
Submitted:
25 April 2025
Posted:
27 April 2025
You are already at the latest version
Abstract
Background: Lycopene exhibits a broad spectrum of biological activities with potential therapeutic applications. Despite its established antioxidant and anti-inflammatory properties, the molecular basis for its pharmacological actions remains incompletely defined. Here we investigated the molecular targets, pharmacodynamic feasibility, and tissue-specific expression of lycopene targets using a network pharmacology approach combined with affinity and protein-protein interaction (PPI) analyses. Methods: Lycopene-associated human protein targets were predicted using Swiss target screening platform. Molecular docking was used to estimate binding affinities, and concentration-affinity (CA) ratios were calculated based on physiologically relevant plasma concentrations (75–210 nM). PPI networks of lycopene targets were constructed to identify highly connected targets, and tissue expression analysis was assessed for high-affinity targets using protein-level data from the Human Protein Atlas database. Results: Of the 94 predicted targets, 37% were nuclear receptors and 18% were Family A GPCRs. Among the top 15 high-affinity targets, nuclear receptors and GPCRs comprised 40% and 26.7% respectively. Twenty targets had affinities 1). PPI analysis revealed 32 targets with high interaction and 9 with significant network connectivity. Seven targets (TRIM24, GRIN1, NTRK1, FGFR1, NTRK3, CHRNB4, and PIK3CD) showed both high affinity and centrality in the interaction network. Expression profiling of submicromolar targets revealed widespread tissue distribution for MAP2K2 and SCN3A, while SCN2A, TOP2A, and TRIM24 showed more restricted expression patterns. Conclusion: This integrative analysis identifies a subset of lycopene targets with both high affinity and pharmacological feasibility, particularly MAP2K2, SCN2A, and TRIM24. Lycopene appears to exert its biological effects through modulation of interconnected signalling networks involving nuclear receptors, GPCRs, and ion channels. These findings support the potential of lycopene as a multi-target therapeutic agent and provide a rationale for future experimental and clinical validation.
Keywords:
lycopene
; network pharmacology
; PI3K/AKT
; MAPK
; oxidative stress
; inflammation
; molecular docking
; systems biology
1. Introduction
Lycopene, a naturally occurring lipophilic dietary carotenoid found predominantly in tomatoes and other red coloured fruits/vegetables, has been extensively studied for its health benefits. Lycopene concentrations per 100 grams for some common sources are as follows; guava: 5,204 micrograms (mcg), cooked tomatoes: 3,041 mcg, watermelon: 4,532 mcg, pink grapefruit: 1,419 mcg, papaya: 1,828 mcg, red bell peppers (cooked): 484 mcg, persimmon: 159 mcg, asparagus (cooked): 30 mcg and red cabbage: 20 mcg. It's important to note that lycopene concentrations can vary based on factors such as ripeness, variety, and growing conditions.[1,2,3] The global lycopene market is valued at approximately Euro 160 million in 2023 and is projected to grow at a compound annual growth rate (CAGR) of 5.1% from 2024 to 2030.[4,5] The average daily lycopene intake ranges from 5 to 11 mg per capita.[4,5] Epidemiological studies have associated high dietary intake of lycopene with reduced risks of chronic diseases such as cardiovascular ailments and certain cancers. These observations have spurred extensive research into the bioactive properties of lycopene, particularly its antioxidant and anti-inflammatory effects.[6,7]
Lycopene was paraphs first isolated from the berries of Tamus communis by a chemist Frederik Anthony Hartsen in 1873.[8] Subsequently, in 1875, it was extracted from tomatoes by a botanist Alexis Millardet as a crude crystalline mixture, who named it as solanorubin. It wasn't until 1903 that chemist Carl A. Schunck identified this red pigment as distinct from other carotenoids and named it "lycopine" probably deriving the term from the tomato's botanical name, Lycopersicon esculentum.[9,10] It isn’t clear how in colloquial use lycopine became lycopene, which tends to be the current nomenclature. Lycopene (PubChem ID 446925; MF: C40H56, MW: 536.9 g/mol) is a carotenoid characterized by its linear, acyclic structure composed of 40 carbon atoms forming a series of 13 double bonds, 11 of which are conjugated.[11,12] This extensive conjugated system imparts lycopene with its deep red colour and potent antioxidant properties.[11,12] This conjugated system allows lycopene to absorb light in the blue and green regions of the visible spectrum, with absorption maxima around 505 nm. As a result, the remaining unabsorbed light is rich in red wavelengths, which are reflected, giving lycopene its characteristic red colour. The number of conjugated double bonds in a molecule directly influences its colour (a greater number of these bonds shifts absorption toward longer wavelengths, resulting in the reflection of colours like red). Unlike many other carotenoids, such as β-carotene, which possess cyclic end groups (rings) at both ends of their structures, lycopene lacks these terminal rings, resulting in its open-chain configuration.[11,12,13] Additionally, lycopene does not exhibit pro-vitamin A activity, distinguishing it functionally from carotenoids like β-carotene that can be converted into vitamin A in the human body.
Lycopene exhibits distinct pharmacokinetic properties in humans. Upon ingestion, lycopene is incorporated into lipid micelles in the small intestine, facilitated by dietary fats and bile salts. Absorption occurs primarily via passive diffusion and is mediated by scavenger receptor class B type 1 (SR-B1).[14,15] Once absorbed, lycopene is transported through the lymphatic system within chylomicrons and subsequently distributed to various tissues, with notable accumulation in the liver, adrenal glands, and testes.[16,17] The compound undergoes extensive isomerization, predominantly converting from the all-trans form to cis-isomers, which are more bioavailable and preferentially accumulate in tissues.[16] Metabolism involves oxidative cleavage by enzymes such as β-carotene oxygenase 2 (BCO2), leading to the formation of polar metabolites excreted via urine. Pharmacokinetic studies have reported following single dose administration (10 to 30 mg) in healthy human adult males, a time to reach maximum plasma concentration (0.08–0.21 uM) (Tmax) ranging from 15 to 33 hours post-ingestion, with a half-life varying between 28 to 62 hours and volume of distribution varying from 250 to 800 litres, indicating prolonged retention in the body.[16,18] These parameters can be influenced by factors including the food matrix, presence of dietary fats, and individual variations in metabolism. Understanding these pharmacokinetic characteristics is essential for optimizing lycopene's therapeutic applications and determining appropriate dosing regimens.
Lycopene has gained significant attention for its broad spectrum of pharmacological activities. Its principal mechanisms of action include potent antioxidant, anti-inflammatory, and anti-proliferative effects, which collectively contribute to its therapeutic potential across a range of human diseases.[7,19,20,21,22] As a strong antioxidant, lycopene efficiently scavenges reactive oxygen species (ROS), thereby protecting cells from oxidative damage, a critical factor in the pathogenesis of cardiovascular diseases, cancers, and neurodegenerative disorders.[7,19,20] Lycopene modulates inflammatory responses by inhibiting pro-inflammatory cytokines and promoting negative feedback mechanisms that suppress chronic inflammation. This anti-inflammatory effect plays a vital role in conditions such as cardiovascular disease, where lycopene improves vascular function and reduces oxidative stress.[23,24] Neuroprotective effects have also been observed, with lycopene helping preserve cognitive function by mitigating oxidative damage and neuroinflammation in disorders like Alzheimer's and Parkinson's disease.[25,26] In cancer prevention, lycopene interferes with growth factor receptor signalling and cell cycle progression, particularly in prostate cancer cells, without inducing toxicity or apoptosis.[7,19] It also impacts pathways such as the mevalonate pathway and Ras activation, further supporting its antitumor activity.[27] In metabolic disorders, lycopene improves insulin sensitivity, lowers blood glucose levels, and protects pancreatic β-cells from oxidative stress, indicating benefits in diabetes and metabolic syndrome.[28,29] At the molecular level, lycopene interacts with a variety of human targets, modulating key signalling pathways including MAPKs, PI3K/Akt, and NF-κB.[30,31] It inhibits IκB kinase to suppress NF-κB signalling and activates PPARγ, a nuclear receptor involved in glucose and lipid metabolism.[32,33] Lycopene also influences EGFR and Protein Kinase C (PKC), affecting cell growth and gene expression.[20,34,35] Despite numerous studies highlighting the beneficial effects of lycopene, the precise molecular mechanisms and biological targets underlying its therapeutic actions remain to have considerable scientific gaps. Traditional pharmacological approaches have identified some pathways influenced by lycopene; however, a comprehensive, systems-level understanding of lycopene pharmacology is lacking. Network pharmacology, which integrates systems biology and computational tools, offers a holistic framework to elucidate the complex interactions between bioactive compounds and biological systems. By constructing and analysing networks of protein-protein interactions (PPIs), this approach can identify key molecular targets and pathways modulated by compounds like lycopene.
