
Submitted:
24 April 2025
Posted:
24 April 2025
You are already at the latest version
Abstract
Lactic acid bacteria are components of the gastrointestinal tract microbiota in both humans and animals and are widely used as probiotics. Lactobacillus is the most related genus to probiotic activity and is able to release membrane microvesicles (MVs), which primary functions include to carry and transmit antigens to host tissues and modulate host defense responses. In the present study, MVs were isolated from Lactobacillus acidophilus resident in the ileum of free-living rats, and its immunostimulant effect was evaluated in two biological models. MVs were characterized using SDS-PAGE electrophoresis, electron microscopy, and nanoparticle tracking analysis. In the first model, the immunostimulant effect of MVs was evaluated on ovine abomasal explants, previously stimulated with MVs and challenged with third-stage larvae of Haemonchus contortus, demonstrating a decrease in the percentage of larvae association, and favoring the migration of inflammatory cells to the infection site. In the second model, macrophage cell line (RAW 264.7) was stimulated with MVs to evaluate the expression of transcripts coding for IL-1β and TNF-α. In both cases, the immunostimulatory effect of MVs was verified, demonstrating that MVs can carry important antigens that could be considered in a formulation to promote the intestinal health in livestock production.
Keywords:
microvesicles
; probiotics
; Lactobacillus acidophilus
; Immunostimulant
; cytokines
1. Introduction
Lactic acid bacteria (LAB) are a broad group of microorganisms commonly used as probiotics. Lactobacillus acidophilus is the most known strain of the Lactobacillus genus, since it has been the most used and studied bacterium over time [1]. LAB widely exist in human and animal mucosae and in fermented foods [2,3]. These microorganisms are associated to immunomodulatory functions and the capacity to reduce the risk of infection and death by pathogens in the gastrointestinal tract (GIT) on mammals [4]. Probiotics are live microorganisms that have beneficial effects on the host when they are administered orally. Scientific evidence shows that the molecules and cellular structures released by these microorganisms have an important probiotic capacity [5,6,7].
All prokaryotic cells have developed numerous ways to transfer proteins to the environment, which largely involve the assistance of devoted protein secretion systems. In that way, the secreted membrane microvesicles (MVs) have become one of the most important mechanisms of bacterial secretion systems and intercellular communication. MVs are blebs from the outer membrane (OMVs) of Gram-negative bacteria and from cytoplasmic membrane (MVs) of Gram-positive bacteria and are produced in all growth phases [8].
Compared to the vast study of OMVs, MVs are of relatively recent interest, so the mechanisms of biogenesis are poorly understood [9]. However, it is believed that the formation of MVs may be stimulated by weakening of the cell envelope by enzymes that hydrolyze the peptidoglycan or by β-lactam antibiotics [9,10,11]. In addition to the above, the use of sublethal concentrations of antibiotics that interfere at the cell wall level and in the protein synthesis, such as penicillin [10], ampicillin [11], and gentamycin [12], shows a significant increase in the production of MVs.
Regarding the genus Lactobacillus, MVs production has been described in L. rhamnosus JB-1 [7], L. plantarum [13], L. acidophilus [14], L. reuteri [15] and L. casei BL23 [16]. Immune regulation by MVs from LAB has been proposed to be involved in signaling between probiotic intestinal bacteria and their mammalian hosts [7]. MVs immunomodulatory effects are related to their interaction with different immune and non-immune cells through pattern recognition receptors, such as Toll-like receptors [TLR] and NOD-like receptors (NLR), as well as different pathogen-associated molecular patterns (PAMPs) [17]. MVs from different bacterial species have shown to increase the production of proinflammatory cytokines. For instance, Staphylococcus aureus MVs [18] stimulate IL-1β and IL-18 production in murine models, Escherichia coli C25 enhances IL-8 production in intestinal epithelial cells [19], and Akkermansia muciniphila induces IL-6 secretion [20]. For LAB-derived MVs, L. casei has been reported to increase IL-6 in human intestinal epithelial cells [21], and L. reuteri DSM-17938 promotes motility in mouse intestines [15]. The important immune response that MVs stimulate in the host has favored the creation of acellular vaccines against pathogenic bacteria such as Neisseria meningitides [22], Vibrio cholera [23], Burkholderia pseudomallei [24], and Mannheimia haemolytica A1 [25], and A2 [26].
The use of probiotics in animal production and health is becoming increasingly relevant [27,28], because the supplementation of probiotics in the diet can improve animal health and performance, through contributions to gut health and nutrient absorption [29,30], as well as to reduce the use of antimicrobials to which microorganisms have become increasingly resistant. The main objective of probiotics is to modulate the host immune system to prevent and treat infections [31]. In livestock industry, it is reported that probiotics administration enhances milk production in Holstein Friesian cows [32,33], while in other species like pigs favors microbiota diversity and weight gain [34,35,36]. In poultry, there are studies who claims that probiotics improve broilers and eggs production [37,38]. Probiotics have not been restricted to animal production, but they are also used in pets such as dogs and cats, in which promote the diversity of the microbiota as well as improve the immune response and help to control diarrhea [29,39,40].
In a previous study, the production and characterization of MVs from LAB isolated from free-living rats GIT were studied [41]. The relevance of working with LAB isolated from this rodent is justified because it is exposed and resistant to many pathogens that infect humans and food animals without developing disease, which can be attributed to its intestinal microbiota [42,43,44]. Lonngi et al. [2022] previously reported a bactericidal effect of 30 µg of MVs from L. acidophilus isolated from the ileum of free-living rats on the growth of Salmonella enterica serovar Typhimurium (ATCC 154) and a field strain of E. coli [41].
On the other hand, parasitic diseases that affect the livestock industry is a persistent and challenging problem due to the resistance that parasites have developed to the different drugs intended for their control. One of the most important ovine gastrointestinal tracts (GIT) diseases is caused by Haemonchus contortus, a hematophagous parasite that causes anemia and death in animals, even when it induces a strong Th2 immune response [45,46,47]. The life cycle of this parasite is direct, and animals become infected when consuming feed contaminated with third-stage larvae [L3], the most pathogenic larval phase. Once in the abomasum, it transforms into L4 and L5 until it reaches adulthood [48]. However, there are no reports of the effects of MVs of probiotic bacteria on L3 of H. contortus.
Although the microbicidal and immunostimulant effects of probiotics are well established, there are few studies on the effect of MVs from L. acidophilus. Nevertheless, there is no information for MVs of L. acidophilus isolated from free-living rats. Bacterial MVs in general have demonstrated their potent ability to carry biologically active antigens, which efficiently stimulate the immune system. The aim of this study was to determine the immunostimulatory capacity of MVs from L. acidophilus on two in vitro biological models: the first was based on using ovine abomasal tissue explants and the second on macrophage cell line cultures (RAW 264.7). The present study is the first report about the MVs effects of L. acidophilus isolated from free-living rats against pathogens that affect animal production and cellular responses.
2. Materials and Methods
2.1. Cells and Strains
Lactobacillus acidophilus ATCC® 314 (KWIK-STIK, Microbiologics) and L. acidophilus, a field isolated from free-living rats ileum, captured in the Metropolitan area in Mexico City, Mexico, and characterized using real time PCR and the API 50CHL system (Biomérieux, France) [41], were used.
The macrophage cell line RAW 264.7, gamma NO (-) (ATCC, USA) was used and third-stage larvae (L3) of H. contortus obtained from two parasite egg donor sheep (23.5 ± 2 kg body weight, BW), previously infected with 350 kg/PC infective larvae (INIFAP, Mexico),. Recovery of eggs from H. contortus was performed following the methodology described by Reséndiz et al., 2022 [49].
2.2. Animals
Ovine Columbia breed Free-grazing male adults were used to obtain larvae and abomasal tissue (23.5± 2 kg of body weight) (INIFAP, México). Sheep were maintained indoors in metabolic cages, and they were supplied with hay and commercial concentrate and water ad libitum. The animals were housed following the care/welfare guidelines of the Mexican Official Rule NOM-051-ZOO-1995 [50].
2.3. Obtaining and Characterization of MVs
Lactobacillus acidophilus strains, both field and reference, were grown in 250 mL of Man, Rogosa, Sharpe (MRS) broth (Becton Dickinson, Mexico) / 37 °C / 24 h, in the absence of CO2. In logarithmic growth phase (3.5 h), 10 µg/mL gentamicin was added to induce the release of MVs and incubated overnight. The culture was centrifuged at 2500 × g / 15 min at 4 °C to remove biomass and recover the supernatant, which was filtered with 0.45 µm and 0.22 µm diameter Millipore membranes to remove whole bacteria. The filtered supernatant (cell-free supernatant, CFS) was ultracentrifuged at 150 000 × g for 3 h at 4 °C. The resulting pellet corresponded to MVs, which were resuspended in 2 mL of 10 mM HEPES and frozen at -70 °C until use. Nanoparticle tracking analysis (NTA) was performed using a Nanosight NS 300 (Malvern Panalytical, UK). The 1 mL sample of MVs was diluted (1:100) in 1X phosphate-buffered saline (PBS) (filtered by 0.22 μm). Three 30 s videos were recorded with an sCMOS camera, and the particle size and concentration were registered.
