
Submitted:
26 April 2025
Posted:
28 April 2025
You are already at the latest version
Abstract
Cadmium(Cd)contamination poses significant risks to agricultural productivity and ecosystems, necessitating effective remediation strategies. This study evaluated the efficacy of iron (Fe)-modified corn straw biochar (FB) in immobilizing Cd and reducing its uptake in lettuce, compared to raw biochar (B). FB was synthesized by pyrolyzing Fe(NO₃)₃-treated corn straw and characterized for physicochemical properties. Pot experiments tested Cd immobilization at 0, 1, and 2 mg kg⁻¹ Cd levels with 0%, 1%, and 3% (w/w) B or FB applications. Results demonstrated that FB exhibited higher specific surface area, pore volume, and oxygen-containing functional groups than B, enhancing Cd adsorption via mechanisms including surface complexation (-OH, Fe-O), precipitation (CdCO₃), and π-π interactions. FB reduced soil bioavailable Cd by 15–28% and shifted Cd speciation from exchangeable to Fe/Mn oxide-bound fractions, outperforming B even at lower doses (1% FB ≈ 3% B). Additionally, FB improved soil fertility, enzyme activities, and enriched Cd-immobilizing bacteria , promoting lettuce growth and reducing Cd content in roots (25–40%) and leaves (33–47%). The findings highlight FB as a cost-effective and sustainable amendment for Cd-contaminated soils, combining enhanced adsorption, microbial modulation, and reduced application rates. This study provides critical insights into Fe-modified biochar’s dual role in soil remediation and crop safety, supporting its practical application in sustainable agriculture.
Keywords:
cadmium
; Fe-modified biochar
; lettuce
; bacterial communities
; im-mobilization
1. Introduction
Soil Cd contamination has become a critical global environmental issue due to its high toxicity, non-degradability, and formidable mobility, threatening ecosystems, agricultural productivity, and human health [1]. Cd enters the soil through anthropogenic activities, such as mining, industrial waste discharge, and the excessive application of fertilizers [2]. Findings have revealed that approximately 7% of China’s arable land is contaminated with Cd, and about 12 million tons of grains are contaminated with heavy metals each year [3]. However, removing Cd from polluted soils is quite difficult. Therefore, the development of effective, environmentally friendly, and low-cost remediation techniques is urgently needed for environmental restoration and to safeguard agricultural productivity.
The in-situ stabilization technique of heavy metals from soil ecosystems by application of soil amendments is eco-friendly and economical, in which biochar is the most extensively researched [4]. Biochar, a carbon-rich material created from the pyrolysis of biomass, is effective for the immobilization of heavy metals and remediation of Cd-contaminated soils. Corn (Zea mays L.) is primarily cultivated in northern China as a staple food, generating over 200 million metric tons of corn straw each year [5]. This straw hardly returned to the field has become one of the largest sources of agricultural solid waste. Therefore, converting crop straw into biochar for use as a soil amendment can enhance carbon sequestration, reduce carbon emissions, and offer ecological and economic benefits. Several studies have found that corn straw biochar can decrease soil Cd availability and plant Cd uptake, and improve soil health and plant growth [6,7]. However, the efficacy of raw corn straw biochar in removing heavy metals is limited by its relatively small specific surface area and few active sites. Considering the costs and limitations of biochar application, enhancing its adsorption efficiency is crucial. Improved adsorption would allow for lower application rates, thereby reducing costs and enhancing its economic viability.
Recently, Fe materials have been widely used in the modification of biochar for their high affinity toward heavy metals, high specific surface area, natural abundance, and environmental friendliness [8]. The maximum adsorption capacity of Cd(II) by K2FeO4 modified biochar reached 330.06 mg g-1, which was 5.15 fold of pristine biochar [9]. Fe-containing biochar is more effective at fixing Cd by forming C-O-Fe structure, iron oxides (Fe3O4, γ-Fe2O3, and Fe-O-Fe), iron-containing functional groups (-Fe-R-COOH and Fe-R-OH) and the mineral crystal XiFeYjOk on biochar. These components can react with Cd to exchange, form complexes, and precipitate, thus improving the removal performance of Cd [10]. Rice husk was modified by Nano-Fe3O4 and applied it to reduce the available Cd content in soil as well as the Cd content in brown rice[11]. It has been found that application of 2% Fe-modified biochar increased the biomass of wheat grains by 148%, decreased the Cd content in wheat grains by 76.3%, and improved the physiology of the plant and the soil nutrition [12]. Besides, Iron is an essential micronutrient in plants, Fe is involved in many plant physiological processes, such as redox reactions, respiration, and chlorophyll biosynthesis [13]. However, most of the previous reports focused on the stabilization effects of biochar on Cd-contaminated soil, the studies on the mechanisms of Fe-modified corn straw biochar in decreasing soil available Cd and the correlation with fractions of Cd and microbial community are few. Furthermore, there is limited research on whether Fe-modified biochar can achieve a comparable level of Cd immobilization with reduced quantities compared to raw biochar, and the measures involved.
Lettuce (Lactuca sativa L.) is a crucial leafy vegetable widely distributed in the world. The accumulation of Cd in the edible tissues of the lettuce exposes humans to Cd toxicity, posing a potential hazard to public health [14,15]. However, the potential of Fe-modified biochar in reducing the Cd uptake in lettuce has not yet been well evaluated. In this study, we hypothesized that the low-temperature Fe-modified biochar has a greater potential to immobilize Cd and reduce Cd uptake in lettuce. The aims of this study were: (1) to synthesize a new biochar based on corn straw loaded with Fe-oxides at a low temperature, and assess its Cd adsorption performance; (2) to evaluate its effects on the availability of Cd in soil, soil physicochemical properties, enzyme activities, and microbial community; (3) to investigate its impacts on lettuce growth and Cd uptake. The obtained results will contribute to the utilization of agricultural waste resource applications and a better understanding of the mechanisms by which Fe-modified biochar immobilizes Cd in soil.
2. Materials and Methods
2.1. Synthesis and Screening of Absorbent
Preparation of biochar: The corn straw used in this experiment was obtained from a commercial company in Jinan, China. Heat the corn straw that had been screened to a diameter of 1 mm in a muffle furnace under oxygen-limited conditions. The temperature should rise from room temperature at a rate of 10℃ min-1 to 400℃ and be maintained for 2 h to obtain raw biochar (B).
Preparation of Fe-modified biochar: B was added to the Fe(NO3)3·9H2O solution according to the setting mass ratio of Fe to B (0.025:1, 0.05:1, 0.075:1, 0.10:1, 0.125:1, and 0.15:1, and named as 0.025FeBC, 0.05FeBC, 0.075FeBC, 0.10FeBC, 0.125FeBC, and 0.15FeBC), and the pH was adjusted to 7 with 1 mol L-1 sodium hydroxide. The mix solution was continuously stirred for 12 h at a constant temperature of 25℃, and then dried at 50℃ on the oven. Then, materials were pyrolyzed at 400℃ for 30 min to obtain Fe-modified biochars. All the experimental biochars were passed through a 100 mesh sieve.
Screening of biochar: Biochar samples (0.025 g) were added into 25 mL of a solution containing 20 mg L-1 Cd2+ (CdCl2·2.5H2O) and shaken at 180 rpm with constant temperature 25℃ for 24 h. The screening experiments were performed under pH = 7.00 ± 0.02. After the experiment, the mixture was filtered with 0.45 µm filters, and the Cd2+ concentration in the filtrate was measured by inductively coupled plasma optical emission spectrometry (ICP-OES Optima 2000, PerkinElmer Co., USA). Then, Fe-modified biochar (FB) with the best Cd2+ removal rate was selected and conducted in the subsequent pot experiment.
2.2. Characterization of Biochar
The ash content of B and FB was measured by combustion at 800℃ to constant weight. The pH was measured using a pH meter (1:20 w/v). The surface elemental distribution of biochars was characterized by Elementar Vario EL cube ( Unicube, Elementar, Germany). The surface morphologies and elemental composition were characterized with SEM-EDS (Regulus 8100, Hitachi, Japan). The characteristics of specific surface and pore structures were characterized with the Micromeritics ASAP 2020 (America). The crystal structure of the materials was analyzed by X-ray diffraction (XRD, D8 Advance, Bruker, Germany). Fourier transform infrared spectra (FTIR, Nicolet iS5, Thermo Scientific, USA) were used to evaluate the functional groups of samples. The elemental combination states and surface compositions of the biochars were characterized by X-ray photoelectron spectrometry (XPS, Escalab 250Xi, Thermo Fischer, USA).
2.3. Pot Experiment
The soil used for the pot experiment was collected from the top layer (0-20 cm) of a non-metal-contaminated yellow-brown soil (Alfisols) field located in Jinan, China. The soil had the following properties: pH 6.68; organic matter (OM), 17.36 g kg−1; AP, 24.61 mg kg−1; AK, 90.32 mg kg−1, CEC, 15.05 cmol kg−1, NH4+-N, 8.15 mg kg−1, NO3--N, 45.42mg kg−1, The pot experiment was performed based on the method described previously [16]. Each pot (20 cm wide at the top, 15 cm wide at the bottom, and 14 cm in height) contained 3 kg of soil supplemented with 0, 1, and 2 mg kg−1 Cd (CdCl2·2.5H2O). For each level of Cd content, we mixed B and FB into the soil homogeneously to achieve the desired doses of 0%, 1%, and 3% (w/w). There were a total of 15 treatments, including (C0B0, C0B1, C0B3, C0FB1, C0FB3, C1B0, C1B1, C1B3, C1FB1, C1FB3, C2B0, C2B1, C2B3, C2FB1, C2FB3) (Table S1), each of which had three replicates. Deionized water was used for irrigating the pots which were placed in a dark environment for three weeks. Nine surface-sterilized lettuce seeds (Shandong Academy of Agricultural Sciences) were sown in each pot, and plants were thinned to four seedlings per pot after 7 days of sowing. During the growth period of lettuce, ensure that the cultivation management is in line with normal production, regularly weed, pest control, and water, with a growth period of 60 days. The pots were allocated throughout the greenhouse at the Qilu University of Technology, China (average temperature 20-25℃, average photoperiod 10-12 h per day, and relative humidity 50-60%) by a completely randomized design protocol.
2.4. Plant Sample Analysis
The harvested plants were washed and divided into roots and shoots to record the fresh weight. The fourth leaves, from top to bottom, were weighed, and then ground to a fine powder in liquid nitrogen for subsequent analyses. The VC content was determined by using the 2,6-dichloroindophenol titrimetric method (AOAC) [17]. The MDA content, SOD, and POD activities were measured with ELISA kits (JM-09865P2, JM-01183P2, JM-01185P2, Jingmei Bio Inc., Jiangsu, China) following the manufacturer’s protocol. Lettuces, including shoots and roots, were oven-dried at 105℃ for 30 min, and then at 55℃ until a constant weight was achieved. The dry weights were recorded. The oven-dried samples were ground to powder and then digested with a mixture of HNO3:H2O2 (v/v, 8:2). The Cd content was measured by ICP-OES.
2.5. Soil Sample Analysis
Soils tightly adhering to the roots of plants in each pot were sampled as rhizosphere soils using the hand-shaking method. Air-dried and sieved soil samples were measured for the physicochemical properties. The soil pH was measured by pH meter (ZD-2A, Shanghai, China) in water : soil ratio of 5:1. OM was measured by potassium dichromate sulfate oxidation method [18], and CEC was assessed by using the 1.0 M ammonium acetate extraction method, followed by titration with 50 mM hydrochloric acid [19]. AK and AP were extracted with 1.0 M NH4OAc (pH 7.0) and 0.5 M NaHCO3, respectively, with a soil: solution ratio of 1:10 and 1:20, respectively [20]. The available Cd of soil samples (DTPA-Cd) was estimated by diethylenetriaminepentaacetic acid (DTPA) solution extraction. Chemical fractionations of Cd were determined using a sequential extraction procedure [21]. These fractions were: exchangeable fraction (EX-Cd), carbonate-bound fraction (CB-Cd), Fe/Mn oxides-bound fraction (OX-Cd), organic-bound fraction (OM-Cd) and residual fraction (RS-Cd). The Cd concentration in the extracts was detected by ICP-OES.
Fresh Soil samples were extracted with 2 M KCl solution (soil : KCl = 1:5), and measured by an Elementary Analyzer (Thermo-Element Flash EA 1112, USA) to determine NH4+-N and NO3−-N contents [22]. Soil urease and sucrase activities were measured with ELISA kits (code JM-06009902, and JM-0602802, Jingmei Bio Inc., Jiangsu, China) according to the manufacturer’s instructions.
2.6. High-Throughput Sequencing
The changes of bacterial community during the culture period were studied. Soil total DNA was extracted from 0.5 g fresh rhizosphere soil using a FastDNA SPIN Kit (MP Biomedicals, Santa Ana, CA, United States). The bacterial 16S rRNA V3-V4 regions were amplified using 515 F/806 R (5’-GTGYCAGCMGCCGCGGTAA-3’; 5’-GGACTACNVGGGTWTCTAAT-3’) [23]. PCR was conducted according to the method of [24]. The amplicon sequencing was performed using anIllumina HiSeq2000 platform (Novogene Bioinformatics Technology Co., Ltd., Tianjin, China). The sequenced data has been uploaded to the NCBI Sequence Read Archive (SRA) database with the accession number SAMN41529245.
