Submitted:
28 April 2025
Posted:
29 April 2025
You are already at the latest version
Abstract
The objective of this study is to isolate, identify, and describe rumen yeast strains, and assess their probiotic potentials and effects on ruminal fermentation in vitro. Yeasts were isolated from ruminal fluids, yielding 59 strains from nine distinct species. A number of tests were conducted to assess their anaerobic traits, growth rate, acid tolerance, lactate utilization ability and a second screening in fresh ruminal fluid to evaluate in vitro pH and acid accumulation was conducted . Finally, Candida rugosa (NJ-5) with good probiotic characteristic was chosen to investigate its effect on ruminal fermentation in vitro.Batch culture technique was used to explore the effect of Candida rugosa (NJ-5) yeast culture on rumen fermentation parameters. By altering the fermentation substrate to a concentrate-to-roughage ratio of 70:30, which simulated a high-concentration diet. Group CON, LYC, MYC and HYC were supplemented with 0%, 1%, 2%, and 5% Candida rugosa (NJ-5) yeast culture (dry matter basis) respectively. The pH value and volatile fatty acid (VFAs) contents was determined at 6, 12 and 24 h after fermentation. The results showed that adding Candida rugosa (NJ-5) yeast culture successfully modulated the in vitro rumen fermentation. Compared to the CON group, HYC groups can mitigate the reduction of pH in fermentation ,resulting in a significant increase in total VFAs and acetate levels (p < 0.05). 16S rRNA sequencing revealed that Candida rugosa (NJ-5) yeast culture supplementation did not significantly alter ruminal bacterial alpha diversity (p > 0.05).At the phylum and genus taxonomic levels Candida rugosa (NJ-5) yeast cultureaddition increased the relative abundance of several functionally important bacterial groups in the rumen microbial community.Compared to the CON group, HYC groups concurrently increased the abundance of Desulfobacterota, Christensenellaceae_R-7_group, F082, and Ruminococcus (p < 0.05), but significantly reduced the abundance of Cyanobacteria, Bdellovibrionota, Succinivibrionaceae_UCG-002, Enterobacter, and Succinivibrio (p < 0.05). The in vitro fermentation experiment domenstrated that the optimal dry matter supplementation of Candida rugosa (NJ-5) into basal diet was 5%, which could be effective to maintain ruminal fermentation stability when ruminants were fed a high-concentrate diet. This study provides empirical support for the use of yeast as a nutritional supplement in ruminant livestock management, as well as a theoretical underpinning for further animal research.
Keywords:
Candida rugosa
; ruminal fermentation
; probiotic yeast
; microbial community composition
1. Introduction
Recent advancements in ruminant production have prominently incorporated yeast products as microbial feed supplements[1]. Numerous studies suggest that adding yeast culture substances into ruminant diets may increase feed intake, improve rumen fermentation, stabilize ruminal pH, and strengthen the animals immune system, and hence promoting production performance[2,3,4]. Moreover, yeast can compete with infections through competitive inhibition pathogen growth in the digestive tract[5,6]. Currently, the majority of commercial yeast products utilized in cattle production are derived from Saccharomyces cerevisiae strains[7]. Nevertheless, these strains were notexplicitly selected for their probiotic characteristics in ruminants. It’s reported that Saccharomyces cerevisiae strains show limited growth in the rumen, and cannot reproduce stably in this environment [8]. Therefore, when selecting potential probiotics, it is crucial to isolate strains from the same ecological niche in which they will be carefully selected suitable for the specific host to ensure reliable probiotic functions[9,10]. Key criteria for selecting a yeast strain includes its ability to thrive and demonstrate vigorous growth in rumen environment with the capacity to tolerate ruminal pH, enhance rumen fermentation, and ultimately improve rumen functionality while maintaining the stability of the microbial community. The in vitro ruminal fermentation method is commonly utilized to assess the nutritional effectiveness of feed additives on rumen fermentation before in vivo studies[11]. Previous studies suggest that Pichia kudriavzevii, Candida rugosa, and Kodamaea ohmeri, derived from ruminal fluid of dairy cow could serve as effective probiotic supplements by regulating lactic acid metabolism in dairy cattle nutrition[12]. Additionally, Candida tropicalis and Candida norvegensis, isolated from rumen significantly improved ruminal fermentation[13]. It’s reported that Candida norvegensis derived from the rumen, significantly improved the in vitro fermentation properties of oat straw, promoting microbial growth and the rate of dry matter decomposition, but greatly reduced methane emissions[14]. Similarly, Pichia kudriavzevii, isolated from ruminal fluid, can influence specific microbial communities to enhance fiber and fat digestion and also promote acetate-type fermentation processes[15]. In this study, the method of in vitro fermentation was employed to evaluate the effect of yeast strains isolated and selected from goat rumen on ruminal fermentation, and 16S rRNA sequencing was used to investigate the influence of the most promising probiotic strains on the bacteria community. The aim of this study was to explore the potential genetic resources from rumen and provide a basic data for these probiotics application for improving animals welfare and productivity in ruminants and non-ruminants.
2. Materials and Methods
The animal experiments were approved by the Institutional Animal Care and Use Committee of Nanjing Agricultural University according to the Guidelines on Ethical Treatment of Experimental Animals (2006) No. 398 set by the Ministry of Science and Technology (2006, Beijing, China) .
2.1. Collection of Rumen Samples for Yeast Isolation and Identification
Fresh ruminal fulid samples were collected to isolate yeasts from the ventral sac of the rumen via the rumen fistula before the morning feeding. The rumen fluid was filtered through four layers of gauze, stored in a CO₂-filled vacuum flask to maintain an anaerobic environment, and swiftly brought to the laboratory for yeast isolation. The filtered rumen fluid was 10 times serially diluted from 10-1 to 10-5 with 0.85% sterile saline. Following dilution, the fluid was inoculated into YPD agar plates and incubated at 39 °C for 48 hours. The YPD agar (1000 mL) consisted of yeast extract 5 g ,peptone 10 g, glucose 20 g and agar 14 g. Morphological observations were performed, and colonies displaying diverse shapes and colors were selected using an inoculation loop. Streak plate method for three rounds of purification was used until the recovered yeast strains exhibited consistent morphological characteristics.
After 48 h of incubation, total 59 yeast isolates were cultured in vitro, and their cellular biomass was collected. Genomic DNA from yeast was extracted using a Yeast Genome DNA Extraction Kit (Beijing Solarbio Science & Technology Co., Ltd., Beijing, China) . The PCR was performed with primers ITS1(5’-TCCGTAGGTGAACCTGCGG-3) and ITS4(5’-TCCTCCGCTTATTGATATGC-3’) [16]. PCR reaction system: 50 μL containing 25 μL of 2 × Taq Master Mix (Nanjing Novozan Biotechnology Co., Ltd., Nanjing, China) , 1 μL template DNA, 1 μL each of forward and reverse primers (10 μM) , and ddH₂O to a final volume of 50 μL. The PCR amplification program : Initial denaturation at 95 °C for 5 min; 30 cycles of 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 30 s; final extension at 72 °C for 10 min. The PCR amplified products were subjected to electrophoresis on a 1% agarose gel. The amplicons were sent to Nanjing Tsingke Biotech Co., Ltd. for DNA sequencing. The sequences were aligned using BLAST on the NCBI GenBank platform. The sequences were acquired and analyzed to construct a phylogenetic tree using MEGA software (version 7.0) .
2.2. Selection of Yeasts with Probiotic Potential
The viability in anaerobic circumstances at 39°C, lactic acid assimilation capacity, and acid tolerance ability was used to assesse the isolated 59 yeast strains for probiotic suitability in ruminants. After each test, only the yeasts that showed the best results were put through the next test.
Initially, to evaluate the proliferation of the isolated 59 yeast strains under anaerobic circumstances. Following three generations of activation, the yeast strains were injected at a 1% (v/v) concentration into YPD liquid medium and cultivated at 39°C, 150 rpm for 48 h. The yeast suspension was subsequently inoculated onto YPD agar plates with an inoculation loop, after which the plates were sealed in anaerobic bags and incubated at 39°C for 48 h. The expansion of colonies was documented and recorded. To identify yeast strains exhibiting accelerated growth rates, we chose those possessing anaerobic capabilities and inoculated them into YPD liquid medium at a 1% (v/v) concentration. Following incubation at 39°C and 150 rpm for 48 h, we collected the fermentation liquid and assessed the absorbance at 600 nm with a microplate reader. Yeast strains with an OD600 value exceeding 1.0 were chosen for additional investigations. Subsequently, yeast strains wih high acid tolerance were selected and then inoculated onto YPD liquid media under pH 5.0, 5.4, and 5.8, respectively utilizing a 1% (v/v) inoculum. Following incubation at 39°C with shaking at 150 rpm for 48 h, The supernatant was sampled, and was quantified at absorbance at 600 nm using a microplate reader. Yeast strains with an OD600 value exceeding 0.8 were selected and preserved for subsequent investigations. Finally, lactic acid was included to modify the YPD liquid medium to a concentration of 15 mmol/L. The chosen yeast strains were inoculated into the medium at a 1% (v/v) concentration and cultivated at 39°C with agitation at 150 rpm for 48 h. The supernatant was sampled, and the level of lactic acid was quantified using a commercial assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, Jiangsu) . The lactic acid consumption rate was determined using the formula: (initial lactic acid concentration in the medium – lactic acid concentration in the fermentation liquid) / initial lactic acid concentration in the medium.