2.4. Transmission Electron Microscopy (TEM)
The MVs were resuspended in cacodylate buffer and 10 to 15 μL placed on nickel grids previously treated with Formvar and shaded with carbon and allowed to set for 15 min (Electron Microscopy Sciences, USA). Afterwards, sample excess was removed with filter paper and the grid was carefully dried, adding 1% phosphotungstic acid (PTA, Electron Microscopy Sciences) at pH 6.0 for 90 s. Finally, samples were observed under a transmission electron microscope (JEM-1400, JEOL, Japan).
2.5. SDS-PAGE
Protein samples (25 µg) from MVs, whole bacteria (L. acidophilus), and CFS were analyzed on a 12% pre-cast acrylamide gel using the Mini-Protean R Tetra Cell 4 Gel System Kit (Bio-Rad, USA), following the manufacturer’s instructions. The dual-color molecular weight marker 1610374TGX (Bio-Rad, USA) was used and run at 100 volts for 2.5 h. Subsequently, the gel was stained using a Silver Stain Pierce kit (Thermo Scientific, USA) following the manufacturer’s instructions. The electrophoretic run was evaluated using the KODAK Gel-Logic 100 system software.
2.6. Obtaining of L3 Larva Stage from H. contortus
Fecal samples were obtained directly from the rectum of a male ovine donor. They were cultured in Petri dishes, following the Corticelli-Lai technique for 7 days [51]. Infective larvae were extracted from the fecal material using the Baermann funnel technique [52]. The larvae were cleaned using a distilled water, centrifuged [5000 g], and exsheathed with 0.187% sodium hypochlorite [52]. Finally, L3 was used for subsequent assays.
2.7. Culture of Abomasal Explants
Abomasal tissue was obtained from grazing ovine by careful washing with warm HANKs medium (CORNING, Spain). Circular sections of approximately 3 cm2 were subsequently cut and placed in six-well culture plates containing 5 mL of warm HANKs, then 5 mL syringes with the needle end removed were placed in the center of each tissue to provide a 13 mm diameter of an isolated zone containing H. contortus larvae, according to the technique described by Jackson et al. in 2004 [53]. MVs (30 μg) in 500 μL of 1X PBS of the field and ATCC strains were added to the mucosal surface within the isolation cylinder and incubated at 37 °C / 1 h. Subsequently, 0.5 mL of 1X PBS / 1,500 H. contortus larvae were introduced into the cylinder, and pressure was applied to the lid of the six-well plate to ensure an effective seal. Tissues with and without larvae were used as positive and negative association controls, respectively. Cultures were incubated for an additional 2 h at 37 °C and 5% CO2.
2.8. Recovery of L3 Larvae of H. contortus
After incubation, the syringe cylinders and tissues were washed with a 1X PBS solution in 50 mL conical tubes (wash tube) to remove all larvae not associated with the mucosa. The tissue was placed in a second tube (digestion tube) and digested in 50 mL of 1% pepsin solution in 1% HCl at 25 °C for 72 h. The number of larvae present in 2% aliquots of each sample was counted using a gridded Petri dish under a stereoscopic microscope (HINOTEK, NSZ-405, China) at 100x [53]. The percent of association in the tissue (digestion tube) was calculated.
2.9. Histopathology
After completion of the experiment, samples were taken from the region delimited by the syringe and fixed in 10% buffered formalin for 24 h. Subsequently, they were subjected to the paraffin technique and stained with hematoxylin and eosin (H&E), for reviewing under a light microscope.
2.10. Larval Mortality Assay
A larval mortality assay was performed using 96-well microtiter plates, employing 30 μg of MVs of both L. acidophilus strains, with distilled water as the negative control and 0.5% ivermectin as the positive control. An aqueous suspension (50 µL) containing 100 ± 15 infective larvae was added to each well. Subsequently, 50 µL of MVs and controls were added to each well individually. The plates were incubated at 25 °C in a humidified chamber for 24 h. The total number of larvae (live or dead) in each well was counted using a microscope [49]. Mortality percentages were estimated based on the criteria used by Olmedo et al. [54].
2.11. RAW 264.7 Cell Cultures
Cells of the RAW 264.7 line (1 X106) were cultured in 24-well culture plates with RPMI 1640 medium high in glucose (ATCC) supplemented with 10% fetal bovine serum (GIBCO, Spain), 2 mM L-glutamine (GIBCO), and 1% antibiotics (CAISSON, Spain). For negative control, cells were maintained without treatment, and as a positive control, 2 μg of LPS from E. coli 0111: B4 (Sigma Aldrich, Germany) were added. MVs (10 μg) of both L. acidophilus strains were used to stimulate the cells and incubated for 1 h at 37 °C in a 5% CO2 atmosphere. Following the incubation period, the cells were detached by raking and suspended in 1 mL of Trizol LS Reagent Ambion (Life Technologies, USA) to extract RNA, following the manufacturer’s instructions.
2.12. qPCR
cDNA was obtained using a NIPPON kit (Genetics EUROPE, Germany) following the instructions of the manufacturer. qPCR for IL-1β and TNF-α was performed using an Mx3005P Thermal Cycler (Agilent Technologies, USA). Primers were designed using Primer 3 software [55], primer sequences are listed in Table 1. The amplification conditions are briefly described: 1 denaturation cycle of 2 min at 95 °C, followed by 10 s 95 °C, 10 s 60 °C and 20 s 72 °C for 40 cycles, and melting curves was for 5 s 95 °C, 10 s 60 °C and 10 s 95 °C. Quantification was performed by relative expression using hypoxanthine-guanine phosphoribosyl-transferase enzyme (HPRT1) as reference gene which alignment temperature was 50 °C.
2.13. Statistical Analysis
Data were analyzed using one-way analysis of variance (ANOVA) and Tukey’s multiple comparison test. All statistical analyses were performed using the GraphPad Prism 8 software (Dotmatics). Statistical significance was established at p<0.05.
3. Results
3.1. Characterization of MVs by NTA and TEM
MVs from the field isolate of L. acidophilus have a size of 165.6 ± 9.2 nm in diameter at a concentration of 6.27 x 108 particles/mL, and MVs from the ATCC strain of L. acidophilus presented a size of 160.7 ± 11.0 nm at a concentration of 1.65 x 1010 particles/ml. Figure 1A and Figure 1B show the representative histograms obtained by nanoparticle tracking analysis (NTA) for both strains, which correspond to a representative sample of the field strain where homogeneity was observed (1C). This can be attributed to the coalescence capacity of the MVs during the NTA procedure. In the TEM image (1D), we observed MVs of the field strain, which showed an individual size of approximately 100 nm in diameter.
3.2. L. acidophilus MVs Express Proteins
SDS-PAGE of proteins found within the MVs, whole bacteria (WB), and cell-free supernatants (CFS) were achieved using a 12% polyacrylamide gel (Figure 2A). The molecular weights of the proteins obtained from the different runs were identified using the KODAK Gel-Logic 100 system software (Figure 2B). In comparison to previous studies [8], proteins associated with probiotic functions (p40 and 045), as well as cell division (FtsZ, DivlB), ABC transporters (ABC transporter ATP-binding and membrane spanning protein, as well as Glutamine ABC transporter permease protein) and possible bacteriocins (bacteriocin like proteins) were potentially identified [8]. Some of these proteins were observed in both the MVs of both strains, the whole bacteria and CFS (Table 2).
3.3. Tissue Stimulated with L. acidophilus MVs Decrease Larvae Association
Experiments were performed by stimulating abomasal tissue explants from grazing sheep with 30 μg of MVs from field and reference L. acidophilus challenged with 1,500 L3 larvae of H. contortus. Figure 3 shows a significant decrease (p<0.0001 and p<0.0085) in the percentage of association in the groups stimulated with MVs of both strains, which was lower than that of the positive control (withouth MVs) when a one-way ANOVA statistical test was applied.
3.4. Stimulation with the Field L. acidophilus MVs Increase the Presence of Inflammatory Cells in the Abomasal Tissue
Histological sections were prepared and stained with H&E to evaluate the explants in contact with MVs and larvae. An abomasal tissue of grazing sheep was observed and presented foci of lymphocytic infiltrate to the mucosa and to a lesser extent macrophages (Figure 4). Histopathological examination of the abomasal mucosa without any treatment shows normal autolysis for the villi due to the past after dead time (Figure 4A). The treated abamosal mucosa with the L3 larvae revealed reactive germinal centers at the submucosal level (Figure 4B) as same as with the MVs from L. acidophilus (Figure 4C), with the difference that under 10x and 40x (Figure 4D and 4E respectively) objectives, is demonstrating the presence of inflammatory cells infiltrating the interstitium of the crypts and villi (Figure 4F).
3.5. L. acidophilus MVs Do Not Kill the L3 Larvae of H. contortus
To determine whether the decrease in the percentage of association observed was due to the effect of L. acidophilus MVs on L3 larvae, a larvae mortality assay was performed. In the positive control group treated with 0.05% ivermectin, the highest percentage of L3 death (98%) was observed, while the negative control group treated with water showed less than 10% (Figure 5). The mortality percentages of the groups treated with MVs from both strains were similar (6% for ATCC and 4% for the field strain) to those of the negative control group, although a significant difference was observed (p<0.0001; one-way ANOVA) with respect to the group treated with ivermectin.