Bioinformatic analysis was performed according to previous studies with some modifications [25]. VSEARCH (version 2.21.1) software was used to perform quality control and operational taxonomic unit (OTU) clustering. The paired-end reads were merged and reads with low quality (expected errors per base > 0.001, containing N, length < 150 bp) were filtered, and then chimeric reads were detected and removed with UCHIME. The high-quality reads were clustered into OTUs by UPARSE (version 7.1) based on 97% similarity [26]. The taxonomy of the bacterial sequence was analyzed by the RDP Classifier algorithm [27]with a 70% confidence threshold against the SILVA database (version 138) and UNITE database (version 8.3), respectively. The alpha diversities were calculated from the representative OTU table in QIIME2 (2019.4). The differences in community compositions were performed by non-metric multidimensional scaling (NMDS) based on the Bray-Curtis distance. Spearman correlation coefficients were calculated using R v.3.3.1 software to analyze the correlation between microbial abundances and soil physicochemical properties
3. Results
3.1. Selection and Characterization of Modified Biochar
The effects of the modifier ratio while removing Cd2+ were shown in Figure S1. Fe-modified biochar could significantly improve the removal rate of Cd2+. By increasing the mass ratio of Fe to raw biochar from 0.025:1 to 0.15:1, the Cd2+ removal rates first increased and then decreased. The maximum removal rate occurred at 0.1FeB, with a removal rate was 84.15%. The removal rates of 0.125FeB and 0.15FeB, at 79.42% and 78.87% respectively, significantly decreased compared to that of 0.1FeB. A rise in the mass ratio may obstruct the porous framework, leading to a decrease in Cd2+ physisorption capacity. The results were consistent with those of [28], which showed that the removal rate of Cd2+ decreased as the proportion of iron-modified biochar increased. Based on these results, 0.1FeB was selected for the following study and given the name FB.
The elemental analysis data of total C, H, O, N, and S contents of B and FB were shown in Table 1. The elemental compositions of B and FB were mainly C and O, which were 61.99% and 45.94% respectively. After modification, C and H contents were reduced, and the O content increased. The H/C atomic ratio decreased, suggesting an increase in the content of aromatic compounds and an improvement in the structural stability of the biochar by the introduction of iron ions. Research indicated that biochar exhibits significant stability and has the potential to persist in the environment for extended periods when its H/C atomic ratio is below 0.6 [29]. The atomic ratio of O/C increased indicating that Fe-modified biochar included more oxygen functional groups (OFGs).
The physicochemical properties of adsorbents were shown in Table 1. The pH of biochar followed the order of B < FB, indicating that Fe(NO3)3 modification enhanced biochar pH. The ash content of FB was 1.78 times greater than that of B, likely due to the introduction of iron ions. The specific surface area (ABET) and total pore volume (Vtot) of FB were higher than those of the B, indicating that the addition of iron oxides can enhance the surface area and total pore volume of B. Fe(NO3)3 can promote oxidative pyrolysis during modification, and Fe oxides increase the surface area and constructed new channels [30]. However, the average pore diameter (Dapd) of FB was lower than that of B, which may be due to metal oxide particles blocking the channels, thus reducing the pore diameter [31]. Typically, biochar with a larger specific surface area offered more active sites, enhancing the efficiency of adsorption reactions.
The morphology and surface elemental distribution of biochar were characterized with SEM-EDS (Figure S2). B exhibited a smooth surface while FB comprised a rough surface with many tiny particles attached, which might be the modifier particles attached to the FB’s surface and increased the roughness. This was the reason that the ABET of B was higher than FB. The EDS analysis confirmed that Fe was successfully loaded on the surface of FB. According to the XRD (Figure S3), the weak diffraction peaks at 2θ=35.6°, and 62.4°, which matched with Fe2O3 were observed in the FB [32,33]. These findings indicated that FB contained a small amount of Fe2O3, suggesting that Fe was successfully loaded on biochar in the form of iron oxides. To identify the functional groups on the surface of biochar, the FTIR spectra were employed (Figure 1A). The broad bands at 3380 cm-1 and 1593 cm-1 were ascribed to -OH and C=C stretching vibration, respectively [34,35]. The typical peak at 2914 cm-1 was assigned to -CH2 [35]. The band observed at 1084 cm-1 was the C-O group in the alcoholic groups [36]. The characteristic peak at 569-580 cm-1 was ascribed to the stretching vibration of Fe-O [37], and it appeared on the FTIR spectra of FB, indicating effective loading of Fe onto the biochar as oxides [38]. This loading may play an important role in Cd immobilization due to the resulting negative charge [39]. The peaks of -OH and C-O-C within FB were strengthened compared with those in B, indicating its higher abundances of OFGs compared to B [34], which could enhance the biochar’s affinity for Cd through increased surface complexation and ion exchange interactions [40]. The XPS result showed that Fe2p peak was present on the FB surface, and the peaks at 713.5 eV and 711.1 eV were attributed to Fe(III) compounds [41] and Fe2O3 [42](Figure 1B and C), thereby affirming that Fe was successfully loaded onto the biochar. In the C 1 s spectra, the ratio of the π-π* satellite band (293.3 eV) increased significantly from 9.5% to 14.4% after iron modification (Figure 1E). This suggested that the iron modification enhanced the π-π* interactions within the biochar, which are generally associated with improved adsorption capabilities [43]. O 1s spectrum peak clearly showed the presence and contents of oxygen-containing compounds (Figure 1F). The ratio of C-OH (533.0-533.4 eV) increased significantly from 16.52% to 37.85% after iron modification, suggesting that more OFGs were found in FB. This result was consistent with FTIR findings [44].
3.2. Mechanism for Cd Immobilization by Fe-Modified Biochar
In this study, iron was successfully loaded in FB. To understand the Cd adsorption mechanism of FB, the FTIR spectrum was used to assess the alterations in the functional groups on FB before and after adsorption (Figure 1A). The FB surface presented more -OH than B, and its peak weakened after Cd2+ adsorption. This might be due to the complex reaction between Cd2+ and -OH on the surface of FB forming -O-Cd. The wave number of peak at 579cm-1 was shifted and sharpened, which implies that the Fe-O groups may have taken part in the Cd2+ adsorption of FB. The aromatic C=C bonds on biochar surfaces generate electron-rich π-systems capable of donating π-electrons for interactions with cations [45]. The peak of C=C on FB weakened and shifted after Cd2+ adsorption, which might be ascribed to cation-π interaction [46]. The above results suggested that the surface complexation between Cd2+ and OFGs (-OH, Fe-O), and cation-π interaction played critical roles in Cd2+ adsorption.
To further elucidate the adsorption mechanism, XPS was employed to examine the shifts in binding energies of the elements in FB pre- and post-Cd adsorption. The new peak, Cd 3d, appeared in the FB after Cd2+ adsorption, suggesting the immobilization of Cd on FB [28] (Figure 1B). The results showed that the XPS high-resolution spectra of Fe 2p, C 1s, O 1s, and Cd 3d regions. The intensity and central peak position of Fe 2p showed significant changes after Cd2+ adsorption (Figure 1C), which confirms that the Fe-O group was involved in the Cd2+ adsorption. In the C 1s XPS spectra (Figure 1E), the π-π* content dramatically decreased from 14.4% to 1.1% after Cd adsorption. This substantial decrease can likely be attributed to the interaction between the biochar’s functional groups and Cd ions, which may lead to the partial disruption or modification of the π-π* structures. Compared with FB, the ratio of -OH in FB-Cd decreased from 37.85% to 22.87% (Figure 1F), suggesting that Cd2+ can react with OFGs. The XPS high-resolution spectra of Cd 3d region showed that the peak at 405.7 eV belonged to CdCO3 (Figure 1D). All these results demonstrated that surface complexation, cation-π interaction, precipitation, and exchange reaction were the main mechanisms of Cd2+ sorption.
3.3. Soil Physiochemical Properties
Effects of biochars on physicochemical properties of lettuce rhizosphere soils were shown in Table S2. Compared with the non-amended soils, the addition of B to soils resulted in a significant increase in pH by 0.51-0.81 units and CEC by 11-15%, while FB resulted in an increase in pH by 0.55-0.87 units and CEC by 12-18%, respectively. The increase in soil pH induced by biochar might be due to the high pH of the two types of biochar [47]. Biochar can develop more OFGs, leading to increased CEC and negative charge of soils [48]. The results obtained in this study agreed with the findings of [49], who found that composted biochar increased the pH and CEC of the rhizosphere soil of Chinese cabbage in a greenhouse experiment. Considering that CEC is the indicator of the capacity of soils to retain nutrients, the biochars we have analyzed may increase the ability of studied soils to hold nutrients [50]. Application of both B and FB significantly increased soil OM, AP, and AK contents compared to the non-amended biochar soils. OM, AP, and AK contents were significantly higher in soils treated with 3%B or 3%FB compared to those treated with 1%B or 1%FB. With increasing application, B and FB significantly increased soil OM, AP, and AK. The application of biochars increased AP, thereby promoting the formation of Cd-phosphate and reducing the availability of Cd. The B and FB applications significantly decreased the NH4+-N content by 16-26% and 18-29%, respectively, and increased the NO3--N content by 23%-49% and 15-51%, respectively, compared with the non-applied treatments. NO3-N content increased significantly. A similar study found that the coastal soil NH4+-N content decreased and NO3--N content increased following the amendment of wood chips biochar [51]. Although the effect of biochar on soil N availability has been extensively studied, the outcomes are not consistent. In a comprehensive meta-analysis comprising data from 124 published articles, application biochar reduced soil NO3--N content by 12% and NH4+-N by 11% [52]. In contrast, some studies have shown significant increases in the content of available N in soil following biochar application [53,54]. Soil enzymes are important evaluation indicators of soil quality and play important roles in soil biological functions. Sucrase decomposes sucrose into glucose and fructose, providing nutrients for soil microorganisms and plants, while urease enhances soil available nitrogen by converting organic nitrogen into a form accessible to plants [55]. In this study, both types biochar application significantly increased soil urease and sucrase activities. Soil enzyme activities were higher in samples treated with 1% FB and 3% FB, respectively than in those treated with 1% B and 3% B. These results concurred with those found by [56]. It is indicated that Fe-modified biochar can promote material transformations and energy flow in the soil system, thus improving soil quality [57].
3.4. Soil Cd Bioavailability and Fractionation
The threat of heavy metal contamination is not only related to the total concentration of heavy metals but also to their chemical forms. Utilizing the chelating agent DTPA as a standard measure facilitates the estimation of Cd’s bioavailable portion in soil [58]. Figure 2A illustrated that amending soils with B and FB led to a reduction in available Cd content by increasing the soil pH. Previous studies have demonstrated the adsorption and precipitation of Cd by soil increased with the rise in soil pH, thereby reducing DTPA-Cd in the soil [59]. This may be due to the reaction of Cd2+ with OH- and CO32- in the soil, which results in the formation of solid compounds as hydroxides and carbonates precipitate [60]. Additionally, improvements in CEC and OM resulting from biochar application contribute to enhanced complexation, thereby reducing soil DTPA-Cd [11,61]. Interestingly, FB presented superior Cd immobilization results compared to B, decreasing DTPA-Cd by 15-28%.
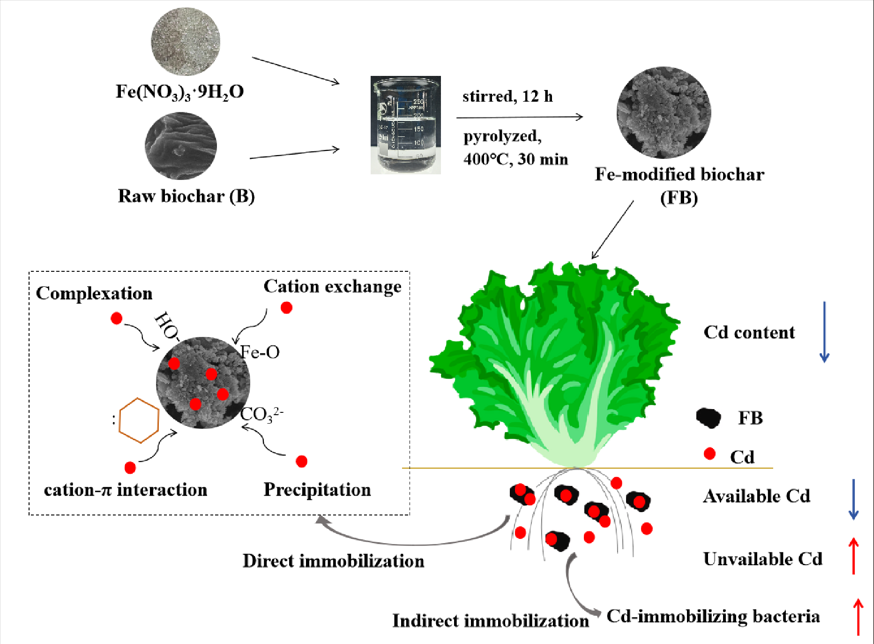
The effect of adding B and FB on the distribution of different forms of Cd in the soil was shown in Figure 2B. It revealed that Cd was predominantly associated with exchangeable and Fe/Mn oxide-bound fractions. Collectively, these fractions constituted over 80% of the total Cd in all soil samples tested. The addition of B and FB altered Cd speciation by shifting it from EX-Cd to OX-Cd, with a decrease in EX-Cd’s percentage by 11-29% and 20-45% and an increase in OX-Cd’s percentage by 9-40% and 21-52%, respectively. This indicated that biochar could convert the state of Cd from an available state to an unavailable state, thus reducing the bioavailability of Cd. This result was similar to the work of [62]. It also has been found that Fe-modified biochar decreased more EX-Cd and increased more OX-Cd in the soil than raw biochar [63]. Notably, the DTPA-Cd, EX-Cd, and OX-Cd levels were comparable between treatments with 3%B and 1%FB, highlighting that iron modification could enhance biochar’s ability to immobilize Cd in soil more efficiently, despite the reduced quantity used. The observed increase in Cd immobilization in soils treated with FB may be attributed to the introduction of iron (a trace element), which can increase the iron oxide content in soil, and potentially enhance its capacity to adsorb Cd. This process facilitated the conversion of available Cd forms into less bioavailable Fe/Mn oxide fractions in soil. Furthermore, iron oxides could adsorb Cd by forming surface precipitates, raising the Cd immobilization potential of the Fe-modified biochar [64]. Additionally, the porous structure of FB provided more physical space for the immobilization of Cd. In summary, FB affected the Cd mobility in the two soils via: (1) indirect interactions, in which FB indirectly enhanced the Cd adsorption in soil by increasing the pH, CEC, and, OM of soils; (2) direct interactions, namely through direct complexation and precipitation with FB, which reduced Cd mobility in the soils. These findings highlighted the potential benefits of iron-modified biochar as a strategy for more effective soil remediation.
