
Submitted:
24 April 2025
Posted:
25 April 2025
You are already at the latest version
Abstract
Glutamatergic pathway abnormalities are increasingly recognised as central to various neuropsychiatric disorders, including autoimmune encephalitis, schizophrenia, and epilepsy, where autoantibodies targeting NMDA, AMPA, and GluR3 receptors are frequently implicated. In systemic lupus erythematosus (SLE), these autoantibodies, along with pro-inflammatory cytokines, are linked to neurocognitive impairments and mood disturbances. Elevated cytokine levels similarly contribute to the pathophysiology of epilepsy and other neurological conditions. Immunotherapies such as intravenous immunoglobulin (IVIG) have shown efficacy in mitigating some autoimmune neurological symptoms, though they offer limited benefit in disorders like amyotrophic lateral sclerosis and autism. Moreover, disruption of the blood-brain barrier—as observed in diseases such as multiple sclerosis and Guillain-Barré syndrome—permits pathogenic immune elements to infiltrate the central nervous system, exacerbating neuroinflammation. These insights highlight the urgent need for precise immunological screening and personalised treatment strategies to address immune-related neurological dysfunctions.
Keywords:
blood-brain barrier
; cytokines
; intravenous immunoglobulins
; neurological diseases
; antibodies
1. Introduction
Glutamatergic function deficits have been implicated in the development of various neuropsychiatric disorders, including schizophrenia. In autoimmune encephalitis, receptors and proteins involved in glutamatergic neurotransmission are often the antigenic targets. Specifically, the N-methyl-D-aspartate (NMDA) and alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors are implicated [1]. Glutamate receptor antibodies, such as anti-AMPA and anti-NMDA, are detectable in a range of neurological and autoimmune disorders, including systemic lupus erythematosus (SLE), Sjogren’s syndrome, schizophrenia, seizure disorders, and mania. These antibodies can downregulate cerebral functions, leading to brain damage and resulting in behavioural, psychiatric, and cognitive abnormalities in animal models. Notably, immunotherapy can reverse these effects in some patients [2,3].
Interestingly, autoantibodies to the “B” peptide (amino acids 372-395) of the glutamate/AMPA receptor subtype 3 (GluR3) have been found in the serum and cerebrospinal fluid of some patients with various types of epilepsy. However, no association has been found between the presence of these antibodies and patients suffering from epilepsy in conjunction with anti-phospholipid syndrome (APS) or Sneddon’s syndrome (SNS), both of which are autoimmune disorders [4]. These findings are significant because they suggest that some neurological disorders may have a basis in clinical immunology. Therefore, screening for specific autoimmune antibodies, immune cells, complement proteins, and cytokines may be necessary. If the autoimmune nature of these neurological or psychiatric problems is confirmed, treatment with immunotherapy, including intravenous immunoglobulins, immunosuppressors, plasmapheresis, rituximab, and cytostatic agents, could be considered. It is important to note that in SLE, neurological and psychiatric manifestations highlight the interrelation between the immune and central nervous systems [5].
The Biological Basis of Some Neuro-Psychiatric Diseases
The biological basis of depression in SLE has recently been corroborated. Biochemical and neurophysiological changes induced by cytokines have been demonstrated to contribute to the development of neuropsychiatric symptoms. Cytokines are known to cause mood swings and depression. The downregulation of the hypothalamic-pituitary-adrenal (HPA) axis correlates with neurophysiological changes involved in depression. Moreover, cerebro-reactive autoantibodies present in the cerebrospinal fluid (CSF), such as anti-NMDA and anti-ribosomal P, can cause significant neuronal damage. This damage is pertinent to mood and behaviour, leading to depressive symptoms [6]. Likewise, epilepsy is a complex and multifactorial condition. Increasing evidence suggests that the immune system plays a crucial role in neuronal excitability and epileptogenesis. Studies on epilepsy patients, including ex vivo investigations, have shown elevated levels of IL-1β, IL-2, IL-5, IL-6, and TNF-α following treatment with carbamazepine, valproic acid, and phenytoin [7].
In Table 1, the association between certain autoantibodies and specific central nervous system diseases is summarised, indicating that these conditions can be classified as autoimmune disorders. This classification has implications for managing these problems, as immunotherapy could be an effective treatment strategy. Table 2 outlines the involvement of cytokines in neuropsychiatric diseases, suggesting a pathogenic role for these molecules in the genesis of these conditions or as a result of interactions between the immune and/or endocrine systems and the brain.
In summary, the interrelationship between the immune system and the central nervous system is evident in various neuropsychiatric and neurological disorders. The presence of specific antibodies and cytokines underscores the potential autoimmune nature of these conditions. Screening for autoimmune markers and implementing immunotherapy may provide significant benefits in managing these disorders. Understanding the role of the immune system in these diseases offers a promising avenue for developing new therapeutic strategies and improving patient outcomes. The ongoing research in this field will undoubtedly continue to shed light on the complex mechanisms underlying these disorders and pave the way for more effective treatments.
Table 2 shows cytokine involvement in neuropsychiatric diseases such as Guillain-Barre syndrome and Schizophrenia.
2. Immunotherapy Used in Neurological Disorders
2.1. The Use of Intravenous Immunoglobulin (IVIG) in Treating Neurological Disorders
Intravenous immunoglobulin (IVIG) is a treatment option used for various neurological disorders. It involves infusing a preparation of immunoglobulins—antibodies extracted from the blood of healthy donors—directly into a patient’s bloodstream. While IVIG has been shown to be effective for certain conditions, the exact mechanisms by which it works are not yet fully understood. This essay explores the potential ways IVIG might benefit neurological conditions and discusses why it is not recommended for some specific disorders. Table 3 shows a list where the use of intravenous immunoglobulins (IVIG) has been used successfully.
2.2. How IVIG Works
IVIG has been observed to help with several neurological disorders, though its precise mechanisms are still being studied. One proposed mechanism is its ability to suppress the immune system. IVIG is thought to have immunosuppressive effects on various immune cells, including B cells, T cells, and antigen-presenting cells [34]. B cells produce antibodies, T cells are involved in directing the immune response, and antigen-presenting cells help other immune cells recognise and respond to foreign substances. By affecting these cells, IVIG may help reduce inappropriate immune reactions that can damage the nervous system. Another key function of IVIG is the down-regulation of complement proteins, which are part of the immune system’s response to infection and injury [35,36,37]. When these proteins are overactive, they can contribute to inflammation and damage in neurological disorders. IVIG may help control this activity, thereby reducing inflammation and associated damage.
IVIG may also influence the NF-kB pathway and the degradation of IκB [35,36,37]. NF-kB is a protein complex that plays a crucial role in regulating inflammation and immune responses. When NF-kB is activated, it can lead to increased inflammation. IVIG might help suppress this activation, thus reducing inflammation in the brain. Additionally, IVIG is believed to affect the idiotypic-anti-idiotypic network, which involves interactions between different antibodies [38]. This network helps regulate the immune response and may be disrupted in autoimmune disorders. By modulating this network, IVIG could help restore immune balance.
IVIG has also been shown to promote the expansion of T regulatory cells [39]. These cells are important for maintaining immune tolerance and preventing autoimmune responses. By increasing their numbers, IVIG might help the body regulate its immune response more effectively. Furthermore, IVIG can neutralise pathogenic autoantibodies [40,41]. In autoimmune disorders, these autoantibodies mistakenly target and damage the body’s own tissues. By neutralising these harmful antibodies, IVIG can potentially reduce tissue damage and improve symptoms.
In addition to these effects, IVIG may support remyelination, which is the process of repairing damaged myelin, the protective sheath around nerve fibres. This can be crucial for conditions where myelin is damaged, such as multiple sclerosis. IVIG might also affect the release of cytokines and cytokine antagonists, which are molecules that help regulate inflammation and immune responses [42,43,44]. By modulating these factors, IVIG could help manage inflammation and promote healing in the nervous system.
2.3. Conditions Treated with IVIG
IVIG has been shown to be effective in several neurological disorders. For instance, it is used in treating conditions like Guillain-Barré syndrome, a disorder where the immune system attacks the peripheral nerves, and chronic inflammatory demyelinating polyneuropathy (CIDP), a condition characterised by progressive weakness and sensory loss due to nerve damage. IVIG helps by modulating the immune response and reducing the inflammation that damages the nerves [35].
2.4. Conditions Where IVIG Is Not Recommended
Despite its benefits, IVIG is not suitable for every neurological disorder. For example, it is generally not recommended for conditions such as paraproteinemic neuropathy (IgM variant), intractable childhood epilepsy, inclusion body myositis, amyotrophic lateral sclerosis (ALS), adrenoleukodystrophy, autism, critical illness polyneuropathy, and POEMS syndrome [45].
