that stimulate the germination of propagules of plant pathogenic fungi. Furthermore, Trichoderma spp inhibit and/or degrade enzymes that are essential for the activities of plant pathogenic fungi [30]. According to Sahebani and Hadavi [31], the inoculation of greenhouse-grown tomato seeds with Trichoderma significantly reduces the level of disease caused by Meloidogyne javanica by affecting nematode establishment, development and reproduction. Similarly, tomato root colonisation by T. harzianum impedes nematode’s ability to invade, parasitize, reproduce and induce galling in the host [32].
Trichoderma spp are also plant growth-promoting fungi. They produce growth factors that enhance the rates of seed germination, plant growth and yield. According to [30], T. harzianum and T. koningii can increase crop productivity by up to 300%. Root colonization by Trichoderma strains enhances root growth and development, crop productivity, resistance to abiotic stresses and uptake of nutrients [30].
This research evaluated the capabilities of Zimbabwean Trichoderma isolates to suppress the FW-RKN disease complex in potatoes. To achieve this, in vitro studies were carried out to assess the effectiveness of three Trichoderma isolates in inhibiting Fusarium oxysporum growth and RKN egg-hatching and inducing juvenile mortality. Additionally, field studies were carried out to evaluate the: (i) efficacy of the Trichoderma isolates in suppressing Fusarium wilt and M. javanica disease prevalence (ii) impact of Trichoderma isolates on potato growth and yield (iii) ideal application technique for the Trichoderma isolates.
2. Materials and Methods
2.1. Study Sites
In vitro studies to evaluate the degree of antagonism caused by Trichoderma isolates against Fusarium oxysporum, the inhibitory effects of isolates against nematode egg hatching, and levels of mortality in M. javanica juveniles, were carried out at the Tobacco Research Board (Kutsaga) in Harare, Zimbabwe. Two field trials for evaluating the efficacies of the Trichoderma isolates against the FW-RKN disease complex and potato plant growth promotion were done at two sites in Gokwe, Midlands Province of Zimbabwe. One of the trial sites was the Department of Research and Specialist Services (DRSS) experimental site in Gokwe South (18°12' S; 28°29' E; 1 237 m.a.s.l) while the other site was at Gwehava village (18°13' S; 28°27' E; 1249 m.a.s.l) in Gokwe North district. Both sites have regosol soils, and receive an average of 800 mm annual rainfall per annum, with a mean annual temperature of between 18 and 22 oC [33].
2.2. Experimental Design and Sources of Trichoderma, Meloidogyne javanica and Fusarium oxysporum Isolates
All in vitro studies were laid out in a completely randomised design with three replications. There were four treatments (Trichoderma isolates) namely T77, Dam 3, T28 and control). The Trichoderma, Meloidogyne javanica and Fusarium oxysporum isolates were sourced from cultures that are maintained in the Plant Health and Agricultural Resilience Division at Kutsaga.
2.3. Experimental Procedures for the Laboratory Experiments
2.3.1. Evaluating the Degree of Antagonism by Trichoderma Isolates in Suppressing Fusarium oxysporum Growth
Potato Dextrose Agar (PDA) plates amended with chloramphenicol were inoculated with Trichoderma and Fusarium oxysporum, with 4 mm agar blocks of the Trichoderma isolates and F. oxysporum placed 1 cm from the edge of the plates on opposite sides. Control plates were inoculated with 4 mm agar blocks of F. oxysporum at the centre of the petri dish containing PDA. All plates were incubated at 28 oC for five days. Mycelial growth inhibition of F. oxysporum by each Trichoderma isolate was calculated as follows:
where:
Tc = Mycelial radius growth in control plate; Tm = Mean mycelial radius growth in test plate
2.3.2. Evaluating the efficacy of Trichoderma isolates in suppressing RKN egg hatching
Tomato plants (cv Rodade) were grown on a site that was naturally infested with RKNs for six weeks after which the plants were uprooted and the roots washed under running water to remove adhering soil particles prior to nematode extraction. The roots were cut into 0.5 cm long pieces and the eggs were extracted according to [34].
Five millilitres of nematode inoculum containing 3,000 eggs was measured out for testing with the Trichoderma isolates. Two millilitres of each Trichoderma isolate solution were mixed with 5 ml of the nematode egg inoculum in a petri dish and were then incubated at room temperature. At 24, 48 and 72 hours, 500 µL of the RKN-Trichoderma isolate mixture were pipetted out into a nematode counting dish and the number of juveniles that hatched was counted under the Leica Wild M3Z light microscope (Leica, USA). The experiment was repeated three times.
2.3.3. Evaluating the Efficacy of Trichoderma Isolates in Causing Mortality in RKN Juveniles
The extracted eggs were incubated in a 500 ml beaker in water for 48 hours at room temperature. The eggs hatched into second-stage juveniles (J2) and a 5 ml solution containing the J2 was measured out and mixed with 2 ml Trichoderma isolates solution and incubated at room temperature. At 24-hour intervals and for 5 days, the numbers of dead juveniles were counted under a stereomicroscope. Juveniles were considered dead when, after being probed with a fine needle, they did not move and the body had become straight. This experiment was also repeated three times.
2.4. Field Evaluation of the Efficacy of Trichoderma Isolates in Controlling FW-RKN Disease Complex and Impact on Plant Growth
2.4.1. Experimental Design
The field trials were laid out as 2 × 6 factorial experiments in a randomized complete block design replicated three times. One factor was the isolate application technique with two levels (seed treatment and furrow application) while the other factor was the isolate treatment with six levels (T77, Dam 3, T28, T77+T28, fenamiphos and control).
2.4.2. Experimental Procedure
In preparing the Trichoderma inocula, eight kilograms of sorghum straws were collected from the field and extraneous matter was removed. The straws were boiled for one hour to kill pathogens and then oven-dried. This process was repeated to ensure complete sterilization of the straws. Two kilograms of the sterilized straws were placed in separate glass jars and inoculated individually with the Trichoderma treatments and F. oxysporum.
Both trial sites were tilled to 30 cm depths, soil clods were broken and plots measuring 9 m x 5 m were marked out. Plots within a block were separated from each other by 0.5 m wide pathways, while blocks were separated from each other by one-metre wide pathways. Sprouted potato tubers (cultivar BP1) were inoculated with the Trichoderma isolate treatments in water suspension at a rate of 10 g/L water some 24 hours before planting. Fenamiphos was applied at 2.5 g/ L water. The potatoes were planted in treated furrows at 75 cm x 25 cm spacing. Furrow treatment with the Trichoderma isolates was done 2 hours before planting at the rate of 10 g/m2, while fenamiphos was applied at the rate of 5 g/m2.
Compound C (6% N: 15% P2O5: 12% K2O) was applied as basal fertilizer at the rate of 1,600 kg/ha. Topdressing with ammonium nitrate (34.5% N) at 100 kg/ha was done as a split application three and five weeks after emergence. The plots were irrigated two days prior to potato planting and subsequent irrigations were applied based on crop-water requirements. Weeds were controlled by hoe weeding. Earthing up was done at 3 and 5 weeks after crop emergence.
At planting, every planting station was inoculated with 5 ml of nematode inoculum. F oxysporum was inoculated two weeks after planting at the rate of 5 ml inoculum solution per plant station. Experimental plots at both trial sites were fumigated with metam sodium after crop harvesting.
2.4.3. Data Collection and Analysis
Potato plant height, chlorophyll content, and leaf area index (LAI) were measured 40 days after planting. A portable Chlorophyll Meter LCPM-A11 (Labtron, United Kingdom) was used to measure chlorophyll content on the two fully-expanded uppermost leaves of the plant. The LAI was measured using a canopy analyser LAI 2000 by using the 180 Sensor. Fusarium wilt disease incidence and severity were measured at 60 days after planting. Disease incidence was measured by counting the number of plants showing wilting and chlorosis symptoms and expressing it as a proportion of the total plot plant population. Laboratory assays to recover and identify F. oxysporum from diseased plants were done. The scale (Table 1) was used in measuring Fusarium wilt disease severity. The average tuber weight per treatment and tuber yield (expressed in tonnes per hectare) were measured at physiological maturity (90 days).
Nematode egg masses per plant were determined using the rating scale described by the Taylor and Sasser eggmass scale [35] (Table 2). The degree of galling was assessed using the Daulton and Nasbaum scale [36] (Table 3).
All data was subjected to analysis of variance using Genstat 18th edition. Where significant differences occurred, the least significant difference (LSD) at p = 0.05 was used to separate means.
3. Results
3.1. Identity of the Trichoderma Isolates Used in this Study
The three Trichoderma isolates that were used in this study were sequenced and identified as Trichoderma spp (T77), Trichoderma asperelloides (Dam 3) and Trichoderma asperellum (T28). The genome sequence data have been deposited in the NCBI BioProject database under accession numbers PRJNA1250815 for T77, PRJNA1246528 for T28 and PRJNA1245440 for Dam3. In this publication, the names of the isolates will be maintained as T77, Dam 3 and T28.
3.2. Antagonistic Effects of Trichoderma Isolates Against F. oxysporum
Trichoderma isolates had a significant (p < 0.05) antagonistic effect on F. oxysporum growth. The T77 isolate had the highest antagonistic effect (61.33%) followed by T28 and Dam 3 at 52.00% and 43.67%, respectively (Figure 1).
3.3. Effect of Isolate Treatments on M. javanica Egg-Hatching
The isolate treatments significantly (p < 0.05) affected M. javanica egg hatching. At Days 2 and 3, nematode egg hatches were statistically similar for all the Trichodermas and they were lower than those of the control treatment. Thereafter, the T77 isolate resulted in fewer egg hatches (8 and 12 egg hatches on Days 4 and 5, respectively) than T28 and Dam 3 (Table 4). There were no statistical differences in the number of eggs that hatched on Days 4 and 5 for isolates T28 and Dam 3.