In this study, we employed network pharmacology methodologies to systematically explore the molecular mechanisms and biological targets of lycopene in humans. By integrating data from open-access databases, we identified lycopene-associated proteins and constructed a PPI network to discern critical nodes and pathways. Furthermore, molecular docking analyses were performed to assess the binding affinities between lycopene and its potential targets, providing insights into the compound's pharmacological interactions. This comprehensive approach aims to bridge the gap between observational studies and mechanistic insights, thereby advancing our understanding of lycopene's therapeutic potential and informing future research directions.
2. Materials and Methods
Chemical structure of lycopene (PubChem ID 446925) was identified in PubChem database and its SMILES ID (CC(=CCC/C(=C/C=C/C(=C/C=C/C(=C/C=C/C=C(/C=C/C=C(/C=C/C=C(/CCC=C(C)C)\C)\C)\C)/C)/C)/C)C) was input into the SwissTargetPrediction web-based tool (http://www.swisstargetprediction.ch/) with Homo sapiens species section to identify all potential targets of lycopene. The target class of the lycopene targets identified were assessed as percent of the total targets for all the targets and as well as top 15 targets. The lycopene targets identified were cross checked for validity in the UniPort database (https://www.uniprot.org/) and their AlphaFold (if available) 3D structure was downloaded in PDB format. For the targets where the AlphaFold was not available, homology modelling as reported before was used to generate a 3D structure.[36,37] The affinity values of lycopene to its identified targets were assessed using AutoDock vina 1.2.0 as reported before for other ligand-receptor combinations.[38,39]
To assess the targetability potential of all identified targets by lycopene, the concentration affinity ratio (CA ratio) for each of the targets was calculated as described before.[36] The following plasma concentration range of lycopene, 75
170 and 210 nM was used for the CA ratio assessment based on the lycopene dose range of 10, 30 and 90 mg/day and its volume of distribution of 520 litres.[16] The CA ratio of lycopene to all its identified targets is presented as a heat map.
To assess the protein-protein interactions between the lycopene targets, protein structure of the targets (PDB files) was imported into Chimera software in pairs and the number of hydrogen bonds (H-bond) formed between each pair of targets at 10 Armstrong (10A) distance was evaluated. The data of these interactions was inputted into the Microsoft Excel sheet and a heat map was generated using the conditional formatting colour scale tool to identify major networks. The networks of the lycopene targets were categorised into the following two group; 1) high interacting networks and 2) significantly interacting networks based on at least ten pairs of targets showing >8000 H-bond between them using the quartile logic. If a minimum of ten pairs of targets showed >15000 H-bond between them, then the target was categorised to have significantly interacting networks. A Venn diagram was plotted using the list of 1) all targets, 2) targets with high interacting networks and 3) targets with significantly interacting networks to illustrate the logical relationships between these three datasets.
All the identified targets of lycopene with affinity values <1uM iwere evaluated for their tissue specific protein expression using the Human Protein Atlas (https://www.proteinatlas.org) database.[36,40] Of the six high affinity targets we could analyse only five as protein expression data of SLC6A5 wasn’t available. The tissue specific protein expression data of the remaining five high affinity lycopene targets (MAP2K2, SCN2A, SCN3A, TOP2A and TRIM24) were plotted using Venn diagram to assess the logical relationships between the expression pattern of the targets and to identify if any specific tissue type is of preference for lycopene pharmacological effects.
3. Results
The target prediction screening of lycopene showed 94 possible targets with affinities varying from 57 nM to 14 mM (Figure 1). A significant proportion of lycopene targets were nuclear receptors (37%) followed by Family A G protein coupled receptors (GPCR 18%), while the rest of the target categories ranged from 2-8% (Figure 1). When only the top 15 targets of the lycopene were assessed, six different target class very identified, of which 40% were nuclear receptors followed by GPCR (26.7%), eraser’s (13%), phosphatases (6.7%), secreted protein (6.7%) and ligand gated ion channels (6.7%). Twenty targets of lycopene were observed to have affinities values <10uM and of these twenty targets the following targets showed affinity values <1uM; MAP2K2, SCN2A, SLC6A5, SCN3A, TOP2A and TRIM24.
To assess the targetability of the lycopene targets, we looked at concentration-affinity (CA) ratios at the following plasma concentration of lycopene 75, 170, 210 nM which was based on the dose rate of 10-90 mg/day in humans. The CA ratio analysis indicated the feasibility of targeting the following three targets MAP2K2, SCN2A and SLC6A5 as their CA ratio was >1, while the rest of the lycopene target showed CA ratio of <0.1, suggesting the affinities values of lycopene to these targets was ten folds greater than plasma concentrations of lycopene.
The protein-protein interaction (PPI) analysis of all lycopene targets indicated 32 high interaction and 9 significant interaction targets (Figure 2). Of the 32 high interaction targets 5 (TRIM24, GRIN1, NTRK1, FGFR1 and NTRK3) showed affinity values to lycopene under 10uM, while among the 9 significant interaction targets only two (CHRNB4 and PIK3CD) showed affinity values to lycopene under 10uM. Although the CA ratios of these lycopene targets with affinity values <10uM ranged from 0.01 to 0.03. Most lycopene targets in addition to showing low affinity values had negligible PPI (<10 hydrogen bonds), suggesting these targets are not of relevance to the pharmacological effects of lycopene or may be indirectly regulated. Based on the PPI and affinity values we observe the three categories of lycopene targets, i.e., 1) targets with high affinity values (<10uM; MAP2K2, SCN2A, SLC6A5, SCN3A, TOP2A, TRIM24, P2RY12, SLC6A1, AURKB, SCN4A, GRIN1, NTRK1, PDGFRA, RXFP1, CHRNB4, PIK3CA, FGFR1, ABCC1, NTRK3 and PIK3CD), 2) targets with high network interactions (TRIM24, GRIN1, NTRK1, CHRNB4, FGFR1, NTRK3, PIK3CD, CLK4, RXRG, CYP2A6, GRK5, RXRA, RXRB, CLK1, NFKB1, MAPK1, PDGFRB, PIK3R1, CXCR4, GLRA1, NR1I2, NR3C2, NFE2L2, NAMPT, PRMT1, METAP2, NR4A1, ADAM10, VDR, HSD17B10, SPHK1 and APEX1) and 3) targets with both high affinity values and network interactions (TRIM24, GRIN1, NTRK1, FGFR1, NTRK3, CHRNB4 and PIK3CD) (Figure 2).