Paraproteinemic neuropathy (IgM variant) is a condition where an abnormal protein in the blood causes nerve damage. Intractable childhood epilepsy involves severe and difficult-to-treat seizures in children. Inclusion body myositis is a muscle disorder that leads to progressive muscle weakness. ALS is a neurodegenerative disease that affects motor neurons, leading to muscle weakness and atrophy. Adrenoleukodystrophy is a genetic disorder affecting the brain and adrenal glands, and autism is a developmental disorder affecting communication and behaviour. Critical illness polyneuropathy occurs in patients who have been critically ill and involves nerve damage. POEMS syndrome is a rare condition characterised by polyneuropathy, organomegaly (enlarged organs), endocrinopathy (hormonal imbalances), oedema (swelling), M-protein (an abnormal protein), and skin abnormalities. In these conditions, IVIG may not be effective due to the specific nature of the disease processes involved. For example, in ALS and inclusion body myositis, the disease mechanisms may not be primarily immune-mediated, making IVIG less useful. Similarly, in conditions like autism, where the underlying pathophysiology is not primarily autoimmune or inflammatory, IVIG’s benefits may be limited. Summary, intravenous immunoglobulin (IVIG) is a valuable treatment for various neurological disorders, primarily due to its ability to modulate the immune response and reduce inflammation. Its effectiveness is attributed to several mechanisms, including immunosuppression, down-regulation of complement proteins, and promotion of T regulatory cells. However, IVIG is not recommended for all neurological conditions and also dermatological disorders [34,35,36,37,38,39,40,41,42,43,44].
3. Disruption of the Blood-Brain Barrier
The blood-brain barrier (BBB) is a sophisticated and selective structure that lines the capillaries throughout the brain, serving as a critical defence mechanism for maintaining the brain’s microenvironment. Under normal conditions, the BBB restricts the passage of large molecular weight substances such as proteins, thereby protecting the brain from potential harmful agents circulating in the blood [44. This barrier is essential for preserving the delicate balance of the brain’s internal environment and ensuring proper neuronal function. The blood-brain barrier (BBB) is an intricate and highly selective structure that plays a pivotal role in maintaining the brain’s homeostasis. This barrier comprises a layer of endothelial cells that line the capillaries throughout the brain, forming a protective shield between the bloodstream and the central nervous system (CNS). Its primary function is to regulate the movement of substances between the blood and the brain, ensuring that the neural environment remains stable and conducive to optimal neuronal function. Under normal physiological conditions, the BBB effectively restricts the passage of large molecular weight substances, such as proteins and other potentially harmful agents, from entering the brain [44].
This selective permeability is crucial for safeguarding the brain from toxins, pathogens, and fluctuations in blood composition that could disrupt neural activities. By limiting the entry of these substances, the BBB helps to maintain the brain’s delicate chemical balance, which is essential for proper neuronal function and overall cognitive health. The integrity of the BBB is maintained by tight junctions between endothelial cells, which form a nearly impermeable barrier. These tight junctions prevent the diffusion of large molecules and most cells across the endothelial layer, while allowing selective transport of essential nutrients and ions through specialised transport mechanisms [44].
This controlled permeability is vital for protecting the brain from potential damage and ensuring that only necessary substances, such as glucose and amino acids, can enter the CNS. Disruption of the BBB can have severe consequences for brain health. Conditions such as multiple sclerosis (MS) and neuropsychiatric systemic lupus erythematosus (NPSLE) are associated with BBB breakdown, which allows immune cells and autoantibodies to infiltrate the brain (Figure 1). This infiltration can lead to inflammation, tissue damage, and a range of neurological symptoms [45,46}.
Understanding the mechanisms that regulate BBB function and its disruption is crucial for developing effective treatments for various neurological and neurodegenerative disorders. In summary, the BBB is a fundamental component of brain protection, maintaining a stable environment for neural function and preventing the entry of potentially harmful substances. Its role in preserving brain homeostasis underscores its importance in both healthy and diseased states, highlighting the need for ongoing research into its mechanisms and potential therapeutic interventions [44].
However, a breach in the BBB can have significant implications for brain health. When the BBB is compromised, circulating antibodies and other immune components may infiltrate the brain. This infiltration is particularly problematic in the context of autoimmune disorders. For instance, in neuropsychiatric systemic lupus erythematosus (NSLE), a condition characterised by systemic autoimmune activity, antibodies that cross-react with neurological tissues can enter the brain. Once inside, these antibodies can interact with neuronal structures, leading to tissue damage and potentially contributing to neurotoxicity. The presence of autoantibodies and pro-inflammatory cytokines within the brain can initiate and exacerbate inflammatory responses. This inflammation is further intensified by the actions of T-helper 1 (TH1) and T-helper 17 (TH17) lymphocytes, which are known to amplify inflammatory reactions and contribute to neurodegeneration. Multiple sclerosis (MS) provides a clear example of how BBB disruption can lead to neurological damage. In MS, the BBB is disrupted, allowing myelin-specific lymphocytes to penetrate the central nervous system (CNS) [47].
These lymphocytes then target and induce demyelination, a process where the protective myelin sheath surrounding nerve fibres is damaged. This demyelination impairs the conduction of electrical signals along the affected nerves, leading to a range of neurological symptoms. The presence of gadolinium (gd)-enhancing lesions on magnetic resonance (MR) imaging is a hallmark of this disease, reflecting areas of active inflammation and BBB breakdown. When the BBB is compromised, immune and accessory cells that typically do not have access to the brain are able to breach this immunological sanctuary. The brain, normally considered an immunologically privileged organ, becomes exposed to these immune components, which can infiltrate the extracellular spaces and cause significant neuronal damage and neurotoxicity. Autoantibodies that specifically target neuronal cells or other components of the CNS can lead to various forms of neuroinflammation and tissue damage. Several autoimmune diseases are associated with such pathological processes. For example, in multiple sclerosis, as mentioned, the breakdown of the BBB allows for the infiltration of myelin-specific lymphocytes that attack and damage the myelin sheath, resulting in impaired nerve signal conduction. Neuromyelitis optica (NMO) is another autoimmune condition where autoantibodies against aquaporin-4, a water channel protein, can cross the compromised BBB and attack astrocytes, leading to severe demyelination and neuronal damage. Guillain-Barré syndrome (GBS) is a peripheral nerve disorder where molecular mimicry between bacterial or viral antigens and peripheral nerve components leads to an autoimmune attack on the myelin sheath. Chronic inflammatory demyelinating polyneuropathy (CIDP) also involves a similar autoimmune attack on the peripheral nerves, causing progressive weakness and sensory impairment due to demyelination. In all these conditions, the common feature is the breach of the BBB, which allows autoimmune factors to enter the CNS and cause damage. The ensuing inflammation and demyelination impair nerve function, leading to a range of neurological deficits. The damage to myelin disrupts the normal conduction of electrical impulses along the nerves, which can manifest as motor and sensory symptoms depending on the specific nerves affected [47,48,49]. Table 4 shows an overview of experimental therapies and their mechanisms of action in neurological autoimmune disorders.
4. Cytokines, Antibodies, and Autoimmune Inflammation
In the study, age-dependent differences in cytokine responses were observed in mice infected with Powassan virus (POWV). In 50-week-old mice, the infection triggered Th1-type cytokines (IFNγ, IL-2, IL-12, IL-4, TNFα, IL-6), indicating a pro-inflammatory, neurodegenerative response. Conversely, 10-week-old mice exhibited Th2-type cytokines (IL-10, TGFβ, IL-4), which are associated with a neuroprotective response. These findings highlight potential targets for therapeutic intervention [62].
This study explores the role of STAT4 in dendritic cell (DC) function, particularly in the context of experimental autoimmune encephalomyelitis (EAE), a model for multiple sclerosis. STAT4, which mediates the effects of proinflammatory cytokines like IL-12 and IL-23, is crucial for DC function. Mice lacking STAT4 in CD11c-expressing cells showed reduced T cell activation and central nervous system inflammation. Restoring EAE susceptibility through transfer of wild-type DCs, but not STAT4- or IL-23R-deficient DCs, indicates that STAT4 is essential for DC-mediated inflammation. Single-cell RNA-sequencing identified STAT4-dependent gene signatures in DCs, aligning with patterns seen in multiple sclerosis (MS) patients [63].
This study compared tocilizumab and conventional immunotherapy for treating refractory acetylcholine receptor antibody-positive (AChR-Ab+) generalized myasthenia gravis (gMG). Conducted at a single centre in China, it included 34 patients, with 20 receiving tocilizumab and 14 on conventional therapy. Tocilizumab resulted in a more significant reduction in the MG activities of daily living (MG-ADL) score at week 4, and these improvements were sustained through week 24. The tocilizumab group showed a higher proportion of patients achieving notable reductions in both MG-ADL and QMG scores. Tocilizumab was also found to be safe, with no severe or unexpected adverse effects reported. These results suggest that tocilizumab is a promising and effective treatment for refractory AChR-Ab+ gMG [64].