3.4. Effect of Trichoderma Isolates on Mortality of M. javanica Juveniles
The isolate treatments had a significant effect (p < 0.05) on the mortality of M. javanica juveniles. At all times, the order of juvenile mortality induced by the Trichodermas was as follows: T77 > Dam 3 > T28. For T77, there was an increase in juvenile mortality over time, with 38% mortality on Day 1 that increased to 96% on Day 5. There were no significant differences in mortality due to T28 and Dam 3 isolates on Days 2, 3 and 4, with significant differences noted on Day1 and 5 where the Dam 3 isolate outperformed the T28 isolate (Table 5).
3.5. Effects of Trichoderma Isolates on Fusarium Wilt and RKN Disease Prevalence in the Field
3.5.1. Effects on Fusarium wilt Disease Incidence and Severity
There was no interaction (p > 0.05) between application technique and isolate treatment on Fusarium wilt disease incidences at both sites. However, significant differences (p < 0.05) were recorded for the effects of the isolate treatment on disease incidences at both sites. All the Trichoderma isolates were either better or comparable to fenamiphos. The T28+T77 isolate treatment resulted in the least disease incidence at both sites. With 14.5% disease incidence at the DRSS site and 16.5% at Gwehava village, it outperformed fenamiphos, which had 16.83% and 18.83% disease incidence at the two sites, respectively (Table 6).
There was also no interaction (p > 0.05) between the application technique and isolate treatments on disease severity at both sites. However, isolate treatments had an effect (p < 0.05) on disease severity at both sites. T77 treatment resulted in the least disease severity at both sites (0.333 at the DRSS site and 0.387 at Gwehava village), followed by Dam 3 (0.500 at DRSS site and 0.550 at Gwehava village), T28+T77 (0.667 at DRSS site and 0.717 at Gwehava village) and T28 (1.000 at DRSS site and 1.05 at Gwehava village). Isolates T77, Dam 3, and T28+T77 were comparable to fenamiphos in suppressing Fusarium wilt disease severity (Table 6).
3.5.2. Effects on the Number of Root Knot Nematode Galls
No interaction (p = 0.142) was observed between the application technique and isolate treatment on the number of galls per plant at the DRSS site. However, isolate treatments had an effect (p < 0.05) on number of galls per plant. All the Trichoderma treatments resulted in significantly fewer galls per plant (1.2 – 1.3) than the control (3.3) and were comparable to fenamiphos (Figure 2).
Significant interaction (p = 0.047) was recorded between the isolate application technique and isolate treatment on the number of RKN galls at Gwehava village. The control treatment at both sites had the highest number of galls (2.00 galls/plant in furrow-applied isolates and 3.00 galls/plant in seed-applied isolates). Furrow-applied Dam 3 isolate, and seed-applied T77 and T28+T77 isolates resulted in the least galling (1.00 galls/plant). The Trichodermas, whether seed or furrow applied were comparable to fenamiphos with regards to the resultant number of galls per plant (Table 7).
3.5.3. Effect on the Number of RKN Egg Masses
There was no interaction (p > 0.05) between the isolate application technique and the isolate treatment on egg mass numbers at both sites. However, significant differences (p < 0.05) were observed for the isolate treatments on egg mass numbers at the DRSS site but not at Gwehava village. The least number of egg masses (1.667) were in plots treated with T28 and Dam 3 isolates. All Trichoderma-based isolate treatments had significantly lower numbers of nematode egg masses than the control and were comparable to the fenamiphos treatment (Figure 3).
3.6. Effects of Isolate Treatments on Potato Plant Growth and Yield
3.6.1. Effects on Plant Height
There was a significant interaction (p = 0.008) between the isolate application techniques and the isolate treatments on plant height at the DRSS site. The tallest plants, at 24 cm, were from furrow- and seed-applied T77 isolate treatments. These plants were 33% taller than those in the control treatments, and 9.7% taller than seed- and furrow- T28 treatments (Figure 4).
At Gwehava village, no interaction (p > 0.05) was observed between the application technique and isolate treatments on plant height. However, isolate treatments had a significant effect (p < 0.05) on plant height, with the tallest plants (24 cm) reported in plots treated with the T77 isolate and those in the control treatment being the shortest (19.33 cm). There were no significant differences in plant height between plants that were treated with the T28 and T28+T77 isolates (Figure 5). The order of plant heights for the isolate treatments was as follows: T77 > Dam 3 > T77+T28 > T28 > fenamiphos > control.
3.6.2. Effects on Potato Chlorophyll Content
There was interaction (p < 0.05) between isolate application techniques and isolate treatments on chlorophyll content at the DRSS site. The highest chlorophyll content (0.98) was recorded in plants where T77 was seed-applied (Figure 6). These plants had 10.11% more chlorophyll than the control treatment (0.89). Amongst the Trichodermas, both furrow- and seed-applied T28 treatments resulted in plants with the least chlorophyll content (0.94 for furrow- and 0.92 for seed-applied).
At Gwehava village, there was no interaction (p = 0.322) between the isolate application techniques and isolate treatments on chlorophyll content. However, isolate treatment had a significant effect (p < 0.05) on chlorophyll content. Plants treated with isolate T28 had the least (0.93) chlorophyll content while those treated with fenamiphos (0.9783) had the highest chlorophyll content. There were no significant differences in chlorophyll content between T77- and fenamiphos-treated plants (Figure 7).
3.6.3. Effects on Leaf Area Index
There was interaction (p < 0.05) between application techniques and isolate treatments on leaf area index (LAI) at both sites. At the DRSS site, Trichoderma-containing isolates had significantly larger LAI than the control and fenamiphos treatments. The largest LAI was recorded in furrow-applied T77+T28 isolate (5.37) and this was 74.92% higher than the control treatment. Amongst the Trichoderma-containing isolates, the lowest LAI was recorded in seed-applied T28 (3.63) and this was still 18.2% higher than the LAI in the control treatment. There were no significant differences in the LAI among the following treatments: seed-applied T77+T28; seed- and furrow-applied T77; and seed- and furrow-applied Dam 3 (Figure 8).
At Gwehava village, the furrow-applied T77+T28 treatment resulted in plants with the largest LAI (5.37). The seed-applied T28 treatment had the least LAI (3.63) amongst the Trichoderma-containing treatments (Figure 9).
3.6.4. Effect on Potato Tuber Size and Yield
No interaction (p > 0.05) was recorded between isolate application techniques and the isolate treatments on potato tuber size at both sites. However, isolate treatments had an effect (p < 0.05) on the tuber size at both sites. After the control treatment, the T28 treatment resulted in the production of the smallest tubers (125.80 g at the DRSS site and 122.4 g at Gwehava village) while the T77+T28 treatment resulted in the production of the largest tubers (130.12 g at DRSS site and 129.23 g at Gwehava village) at both sites (Table 8).
There was also no interaction (p > 0.05) between isolate application techniques and isolate treatments on potato yield at both sites. However, isolate treatment had a significant effect (p < 0.05) on yield, with the highest yield recorded in plots treated with T77 (34.50 t/ha at the DRSS site and 32.5 t/ha at Gwehava village) (Table 8). These yields were at least 100% higher than the yields in the control treatments. Amongst the treatments that contained Trichoderma, the lowest yields were obtained when T28 was applied at both sites (25.13 t/ha at the DRSS site and 28.43 t/ha at Gwehava village).
4. Discussion
4.1. Effects of Trichoderma isolates on F. oxysporum growth and Fusarium wilt disease prevalence
The application of Trichoderma isolates suppressed the growth of F. oxysporum in vitro. It also reduced the prevalence of the Fusarium wilt disease in potatoes in the field. The genus Trichoderma is made up of multi-action fungi that operate by antagonism, competition and mycoparasitism. They antagonize F. oxysporum by producing hydrolytic enzymes that degrade chitin in the pathogen cell wall [29]. Also, Trichoderma compete for nutrients and space against pathogens, parasitize them and secrete antibiotics [27,37]. They produce highly efficient siderophores that chelate iron and stop the growth of other fungi [38].
The Trichoderma isolates differed in their impact on F. oxysporum growth and Fusarium wilt disease prevalence. In this study, the T77 isolate outperformed the other isolates in inhibiting F. oxysporum growth, while the T28+T77 isolate outperformed all the isolates in reducing Fusarium wilt disease incidence and severity. This showed that there are inherent differences among isolates in antagonizing F. oxysporum and suppressing Fusarium wilt disease prevalence. Theradimani et al. [39] attributed such differences to the fact that some Trichoderma species and strains sporulate faster and produce larger quantities of hydrolytic enzymes than others. In this study, the T28+T77 isolate was a combination of T28 and T77 isolates. Its higher efficacy was possibly because of a synergistic effect of the hydrolytic enzymes produced by the individual isolates. In instances where Fusarium wilt is a problem in potato production, application of the T28+T77 isolate either as a seed treatment or in furrows at planting, could reduce disease prevalence.
4.2. Effect of Trichoderma Isolates on Nematode Egg Hatching, Juvenile Mortality and Galling
Trichoderma isolates’ application significantly delayed and inhibited nematode egg hatching. According to [39,41], delayed egg hatching occurs when Trichoderma spp parasitize the gelatinous eggmass matrix. The Trichoderma attaches to the matrix and also utilizes it as a nutrient source. Furthermore, the conidia of Trichoderma attach to the eggshell and parasitize it [42]. Trichoderma spp produce chitinases and proteases which inhibit nematode egg hatching and also degrade eggshells [15,43]. Several enzymes, including chitinases, glucanases, and proteases parasitize nematode juveniles [44].
