
Submitted:
29 April 2025
Posted:
29 April 2025
You are already at the latest version
Abstract
The study on the isolation, identification, and characterization of probiotic properties of lactic acid bacteria strains from the rose blossom of Rosa damascena Mill. is crucial for discovering novel, plant-derived probiotics with potential health benefits and applications in food, medicine, and cosmetics. Nine lactic acid bacteria (LAB) strains were isolated from rose blossom of Rosa damascena Mill. and they were identified to the species level by applying physiological-biochemical (API 50 CHL) and molecular-genetic (16S rRNA gene sequencing). The isolates were identified as representatives of the species Lactobacillus helveticus, Lactobacillus acidophilus and Lactiplantibacillus plantarum. Some probiotic properties of the newly isolated and identified LAB strains were examined: antimicrobial activity against pathogens by the agar-diffusion method with wells and antibiotic resistance profile by the agar-diffusion method with paper discs. The studied LAB strains demonstrated high antimicrobial activity against the pathogenic bacteria Escherichia coli ATCC 25922, Staphylococcus aureus ATCC 25923, Salmonella abony NTCC 6017, Proteus vulgaris J, Listeria monocytogenes ATCC 19115, Enterococcus faecalis ATCC 29212 and were resistant to most of the antibiotics used in clinical practice, which in turn opens up the possibility of their joint inclusion in complex therapy in the composition of probiotics. A batch fermentation process was carried out with Lactiplantibacillus plantarum 5/20 and the kinetic parameters of the fermentation process to obtain a concentrate with a high content of viable cells (1013cfu/cm3) were determined. The obtained concentrate weas freeze-dried and lyophilic preparations with a high content of viable cells (over 1012 cfu/g) were obtained. This research on Rosa damascena Mill.-isolated LAB strains could uncover valuable LAB strains with significant probiotic properties. These strains would be suitable for various applications in the composition of starters for functional foods and beverages as well as probiotic preparations, showcasing the untapped potential of plant-associated microbiota.
Keywords:
lactic acid bacteria
; Rosa damascena Mill.
; isolation
; identification
; sequencing
; antimicrobial activity
; pathogen
; antibiotic
; probiotic
1. Introduction
Lactic acid bacteria (LAB) are recognized for their numerous probiotic properties, which can significantly enhance the health benefits of fermented foods. The isolation of LAB strains from diverse natural environments, such as rose blossom of Rosa damascena Mill., opens up new perspectives for comprehending their probiotic potential and applications [1].
LAB antimicrobial properties are particularly noteworthy [2]. The role of LAB in metabolic processes, particularly in carbohydrate fermentation, further emphasizes their probiotic benefits. LAB convert fermentable sugars into lactic acid and other metabolites, which can lower the gut pH, thus creating an inhospitable environment for pathogens and consequently inhibiting their growth, and enhancing nutrient absorption due to improving digestibility and nutrient bioavailability for the host [3]. Besides lactic acid, LAB also produce various metabolites, such as bacteriocins, hydrogen peroxide, and other organic acids, which can also inhibit the growth of pathogenic microorganisms [2,4,5]. The unique production of these antimicrobial compounds allows LAB to outcompete harmful microorganisms for nutrients and colonization sites within the gut [4,6]. Thus, they not only enhance food safety by preventing spoilage but also contribute to overall health by maintaining a balanced gut microbiota.
The isolation of probiotic LAB strains from Rosa damascena Mill. may present novel opportunities for developing functional foods with enhanced protective effects against spoilage microorganisms and foodborne pathogens. The ability of these strains to produce lactic acid and other antimicrobial agents while demonstrating resistance to adverse conditions suggests their potential utility in the development of novel probiotic-rich foods, such as fortified beverages, fermented vegetables, and even non-dairy alternatives [7]. Therefore, the exploration of LAB from such a unique source as rose blossom not only broadens the scope of probiotic research but also paves the way for innovative applications in functional food industries.
The significance of microbial interactions, particularly between LAB and other microorganisms, can amplify the health benefits provided by these probiotics. The use of LAB as starters in fermentation processes has been associated with improved sensory profiles and biological activity in food products [8].
Furthermore, laboratory screening of various LAB strains for specific probiotic properties can reflect a more directed approach to developing functional foods tailored to consumer health needs. For instance, selecting strains with pronounced antimicrobial activity against common gut pathogens while maintaining gut epithelial adherence can produce more effective probiotics [9,10].
Research into the probiotic potential of natural and indigenous LAB strains is essential as they bear unique genetic makeups that may enhance their functional properties. Cultivating and utilizing strains specifically adapted to local environments could lead to the development of regionally significant functional foods that cater to the specific health needs of local populations [11]. This aligns with broader efforts toward sustainability and resilience in functional food production.
Last but not least, as consumer interest in functional foods continues to rise, the exploration of traditional sources such as Rosa damascena Mill. for LAB isolation could serve both health and economic purposes. Combining ancient ethnobotanical knowledge with modern microbiological techniques may yield new probiotic sources that not only benefit health but also honor cultural practices surrounding food production and fermentation. The exploration of LAB from such unique sources can indeed stimulate innovation in the field while preserving sustainable practices. Continued research is necessary to isolate and identify new LAB strains from plant sources and to explore their probiotic properties in order to identify specific applications in the development of both probiotic formulations and food bio-preservation strategies.
The objectives of present study were to isolate and identify LAB strains from the rose blossom of Rosa damascena Mill. and to examines some of their probiotic properties, namely their antimicrobial activity against spoilage microorganisms and pathogens and their antibiotic resistance profile.
2. Materials and Methods
2.1. Microorganisms
- Lactic acid bacteria. The research in the present work was carried out with 11 LAB strains, designated as: LAB 4/20, LAB 12/20, LAB 5/20, LAB 6/20, LAB 8/20, LAB 9/20, LAB 10/20, LAB 13/20, LAB 16/20, LAB 19/20, LAB 22/20, isolated from rose blossom of Rosa damascena Mill.
- Pathogenic test-microorganisms for the determination of the antimicrobial activity against pathogens. Escherichia coli ATCC 25922, Staphylococcus aureus ATCC 25923, Salmonella abony NTCC 6017, Proteus vulgaris J, Listeria monocytogenes ATCC 19115, Enterococcus faecalis ATCC 29212.
2.2. Nutrient Media:
- MRS-broth (Merck); MRS-agar (Merck); 0.5 % NaCl solution (Merck); LAPTg10-broth; LAPTg10-agar; LBG-broth; LBG-agar
- LAPTg10–broth. Composition (g/dm3): peptone – 15; yeast extract – 10; tryptone – 10; glucose – 10. The pH was adjusted to 6.6 – 6.8 and Tween 80 - 1cm3/dm3 was added. Sterilization - 20 min at 121 °С.
- LAPTg10-agar. Composition (g/dm3): LAPTg10-broth, agar - 20. Sterilization - 20 min at 121 °С.
- LBG-broth. Composition (g/dm3): tryptone - 10; yeast extract - 5; NaCl - 10; glucose - 10 Sterilization - 20 min at 121 °C.
- LBG-agar. Composition (g/dm3): tryptone - 10; yeast extract - 5; NaCl - 10; glucose - 10 agar - 15. Sterilization - 20 min at 121 °C
2.3. Physiological and Biochemical Methods
2.3.1. Isolation of Lactic Acid Bacteria
Lactic acid bacteria were isolated from the rose blosssoms of Rosa damascena Mill. Isolation was carried out using the combined microbiological approach and the use of various selective conditions and nutrient media (MRS-broth and MRS-agar). Isolation of LAB strains was carried out by morphological characteristics and by classical phenotypic methods.
2.3.2. Determination of the Titratable Acidity
The Thörner method was used to determine the acid-forming ability of microorganisms. One °T is equal to 1 cm3 of 0.1 N NaOH, used to neutralize an equivalent amount of organic acid contained in 100 cm3 of culture medium. The method is based on the titration of the sample with 0.1 N NaOH. For this purpose, 10 cm3 of each sample was taken and 20 cm3 of distilled water were added. The mixture was titrated with 0.1 N NaOH using phenolphthalein as an indicator until the appearance of a pale pink color that persists for one minute.
2.3.3. Determination of the Number of Viable Microorganisms
Appropriate tenfold dilutions of the sample for analysis in saline were prepared. 0,1 cm3 from the last three were spread plated on the corresponding solid nutrient medium. The inoculated Petri dishes were cultured for 48 to 72 hours at the optimum temperature for the growth of the corresponding microorganism until the appearance of countable single colonies appear. The number of the single colonies was used to estimate the number of viable microorganisms.
2.3.4. Determination of the Biochemical Profile
The API 50 CHL system (BioMerieux SA, France) was used according to the manusfacturer’s instructions. The results obtained were processed with apiweb® identification software.
2.3.5. Determination of the Antibiotic Susceptibility Profile
The antibiotic susceptibility profile was determined by the disk diffusion method [12]. A fresh 24-hour culture of each strain in MRS-broth was adjusted to a 0.5 McFarland standard and inoculated onto MRS-agar plates.
The paper discs were impregnated with antibiotics targeting:
- Cell wall synthesis: Penicillin, Ampicillin, Oxacillin, Piperacillin, Amoxicillin, Bacitracin, Vancomycin
- Protein synthesis: Tetracycline, Doxycycline, Tobramycin, Amikacin, Gentamicin, Erythromycin, Chloramphenicol, Lincomycin
- DNA synthesis and cell division: Nalidixic acid, Ciprofloxacin, Novobiocin, Rifampin
The Petri dishes were incubated for 48 hours at 37±1 °C. The diameter (in mm) of the formed inhibition zones around each of the antibiotic discs was recorded. The following designations were used when reporting the results: R – resistant (dIZ < 8 mm), SR – intermediately sensitive (dIZ 8 - 16 mm), S – sensitive (dIZ > 16 mm).
2.3.6. Determination of the Antimicrobial Activity Against Pathogens – Agar-Diffusion Method with Wells
To determine the antimicrobial activity of the studied LAB strains against pathogenic microorganisms, 3 samples for each LAB strain were prepared: culture liquid (CL), biomass in saline solution (BSS), cell-free supernatant (CFSN) and neutralized cell-free supernatant (NCFSN) (pH=6.5), obtained from a 24-hour culture of the respective LAB strains. The antimicrobial activity was tested against the following test microorganisms: Escherichia coli ATCC 25922, Staphylococcus aureus ATCC 25923, Salmonella abony NTCC 6017, Proteus vulgaris J, Listeria monocytogenes ATCC 19115, Enterococcus faecalis ATCC 29212.
Each of the test-microorganisms (suspension in saline solution with a concentration of 107 cfu/cm3) was inoculated into Petri dishes with LBG-agar medium and after solidification of the agar, wells (dwell=7 mm) were prepared. 0.1 cm3 of CL, BSS and NCFSN were pipetted into the wells and the Ptri dishes were incubated for 1 h at 4±2ºС for the pipetted samples to diffuse into the medium. Then the Petri dishes with the test-microorganisms were incubated at 37±1 °С for 24 h, after which the diameter of the inhibition zones in mm were recorded.
2.4. Molecular-Genetic Methods
2.4.1. Sequencing of the 16S rRNA Gene
The sequencing of the 16S rRNA gene was performed by the Sanger method by the “Macrogen Europe Laboratory”, the Netherlands [13]. The forward and reverse partial sequences obtained during the sequencing of the 16S rRNA gene were assembled using the CLC Sequence Viewer software. The entire sequences of the 16S rRNA gene were compared with the available sequences in the online GenBank database using the online BLASTn software and the species affiliation of the strains was determined with the corresponding percentage of coincidence between the sequence of the LAB strain and the reference strain from the online database.
2.5. Batch Cultivation
The cultivation of Lactiplantibacillus plantarum 5/20 was carried out in a laboratory bioreactor with a geometric volume of 2 dm3 and a working volume of 1.5 dm3. The control and monitoring of the main fermentation parameters was carried out by a “Sartorius A2” control device, which included control loops for the stirring rate, temperature, pH, etc. The cultivation of lactic acid bacteria was carried out in the following order: the apparatus was washed, then filled with 0.3% neomycin solution for cold sterilization for 24 h. After the sterilization time had elapsed, the apparatus was washed several times with sterile saline solution or distilled water, after which it was ready to carry out the fermentation process. The MRS-broth medium was loaded into the apparatus sterilely, using a peristaltic pump. The process of batch cultivation was carried out in MRS-broth medium. The nutrient medium was sterilized at 121 °С for 20 min. After cooling to 39-40 °С, the prepared medium in the bioreactor was inoculated with 5% (v/v) inoculum from a fresh 24-hour culture of the LAB strain in MRS-broth medium. The cultivation process was carried out at 37 °С, stirring rate of 150 rpm, without aeration. The duration of the cultivation was 24 h, with periodic samples of the culture fluid being taken for analysis of the total number of viable cells of the respective strain (cfu/cm3) and the titratable acidity [14,15].
2.6. Modeling of the Process Kinetics and Identification of the Model Parameters
Since the main target product of the fermentation process is the amount of biomass, the logistic curve equation (equation (1)), which contains a clear biological meaning, was used to model the growth kinetics. The duration of the induction period and the rate constant of adaptation were determined by equation (2).