2.3. The Preparation of Candida Rugosa (NJ-5) Yeast Culture
Candida rugosa (NJ-5) inoculated into YPD liquid medium, then incubated at 39°C with shaking at 150 rpm for 12 h to produce Candida rugosa (NJ-5) seed culture for subsequent application. A 5% inoculum of the yeast seed culture was subsequently introduced to the YPD liquid medium for aerobic fermentation, and the mixture was incubated at 39°C with shaking at 150 rpm for 24 h. Subsequently, the aerobic fermentation liquid was introduced to sterile grain solid medium at a 10% inoculation rate. The solid medium consisting of wheat bran (80 %) , soybean meal (13 %) and corn flour (7 %) .The mixture was enveloped with three layers of sterile gauze and a lid, and then incubated at 30°C for 72 h. The yeast culture was uniformly distributed on the porcelain tray and air-dried at ambient temperature. The desiccated material was pulverized and sifted through a 40-60 mesh to get the yeast culture.
2.4. Experimental Design and In Vitro Culture Procedure
Three healthy goats (average body weight 31 ± 1.3 kg) fitted with permanent rumen cannulas were used as ruminal fluid donors. The goats were fed twice a day, at 8:00 AM and 6:00 PM, with free access to food and water throughout the day. The composition and nutritional components of the daily diet fed to the goats are shown in Table 1. Before morning feeding, ruminal fluids were collected and stored at a well-insulated and securely sealed thermos bottles preheated to 39°C. Carbon dioxide gas was put into the bottle, and the air was evacuated prior to sealing the lid. Ruminal fluids were promptly filtered through four layers of gauze. The fermentation substrates utilized in this experiment included: 0.49 g of maize, 0.21 g of soybean meal, 0.15 g of oats, and 0.15 g of alfalfa, amounting to a total of 1 g. The formula of this fermentation substrate simulates the proportion of high-concentrate diet feeding practise in the intensive system.In vitro rumen fermentation trial 1:The yeast strain group(NJ-5, NJ-12, NJ-14, NJ-36, NJ-46) was supplemented with 1 mL of prepared yeast suspension, achieving a yeast inoculation level of 2×106 CFU/mL, with three replicates per group.In vitro rumen fermentation trial 2: yeast culture concentrations of 0%, 1%, 2%, and 5% based on dry matter were incorporated into the fermentation substrate, establishing the control group (CON) , low-dose group (LYC) , middle-dose group (MYC) , and high-dose group (HYC) , with three replicates for each group. Artificial saliva was prepared as reported previously by Menke et al.[17]. Subsequent to After thorough mixing, the amalgamation was put in a 39°C constant temperature water bath with a continuous supply of CO2. The combination was subsequently blended in a 1:2 ratio of rumen fluid to artificial saliva, and 60 mL of the mixture was transferred to 100 mL fermentation bottles, with continuous CO2 flow to sustain anaerobic conditions, and was cultured at 39°C with shaking at 150 rpm. The supernatant was sampled at incubated for 6, 12, and 24 h for the assessment of pH value and fermentation parameters. For VFAs measurement, samples were centrifuged at 8000×g for 15 minutes at 4°C, then filtered through a 0.45 µm membrane to collect the supernatant.
1.) The premix provided the following per kg of diets:MnSO4 153 mg,ZnSO4 186 mg,FeSO4 125 mg,CoCl2 8.25 mg,KIO3 25 mg,CuSO4 33 mg,NaSeO3 4 mg,VA 15.28 mg,VE 0.47 mg.2) ME was a calculated value, while the others were measured values.
2.5. Assessment of VFAs
The VFAs were measured using a GC-14B (Shimadzu, Shijota,Japan) to assess the amounts of acetic acid, propionic acid, butyric acid, isovaleric acid, and valeric acid as previously[18]. The concentration of lactic acid was quantified by a commercial test kit (A109-21, Nanjing Jiancheng Bioengineering Institute) by measuring the absorbance at 530 nm.
2.6. DNA Extraction and 16S rDNA Sequencing
The CTAB approach, as outlined by Denman et al.[19], was employed to extract total DNA from rumen fluid samples. Following quality control, PCR amplification of the bacterial 16S rRNA gene V4 region was conducted with universal bacterial primers 515F: 5'-GTGCCAGCMGCCGCGG-3' and 806R: 5'-GGACTACHVGGGTWTCTAAT-3'. The PCR products underwent purification, followed by library building and sequencing using the Novaseq 6000 platform (Meiji Biomedical Technology Co., Ltd., Shanghai) . Following rigorous filtering and database comparison, high-quality data were acquired for further research. The sequences were categorized into operational taxonomic units (OTUs) with Uparse software (v7.0.1001) . The operational taxonomic units (OTUs) were annotated utilizing the Mothur methodology and compared against the SILVA138 SSU rRNA database to acquire taxonomic data and assess microbial abundance across the hierarchical levels of kingdom, phylum, class, order, family, genus, and species. The α-diversity analysis (including the Chao1 index, Shannon index, Simpson index, quantity of phyla, and quantity of genera) and principal coordinate analysis (PCoA) were conducted. Tukey's test and the Wilcoxon test were used to evaluate the intergroup differences.
2.7. Data Analysis
Before analysis, all data were tested for normality and homogeneity of variance.All the data were processed using Excel 2016 and analyzed using IBM SPSS Statistics 26 (IBM, Armonk, NY, USA) . One-way analysis of variance (ANOVA) was performed for comparisons of more than two groups, and Duncan's multiple comparison was used to compare the differences between groups. The data are displayed as means and standard errors (SEM) , and p < 0.05 indicated statistical significance.
3. Results
3.1. Isolation and Identification of Yeast Strains
Total 59 yeast strains (NJ 1~59) derived from goat ruminal fluid were isolated under aerobic circumstances as indicated in Table 2, and then the ITS region of the isolated yeast strains was amplified via PCR and subsequently sequenced. A phylogenetic tree was generated by sequencing based on Genbank data(Figure 1) . These 59 isolated yeast strains were classified into 9 distinct species: Candida rugosa (32.2%) , Pichia kudriavzevii (20.3%) , Trichosporon asahii (15.3%) , Candida tropicalis (10.2%) , Magnusiomyces capitatus (6.8%) , Candida pararugosa (6.8%) , Meyerozyma caribbica (5.1%) , Sporidiobolus pararoseus (1.7%) , and Yarrowia lipolytica (1.7%) , respectively.
3.2. Anaerobic Capacity of Yeast Strains
The results showed that 37 out of 59 yeast strains could proliferate in an anaerobic environment, which were chosen for further screening experiments. Figure 2 showed the growth capacity of the selected 37 yeast strains. The OD600 values varied from 0.4 to 1.7, with 18 strains exceeding an OD600 value of 1, representing 48.6% of the total strains, which demonstrated a robust growth potential and chosen for subsequent testing.
3.3. Evaluation of Acid Tolerance of Yeast Strains
The selected 18 yeast strains were cultured at the condition of pH 5.0, 5.4, 5.8, respectively, to evaluate acid tolerance. As shown in Figure 3, all these yeast strains could endure low pH conditions (pH < 5.8) , and 10 yeast strains exhibited OD600 values more than 0.8, which were finally chosen for further assessment.
3.4. Evaluation of Lactate Utilization Capacity of Yeast Strains
In order to measure the lactate utilization capability, the selected 10 yeast strains were cultured in lactate-enriched media. The results showed that all yeast strains demonstrated varying capacities for lactate consumption, with assimilation rates between 16% and 60%. Among these strains, 5 strains exhibited lactate utilization rates exceeding 50%, and then were chosen for the subsequent tests.
3.5. Effect of Yeast Strains On VFAs Production In Vitro Rumen Fermentation
Compared to the control group (CON) , only NJ-14 yeast strain showed a significant increase in total volatile fatty acid (TVFA) concentration (p < 0.05) . The strains of NJ-5 and NJ-12 markedly elevated acetate concentrations (p < 0.05) , and all five strains greatly enhanced the production of propionate (p < 0.05) , but significantly reduced valerate and isobutyrate levels (p < 0.05) . Moreover, the yeast strains of NJ-14, NJ-36 and NJ-46 markedly decreased the ratio of acetate to propionate (p < 0.05) . After 24 h of fermentation, the pH value produced by NJ-5 yeast strain was markedly elevated compared to the CON group (p < 0.05) , thus NJ-5 was chosen for further investigation.
3.6. Preparation of Yeast Culture
3.7. Impact of Yeast Culture Supplementation on pH Value In Vitro Fermentation
As shown in Table 5, compared to CON group, MYC group and HYC group significantly increased pH value in the fermented liquids after 6 h culture, and after 24 h culture, yeast treatment groups all showed higher level of pH compared to CON (p < 0.05) .
3.8. Effect of Yeast Culture on VFAs Concentrations In Vitro Fermentation
Compared to CON group, the total concentration of VFAs in the HYC group was markedly elevated, but the proportion of butyrate was dramatically reduced (p < 0.05) . LYC, MYC and HYC groups exhibited considerably elevated percentages of acetate and the ratio of acetate to propionate (p < 0.05) , however, the proportion of propionate was markedly reduced compared to CON.