The different capacities of the Trichoderma isolates to delay and inhibit egg hatching, and induce juvenile mortality can be attributed to inherent differences among isolates to produce different quantities and types of hydrolytic enzymes [15,45]. In this study, the T77 isolate was most effective at delaying/inhibiting both egg hatching and inducing higher juvenile mortality.
Potato plants that were treated with Trichoderma isolates had significantly fewer galls than those that were not treated. Amongst the Trichoderma isolates, seed- and furrow-applied T77 and Dam 3 treatments resulted in the least galling levels. Trichoderma forms a sheath around the plant root thereby inhibiting nematode penetration into the plant. They parasitize nematodes by using different hydrolytic enzymes thereby leading to suppression of egg-laying and ultimately, galling [22]. Where root knot nematodes are a problem in potatoes, growers can either seed dress or furrow-apply the T77 and Dam 3 isolates.
4.3. Effect of Trichoderma Isolates on Potato Growth and Yield
Application of Trichoderma isolates resulted in significant potato plant growth as was evidenced by increased plant height, chlorophyll content, leaf area index, average tuber size and final tuber yield. The tallest plants, which also had most chlorophyll content, were in plots treated with the T77 isolate while T28-treated plants were the shortest and had the least chlorophyll content. The largest LAI was in plants where the T28+T77 isolate was applied while T28-treated plants had the least LAI. Taller plants with a larger LAI can capture more light for photosynthesis than shorter plants with a smaller LAI, leading to more growth in the taller plants [46]. Similarly, potato plants with higher chlorophyll content produce more photosynthates than those with low chlorophyll content, leading to larger tuber size and higher yield.
The Trichoderma can be applied as a seed dressing or in the furrow at planting. Whether applied as a seed dressing or in the furrow, Trichoderma outcompeted both the root knot nematodes and F. oxysporum for nutrients in the plant′s rhizosphere, thereby reducing disease prevalence and enhancing potato crop growth. Trichoderma is also reported to enhance crop root growth and development thereby leading to efficient nutrient uptake and use which promotes crop growth and development [47]. Increased plant growth due to Trichoderma application could be associated with the secretion of hormones such as auxins, gibberellins and cytokinins that boost root and shoot development as reported by [27].
Different Trichoderma strains and isolates produce different quantities and types of plant growth stimulators [47]. They also have different disease-suppression potentials. The use of these Trichoderma isolates, especially T77 and T28+T77, could help to sustainably suppress diseases [48] and reduce the overuse and misuse of synthetic fertilizers which contribute to eutrophication, soil degradation and the emission of greenhouse gases during their production [49]. Integrating Trichoderma isolates into existing FW-RKN disease complex control practices can contribute to sustainable agriculture and improved crop yield.
5. Conclusions and Recommendations
This study showed that the T77 isolate was most effective in inhibiting F. oxysporum growth, reducing Fusarium wilt prevalence, inducing nematode juvenile mortality and reducing galling in potato plants. The T77 and T28+T77 isolates significantly increased potato plant height, chlorophyll content, leaf area index, tuber size and yield. The isolates can either be seed or furrow-applied. The use of these Trichoderma isolates offers a sustainable option to control the FW-RKN complex and so reduce reliance on synthetic chemicals. Both isolates are recommended for use in sustainable management of the FW-RKN disease complex in potatoes.
Author Contributions
Conceptualization, CC. and HM.; methodology, HM and CC.; data analysis, TM, RM, MC, CK; writing—original draft preparation, HM.; writing—review and editing, CK, CC; supervision, CC, RM, MC; data curation, TM. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The genome sequence data for the Trichoderma isolates have been deposited in the NCBI BioProject database under accession numbers PRJNA1250815 for T77, PRJNA1246528 for T28 and PRJNA1245440 for Dam3.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| DRSS | Department of Research and Specialist Services |
| FW-RKN | Fusarium wilt -root knot nematode |
| LAI | Leaf area index |
| LSD | Least Significant Difference |
| PDA | Potato Dextrose Agar |
| RKN | Root knot nematode |
References
- Devaux, A.; Goffart, J.-P.; Kromann, P.; Andrade-Piedra, J.; Polar, V.; Hareau, G. The potato of the future: opportunities and challenges in sustainable agri-food systems. Potato Res 2021. [Google Scholar] [CrossRef] [PubMed]
- Gikundi, E.K.; Buzera, A.K.; Orina, I.N.; Sila, DN. Storability of Irish potato (Solanum tuberosum L.) varieties grown in Kenya, under different storage conditions. Potato Res 2022, 66, 137–158. [Google Scholar] [CrossRef]
- Ashraf, A.; Rauf, A.; Abbas, M.F.; Rehman, R. Isolation and identification of Verticillium dahliae causes wilt on potato in Pakistan. Pak. J. Phytopathol. 2012, 24, 112–116. [Google Scholar]
- Gondal, A.S.; Javed, N.; Khan, S.A.; Hyder, S. Genotypic diversity of potato germplasm against root knot nematode (Meloidogyne incognita) infection in Pakistan. Int. J. Phytopathol. 2012, 1, 27–38. [Google Scholar] [CrossRef]
- Abbas, M.F.; Aziz-ud-Din, G.A.; Qadir, A.; Ahmed, R. Major potato viruses in potato crops of Pakistan: a brief review. Int. J. Biol. Biotechnol 2013, 10, 425–430. [Google Scholar]
- El-Shennawy, M.Z.; Khalifa, E.Z.; Ammar, M.M.; Mousa, E.M.; Hafez, S.L. Biological control of the disease complex on potato caused by root-knot nematode and Fusarium wilt fungus. Nematol. Mediterr 2012, 40, 169–172. [Google Scholar]
- Lima, F.S.O.; Mattos, V.S.; Silva, E.S.; Carvalho, M.A.S.; Teixeira, R.A.; Silva, J.C.; Correa, V.R. Nematodes affecting potato and sustainable practices for their management. “Potato: from Incas to all over the world. 2018, 107. [Google Scholar] [CrossRef]
- Onkendi, E.M.; Kariuki, G.M.; Marais, M.; Moleleki, L.N. The threat of root-knot nematodes and fitness consequences. J. Exp. Biol. 2014, 211, 1927–1936. [Google Scholar]
- Kassie, Y.G. Status of root-knot nematode (Meloidogyne species) and Fusarium wilt (Fusarium oxysporum) disease complex on tomato (Solanum lycopersicum L.) in the Central Rift Valley, Ethiopia. Agric. Sci 2019, 10, 1090–1103. [Google Scholar] [CrossRef]
- Bhagawati, B.; Das, B.C.; Sinha, A.K. Interaction of Meloidogyne incognita and Rhizoctonia solani on okra. Ann. Plant Prot. Sci. 2007, 1, 533–535. [Google Scholar]
- Manzira, C. Potato production handbook. Potato Seed Association Zimbabwe. Jongwe Printers, Harare, Zimbabwe. 2010.