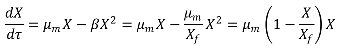
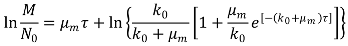 where: µm is the maximum specific growth rate, h-1; Xin and Xf are the initial and final concentration of viable cells, cfu/cm3; β – coefficient of intrapopulation competition cfu/cm3.h; M – current biomass concentration, cfu/cm3; N0 – initial biomass concentration, cfu/cm3; τа – induction period, h; k0 – rate constant of cell adaptation to the environment and cultivation conditions, h-1;
where: µm is the maximum specific growth rate, h-1; Xin and Xf are the initial and final concentration of viable cells, cfu/cm3; β – coefficient of intrapopulation competition cfu/cm3.h; M – current biomass concentration, cfu/cm3; N0 – initial biomass concentration, cfu/cm3; τа – induction period, h; k0 – rate constant of cell adaptation to the environment and cultivation conditions, h-1;
The logistic curve model (1) was solved numerically using the 4th order Runge-Kutta method. For the identification of the model parameters, the Solver function in Excel was used. The model parameters were determined by minimizing the square of the difference between the experimental data and the data obtained from the corresponding model. The parametric identification of the model (2) was performed in the Curve Expert Professional software product through nonlinear regression [16].
2.7. Freeze-Drying
The freeze-dried product was technologically obtained by a combined method of technological treatment - immobilization and freeze-drying, at the following stages of the process - primary treatment, freezing and freeze-drying. The immobilization matrix used was hydrocolloid and contained a 4% solution of highly esterified apple pectin and a 1.2% solution of sodium alginate in a ratio of 1:1, respectively. The mechanical method of immobilization of microbial cells was applied, i.e., cell attachment to the carrier was carried out by mixing the cell suspension with the carrier (hydrocolloid matrix) at the following sequence of technological stages: 1. Obtaining the cell suspension by culturing of the strain.; 2. Mixing of the obtained cultural suspension with the polymer solution at a temperature of 40 – 45 °C and homogenization for 1 hour 30 minutes in a reactor with a stirring speed of 500 rpm [17]. The material thus prepared was subjected to lyophilization in a sublimation installation of the company “Hochvakuum-TG – 16.50” with contact heating of the plates at the Institute of Cryobiology and Food Technology – Sofia at the following parameters: layer thickness – 11 mm; freezing temperature - -30 °C to – 35 °C; freezing by slow method – 0.05 – 0.06 °C/sec; eutectic temperature - – 37 °C; solidification temperature - – 40.00 °C; load on the supporting surface – 10.60 kg/m2; temperature in the desublimator - from – 55 °C to – 60 °C before drying and from - 65.0 °C to – 70 °C during post-drying; partial pressure – 30 Pa to 34 Pa; chamber pressure – 20 Pa to 27 Pa; post-drying temperature – 30 °C; maximum product temperature – 30 °C; residual moisture content – from 1.37 % to 3.09 %; process duration – 14 to 16 hours; storage conditions – in bags of three-layer aluminum foil, sealed under vacuum; stored at a relative humidity of no more than 35 % at 20 – 22 °C.
Before drying, the installation was prepared for loading by cooling the plates to a minimum temperature of –20 °C to -45 °C. The desublimator of the installation was cooled to a minimum temperature of –50 °C to –60 °C, which ensured a sufficiently low partial pressure of water vapor above the desublimator in the initial period and created favorable conditions for mass transfer.
After lyophilization, the lyophilizate was ground in an “Erveka” granulator. Thus processed, the lyophilized bioproduct was packed in a package made of three-layer aluminum foil, hermetically sealed under vacuum.
2.8. Microbiological Studies of the Lyophilized Concentrate
2.8.1. Microbiological Status of Native and Lyophilized Samples
According to, BDS EN ISO 4833: 2004 [18].
Indicators:
- lactic acid bacteria, CFU/g [19];
- total number of mesophilic aerobic and facultative anaerobic bacteria – CFU/g [20];
- Escherichia coli in 0.1 g of the product [21];
- pathogenic microorganisms, including Salmonella sp. in 25.0 g of the product [22];
- coagulase-positive staphylococci in 1.0 g of the product [23];
- sulfite-reducing clostridia in 0.1 g of product [24];
- microscopic mold spores, CFU/g [25];
- yeast, CFU/g [25];
2.9. Processing of the Results
The data from the triplicate experiments were processed using MS Office Excel 2013 software products, using statistical functions to determine the standard deviation and the maximum error of the estimate at significance levels of α<0.05.
3. Results and Discussion
3.1. Isolation, Identification and Selection of LAB Strains from the Rose Blossom of Rosa damascena Mill L.
3.1.1. Isolation and Identification of LAB Strains
- A. Phenotypic characteristics of the newly isolated LAB strains
The characterization of the newly isolated strains began with an assessment of the purity of the culture, macroscopic and microscopic morphological control. The micro - and macromorphological characteristics of the 11 LAB isolates from rose blossom of Rosa damascena Mill. are presented in Table. 1. When cultivated on MRS-agar medium, the isolated strains grew in the form of small, milky-white colonies with a star-shaped or biconvex lenticular shape, which were easily separated from the medium. The isolated strains are Gram-positive (Gr (+), non-spore-forming rods.
- B. Physiological and biochemical characteristics of the isolated strains
After characterization according to the main morphological criteria (Table 1), the newly isolated strains were subjected to determination of their biochemical profile with the rapid identification system API 50 CHL, and their ability to assimilate 49 carbon sources included in the system was determined (Table 2a and Table 2b). Based on the studies conducted with the API 50 CHL kits, the isolated strains were assigned to different LAB species with the corresponding percentage of confidence (Table 3).
The increasing use of molecular methods allows for more accurate, rapid, and reproducible differentiation between closely related species, which are difficult to distinguish based on phenotypic characteristics alone [26,27]. The similarity in the biochemical profiles of phylogenetically close species, as well as the influence of some factors on the metabolic properties of lactic acid bacteria, necessitate additional characterization by applying molecular genetic methods [26,28].
- C. Molecular - taxonomic characterization. Genotypic characteristics of the studied LAB strains. Sequencing of the 16S rRNA gene
For the complete species identification of the newly isolated LAB strains, a genetic method for genotyping was used - sequencing of the 16S rRNA gene. The results of the 16S rDNA sequence analysis refer LAB 4/20 to Lactobacillus helveticus with 99% similarity between the 16S rDNA sequence of LAB 4/20 and the partial 16S rDNA sequence of Lactobacillus helveticus NBRC 15019 (Figure 1, Table 4), LAB 5/20 to Lactiplantibacillus plantarum with 99% similarity between the 16S rDNA sequence of LAB 5/20 and the partial 16S rDNA sequence of Lactiplantibacillus plantarum NBRC 15891 (Figure 2, Table 4), LAB 6/20 to Lactobacillus helveticus with 99% similarity between the 16S rDNA sequence of LAB 6/20 and the partial 16S rDNA sequence of Lactobacillus helveticus NBRC 15019 (Figure 3, Table 4), LAB 8/20 to Lactobacillus helveticus with 99% similarity between the 16S rDNA sequence of LAB 8/20 and the partial 16S rDNA sequence of Lactobacillus helveticus NBRC 15019 (Figure 4, Table 4), LAB 9/20 – to Lactobacillus helveticus with 99% similarity between the 16S rDNA sequence of LAB 9/20 and the partial 16S rDNA sequence of Lactobacillus helveticus NBRC 15019 (Figure 5, Table 4), LAB 10/20 – to Lactobacillus acidophilus with 99% similarity between the 16S rDNA sequence of LAB 10/20 and the partial 16S rDNA sequence of Lactobacillus acidophilus NBRC 13951 (Figure 6, Table 4), LAB 12/20 – to Lactobacillus helveticus with 99% similarity between the 16S rDNA sequence of LAB 12/20 and the partial 16S rDNA sequence of Lactobacillus helveticus NBRC 15019 (Figure 7, Table 4), LAB 13/20 – to Lactiplantibacillus plantarum with 99% similarity between the 16S rDNA sequence of LAB 13/20 and the partial 16S rDNA sequence of Lactiplantibacillus plantarum NBRC 15891 (Figure 8, Table 4), LAB 16/20 – to Leuconostoc mesenteroides with 99% similarity between the 16S rDNA sequence of LAB 19/20 and the partial 16S rDNA sequence of Leuconostoc mesenteroides ATCC 8293 (Figure 9, Table 4); LAB 19/20 – to Lactiplantibacillus plantarum with 99% similarity between the 16S rDNA sequence of LAB 19/20 and the partial 16S rDNA sequence of Lactiplantibacillus plantarum NBRC 15891 (Figure 10, Table 4), LAB 22/20 – to Leuconostoc mesenteroides with 99% similarity between the 16S rDNA sequence of LAB 22/20 and the partial 16S rDNA sequence of Leuconostoc mesenteroides ATCC 8293 (Figure 11, Table 4).
3.2. Characterization of the Probiotic Potential of the Isolated Lactobacilli
3.2.1. Antimicrobial Activity of the Isolated LAB Strains Against Pathogenic Microorganisms
In a series of experiments, the antimicrobial activity against pathogenic microorganisms of the newly isolated LAB strains – Lactobacillus helveticus 4/20, Lactiplantibacillus plantarum 5/20, Lactobacillus helveticus 6/20, Lactobacillus helveticus 8/20, Lactobacillus helveticus 9/20, Lactobacillus acidophilus 10/20, Lactobacillus helveticus 12/20, Lactiplantibacillus plantarum 13/20 and Lactiplantibacillus plantarum 19/20, isolated from rose blossom of Rosa damascena Mill. by the agar diffusion method with wells was studied (Table 5.1 and Table 5.2). All of the studied LAB strains had pronounced antimicrobial activity against the pathogenic bacteria Escherichia coli ATCC 25922, Staphylococcus aureus ATCC 25923, Salmonella abony NTCC 6017, Proteus vulgaris J, Listeria monocytogenes ATCC 19115, Enterococcus faecalis ATCC 29212 (Table 5.1 and Table 5.2). When neutralizing the organic acids released into the medium by the LAB strains, no antimicrobial activity was found on the growth of pathogenic bacteria that cause foodborne infections and intoxications (Table 5.1 and Table 5.2) indicating that the studied LAB strains did not produce any other substances besides organic acids that inhibit the growth of the test-pathogenic bacteria.
Lactobacillus helveticus 4/20, Lactobacillus helveticus 6/20, Lactobacillus helveticus 8/20 and Lactobacillus helveticus 9/20 exhibited the highest antimicrobial activity against Listeria monocytogenes and Enterococcus faecalis; Lactobacillus helveticus 4/20 and Lactobacillus helveticus 12/20 also manifested high antimicrobial activity against Staphylococcus aureus (Table 5.1). Lactiplantibacillus plantarum 5/20 and Lactiplantibacillus plantarum 13/20 showed the highest antimicrobial activity against Listeria monocytogenes, Staphylococcus aureus, Enterococcus faecalis and Proteus vulgaris. Lactobacillus acidophilus 10/20 and Lactiplantibacillus plantarum 19/20 exhibited significantly lower antimicrobial activity than all newly isolated LAB strains (Table 5.2). Of the five Lactobacillus helveticus strains, Lactobacillus helveticus 12/20 demonstrated the highest inhibitory activity against Escherichia coli (Table 5.1). Among the representatives of the species Lactiplantibacillus plantarum, Lactiplantibacillus plantarum 5/20 manifested the highest antimicrobial activity against Escherichia coli (Table 5.2). Of the five Lactobacillus helveticus strains, Lactobacillus helveticus 4/20 exhibited the highest inhibitory activity against Staphylococcus aureus (Table 5.1). Among the representatives of the species Lactiplantibacillus plantarum, Lactiplantibacillus plantarum 5/20 and Lactiplantibacillus plantarum 13/20 showed the highest antimicrobial activity against Escherichia coli (Table 5.2). Of the five strains of Lactobacillus helveticus, Lactobacillus helveticus 4/20 and Lactobacillus helveticus 12/20 demonstrated the highest inhibitory activity against Salmonella abony (Table 5.1).
Among the representatives of the species Lactiplantibacillus plantarum, Lactiplantibacillus plantarum 5/20 exhibited the highest antimicrobial activity against Salmonella abony (Table 5.2). Of the five strains Lactobacillus helveticus, Lactobacillus helveticus 4/20 and Lactobacillus helveticus 12/20 demonstrated the highest inhibitory activity against Proteus vulgaris (Table 5.1). Among the representatives of the species Lactiplantibacillus plantarum, Lactiplantibacillus plantarum 5/20 showed the highest antimicrobial activity against Proteus vulgaris (Table 5.2). Of the five Lactobacillus helveticus strains, Lactobacillus helveticus 6/20, Lactobacillus helveticus 9/20 and Lactobacillus helveticus 12/20 manifested the highest inhibitory activity against Listeria monocytogenes (Table 5.1). Among the representatives of the species Lactiplantibacillus plantarum, Lactiplantibacillus plantarum 5/20 exhibited the highest antimicrobial activity against Listeria monocytogenes (Table 5.2). Of the five Lactobacillus helveticus strains, Lactobacillus helveticus 9/20 and Lactobacillus helveticus 12/20 demonstrated the highest inhibitory activity against Enterococcus faecalis (Table 5.1). Among the representatives of the species Lactiplantibacillus plantarum, Lactiplantibacillus plantarum 5/20 and Lactiplantibacillus plantarum 13/20 exhibited the highest antimicrobial activity against Enterococcus faecalis (Table 5.2). The most pronounced antimicrobial activity against the test microorganisms included in the study was demonstrated by Lactobacillus helveticus 12/20 among the representatives of the species Lactobacillus helveticus and Lactiplantibacillus plantarum 5/20 among the representatives of the species Lactiplantibacillus plantarum (Table 5.1 and Table 5.2.).
When examining the antimicrobial activity of each newly isolated LAB strain against each pathogen included in the experiment, inhibition zones were observed when the culture liquid (CL), biomass in saline solution (BSS) and the cell-free supernatant (CFSN) were pipetted. For some strains, it was found that the CL and BSS had greater antimicrobial activity than the CFSN, which means that LAB suppress pathogenic microorganisms due to both competition for nutrients between them and inhibition by the lactic and other organic acids produced, as a result of which the pH decreases.