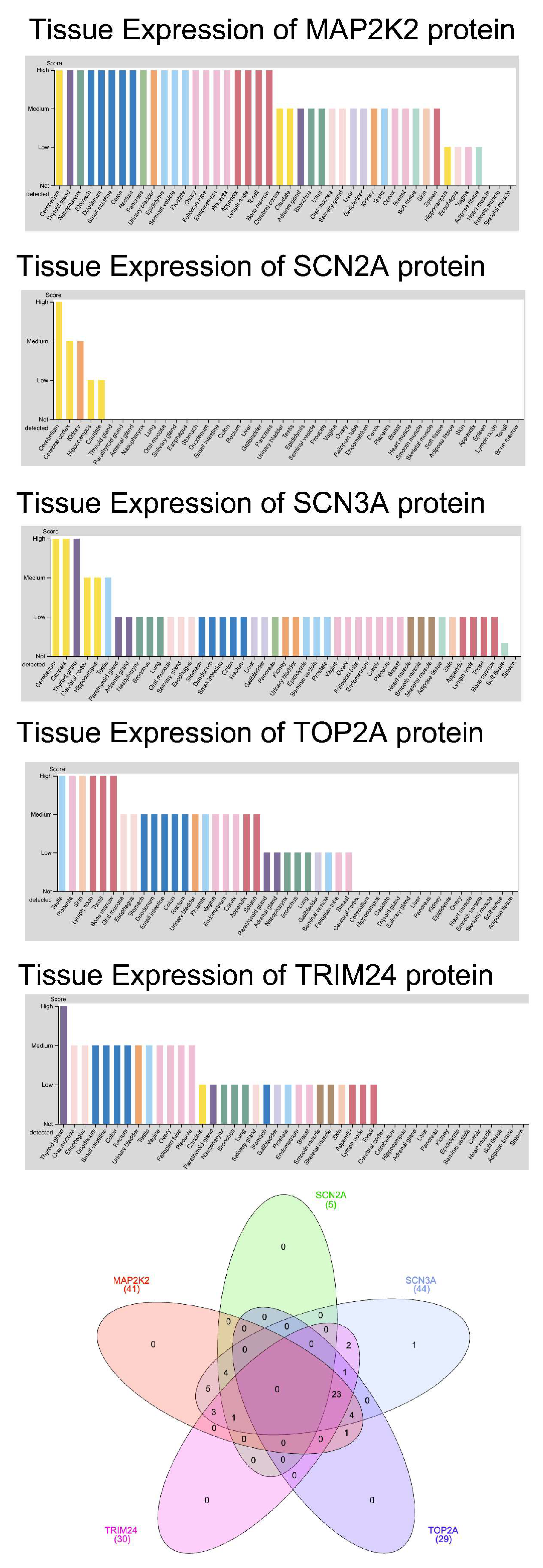
For the expression analysis of the lycopene targets in human tissues, we only looked at high affinity targets with affinities values <1uM as these targets are most likely to be pharmacodynamically preferred. Hence the following targets MAP2K2, SCN2A, SLC6A5, SCN3A, TOP2A and TRIM24 were assessed for their tissue specific expression pattern. As we selectively looked at protein expression pattern, SLC6A5 was excluded as its protein expression data is still pending normal tissue annotation. Moreover, its RNA expression data is limited to the following tissues, lung, spinal cord, cerebellum, pituitary gland, testis and cerebral cortex in humans with the number of transcripts per million (nTPM) value <1. The remaining 5 lycopene targets, except SCN2A (expressed in 5 tissue types) showed extensive tissue expression with MAP2K2 and SCN3A being widely expressed. MAP2K2, SCN3A, TOP2A and TRIM24 were collectively expressed in 23 different tissues. MAP2K2, SCN3A, TOP2A and TRIM24 were all expressed in seminal vesicle, bone marrow, adrenal gland and cervix. SCN3A, TOP2A and TRIM24 were collectively expressed in parathyroid gland only. SCN3A and TRIM24 were collectively expressed in smooth and skeletal muscles. MAP2K2 and TOP2A were collectively expressed in spleen only. MAP2K2, SCN2A, SCN3A and TRIM24 were collectively expressed in caudate nucleus only. MAP2K2, SCN3A and TRIM24 were collectively expressed in thyroid gland, ovary and salivary gland. MAP2K2 and SCN3A were collectively expressed in pancreas, epididymis, liver, soft and adipose tissue. MAP2K2, SCN2A and SCN3A were collectively expressed in cerebellum, cerebral cortex, kidney and hippocampus. SCN3A was exclusively expressed in heart muscle. High expression of MAP2K2 protein was observed in 21 different tissues, while high expression of SCN2A (cerebellum), SCN3A (cerebellum, caudate nucleus and thyroid gland), TOP2A (testis, placenta, skin, lymph node, tonsil and bone marrow) and TRIM24 (thyroid gland) was observed only in one or few tissues.
4. Discussion
This study provides a comprehensive network pharmacology analysis of lycopene, revealing its multi-target interactions and potential pharmacodynamic actions in human tissues. The in-silico target prediction identified 94 candidate targets, with affinity values ranging from 57 nM to 14 mM, highlighting the broad spectrum of lycopene's molecular mechanisms.[20,30,41,42] This is particularly not surprising considering the broad spectrum of health benefits reported following lycopene use.[16,21,43,44] Notably, nuclear receptors constituted the largest proportion (37%) of target classes, consistent with previous findings that carotenoids often modulate gene expression via nuclear receptor pathways.[45,46,47] GPCRs represented the second-largest category (18%), indicating lycopene's potential to modulate signalling at the membrane receptor level as well. Focusing on the top 15 targets of lycopene, six distinct target classes were identified, dominated again by nuclear receptors (40%) and GPCRs (26.7%), which is again consistent with prior reports indicating lycopene’s ability to modulate transcriptional and signalling pathways involved in inflammation, metabolism, and cellular proliferation.[6,34,41]
Among the 20 targets with sub-10 µM affinities, six targets (MAP2K2, SCN2A, SLC6A5, SCN3A, TOP2A, and TRIM24) exhibited high-affinity binding (<1 µM), suggesting these targets are most likely to be pharmacologically modulated at physiological concentrations of lycopene. While lycopene is known for its weak direct activity at most proteins due to its hydrophobic nature and limited aqueous solubility,[48,49] our concentration-affinity (CA) ratio analysis provided critical pharmacodynamic context. Only three targets (MAP2K2, SCN2A, and SLC6A5) showed CA ratios >1 at reported human plasma concentrations (75–210 nM), suggesting these are the most pharmacologically viable targets under physiological conditions. While compounds with CA ratios >1 are more likely to exert biological effects at attainable plasma levels, considering previous studies indicating that lycopene exerts biological activity at nanomolar concentrations,[50,51,52] particularly within membrane-rich environments or lipid rafts where its accumulation is higher,[53,54] it is likely that other lycopene targets with CA ratios <1 can potentially be involved in its pharmacodynamic effects.