- Bairwa, A.; Venkatasalam, E.P.; Mhatre, P.H.; Bhatnagar, A.; Sharma, A.K.; et al. Biology and management of nematodes in potato. In: S Kumar et al. (eds). Sustainable management of potato pests and diseases. 2022. [CrossRef]
- Jahanshir, A.; Dzhalilov Fevzi, S. The effects of fungicides on Fusarium oxysporum f. sp. lycopersici associated with fusarium wilt of tomato. J. Plant Prot. Res. 2010, 50, 172–178. [Google Scholar]
- Barari, H. Biocontrol of tomato Fusarium wilt by Trichoderma species under in vitro and in vivo conditions. Cercetări Agronomice în Moldova 2016, 1, 91–98. [Google Scholar] [CrossRef]
- Feyisa, B.; Lencho, A.; Selvaraj, T.; Getaneh, G. Evaluation of some botanicals and Trichoderma harzianum for the management of tomato root knot nematode (Meloidogyne incognita (Kofoid and White) Chitwood). ACST. 2015, 4, 201. [Google Scholar] [CrossRef]
- Al-Askar, A.A.; Rashad, Y.M. Arbuscular mycorrhizal fungi: a biocontrol agent against common bean Fusarium root rot disease. Plant Pathol. J. 2010, 9, 31–38. [Google Scholar] [CrossRef]
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol. Lett. 2010, 32, 1559–1570. [Google Scholar] [CrossRef]
- Saeki, E.K.; Kobayashi, R.K.T.; Nakazato, G. Quorum sensing system: Target to control the spread of bacterial infections. Microb. Pathog. 2020, 142, 104068. [Google Scholar] [CrossRef]
- Farzand, A.; Moosa, A.; Zubair, M.; Khan, A.R.; Massawe, V.C.; Tahir, H.A.S.; Sheikh, T.M.M.; Ayaz, M.; Gao, X. Suppression of Sclerotinia sclerotiorum by the induction of systemic resistance and regulation of antioxidant pathways in tomato using fengycin produced by Bacillus amyloliquefaciens FZB42. Biomol. 2019, 9, 613. [Google Scholar] [CrossRef]
- Zubair, M.; Farzand, A.; Mumtaz, F.; Khan, A.R.; Sheikh, T.M.M.; Haider, M.S.; Yu, C.; Wang, Y.; Ayaz, M.; Gu, Q.; et al. Novel genetic dysregulations and oxidative damage in Fusarium graminearum induced by plant defense eliciting psychrophilic Bacillus atrophaeus Ts1. Int. J. Mol. Sci 2021, 22, 12094. [Google Scholar] [CrossRef]
- Khan, B.; Yan, W.; Wei, S.; Wang, Z.; Zhao, S.; Cao, L.; et al. Nematicidal metabolites from endophytic fungus Chaetomium globosum YSC5. FEMS. Microb. Lett 2019, 366, fnz169. [Google Scholar] [CrossRef]
- Goswami, B.K.; Pandey, R.K.; Goswami, J.; Tewari, D.D. Management of disease complex caused by root knot nematode and root wilt fungus on pigeon pea through soil organically enriched with vesicular arbuscular mycorrhiza, karanj (Pongamia pinnata) oilseed cake and farmyard manure. J. Environ. Sci. Health Part B. 2007, 8, 899–904. [Google Scholar] [CrossRef]
- Hanawi, M.J. Tagetes erecta with native isolates of Paecilomyces lilacinus and Trichoderma hamatum in controlling root-knot nematode Meloidogyne javanica on tomato. IJAIEM. 2016, 5, 81–88. [Google Scholar]
- Bontempo, A.F.; Lopes, E.A.; Fernandes, R.H.; Freitas, L.G.D.E.; Dallemole-Giaretta, R. Dose-response effect of Pochonia chlamydosporia against Meloidogyne incognita on carrot under field conditions. Rev. Caatinga. 2017, 30, 258–262. [Google Scholar] [CrossRef]
- Jones, E.E.; Rabeendran, N.; Stewart, A. Biocontrol of Sclerotinia sclerotiorum infection of cabbage by Coniothyrium minitans and Trichoderma spp. Biocontrol Sci. Tech. 2014, 24, 1363–1382. [Google Scholar] [CrossRef]
- Khan, M.R.; Ahmad, I.; Ahamad, F. Effect of pure culture and culture filtrates of Trichoderma species on root-knot nematode, Meloidogyne incognita infesting tomato. Indian Phytopathol. 2018, 71, 265–274. [Google Scholar] [CrossRef]
- Verma, N.P.; Kaur, I.; Masih, H.; Singh, A.K.; Singla, A. Efficacy of Trichoderma in controlling Fusarium wilt in tomato (Solanum lycopersicum L.). Res. Environ. Life Sci 2017, 10, 636–639. [Google Scholar]
- Martinuz, A.; Zewdu, G.; Ludwig, N.; Grundler, F.; Sikora, R.A.; Schouten, A. The application of Arabidopsis thaliana in studying tripartite interactions among plants, beneficial fungal endophytes and biotrophic plant-parasitic nematodes. Planta. 2015, 241, 1015–1025. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J. Trichoderma parareesei favors the tolerance of rapeseed (Brassica napus L.) to salinity and drought due to a chorismate mutase. Agron 2020, 10, 118. [Google Scholar] [CrossRef]
- Mohiddin, F.A.; Khan, M.R.; Khan, S.M.; Bhat, B.H. Why Trichoderma considered a super hero (super fungus) against the evil parasites. Plant Pathol. J. 2010, 9, 92–102. [Google Scholar] [CrossRef]
- Sahebani, N.; Hadavi, N. Biological control of the root-knot nematode Meloidogyne javanica by Trichoderma harzianum. Soil Biol. Biochem. 2008, 40, 2016–2020. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Appels, F. V.; van Wees, S.C. Impact of salicylic acid- and jasmonic acid-regulated defences on root colonization by Trichoderma harzianum T-78. Plant Signal. Behav 2017, 12, e1345404. [Google Scholar] [CrossRef]
- Nyamapfene, K.W. Soils of tobacco. Nehanda Publishers. Harare, Zimbabwe. 1991.
- Hussey, R.S.; Barker, K.R. Comparison of methods for collecting inocula of Meloidogyne spp., including a new technique. Plant Dis Rep 1973, 57, 1025–1028. [Google Scholar]
- Colyer, P.D.; Kirkpatrick, T.L.; Caldwell, W.D.; Vernon, P.R. Root-knot nematode reproduction and root galling severity on related conventional and transgenic cotton cultivars. J. Cotton Sci. 2008, 4, 232–236. [Google Scholar]
- Southey, J.F. Laboratory Methods for Work with Plant and Soil Nematodes. Her Majesty’s Stationary Office, London, Great Britain. 1986.
- De Palma, M.; Salzano, M.; Villano, C.; Aversano, R.; Lorito, M.; Ruocco, M.; et al. Transcriptome reprogramming, epigenetic modifications and alternative splicing orchestrate the tomato root response to the beneficial fungus Trichoderma harzianum. Hortic. Res. 2019, 6, 5. [Google Scholar] [CrossRef]
- Khan, A.; Mfarrej, M.F.B.; Nadeem, H.; Ahamad, L.; Hashem, M.; Alamri, S.; Gupta, R.; Ahmad, F. Trichoderma virens mitigates the root-knot disease progression in the chickpea plant. Acta Agri. Scand. Section B — Soil. Plant Sci. 2022, 72, 775–787. [Google Scholar] [CrossRef]
- Theradimani, M.; Thangaselvabai, T.; Swaminathan, V. Management of tuberose root rot caused by Sclerotium rolfsii by biocontrol agents and fungicides. Advances in Floriculture and Urban Horticulture, ICAR, New Delhi. 2018, 308-311.
- Forghani, F.; Hajihassani, A. Recent advances in the development of environmentally benign treatments to control root-knot nematodes. Front. Plant Sci. 2020, 11, 1125. [Google Scholar] [CrossRef] [PubMed]
- Galletti, S.; Paris, R.; Cianchetta, S. Selected isolates of Trichoderma gamsii induce different pathways of systemic resistance in maize upon Fusarium verticillioides challenge. Microbiol. Res. 2020, 233, 126406. [Google Scholar] [CrossRef]
- Sharon, E.; Chet, I.; Viterbo, A.; Bar-Egal, M.; Nagan, H.; Samuels, G.J.; Spiegel, Y. Parasitism of Trichoderma on Meloidogyne javanica and role of the gelatinous matrix. Eur. J. Plant Pathol. 2007, 118, 247–258. [Google Scholar] [CrossRef]
- Bhat, A.A.; Shakeel, A.; Waqar, S.; Handoo, Z.A.; Khan, A.A. Microbes vs. Nematodes: Insights into biocontrol through antagonistic organisms to control root-knot nematodes. Plants 2023, 12, 451. [Google Scholar] [CrossRef]
- Rompali, R.; Mehendrakar, S.S.; Venkata, P.K. Evaluation of biocontrol agents on root knot nematode Meloidogyne incognita and wilt causing fungus Fusarium oxysporum f. sp. conglutinans in vitro. Afr. J. Biotechnol. 2016, 15, 798–805. [Google Scholar] [CrossRef]
- Damodaran, T.; Rajan, S.; Muthukumar, M.; Gopal, R.; Yadav, K.; Kumar, S.; et al. Biological management of banana Fusarium wilt caused by Fusarium oxysporum f. sp. cubense tropical race 4 using antagonistic fungal isolate CSR-T-3 (Trichoderma reesei). Front. Microbiol 2020, 11, 595845. [Google Scholar] [CrossRef]
- Dey, S.; Saha, K.; Dave, R.; Nidhin, P.; Murugesan, A. Estimation of leaf area index of mustard and potato from Sentinel-2 data using parametric, non-parametric and physical retrieval models. RSASE. 2025, 37, 101493. [Google Scholar] [CrossRef]
- Yao, X.; Guo, H.; Zhang, K.; Zhao, M.; Ruan, J.; Chen, J. Trichoderma and its role in biological control of plant fungal and nematode disease. Front. Microbiol. 2023, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ul Haq, I.; Rahim, K.; Yahya, G.; Ijaz, B.; Maryam, S.; Paker, N.P. Eco-smart biocontrol strategies utilizing potent microbes for sustainable management of phytopathogenic diseases. Biotechnol Rep. 2024, 44, e00859. [Google Scholar] [CrossRef]
- Kubheka, B.P.; Ziena, L.W. Trichoderma: A Biofertilizer and a Bio-Fungicide for Sustainable Crop Production. In. Trichoderma - Technology and Uses. F.P. Juliatti (Ed), IntechOpen. 2022. [CrossRef]
Figure 1.
Antagonistic effects of the Trichoderma isolates on F. oxysporum growth.