3.5. Soil Microbial Communities
Soil microbial community composition and structure are often employed as indicators of soil quality for their sensitivity to environmental changes [65,66]. In this study, a total of 4,550,523 high-quality reads were obtained from all soil samples using the Illumina Miseq platform. The characteristics of OTUs are shown in a Venn diagram (Figure S4). The number of bacterial OTU identified in the 0B, 1%B, 3%B, 1%FB, and 3%FB was 13851, 13494, 12131, 12853, and 12879. In total, 4048 bacterial OTUs were common to all samples. The number of bacterial OTUs in the soils treated with B and FB was lower than that in the non-amended soils, indicating that the application of B and FB affected the diversity of soil bacteria.
The changes in species α-diversity and richness during soil remediation using both types of biochars were shown in Table S3. The number of OTU and the Chao index were used to assess microbial richness, while the Shannon and Simpson indices were used to evaluate microbial diversity. Both the OTU numbers and the Chao index suggested that the application of biochars led to lower bacterial richness, and the Shannon and Simpson indices indicated a reduction in bacterial diversity compared to the non-amended groups. Therefore, the addition of raw biochar and Fe-modified biochar resulted in decreased richness and diversity of the microbial communities. This result was consistent with the study of [67], who confirmed that the application of rice straw biochar can suppress the abundance and diversity of bacterial communities by changing the soil’s pH. This may be caused by the extensive use of biochar, which significantly raises the soil’s pH, thereby inhibiting the growth of some bacterial species [68]. This was supported by the decrease in the population of Acidobacteria, which is sensitive to pH shifts (Figure S6) [69]. The differences in the β-diversity of bacterial community among soil samples treated with different types and doses of biochars were evaluated by using non-metric multidimensional scaling plots (NMDS) of the weighted pairwise UniFrac distance ordinations (Figure S5). Greater consistency in the similarity of bacterial community composition is indicated by closeness. The communities in rhizospheric soil treated with different types and doses of biochar were significantly separated. The results indicated that both modification ratio and dose of biochar caused shifts in the bacterial community composition. It has been reported that biochar derived from Mg-Mn-modified corn stover and eggshells altered the bacterial community structure and patterns in the rhizosphere soil of rice [70]. Recently, the effect of biochar on the soil microbial community structure has received considerable attention and has been found to depend largely on the properties of the biochar, the dose of biochar, and the cultivation time [71,72].
The bacterial community structure of all soil samples at the phylum level was shown in Fig. S6. The dominant bacterial phyla included Proteobacteria (22-31%), Actinobacteria (12-16%), Acidobacteria (8-18%), and Firmicutes (4-10%). Compared with the non-amended soils, raw biochar and Fe-modified biochar amended soils increased the abundance of the phyla Proteobacteria, Actinobacteria, Firmicutes, and Bacteroidota, while decreasing the abundance of Acidobacteriota. Proteobacteria is a predominant bacterial group in Cd-contaminated soil and has been demonstrated to play a significant role in the remediation of pollutants [73]. Actinobacteria are known for their potential to immobilize HMs by biosorption, bioaccumulation, and the production of metal-binding compounds[74]. Firmicutes possess a lot of resistant genes of heavy metals, promoting their accelerated spread in heavy metal-contaminated conditions [75]. Bacteroidetes produce sphingolipids to protect cell surfaces and functions, thus facilitating its survival under challenging conditions, especially in metal-rich ecosystems [76]. This suggested that these four bacterial phyla may play important roles in HMs immobilization. Besides, they have been found to play an important role in decomposing organic matter, inhibiting pathogenic organisms, and participating in many soil biogeochemical cycles [77]. Furthermore, the relative abundance of Acidobacteria reduced, which could prove advantageous in mitigating Cd activation, as Acidobacteria are capable of mobilizing Cd by producing acidic compounds [78].
The relative abundance of bacteria at the genus level was shown in Figure S7. The most abundant taxa were genera Pseudarthrobacter (1.3-7.8%), Sphingomonas (1.3-7.8%), Pseudomonas (1.3-7.8%), JG30-KF-CM45 (1.3-7.8%), Vicinamibacteraceae (1.3-7.8%), Subgroup_7 (1.3-7.8%). The relative abundance of Pseudarthrobacter, Sphingomonas, and Pseudomonas in 3%B, 1%FB, and 3%FB was higher than that in 0%B and 1%B (Figure 3). Pseudarthrobacter contains various plant growth-promoting bacteria, and it can also effectively remove soil pollutants [79,80]. Sphingomonas and Pseudomonas have been found to play crucial roles as plant rhizosphere bacteria, enhancing plant growth by fixing nitrogen, suppressing pathogen growth, and producing plant hormones [81,82]. These bacteria are also known for their ability to solubilize insoluble P and K, making these nutrients available to plants [51,83]. This was consistent with the increased content of nutrients such as AP and AK in the biochar-amended soils. Moreover, Sphingomonas and Pseudomonas could immobilize Cd by producing biofilm, extracellular polymeric substances, and arginine decarboxylase [84,85]. Thus, it seems that the application of biochar at higher doses, or the use of Fe-modified biochar, could enhance the population of bacteria that promote plant growth and immobilize Cd.
To understand the effects of soil properties and different fractions of Cd on the relative abundance of the dominant bacteria at both the phylum and genus levels, Spearman’s correlation analysis was performed (Figure 4). The relative abundance of Proteobacteria, Actinobacteria, Firmicutes, and Bacteroidota was significantly positively correlated with AP, AK, pH, OM, CEC, NO3--N and sucrase activity. These bacterial groups could effectively promote plant growth, as evidenced by the strong positive correlations with their relative abundances. At the genus level, the relative abundance of Pseudarthrobacter, Sphingomonas, and Pseudomonas was significantly positively correlated with AP, AK, pH, OM, CEC, NO3--N, urease, and sucrase activities. The plant biomass was significantly positively correlated with the relative abundance of Flavisolibacter, Pseudomonas, Pseudarthrobacter, and Sphingomonas. Flavisolibacter has been previously associated with the process of ammoxidation, which could maintain available N content in the soil [86]. It indicated that Flavisolibacter, Pseudomonas, and Sphingomonas may be effective agents for promoting plant growth by maintaining nutrients in rhizosphere soil. OX-/CB-/OM-Cd were significantly positively correlated with bacterial phyla of Actinobacteria, Bacteroidota, Proteobacteria, and Firmicutes, while showing significant negative correlations with the phyla Verrucomicrobiota, Acidobacteria, Chloroflexi, and Gemmatimonadota. Pseudomonas was found to be significantly positively correlated with OX-/CB-Cd, and significantly negatively correlated with EX-/DTPA-Cd. This suggested that Pseudomonas may be involved in immobilizing Cd by transforming available Cd into unavailable forms. Xu has discovered that Pseudomonas putida X4 can transform 7.5-24.8% of exchangeable Cd primarily into the organic-bound fraction[87].
3.6. Lettuce Growth and Physiological Parameters
As shown in Table 2, the application of biochar materials significantly enhanced lettuce growth, with leaves and roots dry weight increasing by 52-97% and 32-53% regardless of Cd contamination. Besides, compared to the control, the VC content in lettuce leaves significantly increased in B3, FB1, and FB3 treatments. MDA, as a crucial byproduct of membrane lipid peroxidation, serves as an indicative measurement of membrane system impairment in plants under stressful situations. In B3, FB1 and FB3 treatments, the MDA content of lettuce significantly decreased by 21-33% compared to control, indicating that application of 3%B, 1% and 3%FB could mitigate heavy metal stress on lettuce. Compared with control, POD and SOD activities were significantly increased by 27-73% and 33-68% at all biochar treatments with or without Cd stress. However, compared to the control, the application of 1%B had no effects on the VC and MDA contents of lettuce leaves.
Cd toxicity can induce oxidative stress in cells, increase ROS levels, degrade the antioxidative defense, reduce chlorophyll, and impair photosynthesis in plants, thereby decreasing plant yield [88]. Biochar is considered to play a crucial role in enhancing plant growth and mitigating Cd toxic impacts by immobilization of Cd in soil [89,90]. Liu et al. (2022) indicated that pig manure, sewage sludge, and corn straw biochars increased the plant height and fresh weight by lettuce at 1% application rates [91]. In this study, we also found that the application of B and FB remarkably enhanced the biomass of lettuce and mitigated Cd stress. The observed beneficial effects could potentially be due to improved soil qualities, such as increased organic matter, availability of NPK nutrients, activities of soil enzymes, and the amount of plant growth-promoting bacteria (see Table S2 and Figure 3).
3.7. Lettuce Cd Uptake
To further investigate the influence of B and FB on the immobilization of Cd in soil, the uptake of Cd by lettuce under different treatments was studied (Figure 5). Application of both B and FB significantly reduced Cd uptake by lettuce, with Cd content in roots and leaves decreasing as the soil biochar dosage increased. Specifically, FB was shown to be more effective than B in reducing Cd content in roots and leaves of plants by 25-40% and 33-47%, respectively. This differential effectiveness was further emphasized by the fact that lettuce plants showed no significant difference in Cd content when treated with either higher-dosage raw biochar (3%) or lower-dosage FB (1%), indicating that iron modification can enhance biochar’s effectiveness at lower application rates, which is pivotal for efficiently remediating Cd-polluted soils. This phenomenon can predominantly be attributed to the decreased availability and increased immobilization of soil Cd, as indicated by the results (Figure 2). It has been found that Cd content in the wheat and rice grains, and fruits of eggplant and tomato was reduced with the application of biochar, by reducing soil DTPA-Cd and EX-Cd[92,93,94,95]. A recent study reported that Fe-modified biochar significantly reduced DTPA-Cd content more than pristine biochar and was found to be more effective at reducing Cd uptake by Chinese cabbage, due to its higher pH, additional minerals, and oxygen-containing functional groups [96].
4. Discussion
In this study, the results of the SEM-EDS, XRD, FTIR spectra and XPS of biochar indicated that Fe was successfully loaded onto the FB (Figure S2, Figure S3, and Figure 1). Besides, the FTIR spectra provided insights into the changes in functional groups on the surface of FB before and after Cd2+ adsorption (Figure 1A). The weakening of the -OH peak after Cd2+ adsorption indicated that Cd2+ ions form complexes with -OH groups, leading to the formation of -O-Cd bonds [72]. The shift and sharpening of the peak at 579 cm-1 in the FTIR spectrum suggested that Fe-O groups may have participated in the Cd2+ adsorption process. This was likely due to the formation of Fe-Cd complexes, as iron oxides are known to have high affinity for heavy metals [28].The aromatic C=C bonds on biochar surfaces generate electron-rich π-systems capable of donating π-electrons for interactions with cations [45]. The peak of C=C on FB weakened and shifted after Cd2+ adsorption, which might be ascribed to cation-π interaction [46]. To confirm the involvement of these functional groups and the adsorption mechanisms, XPS was used to analyze the binding energy shifts of elements in FB pre- and post-Cd2+ adsorption. A new peak, Cd 3d, which corresponded to CdCO3, appeared in the FB after Cd2+ adsorption, suggesting that the precipitation of CdCO3 was another mechanism by which FB immobilized Cd2+ ions [28](Figure 1B and D). The significant changes in the Fe 2p peak intensity and central position (Figure 1C) confirmed that Fe-O groups were involved in the adsorption process. The C 1s region showed a dramatic decrease in the π-π* content after Cd adsorption (Figure 1E), which can likely be attributed to the interaction between the biochar’s functional groups and Cd2+ ions, leading to the partial disruption or modification of the π-π* structures. The ratio of -OH groups in FB-Cd also decreased (Figure 1F), which further supported the hypothesis that -OH groups form complexes with Cd2+ ions, leading to the formation of -O-Cd bonds [28]. All these results demonstrated that surface complexation, cation-π interaction, precipitation, and exchange reaction were the main mechanisms of Cd2+ sorption.