Protein–protein interaction network analysis further identified a subset of targets with significant biological relevance. We identified 32 high-interaction and 9 significant-interaction proteins, with some of these targets (TRIM24, GRIN1, NTRK1, FGFR1, and NTRK3) exhibiting binding affinities below 10 μM. Although their CA ratios remained low (<0.03), these targets may act as secondary effectors or be relevant in tissues with higher lycopene accumulation, such as the prostate, liver, and adrenal glands.[16,55] TRIM24, GRIN1, NTRK1, FGFR1, NTRK3, CHRNB4, and PIK3CD are key proteins with diverse biological roles, many of which are relevant to disease pathogenesis and potential pharmacological targeting.[56,57,58] TRIM24 is a transcriptional co-regulator that influences nuclear receptor signalling and chromatin remodelling, playing critical roles in cell proliferation and apoptosis and hence it is implicated in cancer, particularly breast and liver malignancies.[59,60] GRIN1 encodes a core subunit of NMDA receptors, essential for calcium ion influx during synaptic transmission. It is central to learning, memory, and neural development, and its dysregulation is associated with neurodegenerative diseases due to excitotoxicity.[61,62] Alterations in NTRK1 is linked to various cancers and sensory disorders, while NTRK3 is associated with breast carcinoma and congenital fibrosarcoma.[63,64] FGFR1, by mediating pathways involved in growth, angiogenesis, and tissue repair contributes to developmental syndromes and several cancers while CHRNB4 is critical for synaptic transmission in the nervous system and is implicated in neurological conditions such as epilepsy, schizophrenia, and nicotine dependence.[65,66,67] PIK3CD is predominantly expressed in immune cells and regulates pathways central to immune function, and its mutations are associated with primary immunodeficiencies like Activated PI3K-Delta Syndrome.[68] Hence PIK3CD is a target of interest in autoimmunity and cancer therapy. These novel observations in our study on the diverse pathophysiological role of lycopene targets identified is consistent with several clinical investigations and reports on potential use of lycopene use in healthcare.[21,69,70]
Expression profiling of the highest affinity targets provided further insight into tissue-specific pharmacodynamics of lycopene. Except for SLC6A5, whose protein expression data remains limited, the remaining high-affinity targets showed expression in a range of tissues. MAP2K2, SCN3A, TOP2A, and TRIM24 are functionally involved in oncogenic signalling, neuronal excitability, and DNA repair, offering diverse but interconnected avenues for pharmacological intervention. Particularly, MAP2K2, SCN3A, TOP2A, and TRIM24 were co-expressed in reproductive and endocrine tissues (seminal vesicle, adrenal gland, and cervix). These results are in line with past transcriptomic studies that observed upregulation of lycopene-responsive genes in prostate and mammary tissues.[54,71,72] MAP2K2 is a critical component of the MAPK/ERK signalling pathway, which regulates cell proliferation, differentiation, and survival. Aberrant activation of MAP2K2 has been linked to various cancers, particularly melanoma and colorectal cancer, making it a validated target for anti-cancer therapies.[73,74] Pharmacological inhibition of MAP2K2 has shown potential in overcoming resistance to upstream oncogenes such as BRAF and RAS and it remains to be tested if lycopene can have synergistic effects with these established MAP2K2 inhibitors to achieve optimal pharmacodynamic outcomes while minimising any potential adverse effects. Interestingly, SCN3A, a voltage-gated sodium channel, showed unique expression in the heart, pointing to possible roles in cardiac excitability supporting prior studies that suggested cardiovascular benefits of lycopene via electrophysiological and anti-inflammatory mechanisms.[75] SCN3A is predominantly expressed in the central nervous system, where it is essential for the initiation and propagation of action potentials in neurons. Dysregulation or mutations in SCN3A are associated with neurological disorders including epilepsy, developmental and epileptic encephalopathies, and cognitive impairments. SCN3A represents a target for anticonvulsant development and therapies aimed at modulating neuronal excitability.[76,77] Based on these there seems to be merit in assessing the potential of lycopene in connecting the heart-brain axis via SCN3A targeting. TOP2A, and TRIM24 have a vital role in DNA replication, transcription, and chromosome segregation and are well-established targets in cancer chemotherapy especially in breast, prostate, and liver cancers.[78,79] Our report of high affinity targeting of TOP2A, and TRIM24 opens avenues for synergistically improving the pharmacodynamics of currently used chemotherapeutics (etoposide and doxorubicin) with lycopene.
Collectively, our findings suggest MAP2K2 and SCN2A as the most pharmacodynamically plausible direct targets of lycopene in humans. Their strong affinity, favourable CA ratios, and wide tissue expression profiles make these targets to have a foundational role in mediating lycopene’s pharmacological actions. Additional targets such as TRIM24, TOP2A, and NTRK family members, while displaying lower CA ratios, may contribute to lycopene’s broader systems-level impact, particularly in the context of inflammation, cancer biology, and neuroprotection. These multi-target interactions are consistent with lycopene’s observed pleiotropic benefits in clinical and preclinical models.[7,30,36,44,80]
In conclusion, our study advances the mechanistic understanding of lycopene pharmacology by integrating affinity profiling, network analysis, and expression mapping. While the hydrophobicity and relatively low systemic concentrations of lycopene pose challenges for classical drug-likeness, its selective interaction with key signalling and nuclear proteins highlights its potential as a nutraceutical agent with systems-level effects which merits advancing its use for therapeutic applications. Future studies should aim to validate these interactions in functional cellular assays and assess tissue-specific accumulation patterns to better delineate lycopene’s therapeutic potential.
Acknowledgments
Research support from University College Dublin-Seed funding/Output Based Research Support Scheme (R19862, 2019), Royal Society-UK (IES\R2\181067, 2018) and Stemcology (STGY2917, 2022) is acknowledged.
Declaration of Interest Statement
none.
References
- Giovannucci, E.; Rimm, E.B.; Liu, Y.; Stampfer, M.J.; Willett, W.C. A Prospective Study of Tomato Products, Lycopene, and. Journal of the National Cancer Institute 2002, 94, 391. [Google Scholar] [CrossRef]
- Rao, A.; Waseem, Z.; Agarwal, S. Lycopene content of tomatoes and tomato products and their contribution to dietary lycopene. Food Research International 1998, 31, 737–741. [Google Scholar] [CrossRef]
- Gerster, H. The potential role of lycopene for human health. Journal of the American College of Nutrition 1997, 16, 109–126. [Google Scholar] [CrossRef]
- Gupta, E.; Mishra, P.; Singh, P.; Mishra, N. Evidence and prospects of lycopene as powerful red superfood: modern approach to food science. Current Traditional Medicine 2024, 10, 50–60. [Google Scholar] [CrossRef]
- Yaqoob, S.; Riaz, M.; Shabbir, A.; Zia-Ul-Haq, M.; Alwakeel, S.S.; Bin-Jumah, M. Commercialization and marketing potential of carotenoids. Carotenoids: structure and function in the human body, 2021; 799–826. [Google Scholar]
- Kulawik, A.; Cielecka-Piontek, J.; Czerny, B.; Kamiński, A.; Zalewski, P. The Relationship Between Lycopene and Metabolic Diseases. Nutrients 2024, 16, 3708. [Google Scholar] [CrossRef]
- Przybylska, S.; Tokarczyk, G. Lycopene in the prevention of cardiovascular diseases. International journal of molecular sciences 2022, 23, 1957. [Google Scholar] [CrossRef]
- Hartsen, F. Beitrage zur Organischen Chemie. Chem. Zentra 1873, 204–207. [Google Scholar]
- Camara Hurtado, R.M.; Cáceres, J.O.; Cámara Hurtado, M.; Manzoor, S.; Fernández Ruiz, V.; Sánchez Mata, M.C. New developments in lycopene analysis by spectroscopic and chromatographic techniques, accompanied by mathematical modelling. 2014.
- Bergougnoux, V. The history of tomato: from domestication to biopharming. Biotechnology advances 2014, 32, 170–189. [Google Scholar] [CrossRef]
- Shi, J.; Maguer, M.L. Lycopene in tomatoes: chemical and physical properties affected by food processing. Critical reviews in food science and nutrition 2000, 40, 1–42. [Google Scholar] [CrossRef]
- Clinton, S.K. Lycopene: chemistry, biology, and implications for human health and disease. Nutrition reviews 1998, 56, 35–51. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, A.K. Lycopene; chemistry, biosynthesis, metabolism and degradation under various abiotic parameters. Journal of Food Science and Technology 2015, 52, 41–53. [Google Scholar] [CrossRef]
- Valacchi, G.; Pecorelli, A. Role of Scavenger Receptor B1 (SR-B1) in Improving Food Benefits for Human Health. Annual Review of Food Science and Technology 2025, 16. [Google Scholar] [CrossRef]
- Moussa, M.; Landrier, J.-F.; Reboul, E.; Ghiringhelli, O.; Coméra, C.; Collet, X.; Fröhlich, K.; Böhm, V.; Borel, P. Lycopene absorption in human intestinal cells and in mice involves scavenger receptor class B type I but not Niemann-Pick C1-like 1. The Journal of nutrition 2008, 138, 1432–1436. [Google Scholar] [CrossRef]
- Arballo, J.; Amengual, J.; Erdman Jr, J.W. Lycopene: A critical review of digestion, absorption, metabolism, and excretion. Antioxidants 2021, 10, 342. [Google Scholar] [CrossRef]
- Botham, K.M.; Bravo, E. Lycopene and chylomicrons. In Lycopene; CRC Press: 2019; pp. 159-182.