3.9. Impact of Yeast Culture on bacteria Community In Vitro Fermentation
3.9.1. Rumen Bacterial Diversity Analysis
In vitro rumen fermentation liquid microbiota was characterized and quantitatively analyzed via 16S rDNA sequencing. Venn diagram showed that there were 3,070 ASV in the CON group, 3,100 in the LYC group, 3,778 in the MYC group and 3,305 in the HYC group,with 489 ASV common to all four groups. (Figure 6A) .The alpha diversity index revealed that the index of Chao1, Shannon ,Pielou_e and Simpson exhibited no statistically significant differences were noted between the groups (p > 0.05) (Figure 6B) . The bacterial communities of the LYC, MYC, and HYC groups exhibited distinct clustering(Figure 6C) , signifying analogous microbial compositions within these groups, and adding yeast culture enriched the bacteria community in vitro fermentation medium.
3.9.2. Impact of Yeast Culture on Bacteria Abundance in vitro Fermentation
As shown in Figure 7, the predominant phyla in the in vitro rumen fermentation fluids were Firmicutes, Bacteroidetes and Proteobacteria(Figure 7 A) .Figure 7 C and D use heatmaps to show the abundance differents of bacterial communities at the phylum and genus levels under different treatment groups.In comparison to the CON group, the LYC, MYC, and HYC groups exhibited a significant increase in the relative abundance of Desulfobacterota (p < 0.05) , but a significant decrease in the relative abundance of Cyanobacteria (p < 0.05) (Figure 7 A and E) . The LYC group exhibited a substantial increase in the relative abundance of Fibrobacterota (p < 0.05) , whereas the MYC and HYC groups had a significant decrease in the relative abundance of Bdellovibrionota (p < 0.05) (Figure 7 A and E) . At the genus level, the LYC and MYC groups considerably augmented the relative abundance of Ruminococcus, Selenomonas, and Rumiobacter compared to the CON group (p < 0.05) , while dramatically reduced the relative abundance of Succinivibrioaceae_UCG-002 and Succinivibrio (p < 0.05) (Figure 7 B and F) . HYC group exhibited a considerable increase in the relative abundance of Christensenell-aceae_R-7_group, F082, and Ruminococcus (p < 0.05) , while considerably decreased the relative abundance of Succinivibrioaceae_UCG-002, E-nterobacter, and Succinivibrio compared to CON (p < 0.05) (Figure 7 B and F) .
Based on the linear discriminant analysis effect size (LEfSe) analysis (p < 0.05, LDA > 2.0) revealed a significant difference in the relative abundance of the 15 detected bacterial taxa (Figure 8) .The order Bacillales, order Bifidobacteriales, family Bifidobacteriaceae, family Enterobacteriaceae, genus Bifidobacterium and genus Enterobacter were the dominant bacterial groups enriched in the CON group.The order Oscillospirales, order Peptostreptococcales_Tissierellales,familyAnaerovoracaceae,genus Family_XIII_AD3011_group ,genus NK4A214_group and genus Prevotellaceae_NK3B31_group were enriched in the LYC group.The family Bacteroidales_RF16_group and genus Bacteroidales_RF16_group were enriched in the HYC group.
4. Discussion
In this study, total 59 yeast strains were successfully isolated from goat ruminal fluids, and classified into 9 yeast species. The isolated yeasts exhibited characteristic morphological features of rumen yeasts, displaying a white to milky-white hue with smooth and slightly convex surfaces. Previous studies showed that yeasts from the genus Candida (Candida albicans, Candida parapsilosis, Candida tropicalis, and Candida rugosa) [20], Trichosporon asahii [21], and Pichia kudriavzevii [22,23] can be isolated from the rumen contents of ruminants. Consistently, in the present study, among the isolated species, Candida parapsilosis, Candida rugosa, and Picha kudriavzevii have been previously recognized for their probiotic potential. Saccharomyces cerevisiae is commonly used as a probiotic in ruminant nutrition [24], unfortunately, which was not isolated in this study. Under optimum conditions, the rumen maintains an anaerobic environment, sustaining a temperature of 38.5 to 40.0℃ and a pH of 6.0 to 7.0. Subacute ruminal acidosis (SARA) arises when the rumen pH declines below 5.8, leading to alterations in the rumen microbial composition. The aim of this study was to develop yeast strains capable of thriving in anaerobic rumen environments, demonstrating vigorous growth at 39°C, adapting to the low pH conditions typical of SARA, and efficiently metabolizing lactate. Ten out of 59 isolated yeast strains demonstrated consistent survival under SARA conditions, as determined by evaluations of anaerobic tolerance, growth potential, and low pH adaptation. The collection included five strains of Candida rugosa, three strains of Picha kudriavzevii, and one strain each of Candida pararugosa,Candida tropicalis, and Trichosporon asahii. Previous studies demonstrate that Candida rugosa, Picha kudriavzevii, and Candida pararugosa exhibit anaerobic tolerance and acid resistance, which was consistent with our findings[21].It’s well known that under SARA conditions, lactic acid accumulates in the rumen, leading to a reduction in pH and negatively impacting rumen health. Therefore, reducing lactic acid levels is crucial for maintaining rumen pH balance, enhancing rumen fermentation and increasing ruminant productivity. Our findings revealed that Candida rugosa, Picha kudriavzevii, Candida tropicalis, and Trichosporon asahii had varying capacity for lactate utilization, with Candida rugosa, Picha kudriavzevii, and Candida pararugosa achieving utilization rates exceeding 50%. Similarly, significant populations of Picha kudriavzevii were detectable in the rumen of dairy cows suffering from SARA, showcasing its strong lactate-utilizing ability[25]. The condition of the rumen is crucial for the general well-being and production of ruminants. The pH of the rumen is a vital indicator of fermentation status, with the optimal range being 6.2~6.8[26,27]. Numerous studies suggest that yeast supplementation may reduce lactate accumulation and enhance microbial diversity, hence increasing rumen pH[28,29]. In our experiment, the addition of five different yeast strains did not significantly alter the pH of in vitro rumen fermentation fluid, as the previous reports [30,31]. It’s reasonable to speculate that dietary supplementation with yeast products derived from ruminal fluids can protect ruminants against SARA suffering due to the higher utilization of lactic acid.
Among the selected strains, Candida rugosa (NJ-5) was identified as the most promising probiotic candidate. This strain has been previously isolated from the rumen of dairy cattle and has exhibited probiotic activity [12,32]. In agreement with the beneficial traits discovered in our study, it’s the first time to report that Candida rugosa (NJ-5) shows potential as a probiotic for alleviating SARA caused by high-concentrate diets . Yeast culture products are generated by first cultivating the yeast strain in a liquid medium, then performing solid-state fermentation, and then drying the outcome. These products consist of living yeast cells, extracellular metabolites (such as vitamins and enzymes) , and the denatured fermentation media [34]. Yeast culture is widely employed as a sustainable feed additive in ruminant production owing to its positive effects on productive performance[33,34]. Currently, most commercial yeast culture products employ Saccharomyces cerevisiae, with improved strains being selected or genetically engineered. Yeasts originating from the rumen grant them more capable of flourishing in ruminal low pH and anaerobic environment and more effective at managing rumen microbial populations. In this study, the yeast strain of Candida rugosa was chosen for its ability to alleviate SARA caused by high-concentrate diets. Under normal conditions, the pH of the rumen is maintained at roughly 6.5. However, when ruminants are fed with a high-concentrate diet, it’s easily led to the accumulation of both VFAs and lactic acid, thus lowering the pH value below 5.8 and finally causing SARA[35]. Our experimental results demonstrated that the addition of yeast culture led to a dose-dependent increase in the pH of fermentation fluid. Adding 5% yeast culture can successfully mitigate the pH decrease, which was consistent with the results conducted on sheep[36].
It’s reported that yeast products can enhance the activity of rumen bacteria in metabolizing lactic acid into VFAs, thereby reducing lactate accumulation and elevating ruminal pH value[37]. The incorporation of yeast culture significantly elevated the total volatile fatty acids (VFAs) and acetate levels, whereas the concentrations of valerate isovalerate and isobutyrate remained relatively unchanged. Propionate, a significant gluconeogenic precursor, substantially influences energy utilization in ruminants[38]. Our study revealed that yeast culture supplementation redirected the fermentation pathway of high-concentrate substrates, transitioning from propionate-dominant fermentation to acetate-dominant fermentation. Han et al.[39]found that the addition of yeast culture in sheep improved the diversity and richness of rumen microbiota. Similarly, it’s reported that adding yeast fermentation products to dairy cows reduced the microbial diversity caused by SARA[40]. However, our results showed that yeast culture did not have a big effect on the variety and abundance of microbiota in the in vitro fermentation system as reported by Dai et al.[41]. The different effects of yeast culture on α-diversity of rumen microbes could be due to different animal models, food types, yeast cultures, and dosages. The primary bacterial phyla in this experiment were Bacteroidota, Firmicutes, and Proteobacteria. The phylum Proteobacteria includes some pathogenic species, such as Escherichia and Salmonella, associated with gastrointestinal diseases and inflammation. The HYC group showed a reduction in Proteobacteria abundance compared to the CON group, suggesting that high-dose yeast cultivation could potentially enhance animal immunity[42]. Fibrobacterota is an essential fibrolytic phylum in the rumen, adept at converting low-quality feed into volatile fatty acids[43]. At the genus level, the abundance of Streptococcus was significantly reduced by three doses of YC supplementation. Meanwhile, Selenomonas, a well-known genus that breaks down lactate, showed a noticeable rise in the LYC and MYC groups and a higher relative abundance in the HYC group compared to the control group. These results indicate that YC may alleviate SARA disease by promoting the growth of bacteria that use lactate as a substrate, such as Selenomonas, and then to decrease the amount of lactate in the rumen. Additionally, YC significantly decreased the abundance of Succinivibrionaceae_UCG-001, which primarily produces succinate and competes with methanogens for hydrogen [44]. The relative abundance of F082 and Fibrobacter was also increased in the YC-supplemented groups compared to the control, indicating that YC may boost fiber degradation by fostering the growth and activity of fibrolytic bacteria, which ultimately improve animal welfare and productive performance.