Multiple sclerosis (MS) is a chronic inflammatory disease of the central nervous system (CNS), affecting an estimated 2.8 million people globally [65]. The condition is thought to result from aberrant adaptive immune responses targeting myelin sheaths, though its exact causes remain unclear. Genetic factors, particularly the HLA-DR*15:01 allele, are strongly linked to MS, suggesting a role for antigen presentation and CD4+ T cell responses [66,67]. Despite this, no consistent autoantigenic targets have been identified, and while myelin-specific T cells are present in MS patients, their frequency and activity vary [68]. Epstein-Barr virus (EBV), a γ-Herpesvirus known for causing infectious mononucleosis (IM) and for its association with B cell tumours, has been implicated in MS pathogenesis. Evidence shows that a history of IM increases MS risk, and EBV seroconversion often precedes disease onset [69,70,71]. Elevated levels of EBNA1 IgG, an EBV latent cycle antigen, are linked to a significantly higher MS risk [72,73]. Moreover, antibodies against EBNA1 have been detected in oligoclonal bands of MS patients and show cross-reactivity with human proteins, suggesting a direct role in neuroimmune mechanisms [74,75,76,77]. The study aimed to expand understanding of EBV-related immune responses by analysing antibody responses to various EBV antigens, including EBNA1, other components of the EBNA complex, and the virus-capsid antigen (VCA), in plasma from individuals with relapsing-remitting MS (RRMS), healthy controls, and those with recent IM. It also investigated EBV-specific T cell responses, particularly cross-reactivity with CNS autoantigens [78,79,80,81]. This research may clarify the role of EBV in MS and identify potential targets for therapeutic intervention.
In autoimmune central nervous system (CNS) disorders, specific antibodies are associated with different clinical manifestations:
Limbic encephalitis is a condition features personality changes, memory deficits, seizures, and psychosis, with MRI changes in the mesial temporal lobes. Antineuronal nuclear antibody (ANNA)-1 and gamma-aminobutyric acid-B receptor (GABAB-R) antibodies are linked to small cell lung cancer (SCLC) and limbic encephalitis [82,83]. Leucine-rich glioma inactivated 1 (LGI1) antibodies are associated with autoimmune limbic encephalitis and faciobrachial dystonic seizures (FBDS), often with rare tumours [84,85]. Other phenotypic descriptions include autoimmune dementia, epilepsy, or movement disorders [86,87,88].
Brainstem encephalitis is characterised by oculomotor abnormalities and cranial nerve dysfunction, associated antibodies include kelch-like protein 11 (KLHL-11), ANNA-2 (anti-Ri), and IgLON5 IgG [89]. Bickerstaff’s Brainstem Encephalitis, linked with ganglioside Q1B (GQ1B) IgG, features multiple cranial nerve abnormalities [90]. IgLON5 autoimmunity may present with sleep disorders and severe cases can lead to cardiorespiratory dysfunction [91,92,93,94].
Extralimbic encephalitis is characterised by symptoms localised in non-limbic structures, anti-GABAA-R antibodies are linked with seizures and multifocal cerebral lesions [95]. Glial Fibrillary Acidic Protein (GFAP) astrocytopathy, affecting deep periventricular white matter, is associated with meningism, parkinsonism, and other CNS symptoms [96,97]. ANNA-1 antibodies can present with extralimbic symptoms, and N-methyl-d-aspartate receptor (NMDA-R) antibodies lead to psychiatric symptoms, psychosis, seizures, and autonomic instability [98,99].
Rapidly Progressive Cerebellar Ataxia is a syndrome often involves gait disorder and limb ataxia with a rapid onset. Antibodies include Purkinje cytoplasmic antibody type 1 (PCA-1, anti-Yo) and glutamic acid decarboxylase (GAD) 65 IgG. Immune therapies may result in stabilization or improvement, with poorer outcomes for antibodies against intracellular antigens, though GAD65 often responds well [100,101].
Autoimmune myelopathy involves ascending sensorimotor deficits and may show sensory levels and upper motor neuron signs. Key antibodies include myelin oligodendrocyte glycoprotein (MOG) and aquaporin-4 (AQP4). Optic neuritis and longitudinally extensive spinal cord lesions are common [102,103,104,105,106]. Autoimmune paraneoplastic myelopathies, such as those with amphiphysin and collapsin-responsive mediator protein (CRMP)-5 IgG, may show tract-specific lesions [107].
Opsoclonus-Myoclonus Syndrome (OMS) involves arrhythmic eye movements and myoclonus. In children, it is associated with neuroblastoma, while in adults, ANNA-2 IgG is most common [108]. Other antibodies such as NMDA-R, GABAB, GAD65, and dipeptidyl-peptidase-like protein (DPPX) are less frequent [109].
6. New approaches in the Treatment of Psychoneuroimmunological Disorders
Chimeric antigen receptor (CAR) T cell therapy targeting B cell maturation antigen (BCMA) shows potential in treating neuromyelitis optica spectrum disorder (NMOSD), a neurological autoimmune condition. This study analysed cerebrospinal fluid (CSF) and blood samples from NMOSD patients treated with anti-BCMA CAR T cells using single-cell multi-omics sequencing. The results highlighted a dominant role for proliferating CD8⁺ CAR T cells with cytotoxic-like profiles in counteracting autoimmunity. These engineered cells demonstrated enhanced chemotactic properties, enabling them to cross the blood-CSF barrier effectively, where they depleted plasmablasts and plasma cells, contributing to the reduction of neuroinflammation. A subset of CAR T cells expressing CD44, indicative of an early memory phenotype, was associated with prolonged cell persistence in patients. Interestingly, compared to CAR T cells used in blood cancers, those from NMOSD patients showed reduced cytotoxic characteristics. These findings advance understanding of CAR T cell behaviour in autoimmune neurological settings and may inform therapeutic improvements [110].
CAR-T cell therapy is an emerging immunotherapy that shows promise in the treatment of neuroimmune diseases. This approach modifies a patient’s T cells to target and eliminate specific immune cells involved in the disease process. Initial studies have shown encouraging results in conditions such as neuromyelitis optica spectrum disorder, where CAR-T cells have been able to cross the blood-brain barrier, reduce inflammation in the central nervous system, and eliminate harmful B cell populations. These findings suggest potential clinical benefits, including symptom relief and reduced disease activity. Despite these advances, the long-term implications of CAR-T therapy in neuroimmune disorders remain uncertain. Potential risks such as cytokine release syndrome, immune system suppression, and unintended neurological effects require further investigation. Additionally, questions around the ideal timing for treatment, long-term persistence of CAR-T cells in the nervous system, and patient selection criteria need to be addressed. More prospective studies with extended follow-up are necessary to determine the full safety and effectiveness of this therapy. In summary, CAR-T cell therapy may represent a new direction in managing neuroimmune diseases, but its routine use will depend on robust clinical evidence and continued monitoring of outcomes [111,112].
7. Conclusions
Intravenous immunoglobulin (IVIG) is a significant advancement in treating neurological disorders due to its ability to modulate immune responses and reduce inflammation. It is especially effective in conditions with autoimmune and inflammatory components. IVIG works by suppressing the activity of immune cells, down-regulating complement proteins, and influencing key signalling pathways like NF-kB, thereby alleviating inflammation. It also enhances the expansion of T regulatory cells, which help maintain immune tolerance, and neutralises pathogenic autoantibodies to reduce tissue damage. Its potential to promote remyelination benefits conditions characterised by demyelination, such as multiple sclerosis (MS). However, IVIG is not universally effective. It does not benefit conditions such as paraproteinemic neuropathy, intractable childhood epilepsy, inclusion body myositis, amyotrophic lateral sclerosis (ALS), adrenoleukodystrophy, autism, critical illness polyneuropathy, and POEMS syndrome, due to their distinct pathophysiological mechanisms. For instance, ALS involves motor neuron degeneration rather than immune-mediated attacks, making IVIG less applicable. The study of age-dependent cytokine responses in Powassan virus infection in mice reveals older mice exhibit a pro-inflammatory Th1 response while younger mice show a neuroprotective Th2 response, suggesting new therapeutic targets. Research into STAT4’s role in dendritic cells highlights its importance in mediating inflammation in multiple sclerosis models. A study comparing tocilizumab to conventional treatments for myasthenia gravis found tocilizumab more effective and safer. MS, a chronic CNS inflammatory disease, affects around 2.8 million people globally. Genetic factors, especially the HLA-DR*15:01 allele, are linked to MS, and Epstein-Barr virus (EBV) may play a role in its development. Autoimmune CNS disorders present various antibody profiles: limbic encephalitis with ANNA-1 and LGI1 antibodies; brainstem encephalitis with KLHL-11 and GQ1B antibodies; extralimbic encephalitis with anti-GABAA-R and GFAP antibodies; and rapidly progressive cerebellar ataxia with PCA-1 and GAD65 antibodies. CAR-T cell therapy is a novel avenue in the treatment of some neurological disorders.