- Gustin, D.M.; Rodvold, K.A.; Sosman, J.A.; Diwadkar-Navsariwala, V.; Stacewicz-Sapuntzakis, M.; Viana, M.; Crowell, J.A.; Murray, J.; Tiller, P.; Bowen, P.E. Single-dose pharmacokinetic study of lycopene delivered in a well-defined food-based lycopene delivery system (tomato paste-oil mixture) in healthy adult male subjects. Cancer Epidemiology Biomarkers & Prevention 2004, 13, 850–860. [Google Scholar]
- Khurana, S.; Sharma, S.; Goyal, P.K. Updates on the Anticancer Profile of Lycopene and its Probable Mechanism Against Breast and Gynecological Cancer. The Natural Products Journal 2025. [Google Scholar] [CrossRef]
- Ozkan, G.; Günal-Köroğlu, D.; Karadag, A.; Capanoglu, E.; Cardoso, S.M.; Al-Omari, B.; Calina, D.; Sharifi-Rad, J.; Cho, W.C. A mechanistic updated overview on lycopene as potential anticancer agent. Biomedicine & Pharmacotherapy 2023, 161, 114428. [Google Scholar]
- Khan, U.M.; Sevindik, M.; Zarrabi, A.; Nami, M.; Ozdemir, B.; Kaplan, D.N.; Selamoglu, Z.; Hasan, M.; Kumar, M.; Alshehri, M.M. Lycopene: food sources, biological activities, and human health benefits. Oxidative medicine and cellular longevity 2021, 2021, 2713511. [Google Scholar] [CrossRef]
- Heber, D.; Lu, Q.-Y. Overview of mechanisms of action of lycopene. Experimental biology and medicine 2002, 227, 920–923. [Google Scholar] [CrossRef]
- Mozos, I.; Stoian, D.; Caraba, A.; Malainer, C.; Horbańczuk, J.O.; Atanasov, A.G. Lycopene and vascular health. Frontiers in pharmacology 2018, 9, 521. [Google Scholar] [CrossRef]
- Lee, W.; Ku, S.-K.; Bae, J.W.; Bae, J.-S. Inhibitory effects of lycopene on HMGB1-mediated pro-inflammatory responses in both cellular and animal models. Food and Chemical Toxicology 2012, 50, 1826–1833. [Google Scholar] [CrossRef]
- Prema, A.; Janakiraman, U.; Manivasagam, T.; Thenmozhi, A.J. Neuroprotective effect of lycopene against MPTP induced experimental Parkinson’s disease in mice. Neuroscience letters 2015, 599, 12–19. [Google Scholar] [CrossRef]
- Kaur, H.; Chauhan, S.; Sandhir, R. Protective effect of lycopene on oxidative stress and cognitive decline in rotenone induced model of Parkinson’s disease. Neurochemical research 2011, 36, 1435–1443. [Google Scholar] [CrossRef]
- Palozza, P.; Simone, R.; Catalano, A.; Parrone, N.; Monego, G.; Ranelletti, F.O. Lycopene regulation of cholesterol synthesis and efflux in human macrophages. The Journal of nutritional biochemistry 2011, 22, 971–978. [Google Scholar] [CrossRef]
- Özmen, Ö.; Şahi̇n, M.; Topsakal, Ş.; Taşan, Ş.; Şahi̇n, U. Lycopene ameliorates diabetes-induced pancreatic, hepatic, and renal damage by modulating the JAK/STAT/SOCS signaling pathway in rats. Iranian Journal of Basic Medical Sciences 2025, 28, 461. [Google Scholar]
- Liu, Y.; Tian, Y.; Dai, X.; Liu, T.; Zhang, Y.; Wang, S.; Shi, H.; Yin, J.; Xu, T.; Zhu, R. Lycopene ameliorates islet function and down-regulates the TLR4/MyD88/NF-κB pathway in diabetic mice and Min6 cells. Food & Function 2023, 14, 5090–5104. [Google Scholar]
- Tripathi, A.K.; Das, R.; Ray, A.K.; Mishra, S.K.; Anand, S. Recent insights on pharmacological potential of lycopene and its nanoformulations: an emerging paradigm towards improvement of human health. Phytochemistry Reviews 2024, 1–28. [Google Scholar] [CrossRef]
- Fang, Y.; Ou, S.; Wu, T.; Zhou, L.; Tang, H.; Jiang, M.; Xu, J.; Guo, K. Lycopene alleviates oxidative stress via the PI3K/Akt/Nrf2pathway in a cell model of Alzheimer’s disease. PeerJ 2020, 8, e9308. [Google Scholar] [CrossRef]
- Assar, E.A.; Vidalle, M.C.; Chopra, M.; Hafizi, S. Lycopene acts through inhibition of IκB kinase to suppress NF-κB signaling in human prostate and breast cancer cells. Tumor Biology 2016, 37, 9375–9385. [Google Scholar] [CrossRef]
- Simone, R.E.; Russo, M.; Catalano, A.; Monego, G.; Froehlich, K.; Boehm, V.; Palozza, P. Lycopene inhibits NF-kB-mediated IL-8 expression and changes redox and PPARγ signalling in cigarette smoke–stimulated macrophages. PloS one 2011, 6, e19652. [Google Scholar] [CrossRef]
- Marzocco, S.; Singla, R.K.; Capasso, A. Multifaceted effects of lycopene: a boulevard to the multitarget-based treatment for cancer. Molecules 2021, 26, 5333. [Google Scholar] [CrossRef]
- Han, H.; Lim, J.W.; Kim, H. Lycopene inhibits activation of epidermal growth factor receptor and expression of cyclooxygenase-2 in gastric cancer cells. Nutrients 2019, 11, 2113. [Google Scholar] [CrossRef]
- Kumar, A.H. Network Proteins of Human Sortilin1, Its Expression and Targetability Using Lycopene. Life 2024, 14, 137. [Google Scholar] [CrossRef]
- Khosravi, Z.; Kumar, A.H. Analysing the role of SERPINE1 network in the pathogenesis of human glioblastoma. Journal of Cancer Research 2023, 1, 1–6. [Google Scholar]
- Friend, J.; Kumar, A.H.S. A Network Pharmacology Approach to Assess the Comparative Pharmacodynamics of Pharmaceutical Excipient Trehalose in Human, Mouse, and Rat. Nature Cell Sci 2023, 1, 33–43. [Google Scholar] [CrossRef]
- Kumar, A.H. Pharmacological targets of Asundexian relevant to its therapeutic efficacy in treating cardiovascular diseases. Biology, Engineering, Medicine and Science Reports 2022, 8, 24–27. [Google Scholar] [CrossRef]
- Singh NK; Dhanasekaran M; AHS. K. Pharmacology of Berberine and Its Metabolites, Is It the Natures Ozempic or Imatinib? Arch Pharmacol Ther 2023, 5, 67–81. [Google Scholar] [CrossRef]
- Landrier, J.-F.; Breniere, T.; Sani, L.; Desmarchelier, C.; Mounien, L.; Borel, P. Effect of tomato, tomato-derived products and lycopene on metabolic inflammation: from epidemiological data to molecular mechanisms. Nutrition Research Reviews 2023, 1–17. [Google Scholar]
- Puah, B.-P.; Jalil, J.; Attiq, A.; Kamisah, Y. New insights into molecular mechanism behind anti-cancer activities of lycopene. Molecules 2021, 26, 3888. [Google Scholar] [CrossRef] [PubMed]