Application of B and FB significantly increased soil pH and CEC (Table S2). The results obtained in this study agreed with the findings of [49], who found that composted biochar increased the pH and CEC of the rhizosphere soil of Chinese cabbage in a greenhouse experiment. The increase in soil pH induced by biochar might be due to the high pH of the two types of biochar [47]. Biochar can develop more OFGs, leading to increased CEC and negative charge of soils [48]. Considering that CEC is the indicator of the capacity of soils to retain nutrients, the biochars we have analyzed may increase the ability of studied soils to hold nutrients [50]. We found that B and FB significantly decreased the soil NH4+-N content and increased the NO3--N content (Table S2). A similar study reported that the coastal soil NH4+-N content decreased, while the NO3--N content increased, following the amendment of wood chips biochar [51]. Although the effect of biochar on soil N availability has been extensively studied, the outcomes are not consistent. In a comprehensive meta-analysis comprising data from 124 published articles, application biochar reduced soil NO3--N content by 12% and NH4+-N by 11% [52]. In contrast, some studies have shown significant increases in the content of available N in soil following biochar application [53,54]. The observed increase in soil nitrogen forms (NO3--N) in our study may be due to the biochar’s ability to promote the nitrification process. Soil enzymes are important evaluation indicators of soil quality and play important roles in soil biological functions. Sucrase decomposes sucrose into glucose and fructose, providing nutrients for soil microorganisms and plants, while urease enhances soil available nitrogen by converting organic nitrogen into a form accessible to plants [55]. We found that FB significantly increased soil urease and sucrase activities (Table S2), which are important for soil biological functions. These results are consistent with those of Sun et al. (2021), who found that Fe-modified biochar can promote material transformations and energy flow in the soil system, thus improving soil quality [57].
In this study, the reduction in DTPA-Cd and the shift from EX-Cd to OX-Cd were critical outcomes for soil remediation (Figure 2). This result was similar to the work of Liu et al. (2019). We found that B and FB decreased available Cd content by increasing the soil pH, CEC and OM (Table S2). Previous studies have demonstrated the adsorption and precipitation of Cd by soil increased with the rise in soil pH, thereby reducing DTPA-Cd in the soil [59]. This may be due to the reaction of Cd2+ with OH- and CO32- in the soil, which results in the formation of solid compounds as hydroxides and carbonates precipitate [60]. Future studies should include the measurement of carbonate content in the soil to definitively confirm the role of precipitation in Cd removal. This will provide a more comprehensive understanding of the mechanisms involved. Additionally, improvements in CEC and OM resulting from biochar application contribute to enhanced complexation, thereby reducing soil DTPA-Cd [11,61] Interestingly, FB was particularly effective in shifting the state of Cd from an available state to an unavailable state. It also has been found that Fe-modified biochar decreased more EX-Cd and increased more OX-Cd in the soil than raw biochar [63]. The observed increase in Cd immobilization in soils treated with FB may be attributed to the introduction of available Fe, which can increase the iron oxide content in soil, and potentially enhance its capacity to adsorb Cd [16]. Future studies should include measurements of available Fe to further elucidate the mechanisms of Cd immobilization in soil. This process facilitated the conversion of available Cd forms into less bioavailable Fe/Mn oxide fractions in soil. Furthermore, iron oxides could adsorb Cd by forming surface precipitates, raising the Cd immobilization potential of the Fe-modified biochar [64]. Additionally, the porous structure of FB provided more physical space for the immobilization of Cd. In summary, FB affected the Cd mobility in the two soils via: (1) indirect interactions, in which FB indirectly enhanced the Cd adsorption in soil by increasing the pH, CEC, and, OM of soils; (2) direct interactions, namely through direct complexation and precipitation with FB, which reduced Cd mobility in the soils. Future studies should focus on characterizing the soil environment in greater detail to conclusively identify the mechanisms responsible for Cd immobilization. This will aid in the development of more effective strategies for soil remediation.
Soil microbial community composition and structure are often employed as indicators of soil quality for their sensitivity to environmental changes [65,66]. The results of this study indicated that the application of B and FB significantly affected the diversity and composition of soil microbial communities. The reduction in bacterial richness and diversity suggested that biochar amendments could alter the microbial community structure (Table S3).This result was consistent with the study of Xu et al. (2022), who confirmed that the application of rice straw biochar can suppress the abundance and diversity of bacterial communities by changing the soil’s pH. This may be caused by the use of biochar, which significantly raises the soil’s pH, thereby inhibiting the growth of some bacterial species [68]. This was supported by the decrease in the population of Acidobacteria, which is sensitive to pH shifts (Figure S6) [69]. The soil bacterial composition treated with different types and doses of biochar was also significantly separated (Figure S5). It has been reported that biochar derived from Mg-Mn-modified corn stover and eggshells altered the bacterial community structure and patterns in the rhizosphere soil of rice[70]. Recently, the effect of biochar on the soil microbial community structure has received considerable attention and has been found to depend largely on the properties of the biochar, the dose of biochar, and the cultivation time [71,72].
The significant increase in the relative abundance of Proteobacteria, Actinobacteria, Firmicutes, and Bacteroidota in biochar-amended soils (Figure S6) suggested that these bacterial phyla may play important roles in the remediation of HMs and the promotion of plant growth. Proteobacteria is a predominant bacterial group in Cd-contaminated soil and has been demonstrated to play a significant role in the remediation of pollutants [73]. Actinobacteria are known for their potential to immobilize HMs by biosorption, bioaccumulation, and the production of metal-binding compounds [74]. Firmicutes possess a lot of resistant genes of heavy metals, promoting their accelerated spread in heavy metal-contaminated conditions [75]. Bacteroidetes produce sphingolipids to protect cell surfaces and functions, thus facilitating its survival under challenging conditions, especially in metal-rich ecosystems [76]. These bacterial groups are also involved in decomposing organic matter, inhibiting pathogenic organisms, and participating in many soil biogeochemical cycles [77]. Furthermore, the relative abundance of Acidobacteria reduced, which could prove advantageous in mitigating Cd activation, as Acidobacteria are capable of mobilizing Cd by producing acidic compounds [78].
Interestingly, the relative abundance of Pseudarthrobacter, Sphingomonas, and Pseudomonas was significantly higher in the 3%B, 1%FB, and 3%FB treatments (Figure 3). Pseudarthrobacter contains various plant growth-promoting bacteria, and it can also effectively remove soil pollutants [79,80]. Sphingomonas and Pseudomonas have been found to play crucial roles as plant rhizosphere bacteria, enhancing plant growth by fixing nitrogen, suppressing pathogen growth, and producing plant hormones [81,82]. These bacteria are also known for their ability to solubilize insoluble P and K, making these nutrients available to plants [51,83]. This was consistent with the increased content of nutrients such as AP and AK in the biochar-amended soils. Moreover, Sphingomonas and Pseudomonas could immobilize Cd by producing biofilm, extracellular polymeric substances, and arginine decarboxylase [84,85]. Thus, it seemed that the application of biochar at higher doses, or the use of Fe-modified biochar, could enhance the population of bacteria that promote plant growth and immobilize Cd.
Furthermore, Spearman’s correlation analysis revealed that the relative abundance of Pseudomonas was significantly positively correlated with OX-/CB-Cd and negatively correlated with EX-/DTPA-Cd (Figure 4). This suggested that Pseudomonas may be involved in immobilizing Cd by transforming available Cd into unavailable forms. Xu et al. (2012) has discovered that Pseudomonas putida X4 can transform 7.5-24.8% of exchangeable Cd primarily into the organic-bound fraction.
Cd toxicity can induce oxidative stress in cells, increase ROS levels, degrade the antioxidative defense, reduce chlorophyll, and impair photosynthesis in plants, thereby decreasing plant yield [88]. Biochar is considered to play a crucial role in enhancing plant growth and mitigating Cd toxic impacts by immobilization of Cd in soil [89][90]. Liu et al. (2022) indicated that pig manure, sewage sludge, and corn straw biochars increased the plant height and fresh weight by lettuce at 1% application rates. In this study, we found that the application of B and FB remarkably enhanced the biomass of lettuce and mitigated Cd stress (Table 2). The observed beneficial effects could potentially be due to improved soil qualities, such as increased organic matter, availability of NPK nutrients, activities of soil enzymes, and the amount of plant growth-promoting bacteria (see Table S2 and Figure 3). We also found the significant reduction in Cd uptake by lettuce in both B and FB treatments highlights the potential of biochar in soil remediation. This phenomenon can predominantly be attributed to the decreased availability and increased immobilization of soil Cd, as indicated by the results (Figure 2). It has been found that Cd content in the wheat and rice grains, and fruits of eggplant and tomato was reduced with the application of biochar, by reducing soil DTPA-Cd and EX-Cd [92,93,94,95]. Specifically, The higher effectiveness of FB in reducing Cd content in lettuce roots and leaves by 25-40% and 33-47%, respectively, compared to raw biochar, can be attributed to the additional properties imparted by iron modification. A recent study reported that Fe-modified biochar significantly reduced DTPA-Cd content more than pristine biochar and was found to be more effective at reducing Cd uptake by Chinese cabbage, due to its higher pH, additional minerals, and oxygen-containing functional groups [96]. The similar Cd reduction in lettuce plants treated with higher-dosage B (3%) and lower-dosage FB (1%) underscores the economic and practical benefits of using iron-modified biochar at lower concentrations. This is particularly important for efficiently remediating Cd-polluted soils, as it reduces the amount of biochar needed and the associated costs.
5. Conclusions
In this study, a novel functional corn straw biochar (FB) prepared through iron modification was used for the remediation of Cd-contaminated soil. We found that Fe was loaded onto the biochar as iron oxides, and FB had a larger specific surface area, more pore volume, OFGs, and π-conjugated aromatic structures compared to B. The application of B and FB improved soil CEC, nutrient availability (AP and AK), soil enzymatic activities, and plant growth-promoting bacteria linked to increased lettuce biomass. The addition of B and FB converted available Cd into unavailable Cd by increasing the pH and CEC of the soils, resulting in a significant decrease in the content of bioavailable Cd. As a result, the Cd content of lettuce was reduced. Interestingly, FB was found to be more effective than B in immobilizing Cd. These significantly positive effects were achieved by establishing that a low dose (1%FB) was comparable to a high dose (3%B) in terms of Cd immobilization, thus eliminating concerns over the high costs associated with the application of biochar. The main mechanisms involved were direct precipitation, complexation, cation-π interaction, and exchange reaction of Cd with FB, and indirectly increasing Cd-immobilizing bacteria. Overall, these findings suggest that iron-modified biochar is a more cost-effective and environmentally friendly technique for the remediation of Cd-contaminated soil and reducing the risk of Cd in agriculture. It is necessary to further study the long-term effects of this FB under field conditions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author on reasonable request.
Acknowledgments
The National Natural Science Foundation of China (No. 42007140), the Natural Science Foundation of Shandong Province (ZR2020QD121), the National Natural Science Foundation of China (No. 22176104), the Integration of Science and Education Program Foundation for the Talents by the Qilu University of Technology (2023RCK217), Shandong Province Technology Innovation Guidance Project (YDZX2023006).
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Hamid: Y.; Tang, L.; Hussain, B.; Usman, M.; Lin, Q.Rashid, M.S.;He, Z.; Yang, X. Organic soil additives for the remediation of cadmium contaminated soils and their impact on the soil-plant system: A review. Science of the Total Environment. 2020, 707: 136121.