5. Conclusion
In this study, total 59 yeast strains were successfully isolated from goat ruminal fluids, and Candida rugosa (NJ-5) was selected as the most promising probiotic yeast to improve fermented environment in the rumen particularly under high-concentrate diets. In fact, adding Candida rugosa (NJ-5) yeast culture can mitigate the reduction of pH in fermentation , increased total VFA and acetate levels through remodeling rumen microbial composition with higher abundance of Desulfobacterota, Christensenellaceae_R-7_group, F082 and Ruminococcus, but lower level of Cyanobacteria, Bdellovibrionota, Succinivibrioaceae_UCG-002, Enterobacter and Succinivibrio.
Author Contributions
P.S., X.Y. and Y.N. designed the research. P.S. performed the experiment. The data collection, analysis, and article writing were completed by P.S., X.Y., M.H., Y.C. and L.L. Y.F. and Y.N. revised the article. All authors have read and agreed to the published version of the manuscript
Funding
This research was funded by the National Natural Science Foundation of China (Grant No.32172810) .
Institutional Review Board Statement
The animal experimental procedures of this study were approved by Institutional Animal Care and Use Committee of Nanjing Agricultural University (NJAU.No20221101207) .
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to privacy.
Acknowledgments
We sincerely thank all members for assisting in conducting the experiments, technical support, and data analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Campanile, G.; Zicarelli, F.; Vecchio, D.; Pacelli, C.; Neglia, G.; Balestrieri, A.; Di Palo, R.; Infascelli, F. Effects of Saccharomyces Cerevisiae on in Vivo Organic Matter Digestibility and Milk Yield in Buffalo Cows. Livestock Science 2008, 114, 358–361. [Google Scholar] [CrossRef]
- Mitchell, L.K.; Heinrichs, A.J. Feeding Various Forages and Live Yeast Culture on Weaned Dairy Calf Intake, Growth, Nutrient Digestibility, and Ruminal Fermentation. Journal of Dairy Science 2020, 103, 8880–8897. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Su, M.; Wang, C.; Li, D.; Li, Q.; Liu, Z.; Qi, X.; Wu, Y.; Zhao, Y.; Li, T.; et al. Yeast Culture Repairs Rumen Epithelial Injury by Regulating Microbial Communities and Metabolites in Sheep. Front. Microbiol. 2023, 14, 1305772. [Google Scholar] [CrossRef]
- Halfen, J.; Carpinelli, N.; Del Pino, F.A.B.; Chapman, J.D.; Sharman, E.D.; Anderson, J.L.; Osorio, J.S. Effects of Yeast Culture Supplementation on Lactation Performance and Rumen Fermentation Profile and Microbial Abundance in Mid-Lactation Holstein Dairy Cows. Journal of Dairy Science 2021, 104, 11580–11592. [Google Scholar] [CrossRef]
- Bach, A.; Guasch, I.; Elcoso, G.; Chaucheyras-Durand, F.; Castex, M.; Fàbregas, F.; Garcia-Fruitos, E.; Aris, A. Changes in Gene Expression in the Rumen and Colon Epithelia during the Dry Period through Lactation of Dairy Cows and Effects of Live Yeast Supplementation. Journal of Dairy Science 2018, 101, 2631–2640. [Google Scholar] [CrossRef]
- Ghazanfar, S. Understanding the Mechanism of Action of Indigenous Target Probiotic Yeast: Linking the Manipulation of Gut Microbiota and Performance in Animals. In Saccharomyces; Peixoto Basso, T., Carlos Basso, L., Eds.; IntechOpen, 2021 ISBN 978-1-83968-789-1.
- Dias, J.D.L.; Silva, R.B.; Fernandes, T.; Barbosa, E.F.; Graças, L.E.C.; Araujo, R.C.; Pereira, R.A.N.; Pereira, M.N. Yeast Culture Increased Plasma Niacin Concentration, Evaporative Heat Loss, and Feed Efficiency of Dairy Cows in a Hot Environment. Journal of Dairy Science 2018, 101, 5924–5936. [Google Scholar] [CrossRef] [PubMed]
- Kung, L.; Kreck, E.M.; Tung, R.S.; Hession, A.O.; Sheperd, A.C.; Cohen, M.A.; Swain, H.E.; Leedle, J.A.Z. Effects of a Live Yeast Culture and Enzymes on In Vitro Ruminal Fermentation and Milk Production of Dairy Cows. Journal of Dairy Science 1997, 80, 2045–2051. [Google Scholar] [CrossRef]
- Bajagai. Yadav S Probiotics in Animal Nutrition: Production, Impact and Regulation; FAO animal production and health paper; Food and Agricultural Organization of the United Nations: Rome, 2016; ISBN 978-92-5-109333-7. [Google Scholar]
- De Melo Pereira, G.V.; De Oliveira Coelho, B.; Magalhães Júnior, A.I.; Thomaz-Soccol, V.; Soccol, C.R. How to Select a Probiotic? A Review and Update of Methods and Criteria. Biotechnology Advances 2018, 36, 2060–2076. [Google Scholar] [CrossRef]
- Tunkala, B.Z.; DiGiacomo, K.; Alvarez Hess, P.S.; Gardiner, C.P.; Suleria, H.; Leury, B.J.; Dunshea, F.R. Evaluation of Legumes for Fermentability and Protein Fractions Using in Vitro Rumen Fermentation. Animal Feed Science and Technology 2023, 305, 115777. [Google Scholar] [CrossRef]
- Sirisan, V.; Pattarajinda, V.; Vichitphan, K.; Leesing, R. Isolation, Identification and Growth Determination of Lactic Acid-Utilizing Yeasts from the Ruminal Fluid of Dairy Cattle. Lett Appl Microbiol 2013, 57, 102–107. [Google Scholar] [CrossRef]
- Marrero, Y.; Castillo, Y.; Ruiz, O.; Burrola, E.; Angulo, C. Feeding of Yeast (Candida Spp.) Improves in Vitro Ruminal Fermentation of Fibrous Substrates. Journal of Integrative Agriculture 2015, 14, 514–519. [Google Scholar] [CrossRef]
- Ruiz, O.; Castillo, Y.; Arzola, C.; Burrola, E.; Salinas, J.; Corral, A.; Hume, M.E.; Murillo, M.; Itza, M. Effects of Candida Norvegensis Live Cells on In Vitro Oat Straw Rumen Fermentation. Asian Australas. J. Anim. Sci 2015, 29, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Z.; Jin, W.; Mao, S. Isolation and Characterization of Ruminal Yeast Strain with Probiotic Potential and Its Effects on Growth Performance, Nutrients Digestibility, Rumen Fermentation and Microbiota of Hu Sheep. JoF 2022, 8, 1260. [Google Scholar] [CrossRef]
- Carvalho, B.F.; Ávila, C.L.S.; Bernardes, T.F.; Pereira, M.N.; Santos, C.; Schwan, R.F. Fermentation Profile and Identification of Lactic Acid Bacteria and Yeasts of Rehydrated Corn Kernel Silage. J Appl Microbiol 2017, 122, 589–600. [Google Scholar] [CrossRef]
- Menke, H. H; Steingass, H Estimation of the Energetic Feed Value Obtained from Chemical Analysis and in Vitro Gas Production Using Rumen Fluid. Animal Research and Development 1988, 28, 7–55. [Google Scholar]
- Xue, Y.; Lin, L.; Hu, F.; Zhu, W.; Mao, S. Disruption of Ruminal Homeostasis by Malnutrition Involved in Systemic Ruminal Microbiota-Host Interactions in a Pregnant Sheep Model. Microbiome 2020, 8, 138. [Google Scholar] [CrossRef] [PubMed]
- Denman, S.E.; McSweeney, C.S. Development of a Real-Time PCR Assay for Monitoring Anaerobic Fungal and Cellulolytic Bacterial Populations within the Rumen: Real-Time PCR Assay of the Rumen Anaerobic Fungal Population. FEMS Microbiology Ecology 2006, 58, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Suntara, C.; Cherdthong, A.; Wanapat, M.; Uriyapongson, S.; Leelavatcharamas, V.; Sawaengkaew, J.; Chanjula, P.; Foiklang, S. Isolation and Characterization of Yeasts from Rumen Fluids for Potential Use as Additives in Ruminant Feeding. Veterinary Sciences 2021, 8, 52. [Google Scholar] [CrossRef]