- Tufail, T.; Bader Ul Ain, H.; Noreen, S.; Ikram, A.; Arshad, M.T.; Abdullahi, M.A. Nutritional benefits of lycopene and beta-carotene: A comprehensive overview. Food Science & Nutrition 2024, 12, 8715–8741. [Google Scholar]
- Shafe, M.O.; Gumede, N.M.; Nyakudya, T.T.; Chivandi, E. Lycopene: A Potent Antioxidant with Multiple Health Benefits. Journal of Nutrition and Metabolism 2024, 2024, 6252426. [Google Scholar]
- Wu, S.; Chen, R.; Chen, J.; Yang, N.; Li, K.; Zhang, Z.; Zhang, R. Study of the anti-inflammatory mechanism of β-carotene based on network pharmacology. Molecules 2023, 28, 7540. [Google Scholar] [CrossRef]
- Mukherjee, S.; Yun, J.W. β-Carotene stimulates browning of 3T3-L1 white adipocytes by enhancing thermogenesis via the β3-AR/p38 MAPK/SIRT signaling pathway. Phytomedicine 2022, 96, 153857. [Google Scholar] [CrossRef]
- Moran, N.E.; Thomas-Ahner, J.M.; Smith, J.W.; Silva, C.; Hason, N.A.; Erdman Jr, J.W.; Clinton, S.K. β-Carotene oxygenase 2 genotype modulates the impact of dietary lycopene on gene expression during early TRAMP prostate carcinogenesis. The Journal of nutrition 2022, 152, 950–960. [Google Scholar] [CrossRef]
- Tan, H.; Wang, F.; Hu, J.; Duan, X.; Bai, W.; Wang, X.; Wang, B.; Su, Y.; Hu, J. Inhibitory interaction of flavonoids with organic cation transporter 2 and their structure–activity relationships for predicting nephroprotective effects. Journal of Applied Toxicology 2023, 43, 1421–1435. [Google Scholar] [CrossRef]
- Mittal, S.; Sawarkar, S.; Doshi, G.; Pimple, P.; Shah, J.; Bana, T. Pharmacokinetics and bioavailability of nutraceuticals. Industrial Application of Functional Foods, Ingredients and Nutraceuticals, 2023; 725–783. [Google Scholar]
- Neves-Silva, S.; Xavier-de-Britto, I.; Gomes-da-Silva, N.C.; França, Á.R.S.; Pedrochi, F.; Queiroz, M.N.; Moura-Silva, J.; Majerowicz, D.; Ricci-Junior, E.; Ferreira, T.P.T. Enhancing therapeutic efficacy: In vivo mechanisms and biochemical effects of lycopene encapsulated in nanomicelles for acute inflammation and lipid metabolism. European Journal of Pharmaceutics and Biopharmaceutics 2025, 207, 114585. [Google Scholar]
- Báo, S.N.; Machado, M.; Da Silva, A.L.; Melo, A.; Cunha, S.; Sousa, S.S.; Malheiro, A.R.; Fernandes, R.; Leite, J.R.S.; Vasconcelos, A.G. Potential biological properties of lycopene in a self-emulsifying drug delivery system. Molecules 2023, 28, 1219. [Google Scholar] [CrossRef]
- Binsuwaidan, R.; Sultan, A.A.; Negm, W.A.; Attallah, N.G.; Alqahtani, M.J.; Hussein, I.A.; Shaldam, M.A.; El-Sherbeni, S.A.; Elekhnawy, E. Bilosomes as nanoplatform for oral delivery and modulated in vivo antimicrobial activity of lycopene. Pharmaceuticals 2022, 15, 1043. [Google Scholar] [CrossRef]
- Guo, Z.; Liu, Y.; Luo, Y. Mechanisms of carotenoid intestinal absorption and the regulation of dietary lipids: lipid transporter-mediated transintestinal epithelial pathways. Critical reviews in food science and nutrition 2024, 64, 1791–1816. [Google Scholar] [PubMed]
- Li, Y.; Zhan, M.; Li, J.; Zhang, W.; Shang, X. Lycopene alleviates lipopolysaccharide-induced testicular injury in rats by activating the PPAR signaling pathway to integrate lipid metabolism and the inflammatory response. Translational Andrology and Urology 2023, 12, 271. [Google Scholar] [PubMed]
- Georgiadou, E.C.; Antoniou, C.; Majak, I.; Goulas, V.; Filippou, P.; Smolińska, B.; Leszczyńska, J.; Fotopoulos, V. Tissue-specific elucidation of lycopene metabolism in commercial tomato fruit cultivars during ripening. Scientia Horticulturae 2021, 284, 110144. [Google Scholar] [CrossRef]
- Borrageiro, G.; Haylett, W.; Seedat, S.; Kuivaniemi, H.; Bardien, S. A review of genome-wide transcriptomics studies in Parkinson's disease. European Journal of Neuroscience 2018, 47, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Shi, E.; Chmielecki, J.; Tang, C.-M.; Wang, K.; Heinrich, M.C.; Kang, G.; Corless, C.L.; Hong, D.; Fero, K.E.; Murphy, J.D. FGFR1 and NTRK3 actionable alterations in “Wild-Type” gastrointestinal stromal tumors. Journal of translational medicine 2016, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Amatu, A.; Sartore-Bianchi, A.; Siena, S. NTRK gene fusions as novel targets of cancer therapy across multiple tumour types. ESMO open 2016, 1, e000023. [Google Scholar] [CrossRef]
- Appikonda, S.; Thakkar, K.N.; Barton, M.C. Regulation of gene expression in human cancers by TRIM24. Drug Discovery Today: Technologies 2016, 19, 57–63. [Google Scholar] [CrossRef]
- Chambon, M.; Orsetti, B.; Berthe, M.-L.; Bascoul-Mollevi, C.; Rodriguez, C.; Duong, V.; Gleizes, M.; Thénot, S.; Bibeau, F.; Theillet, C. Prognostic significance of TRIM24/TIF-1α gene expression in breast cancer. The American journal of pathology 2011, 178, 1461–1469. [Google Scholar] [CrossRef]
- Venkatesan, S.; Binko, M.A.; Mielnik, C.A.; Ramsey, A.J.; Lambe, E.K. Deficits in integrative NMDA receptors caused by Grin1 disruption can be rescued in adulthood. Neuropsychopharmacology 2023, 48, 1742–1751. [Google Scholar] [CrossRef]
- Xu, Y.; Song, R.; Chen, W.; Strong, K.; Shrey, D.; Gedela, S.; Traynelis, S.F.; Zhang, G.; Yuan, H. Recurrent seizure-related GRIN1 variant: molecular mechanism and targeted therapy. Annals of clinical and translational neurology 2021, 8, 1480–1494. [Google Scholar] [CrossRef]
- Mohammadi, R.; Ghiasi, M.; Mehdizadeh, S.; Mohammadi, J.; Mohammad Ganji, S. A comprehensive bioinformatic evaluation of the NTRK family’s potential as prognostic biomarkers in breast cancer. Bioinformatics Advances 2025, 5, vbaf030. [Google Scholar] [CrossRef]