The present study highlights the significant antimicrobial potential of LAB strains isolated from the rose blossoms of Rosa damascena Mill., as evaluated through the well diffusion method. All tested LAB strains exhibited pronounced inhibitory activity against a spectrum of foodborne pathogenic bacteria, including Escherichia coli, Staphylococcus aureus, Salmonella abony, Proteus vulgaris, Listeria monocytogenes, and Enterococcus faecalis. These findings reinforce the growing body of evidence supporting LAB as effective bio-preservatives and potential candidates for the development of functional foods and probiotics.
A key observation from this study is that the antimicrobial activity of the LAB strains was abolished upon neutralization of their organic acid content. This clearly suggests that the antimicrobial effects observed are primarily attributable to organic acid production—mainly lactic acid—and not due to bacteriocins, hydrogen peroxide, or other antimicrobial metabolites. This mode of inhibition, largely through acidification of the medium, aligns with previous findings reported for Lactobacillus and Lactiplantibacillus species, where the decrease in environmental pH creates unfavorable conditions for pathogen proliferation.
Among the tested strains, Lactobacillus helveticus 12/20 and Lactiplantibacillus plantarum 5/20 consistently demonstrated the most potent antimicrobial activity against nearly all tested pathogens. Notably, L. helveticus 12/20 exhibited strong inhibition against E. coli, S. aureus, Salmonella abony, Proteus vulgaris, L. monocytogenes, and E. faecalis, indicating its broad-spectrum efficacy. Likewise, Lp. plantarum 5/20 was particularly effective, showing significant inhibitory activity against S. aureus, L. monocytogenes, and P. vulgaris. These results underscore the strain-specific nature of antimicrobial efficacy, even within the same species, further emphasizing the necessity of individual assessment of LAB candidates for food and health applications.
The comparative analysis of different LAB species and strains revealed a pattern: L. helveticus strains, particularly L. helveticus 4/20, L. helveticus 6/20, L. helveticus 9/20, and L. helveticus 12/20, were more effective against Gram-positive bacteria such as L. monocytogenes and E. faecalis, while Lp. plantarum strains, especially Lp. plantarum 5/20 and Lp. plantarum 13/20, showed a broader spectrum, with potent activity against both Gram-positive and Gram-negative bacteria, including E. coli and S. abony. This strain-level variability could be attributed to differences in organic acid profiles, acid tolerance, and metabolic capabilities.
The findings also underscore the potential of underexplored natural sources, such as the rose blossom of Rosa damascena Mill., as reservoirs of beneficial microorganisms. These results are especially relevant for the food industry, where there is a growing demand for natural preservatives and health-promoting microbial strains. The ability of certain strains to effectively inhibit foodborne pathogens suggests their promising application as components in biopreservatives, starter cultures, or next-generation probiotics.
The present study contributes valuable insights into the antimicrobial potential of novel LAB strains, reaffirming the critical importance of strain-specific evaluation. Future research should aim to characterize the organic acid profiles of these strains, explore their performance in food matrix systems, and evaluate their safety and efficacy in vivo.
3.2.2. Antibiotic Resistance of the Isolated LAB Strains
Knowledge of the antibiotic resistance of LAB with probiotic potential is essential. On the one hand, this can be considered as a significant criterion in the selection of probiotic cultures, due to the possibility of conducting combined therapy with an antibiotic and a probiotic, in order to restore the normal microflora of the gastrointestinal tract and/or urogenital tract [29]. On the other hand, a number of authors express the hypothesis that bacteria normally found in the body, including LAB, can serve as a source of antibiotic resistance genes, transferring them to various pathogenic microorganisms [30]. Therefore, one of the conditions for the selection of potentially probiotic strains is knowledge of their antibiotic sensitivity.
Thus, 20 antibiotics with different mechanisms of action from the main groups used in medical practice were selected and the sensitivity of the newly isolated LAB strains was tested. The results obtained from the studies conducted using the agar diffusion method of Bauer et al., 1966 [12,31] for 24 h are summarized in Table 6.1 and Table 6.2.
The antibiotic susceptibility patterns observed among the Lactobacillus helveticus and Lactiplantibacillus plantarum strains tested in this study revealed substantial variability, indicating the strain-specific nature of antibiotic resistance and susceptibility in this species. Given the increasing interest in LAB as probiotics and components of food starters, understanding their antibiotic resistance profiles is critical from both safety and therapeutic standpoints (Table 6.1 and Table 6.2).
When it comes to the Lactobacillus helveticus strains, among the antibiotics targeting cell wall synthesis, a heterogeneous resistance pattern was observed. L. helveticus 4/20 and L. helveticus 9/20 exhibited resistance to multiple β-lactam antibiotics including penicillin, ampicillin, and oxacillin, while strains such as L. helveticus 8/20 demonstrated broad sensitivity within this group (Table 6.1). Interestingly, vancomycin resistance was detected in L. helveticus 4/20, L. helveticus 9/20, and L. helveticus 12/20, which is noteworthy since LAB are often intrinsically resistant to glycopeptides due to the absence of the D-Ala-D-Lac target in their peptidoglycan precursors. The sensitivity of L. helveticus 6/20 to vancomycin may suggest a deviation from typical intrinsic resistance or experimental variability, warranting further genetic investigation. Regarding protein synthesis inhibitors, all L. helveticus strains showed resistance to lincomycin and most to amikacin, consistent with known intrinsic resistance mechanisms in LAB, including poor drug uptake or active efflux. However, variable responses to tetracyclines were observed: while L. helveticus 4/20 and L. helveticus 6/20 were resistant, L. helveticus 12/20 was sensitive.
Chloramphenicol sensitivity was broadly maintained across strains, aligning with its known efficacy against a wide range of Gram-positive bacteria. Erythromycin sensitivity in L. helveticus 6/20, L. helveticus 8/20, and L. helveticus 12/20 is particularly important, as macrolides are frequently used in clinical settings; resistance in L. helveticus 9/20 could reflect either acquired resistance or a variation in ribosomal binding sites. In the group of antibiotics that inhibit DNA synthesis or cell division, ciprofloxacin and nalidixic acid resistance was widespread, except in L. helveticus 8/20, which showed intermediate susceptibility. These findings may be due to mutations in the quinolone resistance-determining regions (QRDRs) of DNA gyrase or topoisomerase IV, commonly implicated in fluoroquinolone resistance. Sensitivity to novobiocin and rifampin was more variable, with L. helveticus 6/20 being uniquely sensitive to rifampin, suggesting differential mechanisms of resistance, potentially at the level of RNA polymerase structure or function. The strain-dependent differences in antibiotic resistance profiles observed underscore the importance of individual strain screening, especially when considering LAB strains for probiotic or food industry applications. Moreover, the presence of resistance to clinically relevant antibiotics such as vancomycin and ciprofloxacin raises potential concerns regarding horizontal gene transfer, particularly in environments where LAB co-exist with pathogenic bacteria. Further genomic analysis is warranted to determine whether the resistance observed is due to intrinsic mechanisms or the acquisition of resistance genes. This is especially important given the safety implications of using L. helveticus strains in food or pharmaceutical products (Table 6.1).
When it comes to the Lactiplantibacillus plantarum strains, among the cell wall synthesis inhibitors, resistance to β-lactam antibiotics was consistently observed across all Lactiplantibacillus plantarum strains. All three strains showed resistance to penicillin (Table 6.2), ampicillin, oxacillin, and vancomycin — an expected finding considering the intrinsic resistance of many LAB to β-lactams and glycopeptides due to variations in their cell wall structure and low-affinity penicillin-binding proteins. However, strain-specific differences were noted in response to bacitracin, piperacillin, and amoxicillin. For instance, Lactiplantibacillus plantarum 5/20 was sensitive only to amoxicillin, whereas Lactiplantibacillus plantarum 13/20 and Lactiplantibacillus plantarum 19/20 demonstrated intermediate susceptibility to amoxicillin and bacitracin, respectively. These subtle differences could reflect either adaptive phenotypic variation or distinct underlying resistance mechanisms, such as efflux pump activity or cell envelope permeability. In terms of protein synthesis inhibitors, tobramycin and amikacin resistance was universal across all Lactiplantibacillus plantarum strains, aligning with known intrinsic resistance in LAB to aminoglycosides due to their inability to penetrate the thick peptidoglycan layer in the absence of oxygen. Lincomycin resistance was also observed in all three Lactiplantibacillus plantarum strains, suggesting possible intrinsic mechanisms or conserved ribosomal protection elements. However, the response to other antibiotics such as tetracycline, doxycycline, erythromycin, and gentamicin varied between Lactiplantibacillus plantarum strains. Lactiplantibacillus plantarum 13/20 was sensitive to tetracycline and erythromycin, while Lactiplantibacillus plantarum 5/20 and Lactiplantibacillus plantarum 19/20 were resistant. Notably, chloramphenicol sensitivity was retained in all strains, a favorable observation given its historical effectiveness against Gram-positive bacteria and its use as a marker in probiotic screening. The intermediate responses to gentamicin and doxycycline across multiple strains suggest either partial resistance mechanisms or borderline susceptibility thresholds that warrant further MIC-based analysis. For DNA synthesis and cell division inhibitors, resistance to nalidixic acid was observed in all Lactiplantibacillus plantarum strains, which is consistent with previously reported resistance in LAB due to mutations in DNA gyrase or topoisomerase IV. Ciprofloxacin resistance was also prevalent but exhibited intermediate susceptibility in strains Lactiplantibacillus plantarum 13/20 and Lactiplantibacillus plantarum 19/20, suggesting potential variability in fluoroquinolone target affinity or efflux mechanisms. Rifampin sensitivity in Lactiplantibacillus plantarum 13/20 and intermediate susceptibility in others points toward differing rpoB gene sequences or promoter activity. Resistance to novobiocin in Lactiplantibacillus plantarum 13/20 and intermediate responses in the other two strains align with the strain-dependent nature of resistance against this DNA gyrase inhibitor. The data reveal that while Lp. plantarum strains share common resistance traits—particularly against aminoglycosides, β-lactams, and quinolones — there is clear inter-strain heterogeneity that could impact their use in functional foods or clinical applications. Once again, the occurrence of resistance to clinically important antibiotics such as erythromycin and ciprofloxacin in some strains is a potential concern, especially in the context of horizontal gene transfer within the gut microbiota. These findings underscore the necessity of thorough safety assessments for each L. plantarum strain before inclusion in probiotic formulations, particularly when these strains may encounter antibiotic-selective environments in the host. Whole-genome sequencing, plasmid analysis, and conjugation assays would provide deeper insight into the genetic basis of resistance and the potential risk of horizontal gene transfer (Table 6.2).
Resistance to β-lactam antibiotics, including penicillin, ampicillin, oxacillin, and amoxicillin, was evident in L. acidophilus 10/20, which aligns with previously reported intrinsic resistance mechanisms in lactobacilli (Table 6.2). These mechanisms are thought to involve low-affinity penicillin-binding proteins (PBPs) and limited cell wall permeability to β-lactams. Interestingly, this strain showed sensitivity to piperacillin and intermediate sensitivity to bacitracin, suggesting partial or incomplete resistance mechanisms to certain β-lactams and polypeptide antibiotics.
This variability may reflect differences in the structural targets of these drugs or differential expression of resistance genes. In the category of protein synthesis inhibitors, L. acidophilus 10/20 demonstrated resistance to several commonly used antibiotics, including tetracycline, tobramycin, amikacin, lincomycin, and erythromycin. Resistance to aminoglycosides such as tobramycin and amikacin is well-documented in Lactobacillus species and is generally considered intrinsic due to their anaerobic or microaerophilic growth, which limits aminoglycoside uptake. Resistance to macrolides (erythromycin) and lincosamides (lincomycin), however, may be mediated by acquired genes such as erm(B) or lnu(A) and warrants further investigation to determine the risk of horizontal gene transfer. The strain retained sensitivity to chloramphenicol, a broad-spectrum antibiotic, and showed intermediate susceptibility to doxycycline and gentamicin, suggesting that some protein synthesis pathways remain vulnerable to inhibition. The intermediate susceptibility may indicate reduced expression of resistance mechanisms, low-level efflux activity, or partial permeability to these compounds. With respect to DNA synthesis and cell division inhibitors, L. acidophilus 10/20 was resistant to nalidixic acid and demonstrated intermediate sensitivity to ciprofloxacin, rifampin, and novobiocin. Resistance to quinolones such as nalidixic acid is common in lactobacilli and may arise from mutations in DNA gyrase or topoisomerase IV. The partial susceptibility to ciprofloxacin and rifampin may suggest incomplete resistance or adaptive tolerance, but further genomic analysis would be required to confirm whether these resistance traits are chromosomal or potentially mobile. The antibiotic profile of L. acidophilus 10/20 indicates a predominance of resistance to antibiotics across all three functional groups—cell wall synthesis, protein synthesis, and DNA replication inhibitors—with a few key exceptions. The retention of sensitivity to chloramphenicol and piperacillin, and intermediate sensitivity to several others, offers a promising basis for safe probiotic application, provided these traits are confirmed to be non-transferable (Table 6.2)
3.3. Batch Cultivation of Lactiplantibacillus plantarum 5/20 in a Laboratory Bioreactor with Continuous Stirring in MRS-Broth Medium. Determination of the Process Kinetics and Identification of Model Parameters
The possibility of obtaining a probiotic concentrate of active cells of Lactiplantibacillus plantarum 5/20 in MRS-broth medium was investigated. For this purpose, batch cultivation of the studied strain was carried out with correction and maintenance of pH=5.7 in a laboratory bioreactor with mechanical stirring. The dynamics of biomass accumulation during the process was monitored (Figure 12). The lag phase of the strain was extremely short and difficult to determine. Therefore, according to equation 2, the induction period (the time from the lag phase during which the cells begin to synthesize the necessary cellular structures and enzymes and to transition from a non-adapted to an adapted state to the composition of the medium and the cultivation conditions) and the adaptation rate constant k0 were determined. According to the model, for this strain the induction period (τ0) was 0.835 h, which was about 50 minutes. The adaptation rate constant (k0) for this strain was relatively high – 0.500 h-1. The short induction period and the high adaptation rate constant show that the strain quickly and easily adapted to the medium and cultivation conditions. From the 2nd to the 8th hour, intensive growth of the culture in the exponential phase was observed. From the 8th to the 12th hour, the strain entered the phase of slower growth, and at the 12th hour it entered the stationary phase, when a high concentration of viable cells of 7.1.1012 cfu/cm3 was achieved. From the 12th to the 24th hour the concentration of viable cells increased slightly and was about 2.4.1013 cfu/cm3.