- Sales, G.F.C.; Carvalho, B.F.; Schwan, R.F.; Pereira, M.N.; Ávila, C.L.S. Diversity and Probiotic Characterisation of Yeast Isolates in the Bovine Gastrointestinal Tract. Antonie van Leeuwenhoek 2023, 116, 1123–1137. [Google Scholar] [CrossRef]
- Rahbar Saadat, Y.; Yari Khosroushahi, A.; Movassaghpour, A.A.; Talebi, M.; Pourghassem Gargari, B. Modulatory Role of Exopolysaccharides of Kluyveromyces Marxianus and Pichia Kudriavzevii as Probiotic Yeasts from Dairy Products in Human Colon Cancer Cells. Journal of Functional Foods 2020, 64, 103675. [Google Scholar] [CrossRef]
- Chelliah, R.; Ramakrishnan, S.R.; Prabhu, P.R.; Antony, U. Evaluation of Antimicrobial Activity and Probiotic Properties of Wild-strain Pichia Kudriavzevii Isolated from Frozen Idli Batter. Yeast 2016, 33, 385–401. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Ishaq, S.L.; Rodriguez-Herrera, M.V.; Garcia-Hernandez, C.A.; Kawas, J.R.; Nagaraja, T.G. Review: Are There Indigenous Saccharomyces in the Digestive Tract of Livestock Animal Species? Implications for Health, Nutrition and Productivity Traits. Animal 2020, 14, 22–30. [Google Scholar] [CrossRef]
- Wang, D.S.; Zhang, R.Y.; Zhu, W.Y.; Mao, S.Y. Effects of Subacute Ruminal Acidosis Challenges on Fermentation and Biogenic Amines in the Rumen of Dairy Cows. Livestock Science 2013, 155, 262–272. [Google Scholar] [CrossRef]
- Russell, J.B. The Importance of pH in the Regulation of Ruminal Acetate to Propionate Ratio and Methane Production In Vitro. Journal of Dairy Science 1998, 81, 3222–3230. [Google Scholar] [CrossRef]
- Hou, M.; Song, P.; Chen, Y.; Yang, X.; Chen, P.; Cao, A.; Ni, Y. Bile Acids Supplementation Improves Colonic Mucosal Barrier via Alteration of Bile Acids Metabolism and Gut Microbiota Composition in Goats with Subacute Ruminal Acidosis (SARA). Ecotoxicology and Environmental Safety 2024, 287, 117313. [Google Scholar] [CrossRef] [PubMed]
- Silberberg, M.; Chaucheyras-Durand, F.; Commun, L.; Mialon, M.M.; Monteils, V.; Mosoni, P.; Morgavi, D.P.; Martin, C. Repeated Acidosis Challenges and Live Yeast Supplementation Shape Rumen Microbiota and Fermentations and Modulate Inflammatory Status in Sheep. Animal 2013, 7, 1910–1920. [Google Scholar] [CrossRef] [PubMed]
- Thrune, M.; Bach, A.; Ruiz-Moreno, M.; Stern, M.D.; Linn, J.G. Effects of Saccharomyces Cerevisiae on Ruminal pH and Microbial Fermentation in Dairy Cows. Livestock Science 2009, 124, 261–265. [Google Scholar] [CrossRef]
- Ferraretto, L.F.; Shaver, R.D.; Bertics, S.J. Effect of Dietary Supplementation with Live-Cell Yeast at Two Dosages on Lactation Performance, Ruminal Fermentation, and Total-Tract Nutrient Digestibility in Dairy Cows. Journal of Dairy Science 2012, 95, 4017–4028. [Google Scholar] [CrossRef]
- Bayat, A.R.; Kairenius, P.; Stefański, T.; Leskinen, H.; Comtet-Marre, S.; Forano, E.; Chaucheyras-Durand, F.; Shingfield, K.J. Effect of Camelina Oil or Live Yeasts (Saccharomyces Cerevisiae) on Ruminal Methane Production, Rumen Fermentation, and Milk Fatty Acid Composition in Lactating Cows Fed Grass Silage Diets. Journal of Dairy Science 2015, 98, 3166–3181. [Google Scholar] [CrossRef]
- Fernandes, T.; Carvalho, B.F.; Mantovani, H.C.; Schwan, R.F.; Ávila, C.L.S. Identification and Characterization of Yeasts from Bovine Rumen for Potential Use as Probiotics. J Appl Microbiol 2019, 127, 845–855. [Google Scholar] [CrossRef]
- Pang, Y.; Zhang, H.; Wen, H.; Wan, H.; Wu, H.; Chen, Y.; Li, S.; Zhang, L.; Sun, X.; Li, B.; et al. Yeast Probiotic and Yeast Products in Enhancing Livestock Feeds Utilization and Performance: An Overview. JoF 2022, 8, 1191. [Google Scholar] [CrossRef] [PubMed]
- Bass, B.E.; Tsai, T.-C.; Yang, H.; Perez, V.; Holzgraefe, D.; Chewning, J.; Frank, J.W.; Maxwell, C.V. Influence of a Whole Yeast Product ( Pichia Guilliermondii ) Fed throughout Gestation and Lactation on Performance and Immune Parameters of the Sow and Litter. Journal of Animal Science 2019, 97, 1671–1678. [Google Scholar] [CrossRef]
- Ma, J.; Shah, A.M.; Wang, Z.; Fan, X. Potential Protective Effects of Thiamine Supplementation on the Ruminal Epithelium Damage during Subacute Ruminal Acidosis. Animal Science Journal 2021, 92, e13579. [Google Scholar] [CrossRef]
- Wang, H.; Liu, G.; Zhou, A.; Yang, H.; Kang, K.; Ahmed, S.; Li, B.; Farooq, U.; Hou, F.; Wang, C.; et al. Effects of Yeast Culture on in Vitro Ruminal Fermentation and Microbial Community of High Concentrate Diet in Sheep. AMB Expr 2024, 14, 37. [Google Scholar] [CrossRef] [PubMed]
- Desnoyers, M.; Giger-Reverdin, S.; Bertin, G.; Duvaux-Ponter, C.; Sauvant, D. Meta-Analysis of the Influence of Saccharomyces Cerevisiae Supplementation on Ruminal Parameters and Milk Production of Ruminants. Journal of Dairy Science 2009, 92, 1620–1632. [Google Scholar] [CrossRef]
- Lu, G.; Li, Y.; Mao, K.; Zang, Y.; Zhao, X.; Qiu, Q.; Qu, M.; Ouyang, K. Effects of Rumen-Protected Creatine Pyruvate on Meat Quality, Hepatic Gluconeogenesis, and Muscle Energy Metabolism of Long-Distance Transported Beef Cattle. Front. Anim. Sci. 2022, 3, 904503. [Google Scholar] [CrossRef]
- Han, G.; Gao, X.; Duan, J.; Zhang, H.; Zheng, Y.; He, J.; Huo, N.; Pei, C.; Li, H.; Gu, S. Effects of Yeasts on Rumen Bacterial Flora, Abnormal Metabolites, and Blood Gas in Sheep with Induced Subacute Ruminal Acidosis. Animal Feed Science and Technology 2021, 280, 115042. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, Z.; Guan, L.L.; Yoon, I.; Plaizier, J.C.; Khafipour, E. Postbiotics from Saccharomyces Cerevisiae Fermentation Stabilize Microbiota in Rumen Liquid Digesta during Grain-Based Subacute Ruminal Acidosis (SARA) in Lactating Dairy Cows. J Animal Sci Biotechnol 2024, 15, 101. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Liu, Y.; Kong, F.; Guo, C.; Dong, C.; Xu, X.; Li, S.; Wang, W. Saccharomyces Cerevisiae Culture’s Dose–Response Effects on Ruminal Nutrient Digestibility and Microbial Community: An In Vitro Study. Fermentation 2023, 9, 411. [Google Scholar] [CrossRef]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial Signature of Dysbiosis in Gut Microbiota. Trends in Biotechnology 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Li, Q.S.; Wang, R.; Ma, Z.Y.; Zhang, X.M.; Jiao, J.Z.; Zhang, Z.G.; Ungerfeld, E.M.; Le Yi, K.; Zhang, B.Z.; Long, L.; et al. Dietary Selection of Metabolically Distinct Microorganisms Drives Hydrogen Metabolism in Ruminants. The ISME Journal 2022, 16, 2535–2546. [Google Scholar] [CrossRef] [PubMed]
- McCabe, M.S.; Cormican, P.; Keogh, K.; O’Connor, A.; O’Hara, E.; Palladino, R.A.; Kenny, D.A.; Waters, S.M. Illumina MiSeq Phylogenetic Amplicon Sequencing Shows a Large Reduction of an Uncharacterised Succinivibrionaceae and an Increase of the Methanobrevibacter Gottschalkii Clade in Feed Restricted Cattle. PLoS ONE 2015, 10, e0133234. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic tree construction of yeast strains strains.