Author Contributions
This manuscript was conceptualised by A.J.-V. and planning and discussions were conducted by all authors. All authors participated in writing the initial draft of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study did not receive any external funding.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
The dataset supporting the findings of this study is included within the manuscript and its referenced sources, ensuring comprehensive access to the relevant data for further examination and analysis.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kao, Y.C.; Lin, M.I.; Weng, W.C.; Lee, W.T. Neuropsychiatric disorders due to limbic encephalitis: Immunologic aspect. Int. J. Mol. Sci. 2020, 22, 389. [CrossRef]
- Taiwo, R.O.; Goldberg, H.S.; Ilouz, N.; Singh, P.K.; Shekh-Ahmad, T.; Levite, M. Enigmatic intractable Epilepsy patients have antibodies that bind glutamate receptor peptides, kill neurons, damage the brain, and cause Generalized Tonic Clonic Seizures. J. Neural Transm. 2025, 1–26. [CrossRef]
- Husari, K.S.; Dubey, D. Autoimmune epilepsy. Neurotherapeutics 2019, 16, 685–702.
- Levite, M.; Goldberg, H. Autoimmune epilepsy-novel multidisciplinary analysis, discoveries and insights. Front. Immunol. 2022, 12, 762743.
- Appenzeller, S.; Andrade, S.D.O.; Bombini, M.F.; Sepresse, S.R.; Reis, F.; França, M.C., Jr. Neuropsychiatric manifestations in primary Sjogren syndrome. Expert Rev. Clin. Immunol. 2022, 18, 1071–1081. [CrossRef]
- Dima, A.; Caraiola, S.; Delcea, C.; Ionescu, R.A.; Jurcut, C.; Badea, C. Self-reported disease severity in women with systemic lupus erythematosus. Rheumatol. Int. 2019, 39, 533–539. [CrossRef]
- Gremke, N.; Printz, M.; Möller, L.; Ehrenberg, C.; Kostev, K.; Kalder, M. Association between anti-seizure medication and the risk of lower urinary tract infection in patients with epilepsy. Epilepsy Behav. 2022, 135, 108910. [CrossRef]
- Kamyshna, I.I.; Pavlovych, L.B.; Kamyshnyi, A.M. Prediction of the cognitive impairment development in patients with autoimmune thyroiditis and hypothyroidism. Endocr. Regul 2022, 56, 178–189. [CrossRef]
- Manocchio, N.; Magro, V.M.; Massaro, L.; Sorbino, A.; Ljoka, C.; Foti, C. Hashimoto’s Encephalopathy: Clinical Features, Therapeutic Strategies, and Rehabilitation Approaches. Biomedicines 2025, 13, 726.
- Shojima, Y.; Nishioka, K.; Watanabe, M.; Jo, T.; Tanaka, K.; Takashima, H.; Nodayes, K.; Okuma, Y.; Urabe, T.; Yokoyama, K.; et al. Clinical characterization of definite autoimmune limbic encephalitis: A 30-case series. Intern. Med. 2019, 58, 3369–3378. [CrossRef]
- Mangnus, T.J.; Dirckx, M.; Huygen, F.J. Different types of pain in complex regional pain syndrome require a personalized treatment strategy. J. Pain Res. 2023, 4379–4391. [CrossRef]
- Dziadkowiak, E.; Moreira, H.; Buska-Mach, K.; Szmyrka, M.; Budrewicz, S.; Barg, E.; Janik, M.; Pokryszko-Dragan, A. Occult Autoimmune Background for Epilepsy—The Preliminary Study on Antibodies Against Neuronal Surface Antigens. Front. Neurol. 2021, 12, 660126. [CrossRef]
- Vasilevska, V.; Guest, P.C.; Schlaaff, K.; Incesoy, E.I.; Prüss, H.; Steiner, J. Potential cross-links of inflammation with schizophreniform and affective symptoms: A review and outlook on autoimmune encephalitis and COVID-19. Front. Psychiatry 2021, 12, 729868. [CrossRef]
- Murashko, A.A.; Pavlov, K.A.; Pavlova, O.V.; Gurina, O.I.; Shmukler, A. Antibodies against N-Methyl D-aspartate receptor in psychotic disorders: A systematic review. Neuropsychobiology 2022, 81, 1–18. [CrossRef]
- Zádor, F.; Nagy-Grócz, G.; Kekesi, G.; Dvorácskó, S.; Szűcs, E.; Tömböly, C.; Horvath, G.; Benyhe, S.; Vécsei, L. Kynurenines and the endocannabinoid system in schizophrenia: Common points and potential interactions. Molecules 2019, 24, 3709. [CrossRef]
- Howes, O.D.; Bukala, B.R.; Beck, K. Schizophrenia: From neurochemistry to circuits, symptoms and treatments. Nat. Rev. Neurol. 2024, 20, 22–35. [CrossRef]
- Kitanosono, H.; Motomura, M.; Tomita, H.; Iwanaga, H.; Iwanaga, N.; Irioka, T.; Shiraishi, H.; Tsujino, A.. Paraneoplastic cerebellar degeneration with lambert-eaton myasthenic syndrome: A report of an effectively treated case and systematic review of Japanese cases. Brain Nerve = Shinkei Kenkyu No Shinpo 2019, 71, 167–174.
- Takamori, M. Myasthenia gravis: From the viewpoint of pathogenicity focusing on acetylcholine receptor clustering, trans-synaptic homeostasis and synaptic stability. Front. Mol. Neurosci. 2020, 13, 86. [CrossRef]
- Justiz-Vaillant, A.A.; Gopaul, D.; Soodeen, S.; Arozarena-Fundora, R.; Barbosa, O.A.; Unakal, C.; Thompson, R.; Pandit, B.; Umakanthan, S.; Akpaka, P.E. Neuropsychiatric Systemic Lupus Erythematosus: Molecules Involved in Its Imunopathogenesis, Clinical Features, and Treatment. Molecules 2024, 29, 747. [CrossRef]
- Ferrazzano, G.; Crisafulli, S.G.; Baione, V.; Tartaglia, M.; Cortese, A.; Frontoni, M.; Altieri, M.; Pauri, F.; Millefiorini, E.; Conte, A. Early diagnosis of secondary progressive multiple sclerosis: Focus on fluid and neurophysiological biomarkers. J. Neurol. 2021, 268, 3626–3645. [CrossRef]
- Li, H.; Liu, S.; Han, J.; Li, S.; Gao, X.; Wang, M.; Zhu, J.; Jin, T. Role of toll-like receptors in neuroimmune diseases: Therapeutic targets and problems. Front. Immunol. 2021, 12, 777606. [CrossRef]
- Bryll, A.; Skrzypek, J.; Krzyściak, W.; Szelągowska, M.; Śmierciak, N.; Kozicz, T.; Popiela, T. Oxidative-antioxidant imbalance and impaired glucose metabolism in schizophrenia. Biomolecules 2020, 10, 384. [CrossRef]
- Gagné, A.M.; Moreau, I.; St-Amour, I.; Marquet, P.; Maziade, M. Retinal function anomalies in young offspring at genetic risk of schizophrenia and mood disorder: The meaning for the illness pathophysiology. Schizophr. Res. 2020, 219, 19–24. [CrossRef]
- Slavov, G. Changes in serum cytokine profile and deficit severity in patients with relapsing-remitting multiple sclerosis. Folia Medica 2023, 65, 625–630. [CrossRef]
- Velikova, T.; Sekulovski, M.; Bogdanova, S.; Vasilev, G.; Peshevska-Sekulovska, M.; Miteva, D.; Georgiev, T. Intravenous immunoglobulins as immunomodulators in autoimmune diseases and reproductive medicine. Antibodies 2023, 12, 20. [CrossRef]
- Conti, F.; Moratti, M.; Leonardi, L.; Catelli, A.; Bortolamedi, E.; Filice, E.; Fetta, A.; Fabi, M.; Facchini, E.; Cantarini, M.E.; et al. Anti-inflammatory and immunomodulatory effect of high-dose immunoglobulins in children: From approved indications to off-label use. Cells 2023, 12, 2417. [CrossRef]
- Manganotti, P.; Garascia, G.; Furlanis, G.; Buoite Stella, A. Efficacy of intravenous immunoglobulin (IVIg) on COVID-19-related neurological disorders over the last 2 years: An up-to-date narrative review. Front. Neurosci. 2023, 17, 1159929. [CrossRef]