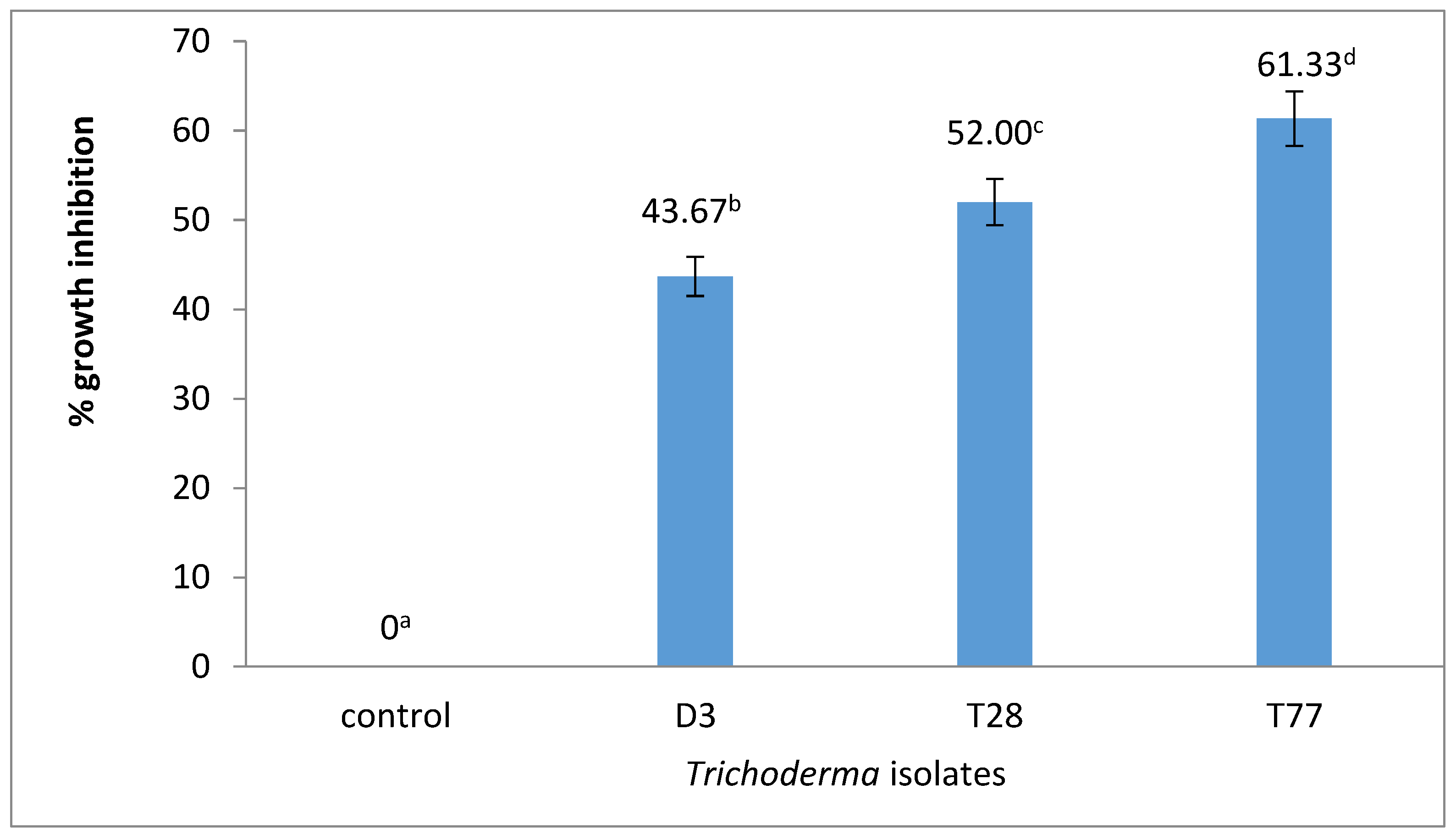
Figure 2.
Effect of Trichoderma isolates on galls per plant at the DRSS site.
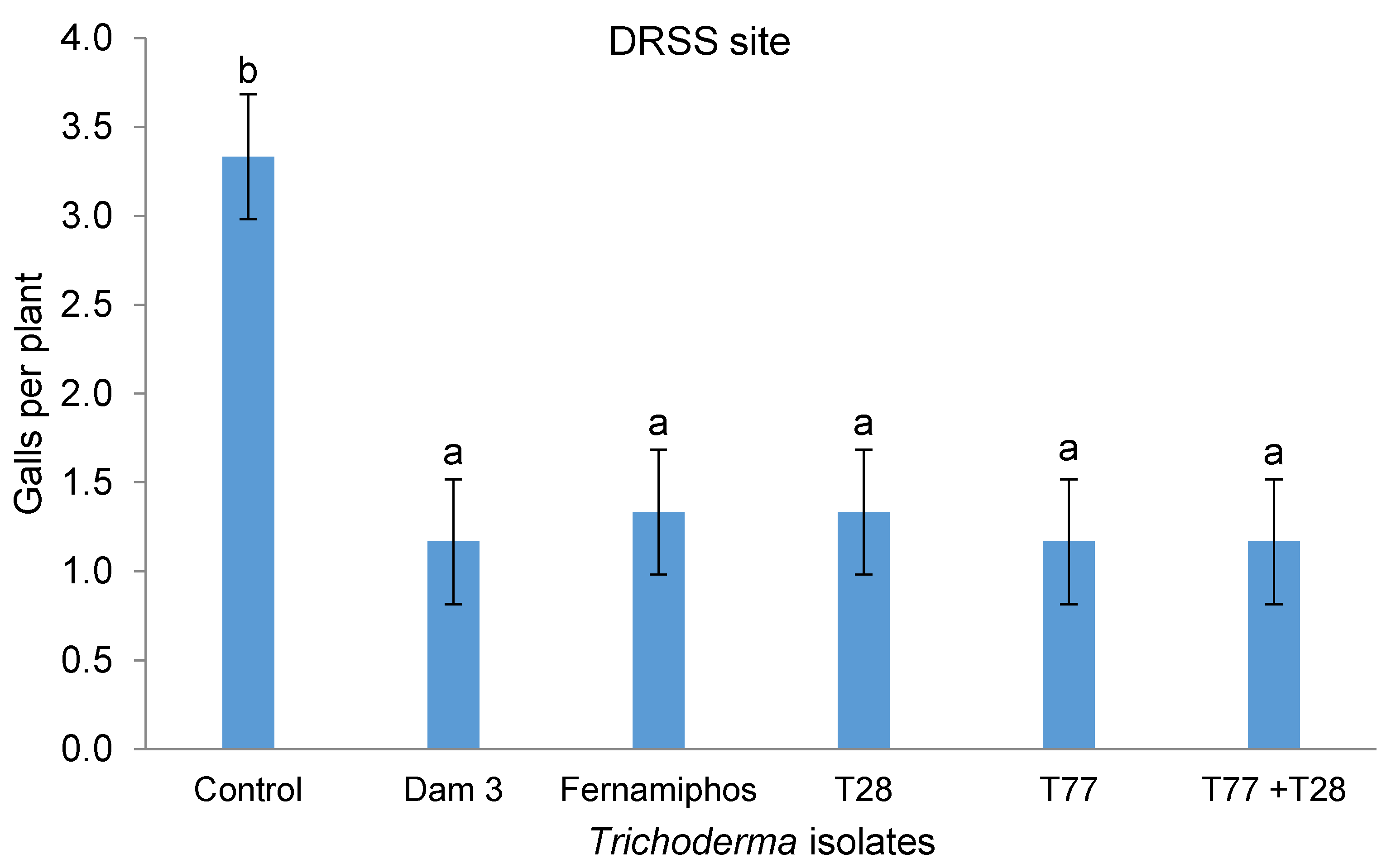
Figure 3.
Effect of Isolate treatments on nematode egg masses at DRSS site.
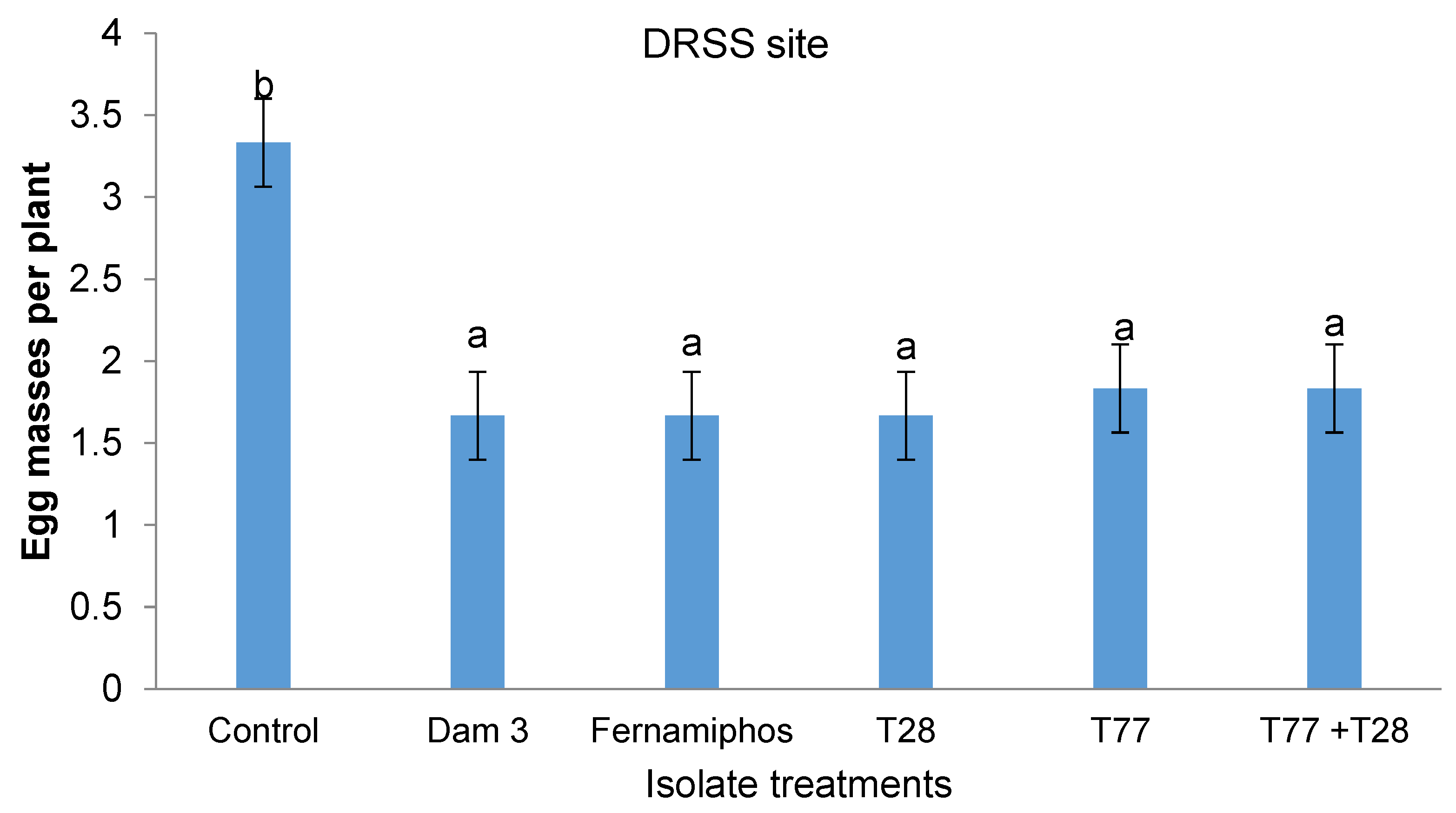
Figure 4.