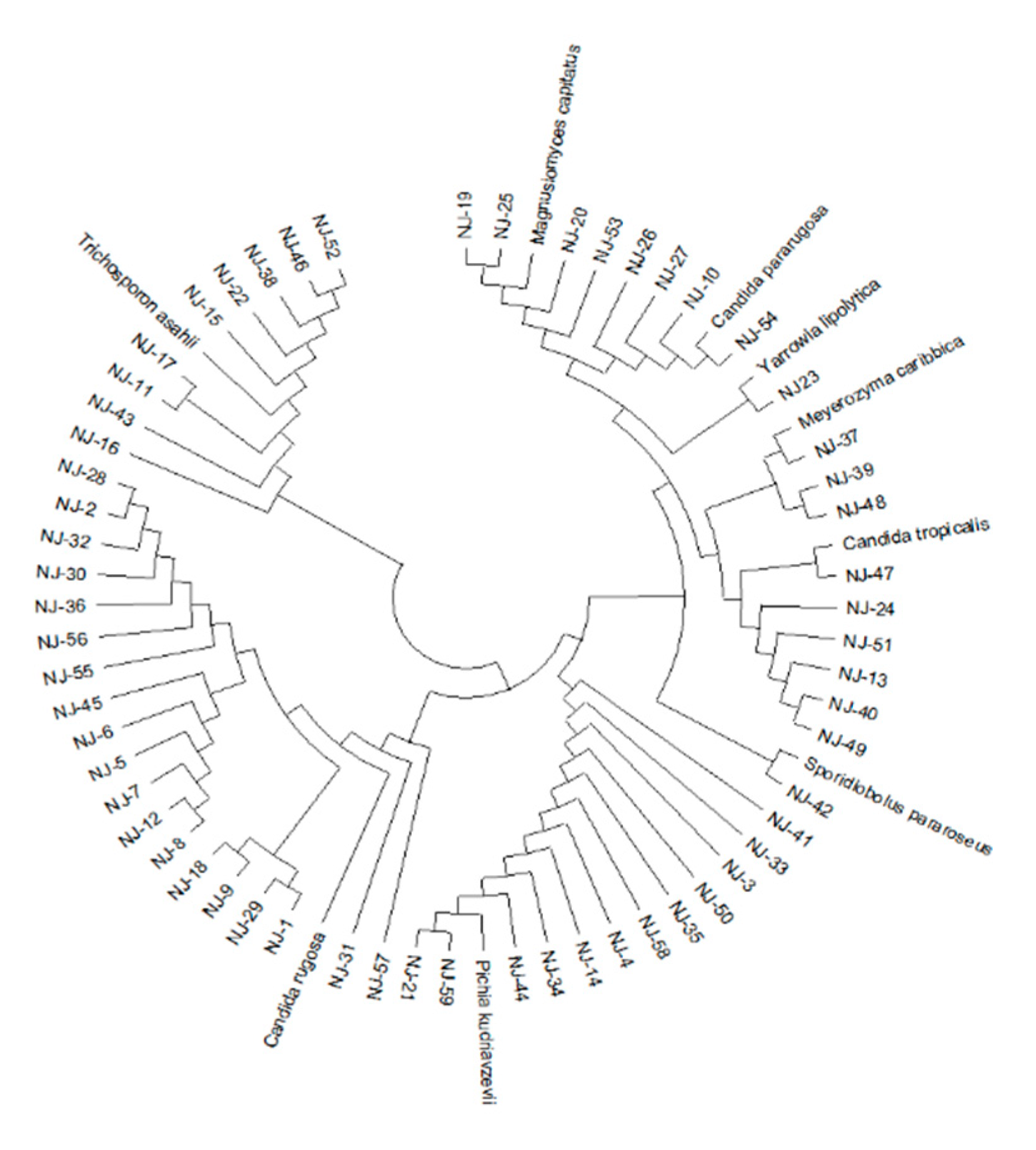
Figure 2.
Growth capacity of yeast strains.
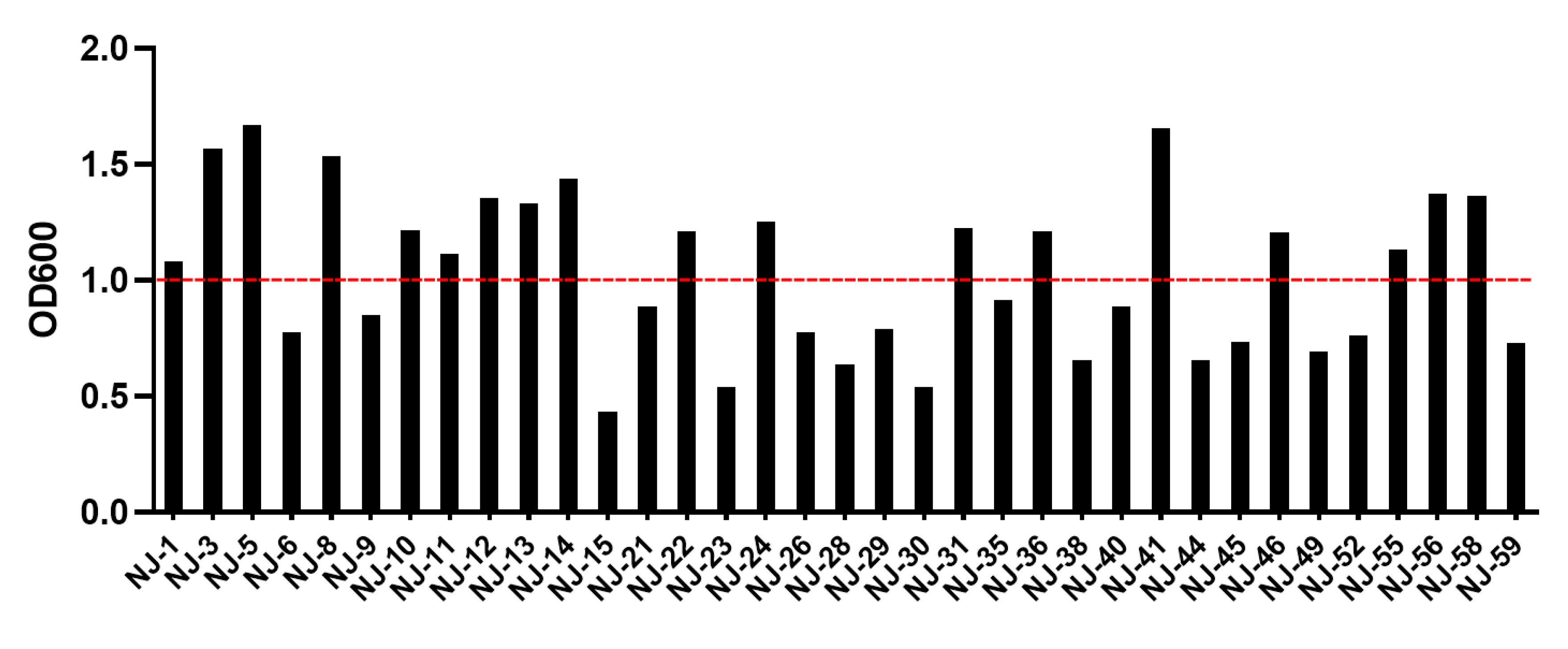
Figure 3.
Acid resistance capacity of yeast strains.
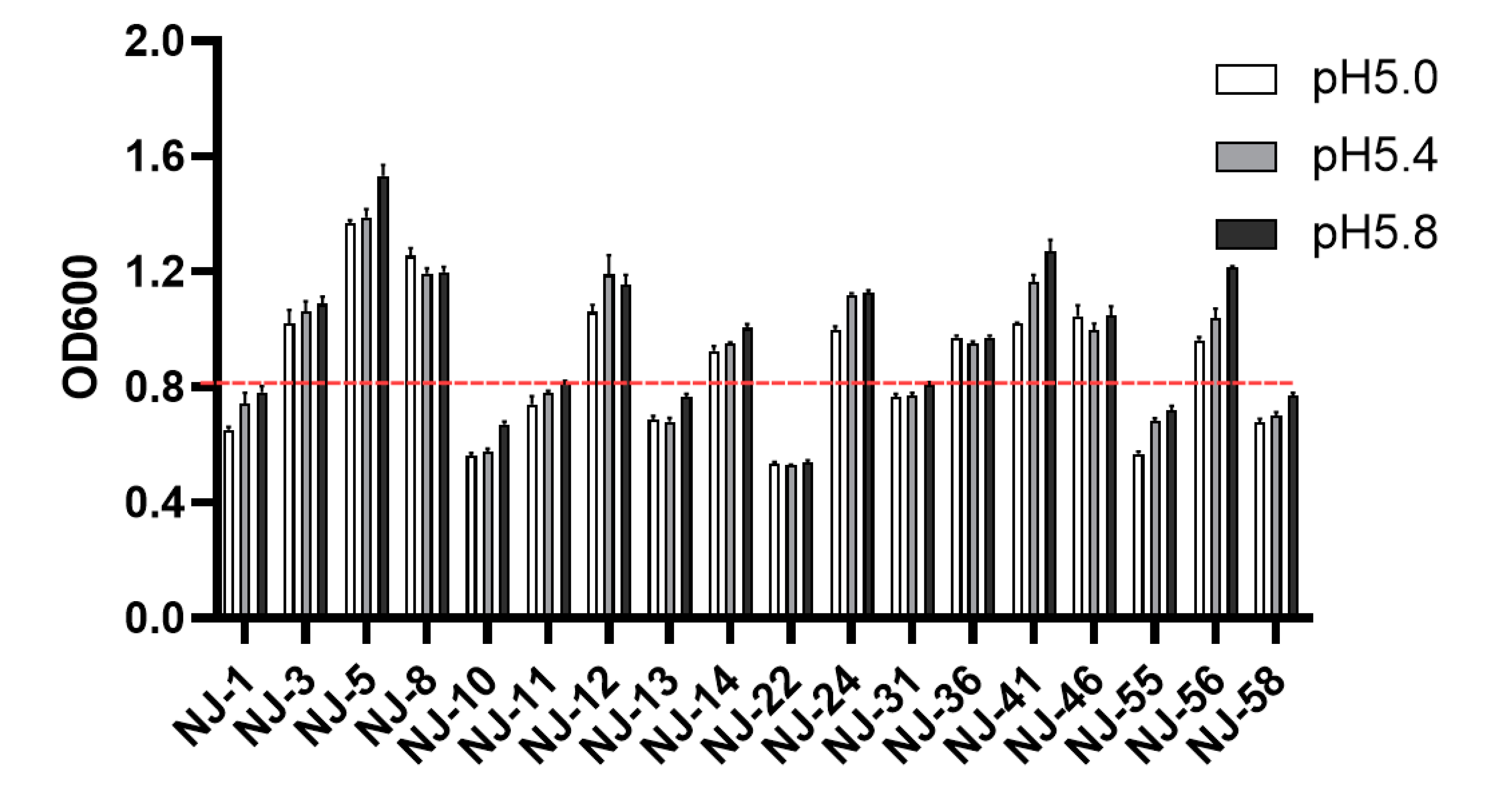
Figure 4.