- Haider,F.U.; Liqun, C.; Coulter, J.A.; Cheema, S.A.; Wu, J.; Zhang, R.; Wenjun, M.; Farooq, M.Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicology and Environmental Safety. 2021, 211: 111887. [CrossRef]
- Wang, C.C.; Zhang, Q.C.; Yan, C.A.; Tang, G.Y.; Zhang, M.Y.; Ma, L.Q.; Gu, R.H.; Xiang, P.Heavy metal(loid)s in agriculture soils, rice, and wheat across China: Status assessment and spatiotemporal analysis. Science of the Total Environment, 2023a. 882: 163361. [CrossRef]
- Xu, D.M.; Fu, R.B.; Wang, J.X.; Shi, Y.X.; Guo, X.P.Chemical stabilization remediation for heavy metals in contaminated soils on the latest decade: Available stabilizing materials and associated evaluation methods-A critical review. Journal of Cleaner Production. 2021. 321: 128730. [CrossRef]
- Gao, Y.; Zhang, J.; Chen, C.; Du, Y.; Teng, G.; Wu, Z.Functional biochar fabricated from waste red mud and corn straw in China for acidic dye wastewater treatment. Journal of Cleaner Production. 2021, 320: 128887. [CrossRef]
- Qianqian, M.; Haider, F.U.; Farooq, M.; Adeel, M.; Shakoor, N.; Jun, W.; Jiaying, X.; Wang, X.W.; Panjun, L.; Cai, L.Selenium treated foliage and biochar treated soil for improved lettuce (Lactuca sativa L.) growth in Cd-polluted soil. Journal of Cleaner Production. 2022, 335: 130267. [CrossRef]
- Wang, Y.M.; Liu, Q.; Li, M.; Yuan, X.Y.; Uchimiya, M.; Wang, S.W.; Zhang, Z.Y.; Ji, T.; Wang, Y.; Zhao, Y.Y.Rhizospheric pore-water content predicts the biochar-attenuated accumulation, translocation, and toxicity of cadmium to lettuce. Ecotoxicology and Environmental Safety. 2021b, 208: 111675. [CrossRef]
- Shaheen, S.M.; Mosa, A.; Niazi, N.K.; Antoniadis, V.; Shahid, M.; Song, H.; Kwon, E.E.; Rinklebe, J.Removal of toxic elements from aqueous environments using nano zero-valent iron- and iron oxide-modified biochar: a review. Biochar. 2022, 4: 24. [CrossRef]
- Qiao, H.; Zhang, S.; Liu, X.; Wang, L.; Zhu, L.; Wang, Y.Adsorption characteristics and mechanisms of Cd(II) from wastewater by modified chicken manure biochar. Environmental Science and Pollution Research. 2024, 31: 3800-3814. [CrossRef]
- Yuan, S.; Hong, M.; Li, H.; Ye, Z.; Gong, H.; Zhang, J.; Huang, Q.; Tan, Z.Contributions and mechanisms of components in modified biochar to adsorb cadmium in aqueous solution. Science of the Total Environment. 2020, 733: 139320. [CrossRef]
- Zhang, J.Y.; Zhou, H.; Gu, J.F.; Huang, F.; Yang, W.J.; Wang, S.L.; Yuan, T.Y.; Liao, B.H.Effects of nano-Fe3O4-modified biochar on iron plaque formation and Cd accumulation in rice (Oryza sativa L.). Environmental Pollution. 2020a, 260: 113970. [CrossRef]
- Algethami, J.S.; Irshad, M.K.; Javed, W.; Alhamami, M.A.; Ibrahim, M.Iron-modified biochar improves plant physiology, soil nutritional status and mitigates Pb and Cd-hazard in wheat (Triticum aestivum L.). Frontiers in Plant Science. 2023, 14. [CrossRef]
- Li, P.; Wang, A.; Du, W.; Mao, L.; Wei, Z.; Wang, S.; Yuan, H.; Ji, R.; Zhao, L.Insight into the interaction between Fe-based nanomaterials and maize (Zea mays) plants at metabolic level. Science of the Total Environment. 2020b, 738: 139795. [CrossRef]
- Li, J.; Qiu, Y.; Zhao, Q.; Chen, D.; Wu, Z.; Peng, A.; Niazi, N.K.; Trakal, L.; Sakrabani, R.; Gao, B.; Wang, H.; Wu, W.Lead and copper-induced hormetic effect and toxicity mechanisms in lettuce (Lactuca sativa L.) grown in a contaminated soil. Science of the Total Environment. 2020a, 741: 140440. [CrossRef]
- Zhao, H.; Huang, X.; Liu, F.; Hu, X.; Zhao, X.; Wang, L.; Gao, P.; Li, J.; Ji, P.Potential of a novel modified gangue amendment to reduce cadmium uptake in lettuce (Lactuca sativa L.). Journal of Hazardous Materials. 2021, 410: 124543. [CrossRef]
- Wang, X.; Dong, G.; Liu, X.; Zhang, S.; Li, C.; Lu, X.; Xia, T.Poly-γ-glutamic acid-producing bacteria reduced Cd uptake and effected the rhizosphere microbial communities of lettuce. Journal of Hazardous Materials. 2020, 398: 123146. [CrossRef]
- Hernández, Y.; Lobo, M.G.; González, M.Determination of vitamin C in tropical fruits: A comparative evaluation of methods. Food Chemistry. 2006, 96: 654-664. [CrossRef]
- Kayaalp, N.; Ersahin, M.E.; Ozgun, H.; Koyuncu, I.; Kinaci, C.A new approach for chemical oxygen demand (COD) measurement at high salinity and low organic matter samples. Environmental Science and Pollution Research. 2010, 17: 1547-1552. [CrossRef]
- He, K.; He, G.; Wang, C.; Zhang, H.; Xu, Y.; Wang, S.; Kong, Y.; Zhou, G.; Hu, R.Biochar amendment ameliorates soil properties and promotes Miscanthus growth in a coastal saline-alkali soil. Applied Soil Ecology. 2020, 155: 103674. [CrossRef]
- Hu, X.; Huang, X.; Zhao, H.; Liu, F.; Wang, L.; Zhao, X.; Gao, P.; Li, X.; Ji, P.Possibility of using modified fly ash and organic fertilizers for remediation of heavy-metal-contaminated soils. Journal of Cleaner Production. 2021, 284: 124713. [CrossRef]
- Tessier, A.; Campbell, PG.; Bisson, M.Sequential extraction procedure for the speciation of particulate trace metals. Analytical chemistry. 1979, 51: 844-851. [CrossRef]
- Bremner, J.M.; Keeney, D.R.Steam distillation methods for determination of ammonium, nitrate and nitrite. Analytica Chimica Acta. 1965, 32: 485-495. [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R.Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proceedings of the National Academy of Sciences. 2011, 108: 4516-4522. [CrossRef]
- Sun. R.; Ding, J.; Li, H.; Wang, X.; Li, W.; Li, K.; Ye, X.; Sun, S. Mitigating nitrate leaching in cropland by enhancing microbial nitrate transformation through the addition of liquid biogas slurry. Agriculture, Ecosystems & Environment. 2023, 345: 108324. [CrossRef]
- Sun, R.; Zhang, W.; Liu, Y.; Yun, W.; Luo, B.; Chai, R.; Zhang, C.; Xiang, X.; Su, X. Changes in phosphorus mobilization and community assembly of bacterial and fungal communities in rice rhizosphere under phosphate deficiency. Frontiers in Microbiology. 2022b, 13:953340. [CrossRef]
- Liu, D.; Tong, C. Bacterial community diversity of traditional fermented vegetables in China. LWT. 2017, 86: 40-48. [CrossRef]
- Wang, Q. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73:5261 - 5267. [CrossRef]
- Yang. T.; Xu, Y.; Huang, Q.; Sun, Y.; Liang, X.; Wang, L.; Qin, X.; Zhao, L. Adsorption characteristics and the removal mechanism of two novel Fe-Zn composite modified biochar for Cd(II) in water. Bioresource Technology. 2021b, 333: 125078. [CrossRef]
- Cantrell, K.B.; Hunt, P.G.; Uchimiya, M.; Novak, J.M. Impact of pyrolysis temperature and manure source on physicochemical characteristics of biochar. Bioresource Technology. 2012, 107: 419-428. [CrossRef]
- Wang, X.; Yu, H.Y.; Li, F.; Liu, T.; Wu, W.; Liu, C.; Liu, C.; Zhang, X. Enhanced immobilization of arsenic and cadmium in a paddy soil by combined applications of woody peat and Fe(NO3)3: Possible mechanisms and environmental implications. Science of the Total Environment. 2019b, 649: 535-543. [CrossRef]
- Yin, G.; Song, X.; Tao, L.; Sarkar, B.; Sarmah, A.K.; Zhang, W.; Lin, Q.; Xiao, R.; Liu, Q.; Wang, H. Novel Fe-Mn binary oxide-biochar as an adsorbent for removing Cd(II) from aqueous solutions. Chemical Engineering Journal. 2020, 389: 124465. [CrossRef]
- Al-Swadi, H.A.; Al-Farraj, A.S.; Al-Wabel, M.I.; Ahmad, M.; Usman, A.R.; Ahmad. J.; Mousa, M.A.; Rafique, M.I.; Impacts of kaolinite enrichment on biochar and hydrochar characterization, stability, toxicity, and maize germination and growth. Scientific Reports. 2024, 14: 1259.
- Zhang, Y.; Wang, Q.; Wang, X.; Xiong, R.; Fu, D.; He, C.; Lai, B.; Ma, J. Enhanced degradation of Bisphenol AF by Fe/Zn modified biochar/ferrate(VI): Performance and enhancement mechanism. Journal of Environmental Chemical Engineering. 2023, 11: 111582. [CrossRef]
- Wan, X.; Li, C.; Parikh, S.J. Simultaneous removal of arsenic, cadmium, and lead from soil by iron-modified magnetic biochar. Environmental Pollution. 2020, 261: 114157. [CrossRef]
- Zeng, B.; Xu, W.; Khan, S.B.; Wang, Y.; Zhang, J.; Yang, J.; Su, X.; Lin, Z. Preparation of sludge biochar rich in carboxyl/hydroxyl groups by quenching process and its excellent adsorption performance for Cr(VI). Chemosphere. 2021, 285: 131439. [CrossRef]
- Zhao, N.; Zhao, C.; Zhang, W.; Du, Y.; Hao, Z.; Zhang, J. Adsorption and coadsorption mechanisms of Cr(VI) and organic contaminants on H3PO4 treated biochar. Chemosphere. 2017, 186: 422-429. [CrossRef]
- Fu, H.; Ma, S.; Xu, S.; Duan, R.; Cheng, G.; Zhao, P. Hierarchically porous magnetic biochar as an efficient amendment for cadmium in water and soil: Performance and mechanism. Chemosphere, 2021. 281: 130990. [CrossRef]
- Xu, Y.; Luo, G.; He, S.; Deng, F.; Pang, Q.; Xu, Y.; Yao, H. Efficient removal of elemental mercury by magnetic chlorinated biochars derived from co-pyrolysis of Fe(NO3)3-laden wood and polyvinyl chloride waste. Fuel. 2019, 239: 982-990. [CrossRef]
- Yang, F.; Zhang, S.; Li, H.; Li, S.; Cheng, K.; Li, J.S.; Tsang, D.C.W. Corn straw-derived biochar impregnated with α-FeOOH nanorods for highly effective copper removal. Chemical Engineering Journal, 2018. 348: 191-201. [CrossRef]
- Zhou Z, Xu Z, Feng Q, Yao D, Yu J, Wang D, Liu Y, Zhou N, Zhong M (2018) Effect of pyrolysis condition on the adsorption mechanism of lead, cadmium and copper on tobacco stem biochar. Journal of Cleaner Production 187: 996-1005.