Based on these experimental data, mathematical modeling of the growth kinetics was performed and the maximum growth rate µm and the coefficient of intrapopulation competition β, which are important bioprocess parameters of the culture, were determined. A comparison was made between the experimental data and those obtained from the models. The results of these studies are presented in Figure 12, and the values of the kinetic parameters are shown in Table 7.
From the data presented in Figure 12 it is clear that the models were very well in agreement with the experimental results. This was also confirmed by the high correlation coefficient and the low value of the identification error R2 (Table 7). In addition, the model gave a theoretical value of the maximum biomass concentration of 13.63 LogN, which was very close to the experimental one - 13.36 LogN. This allows the derived mathematical model and the identified kinetic parameters in it to be used for forecasting and prediction of the fermentation process in the studied time interval, as well as for its scaling up and control. The data presented in Table 8 indicate that Lactiplantibacillus plantarum 5/20 was characterized by a relatively high maximum specific growth rate - 0.130 h-1 and a low coefficient of intrapopulation competition - 0.0095 cm3/(CFU.h). The relatively high maximum specific growth rate and low coefficient of intrapopulation competition indicate that optimal conditions had been created for the growth of the studied strain.
Figure 13 presents the change in pH and oxidation-reduction potential. The data presented in Figure 9 show that the pH value rapidly decreased from 6.19 at the beginning of the process to 5.7 at the 4th hour, which once again testified to the rapid growth of the strain. After that, the pH value was maintained constant until the end of the fermentation process. The oxidation-reduction potential was relatively constant until the 4th hour and ranged between 114 – 118 mV, after which it began to decrease, and this trend was maintained until the end of the fermentation process, when its value was -89 mv.
3.4. Freeze-Drying
The freeze-drying method is appropriate when the moisture content of the raw materials is over 30%. The most common food products contain significant amounts of free water (from 70 to 90%) for different types of products. Like all preservation methods, freeze-drying (lyophilization) requires the products to be of high quality. Pre-treatment of production-valuable strains or mixed cultures is especially necessary. Pre-treatment aims to create conditions that guarantee maximum survival of bacterial cultures during freeze-drying. The vitality of bacterial cultures in the freeze-dried preparation is determined by the ability of bacterial cells to restore their initial biochemical properties upon rehydration. For lactic acid bacteria, these are fermentative, proteolytic, aroma-forming, antimicrobial and other activities. When microorganisms are stored in a dry state, they are in anabiosis. In this state, it is assumed that the metabolic processes are greatly slowed down, close to the cessation of all life processes - nutrition, respiration and reproduction. Since the cells are deprived of water and oxygen, metabolic processes are also terminated. The survival of different types of microorganisms subjected to freeze-drying is different. The LAB viability and in particular the microflora of Bulgarian yogurt, show that the survival of the studied and selected LAB strains during freeze-drying was also influenced by the growth stage at the time of drying [32]. The survival of bacterial cells also depends on the environment in which they are in the drying process. A number of substances contained in the nutrient media for cultivating microorganisms have a protective effect and preserve bacterial cells during drying. The concentration of bacterial cells also affects their survival during freeze-drying. At a concentration of cultures of lactic acid bacteria of 1010CFU/cm3 and higher, greater survival is observed [33]. Of essential importance for the survival of microorganisms is the method of supplying heat to freeze-dried cultures; with conductive heating the survival of lactic acid streptococci is high, and of L. bulgaricus, which is very sensitive to freeze-drying, it is low [34]. After completion of the fermentation process, the biomass was immobilized in a hydrocolloid matrix. After freezing to a temperature of -40°C, it was subjected to sublimation. The microbiological status of the obtained lyophilisate was determined (Table 8 and Figure 14).
The titer of active cells in the dry product after freeze-drying was determined and the concentration of active cells was found to be 2.3x1012 cfu/g, with a survival rate of 92.52% (Figure 14 and Table 8). The high survival rate indicates that the strain was xerotolerant and was little affected by the freeze-drying conditions. The resistance of the strain to damaging factors during freeze-drying makes it valuable for production purposes, since not all lactobacilli show such high survival and resistance to production processes.
4. Conclusions
11 strains were isolated from rose blossom of Rosa damascena Mill. They were identified to species level by applying cultural, microbiological, physiological-biochemical (API 50 CHL) and molecular-genetic (16S rRNA gene sequencing) identification methods. Among the isolates, there were 5 strains of the species Lactobacillus helveticus (4/20; 6/20; 8/20; 9/20 and 12/20), 3 strains of the species Lactiplantibacillus plantarum (5/20; 13/20 and 19/20), 1 strain of Lactobacillus acidophilus (10/20) and 2 strains of Leuconostoc mesenteroides (16/20 and 22/20). All newly isolated LAB strains exhibited high antimicrobial activity against the pathogenic bacteria Escherichia coli, Staphylococcus aureus, Salmonella abony, Proteus vulgaris, Listeria monocytogenes, Enterococcus faecalis. The strains Lactiplantibacillus plantarum 5/20, Lactobacillus helveticus 6/20 and Lactobacillus acidophilus 10/20 were resistant to most of the antibiotics used in clinical practice, which in turn opens up the possibility of their joint inclusion in the complex therapy in the composition of probiotics that would be administered together with the corresponding antibiotic. A batch fermentation process was carried out with Lactiplantibacillus plantarum 5/20 and the kinetic parameters of the fermentation process to obtain a concentrate with a high content of viable cells (1013CFU/cm3) were determined. A freeze-dried preparation with a high content of viable cells (over 1012CFU/g) was obtained. After additional studies of the probiotic properties of the newly isolated LAB strains, they can be included in the development of probiotic preparations and starter cultures for various types of functional foods and beverages.
Author Contributions
Conceptualization, Z. D. and R.D-K.; methodology, R.D-K. and B. G. .; software, R. D-K., G. K. and B. G..; validation, Z. D., R.D-K. and G. K.; formal analysis, P. Z., Y. G., B. G. and V. S; resources, Z. D.; data curation, R. D.-K and G. K.; writing—original draft preparation, R. D.-K., V. S., P. Z., B. G., Y. G.; writing—review and editing, R. D.-K., Z. D. and G. K; supervision, Z. D.; funding acquisition, G. K and P. Z.
Funding
The research in this article was funded by the project "Investigation of the synergistic action of plant extracts and lactic acid bacteria and ensuring sustainability in the production of fermented foods", National Program "Young scientists and postdoctoral students - 2", Ministry of Education and Science, Bulgaria
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Yu, A.O.; Leveau, J.H.; Marco, M.L. Abundance, diversity and plant-specific adaptations of plant-associated lactic acid bacteria. Environ. Microbiol. Rep. 2020, 12, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.A.; Ayivi, R.D.; Zimmerman, T.; Siddiqui, S.A.; Altemimi, A.B.; Fidan, H.; Esatbeyoglu, T.; Bakhshayesh, R.V. Lactic acid bacteria as antimicrobial agents: Food safety and microbial food spoilage prevention. Foods 2021, 10, 3131. [Google Scholar] [CrossRef]
- Lingga, R.; Astuti, R.; Bahtera, N.; Adibrata, S.; Nurjannah, N.; Ahsaniyah, S.; Irawati, I. Effect of additional probiotic in cattle feed on cattle's consumption and growth rate. Int. J. Sci. Technol. Manag. 2022, 3, 1418–1425. [Google Scholar] [CrossRef]
- Shivsharan, U.; Bhitre, M. Antimicrobial activity of lactic acid bacterial isolates. Int. Res. J. Pharm. 2013, 4, 197–201. [Google Scholar] [CrossRef]
- Nadirova, S.; Sinyavskiy, Y. Biotechnological approaches to the creation of new fermented dairy products. Eurasian J. Appl. Biotechnol. 2023, 4, 128–136. [Google Scholar] [CrossRef]
- Bayu, H.; Irwanto, R.; Dalimunthe, N.; Lingga, R. Isolation and identification of lactic acid bacteria from Channa sp. as potential probiotic. J. Pembelajaran Biol. Nukleus 2023, 9, 75–84. [Google Scholar] [CrossRef]
- Ar, I.; Laksmi, B.; Astawan, M.; Fujiyama, K.; Arief, I. Identification and probiotic characteristics of lactic acid bacteria isolated from Indonesian local beef. Asian J. Anim. Sci. 2014, 9, 25–36. [Google Scholar] [CrossRef]
- Pamungkaningtyas, F.; Mariyatun, M.; Kamil, R.; Setyawan, R.; Hasan, P.; Wiryohanjoyo, D.; Rahayu, E. Sensory evaluation of yogurt-like set and yogurt-like drink produced by indigenous probiotic strains for market test. Indones. Food Nutr. Prog. 2018, 15, 1. [Google Scholar] [CrossRef]
- Pundir, R.K.; Rana, S.; Kashyap, N.; Kaur, A. Probiotic potential of lactic acid bacteria isolated from food samples: an in vitro study. J. Appl. Pharm. Sci. 2013, 3, 85–93. [Google Scholar] [CrossRef]
- Wang, J.; Lu, C.; Xu, Q.; Li, Z.; Song, Y.; Zhou, S.; Luo, X. Bacterial diversity and lactic acid bacteria with high alcohol tolerance in the fermented grains of soy sauce aroma type baijiu in North China. Foods 2022, 11, 1794. [Google Scholar] [CrossRef]
- Aritonang, S.; Roza, E.; Rossi, E.; Purwati, E.; Husmaini, H. Isolation and identification of lactic acid bacteria from okara and evaluation of their potential as candidate probiotics. Pak. J. Nutr. 2017, 16, 618–628. [Google Scholar] [CrossRef]
- Hudzicki, J. Kirby-Bauer disk diffusion susceptibility test protocol. Am. Soc. Microbiol. 2009, 15, 1. Available online: https://asm.org/protocols/kirby-bauer-disk-diffusion-susceptibility-test-pro (accessed on 27 April 2025).
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Kostov, G.; Denkova, R.; Ivanova, P.; Denkova, I. Kinetics of batch fermentation in the cultivation of a probiotic strain of lactic acid bacteria. Acta Univ. Cibiniensis Ser. E Food Technol. 2015, 19, 61–72. Available online: https://saiapm.ulbsibiu.ro/cercetare/ACTA_E/AUCFT2015_19(1)_61-72.pdf (accessed on 27 April 2025).
- Kostov, G.; Denkova, R.; Ivanova, P.; Denkova, I. Microbial growth of Lactobacillus delbrueckii ssp. bulgaricus B1 in a complex nutrient medium (MRS-broth). ECMS Proc. 2022, pp. 135–142. Available online: https://www.scs-europe.net/dlib/2022/ecms2022acceptedpapers/0135_simo_ecms2022_0028.pdf (accessed on 27 April 2025).
- Kostov, G.; Denkova-Kostova, R.; Ivanova, P.; Denkova, I. New approach to modelling the kinetics of the fermentation process in cultivation of lactic acid bacteria. Proc. Eur. Conf. Model. Simul. 2018, 212–218. Available online: https://www.scs-europe.net/dlib/2018/2018-0212.htm (accessed on 27 April 2025).
- Denkova, R.; Denkova, Z.; Yanakieva, V.; Georgieva, L. Highly active freeze-dried probiotic concentrates with long shelf life. Food Environ. Saf. 2013, 12, 225. Available online: https://fens.usv.ro/index.php/FENS/article/download/169/167 (accessed on 27 April 2025).
- Bulgarian State Standard (BDS) EN ISO 4833:2004. Microbiology of food and animal feeding stuffs—Horizontal method for the enumeration of microorganisms—Colony-count technique at 30 °C. Bulgarian Institute for Standardization: Sofia, Bulgaria, 2004.
- Bulgarian State Standard (BDS) ISO 15214:2002. Microbiology of food and animal feeding stuffs—Horizontal method for the enumeration of mesophilic lactic acid bacteria—Colony-count technique at 30 degrees C. Bulgarian Institute for Standardization: Sofia, Bulgaria, 2002.
- Bulgarian State Standard (BDS) EN ISO 4833-1:2013/A1:2022. Microbiology of the food chain—Horizontal method for the enumeration of microorganisms—Part 1: Colony count at 30 degrees C by the pour plate technique; Amendment 1; Bulgarian Institute for Standardization: Sofia, Bulgaria, 2022. [Google Scholar]
- Bulgarian State Standard (BDS) ISO 16649-2:2014. Microbiology of the food chain—Horizontal method for the enumeration of β-glucuronidase-positive Escherichia coli—Part 2: Colony-count technique at 44 °C using 5-bromo-4-chloro-3-indolyl β-D-glucuronide. Bulgarian Institute for Standardization: Sofia, Bulgaria, 2014.