Lactate utilization capacity of yeast strains.
Figure 4.
The picture of Yeast culture sample.

Figure 6.
Rumen bacterial diversity analysis.(A) Venn diagram showing the number of ASV. (B) Alpha diversity including Chao1 , Shannon, Pielou_e and Simpson diversity index on ASV. (C) Beta diversity analysis based on PLS-DA analysis on ASV.
Figure 6.
Rumen bacterial diversity analysis.(A) Venn diagram showing the number of ASV. (B) Alpha diversity including Chao1 , Shannon, Pielou_e and Simpson diversity index on ASV. (C) Beta diversity analysis based on PLS-DA analysis on ASV.
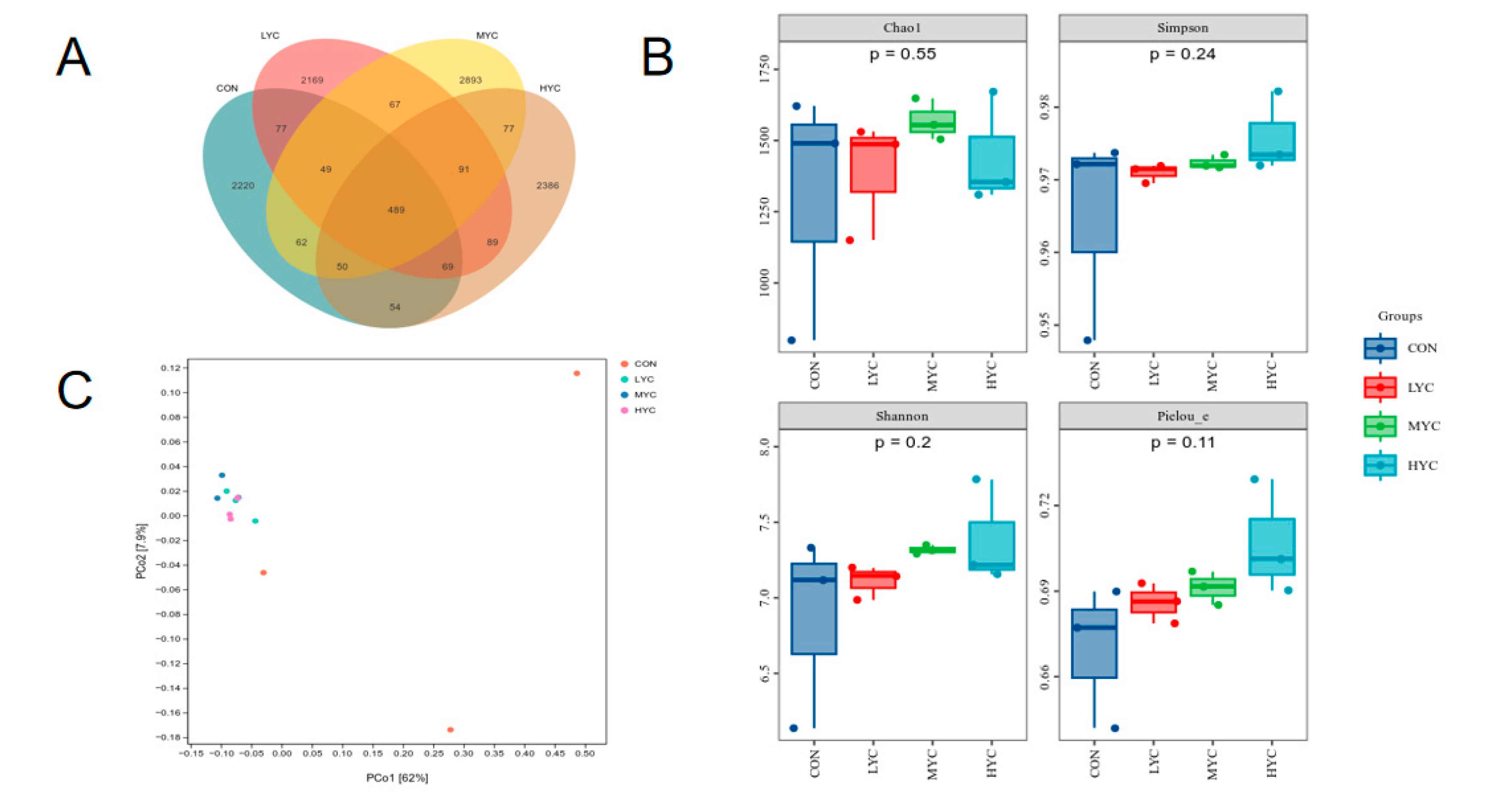
Figure 7.
Effects of adding yeast culture on the bacterial community in goat in vitro rumen fermentation fluid.Relative abundance of bacteria at the phylum level (A) and the genus level (B) ; Heatmap of species composition at the phylum level (C) and the genus level (D) ; Differential abundance at the phylum level (E) and the genus level (F) .
Figure 7.
Effects of adding yeast culture on the bacterial community in goat in vitro rumen fermentation fluid.Relative abundance of bacteria at the phylum level (A) and the genus level (B) ; Heatmap of species composition at the phylum level (C) and the genus level (D) ; Differential abundance at the phylum level (E) and the genus level (F) .
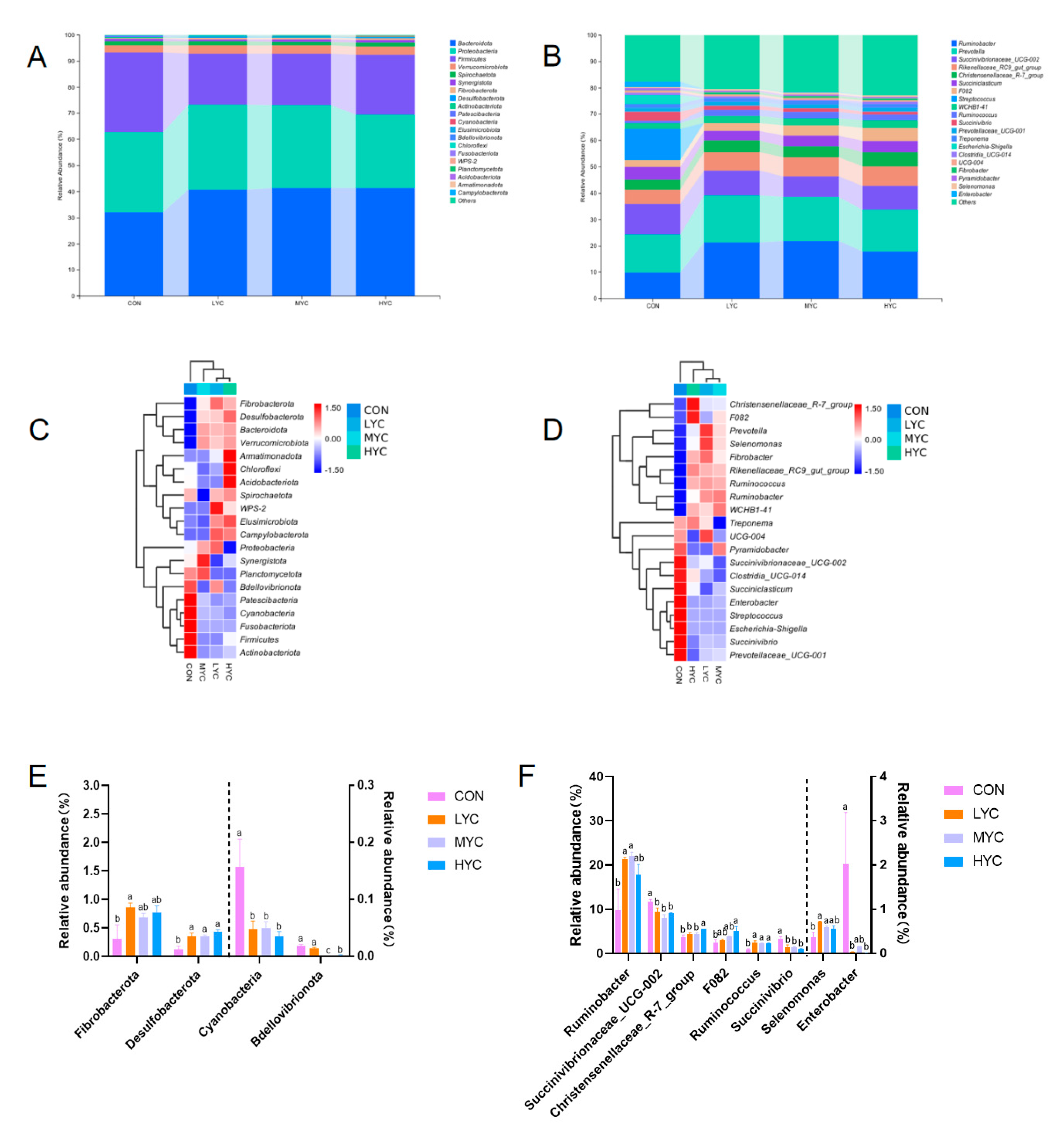
Figure 8.