- Bayry, J.; Ahmed, E.A.; Toscano-Rivero, D.; Vonniessen, N.; Genest, G.; Cohen, C.G.; Dembele, M.; Kaveri, S.V.; Mazer, B.D. Intravenous immunoglobulin: Mechanism of action in autoimmune and inflammatory conditions. J. Allergy Clin. Immunol. Pract. 2023, 11, 1688–1697. [CrossRef]
- Shock, A.; Humphreys, D.; Nimmerjahn, F. Dissecting the mechanism of action of intravenous immunoglobulin in human autoimmune disease: Lessons from therapeutic modalities targeting Fcγ receptors. J. Allergy Clin. Immunol. 2020, 146, 492–500. [CrossRef]
- Ashton, C.; Paramalingam, S.; Stevenson, B.; Brusch, A.; Needham, M. Idiopathic inflammatory myopathies: A review. Intern. Med. J. 2021, 51, 845–852. [CrossRef]
- Zeng, R.; Glaubitz, S.; Schmidt, J. Antibody therapies in autoimmune inflammatory myopathies: Promising treatment options. Neurotherapeutics 2022, 19, 911–921. [CrossRef]
- Gandiga, P.C.; Ghetie, D.; Anderson, E.; Aggrawal, R. Intravenous immunoglobulin in idiopathic inflammatory myopathies: A practical guide for clinical use. Curr. Rheumatol. Rep. 2023, 25, 152–168. [CrossRef]
- Sanchis, P.; Fernández-Gayol, O.; Comes, G.; Escrig, A.; Giralt, M.; Palmiter, R.D.; Hidalgo, J. Interleukin-6 derived from the central nervous system may influence the pathogenesis of experimental autoimmune encephalomyelitis in a cell-dependent manner. Cells 2020, 9, 330. [CrossRef]
- Danieli, M.G.; Antonelli, E.; Auria, S.; Buti, E.; Shoenfeld, Y. Low-dose intravenous immunoglobulin (IVIg) in different immune-mediated conditions. Autoimmun. Rev. 2023, 22, 103451. [CrossRef]
- Gillespie, E.R.; Ruitenberg, M.J. Neuroinflammation after SCI: Current insights and therapeutic potential of intravenous immunoglobulin. J. Neurotrauma 2022, 39, 320–332. [CrossRef]
- Mroué, M.; Bessaguet, F.; Nizou, A.; Richard, L.; Sturtz, F.; Magy, L.; Bourthoumieu, S.; Danigo, A.; Demiot, C. Neuroprotective Effect of Polyvalent Immunoglobulins on Mouse Models of Chemotherapy-Induced Peripheral Neuropathy. Pharmaceutics 2024, 16, 139. [CrossRef]
- Ren, X.; Zhang, M.; Zhang, X.; Zhao, P.; Zhai, W. Can low-dose intravenous immunoglobulin be an alternative to high-dose intravenous immunoglobulin in the treatment of children with newly diagnosed immune thrombocytopenia: A systematic review and meta-analysis. BMC Pediatr. 2024, 24, 199. [CrossRef]
- Hoffmann, J.H.; Enk, A.H. High-dose intravenous immunoglobulin in skin autoimmune disease. Front. Immunol. 2019, 10, 1090. [CrossRef]
- Nadig, P.L.; Joshi, V.; Pilania, R.K.; Kumrah, R.; Kabeerdoss, J.; Sharma, S.; Suri, D.; Rawat, A.; Singh, S.. Intravenous immunoglobulin in Kawasaki disease—Evolution and pathogenic mechanisms. Diagnostics 2023, 13, 2338. [CrossRef]
- de Carvalho, J.F.; Skare, T.L. Rituximab combined with intravenous immunoglobulin in autoimmune diseases: A systematic review. Adv. Rheumatol. 2025, 65, 1–10.
- N’kaoua, E.; Attarian, S.; Delmont, E.; Campana-Salort, E.; Verschueren, A.; Grapperon, A.-M.; Mestivier, E.; Roche, M. Immunoglobulin shortage: Practice modifications and clinical outcomes in a reference centre. Rev. Neurol. 2022, 178, 616–623. [CrossRef]
- Barthel, C.; Musquer, M.; Veyrac, G.; Bernier, C. Delayed eczematous skin reaction as an adverse drug reaction to immunoglobulin infusions: A case series. In Annales de Dermatologie et de Vénéréologie; Elsevier Masson: Amsterdam, The Netherlands, 2022; Volume 149, pp. 264–270. [CrossRef]
- Ozen, S.; Esenboga, S. Alternative Therapies for Cytokine Storm Syndromes. Cytokine Storm Syndr. 2019, 581–593.
- Kadry, H.; Noorani, B.; Cucullo, L. A blood–brain barrier overview on structure, function, impairment, and biomarkers of integrity. Fluids Barriers CNS 2020, 17, 1–24.
- Justiz-Vaillant, A.; Gopaul, D.; Soodeen, S.; Unakal, C.; Thompson, R.; Pooransingh, S.; Arozarena-Fundora, R.; Asin-Milan, O.; Akpaka, P.E. Advancements in Immunology and Microbiology Research: A Comprehensive Exploration of Key Areas. Microorganisms 2024, 12, 1672. [CrossRef]
- Justiz-Vaillant, A.A.; Gopaul, D.; Soodeen, S.; Arozarena-Fundora, R.; Barbosa, O.A.; Unakal, C.; Thompson, R.; Pandit, B.; Umakanthan, S.; Akpaka, P.E. Neuropsychiatric Systemic Lupus Erythematosus: Molecules Involved in Its Imunopathogenesis, Clinical Features, and Treatment. Molecules 2024, 29, 747. [CrossRef]
- Wesselingh, R.; Butzkueven, H.; Buzzard, K.; Tarlinton, D.; O’Brien, T.J.; Monif, M. Innate immunity in the central nervous system: A missing piece of the autoimmune encephalitis puzzle? Front. Immunol. 2019, 10, 2066. [CrossRef]
- Radetz, A.; Groppa, S. White Matter Pathology. In Translational Methods for Multiple Sclerosis Research; Springer: New York, NY, USA, 2021; pp. 29–46.
- Dhaiban, S.; Al-Ani, M.; Elemam, N.M.; Al-Aawad, M.H.; Al-Rawi, Z.; Maghazachi, A.A. Role of peripheral immune cells in multiple sclerosis and experimental autoimmune encephalomyelitis. Sci 2021, 3, 12. [CrossRef]
- Bordet, R.; Camu, W.; De Seze, J.; Laplaud, D.A.; Ouallet, J.C.; Thouvenot, E. Mechanism of action of s1p receptor modulators in multiple sclerosis: The double requirement. Rev. Neurol. 2020, 176, 100–112. [CrossRef]
- Yang, T.; Tian, X.; Chen, C.; Ma, L.; Zhou, S.; Li, M.; Wu, Y.; Zhou, Y.; Cui, Y.. The efficacy and safety of fingolimod in patients with relapsing multiple sclerosis: A meta-analysis. Br. J. Clin. Pharmacol. 2020, 86, 637–645. [CrossRef]
- Stascheit, F.; Li, L.; Mai, K.; Baum, K.; Siebert, E.; Ruprecht, K. Delayed onset hypophysitis after therapy with daclizumab for multiple sclerosis–A report of two cases. J. Neuroimmunol. 2021, 351, 577469. [CrossRef]
- Kim, W.; Patsopoulos, N.A. Genetics and functional genomics of multiple sclerosis. In Seminars in Immunopathology; Springer: Berlin/Heidelberg, Germany, 2022; Volume 44, pp. 63–79.
- Cohan, S.L.; Lucassen, E.B.; Romba, M.C.; Linch, S.N. Daclizumab: Mechanisms of action, therapeutic efficacy, adverse events and its uncovering the potential role of innate immune system recruitment as a treatment strategy for relapsing multiple sclerosis. Biomedicines 2019, 7, 18. [CrossRef]
- Rothhammer, V., Kenison, J. E., Li, Z., Tjon, E., Takenaka, M. C., Chao, C. C., ... & Quintana, F. J. Aryl hydrocarbon receptor activation in astrocytes by laquinimod ameliorates autoimmune inflammation in the CNS. Neurology: Neuroimmunology & Neuroinflammation, 2021, 8(2), e946. [CrossRef]
- Biernacki, T.; Sandi, D.; Bencsik, K.; Vécsei, L. Kynurenines in the Pathogenesis of Multiple Sclerosis: Therapeutic Perspectives. Cells 2020, 9, 1564. [CrossRef]
- Comi, G.; Dadon, Y.; Sasson, N.; Steinerman, J.R.; Knappertz, V.; Vollmer, T.L.; Boyko, A.; Vermersch, P.; Ziemssen, T.; Montalban, X.; et al. CONCERTO: A randomized, placebo-controlled trial of oral laquinimod in relapsing-remitting multiple sclerosis. Mult. Scler. J. 2022, 28, 608–619.