- Zhu, Y.; Yi, B.; Hu, H.; Zong, Z.; Chen, M.; Yuan, Q. The relationship of structure and organic matter adsorption characteristics by magnetic cattle manure biochar prepared at different pyrolysis temperatures. Journal of Environmental Chemical Engineering. 2020, 8: 104112. [CrossRef]
- Zhang, Y.; Zhao, L.; Yang, Y.; Sun, P. Fenton-like oxidation of antibiotic ornidazole using biochar-supported nanoscale zero-valent iron as heterogeneous hydrogen peroxide activator. International Journal of Environmental Research and Public Health .2020b, 17: 1324. [CrossRef]
- Zhang, X.; Li, Y.; Wu, M.; Pang, Y.; Hao, Z.; Hu, M.; Qiu, R.; Chen, Z. Enhanced adsorption of tetracycline by an iron and manganese oxides loaded biochar: Kinetics, mechanism and column adsorption. Bioresource Technology. 2021, 320: 124264. [CrossRef]
- Wang, P.; Ding, F.; Huang, Z.; Fu, Z.; Zhao, P.; Men, S. Adsorption behavior and mechanism of Cd (II) by modified coal-based humin. Environmental Technology & Innovation. 2021a, 23: 101699. [CrossRef]
- Deng, Y.; Huang, S.; Laird, D.A.; Wang, X.; Meng, Z. Adsorption behaviour and mechanisms of cadmium and nickel on rice straw biochars in single- and binary-metal systems. Chemosphere. 2019, 218: 308-318. [CrossRef]
- Teng, D.; Zhang, B.; Xu, G.; Wang, B.; Mao, K.; Wang, J.; Sun, J.; Feng, X.; Yang, Z.; Zhang, H. Efficient removal of Cd(II) from aqueous solution by pinecone biochar: Sorption performance and governing mechanisms. Environmental Pollution. 2020, 265: 115001. [CrossRef]
- Fan, J.; Cai, C.; Chi, H.; Reid, B.J.; Coulon, F.; Zhang, Y.; Hou, Y. Remediation of cadmium and lead polluted soil using thiol-modified biochar. Journal of Hazardous Materials. 2020, 388: 122037. [CrossRef]
- Li, Y.; Liu, X.; Zhang, P.; Wang, X.; Cao, Y.; Han, L. Qualitative and quantitative correlation of physicochemical characteristics and lead sorption behaviors of crop residue-derived chars. Bioresource Technology. 2018, 270: 545-553. [CrossRef]
- Awasthi. M.K.; Wang, Q.; Chen, H.; Liu, T.; Awasthi, S.K.; Duan, Y.; Varjani, S.; Pandey, A.; Zhang, Z. Role of compost biochar amendment on the (im)mobilization of cadmium and zinc for Chinese cabbage (Brassica rapa L.) from contaminated soil. Journal of Soils and Sediments. 2019, 19: 3883-3897. [CrossRef]
- Azhar, M.; Zia ur Rehman, M.; Ali, S.; Qayyum, M.F.; Naeem, A.; Ayub, M.A.; Anwar, M.; Iqbal, A.; Rizwan, M. Comparative effectiveness of different biochars and conventional organic materials on growth, photosynthesis and cadmium accumulation in cereals. Chemosphere. 2019, 227: 72-81. [CrossRef]
- You, X.; Yin, S.; Suo, F.; Xu, Z.; Chu, D.; Kong, Q.; Zhang, C.; Li, Y.; Liu, L. Biochar and fertilizer improved the growth and quality of the ice plant (Mesembryanthemum crystallinum L.) shoots in a coastal soil of Yellow River Delta, China. Science of the Total Environment. 2021, 775: 144893. [CrossRef]
- Gao, S.; DeLuca, T.H.; Cleveland, C.C. Biochar additions alter phosphorus and nitrogen availability in agricultural ecosystems: A meta-analysis. Science of the Total Environment. 2019, 654: 463-472. [CrossRef]
- Liu, Z.; Xu, N.; Cao, T.; An, Z.; Yang, X.; He, T.; Yang, T.; Meng, J. Maize straw is more effective than maize straw biochar at improving soil N availability and gross N transformation rate. European Journal of Soil Science, 2023. 74: 13403. [CrossRef]
- Ren, H.; Huang, B.; Fernández-García, V.; Miesel, J.; Yan, L. Biochar and Rhizobacteria amendments improve several soil properties and bacterial diversity, Microorganisms. 2020, 8: 502. [CrossRef]
- Tang, J.; Zhang, L.; Zhang, J.; Ren, L.; Zhou, Y.; Zheng, Y.; Luo, L.; Yang, Y.; Huang, H.; Chen, A. Physicochemical features, metal availability and enzyme activity in heavy metal-polluted soil remediated by biochar and compost. Science of the Total Environment. 2020, 701: 134751. [CrossRef]
- Sun, T.; Xu, Y.; Sun, Y.; Wang, L.; Liang, X.; Zheng, S. Cd immobilization and soil quality under Fe–modified biochar in weakly alkaline soil. Chemosphere. 2021, 280: 130606. [CrossRef]
- Si, T.; Yuan, R.; Qi, Y.; Zhang, Y.; Wang, Y.; Bian, R.; Liu, X.; Zhang, X.; Joseph, S.; Li, L.; Pan, G. Enhancing soil redox dynamics: Comparative effects of Fe-modified biochar (N–Fe and S–Fe) on Fe oxide transformation and Cd immobilization. Environmental Pollution. 2024, 347: 123636. [CrossRef]
- Yang, Q.; Yang, C.; Yu, H.; Zhao, Z.; Bai, Z. The addition of degradable chelating agents enhances maize phytoremediation efficiency in Cd-contaminated soils. Chemosphere. 2021, 269: 129373. [CrossRef]
- Sun, L.; Gong, P.; Sun, Y.; Qin, Q.; Song, K.; Ye, J.; Zhang, H.; Zhou, B.; Xue, Y. Modified chicken manure biochar enhanced the adsorption for Cd2+ in aqueous and immobilization of Cd in contaminated agricultural soil. Science of the Total Environment. 2022a, 851: 158252. [CrossRef]
- Yang, S.; Xiao, Q.; Li, B.; Zhou, T.; Cen, Q.; Liu, Z.; Zhou, Y. Insights into remediation of cadmium and lead contaminated-soil by Fe-Mn modified biochar. Journal of Environmental Chemical Engineering. 2024, 12: 112771. [CrossRef]
- Meng, J.; Tao, M.; Wang, L.; Liu, X.; Xu, J. Changes in heavy metal bioavailability and speciation from a Pb-Zn mining soil amended with biochars from co-pyrolysis of rice straw and swine manure. Science of the Total Environment. 2018, 633: 300-307. [CrossRef]
- Liu, Q.; Li, X.; Tang, J.; Zhou, Y.; Lin, Q.; Xiao, R.; Zhang, M. Characterization of goethite-fulvic acid composites and their impact on the immobility of Pb/Cd in soil. Chemosphere. 2019, 222: 556-563. [CrossRef]
- Moradi, N.; Karimi, A. Fe-Modified common reed biochar reduced cadmium (Cd) mobility and enhanced microbial activity in a contaminated calcareous soil. Journal of Soil Science and Plant Nutrition. 2021, 21: 329-340. [CrossRef]
- Yang, T.; Xu, Y.; Huang, Q.; Sun, Y.; Liang, X.; Wang, L.; Qin, X.; Zhao, L. An efficient biochar synthesized by iron-zinc modified corn straw for simultaneously immobilization Cd in acidic and alkaline soils. Environmental Pollution. 2021c, 291: 118129. [CrossRef]
- Herrmann, L.; Lesueur, D.; Robin, A.; Robain, H.; Wiriyakitnateekul, W.; Bräu, L. Impact of biochar application dose on soil microbial communities associated with rubber trees in North East Thailand. Science of the Total Environment. 2019, 689: 970-979. [CrossRef]
- Li, Q.; Song, X.; Yrjälä, K.; Li, Y.; Wu, J.; Qin, H. Biochar mitigates the effect of nitrogen deposition on soil bacterial community composition and enzyme activities in a Torreya grandis orchard. Forest Ecology and Management. 2020c, 457: 117717. [CrossRef]
- Xu, M.; Dai, W.; Zhao, Z.; Zheng, J.; Huang, F.; Mei, C.; Huang, S.; Liu, C.; Wang, P.; Xiao, R. Effect of rice straw biochar on three different levels of Cd-contaminated soils: Cd availability, soil properties, and microbial communities. Chemosphere. 2022, 301: 134551. [CrossRef]
- Liu, Q.; Sheng, Y.; Wang, W.; Liu, X. Efficacy and microbial responses of biochar-nanoscale zero-valent during in-situ remediation of Cd-contaminated sediment. Journal of Cleaner Production. 2021, 287: 125076. [CrossRef]
- Whitman, T.; Pepe-Ranney, C.; Enders, A.; Koechli, C.; Campbell, A.; Buckley, D.H.; Lehmann, J. Dynamics of microbial community composition and soil organic carbon mineralization in soil following addition of pyrogenic and fresh organic matter. The ISME Journal. 2016, 10: 2918-2930. [CrossRef]
- Islam, M.S.; Zhu, J.; Xiao, L.; Khan, Z.H.; Saqib, H.; Gao, M.; Song, Z. Enhancing rice quality and productivity: Multifunctional biochar for arsenic, cadmium, and bacterial control in paddy soil. Chemosphere. 2023, 342: 140157. [CrossRef]
- Chen, Z.; Pei, J.; Wei, Z.; Ruan, X.; Hua, Y.; Xu, W.; Zhang, C.; Liu, T.; Guo, Y. A novel maize biochar-based compound fertilizer for immobilizing cadmium and improving soil quality and maize growth. Environmental Pollution. 2021, 277: 116455. [CrossRef]
- Huang, D.; Liu, L.; Zeng, G.; Xu, P.; Huang, C.; Deng, L.; Wang, R.; Wan, J. The effects of rice straw biochar on indigenous microbial community and enzymes activity in heavy metal-contaminated sediment. Chemosphere. 2017, 174: 545-553. [CrossRef]
- Liu, H.; Wang, C.; Xie, Y.; Luo, Y.; Sheng, M.; Xu, F.; Xu, H. Ecological responses of soil microbial abundance and diversity to cadmium and soil properties in farmland around an enterprise-intensive region. Journal of Hazardous Materials. 2020, 392: 122478. [CrossRef]
- Alvarez, A.; Saez, J.M.; Davila Costa, J.S.; Colin, V.L.; Fuentes, M.S.; Cuozzo, S.A.; Benimeli, C.S.; Polti, M.A.; Amoroso, M.J. Actinobacteria: Current research and perspectives for bioremediation of pesticides and heavy metals. Chemosphere. 2017, 166: 41-62. [CrossRef]
- Sazykin, I.; Khmelevtsova, L.; Azhogina, T.; Sazykina, M. Heavy metals influence on the bacterial community of soils: A review, Agriculture. 2023, 13: 653. ttps://doi.org/10.3390/agriculture13030653.
- Fan, Q.; Chen, Y.; Xu, R.; Guo, Z. Characterization of keystone taxa and microbial metabolic potentials in copper tailing soils. Environmental Science and Pollution Research. 2023, 30: 1216-1230. [CrossRef]
- Fuke, P.; Kumar, M.; Sawarkar, A.D.; Pandey, A.; Singh, L. Role of microbial diversity to influence the growth and environmental remediation capacity of bamboo: A review. Industrial Crops and Products. 2021, 167: 113567. [CrossRef]
- Yuan, B.; Huang, L.; Liu, X.; Bai, L.; Liu, H.; Jiang, H.; Zhu, P.; Xiao, Y.; Geng, J.; Liu, Q.; Hao, X. Application of mixotrophic acidophiles for the bioremediation of cadmium-contaminated soils elevates cadmium removal, soil nutrient availability, and rice growth. Ecotoxicology and Environmental Safety. 2022, 236: 113499. [CrossRef]
- Ham, S.H.; Yoon, A.R.; Park, Y.G. Plant growth-promoting microorganism Pseudarthrobacter sp. NIBRBAC000502770 enhances the growth and flavonoid content of Geum aleppicum, Microorganisms. 2022, 10: 1241.
- Li, J.; Peng, W.; Yin, X.; Wang, X.; Liu, Z.; Liu, Q.; Deng, Z.; Lin, S.; Liang, R. Identification of an efficient phenanthrene-degrading Pseudarthrobacter sp. L1SW and characterization of its metabolites and catabolic pathway. Journal of Hazardous Materials. 2024, 465: 133138. [CrossRef]
- Al-Karablieh, N.; Al-Shomali, I.; Al-Elaumi, L.; Hasan, K. Pseudomonas fluorescens NK4 siderophore promotes plant growth and biocontrol in cucumber. Journal of Applied Microbiology. 2022, 133: 1414-1421. [CrossRef]
- Asaf, S.; Numan, M.; Khan, A.L.; Al-Harrasi, A. Sphingomonas: from diversity and genomics to functional role in environmental remediation and plant growth. Critical Reviews in Biotechnology. 2020, 40: 138-152. [CrossRef]
- Wang, G.; Govinden, R.; Chenia, H.Y.; Ma, Y.; Guo, D.; Ren, G. Suppression of Phytophthora blight of pepper by biochar amendment is associated with improved soil bacterial properties. Biology and Fertility of Soils. 2019a, 55: 813-824. [CrossRef]
- Chellaiah, E.R. Cadmium (heavy metals) bioremediation by Pseudomonas aeruginosa: a minireview. Applied Water Science. 2018, 8: 154. [CrossRef]
- Cheng. C.; Wang, R.; Sun, L.; He, L.; Sheng, X. Cadmium-resistant and arginine decarboxylase-producing endophytic Sphingomonas sp. C40 decreases cadmium accumulation in host rice (Oryza sativa Cliangyou 513). Chemosphere. 2021, 275: 130109. [CrossRef]
- Ding, Y.; Xiong, J.; Zhou, B.; Wei, J.; Qian, A.; Zhang, H.; Zhu, W.; Zhu, J. Odor removal by and microbial community in the enhanced landfill cover materials containing biochar-added sludge compost under different operating parameters. Waste Management. 2019, 87: 679-690. [CrossRef]
- Xu, X.; Huang, Q.; Huang, Q.; Chen, W. Soil microbial augmentation by an EGFP-tagged Pseudomonas putida X4 to reduce phytoavailable cadmium. International Biodeterioration & Biodegradation. 2012, 71: 55-60.
- Awan, S.A.; Khan, I.; Rizwan, M.; Irshad, M.A.; Xiaosan, W.; Zhang, X.; Huang, L. Reduction in the cadmium (Cd) accumulation and toxicity in pearl millet (Pennisetum glaucum L.) by regulating physio-biochemical and antioxidant defense system via soil and foliar application of melatonin. Environmental Pollution. 2023, 328: 121658. [CrossRef]
- Irshad,M.K.; Noman, A.; Alhaithloul, H.A.; Adeel, M.; Rui, Y.; Shah, T.; Zhu, S.; Shang, J. Goethite-modified biochar ameliorates the growth of rice (Oryza sativa L.) plants by suppressing Cd and As-induced oxidative stress in Cd and As co-contaminated paddy soil. Science of the Total Environment. 2020, 717: 137086. [CrossRef]
- Kamran. M.; Malik, Z.; Parveen, A.; Zong, Y.; Abbasim G,H,l Rafiqm M,T.; Shaaban, M.; Mustafa, A.; Bashir, S.; Rafay, M.; Mehmood, S.; Ali, M. Biochar alleviates Cd phytotoxicity by minimizing bioavailability and oxidative stress in pakchoi (Brassica chinensis L.) cultivated in Cd-polluted soil. Journal of Environmental Management. 2019, 250: 109500.
- Liu, Q.; Huang, L.; Chen, Z.; Wen, Z.; Ma, L.; Wu, Y.; Liu, Y.; Feng, Y. Biochar and its combination with inorganic or organic amendment on growth, uptake and accumulation of cadmium on lettuce. Journal of Cleaner Production. 2022, 370: 133610. [CrossRef]
- Ghani, M.I.; Ahanger ,M.A.; Sial, T.A.; Haider, S.; Siddique, J.A.; Fan, R.; Liu, Y.; Ali, E.F.; Kumar, M.; Yang, X.; Rinklebe, J.; Chen, X.; Lee, S.S.; Shaheen, S.M. Almond shell-derived biochar decreased toxic metals bioavailability and uptake by tomato and enhanced the antioxidant system and microbial community. Science of the Total Environment. 2024, 929: 172632. [CrossRef]
- Jing. F.; Chen, C.; Chen, X.; Liu, W.; Wen, X.; Hu, S.; Yang, Z.; Guo, B.; Xu, Y.; Yu, Q. Effects of wheat straw derived biochar on cadmium availability in a paddy soil and its accumulation in rice. Environmental Pollution. 2020, 257: 113592. [CrossRef]
- Li, Z.; Q,i X.; Fan, X.; Du, Z.; Hu, C.; Zhao, Z.; Liu, Y. Amending the seedling bed of eggplant with biochar can further immobilize Cd in contaminated soils. Science of the Total Environment. 2016, 572: 626-633. [CrossRef]
- Wang, Q.; Duan, C.; Liang, H.X.; Ren, J.W.; Geng, Z.C.; Xu, C.Y. Phosphorus acquisition strategies of wheat are related to biochar types added in cadmium-contaminated soil: Evidence from soil zymography and root morphology. Science of the Total Environment. 2023b, 856: 159033. [CrossRef]
- Da, Y.; Xu, M.; Ma, J.; Gao, P.; Zhang, X.; Yang, G.; Wu, J.; Song, C.; Long, L.; Chen, C. Remediation of cadmium contaminated soil using K2FeO4 modified vinasse biochar. Ecotoxicology and Environmental Safety. 2023, 262: 115171. [CrossRef]
Figure 1.