3.6. L. acidophilus MVs Activate Macrophages (RAW 264.7)
When macrophages of the RAW 264.7 cell line (Figure 6A) stimulated with MVs of both L. acidophilus strains were cultured, we observed that the cells modified their morphology, showing a greater number of pseudopodia (Figure 6B), suggesting macrophage activation. After incubating the cells for 1 h, RNA was obtained, and qPCR was performed. An increase in the expression of IL-1β (Figure 6C) and TNF-α (Figure 6D) was found, compared with the positive control group, which received LPS. In addition, relatively higher messenger expression was observed upon stimulation with L. acidophilus MVs from the field strain compared to those from the ATCC strain.
4. Discussion
Rats are importat carriers of pathogenic bacteria that affect both humans and animals, such as Salmonella, Escherichia coli, Campylobacter, Listeria, among others [56]. However, in most cases they do not show signs or symptoms of infection, due in part to their microbiota [56,57]. Studies focused on the microbiota of both free-living and laboratory rats describe Lactobacillus as one of the most abundant genera along the gastrointestinal tract [58,59], due to the importance of this genera it was used in the present study.
Numerous studies have reported beneficial effects of L. acidophilus [1,4,5,8,27,28,36]. In the present study, we have demonstrated that MVs from this bacterium could stimulate an immune response in the analyzed biological models.
The production of L. acidophilus MVs was confirmed by TEM and NTA, with diameters of approximately 160 nm, like those previously reported by Dean et al. [14] for L. acidophilus ATCC 53544, ranging from 100–150 nm. Studies analyzing proteins carried in MVs of different bacterial genera, both Gram-positive and Gram-negative, have demonstrated the great capacity of these structures to conform to proteins from the wall or the external membrane of origin, as well as proteins from the cytoplasm or in transit between the periplasm and lumen of MVs during formation [6,60,61]. The proteins carried in the MVs of both L. acidophilus strains were analyzed by SDS-PAGE, which demonstrated that a significant number of protein bands of different molecular weights were shared between them. Dean et al., [14] characterized L. acidophilus ATCC 53544 MVs and identified more than 80 protein components, including bacteriocins and ABC transporters. In the present study, low-molecular-weight proteins ranging from 10 kDa to 6 kDa were identified, like those reported by Dean as possible bacteriocins, which were named bacteriocin-like substances (BLS) [14,62]. In addition, proteins of 32 kDa and 64 kDa that, due to their molecular weight, coincided with the cell division protein (DivIB) and FtsZ protein [homologous to eukaryotic tubulin], respectively, which are known to interfere with cell division [14,63]. The functions of the 40 kDa and 75 kDa proteins in the complete bacterium are reported to be p40 and p75, respectively, which are associated with cell communication, epithelial adhesion, anti-apoptotic effects, and immunomodulatory capacity [64,65,66]. Likewise, proteins of 54 kDa and 59 kDa were identified that could correspond to ABC transporters, ABC transporter ATP-binding, membrane-spanning protein, and Glutamine ABC transporter permease protein, respectively, all of which are important for bacterial metabolism [14,62]. However, none of the proteins reported were sequenced and identified in this study.
In explants of ovine abomasal tissue, the association of L3 of H. contortus with MVs was evaluated. Over the 3-hour duration of the experiment, penetration of the larvae into the mucosa was not observed; therefore, for this study, we will only refer to an association phenomenon. L. acidophilus MVs from a field isolate and an ATCC reference strain inhibited the association of L3 larvae of H. contortus to the ovine abomasal explant tissue. Huang (2021), Martin (2020), and Gao (2009) reported that bacteriocins purified from both Gram-positive and Gram-negative bacteria affected other pathogenic bacterial genera and inhibit viruses and parasites [60,67,68]. For example, AS-48 is a peptide synthesized by Enterococcus faecalis that has a bactericidal effect on many bacteria and reduces the number of Trypanosoma cruzi by causing depolarization of the mitochondrial membrane and favoring the production of reactive oxygen species in the parasite [60,67,68]. Importantly, no reports have been recorded on the effects of the bacteriocins present in L. acidophilus MVs on any parasite.
To test the effect of LAB MVs on the viability of L3 of H. contortus, a larval mortality assay was performed. The MVs did not affect the larva viability, as the percentage of mortality was the same as that of the negative control, all the contrary to what was observed with ivermectin, which showed a mortality rate of 98% in the larvae, since it is a potent antiparasitic effective against H. contortus [69]. This indicates that the decrease in larval association with abomasal tissue could be due to MVs stimulation of the abomasal mucosa rather than their direct effect on the larvae. The metabolic derivatives of LAB contain various biologically active antigens, such as antimicrobial peptides and immune system signaling compounds (PAMPs) [70,71,72], to which the observed effects can be attributed. In 2007, Terefe et al. reported that the abomasal mucosa can synthesize pro-inflammatory cytokines (IL-4, IL-5, and IL-13) upon H. contortus infection [73]. IL-13 favors mucosal contractility and mucus production in the tissue, reducing the mobility of L3 larvae, which contributes to the non-association of the parasite [74].
Ovine abomasal tissue analyzed by histopathology showed the presence of mononuclear cells in the interstitium of the crypts and base of the villi in explants previously stimulated with L. acidophilus MVs. Simpson et al. [75] demonstrated from histopathological evaluation of abomasal tissue sections from animals infected with L3 of H. contortus the presence of foci of inflammatory infiltrates confined to the submucosa. Ex vivo studies have described the presence of immune cells in the submucosa of the abomasum without the infiltrate moving to the mucosa. In contrast, the capacity of the MVs in this study to stimulate immune cells favored their migration, which is like the report by Li (2005), where the supernatants of L. acidophilus cultures contained only cellular structures and metabolic derivatives that favored chemotaxis and proliferation of immune cells as well as angiogenesis in chicken embryonic tissues in vitro [76].
Ovine are generally prone to gastrointestinal parasitism because they feed on pastures contaminated with L3 stage larvae of H. contortus. In Mexico, its prevalence ranges from 17.5% to 57% among free-grazing animals [77,78,79]. In the animals used in the present study, adult nematodes of H. contortus were found in the abomasum of grazing animals, which explains the abundant inflammatory infiltrates found in the histological sections [80]. Constant exposure to H. contortus in ovine enhances the development of resistance to subsequent infections by this and other parasites. This is because the host immune system rapidly rejects the association of L3 by a phenomenon known as immune exclusion, wherein immune cells such as T lymphocytes and mast cells release protein compounds such as galectin and histamine that prevent the establishment of L3 in the crypts of the mucosa [80,81,82] by immunomodulation mechanisms. This phenomenon could explain the observed percentage of L3 association described in the present study, as well as that reported in others in vivo [83], in vitro [53], and ex vivo [81] studies.
The stimulation of the host immune system is one of the main functions of probiotics and their derivatives. In the present study, it was demonstrated that L. acidophilus MVs from the field strain can stimulate the RAW 264.7 murine cell macrophages. Morphological changes were observed in cells stimulated with both LPS (control +) and MVs, suggesting cell activation, such as the extension of pseudopodia, which is characteristic of this cell line [84]. The expression of transcripts encoding pro-inflammatory cytokines, such as IL-1β and TNF-α, was identified. IL-1β and TNF-α are secreted mainly by macrophages and have a synergistic effect on inflammation in infectious and non-infectious processes. Their main effects are chemotaxis of immune cells, cell activation, and stimulation of the production of cytokines such as IL-6, IL-8, and MCP-1 [85].
The presence of complete bacteria and some antigens secreted by them activates macrophages to synthesize proinflammatory cytokines [86,87,88]. One of the most studied components is peptidoglycan, the main component of the cell walls of Gram-positive bacteria. As MVs are formed from the elongation of the membrane, they carry structural components, including those of the cell wall [14], which act as PAMPs responsible for stimulating the cells in vitro. The whole cell of L. acidophilus has been reported to favor the expression of IL-1β in macrophages in vitro [90,91]. However, Li et al. in 2005 found that CFS of L. acidophilus cells could increase TNF-α expression in chick embryonic tissues [71]. Studies have been performed with the MVs of other strains of Lactobacillus, as is the case of Vargoorani, who used MVs derived from L. casei, observing an increase in the production of IL-6 and not IL-1β in cultures of human intestinal epithelial cells [21]. In mice orally administered with L. plantarum, TNF-α and IL-6 levels decreased [92]. These studies demonstrate that probiotic bacteria and their derivatives can modulate the immune system and the inflammatory response by increasing or decreasing the production of cytokines. Although they all belong to the same genus not all species act in the same manner.
The results obtained in this study demonstrate that the MVs of the field isolate of L. acidophilus have a probiotic capacity by stimulating the host immune system, similar to that reported with the use of whole bacteria. This allowed us to consider the possibility of using these compounds to prevent and control infections. MVs have the advantage of not having the capacity to replicate; therefore, they cannot cause infection and can be administered to immunocompromised patients or patients treated with antibiotics, contrary to what has been reported with the use of probiotic bacteria, where sepsis occurs even after their administration [70,92,93].