- Bulgarian State Standard (BDS) EN ISO 6579-1:2017/A1:2020. Microbiology of the food chain—Horizontal method for the detection, enumeration and serotyping of Salmonella—Part 1: Detection of Salmonella spp.; Amendment 1: Broader range of incubation temperatures and clarification of testing of samples with high background flora; Bulgarian Institute for Standardization: Sofia, Bulgaria, 2020. [Google Scholar]
- Bulgarian State Standard (BDS) EN ISO 6888-1:2021/A1:2023. Microbiology of the food chain—Horizontal method for the enumeration of Staphylococcus aureus—Part 1: Technique using Baird-Parker agar medium; Amendment 1.; Bulgarian Institute for Standardization: Sofia, Bulgaria, 2023. [Google Scholar]
- Bulgarian State Standard (BDS) ISO 15213:2023. Microbiology of the food chain—Horizontal method for the detection and enumeration of Clostridium spp.—Colony-count technique. Bulgarian Institute for Standardization: Sofia, Bulgaria, 2023.
- Bulgarian State Standard (BDS) ISO 6611:2006. Milk and milk products—Enumeration of colony-forming units of yeasts and/or moulds—Colony-count technique at 25 degrees C. Bulgarian Institute for Standardization: Sofia, Bulgaria, 2006.
- Majhenič, A.Č.; Lorbeg, P.M.; Treven, P. Enumeration and identification of mixed probiotic and lactic acid bacteria starter cultures. In Probiotic Dairy Products; CRC Press: Boca Raton, FL, USA, 2017; pp. 207–251. [Google Scholar]
- Singh, S.; Goswami, P.; Singh, R.; Heller, K.J. Application of molecular methods for identification of lactic acid bacteria. LWT—Food Sci. Technol. 2009, 42, 448–457. [Google Scholar] [CrossRef]
- Song, Y.L.; Kato, N.; Matsumiya, Y.; Liu, C.X.; Kato, H.; Watanabe, K. Identification of Lactobacillus species of human origin by a commercial kit, API50CHL. Rinsho Biseibutsu Jinsoku Shindan Kenkyukai Shi 1999, 10, 77–82. [Google Scholar]
- Charteris, W.P.; Kelly, P.M.; Morelli, L.; Collins, J.K. Development and application of an in vitro methodology to determine the transit tolerance of potentially probiotic Lactobacillus and Bifidobacterium species in the upper human gastrointestinal tract. J. Appl. Microbiol. 1998, 84, 759–768. [Google Scholar] [CrossRef]
- Mathur, S.; Singh, R. Antibiotic resistance in food lactic acid bacteria—A review. Int. J. Food Microbiol. 2005, 105, 281–295. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, F.; Cenard, S.; Passot, S. Freeze-drying of lactic acid bacteria. In Methods in Molecular Biology; Humana Press: New York, NY, USA, 2015; Volume 1257, pp. 477–488. [Google Scholar] [CrossRef]
- Tsvetkov, T.; Brankova, R. Viability of micrococci and lactobacilli upon freezing and freeze-drying in the presence of different cryoprotectants. Cryobiology 1983, 20, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Montel Mendoza, G.; Pasteris, S.E.; Otero, M.C.; Nader-Macías, M.E.F. Survival and beneficial properties of lactic acid bacteria from raniculture subjected to freeze-drying and storage. Benef. Microbes 2014, 5, 381–390. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 4/20 and the partial nucleotide sequence of the 16S rDNA of Lactobacillus helveticus NBRC 15019.
Figure 1.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 4/20 and the partial nucleotide sequence of the 16S rDNA of Lactobacillus helveticus NBRC 15019.

Figure 2.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 5/20 and the partial nucleotide sequence of the 16S rDNA of Lactiplantibacillus plantarum NBRC 15891.
Figure 2.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 5/20 and the partial nucleotide sequence of the 16S rDNA of Lactiplantibacillus plantarum NBRC 15891.

Figure 3.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 6/20 and the partial nucleotide sequence of the 16S rDNA of Lactobacillus helveticus NBRC 15019.
Figure 3.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 6/20 and the partial nucleotide sequence of the 16S rDNA of Lactobacillus helveticus NBRC 15019.

Figure 4.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 8/20 and the partial nucleotide sequence of the 16S rDNA of Lactobacillus helveticus NBRC 15019.
Figure 4.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 8/20 and the partial nucleotide sequence of the 16S rDNA of Lactobacillus helveticus NBRC 15019.
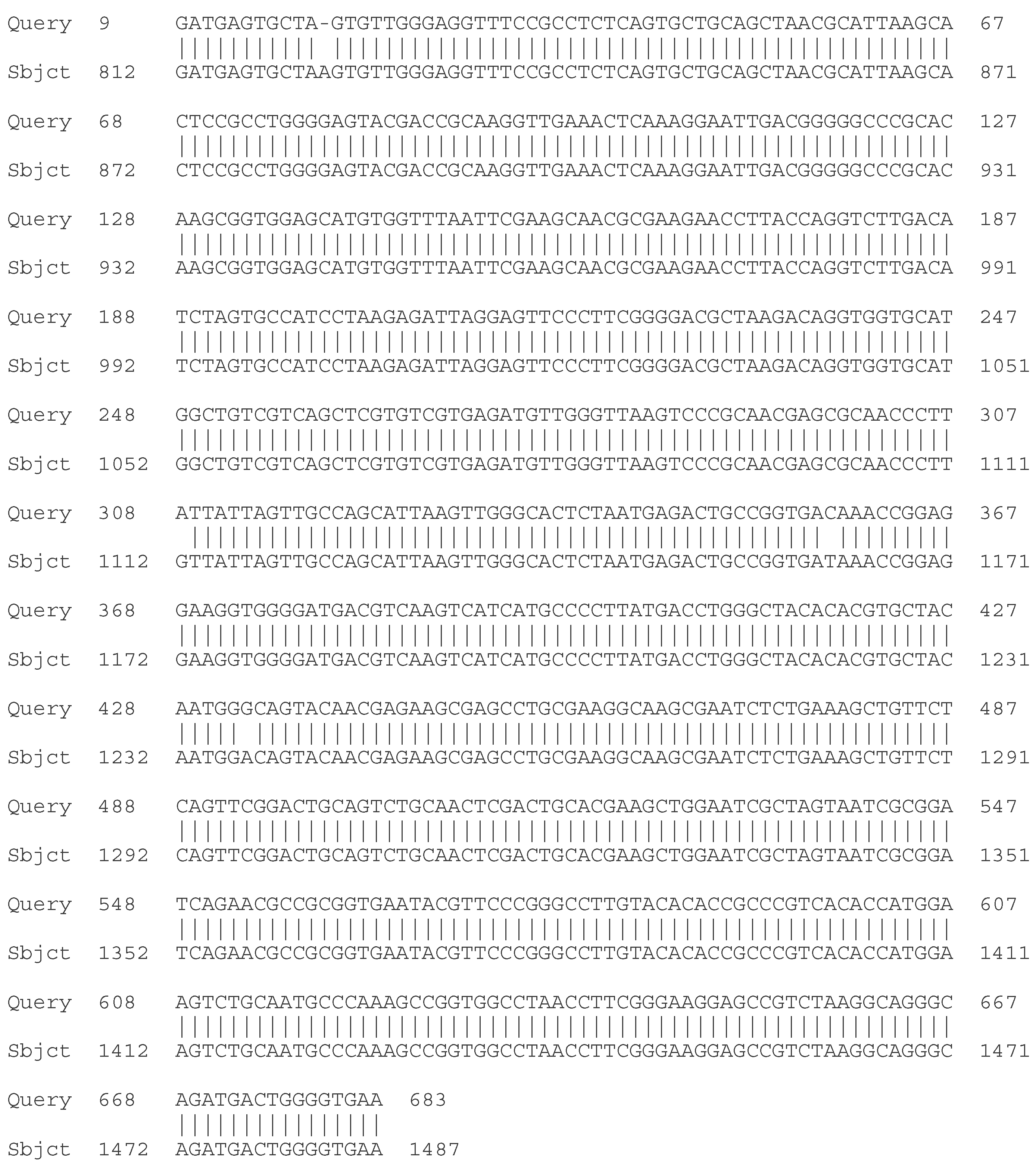
Figure 5.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 9/20 and the partial nucleotide sequence of the 16S rDNA of Lactobacillus helveticus NBRC 15019.
Figure 5.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 9/20 and the partial nucleotide sequence of the 16S rDNA of Lactobacillus helveticus NBRC 15019.

Figure 6.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 10/20 and the partial nucleotide sequence of the 16S rDNA of Lactobacillus acidophilus NBRC 13951.
Figure 6.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 10/20 and the partial nucleotide sequence of the 16S rDNA of Lactobacillus acidophilus NBRC 13951.
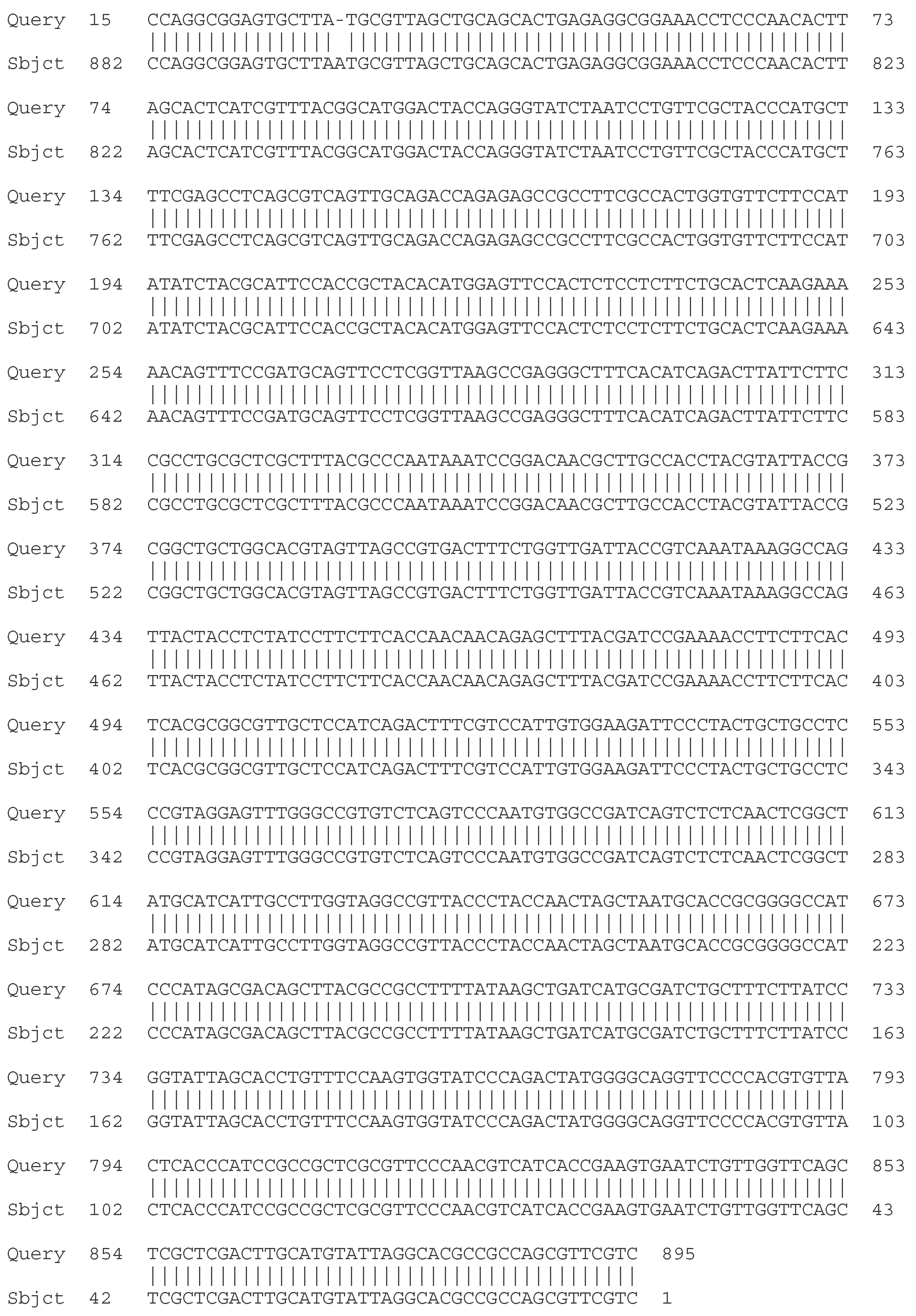
Figure 7.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 12/20 and the partial nucleotide sequence of the 16S rDNA of Lactobacillus helveticus NBRC 15019.
Figure 7.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 12/20 and the partial nucleotide sequence of the 16S rDNA of Lactobacillus helveticus NBRC 15019.

Figure 8.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 13/20 and the partial nucleotide sequence of the 16S rDNA of Lactiplantibacillus plantarum NBRC 15891.
Figure 8.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 13/20 and the partial nucleotide sequence of the 16S rDNA of Lactiplantibacillus plantarum NBRC 15891.

Figure 9.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 16/20 and the partial nucleotide sequence of the 16S rDNA of Leuconostoc mesenteroides ATCC 8293.
Figure 9.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 16/20 and the partial nucleotide sequence of the 16S rDNA of Leuconostoc mesenteroides ATCC 8293.

Figure 10.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 19/20 and the partial nucleotide sequence of the 16S rDNA of Lactiplantibacillus plantarum NBRC 15891.
Figure 10.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 19/20 and the partial nucleotide sequence of the 16S rDNA of Lactiplantibacillus plantarum NBRC 15891.