(A): FTIR spectrums of three different types of biochar; (B): XPS of a full scan spectrum of three different types of biochar; (C): Fe 2p XPS spectra of FB before and after adsorption of Cd2+; (D): Cd 3d XPS spectra of FB after adsorption of Cd2+; (E): C 1s XPS spectra of three different types of biochar; (F): O1s XPS spectra of three different types of biochar.
Figure 1.
(A): FTIR spectrums of three different types of biochar; (B): XPS of a full scan spectrum of three different types of biochar; (C): Fe 2p XPS spectra of FB before and after adsorption of Cd2+; (D): Cd 3d XPS spectra of FB after adsorption of Cd2+; (E): C 1s XPS spectra of three different types of biochar; (F): O1s XPS spectra of three different types of biochar.
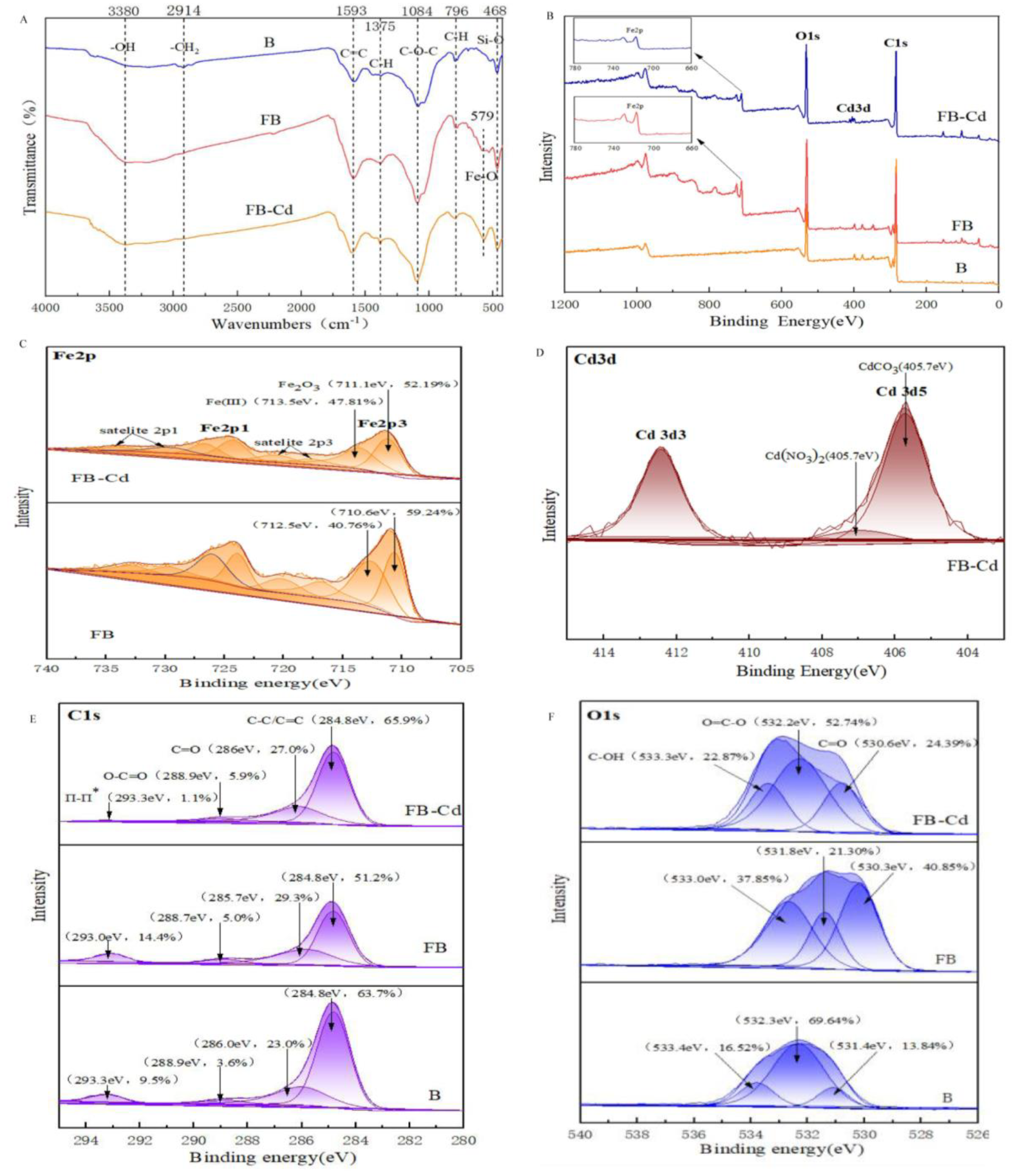
Figure 2.
The influences of different types of biochar on soil available Cd content (A) and Cd fractionation (B). EX-Cd, exchangeable Cd; CB-Cd, carbonate bound Cd; OX-Cd, Fe/Mn oxide bound Cd; OM-Cd, organic matter bound Cd; RS-Cd, residual Cd. Bars (n = 3) with different letters indicate significant differences at P < 0.05 according to the Dunn test.
Figure 2.
The influences of different types of biochar on soil available Cd content (A) and Cd fractionation (B). EX-Cd, exchangeable Cd; CB-Cd, carbonate bound Cd; OX-Cd, Fe/Mn oxide bound Cd; OM-Cd, organic matter bound Cd; RS-Cd, residual Cd. Bars (n = 3) with different letters indicate significant differences at P < 0.05 according to the Dunn test.
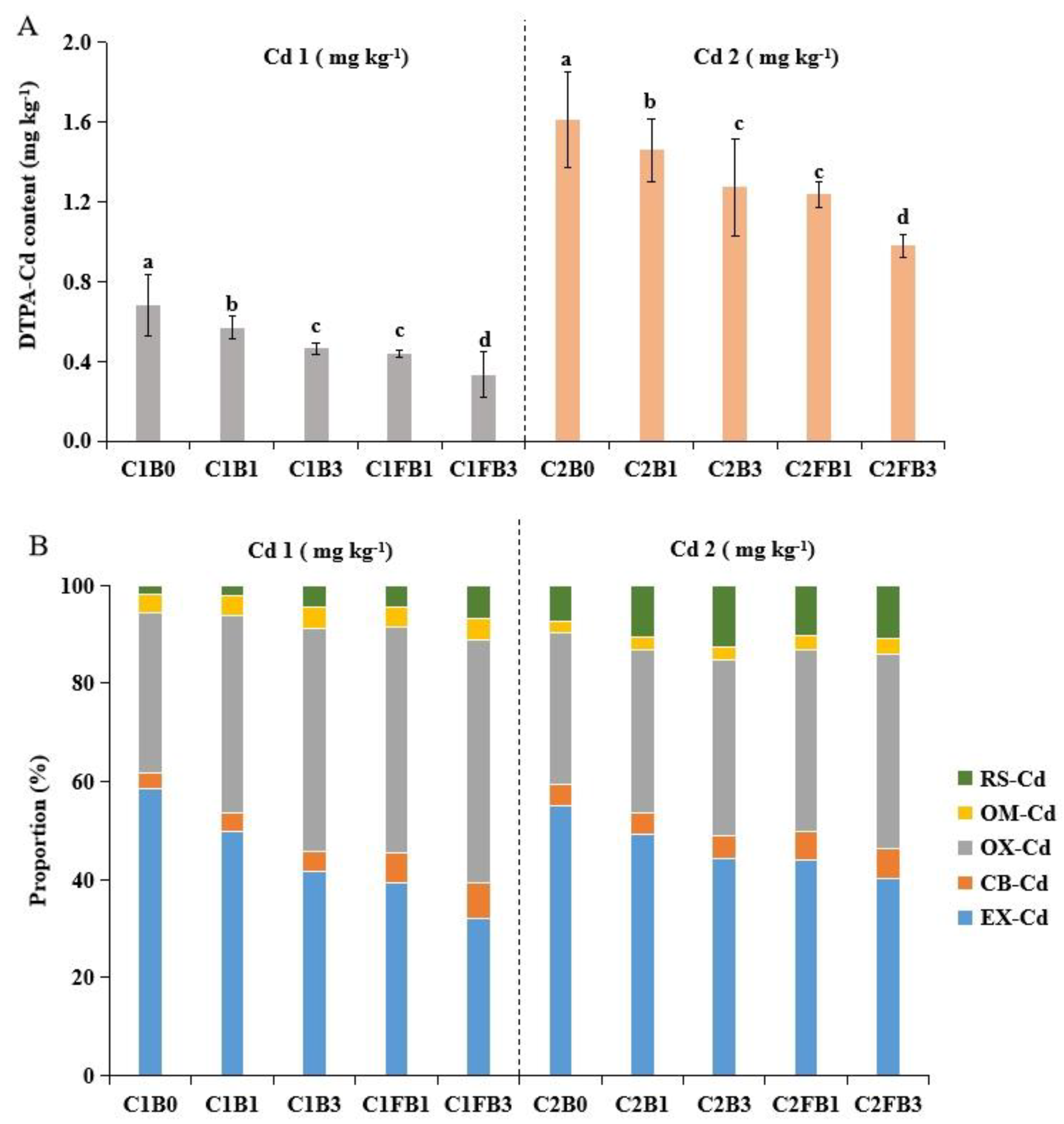
Figure 3.
The influences of different types of biochar on the relative abundances of the top 3 bacterial genera: Pseudarthrobacter (A), Sphingomonas (B), Pseudomonas (C). Bars (n = 3) with different letters indicate significant differences at P < 0.05 according to the Dunn test.
Figure 3.
The influences of different types of biochar on the relative abundances of the top 3 bacterial genera: Pseudarthrobacter (A), Sphingomonas (B), Pseudomonas (C). Bars (n = 3) with different letters indicate significant differences at P < 0.05 according to the Dunn test.
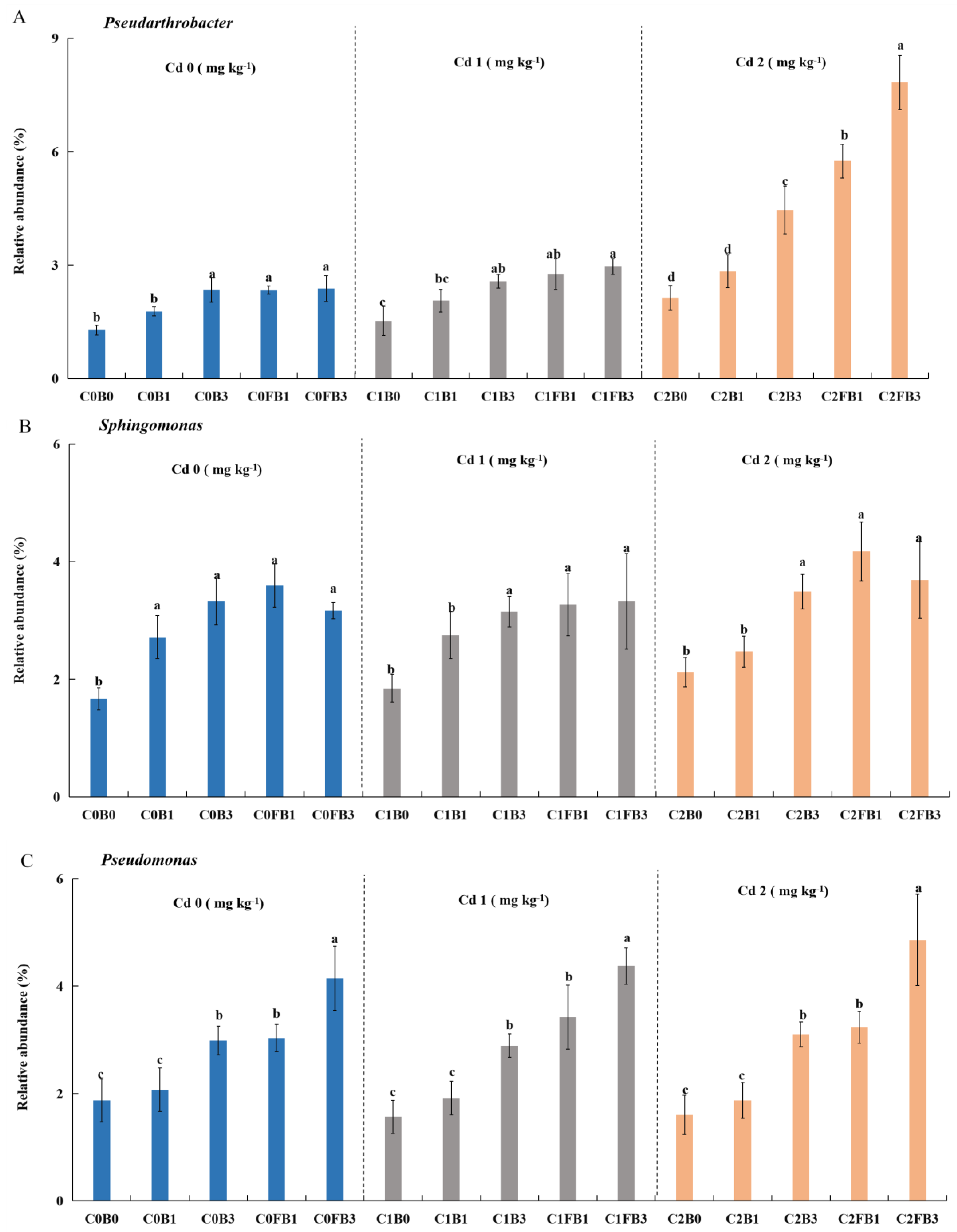
Figure 4.