5. Conclusions
Lactobacillus acidophilus MVs isolated from free-living rats were shown to have probiotic and immunostimulatory effects on abomasal ovine tissue, decreasing the association with L3 larvae of H. contortus. While in cell culture of a murine macrophage line (RAW 274.6), MVs activated the cells by increasing the expression of proinflammatory cytokine transcripts IL-1β and TNF-α. These results suggest that L. acidophilus MVs possess the similar immunostimulatory effects as those reported for the parent bacteria, opening new perspectives for treatments that could improve the intestinal health in other mammals including humans. The expression and further encapsulation of proteins into MVs of Generally Recognized as Safe bacteria could represent a scientific novelty, with applications in food, nutraceuticals, and clinical therapies.
Author Contributions
Conceptualization, C.G.R; methodology, P.P.M(lead), V.G.E, C.A.G, C.L.S, R.H.P and F.G.D; software, A.V.R; validation, P.P.M and C.G.R; formal analysis, P.P.M and E.M.F; investigation, P.P.M; resources, C.G,R H.R.A, R.H.P and M.G.A; data curation, P.P.M; writing—original draft preparation, C.G.R; writing—review and editing, P.P.M, H.R.A and C.G.R; visualization, P.P.M; supervision, C.G.R and H.R.A; project administration, C.G.R; funding acquisition, C.G.R.
Funding
This research was funded by the Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT/ IT201521) and the Programa Interno de Cátedras de Investigación 2024 / PICI-CI2465. Dr. Pamela Izaret Pérez Martínez was beneficiary of a postdoctoral fellowship from the Dirección General de Asuntos del Personal Académico (DGAPA), all programs of the Universidad Nacional Autónoma de México (UNAM).
Institutional Review Board Statement
The animal study protocol was approved by the Internal Committee for the Care and Use of Experimental Animals of the Facultad de Estudios Superiores Cuautitlán [CICUAE-FESC] of UNAM [registration number: C 23_12].
Data Availability Statement
The data from the results obtained in this investigation are available on ZENODO repository with the DOI: 10.5281/zenodo.10999677.
Acknowledgments
We thank the biologists. María de Lourdes Rojas Morales from the Advanced Microscopy Laboratory for her support with the transmission electron microscopy micrographs, as well as the Cell Biology Department where the cell cultures were performed, both from to the Centro de Investigación y de Estudios Avanzados del IPN (CINVESTAV).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MVs | Microvesicles |
| GIT | Gastrointestinal tract |
| L3 | Stage 3 larva |
| WB | Whole bacteria |
| CFS | Cell-free supernatant |
| BLS | Bacteriocin-like substances |
| FI | Field Isolated |
References
- Santacroce, L.; Charitos, I.A.; Bottalico, L. A successful history: probiotics and their potential as antimicrobials. Expert Rev. Anti Infect. Ther 2019, 17, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Ibnou-Zekri, N.; Blum, S.; Schiffrin, E.J.; von der Weid, T. Divergent patterns of colonization and immune response elicited from two intestinal Lactobacillus strains that display similar properties in vitro. Infect Immun 2013, 71, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Behnsen, J.; Sassone-Corsi, M.; Raffatellu, M. Probiotics: Properties, Examples, and Specific Applications. Cold Spring Harb Perspect Med 2013, 3, a010074. [Google Scholar] [CrossRef] [PubMed]
- Morelli, L.; Capurso, L. FAO/WHO Guidelines on Probiotics: 10 Years Later. J Clin Gastroenterol 2001, 46, S1–S2. [Google Scholar] [CrossRef]
- Corthesy, B.; Gaskins, H.R.; Mercenier, A. Cross-talk between probiotic bacteria and the host immune system. J Nutr 2007, 137(3 Suppl 2), 781S–90S. [Google Scholar] [CrossRef]
- Ruiz, L.; Hevia, A.; Bernardo, D.; Margolles, A.; Sánchez, B. Extracellular molecular effectors mediating probiotic attributes. FEMS Microbiol Lett 2014, 359, 1–11. [Google Scholar] [CrossRef]
- Al-Nedawi, K.; Mian, M.F.; Hossain, N.; Karimi, K.; Mao, Y.K.; Forsythe, P.; Min, K.K.; Stanisz, A.M.; Kunze, W.A.; Bienenstock, J. Gut commensal microvesicles reproduce parent bacterial signals to host immune and enteric nervous systems. FASEB J 2015, 29, 684–695. [Google Scholar] [CrossRef]
- Kulp, A.; Kuehn, M.J. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu Rev Microbiol 2010, 64, 163–184. [Google Scholar] [CrossRef]
- Toyofuku, M.; Nomura, N.; Eberl, L. Types and origins of bacterial membrane vesicles. Nat Rev Microbiol 2019, 17, 13–24. [Google Scholar] [CrossRef]
- Wang, X.; Eagen, W.J.; Lee, J.C. Orchestration of human macrophage NLRP3 inflammasome activation by Staphylococcus aureus extracellular vesicles. Proc Natl Acad Sci U S A 2020, 117, 3174–3184. [Google Scholar] [CrossRef]
- Choi, J.H.; Moon, C.M.; Shin, T.S.; Kim, E.K.; McDowell, A.; Jo, M.K.; Joo, Y.H.; Kim, S.E.; Jung, H.K.; Shim, K.N.; Jung, S.A.; Kim, Y.K. Lactobacillus paracasei-derived extracellular vesicles attenuate the intestinal inflammatory response by augmenting the endoplasmic reticulum stress pathway. Exp Mol Med 2020, 52, 423–437. [Google Scholar] [CrossRef] [PubMed]
- May, A. L.; Corona, A. I.; Jiménez, A. L. L.; Cortés, N. G.; Vera, R. J. Sensibilidad y Resistencia a Antibióticos de Cepas Probióticas Empleadas en Productos Comerciales. ESJ 2020, 16, 43. [Google Scholar] [CrossRef]
- Hao, H.; Zhang, X.; Tong, L.; Liu, Q.; Liang, X.; Bu, Y.; Gong, P.; Liu, T.; Zhang, L.; Xia, Y.; Ai, L.; Yi, H. Effect of Extracellular Vesicles Derived From Lactobacillus plantarum Q7 on Gut Microbiota and Ulcerative Colitis in Mice. Front Immunol 2021, 2, 12–777147. [Google Scholar] [CrossRef] [PubMed]
- Dean, S.N.; Leary, D.H.; Sullivan, C.J.; Oh, E.; Walper, S.A. Isolation and characterization of Lactobacillus-derived membrane vesicles. Sci Rep 2019, 9, 877. [Google Scholar] [CrossRef]
- West, C.L.; Stanisz, A.M.; Mao, Y.K.; Champagne-Jorgensen, K.; Bienenstock, J.; Kunze, W.A. Microvesicles from Lactobacillus reuteri (DSM-17938) completely reproduce modulation of gut motility by bacteria in mice. PLoS One 2020, 7;15(1), e0225481. [CrossRef]
- Domínguez, A.P.; Martínez, J.H.; Martínez, D.C.; Coluccio, F.; Piuri, M.; Pérez, O.E. Lactobacillus casei BL23 Produces Microvesicles Carrying Proteins That Have Been Associated with Its Probiotic Effect. Front Microbiol 2017, 20, 8–1783. [Google Scholar] [CrossRef]
- Jan, A.T. Outer Membrane Vesicles (OMVs) of Gram-negative Bacteria: A Perspective Update. Front. Microbiol 2017, 8, 1053. [Google Scholar] [CrossRef]
- Wang, X.; Thompson, C.D.; Weidenmaier, C.; Lee, J.C. Release of Staphylococcus aureus extracellular vesicles and their application as a vaccine platform. Nat. Commun 2018, 9, 1379. [Google Scholar] [CrossRef]
- Patten, D.A; Hussein, E.; Davies, S.P.; Humphreys, P.N.; Collett, A. Commensal-derived OMVs elicit a mild proinflammatory response in intestinal epithelial cells. Microbiology (Reading) 2017, 163(5), 702–711. [Google Scholar] [CrossRef]
- Kang, C.S.; Ban, M.; Choi, E.J.; Moon, H.G.; Jeon, J.S.; Kim, D.K.; Park, S.K.; Jeon, S.G.; Roh, T.Y.; Myung, S.J.; Gho, Y.S.; Kim, J.G.; Kim, Y.K. Extracellular vesicles derived from gut microbiota, especially Akkermansia muciniphila, protect the progression of dextran sulfate sodium-induced colitis. PLoS One 2018, 8, e76520. [Google Scholar] [CrossRef]
- Vargoorani, M.E.; Modarressi, M.H.; Vaziri, F.; Motevaseli, E.; Siadat, S.D. Stimulatory effects of Lactobacillus casei derived extracellular vesicles on toll-like receptor 9 gene expression and cytokine profile in human intestinal epithelial cells. IJDMD 2020, 19, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.M.; Awanye, A.M.; Marsay, L.; Dold, C.; Pollard, A.J.; Rollier, C.S.; Feavers, I.M.; Maiden, M.C.J.; Derrick, J.P. Application of a Neisseria meningitidis antigen microarray to identify candidate vaccine proteins from a human Phase I clinical trial. Vaccine 2022, 40, 3835–3842. [Google Scholar] [CrossRef] [PubMed]
- Leitner, D.R.; Lichtenegger, S.; Temel, P.; Zingl, F.G.; Ratzberger, D.; Roier, S.; Schild-Prüfert, K.; Feichter, S.; Reidl, J.; Schild, S. A. A combined vaccine approach against Vibrio cholerae and ETEC based on outer membrane vesicles. Front. Microbiol 2015, 6, 823. [Google Scholar] [CrossRef]
- Nieves, W.; Petersen, H.; Judy B. M., A. Burkholderia pseudomallei outer membrane vesicle vaccine provides protection against lethal sepsis. Clin. Vaccine Immunol 2014, 21, 747–754. [Google Scholar] [CrossRef]
- Ayalew, S.; Confer, A.W.; Shrestha, B.; Wilson, A.E.; Montelongo, M. Proteomic analysis and immunogenicity of Mannheimia haemolytica vesicles. Clin. Vaccine Immunol 2013, 20, 191–196. [Google Scholar] [CrossRef]
- De la Garza, M.; González, C.; Suárez, F.; Tenorio, V.; Trigo, F. Biológico vacunal elaborado a partir de Microvesículas (MVs) de Mannheimia haemolytica serotipo A2 de administración intranasal en ovinos. (Mexico pathent Nomber 341611). IMPI 2016. https://siga.impi.gob.mx/newSIGA/content/common/principal.jsf.