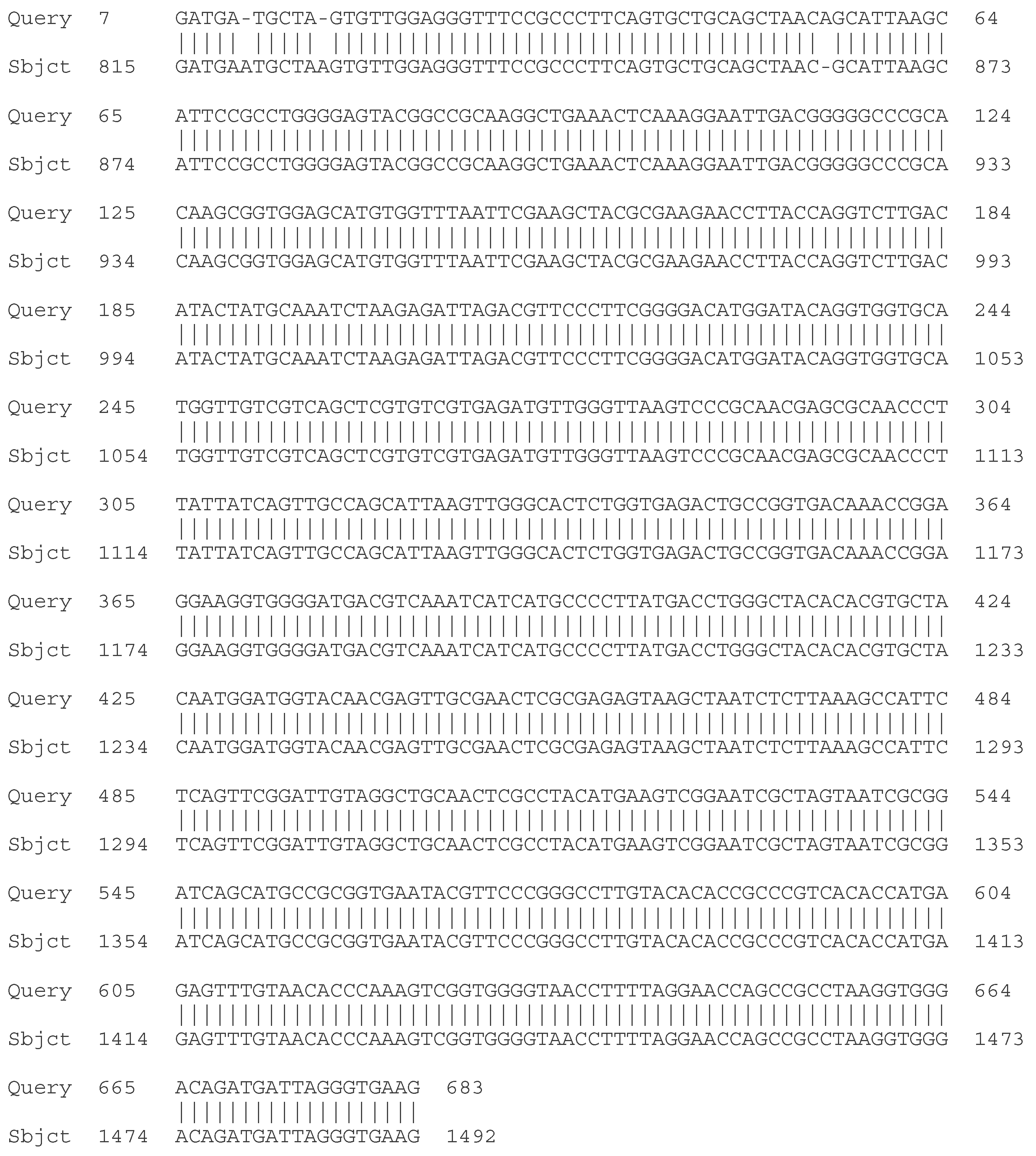
Figure 11.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 22/20 and the partial nucleotide sequence of the 16S rDNA of Leuconostoc mesenteroides ATCC 8293.
Figure 11.
Comparison between the partial nucleotide sequence of the 16S rDNA of LAB 22/20 and the partial nucleotide sequence of the 16S rDNA of Leuconostoc mesenteroides ATCC 8293.

Figure 12.
Comparison of experimental data with the logistic curve model for biomass accumulation during batch cultivation of Lactiplantibacillus plantarum 5/20 in a bioreactor with mechanical stirring at 37 °C.
Figure 12.
Comparison of experimental data with the logistic curve model for biomass accumulation during batch cultivation of Lactiplantibacillus plantarum 5/20 in a bioreactor with mechanical stirring at 37 °C.
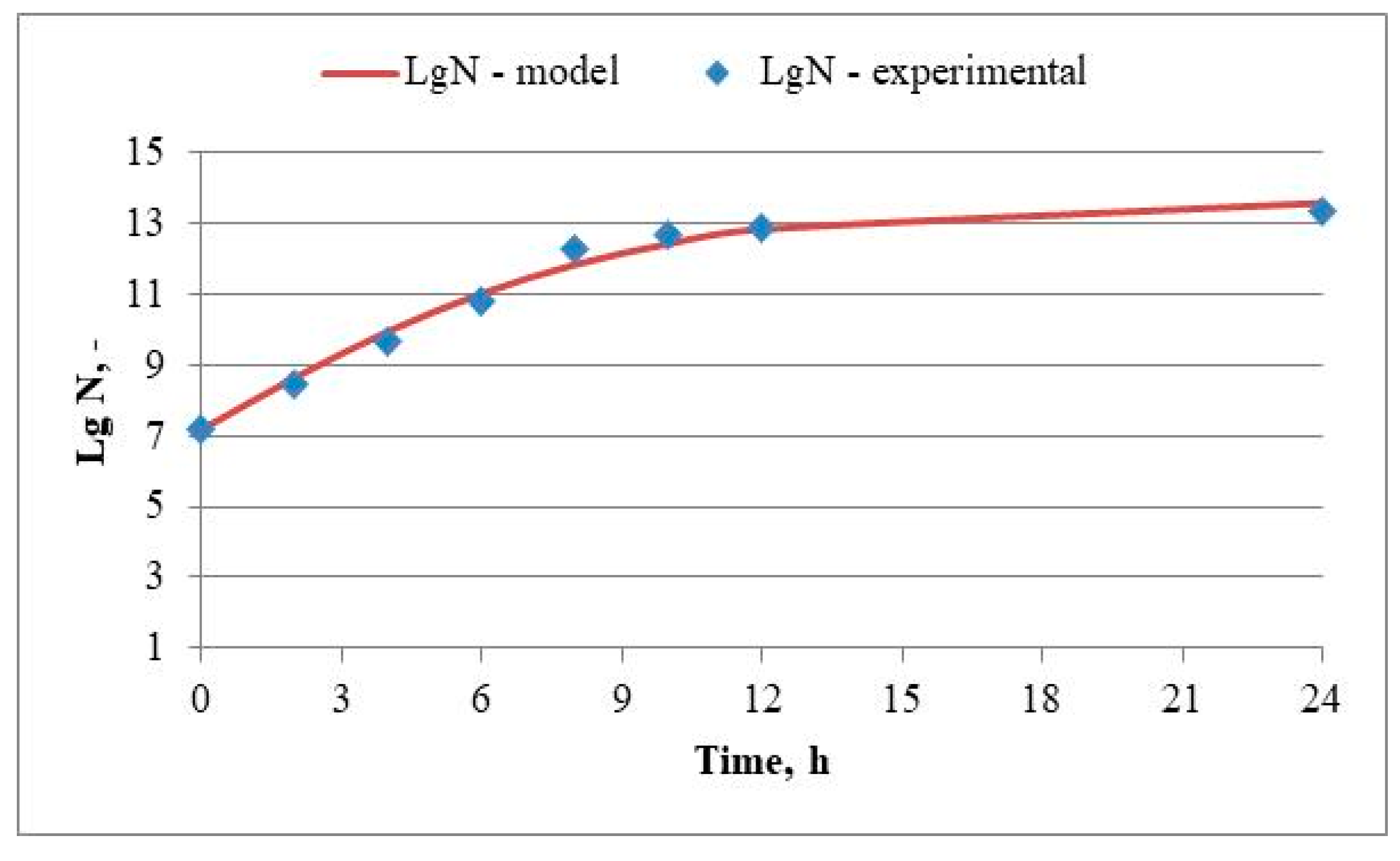
Figure 13.
Change in the oxidation-reduction potential and the pH in the batch cultivation of Lactiplantibacillus plantarum 5/20 in a bioreactor with mechanical stirring at 37 °C.
Figure 13.
Change in the oxidation-reduction potential and the pH in the batch cultivation of Lactiplantibacillus plantarum 5/20 in a bioreactor with mechanical stirring at 37 °C.
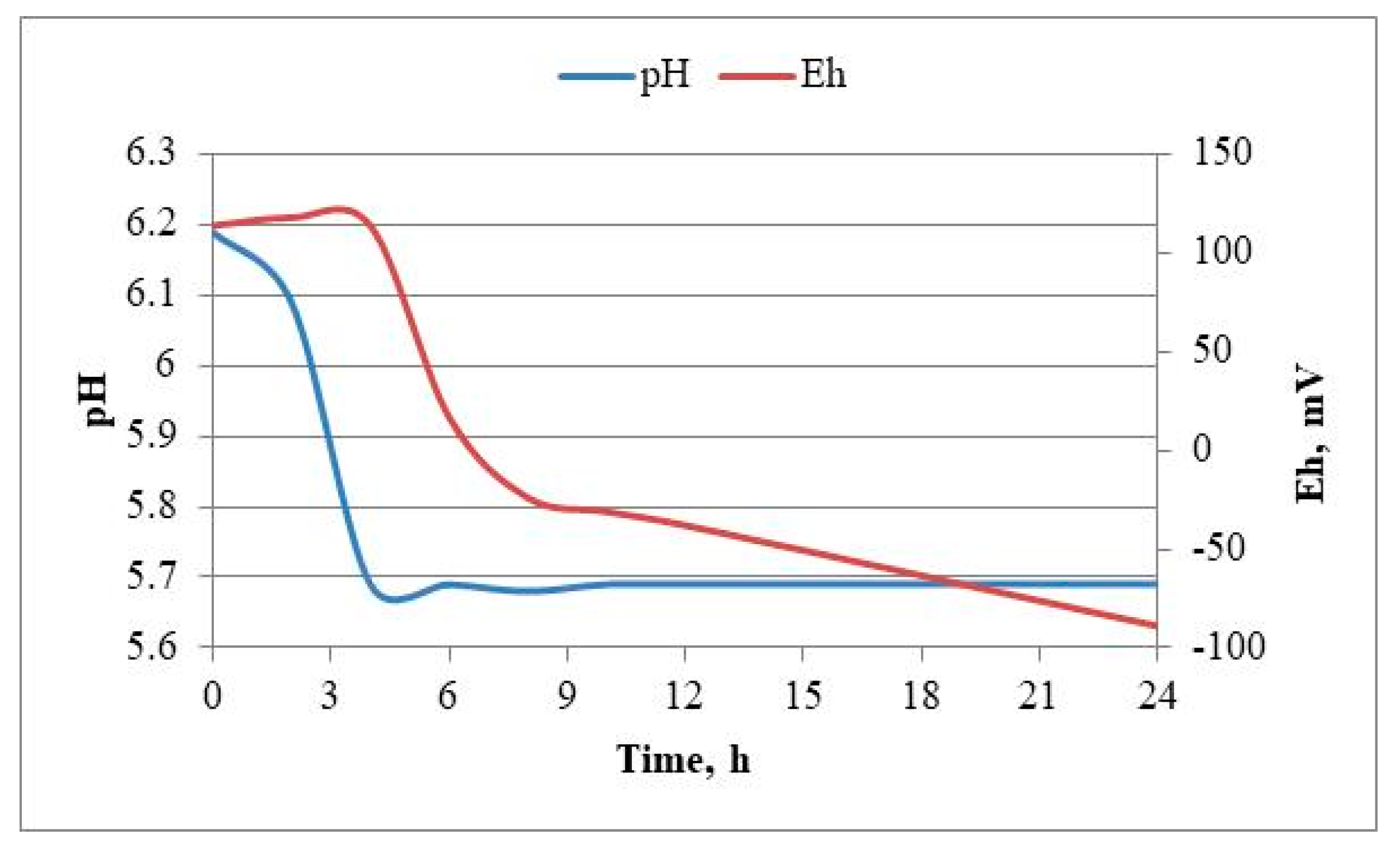
Figure 14.
Survival of Lactiplantibacillus plantarum 5/20 cells during the freeze-drying process.