- Ruetsch-Chelli, C.; Bresch, S.; Seitz-Polski, B.; Rosenthal, A.; Desnuelle, C.; Cohen, M.; Brglez, V.; Ticchioni, M.; Lebrun-Frenay, C. Memory B cells predict relapse in rituximab-treated myasthenia gravis. Neurotherapeutics 2021, 18, 938–948. [CrossRef]
- Mantegazza, R.; Antozzi, C. When myasthenia gravis is deemed refractory: Clinical signposts and treatment strategies. Ther. Adv. Neurol. Disord. 2018, 11, 1756285617749134. [CrossRef]
- Chan, A.M.; Baehring, J.M. Paraneoplastic neurological syndromes: A single institution 10-year case series. J. Neurooncol. 2019, 141, 431–439. [CrossRef]
- Rosenfeld, M.R.; Dalmau, J. Paraneoplastic neurologic syndromes. Neurol. Clin. 2018, 36, 675–685.
- Mladinich, M.C.; Himmler, G.E.; Conde, J.N.; Gorbunova, E.E.; Schutt, W.R.; Sarkar, S.; Tsirka, S.-A.E.; Kim, H.K.; Mackow, E.R. Age-dependent Powassan virus lethality is linked to glial cell activation and divergent neuroinflammatory cytokine responses in a murine model. J. Virol. 2024, e0056024. [CrossRef]
- Alakhras, N.S.; Zhang, W.; Barros, N.; Sharma, A.; Ropa, J.; Priya, R.; Yang, X.F.; Kaplan, M.H. An IL-23-STAT4 pathway is required for the proinflammatory function of classical dendritic cells during CNS inflammation. Proc. Natl. Acad. Sci. USA 2024, 121, e2400153121. [CrossRef]
- Ruan, Z.; Tang, Y.; Gao, T.; Li, C.; Guo, R.; Sun, C.; Huang, X.; Li, Z.; Chang, T. Efficacy and safety of tocilizumab in patients with refractory generalized myasthenia gravis. CNS Neurosci. Ther. 2024, 30, e14793. [CrossRef]
- Walton, C.; King, R.; Rechtman, L.; Kaye, W.; Leray, E.; Marrie, R.A.; Robertson, N.; La Rocca, N.; Uitdehaag, B.; Van Der Mei, I.; et al. Rising prevalence of multiple sclerosis worldwide: Insights from the Atlas of MS, third edition. Mult. Scler. J. 2020, 26, 1816–1821. [CrossRef]
- Heitmann, H.; Andlauer, T.F.; Korn, T.; Mühlau, M.; Henningsen, P.; Hemmer, B.; Ploner, M. Fatigue, depression, and pain in multiple sclerosis: How neuroinflammation translates into dysfunctional reward processing and anhedonic symptoms. Mult. Scler. J. 2022, 28, 1020–1027. [CrossRef]
- Zarghami, A.; Li, Y.; Claflin, S.B.; van der Mei, I.; Taylor, B.V. Role of environmental factors in multiple sclerosis. Expert Rev. Neurother. 2021, 21, 1389–1408. [CrossRef]
- Cruciani, C.; Puthenparampil, M.; Tomas-Ojer, P.; Jelcic, I.; Docampo, M.J.; Planas, R.; Manogaran, P.; Opfer, R.; Wicki, C.; Reindl, M.; et al. T-cell specificity influences disease heterogeneity in multiple sclerosis. Neurol.-Neuroimmunol. Neuroinflamm. 2021, 8, e1075. [CrossRef]
- Long, H.M.; Meckiff, B.J.; Taylor, G.S. The T-cell response to Epstein-Barr virus–new tricks from an old dog. Front. Immunol. 2019, 10, 2193. [CrossRef]
- Bjornevik, K.; Cortese, M.; Healy, B.C.; Kuhle, J.; Mina, M.J.; Leng, Y.; Elledge, S.J.; Niebuhr, D.W.; Scher, A.I.; Munger, K.L.; et al. Longitudinal analysis reveals high prevalence of Epstein-Barr virus associated with multiple sclerosis. Science 2022, 375, 296–301. [CrossRef]
- Deeba, E.; Koptides, D.; Gaglia, E.; Constantinou, A.; Lambrianides, A.; Pantzaris, M.; Krashias, G.; Christodoulou, C. Evaluation of Epstein-Barr virus-specific antibodies in Cypriot multiple sclerosis patients. Mol. Immunol. 2019, 105, 270–275. [CrossRef]
- Bjornevik, K.; Cortese, M.; Healy, B.C.; Kuhle, J.; Mina, M.J.; Leng, Y.; Elledge, S.J.; Niebuhr, D.W.; Scher, A.I.; Munger, K.L.; et al. Longitudinal analysis reveals high prevalence of Epstein-Barr virus associated with multiple sclerosis. Science 2022, 375, 296–301. [CrossRef]
- Huang, J.; Tengvall, K.; Lima, I.B.; Hedström, A.K.; Butt, J.; Brenner, N.; Gyllenberg, A.; Stridh, P.; Khademi, M.; Ernberg, I.; et al. Genetics of immune response to Epstein-Barr virus: Prospects for multiple sclerosis pathogenesis. Brain 2024, 147, 3573–3582. [CrossRef]
- Jog, N.R.; McClain, M.T.; Heinlen, L.D.; Gross, T.; Towner, R.; Guthridge, J.M.; Axtell, R.C.; Pardo, G.; Harley, J.B.; James, J.A. Epstein Barr virus nuclear antigen 1 (EBNA-1) peptides recognized by adult multiple sclerosis patient sera induce neurologic symptoms in a murine model. J. Autoimmun. 2020, 106, 102332. [CrossRef]
- Bjornevik, K.; Cortese, M.; Healy, B.C.; Kuhle, J.; Mina, M.J.; Leng, Y.; Elledge, S.J.; Niebuhr, D.W.; Scher, A.I.; Munger, K.L.; et al. Longitudinal analysis reveals high prevalence of Epstein-Barr virus associated with multiple sclerosis. Science 2022, 375, 296–301. [CrossRef]
- Tengvall, K.; Huang, J.; Hellström, C.; Kammer, P.; Biström, M.; Ayoglu, B.; Bomfim, I.L.; Stridh, P.; Butt, J.; Brenner, N.; et al. Molecular mimicry between Anoctamin 2 and Epstein-Barr virus nuclear antigen 1 associates with multiple sclerosis risk. Proc. Natl. Acad. Sci. USA 2019, 116, 16955–16960. [CrossRef]
- Wang, Z.; Kennedy, P.G.; Dupree, C.; Wang, M.; Lee, C.; Pointon, T.; Langford, T.D.; Graner, M.W.; Yu, X. Antibodies from Multiple Sclerosis Brain Identified Epstein-Barr Virus Nuclear Antigen 1 & 2 Epitopes which Are Recognized by Oligoclonal Bands. J. Neuroimmune Pharmacol. 2021, 16, 567–580. [CrossRef]
- Neves, M.; Marinho-Dias, J.; Ribeiro, J.; Sousa, H. Epstein-Barr virus strains and variations: Geographic or disease-specific variants? J. Med. Virol. 2017, 89, 373–387. [CrossRef]
- Thomas, O.G.; Bronge, M.; Tengvall, K.; Akpinar, B.; Nilsson, O.B.; Holmgren, E.; Hessa, T.; Gafvelin, G.; Khademi, M.; Alfredsson, L.; et al. Cross-reactive EBNA1 immunity targets alpha-crystallin B and is associated with multiple sclerosis. Sci. Adv. 2023, 9, eadg3032. [CrossRef]
- Lanz, T.V.; Brewer, R.C.; Ho, P.P.; Moon, J.-S.; Jude, K.M.; Fernandez, D.; Fernandes, R.A.; Gomez, A.M.; Nadj, G.-S.; Bartley, C.M.; et al. Clonally expanded B cells in multiple sclerosis bind EBV EBNA1 and GlialCAM. Nature 2022, 603, 321–327. [CrossRef]
- Telford, M.; Hughes, D.A.; Juan, D.; Stoneking, M.; Navarro, A.; Santpere, G. Expanding the geographic characterisation of Epstein–Barr virus variation through gene-based approaches. Microorganisms 2020, 8, 1686. [CrossRef]
- Romero, C.; Quijada, A.; Abudinén, G.; Céspedes, C.; Aguilera, L. Opercular myoclonic-anarthric status (OMASE) secondary to anti-Hu paraneoplastic neurological syndrome. Epilepsy Behav. Rep. 2024, 27, 100703. [CrossRef]
- Zhu, F.; Shan, W.; Lv, R.; Li, Z.; Wang, Q. Clinical characteristics of anti-GABA-B receptor encephalitis. Front. Neurol. 2020, 11, 403. [CrossRef]
- Tsang-Shan, C.; Ming-Chi, L.; Huang, H.Y.I.; Chin-Wei, H.; Immunity, Ion Channels and Epilepsy. Int. J. Mol. Sci. 2022, 23, 6446.