Spearman correlation heatmaps reveal the correlations of plant biomass, soil properties, Cd fractions, and the relative abundances of top 10 dominant phyla (A and B) and top 15 dominant genera (C and D) of bacteria. The color key for correlation values is illustrated on the right panel inset. Positive correlations are shown in red text, and negative correlations are shown in blue (* 0.01 ≤ P < 0.05, **0.001 ≤ P < 0.01).
Figure 4.
Spearman correlation heatmaps reveal the correlations of plant biomass, soil properties, Cd fractions, and the relative abundances of top 10 dominant phyla (A and B) and top 15 dominant genera (C and D) of bacteria. The color key for correlation values is illustrated on the right panel inset. Positive correlations are shown in red text, and negative correlations are shown in blue (* 0.01 ≤ P < 0.05, **0.001 ≤ P < 0.01).
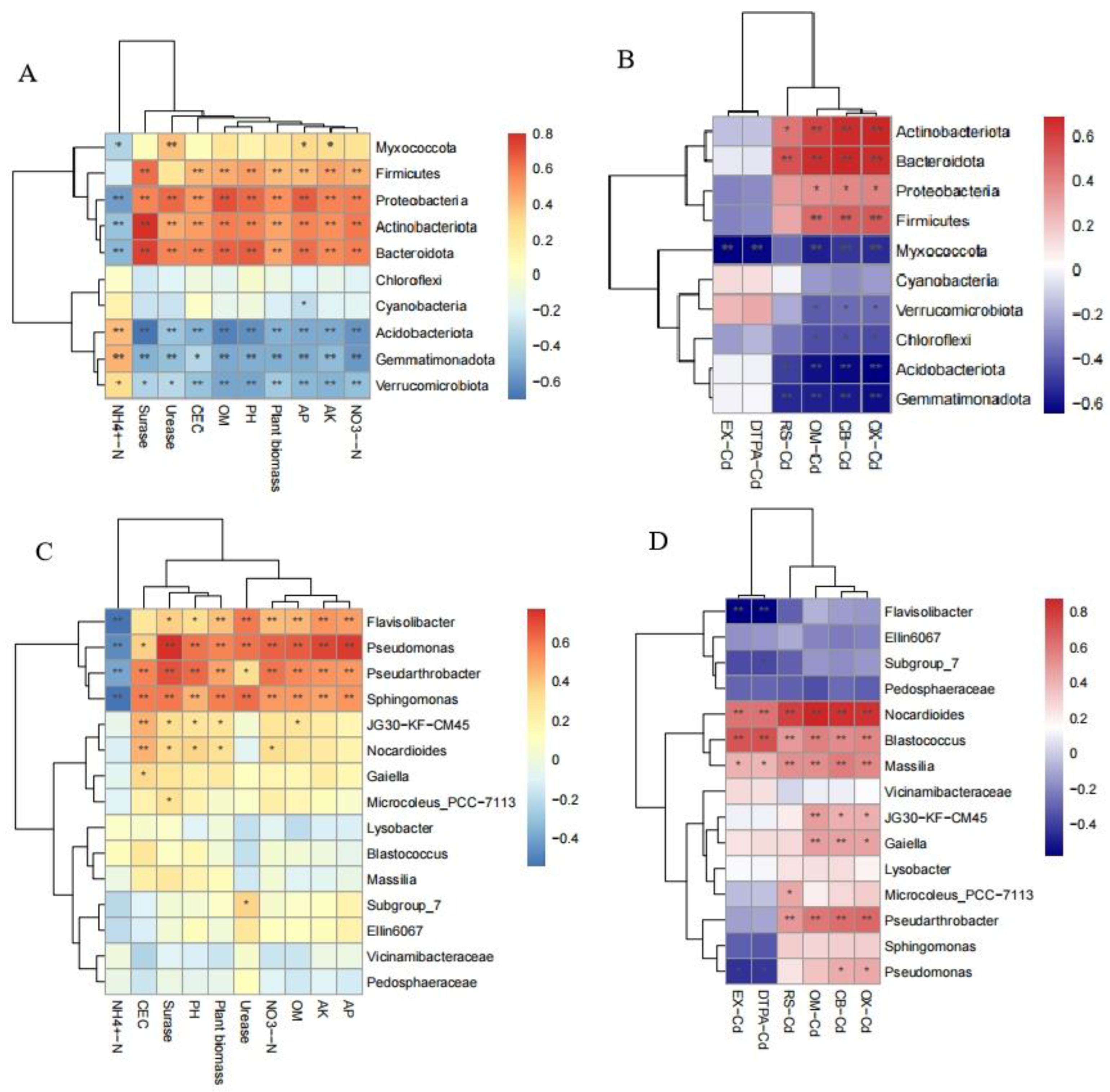
Figure 5.
The influences of different types of biochar on root (A) and leaf (B) Cd content of lettuce. Bars (n = 3) with different letters indicate significant differences at P < 0.05 according to the Dunn test.
Figure 5.
The influences of different types of biochar on root (A) and leaf (B) Cd content of lettuce. Bars (n = 3) with different letters indicate significant differences at P < 0.05 according to the Dunn test.
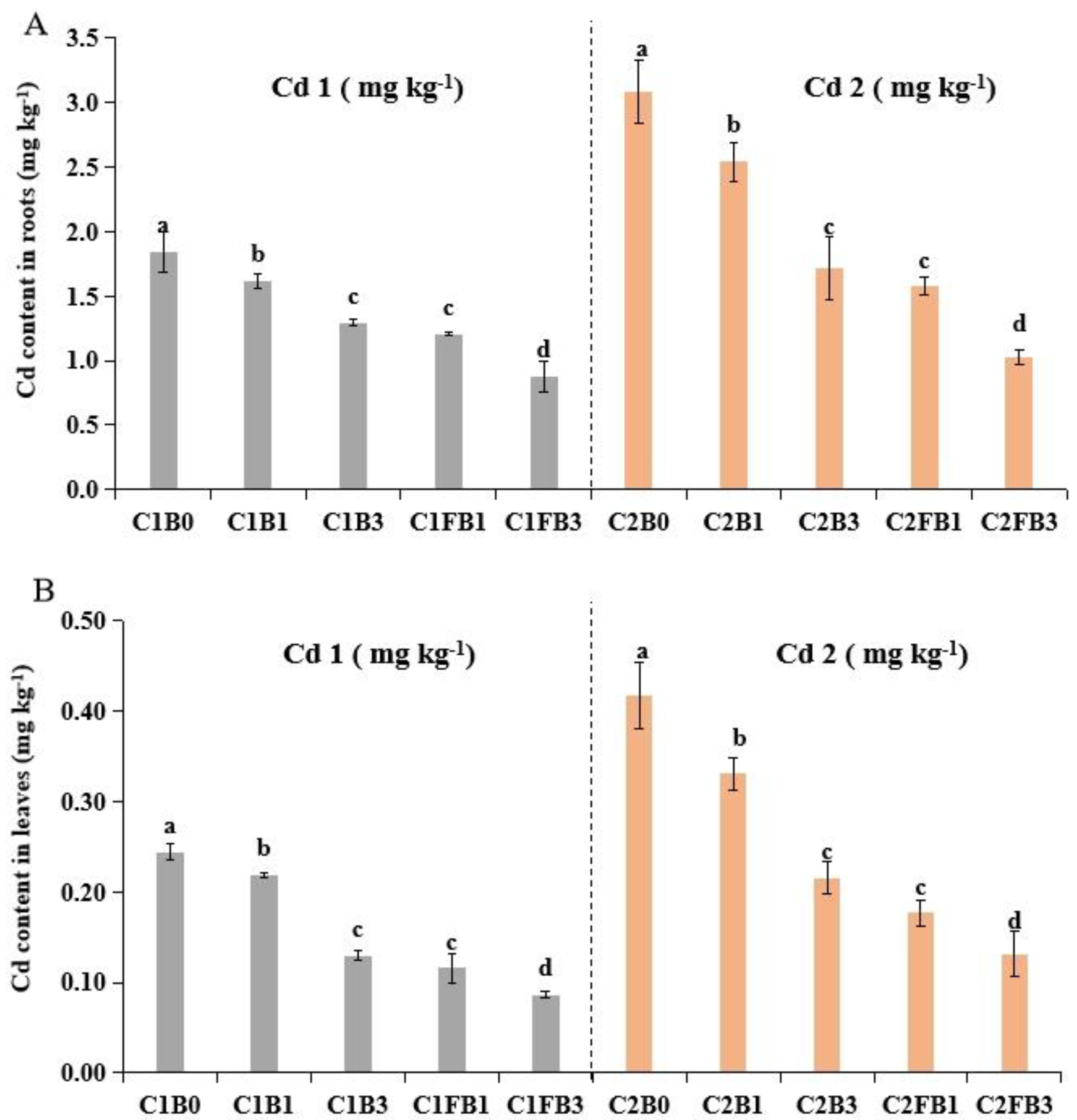

Table 1.
The basic physicochemical characteristics of different biochar materials.
| Adsorbents | Yield (%) | Ash (%) | pH | ABET (m2 g-1) |
Vtot (cm3 g-1) |
Dapd (nm) |
Elemental Content (%) | Atomicratio(%) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | H | O | N | S | H/C | O/C | |||||||
| BC | 51.6% | 20.7% | 9.45 | 7.17 | 0.0084 | 30.91 | 49 | 2.82 | 12.99 | 1.44 | 0.18 | 0.06 | 0.27 |
| FB | 93.7% | 36.8% | 10.48 | 17.53 | 0.0506 | 16.03 | 26.15 | 1.15 | 19.34 | 1.82 | 0.15 | 0.04 | 0.74 |
Table 2.
Effects of different treatments on the biomass, VC and MDA contents, POD and SOD activities of lettuce.
Table 2.
Effects of different treatments on the biomass, VC and MDA contents, POD and SOD activities of lettuce.
| Treatments | Lettuce Leaves Dry Weight (g) | Lettuce Roots Dry Weight (g) | VC (mg g-1) |
MDA (μmol g-1 ) |
POD (U mg-1) |
SOD (U mg-1) |
|---|---|---|---|---|---|---|
| Cd 0 | ||||||
| C0B0 | 0.82±0.14b | 0.27±0.03b | 115.5±4.82c | 6.33±0.26a | 0.399±0.029b | 0.249±0.010b |
| C0B1 | 1.25±0.25a | 0.35±0.02a | 137.5±12.35c | 5.70±0.44a | 0.479±0.036a | 0.333±0.053a |
| C0B3 | 1.33±0.14a | 0.35±0.02a | 185.2±17.18b | 4.98±0.70b | 0.549±0.061a | 0.374±0.044a |
| C0FB1 | 1.29±0.20a | 0.36±0.02a | 177.9±19.24b | 4.73±0.51b | 0.523±0.053a | 0.359±0.018a |
| C0FB3 | 1.36±0.26a | 0.39±0.03a | 221.8±19.81a | 5.03±0.59b | 0.535±0.095a | 0.328±0.048a |
| Cd 1 | ||||||
| C1B0 | 0.80±0.14b | 0.25±0.04b | 108.1±12.17c | 6.52±0.51a | 0.364±0.055b | 0.246±0.005b |
| C1B1 | 1.28±0.24a | 0.36±0.03a | 131.0±21.55c | 5.34±0.24a | 0.462±0.045a | 0.338±0.043a |
| C1B3 | 1.37±0.20a | 0.37±0.04a | 177.8±11.37b | 4.63±0.53b | 0.562±0.077a | 0.347±0.019a |
| C1FB1 | 1.36±0.23a | 0.38±0.03a | 184.2±12.82b | 4.85±0.40b | 0.511±0.053a | 0.345±0.055a |
| C1FB3 | 1.43±0.25a | 0.38±0.06a | 228.2±17.91a | 4.47±0.20b | 0.568±0.073a | 0.343±0.070a |
| Cd 2 | ||||||
| C2B0 | 0.72±0.11b | 0.27±0.05b | 100.1±1.40c | 8.33±0.32a | 0.343±0.010b | 0.232±0.009b |
| C2B1 | 1.24±0.20a | 0.37±0.05a | 122.6±7.89c | 5.62±0.97a | 0.420±0.037a | 0.346±0.041a |
| C2B3 | 1.36±0.17a | 0.37±0.01a | 163.5±23.08b | 4.70±0.33b | 0.459±0.073a | 0.379±0.029a |
| C2FB1 | 1.32±0.26a | 0.38±0.03a | 181.0±6.06a | 4.63±0.30b | 0.44±0.0260a | 0.351±0.045a |
| C2FB3 | 1.42±0.07a | 0.38±0.03a | 197.3±8.79a | 4.69±0.10b | 0.495±0.022a | 0.389±0.021a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