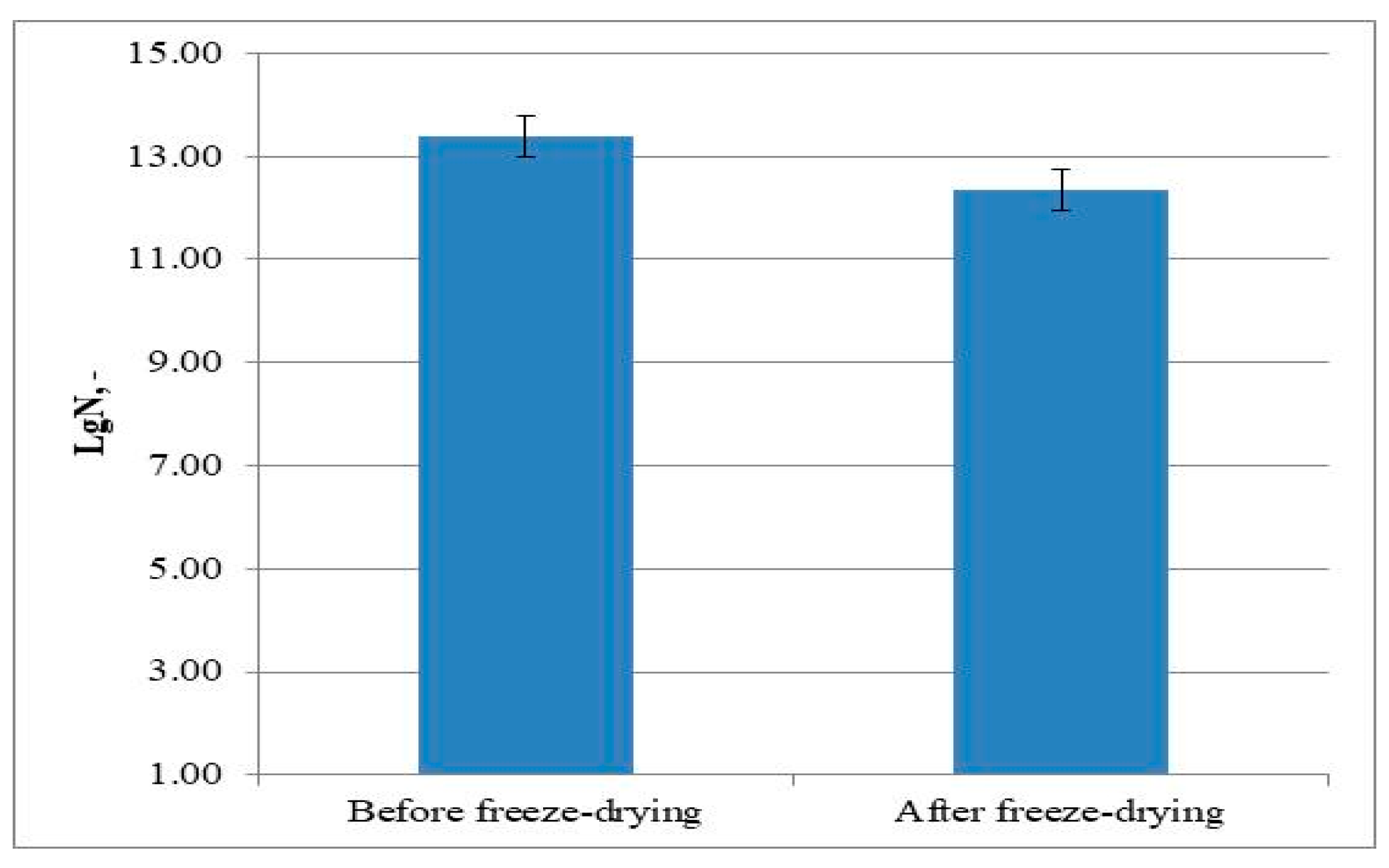

Table 1.
Colonial characteristics and cell morphology of the isolated strains.
| Strain | Cellular morphology | Colonial characteristics | ||
| Description | Visualization | Description | Visualization | |
| LAB 4/20 | Long, thickened rods with rounded ends, arranged singly, in pairs and in short chains |  |
snowflake-shaped colonies, serrated ends, 2-3 mm in diameter |  |
| LAB 5/20 | Fine, short rods with rounded ends, arranged in pairs and in short chains |  |
round colonies, with wavy ends, soft consistency, 2-3 mm in diameter |  |
| LAB 6/20 | Long, thin rods with rounded ends, arranged singly and in long chains |  |
round colonies with wavy ends, soft consistency, 2-3 mm in diameter |  |
| LAB 8/20 | Long, thin rods with rounded ends, arranged singly and in long chains |  |
round colonies with wavy ends, soft consistency, 2-3 mm in diameter |  |
| LAB 9/20 | Short, thickened rods with rounded ends, arranged singly and in clusters |  |
round colonies with wavy ends, soft consistency, 2-3 mm in diameter |  |
| LAB 10/20 | Long rods with rounded ends, arranged singly and in short chains |  |
snowflake-shaped colonies with wavy ends, soft consistency, 2-3 mm in diameter |  |
| LAB 12/20 | Long thin rods with rounded ends, arranged singly and in short chains |  |
snowflake-shaped colonies with serrated ends, 2-3 mm in diameter |  |
| LAB 13/20 | Short thickened rods with rounded ends, arranged singly, in pairs and in clusters |  |
round colonies with even ends, soft consistency, 2-3 mm in diameter |  |
| LAB 16/20 | Cocci, arranged in pairs and in groups |  |
round colonies with even ends, soft consistency, 2-3 mm in diameter |  |
| LAB 19/20 | Very short rods with rounded ends, arranged singly and in groups |  |
round colonies with even ends, soft consistency, 2-3 mm in diameter |  |
| LAB 22/20 | Short rods with rounded ends, arranged singly and in groups round, flat ends, soft consistency, 2-3 mm |  |
round colonies with even ends, soft consistency, 2-3 mm in diameter |  |
Table 2a.
Ability of strains LAB 4/20, LAB 5/20, LAB 6/20, LAB 8/20, LAB 9/20, LAB 10/20 to assimilate 49 carbon sources included in the API 50 CHL identification system.
Table 2a.
Ability of strains LAB 4/20, LAB 5/20, LAB 6/20, LAB 8/20, LAB 9/20, LAB 10/20 to assimilate 49 carbon sources included in the API 50 CHL identification system.
| # | Carbohydrates | LAB 4/20 | LAB 5/20 | LAB 6/20 | LAB 8/20 | LAB 9/20 | LAB 10/20 |
| 1 | Glycerol | - | - | - | - | - | - |
| 2 | Erythriol | - | - | - | - | - | - |
| 3 | D-arabinose | - | - | - | - | - | - |
| 4 | L-arabinose | - | + (90-100%) | - | - | - | + (90-100%) |
| 5 | Ribose | - | + (90-100%) | + (90-100%) | - | - | + (90-100%) |
| 6 | D-xylose | - | - | - | - | - | + (90-100%) |
| 7 | L-xylose | - | - | - | - | - | - |
| 8 | Adonitol | - | - | - | - | - | - |
| 9 | β -metil-D-xyloside | - | - | - | - | - | - |
| 10 | Galactose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 11 | D-glucose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 12 | D-fructose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 13 | D-mannose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 14 | L-sorbose | - | - | - | + (90-100%) | - | - |
| 15 | Rhamnose | - | - | - | + (90-100%) | - | - |
| 16 | Dulcitol | - | - | - | - | - | - |
| 17 | Inositol | - | - | - | - | - | - |
| 18 | Manitol | - | + (90-100%) | + (90-100%) | + (90-100%) | - | + (90-100%) |
| 19 | Sorbitol | - | + (90-100%) | + (90-100%) | + (90-100%) | - | + (90-100%) |
| 20 | α -methyl-D-mannoside | - | + (90-100%) | + (90-100%) | - | - | + (90-100%) |
| 21 | α -methyl-D-glucoside | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 22 | N-acetyl-glucosamine | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 23 | Amigdalin | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 24 | Arbutin | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 25 | Esculin | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 26 | Salicin | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 27 | Cellobiose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 28 | Maltose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 29 | Lactose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 30 | Melibiose | - | + (90-100%) | + (90-100%) | + (90-100%) | - | + (90-100%) |
| 31 | Saccharose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 32 | Trehalose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 33 | Inulin | - | - | + (90-100%) | - | - | - |
| 34 | Melezitose | - | + (90-100%) | + (90-100%) | + (90-100%) | - | + (90-100%) |
| 35 | D-raffinose | - | - | + (90-100%) | - | - | + (90-100%) |
| 36 | Amidon | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | - |
| 37 | Glycogen | - | - | - | - | - | - |
| 38 | Xylitol | - | - | - | - | - | - |
| 39 | β -gentiobiose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 40 | D-turanose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 41 | D-lyxose | - | - | - | - | - | - |
| 42 | D-tagarose | - | - | - | - | - | - |
| 43 | D-fuccose | - | - | - | - | - | - |
| 44 | L-fuccose | - | - | - | - | - | - |
| 45 | D-arabitol | - | - | - | - | - | - |
| 46 | L-arabitol | - | - | - | - | - | - |
| 47 | Gluconate | - | + (90-100%) | + (90-100%) | - | - | + (90-100%) |
| 48 | 2-keto-gluconate | - | - | - | - | - | - |
| 49 | 5-keto-gluconate | - | - | - | - | - | - |
Table 2b.
Ability of strains LAB 12/20, LAB 13/20, LAB 16/20, LAB 19/20, LAB 22/20 to assimilate 49 carbon sources included in the API 50 CHL identification system.
Table 2b.
Ability of strains LAB 12/20, LAB 13/20, LAB 16/20, LAB 19/20, LAB 22/20 to assimilate 49 carbon sources included in the API 50 CHL identification system.
| # | Sugars | LAB 12/20 | LAB 13/20 | LAB 16/20 | LAB 19/20 | LAB 22/20 |
| 1 | Glycerol | - | - | - | - | - |
| 2 | Erythriol | - | - | - | - | - |
| 3 | D-arabinose | - | - | - | - | - |
| 4 | L-arabinose | - | + (90-100%) | + (90-100%) | - | - |
| 5 | Ribose | - | + (90-100%) | + (90-100%) | - | - |
| 6 | D-xylose | - | - | + (90-100%) | - | - |
| 7 | L-xylose | - | - | - | - | - |
| 8 | Adonitol | - | - | - | - | - |
| 9 | β -metil-D-xyloside | - | - | - | - | - |
| 10 | Galactose | + (90-100%) | + (90-100%) | + (90-100%) | - | + (90-100%) |
| 11 | D-glucose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 12 | D-fructose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 13 | D-mannose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 14 | L-sorbose | - | - | - | - | - |
| 15 | Rhamnose | - | - | - | - | - |
| 16 | Dulcitol | - | - | - | - | - |
| 17 | Inositol | - | - | - | - | - |
| 18 | Manitol | - | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 19 | Sorbitol | - | + (90-100%) | - | + (90-100%) | + (90-100%) |
| 20 | α -methyl-D-mannoside | - | + (90-100%) | + (90-100%) | - | - |
| 21 | α -methyl-D-glucoside | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 22 | N-acetyl-glucosamine | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 23 | Amigdalin | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 24 | Arbutin | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 25 | Esculin | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 26 | Salicin | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 27 | Cellobiose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 28 | Maltose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 29 | Lactose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 30 | Melibiose | - | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 31 | Saccharose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 32 | Trehalose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 33 | Inulin | - | - | + (90-100%) | - | - |
| 34 | Melezitose | - | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 35 | D-raffinose | - | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 36 | Amidon | + (90-100%) | - | - | - | - |
| 37 | Glycogen | - | - | - | - | - |
| 38 | Xylitol | - | - | - | - | - |
| 39 | β -gentiobiose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | - |
| 40 | D-turanose | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) | + (90-100%) |
| 41 | D-lyxose | - | - | - | - | - |
| 42 | D-tagarose | - | - | - | - | - |
| 43 | D-fuccose | - | - | - | - | - |
| 44 | L-fuccose | - | - | - | - | - |
| 45 | D-arabitol | - | - | - | - | - |
| 46 | L-arabitol | - | - | - | - | - |
| 47 | Gluconate | - | + (90-100%) | - | + (90-100%) | - |
| 48 | 2-keto-gluconate | - | - | - | - | - |
| 49 | 5-keto-gluconate | - | - | - | - | - |
Table 3.
Species identification of the newly isolated LAB strains after processing the API 50 CHL results with apiweb®.
Table 3.
Species identification of the newly isolated LAB strains after processing the API 50 CHL results with apiweb®.
| Strain | Species affiliation | Reliability, % |
| LAB 4/20 | Lactobacillus crispatus | 65.3 |
| LAB 5/20 | Lactiplantibacillus plantarum | 99.9 |
| LAB 6/20 | Lactiplantibacillus plantarum | 99.9 |
| LAB 8/20 | Lactiplantibacillus plantarum | 99,9 |
| LAB 9/20 | Lactobacillus crispatus | 65,3 |
| LAB 10/20 | Lactiplantibacillus plantarum | 99.9 |
| LAB 12/20 | Lactobacillus crispatus | 65.3 |
| LAB 13/20 | Lactiplantibacillus plantarum | 99.9 |
| LAB 16/20 | Lactiplantibacillus plantarum | 94,1 |
| LAB 19/20 | Lactiplantibacillus plantarum | 99.9 |
| LAB 22/20 | Lactiplantibacillus plantarum | 99,9 |
Table 4.
Species identification of the newly isolated LAB strains, determined as a result of the 16S rRNA gene sequence analysis.
Table 4.
Species identification of the newly isolated LAB strains, determined as a result of the 16S rRNA gene sequence analysis.
| Strain | Species affiliation | Similarity, % |
| LAB 4/20 | Lactobacillus helveticus | 99% |
| LAB 5/20 | Lactiplantibacillus plantarum | 99% |
| LAB 6/20 | Lactobacillus helveticus | 99% |
| LAB 8/20 | Lactobacillus helveticus | 99% |
| LAB 9/20 | Lactobacillus helveticus | 99% |
| LAB 10/20 | Lactobacillus acidophilus | 99% |
| LAB 12/20 | Lactobacillus helveticus | 99% |
| LAB 13/20 | Lactiplantibacillus plantarum | 99% |
| LAB 16/20 | Leuconostoc mesenteroides | 99% |
| LAB 19/20 | Lactiplantibacillus plantarum | 99% |
| LAB 22/20 | Leuconostoc mesenteroides | 99% |
Table 5.1.
Antimicrobial activity of Lactobacillus helveticus 4/20, Lactobacillus helveticus 6/20, Lactobacillus helveticus 8/20, Lactobacillus helveticus 9/20, Lactobacillus helveticus 12/20 against pathogenic bacteria. dwell=7mm. The LAB concentration was 1011 – 1012 CFU/cm3. “-” – no inhibition of the growth of pathogenic microorganisms by the LAB isolates was found.
Table 5.1.