Effect of isolate application method and isolate treatments on potato plant height at the DRSS site.
Figure 4.
Effect of isolate application method and isolate treatments on potato plant height at the DRSS site.
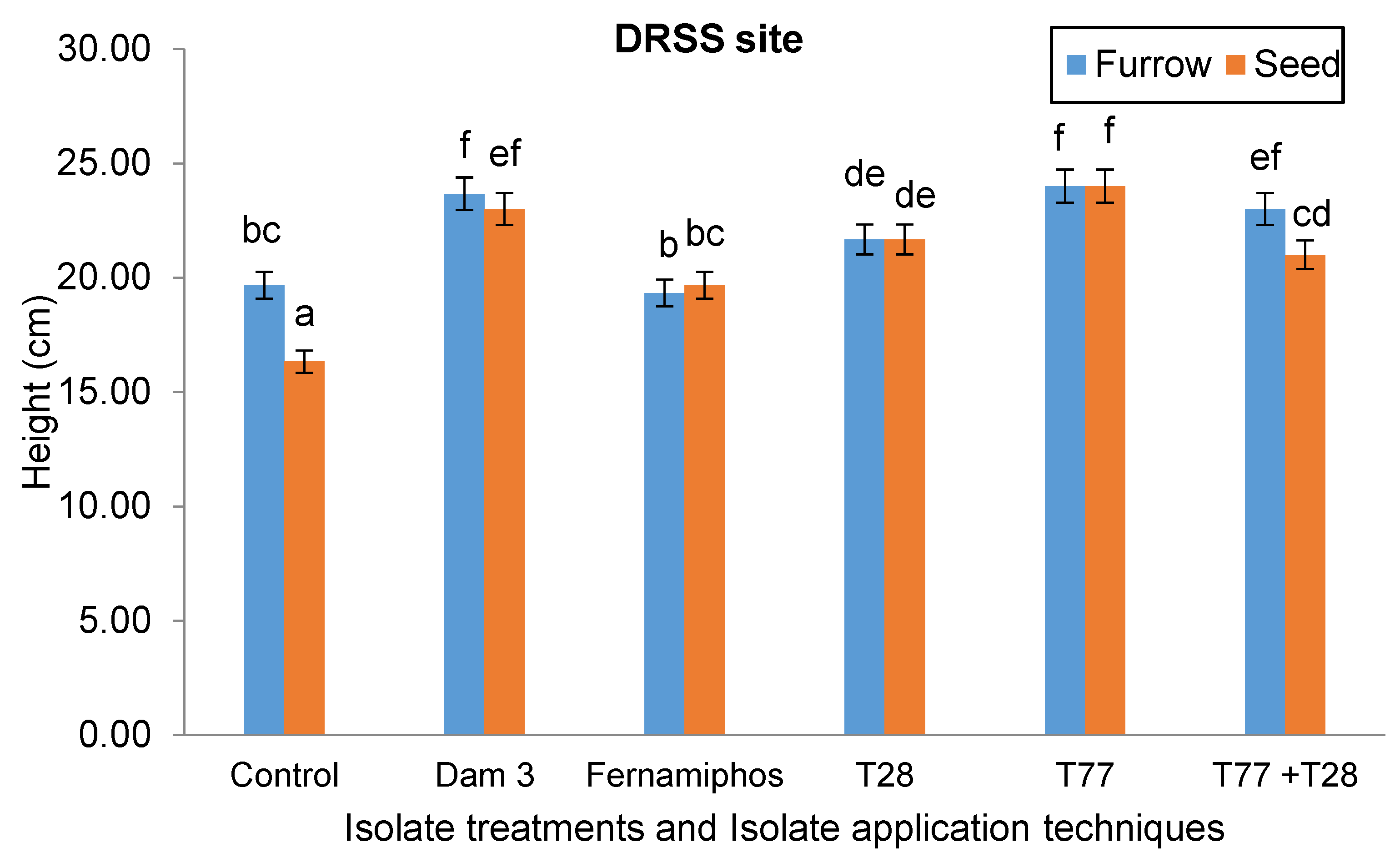
Figure 5.
Effect of Isolate treatments on plant height at Gwehava village.
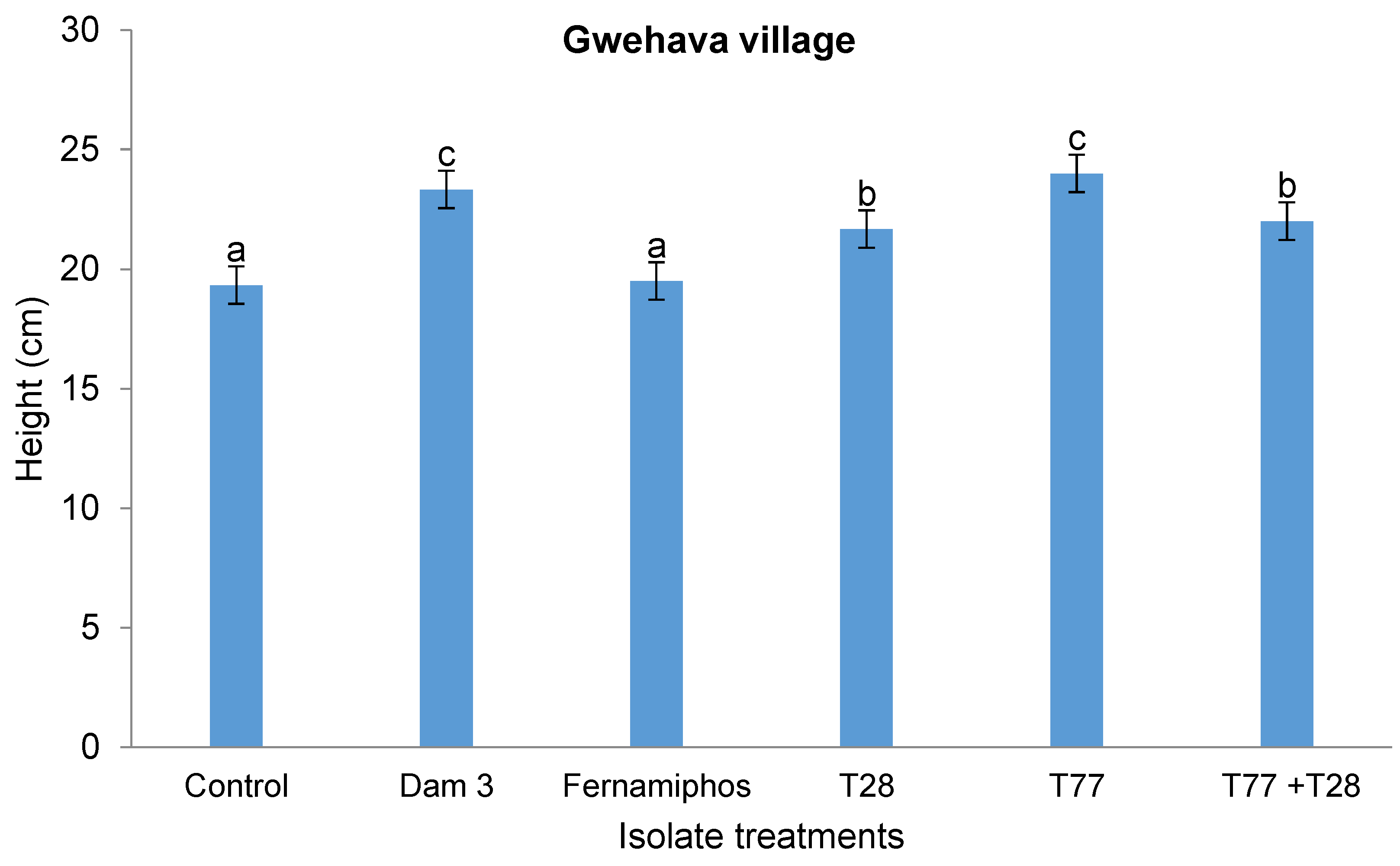
Figure 6.
Effect of isolates application techniques and isolate treatment type on chlorophyll content at DRSS site.
Figure 6.
Effect of isolates application techniques and isolate treatment type on chlorophyll content at DRSS site.