- Gilligan, M.; McGuigan, C.; McKeon, A. Paraneoplastic neurologic disorders. Curr. Neurol. Neurosci. Rep. 2023, 23, 67–82.
- Gilligan, M.; McGuigan, C.; McKeon, A. Autoimmune central nervous system disorders: Antibody testing and its clinical utility. Clin. Biochem. 2024, 126, 110746. [CrossRef]
- Zhou, K.; Zhang, L.; Shen, S.; Lin, J.-F.; Wang, J.-R.; Zhou, D.; Li, J.-M.; Sima, X. Neurological autoantibody prevalence in chronic epilepsy: Clinical and neuropathologic findings. Seizure: Eur. J. Epilepsy 2024, 115, 28–35. [CrossRef]
- Dubey, D.; Kothapalli, N.; McKeon, A.; Flanagan, E.P.; Lennon, V.A.; Klein, C.J.; Britton, J.W.; So, E.; Boeve, B.F.; Tillema, J.-M.; et al. Predictors of neural-specific autoantibodies and immunotherapy response in patients with cognitive dysfunction. J. Neuroimmunol. 2018, 323, 62–72. [CrossRef]
- Orozco, E.; Valencia-Sanchez, C.; Britton, J.; Dubey, D.; Flanagan, E.P.; Lopez-Chiriboga, A.S.; Zalewski, N.; Zekeridou, A.; Pittock, S.J.; McKeon, A. Autoimmune encephalitis criteria in clinical practice. Neurol. Clin. Pract. 2023, 13, e200151. [CrossRef]
- Abide, Z.; Nasr, K.S.; Kaddouri, S.; Edderai, M.; Elfenni, J.; Salaheddine, T. Bickerstaff brainstem encephalitis: A case report. Radiol. Case Rep. 2023, 18, 2704–2706. [CrossRef]
- rozco, E.; Guo, Y.; Chen, J.J.; Dubey, D.; Howell, B.; Moutvic, M.; Louis, E.K.S.; McKeon, A. Clinical reasoning: A 43-year-old man with subacute onset of vision disturbances, jaw spasms, and balance and sleep difficulties. Neurology 2022, 99, 387–392. [CrossRef]
- Tisavipat, N.; Chang, B.K.; Ali, F.; Pittock, S.J.; Kammeyer, R.; Declusin, A.; Cohn, S.J.; Flanagan, E.P. Subacute horizontal diplopia, jaw dystonia, and laryngospasm. Neurol. Neuroimmunol. Neuroinflamm. 2023, 10, e200128. [CrossRef]
- Lana-Peixoto, M.A.; Talim, N. Neuromyelitis optica spectrum disorder and anti-MOG syndromes. Biomedicines 2019, 7, 49. [CrossRef]
- Blattner, M.S.; Day, G.S. Sleep disturbances in patients with autoimmune encephalitis. Curr. Neurol. Neurosci. Rep. 2020, 20, 28. [CrossRef]
- O’Connor, K.; Waters, P.; Komorowski, L.; Zekeridou, A.; Guo, C.Y.; Mgbachi, V.C.; Probst, C.; Mindorf, S.; Teegen, B.; Gelfand, J.M.; et al. GABA(A) receptor autoimmunity: A multicenter experience. Neurol. Neuroimmunol. Neuroinflamm. 2019, 6, e552.
- Gravier-Dumonceau, A.; Ameli, R.; Rogemond, V.; Ruiz, A.; Joubert, B.; Muniz-Castrillo, S.; Vogrig, A.; Picard, G.; Ambati, A.; Benaiteau, M.; et al. Glial fibrillary acidic protein autoimmunity: A French cohort study. Neurology 2022, 98, e653–e668.
- Flanagan, E.P.; Hinson, S.R.; Lennon, V.A.; Fang, B.; Aksamit, A.J.; Morris, P.P.; Basal, E.; Honorat, J.A.; Alfugham, N.B.; Linnoila, J.J.; et al. Glial fibrillary acidic protein immunoglobulin G as biomarker of autoimmune astrocytopathy: Analysis of 102 patients. Ann. Neurol. 2017, 81, 298–309. [CrossRef]
- Guasp, M.; Dalmau, J. Encephalitis associated with antibodies against the NMDA receptor. Med. Clin. 2018, 151, 71–79. [CrossRef]
- Budhram, A.; Sharma, M.; Young, G.B. Seizures in anti-hu-associated extra-limbic encephalitis: Characterization of a unique disease manifestation. Epilepsia 2022, 63, e172–e177.
- Liu, M.; Ren, H.; Wang, L.; Fan, S.; Bai, L.; Guan, H. Prognostic and relapsing factors of primary autoimmune cerebellar ataxia: A prospective cohort study. J. Neurol. 2024, 271, 1072–1079. [CrossRef]
- Hadjivassiliou, M.; Graus, F.; Honnorat, J.; Jarius, S.; Titulaer, M.; Manto, M.; Hoggard, N.; Sarrigiannis, P.; Mitoma, H. Diagnostic criteria for primary autoimmune cerebellar ataxia-guidelines from an international task force on immune-mediated cerebellar ataxias. Cerebellum 2020, 19, 605–610. [CrossRef]
- Banwell, B.; Bennett, J.L.; Marignier, R.; Kim, H.J.; Brilot, F.; Flanagan, E.P.; Ramanathan, S.; Waters, P.; Tenembaum, S.; Graves, J.S.; et al. Diagnosis of myelin oligodendrocyte glycoprotein antibody-associated disease: International MOGAD panel proposed criteria. Lancet Neurol. 2023, 22, 268–282. [CrossRef]
- Valencia-Sanchez, C.; Flanagan, E.P. Uncommon inflammatory/immune-related myelopathies. J. Neuroimmunol. 2021, 361, 577750. [CrossRef]
- Chiriboga, S.L.; Flanagan, E.P. Myelitis and other autoimmune myelopathies. Contin. Lifelong Learn. Neurol. 2021, 27, 62–92. [CrossRef]
- Banks, S.A.; Morris, P.P.; Chen, J.J.; Pittock, S.J.; Sechi, E.; Kunchok, A.; Tillema, J.-M.; Fryer, J.P.; Weinshenker, B.G.; Krecke, K.N.; et al. Brainstem and cerebellar involvement in MOG-IgG-associated disorder versus aquaporin-4-IgG and MS. J. Neurol. Neurosurg. Psychiatry 2021, 92, 384–390. [CrossRef]
- Ciron, J.; Cobo-Calvo, A.; Audoin, B.; Bourre, B.; Brassat, D.; Cohen, M.; Collongues, N.; Deschamps, R.; Durand-Dubief, F.; Laplaud, D.; et al. Frequency and characteristics of short versus longitudinally extensive myelitis in adults with MOG antibodies: A retrospective multicentric study. Mult. Scler. J. 2020, 26, 936–944. [CrossRef]
- Cacciaguerra, L.; Sechi, E.; Rocca, M. A.; Filippi, M.; Pittock, S. J.; & Flanagan, E. P. Neuroimaging features in inflammatory myelopathies: a review. Frontiers in Neurology, 2022, 13, 993645. [CrossRef]
- McKeon, A.; Lesnick, C.; Vorasoot, N.; Buckley, M. W.; Dasari, S.; Flanagan, E. P.; Mills, J.; et al. Utility of protein microarrays for detection of classified and novel antibodies in autoimmune neurologic disease. Neurology: Neuroimmunology & Neuroinflammation, 2023, 10(5), e200145. [CrossRef]
- Fonseca, E.; Varas, R.; Godoy-Santín, J.; Valenzuela, R.; Sandoval, P. Opsoclonus-myoclonus syndrome associated with anti Kelch-like protein-11 antibodies in a young female patient without cancer. J. Neuroimmunol. 2021, 355, 577570. [CrossRef]
- Qin, C.; Zhang, M.; Mou, D. P.; Zhou, L. Q.; Dong, M. H.; Huang, L.; Wang, W.; Cai, S. B.; You, Y. F.; Shang, K.; Xiao, J.; Wang, D.; Li, C. R.; Hao, Y.; Heming, M.; Wu, L. J.; Meyer Zu Hörste, G.; Dong, C.; Bu, B. T.; Tian, D. S.; … Wang, W. Single-cell analysis of anti-BCMA CAR T cell therapy in patients with central nervous system autoimmunity. Science immunology, 2024, 9(95), eadj9730. [CrossRef]
- Vukovic, J.; Abazovic, D.; Vucetic, D.; Medenica, S. CAR-engineered T cell therapy as an emerging strategy for treating autoimmune diseases. Front Med (Lausanne). 2024; 11:1447147. [CrossRef]
- Li, Y. R.; Lyu, Z.; Chen, Y.; Fang, Y.; & Yang, L. Frontiers in CAR-T cell therapy for autoimmune diseases. Trends in pharmacological sciences, 2024, 45(9), 839–857. [CrossRef]
Figure 1.