- Dhama, K.; Verma, V.; Sawant, P.; Tiwari, R.; Vaid, R.; Chauhan, R. Applications of probiotics in poultry: enhancing immunity and beneficial effects on production performances and health: a review. Immunol. Immunopathol 2011, 13, 1–19. [Google Scholar]
- Hoseinifar, S.H.; Sun, Y.Z.; Wang, A.; Zhou, Z. Probiotics as means of diseases control in aquaculture, a review of current knowledge and future perspectives. Front. Micrrobiol 2018, 9, 2429. [Google Scholar] [CrossRef]
- Di Gioia, D.; Biavati, B. Probiotics and Prebiotics in Animal Health and Food Safety Switzerland. Springer International Publishing 2018, 273. [Google Scholar] [CrossRef]
- Zhong, Y.; Wang, S.; Di, H.; Deng, Z.; Liu, J.; Wang, H. Gut health benefit and application of postbiotics in animal production. J. Anim. Sci. Biotechnol 2022, 13, 38. [Google Scholar] [CrossRef]
- Raheem, A.; Liang, L.; Zhang, G.; Cui, S. Modulatory Effects of Probiotics During Pathogenic Infections with Emphasis on Immune Regulation. Front. Immunol 2021, 12, 616713. [Google Scholar] [CrossRef]
- Zhang, R.; Zhou, M.; Tu, Y.; Zhang, N.F.; Deng, K.D.; Ma, T.; Diao, Q.Y. Effect of oral administration of probiotics on growth performance, apparent nutrient digestibility and stress-related indicators in Holstein calves. J. Anim. Physiol. Anim. Nutr 2015, 100, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Huang, W.; Hou, Q.; Kwok, L.Y.; Sun, Z.; Ma, H.; Zhao, F.; Lee, Y.K.; Zhang, H. The effects of probiotics administration on the milk production, milk components and fecal bacteria microbiota of dairy cows. Sci. Bull. 2017, 62, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.Q.; Chang, J.; Zuo, R.Y.; Chen, L.Y.; Chen, Q.X.; Wei, X.Y.; Guan, Q.F.; Sun, J.W.; Zheng, Q.H.; Yang, X.; Ren, G.Z. Effect of the transformed lactobacillus with phytase gene on pig production performance, nutrient digestibility, gut microbes, and serum biochemical indexes. AJAS 2010, 23, 246–252. [Google Scholar] [CrossRef]
- Xin, J.; Zeng, D.; Wang, H.; Sun, N.; Zhao, Y.; Dan, Y.; Pan, K.; Jing, B.; Ni, X. Probiotic Lactobacillus johnsonii BS15 Promotes Growth Performance, Intestinal Immunity, and Gut Microbiota in Piglets. Probiotics Antimicro 2020, 12, 184–193. [Google Scholar] [CrossRef]
- Yang, J.; Wang, C.; Huang, K.; Zhang, M.; Wang, J.; Pan, X. Compound Lactobacillus sp. administration ameliorates stress and body growth through gut microbiota optimization on weaning piglets. Appl. Microbiol. Biotechnol 2020, 104, 6749–6765. [Google Scholar] [CrossRef]
- Iraqi, E.K.G.; Fayed, R.H. Effect of yeast as feed supplement on behavioural and productive performance of broiler chickens. J. Life Sci 2012, 9, 4026–4031. [Google Scholar]
- Forte, C.; Manuali, E.; Abbate, Y.; Papa, P.; Vieceli, L.; Tentellini, M.; Trabalza, M.; Moscati, L. Dietary Lactobacillus acidophilus positively influences growth performance, gut morphology, and gut microbiology in rurally reared chickens. Poult. Sci 2018, 97, 930–936. [Google Scholar] [CrossRef]
- Anee, I.J.; Alam, S.; Begum, R.A.; Shahjahan, R.; Khandaker, A. The role of probiotics on animal health and nutrition. JOBAZ 2021, 82, 52. [Google Scholar] [CrossRef]
- Jensen, H.; Grimmer, S.; Naterstad, K.; Axelsson, L. In vitro testing of commercial and potential probiotic lactic acid bacteria. Int. J. Food Microbiol 2012, 153, 216–222. [Google Scholar] [CrossRef]
- Lonngi, C. Evaluación del efecto de microvesículas (MVs) de bacterias ácido lácticas (BAL), en cultivos de Salmonella typhimurium ATCC 154 y Escherichia coli de campo. Bachelor’s degree, Universidad Nacional Autónoma de México, State of Mexico 2022. Avaiable on the institutional repository of the UNAM https://tesiunam.dgb.unam.mx/F/UE2HK7UPD3Q2NV8BG4VCT8GNALMUPXXVYPVIFF5225NLCRDT3A-40114?
- Zhang, S.D.; Lin, G.H.; Han, J.R.; Lin, Y.W.; Wang, F.Q.; Lu, D.C.; Xie, J.X.; Zhao, J.X. Digestive Tract Morphology and Gut Microbiota Jointly Determine an Efficient Digestive Strategy in Subterranean Rodents: Plateau Zokor. Animals (Basel) 2022, 12, 2155. [Google Scholar] [CrossRef]
- He, W.Q.; Xiong, Y.Q.; Ge, J.; Chen, Y.X.; Chen, X.J.; Zhong, X.S.; Ou, Z.J.; Gao, Y.H.; Cheng, M.J.; Mo, Y.; Wen, Y.Q.; Qiu, M.; Huo, S.T.; Chen, S.W.; Zheng, X.Y.; He, H.; Li, Y.Z.; You, F.F.; Zhang, M.Y.; Chen, Q. Composition of gut and oropharynx bacterial communities in Rattus norvegicus and Suncus murinus in China. BMC Vet. Res 2020, 16, 413. [Google Scholar] [CrossRef] [PubMed]
- Desvars, A.; Ruppitsch, W.; Lepuschitz, S.; Szostak, M.P.; Spergser, J.; Feßler, A.T.; Schwarz, S.; Monecke, S.; Ehricht, R.; Walzer, C.; Loncaric, I. Urban brown rats (Rattus norvegicus) as possible source of multidrug-resistant Enterobacteriaceae and meticillin-resistant Staphylococcus spp. Vienna, Austria, 2016 and 2017. Euro Surveill 2019, 24, 1900149. [Google Scholar] [CrossRef]
- Arsenopoulos, K.V.; Fthenakis, G.C.; Katsarou, E.I.; Papadopoulos, E. Haemonchosis: A challenging parasitic infection of sheep and goats. Animals 2021, 11, 363. [Google Scholar] [CrossRef] [PubMed]
- Besier, R.B.; Kahn, L.P.; Sargison, N.D.; Van, J.A. The pathophysiology, ecology and epidemiology of Haemonchus contortus infection in small ruminants. Adv. Parasitol 2016, 93, 95–143. [Google Scholar] [CrossRef]
- Emery, D.L.; Hunt, P.W.; Leo, F.L.J. Haemonchus contortus: The then and now, and where to from here? Int. J. Parasitol 2016, 46, 755–769. [Google Scholar] [CrossRef]
- Adduci, I.; Sajovitz, F.; Hinney, B.; Lichtmannsperger, K.; Joachim, A.; Wittek, T.; Yan, S. Haemonchosis in Sheep and Goats, Control Strategies and Development of Vaccines against Haemonchus contortus. Animals (Basel) 2022, 12, 2339. [Google Scholar] [CrossRef]
- Reséndiz, G.; Higuera, R.I.; Lara, A.; González, R.; Cortes, J.A.; González, M.; Mendoza, P.; Romero, S.G.; Olmedo, A. In Vitro Anthelmintic Activity of a Hydroalcoholic Extract from Guazuma ulmifolia Leaves against Haemonchus contortus. Pathog 2022, 11, 1160. [Google Scholar] [CrossRef]
- Gobierno de México/ SENASICA. Available online: https://www.gob.mx/senasica/documentos/nom-051-zoo-1995 (accessed on 26/03/2025).