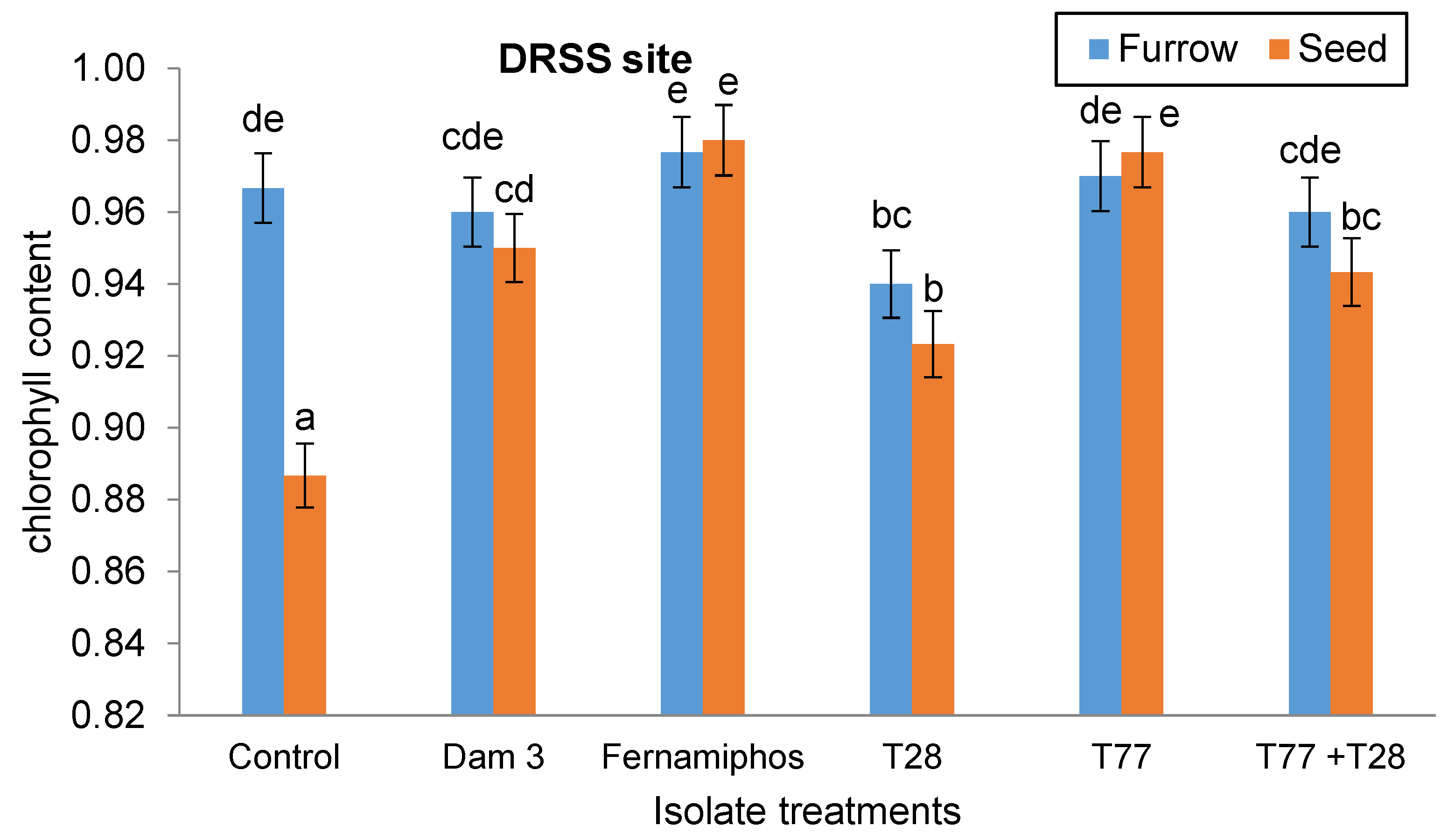
Figure 7.
Effect of Isolate treatments on chlorophyll content at Gwehava village.
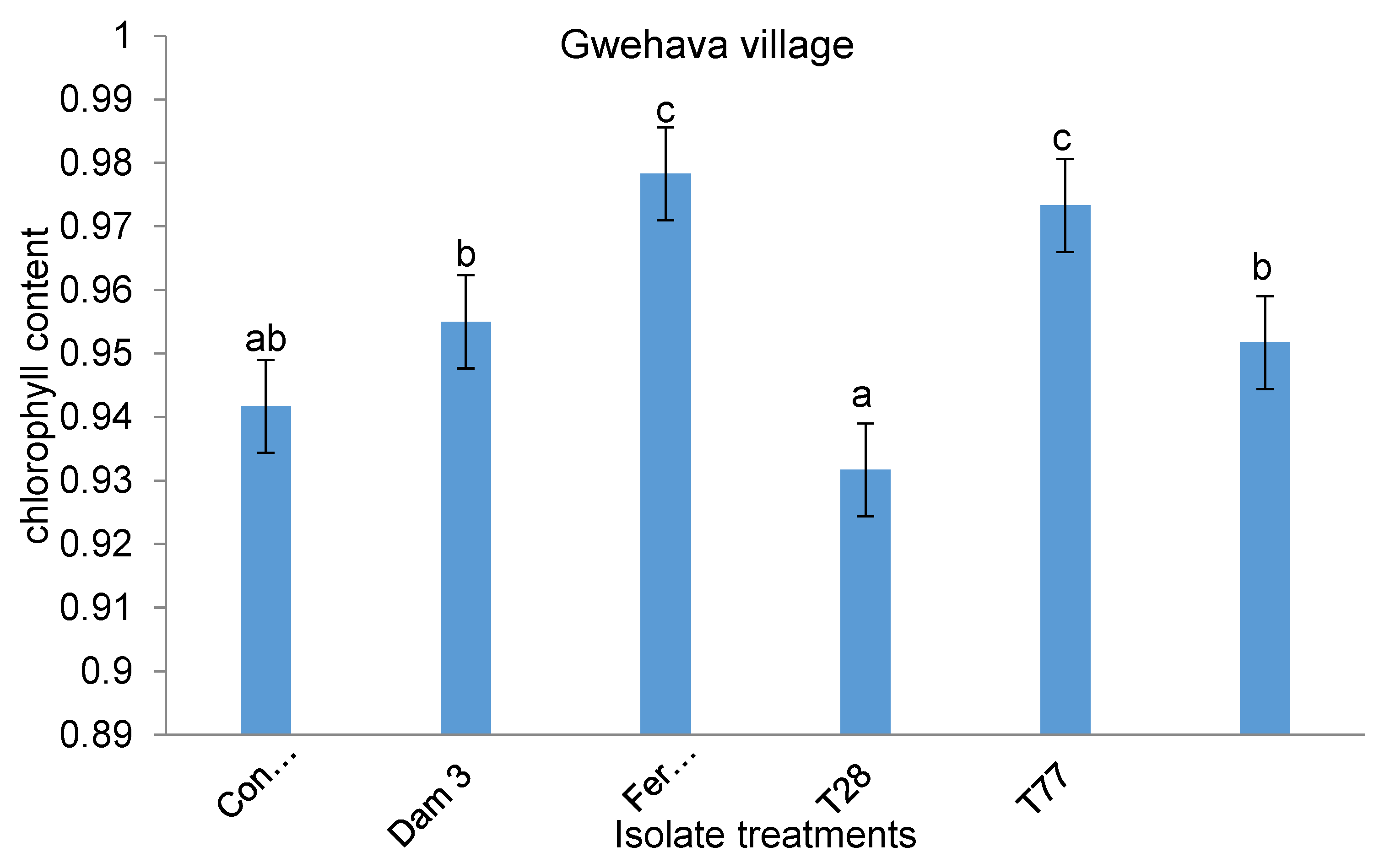
Figure 8.
Effect of application technique and isolate treatments on leaf area index at the DRSS site.
Figure 8.
Effect of application technique and isolate treatments on leaf area index at the DRSS site.
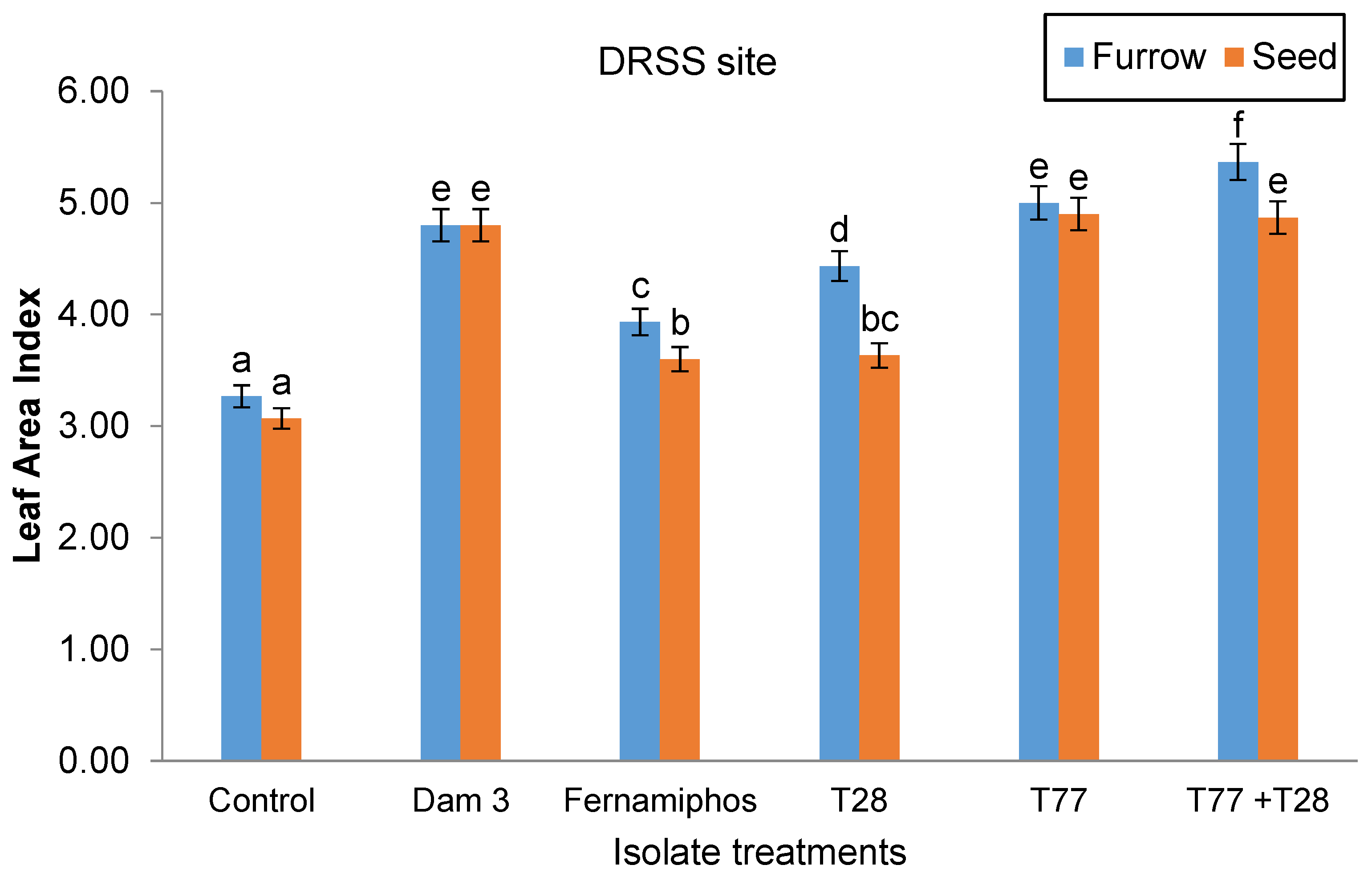
Figure 9.