The Blood-brain barrier (BBB) and the immune system play a role in the immunopathogenesis of NPSLE [46].
Figure 1.
The Blood-brain barrier (BBB) and the immune system play a role in the immunopathogenesis of NPSLE [46].
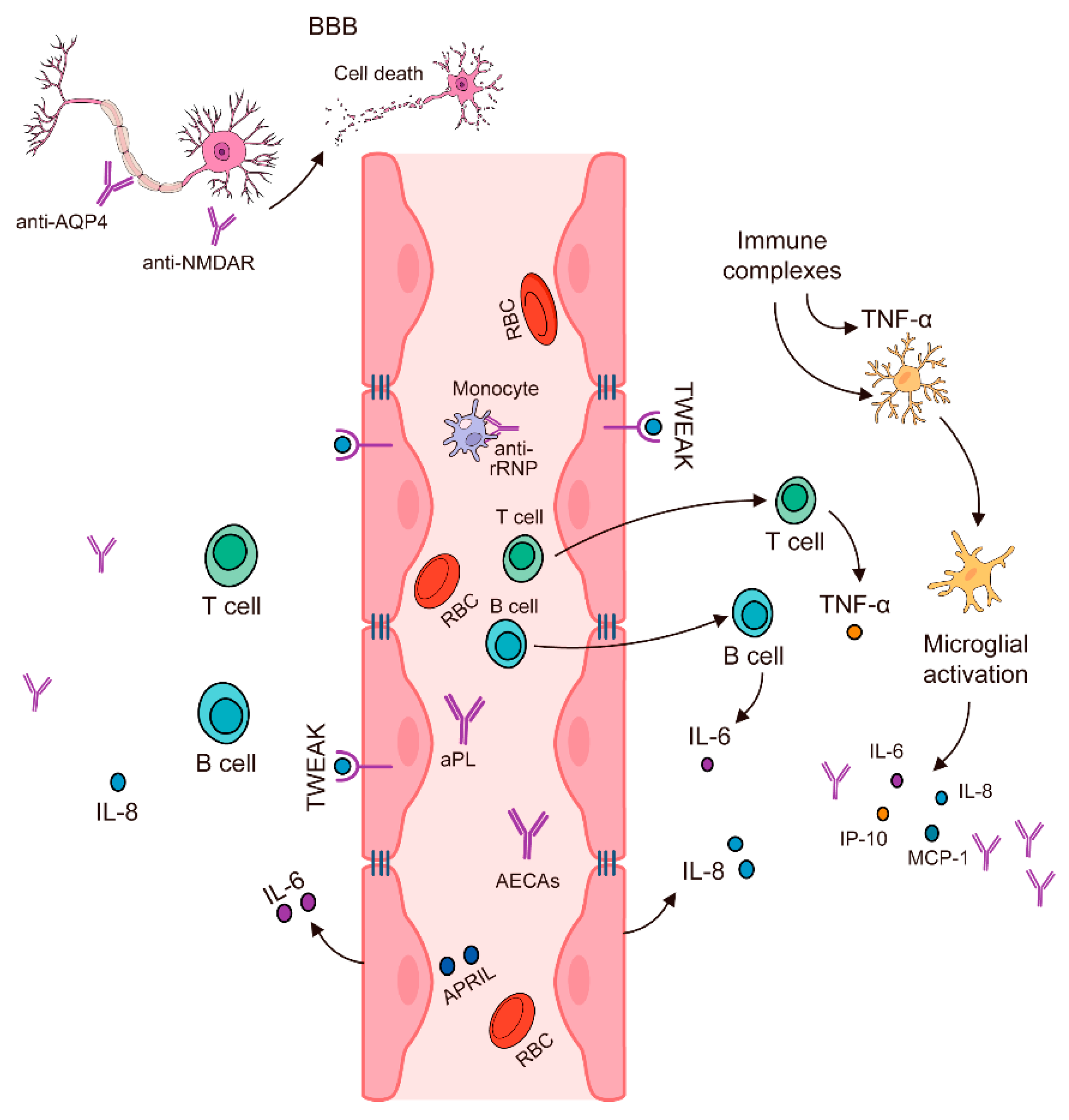

Table 1.
Presence of auto-antibodies in neurological and psychiatric disorders.
| Disease | Autoantibody | Reference | ||
|---|---|---|---|---|
| Cognitive and affective dysfunctions in autoimmune thyroiditis | Anti-thyroid peroxidase Ab, anti-central nervous system Ab | [8] | ||
| Hashimoto’s encephalopathy (HE) | Anti-α-enolase Ab, anti-thyroid peroxidase Ab |
[8,9] | ||
| Limbic encephalitis multiple sclerosis | Anti-N-methyl D-aspartate-type glutamate receptor Ab | [1,10] | ||
| Complex regional syndrome | Anti-nuclear Ab (ANA), anti-neuronal Ab | [11] | ||
| Idiopathic and symptomatic epilepsies | Neurotropic Abs to NF-200, GFAP, MBP and S100β, and to receptors of neuromediators (glutamate, GABA, dopamine, serotonin and choline-receptors | [12] | ||
| Schizophrenia | Autoantibodies against glutamate, dopamine, acetylcholine and serotonin receptors, and antineuronal antibodies against synaptic biomolecules | [13,14,15,16] | ||
| Lambert-Eaton myasthenic syndrome | Autoantibodies against P/Q-type voltage-gated calcium channels | [17] | ||
| Myasthenia gravis | Auto-Ab to tyrosine kinase | muscle-specific | [18] | |
Table 2.
Cytokine involvement in neuropsychiatric diseases.
| Disease | Cytokine Involved | Reference |
|---|---|---|
| Neuropsychiatric systemic lupus erythematosus | Elevated interleukin (IL)-17, IL-2, interferon- gamma (IFN-γ), IL-5, basic fibroblast growth factor (FGF) and IL-15 |
[19] |
| Relapsing remitting multiple sclerosis | Elevated IL-17 and INF-gamma and decreased transforming growth factor- beta (TGF-beta 1) levels | [20] |
| Guillain-Barre syndrome | Elevated TNFα and IL-10 | [21] |
| Schizophrenia | Increased interleukin (IL)-1, IL-6, and TGF-β appear to be state markers, whereas IL-12, interferon-gamma (IFN-γ), TNF-α, and soluble IL-2 receptor appear to be trait markers | [22,23] |
| Multiple sclerosis (MS) | IL-17 plays an important role in the inflammatory phase of relapsing- remitting MS | [24] |
Table 3.
List of neurological diseases where the use of intravenous immunoglobulins (IVIG) has been used successfully.
Table 3.
List of neurological diseases where the use of intravenous immunoglobulins (IVIG) has been used successfully.
| Diseases | References |
|---|---|
| Guillain-Barré syndrome | [25,26,27,28,29] |
| Chronic inflammatory demyelinating polyneuropathy | [25,26,27,28,29] |
| Multiple motor neuropathy | [26,27] |
| Multiple sclerosis | [26,27] |
| Myasthenia gravis | [26,27,28] |
| Acute disseminated encephalomyelitis | [27] |
| Diabetic neuropathy | [27] |
| Lambert-Eaton myasthenic syndrome | [27] |
| Opsoclonus-myoclonus | [27] |
| Pediatric autoimmune neuropsychiatric disorders associated with streptococcal infections | [27] |
| Polymyositis | [27,30,31,32] |
| Rasmussen’s encephalitis | [27] |
| Multiple sclerosis | [33] |
Table 4.
Presents an overview of experimental therapies and their mechanisms of action in neurological autoimmune disorders. These therapies aim to address the underlying autoimmune processes, restore BBB integrity, and mitigate inflammation and neurotoxicity.
Table 4.
Presents an overview of experimental therapies and their mechanisms of action in neurological autoimmune disorders. These therapies aim to address the underlying autoimmune processes, restore BBB integrity, and mitigate inflammation and neurotoxicity.
| Disease | Drug used | Mechanism of Action | Reference |
|---|---|---|---|
| Multiple sclerosis | Oral fingolimod | It Inhibits egress of lymphocytes from lymph nodes and their recirculation | [50,51] |
| Multiple sclerosis | Daclizumab | It is a humanized neutralizing monoclonal antibody against the α-chain of the interleukin-2 receptor |
[52,53,54] |
| Experimental autoimmune encephalomyelitis | Laquinimod | Modulates adaptive T cell immune responses via its effects on cells of the innate immune system and may not influence T cells directly | [55,56,57] |
| Myasthenia gravis | Rituximab | A chimeric IgG k monoclonal antibody that target CD20 on B cells | [58,59] |
| Guillain-Barré syndrome | Plasma exchange | Deplete pathogenic autoantibody | [60] |
| Paraneoplastic neurological disorders | IVIG, plasma exchange | Immunomodulator, deplete auto-Ab | [61] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