- Corticelli, B.; Lai, M. Ricerche sulla tecnica di coltura delle larve infestive degli strongili gastro-intestinali del bovino. Acta Med. Vet 1963, 9, 347–357. [Google Scholar]
- Mesquita, J.R.; Mega, C.; Coelho, C.; Cruz, R.; Vala, H.; Estebez, S.; Santos, C.; Vasconcelos, C. EBC series on diagnostic parasitology part 3: The Baermann technique. Vet. Nurs. J 2017. [Google Scholar] [CrossRef]
- Jackson, F.; Greer, A.W.; Huntley, J.; McAnulty, R.W.; Bartley, D.J.; Stanley, A.; Stenhouse, L.; Stankiewicz, M.; Sykes, A.R. Studies using Teladorsagia circumcincta in an in vitro direct challenge method using abomasal tissue explants. Vet. Parasitol 2004, 124, 73–89. [Google Scholar] [CrossRef]
- Olmedo, A.; Rojo, R.; Zamilpa, A.; Mendoza, P.; Arece, J.; López, M.E.; von Son-de Fernex, E. In vitro larvicidal effect of a hydroalcoholic extract from Acacia cochliacantha leaf against ruminant parasitic nematodes. Vet. Res. Commun 2017, 41, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol 2000, 132, 365–86. [Google Scholar] [CrossRef] [PubMed]
- Meerburg, B. G.; Singleton, G. R.; Kijlstra, A. Rodent-borne diseases and their risks for public health. Crit Rev Microbiol 2009, 35, 221–270. [Google Scholar] [CrossRef] [PubMed]
- Čoklo, M.; Maslov, D.R.; Kraljević, S. Modulation of gut microbiota in healthy rats after exposure to nutritional supplements. Gut Microbes, 2020; 12, 1–28. [Google Scholar] [CrossRef]
- Brooks, S.P.; McAllister, M.; Sandoz, M.; Kalmokoff, M.L. Culture-independent phylogenetic analysis of the faecal flora of the rat. Can. J. Microbiol 2003, 49, 589–601. [Google Scholar] [CrossRef]
- Li, D.; Chen, H.; Mao, B.; Yang, Q.; Zhao, J.; Gu, Z.; Zhang, H.; Chen, Y.Q.; Chen, W. Microbial Biogeography and Core Microbiota of the Rat Digestive Tract. Sci. Rep 2017, 4;8, 45840. Sci. Rep. [CrossRef]
- Gao, B.; Rodriguez, M.d.C.; Lanz, H.; Zhu, S. AdDLP, a bacterial defensin-like peptide, exhibits anti-Plasmodium activity. BBRC 2009, 387, 393–398. [Google Scholar] [CrossRef]
- Collins, S.M.; Nice, J.B.; Chang, E.H.; Brown, A.C. Size Exclusion Chromatography to Analyze Bacterial Outer Membrane Vesicle Heterogeneity. Journal of Visualized Experiments. Vis Exp 2021, 169. [Google Scholar] [CrossRef]
- Dean, S.N.; Rimmer, M.A.; Turner, K.B.; Phillips, D.A.; Caruana, J.C.; Hervey, W. J 4th.; Leary, D.H.; Walper, SA. Lactobacillus acidophilus Membrane Vesicles as a Vehicle of Bacteriocin Delivery. Front. Microbiol 2020, 11, 710. [Google Scholar] [CrossRef]
- Margolin, W. FtsZ and the division of prokaryotic cells and organelles. Nat. Rev. Mol. Cell Biol 2005, 6, 862–871. [Google Scholar] [CrossRef]
- Bäuerl, C.; Coll, J. M.; Tarazona, C.; Pérez, G. Lactobacillus casei extracellular vesicles stimulate EGFR pathway likely due to the presence of proteins P40 and P75 bound to their surface. Sci. Rep 2020, 10. [Google Scholar] [CrossRef]
- Raman, M.; Ambalam, P.; Kondepudi, K.K.; Pithva, S.; Kothari, C.; Patel, A.T.; Purama, R.K.; Dave, J.M.; Vyas, B.R. Potential of probiotics, prebiotics and synbiotics for management of colorectal cancer. Gut Microbes 2013, 4, 181–92. [Google Scholar] [CrossRef]
- Claes I., J.; Schoofs, G.; Regulski, K.; Courtin, P.; Chapot, M.P.; Rolain, T.; Hols, P.; von Ossowski, I.; Reunanen, J.; de Vos, W.M.; Palva, A.; Vanderleyden, J.; De Keersmaecker, S.C.; Lebeer, S. Genetic and biochemical characterization of the cell wall hydrolase activity of the major secreted protein of Lactobacillus rhamnosus GG. PLoS One 2012, 7, e31588. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Teng, K.; Liu, Y. Bacteriocins: Potential for Human Health. Oxid. Med. Cell. Longev 2021, 10, 5518825. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Cebrián, R.; Maqueda, M.; Romero, D.; Rosales, M.J.; Sánchez, M.; Marín, C. Assessing the effectiveness of AS-48 in experimental mice models of Chagas’ disease. J. Antimicrob. Chemother 2020, 75, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Puspitasari, S.; Farajallah, A.; Sulistiawati, E.; Muladno. Effectiveness of Ivermectin and Albendazole against Haemonchus contortus in Sheep in West Java, Indonesia. Trop. Life. Sci. Res 2016, 27, 135–44. [Google Scholar]
- Moradi, M.; Kousheh, S.A.; Almasi, H.; Alizadeh, A.; Guimarães, J.T.; Yılmaz, N.; Lotfi, A. Postbiotics produced by lactic acid bacteria: The next frontier in food safety. Compr. Rev. Food. Sci. Food Saf 2020, 19, 3390–3415. [Google Scholar] [CrossRef]
- Moradi, M.; Mardani, K.; Tajik, H. Characterization and application of postbiotics of Lactobacillus spp. on Listeria monocytogenes in vitro and in food models. LWT 2019, 111, 457–464. [Google Scholar] [CrossRef]
- Rajakovich, L. J.; Balskus, E. P. Metabolic functions of the human gut microbiota: The role of metalloenzymes. Nat. Prod. Rep 2019, 36, 593–625. [Google Scholar] [CrossRef]
- Terefe, G.; Lacroux, C.; Andreoletti, O.; Grisez, C.; Prevot, F.; Bergeaud, J.P.; Penicaud, J.; Rouillon, V.; Gruner, L.; Brunel, J.C.; Francois, D.; Bouix, J.; Dorchies, P.; Jacquiet, P. Immune response to Haemonchus contortus infection in susceptible (INRA 401) and resistant (Barbados Black Belly) breeds of lambs. Parasite Immunol 2007, 29, 415–24. [Google Scholar] [CrossRef]
- Shepherd, E.; Greiner, S.P.; Russ, B.; Bowdridge, S.A. Interleukin-13 induces paralysis of Haemonchus contortus larvae in vitro. Parasite Immunol 2020, 42, e12758. [Google Scholar] [CrossRef]
- Simpson, H.V.; Umair, S.; Hoang, V.C.; Savoian, M.S. Histochemical study of the effects on abomasal mucins of Haemonchus contortus or Teladorsagia circumcincta infection in lambs. Vet. Parasitol 2016, 226, 210–221. [Google Scholar] [CrossRef]
- Li, W.I.; Brackett, B.G.; Halper, J. Culture supernatant of Lactobacillus acidophilus stimulates proliferation of embryonic cells. Exp. Biol. Med. (Maywood) 2005, 230, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Hernández, S.; Segura, I.; Olivares, P.J.; Almazán, M.T. Prevalencia de nematodos gastrointestinales en ovinos en pastoreo en la parte alta del MPIO. de Cuetzala del Progreso, Guerrero-México. Redvet 2007, 8, 1–7. [Google Scholar]
- Solís, J.; Gaxiola, S.; Enríquez, I.; Portillo, J.; López, G.; Castro, N. Factores ambientales asociados a la prevalencia de Haemonchus spp en corderos de la zona centro de Sinaloa. Abanico veterinario 2021, 11. [Google Scholar] [CrossRef]
- González, R.; Córdova, C.; Torres, G.; Mendoza, P.; Arece, J. Prevalencia de parásitos gastrointestinales en ovinos sacrificados en un rastro de Tabasco, México. Veterinaria México 2011, 42, 125–135. [Google Scholar]
- Balic. A.; Cunningham, C.P.; Meeusen, E.N. Eosinophil interactions with Haemonchus contortus larvae in the ovine gastrointestinal tract. Parasite Immunol 2006, 28, 107–115. [Google Scholar] [CrossRef]
- Kemp, J.M.; Robinson, N.A.; Meeusen, E.N.; Piedrafita, D.M. The relationship between the rapid rejection of Haemonchus contortus larvae with cells and mediators in abomasal tissues in immune sheep. Int. J. Parasitol 2009, 39, 1589–1594. [Google Scholar] [CrossRef]
- Yang, X.; Khan, S.; Zhao, X.; Zhang, J.; Nisar, A.; Feng, X. Suppression of hyaluronidase reduces invasion and establishment of Haemonchus contortus larvae in sheep. J. Vet. Res 2020, 51, 106. [Google Scholar] [CrossRef]
- Miller. H.R.; Jackson. F.; Newlands. G.; Appleyard. W.T. Immune exclusion, a mechanism of protection against the ovine nematode Haemonchus contortus. Res. Vet. Sci 1983, 35, 357–363.