Figure 8. Differences in the compositon of the ruminal microbiome among different groups. The differential abundant bacteria among the groups are presented in the LDA cladogram generated via LEfSe analysis (p < 0.05, LDA > 2.0) (A and B) . Heatmap showing significantly different microbiota at the order, family and genus levels (C) .
Figure 8.
Figure 8. Differences in the compositon of the ruminal microbiome among different groups. The differential abundant bacteria among the groups are presented in the LDA cladogram generated via LEfSe analysis (p < 0.05, LDA > 2.0) (A and B) . Heatmap showing significantly different microbiota at the order, family and genus levels (C) .
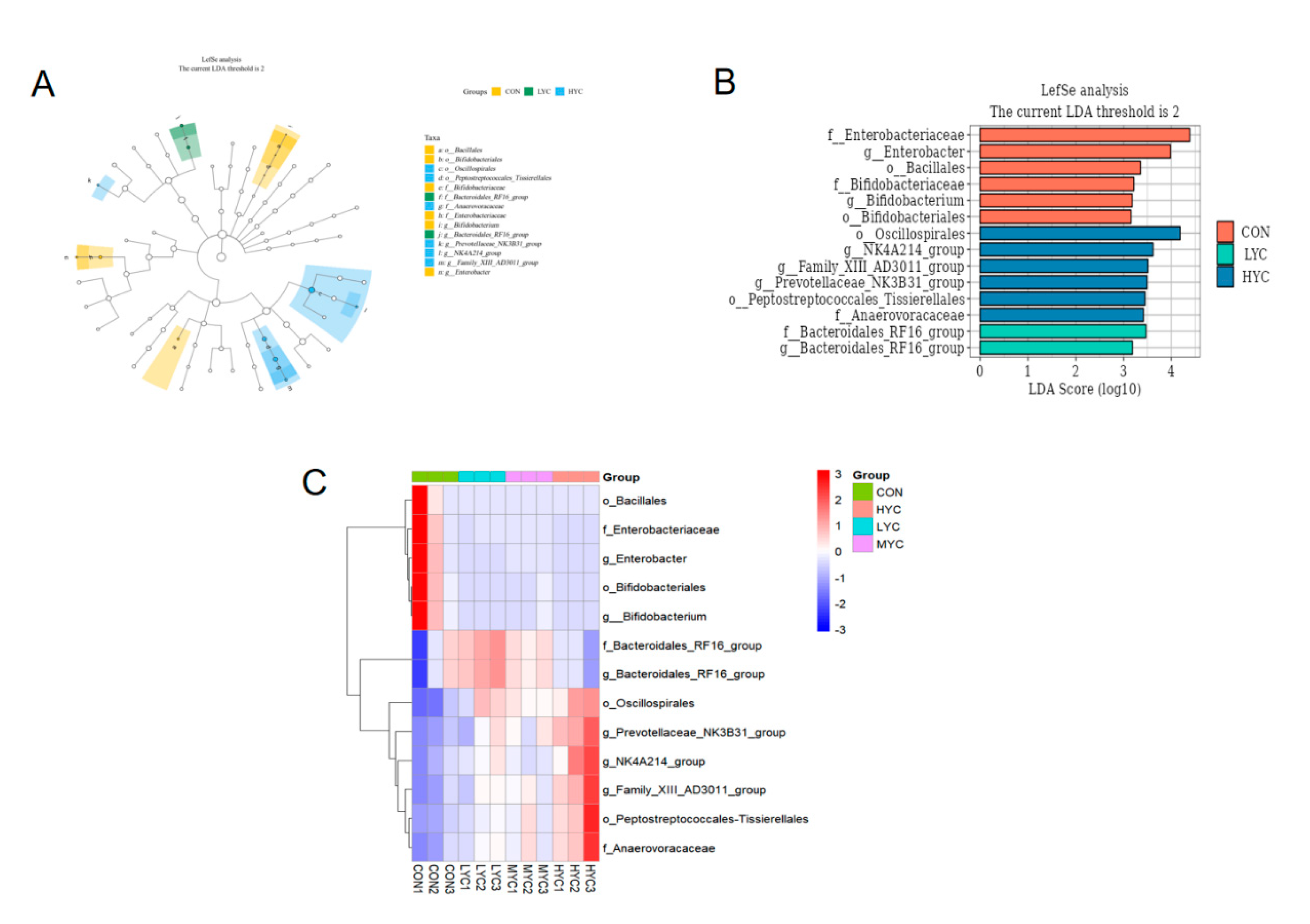

Table 1.
Composition and nutrient levels of the basal diet (DM basis) .
| Diet ingredients % | Nutritional Composition | ||
|---|---|---|---|
| Alfalfa | 70 | ME (MJ/kg) | 8.81 |
| Corn | 14 | CP(%) | 10.81 |
| Soybean meal | 13 | NDF(%) | 44.82 |
| Calcium hydrophosphate | 1.42 | ADF(%) | 23.89 |
| Limestone | 0.58 | Starch(%) | 23.16 |
| NaCl | 0.5 | Calcium(%) | 0.81 |
| Premix1 | 0.5 | Phosphorus(%) | 0.47 |
| Total | 100 |
Table 2.
The 59 yeast strains investigated in the present study.
| Yeast species | Number of isolates | Strain ID |
|---|---|---|
| Candida rugosa | 19 | NJ(1, 2, 5-9, 12, 18, 28-32, 36, 45, 55-57) |
| Pichia kudriavzevii | 12 | NJ(3, 4, 14, 21, 33-35, 41, 44, 50, 58, 59) |
| Trichosporon asahii | 9 | NJ(11, 15-17, 22, 38, 43, 46, 52) |
| Candida tropicalis | 6 | NJ(13, 24, 40, 47, 49, 51) |
| Magnusiomyces capitatus | 4 | NJ(19, 20) 25, 53) |
| Candida pararugosa | 4 | NJ(10, 26, 27, 54) |
| Meyerozyma caribbica | 3 | NJ(37, 39, 48) |
| Sporidiobolus pararoseus | 1 | NJ(42) |
| Yarrowia lipolytica | 1 | NJ(23) |
Table 3.
Effects of 5 different strains of yeast on rumen fermentation parameters.
| Culture time (h) | Groups | SEM | p-values | |||||
|---|---|---|---|---|---|---|---|---|
| CON | NJ-5 | NJ-12 | NJ-14 | NJ-36 | NJ-46 | |||
| pH | 5.58b | 5.65a | 5.61ab | 5.57b | 5.61ab | 5.58b | 0.05 | 0.040 |
| Total VFA, mmol/L | 100.13bc | 101.02ab | 100.82abc | 101.41a | 100.64abc | 100.06c | 0.28 | 0.003 |
| Acetate(%) | 52.61c | 53.34a | 53.23ab | 52.72bc | 52.73bc | 52.84abc | 0.16 | 0.003 |
| Propionate(%) | 27.7c | 28.37b | 28.39ab | 28.81a | 28.45ab | 28.69ab | 0.13 | 0.000 |
| Butynate(%) | 13.87 | 13.52 | 13.56 | 13.47 | 13.66 | 13.40 | 0.17 | 0.155 |
| Isobutyrate(%) | 1.62a | 1.40b | 1.38b | 1.42b | 1.45b | 1.45b | 0.05 | 0.002 |
| Valerate(%) | 2.63a | 1.98c | 1.97c | 2.08bc | 2.19b | 2.04bc | 0.06 | 0.000 |
| Isovalerate(%) | 1.56 | 1.51 | 1.51 | 1.50 | 1.52 | 1.50 | 0.04 | 0.654 |
| Acetate/Propionate ratio | 1.90a | 1.88ab | 1.88ab | 1.83c | 1.85bc | 1.84bc | 0.01 | 0.000 |
Table 3.
Effect of yeast culture on in vitro fermentation fluid pH value of goat.
| Culture time (h) | Groups | SEM | p-values | |||
|---|---|---|---|---|---|---|
| CON | LYC | MYC | HYC | |||
| 6 | 5.89b | 5.92ab | 6.07a | 6.03a | 0.06 | 0.038 |
| 12 | 5.85b | 5.84b | 5.85b | 6.00a | 0.03 | 0.001 |
| 24 | 5.69b | 5.84a | 5.82a | 5.88a | 0.05 | 0.040 |
Table 4.
Effect of yeast culture on fermentation parameters in rumen fermentation broth of goat in vitro.
Table 4.
Effect of yeast culture on fermentation parameters in rumen fermentation broth of goat in vitro.
| Culture time (h) | Groups | SEM | p-values | |||
|---|---|---|---|---|---|---|
| CON | LYC | MYC | HYC | |||
| Total VFA,mmol/L | 75.69b | 76.00ab | 76.09ab | 76.36a | 0.16 | 0.018 |
| Acetate(%) | 53.53c | 54.56b | 55.09b | 56.01a | 0.16 | 0.000 |
| Propionate(%) | 28.62a | 27.41b | 27.19b | 26.70c | 0.10 | 0.000 |
| Butynate(%) | 14.46a | 14.62a | 14.43a | 14.09b | 0.10 | 0.005 |
| Isobutyrate(%) | 1.12 | 1.08 | 1.09 | 1.04 | 0.04 | 0.214 |
| Valerate(%) | 1.20 | 1.29 | 1.21 | 1.12 | 0.05 | 0.090 |
| Isovalerate(%) | 1.05 | 1.03 | 0.99 | 1.03 | 0.04 | 0.543 |
| Acetate/Propionate ratio | 1.97 | 1.99 | 1.92 | 1.91 | 0.01 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s) . MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