- Stravodimou, A.; Voutsadakis, I.A. Neurotrophic receptor tyrosine kinase family members in secretory and non-secretory breast carcinomas. World journal of clinical oncology 2022, 13, 135. [Google Scholar] [CrossRef]
- Becchetti, A.; Grandi, L.C.; Cerina, M.; Amadeo, A. Nicotinic acetylcholine receptors and epilepsy. Pharmacological Research 2023, 189, 106698. [Google Scholar] [CrossRef]
- Kanda, M.; Shimizu, D.; Nakamura, S.; Sawaki, K.; Umeda, S.; Miwa, T.; Tanaka, H.; Inokawa, Y.; Hattori, N.; Hayashi, M. Blockade of CHRNB2 signaling with a therapeutic monoclonal antibody attenuates the aggressiveness of gastric cancer cells. Oncogene 2021, 40, 5495–5504. [Google Scholar] [CrossRef] [PubMed]
- Labate, A.; Mumoli, L.; Fratto, A.; Quattrone, A.; Gambardella, A. Hippocampal sclerosis worsens autosomal dominant nocturnal frontal lobe epilepsy (ADNFLE) phenotype related to CHRNB2 mutation. European Journal of Neurology 2013, 20, 591–593. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Zhang, H.; Cheng, H.; Wen, J.; Li, D. PIK3CD correlates with prognosis, epithelial–mesenchymal transition and tumor immune infiltration in breast carcinoma. Discover Oncology 2023, 14, 187. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.-R.; Lin, Q.-Y.; Adu, M.; Huang, H.-L.; Yan, Z.-H.; Shao, F.; Zhong, G.-Y.; Zhang, Z.-L.; Sang, Z.-P.; Cao, L. The sources, properties, extraction, biosynthesis, pharmacology, and application of lycopene. Food & Function 2023, 14, 9974–9998. [Google Scholar]
- Yeung, A.W.K.; Choudhary, N.; Tewari, D.; El-Demerdash, A.; Tomczyk, M.; Das, N.; Pirgozliev, V.; Lucarini, M.; Durazzo, A.; Souto, E.B. Lycopene: total-scale literature landscape analysis of a valuable nutraceutical with numerous potential applications in the promotion of human and animal health. Animal Science Papers and Reports 2022, 40, 119–134. [Google Scholar]
- Tang, J.; Li, Y.; Liu, Z.; Wei, M.; Shi, Q.; Yang, F. Integrated transcriptomics and metabolomics analyses reveal the molecular mechanisms of red-light on carotenoids biosynthesis in tomato fruit. Food Quality and Safety 2022, 6, fyac009. [Google Scholar] [CrossRef]
- Sun, S.; Meng, Q.; Bai, Y.; Cao, C.; Li, J.; Cheng, B.; Shi, B.; Shan, A. Lycopene improves maternal reproductive performance by modulating milk composition and placental antioxidative and immune status. Food & Function 2021, 12, 12448–12467. [Google Scholar]
- Fernandes, M.; Barcelos, D.; Carapeto, F.C.L.; Cardili, L.; Comodo, A.N.; Mazloum, S.F.; Marins, M.M.; Mendes, A.R.; Pesquero, J.B.; Landman, G. Evaluation of Heterogeneity in the Coding Region of BRAF, MAP2K1, and MAP2K2 Genes in Primary and Metastatic Melanomas. Journal of Cutaneous Pathology 2024. [Google Scholar] [CrossRef]
- Mizuno, S.; Ikegami, M.; Koyama, T.; Sunami, K.; Ogata, D.; Kage, H.; Yanagaki, M.; Ikeuchi, H.; Ueno, T.; Tanikawa, M. High-throughput functional evaluation of MAP2K1 variants in cancer. Molecular Cancer Therapeutics 2023, 22, 227–239. [Google Scholar] [CrossRef]
- Tzialla, C.; Arossa, A.; Mannarino, S.; Orcesi, S.; Veggiotti, P.; Fiandrino, G.; Zuffardi, O.; Errichiello, E. SCN2A and arrhythmia: A potential correlation? A case report and literature review. European Journal of Medical Genetics 2022, 65, 104639. [Google Scholar] [CrossRef] [PubMed]
- Asadollahi, R.; Delvendahl, I.; Muff, R.; Tan, G.; Rodríguez, D.G.; Turan, S.; Russo, M.; Oneda, B.; Joset, P.; Boonsawat, P. Pathogenic SCN2A variants cause early-stage dysfunction in patient-derived neurons. Human Molecular Genetics 2023, 32, 2192–2204. [Google Scholar] [CrossRef] [PubMed]
- Brunklaus, A.; Brünger, T.; Feng, T.; Fons, C.; Lehikoinen, A.; Panagiotakaki, E.; Vintan, M.-A.; Symonds, J.; Andrew, J.; Arzimanoglou, A. The gain of function SCN1A disorder spectrum: novel epilepsy phenotypes and therapeutic implications. Brain 2022, 145, 3816–3831. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.; Pour Abbasi, M.; Khorrami, S.; Khodamoradi, S.; Mohammadi Goldar, Z.; Ebrahimzadeh, F. The TRIM proteins in cancer: from expression to emerging regulatory mechanisms. Clinical and Translational Oncology 2022, 1–11. [Google Scholar] [CrossRef]
- Boyson, S.P.; Gao, C.; Quinn, K.; Boyd, J.; Paculova, H.; Frietze, S.; Glass, K.C. Functional roles of bromodomain proteins in cancer. Cancers 2021, 13, 3606. [Google Scholar] [CrossRef]
- Kulawik, A.; Cielecka-Piontek, J.; Zalewski, P. The importance of antioxidant activity for the health-promoting effect of lycopene. Nutrients 2023, 15, 3821. [Google Scholar] [CrossRef]
Figure 1.
The chemical structure of lycopene to outline it linear nature is shown. The target class of all and top 15 lycopene targets are shown as pie chart along with the relative percent of each of the target categories. The heat map shows the affinity and concentration-affinity (CA) ratio values (for plasma concentration of lycopene of 75, 170 and 210 nM) for each of the lycopene targets. The lycopene targets are represented by their UniProt ID. The lycopene targets highlighted in green are high affinity targets all will affinity values <10uM. The heat map scale shown is from high (red) to low (green) CA ratio.
Figure 1.
The chemical structure of lycopene to outline it linear nature is shown. The target class of all and top 15 lycopene targets are shown as pie chart along with the relative percent of each of the target categories. The heat map shows the affinity and concentration-affinity (CA) ratio values (for plasma concentration of lycopene of 75, 170 and 210 nM) for each of the lycopene targets. The lycopene targets are represented by their UniProt ID. The lycopene targets highlighted in green are high affinity targets all will affinity values <10uM. The heat map scale shown is from high (red) to low (green) CA ratio.
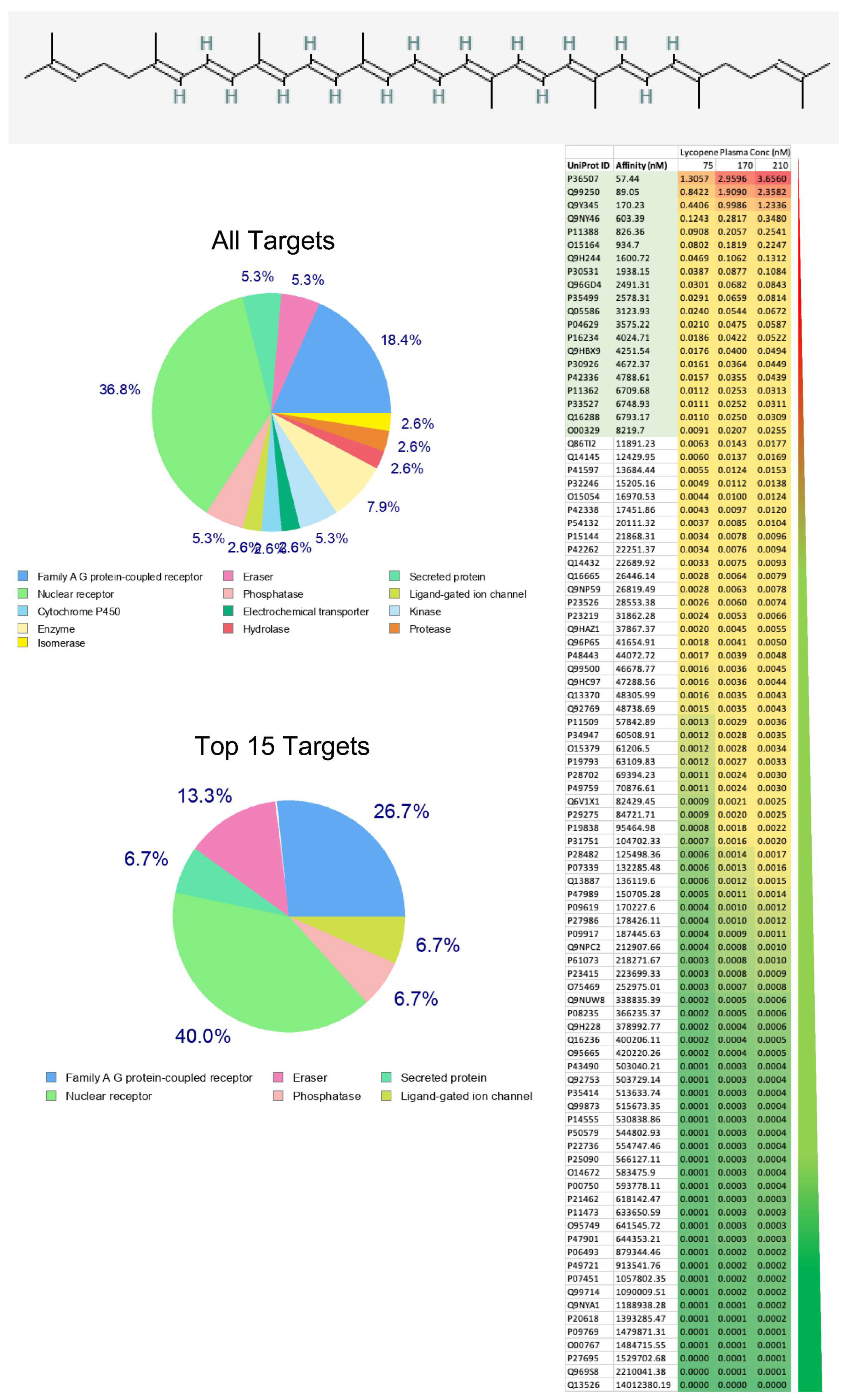
Figure 2.