- Xiaoyan, X.; Hongxia, S.; Jiamin, G.; Huicheng, C.; Ye, L.; Qiang, X. Antimicrobial peptide HI-3 from Hermetia illucens alleviates inflammation in lipopolysaccharide-stimulated RAW 264.7 cells via suppression of the nuclear factor kappa-B signaling pathway. Microbiol. Immunol 2023, 67, 32–43. [Google Scholar] [CrossRef]
- Weber, A.; Wasiliew, P.; Kracht, M. Interleukin-1 (IL-1) pathway. Sci. Signaling 2020, 105, cm1. [Google Scholar] [CrossRef]
- Ellis, T.N.; Leiman, S.A.; Kuehn, M.J. Naturally produced outer membrane vesicles from Pseudomonas aeruginosa elicit a potent innate immune response via combined sensing of both lipopolysaccharide and protein components. Infect. Immun 2010, 78, 3822–31. [Google Scholar] [CrossRef]
- Briaud, P.; Carroll, R.K. Extracellular Vesicle Biogenesis and Functions in Gram-Positive Bacteria. Infect Immun 2020, 88, e00433–20. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Descripción general de la familia IL-1 en la inflamación innata y la inmunidad adquirida. Immunol. Rev 2018, 281, 8–27. [Google Scholar] [CrossRef]
- Matsubara, V.H.; Ishikawa, K.H.; Ando, E.S.; Bueno, B.; Nakamae, A.E.M.; Mayer, M.P.A. Probiotic Bacteria Alter Pattern-Recognition Receptor Expression and Cytokine Profile in a Human Macrophage Model Challenged with Candida albicans and Lipopolysaccharide. Front. Microbiol. 2017, 8, 2280. [Google Scholar] [CrossRef]
- Yanagihara, S.; Goto, H.; Maywood, Hirota, T. ; Fukuda, S.; Ohno, H.; Yamamoto, N. Lactobacillus acidophilus L-92 Cells Activate Expression of Immunomodulatory Genes in THP-1 Cells. Biosci. Microbiota Food Health 2014, 33, 157–164. [Google Scholar] [CrossRef]
- Le, B.; Anh, P.T.N.; Yang, S.H. Enhancement of the anti-inflammatory effect of mustard kimchi on RAW 264.7 macrophages by the Lactobacillus plantarum fermentation-mediated generation of phenolic compound derivatives. Foods 2020, 9, 181. [Google Scholar] [CrossRef]
- Vinderola, G.; Sanders, M.E.; Salminen, S. The Concept of Postbiotics. Foods 2022, 11, 1077. [Google Scholar] [CrossRef]
- Tsilingiri, K.; Barbosa, T.; Penna, G.; Caprioli, F.; Sonzogni, A.; Viale, G.; Rescigno, M. Probiotic and postbiotic activity in health and disease: comparison on a novel polarised ex-vivo organ culture model, Gut 2012, 61, 1007–1015. [CrossRef]
Figure 1.
Characterization of extracellular microvesicles (MVs) secreted by Lactobacillus acidophilus. Concentration and size distribution [nm] of MVs produced by field (A) and ATCC (B) L. acidophilus. (C) Representative frame of one of the videos of L. acidophilus in NanoSight where MVs are observed (white arrow). (D) TEM micrograph of MVs produced by field L. acidophilus stained with phosphotungstic acid (white arrow).
Figure 1.
Characterization of extracellular microvesicles (MVs) secreted by Lactobacillus acidophilus. Concentration and size distribution [nm] of MVs produced by field (A) and ATCC (B) L. acidophilus. (C) Representative frame of one of the videos of L. acidophilus in NanoSight where MVs are observed (white arrow). (D) TEM micrograph of MVs produced by field L. acidophilus stained with phosphotungstic acid (white arrow).
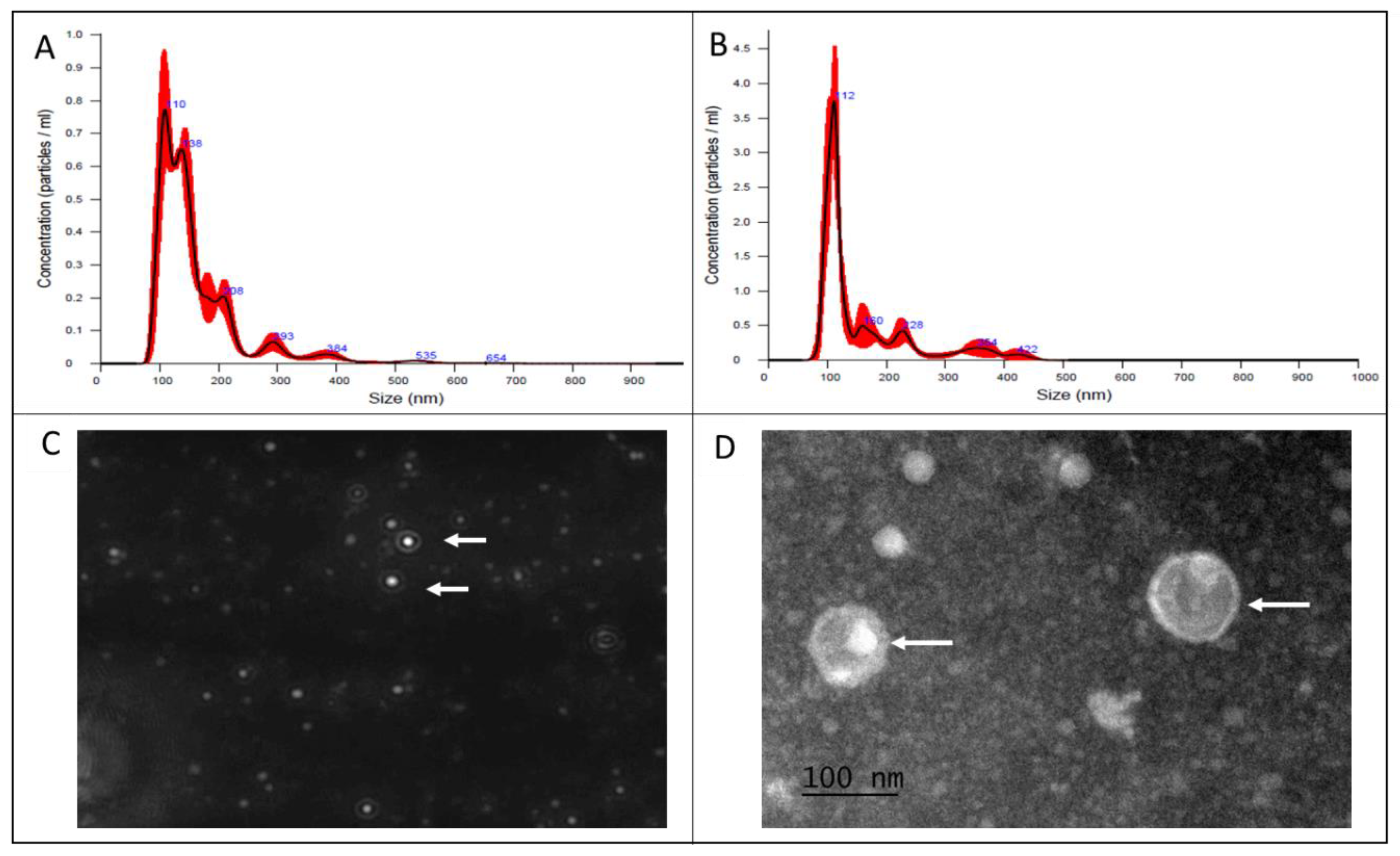
Figure 2.
Protein composition of different cell fractions of Lactobacillus acidophilus (A) SDS-PAGE gel (12%), stained with silver, of proteins from both field (FI) and ATCC strains of L. acidophilus from MVs (FI MVS and ATCC MVS) whole bacteria (FI WB and ATCC WB) and cell-free supernatant (FI CFS and ATCC CFS). In all lanes, 25 μg of protein was placed. (B) SDS-PAGE 12% gel analysis, with KODAK Gel-Logic 100 system software, where proteins detected by the software are marked with white lines.
Figure 2.