Antimicrobial activity of Lactobacillus helveticus 4/20, Lactobacillus helveticus 6/20, Lactobacillus helveticus 8/20, Lactobacillus helveticus 9/20, Lactobacillus helveticus 12/20 against pathogenic bacteria. dwell=7mm. The LAB concentration was 1011 – 1012 CFU/cm3. “-” – no inhibition of the growth of pathogenic microorganisms by the LAB isolates was found.
| dIZ, [mm] |
Escherichia coli ATCC 25922, 6x109cfu/cm3 |
Staphylococcus aureus ATCC 25093, 1,7x1012 cfu/cm3 |
Salmonella abony NTCC 6017, 2x1012 cfu/cm3 |
Proteus vulgaris J, 1,5x1012 cfu/cm3 |
Listeria monocytogenes ATCC 19115, 4x1012 cfu/cm3 |
Enterococcus faecalis ATCC 29212, 4,2x1012 cfu/cm3 |
|
| LAB strain | |||||||
| 4/20 | CL | 12,67±0,47 | 17,17±0,24 | 14,67±0,47 | 15,50±0,41 | 17,00±0,00 | 16,67±0,47 |
| BSS | 11,17±0,24 | 20,17±0,24 | 9,67±0,47 | 12,33±0,47 | 14,17±0,24 | 13,33±0,47 | |
| CFSN | 10,50±0,41 | 10,17±0,24 | 10,17±0,24 | 12,00±0,00 | 15,17±0,24 | 11,33±0,47 | |
| NCFSN | - | - | - | - | - | - | |
| 6/20 | CL | 12,17±0,24 | 15,00±0,00 | 11,33±0,47 | 14,33±0,47 | 18,33±0,47 | 16,50±0,41 |
| BSS | 9,17±0,24 | 16,17±0,24 | 8,00±0,00 | 9,17±0,24 | 14,00±0,00 | 13,17±0,24 | |
| CFSN | 10,17±0,24 | 13,67±0,47 | 9,50±0,41 | 12,00±0,00 | 15,67±0,47 | 13,00±0,00 | |
| NCFSN | - | - | - | - | - | - | |
| 8/20 | CL | 11,17±0,24 | 13,50±0,41 | 10,17±0,24 | 13,17±0,24 | 16,50±0,41 | 16,00±0,00 |
| BSS | 8,00±0,00 | 12,17±0,24 | 8,33±0,47 | 8,17±0,24 | 13,17±0,24 | 12,17±0,24 | |
| CFSN | 10,50±0,41 | 10,50±0,41 | 8,00±0,00 | 10,00±0,00 | 14,67±0,47 | 13,17±0,24 | |
| NCFSN | - | - | - | - | - | - | |
| 9/20 | CL | 11,50±0,41 | 16,00±0,00 | 11,67±0,47 | 12,50±0,41 | 18,67±0,47 | 18,67±0,47 |
| BSS | 8,00±0,00 | 10,17±0,24 | 8,17±0,24 | 9,00±0,00 | 12,17±0,24 | 15,00±0,00 | |
| CFSN | 10,00±0,00 | 10,67±0,47 | 9,50±0,41 | 10,00±0,00 | 15,17±0,24 | 14,00±0,00 | |
| NCFSN | - | - | - | - | - | - | |
| 12/20 | CL | 14,67±0,47 | 16,17±0,24 | 14,00±0,00 | 15,00±0,00 | 18,00±0,82 | 18,17±0,24 |
| BSS | 9,17±0,24 | 12,33±0,47 | 10,00±0,00 | 10,00±0,00 | 15,17±0,24 | 13,17±0,24 | |
| CFSN | 12,00±0,00 | 13,17±0,24 | 10,17±0,24 | 13,50±0,41 | 15,67±0,47 | 13,00±0,00 | |
| NCFSN | - | - | - | - | - | 8,00±0,00 |
Table 5.2.
Antimicrobial activity of Lactiplantibacillus plantarum 5/20, Lactiplantibacillus plantarum 13/20, Lactiplantibacillus plantarum 19/20, Lactobacillus acidophilus 10/20 against pathogenic bacteria. dwell=7mm. The LAB concentration was 1011 – 1012 CFU/cm3. “-” – no inhibition of the growth of pathogenic microorganisms by the LAB isolates was found.
Table 5.2.
Antimicrobial activity of Lactiplantibacillus plantarum 5/20, Lactiplantibacillus plantarum 13/20, Lactiplantibacillus plantarum 19/20, Lactobacillus acidophilus 10/20 against pathogenic bacteria. dwell=7mm. The LAB concentration was 1011 – 1012 CFU/cm3. “-” – no inhibition of the growth of pathogenic microorganisms by the LAB isolates was found.
| dIZ, [mm] |
Escherichia coli ATCC 25922, 6x109cfu/cm3 |
Staphylococcus aureus ATCC 25093, 1,7x1012 cfu/cm3 |
Salmonella abony NTCC 6017, 2x1012 cfu/cm3 |
Proteus vulgaris J, 1,5x1012 cfu/cm3 |
Listeria monocytogenes ATCC 19115, 4x1012 cfu/cm3 |
Enterococcus faecalis ATCC 29212, 4,2x1012 cfu/cm3 |
|
| LAB strain | |||||||
| 5/20 | CL | 13,67±0,47 | 19,67±0,47 | 13,33±0,47 | 17,00±0,00 | 20,00±0,00 | 18,50±0,41 |
| BSS | 9,17±0,24 | 17,33±0,47 | 9,17±0,24 | 10,00±0,00 | 16,17±0,24 | 14,67±0,47 | |
| CFSN | 11,17±0,24 | 14,67±0,47 | 11,33±0,47 | 13,17±0,24 | 17,67±0,47 | 13,50±0,41 | |
| NCFSN | 8,17±0,24 | - | - | - | 9,17±0,24 | - | |
| 13/20 | CL | 11,17±0,24 | 19,50±0,41 | 10,17±0,24 | 14,50±0,41 | 17,17±0,24 | 17,67±0,47 |
| BSS | 9,00±0,00 | 17,17±0,24 | 8,00±0,00 | 10,17±0,24 | 15,00±0,00 | 14,17±0,24 | |
| CFSN | 10,50±0,41 | 13,00±0,00 | 9,00±0,00 | 13,17±0,24 | 15,17±0,24 | 13,00±0,00 | |
| NCFSN | - | - | - | - | 8,00±0,00 | - | |
| 19/20 | CL | 10,17±0,24 | 10,17±0,24 | 9,67±0,47 | 10,67±0,47 | 11,00±0,00 | 10,17±0,24 |
| BSS | 8,00±0,00 | 9,17±0,24 | 8,00±0,00 | 8,17±0,24 | 8,00±0,00 | 8,67±0,47 | |
| CFSN | 8,67±0,47 | 8,00±0,00 | 8,50±0,41 | 9,00±0,00 | 9,00±0,00 | 9,17±0,24 | |
| NCFSN | - | - | - | - | - | - | |
| 10/20 | CL | 10,17±0,24 | 8,00±0,00 | 9,00±0,00 | 10,17±0,24 | 9,67±0,47 | 8,17±0,24 |
| BSS | 8,17±0,24 | 8,17±0,24 | 8,17±0,24 | 8,17±0,24 | 8,17±0,24 | 8,17±0,24 | |
| CFSN | 9,17±0,24 | 8,00±0,00 | 9,00±0,00 | 9,17±0,24 | 9,17±0,24 | 8,00±0,00 | |
| NCFSN | - | - | - | - | - | - |
Table 6.1.
Antibiotic resistance of Lactobacillus helveticus 4/20, Lactobacillus helveticus 6/20, Lactobacillus helveticus 8/20, Lactobacillus helveticus 9/20, Lactobacillus helveticus 12/20.
Table 6.1.
Antibiotic resistance of Lactobacillus helveticus 4/20, Lactobacillus helveticus 6/20, Lactobacillus helveticus 8/20, Lactobacillus helveticus 9/20, Lactobacillus helveticus 12/20.
| # | Mechanism of action | Antibiotic | Concentration | 4/20 | 6/20 | 8/20 | 9/20 | 12/20 | |
| 1 | Inhibitor of cell wall synthesis | Penicillin | P | 10 E/disc | R | S | S | R | R |
| 2 | Bacitracin | Cm | 0,07 E/disc | R | S | S | SR | SR | |
| 3 | Piperacillin | P | 100 µg/disc | S | R | S | S | SR | |
| 4 | Ampicillin | A | 10 µg/disc | R | R | S | R | R | |
| 5 | Oxacillin | O | 1 µg/disc | R | S | R | R | R | |
| 6 | Amoxicillin | Ax | 25 µg/disc | S | S | S | R | SR | |
| 7 | Vancomycin | V | 30 µg/disc | R | S | S | R | R | |
| 8 | Inhibitor of protein synthesis | Tetracycline | T | 30 µg/disc | R | R | S | R | S |
| 9 | Doxycycline | D | 30 µg/disc | R | SR | SR | SR | S | |
| 10 | Gentamicin | G | 10 µg/disc | SR | R | S | SR | SR | |
| 11 | Tobramycin | Tb | 10 µg/disc | R | S | S | R | R | |
| 12 | Amikacin | Am | 30 µg/disc | R | R | SR | R | SR | |
| 13 | Lincomycin | L | 15 µg/disc | R | R | SR | R | SR | |
| 14 | Chloramphenicol | C | 30 µg/disc | S | SR | S | S | S | |
| 15 | Erythromycin | E | 15 µg/disc | SR | S | S | R | S | |
| 16 | Inhibitor of the DNA synthesis and/or of cell division | Novobiocin | Nb | 5 µg/disc | S | R | S | SR | R |
| 17 | Nalidixic acid | Nx | 30 µg/disc | R | R | SR | R | R | |
| 18 | Rifampin | R | 5 µg/disc | SR | S | S | SR | S | |
| 19 | Ciprofloxacin | Cp | 5 µg/disc | R | R | SR | SR | SR | |
At dIZ < 8 mm – R, at 8 < dIZ < 16 mm – SR, at dIZ > 16 mm – S.
Table 6.2.
Antibiotic resistance of Lactiplantibacillus plantarum 5/20, Lactiplantibacillus plantarum 13/20, Lactiplantibacillus plantarum 19/20, Lactobacillus acidophilus 10/20.
Table 6.2.
Antibiotic resistance of Lactiplantibacillus plantarum 5/20, Lactiplantibacillus plantarum 13/20, Lactiplantibacillus plantarum 19/20, Lactobacillus acidophilus 10/20.
| # | Mechanism of action | Antibiotic | Concentration | 5/20 | 13/20 | 19/20 | 10/20 | |
| 1 | Inhibitor of cell wall synthesis | Penicillin | P | 10 E/disc | R | R | R | R |
| 2 | Bacitracin | Cm | 0,07 E/disc | R | SR | SR | SR | |
| 3 | Piperacillin | P | 100 µg/disc | R | SR | S | S | |
| 4 | Ampicillin | A | 10 µg/disc | R | R | R | R | |
| 5 | Oxacillin | O | 1 µg/disc | R | R | R | R | |
| 6 | Amoxicillin | Ax | 25 µg/disc | S | SR | R | R | |
| 7 | Vancomycin | V | 30 µg/disc | R | R | R | R | |
| 8 | Inhibitor of protein synthesis | Tetracycline | T | 30 µg/disc | SR | S | R | R |
| 9 | Doxycycline | D | 30 µg/disc | S | S | SR | SR | |
| 10 | Gentamicin | G | 10 µg/disc | SR | SR | SR | SR | |
| 11 | Tobramycin | Tb | 10 µg/disc | R | R | R | R | |
| 12 | Amikacin | Am | 30 µg/disc | R | SR | R | R | |
| 13 | Lincomycin | L | 15 µg/disc | R | SR | R | R | |
| 14 | Chloramphenicol | C | 30 µg/disc | S | S | S | S | |
| 15 | Erythromycin | E | 15 µg/disc | R | S | R | R | |
| 16 | Inhibitor of the DNA synthesis and/or of cell division | Novobiocin | Nb | 5 µg/disc | SR | R | SR | SR |
| 17 | Nalidixic acid | Nx | 30 µg/disc | R | R | R | R | |
| 18 | Rifampin | R | 5 µg/disc | SR | S | SR | SR | |
| 19 | Ciprofloxacin | Cp | 5 µg/disc | R | SR | SR | SR | |
At dIZ < 8 mm – R, at 8 < dIZ < 16 mm – SR, at dIZ > 16 mm – S.
Table 7.
Kinetic parameters of the process of batch cultivation of Lactiplantibacillus plantarum 5/20 in a bioreactor with mechanical stirring at 37 °C.
Table 7.
Kinetic parameters of the process of batch cultivation of Lactiplantibacillus plantarum 5/20 in a bioreactor with mechanical stirring at 37 °C.
| Batch cultivation of Lactiplantibacillus plantarum 5/20 | ||||
| µ, h-1 | β, cm3/(CFU.h) | Xкр, LogN | R2 | Error |
| 0,130 | 0,0095 | 13,63 | 0,9995 | 0,778 |
Table 8.
Microbiological status of the lyophilized Lactiplantibacillus plantarum 5/20.
| Type of pathogenic microorganisms tested | According to BDS | Analysis result |
| Total number of mesophilic aerobic and facultative anaerobic bacteria | No more than 800 | 180 |
| Escherichia coli in 0.l g of the product | Not to be detected any | Not detected |
| Sulphite-reducing clostridia in 0.l g of the product | Not to be detected any | Not detected |
| Salmonella sp. in 25.0 g of the product | Not to be detected any | Not detected |
| Coagulase-positive staphylococci in 1.0 g of the product | Not to be detected any | Not detected |
| Spores of microscopic molds, CFU/g | No more than 100 | Not detected |
| Yeasts, CFU/g | No more than 100 | Not detected |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