Heat map of the protein-protein interaction networks of all lycopene targets, based on the number of hydrogen bonds formed between a pair of targets at 10 Armstrong distance. Venn diagram in the middle shows the logical relationships between all, high (at least ten pairs of targets showing >8000 H-bond between them) and significant (a minimum of ten pairs of targets showed >15000 H-bond between them) network targets of lycopene. The Venn diagram below shows the logical relationships between high affinity and high network targets of lycopene. The three major categories of lycopene targets are shown i.e., 1) high affinity targets (orange), 2) high network targets (green) and 3) targets with high affinity and networks (olive).
Figure 2.
Heat map of the protein-protein interaction networks of all lycopene targets, based on the number of hydrogen bonds formed between a pair of targets at 10 Armstrong distance. Venn diagram in the middle shows the logical relationships between all, high (at least ten pairs of targets showing >8000 H-bond between them) and significant (a minimum of ten pairs of targets showed >15000 H-bond between them) network targets of lycopene. The Venn diagram below shows the logical relationships between high affinity and high network targets of lycopene. The three major categories of lycopene targets are shown i.e., 1) high affinity targets (orange), 2) high network targets (green) and 3) targets with high affinity and networks (olive).
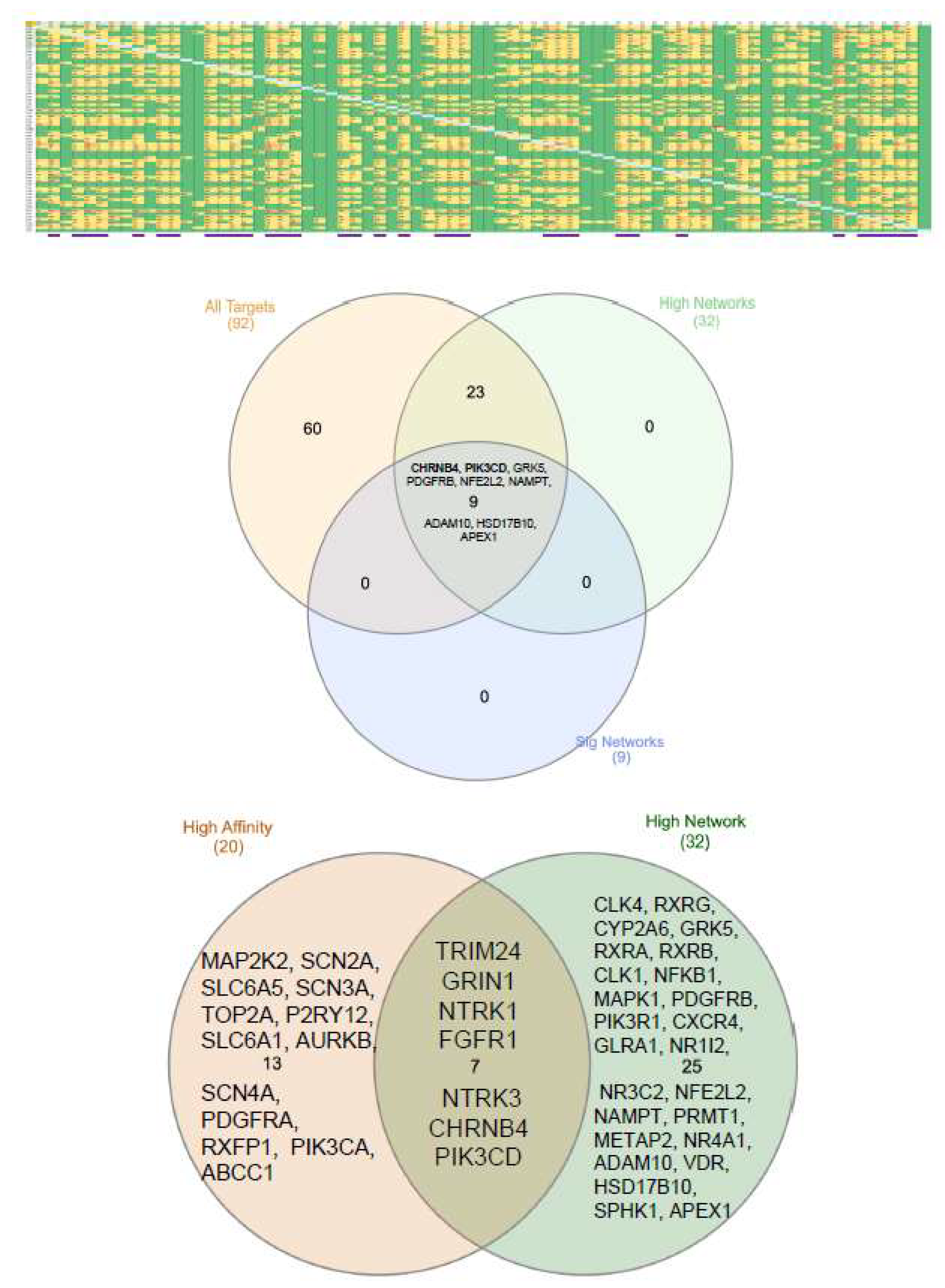
Figure 3.
Tissue specific protein expression of the top five high affinity targets (MAP2K2, SCN2A, SCN3A, TOP2A and TRIM24) of lycopene in humans. Expression of MAP2K2, SCN2A, SCN3A, TOP2A and TRIM24 was observed in 41, 5, 44, 29 and 30 different tissues. The Venn diagram shows the synergistic relationship between the tissues expressing top five high affinity targets of lycopene. MAP2K2, SCN3A, TOP2A and TRIM24 were collectively expressed in 23 different tissues, while SCN3A was the only lycopene target expressed in heart muscle.
Figure 3.
Tissue specific protein expression of the top five high affinity targets (MAP2K2, SCN2A, SCN3A, TOP2A and TRIM24) of lycopene in humans. Expression of MAP2K2, SCN2A, SCN3A, TOP2A and TRIM24 was observed in 41, 5, 44, 29 and 30 different tissues. The Venn diagram shows the synergistic relationship between the tissues expressing top five high affinity targets of lycopene. MAP2K2, SCN3A, TOP2A and TRIM24 were collectively expressed in 23 different tissues, while SCN3A was the only lycopene target expressed in heart muscle.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