Effect of application techniques and isolate treatments on leaf area index at Gwehava village.
Figure 9.
Effect of application techniques and isolate treatments on leaf area index at Gwehava village.
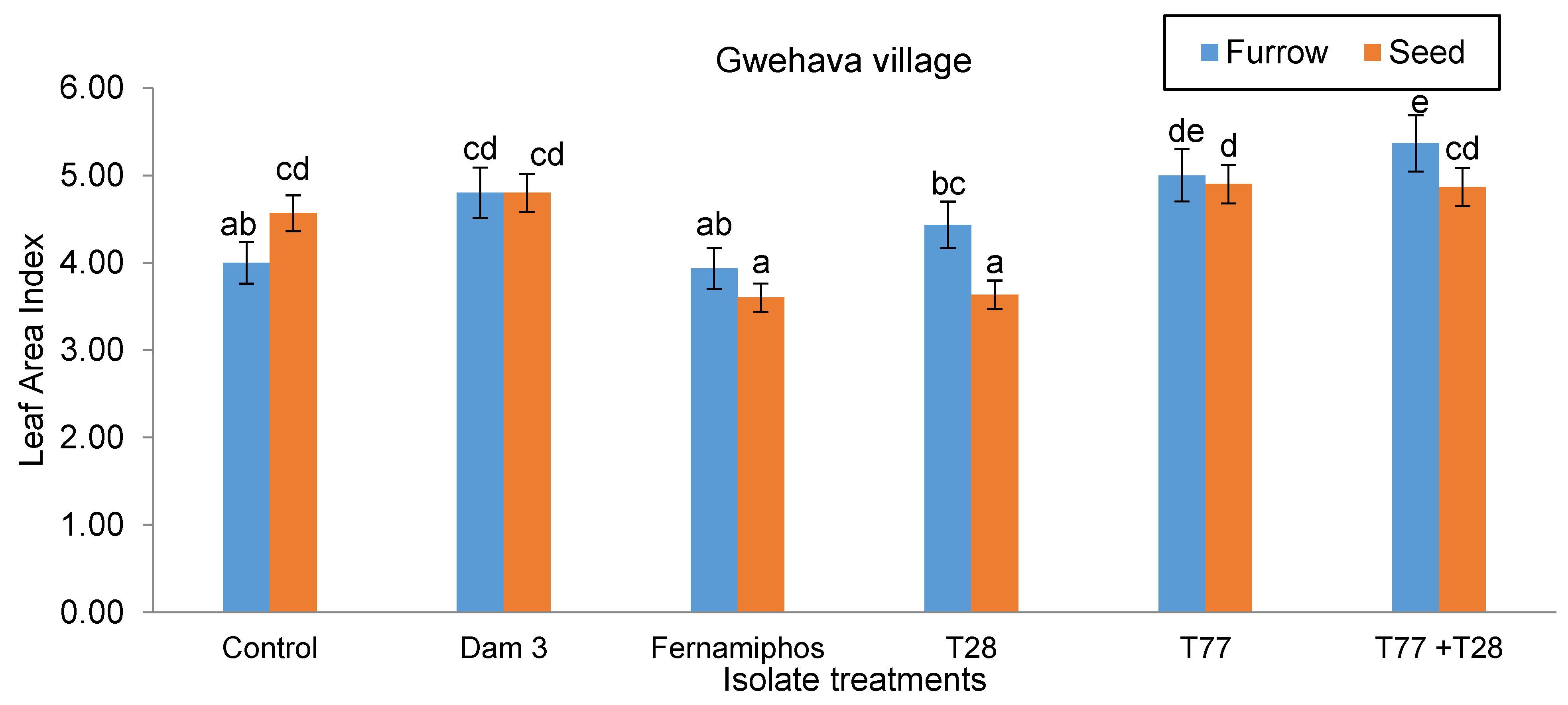

Table 1.
Fusarium wilt disease severity scale.
| Score | Degree of severity |
| 0 | No symptoms |
| 1 | 5% leaves yellow and wilted, very limited |
| 2 | 6-10% leaves yellow and wilted, limited wilting |
| 3 | 11-20% leaves yellow and wilted, moderate wilting |
| 4 | 21-50% leaves yellow and severe wilting |
| 5 | More than 50% of leaves are yellow, severe wilting and/or plant deaths |
Table 2.
Taylor and Sasser egg mass scale [35].
Table 2.
Taylor and Sasser egg mass scale [35].
| Score | Number of egg masses |
| 0 | 0 |
| 1 | 1-2 |
| 2 | 3-10 |
| 3 | 11-30 |
| 4 | 31-100 |
| 5 | 100 |
Table 3.
Daulton and Nasbaum scale [36].
Table 3.
Daulton and Nasbaum scale [36].
| Score | Degree of galling |
| 0 | Free from galls |
| 1 | < 5 galls |
| 2 | Trace to 25 galls |
| 3 | 20-100 galls |
| 4 | Numerous galls, mostly discrete |
| 5 | Numerous galls, many coalesced |
| 6 | Heavy, mostly coalesced |
| 7 | Very heavy, mass invasion, slight root growth |
| 8 | Mass invasion, no root development |
Table 4.
Effect of Trichoderma isolates on egg hatching of M. javanica.
| Isolate treatment | Number of hatched eggs per 1.0 ml treatment solution | ||||
| Day 2 | Day 3 | Day 4 | Day 5 | ||
| T77 T28 Dam 3 Control |
0a 0a 0a 69.34b |
4.00a 6.66a 10.66a 89.34b |
8.00a 16.00b 18.00b 132.66c |
12.00a 20.00b 24.00b 193.34c |
|
| P value | <0.001 | <0.001 | <0.001 | <0.001 | |
| LSD0.05 | 9.476 | 9.964 | 5.752 | 4.348 | |
| CV (%) | 29.00 | 19.10 | 7.10 | 3.70 | |
Table 5.
Effect of isolate treatments on mortality of M. javanica juveniles.
| Isolate | Nematode mortality (%) | ||||
| Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | |
| T77 T28 Dam 3 Control |
38.00c 26.67a 32.67b 0d |
50.00b 41.33a 44.00a 0d |
66.67a 55.33a 60.00a 0d |
80.67b 70.67a 75.33a 0d |
96.00c 92.67a 94.00b 0d |
| P-value | < 0.001 | < 0.001 | < 0.001 | < 0.001 | < 0.001 |
| LSD0.05 | 4.482 | 4.348 | 13.13 | 6.79 | 1.087 |
| CV (%) | 9.8 | 6.80 | 15.30 | 6.40 | 0.80 |
Table 6.
Disease prevalence of Fusarium wilt at the DRSS and Gwehava village sites.
| Isolate treatment | Disease incidence (%) | Disease severity | ||
| DRSS site | Gwehava village | DRSS site | Gwehava village | |
| Control Dam 3 Fenamiphos T28 T77 T28 + T77 |
27.83d 15.17abc 16.83bc 17.17c 14.83ab 14.50a |
21.17c 17.17a 18.83b 19.17b 16.83a 16.50a |
3.167c 0.500ab 0.667ab 1.000b 0.333a 0.667ab |
2.000c 0.550ab 0.717ab 1.050b 0.387a 0.717ab |
| P-value | < 0.001 | < 0.001 | < 0.001 | < 0.001 |
| LSD0.05 | 2.183 | 1.798 | 0.6564 | 0.6136 |
| CV (%) | 10.30 | 9.00 | 21.90 | 19.50 |
Table 7.
Interaction table of isolate treatment and isolate application technique on RKN galling at Gwehava village.
Table 7.
Interaction table of isolate treatment and isolate application technique on RKN galling at Gwehava village.
| Isolate application technique (number of galls per plant) | ||
| Isolate treatment | Furrow | Seed |
| Control Dam 3 Fenamiphos T28 T77 T28 + T77 |
2.000b 1.000a 1.333a 1.333a 1.333a 1.333a |
3.000c 1.333a 1.667a 1.330a 1.000a 1.000a |
| P-value | 0.047 | |
| LSD0.05 | 0.6368 | |
| CV (%) | 25.5 | |
Table 8.
Effect of isolate treatments on average potato tuber weight and final yield.
| Isolate Treatment | Tuber size (g) | Tuber yield (t/ha) | ||
| DRSS site | Gwehava village | DRSS site | Gwehava village | |
| Control Dam 3 Fenamiphos T28 T77 T28 + 77 |
119.17a 127.33bc 129.27cd 125.80b 129.20cd 130.12d |
115.50a 123.67b 126.13c 122.40b 128.12d 129.23e |
12.08a 27.43bc 30.58cd 25.13b 34.50e 33.40de |
15.18a 29.54bc 32.08c 28.43b 32.50c 32.00c |
| P value | < 0.001 | < 0.001 | < 0.001 | < 0.001 |
| LSD | 2.201 | 1.904 | 3.165 | 3.128 |
| CV (%) | 14.00 | 12.00 | 10.50 | 9.90 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