Protein composition of different cell fractions of Lactobacillus acidophilus (A) SDS-PAGE gel (12%), stained with silver, of proteins from both field (FI) and ATCC strains of L. acidophilus from MVs (FI MVS and ATCC MVS) whole bacteria (FI WB and ATCC WB) and cell-free supernatant (FI CFS and ATCC CFS). In all lanes, 25 μg of protein was placed. (B) SDS-PAGE 12% gel analysis, with KODAK Gel-Logic 100 system software, where proteins detected by the software are marked with white lines.
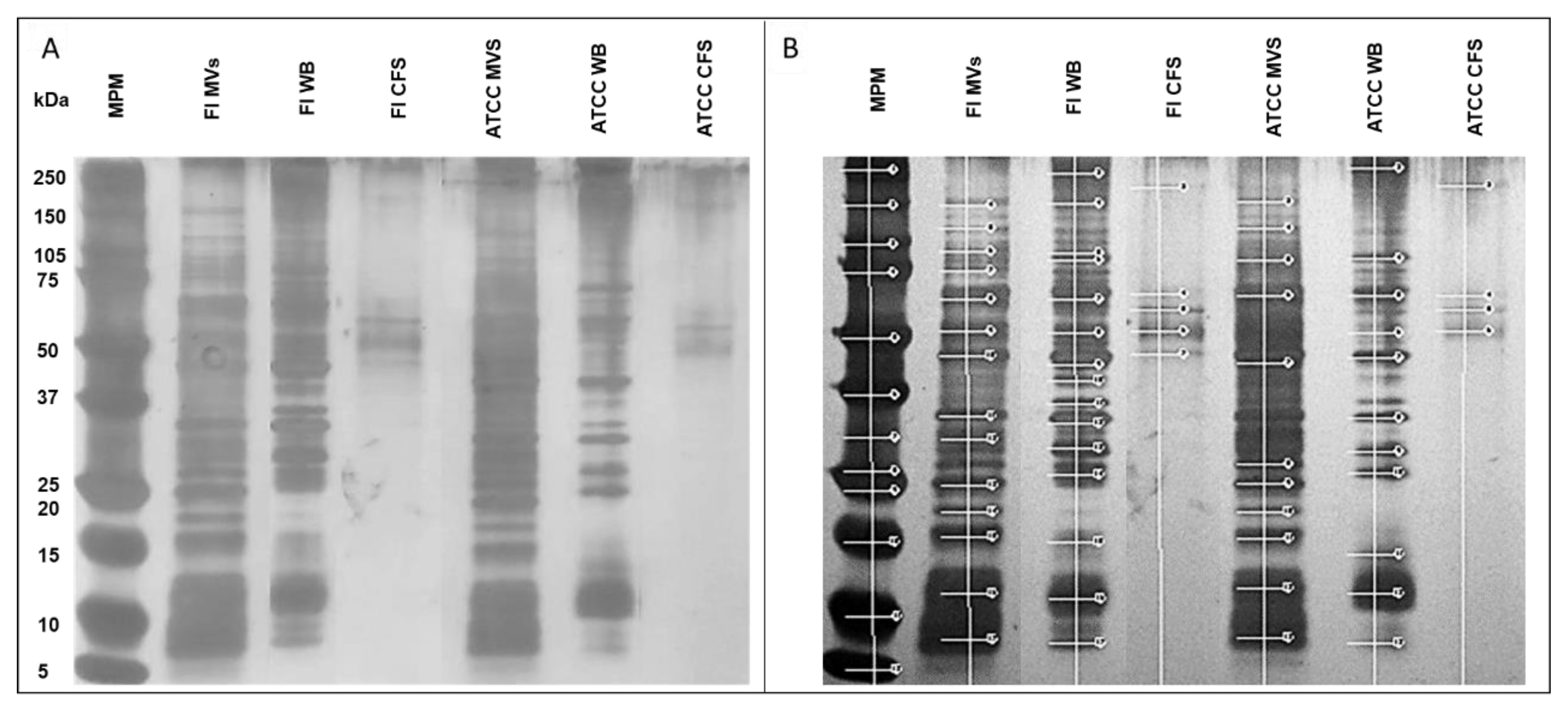
Figure 3.
Percentage of Haemonchus contortus L3 association in ovine abomasal tissue. Association of H. contortus L3 larvae in abomasal tissue of grazing ovine where abomasal mucosa only (CTRL-), without stimulation of MVs (CTRL+) and stimulated with 30μg of MVs of L. acidophilus field strain (FI) and ATCC strain, a significant decrease in the percentage of association is appreciated. One-way ANOVA p=0.001(****).
Figure 3.
Percentage of Haemonchus contortus L3 association in ovine abomasal tissue. Association of H. contortus L3 larvae in abomasal tissue of grazing ovine where abomasal mucosa only (CTRL-), without stimulation of MVs (CTRL+) and stimulated with 30μg of MVs of L. acidophilus field strain (FI) and ATCC strain, a significant decrease in the percentage of association is appreciated. One-way ANOVA p=0.001(****).
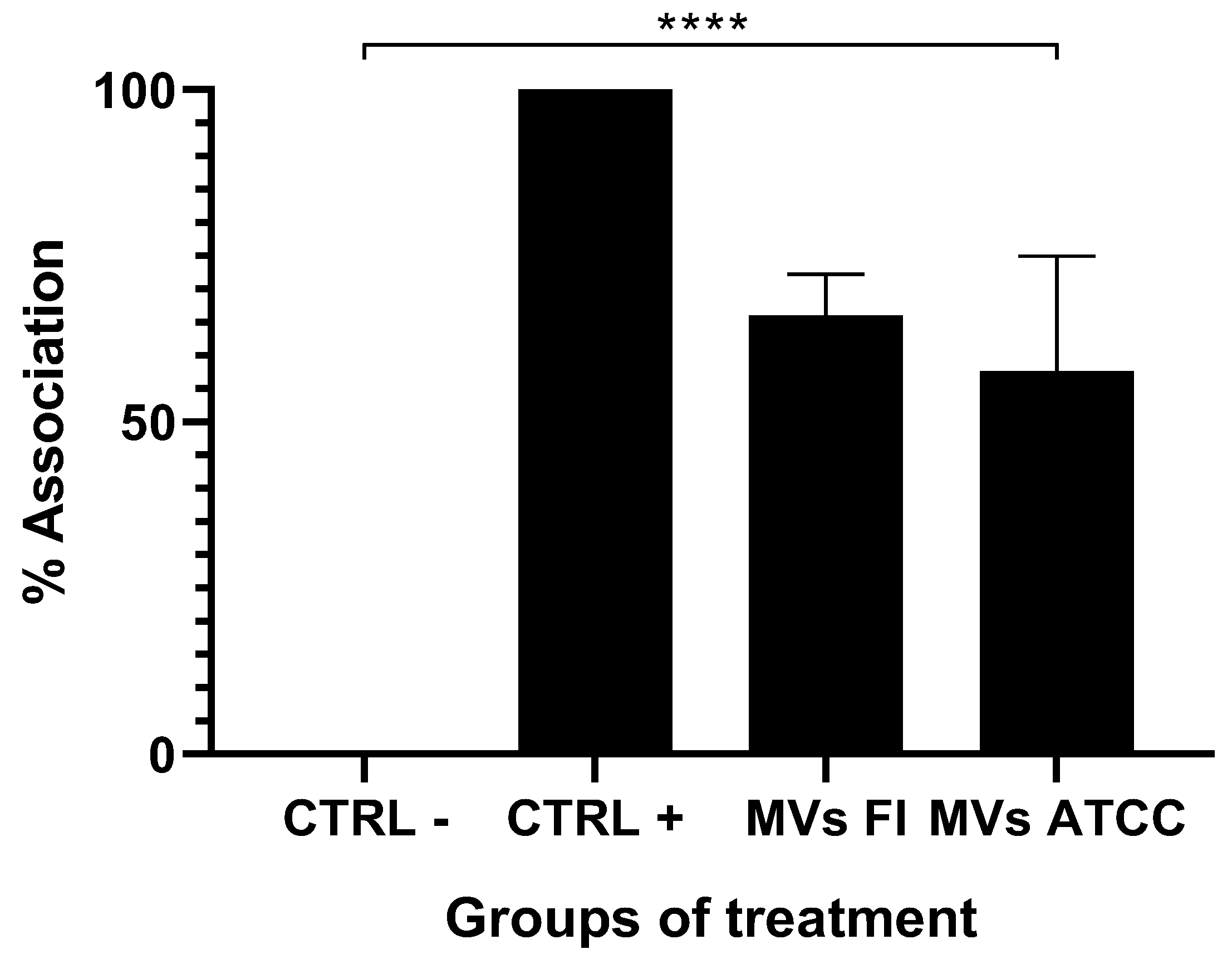
Figure 4.
Sections of ovine abomasal tissue stimulated with MV of Lactobacillus acidophilus stained with H&E. Abomasal tissue from free grazing ovine. (A) Unstimulated tissue (Ctrl -), (B) Tissue challenged with 1500 L3 of H. contortus (Ctrl +), (C) Tissue stimulated with MVS of L. acidophilus field strain and ATCC, (D) Tissue stimulated with MVS of L. acidophilus field strain and ATCC. Mucosal-associated lymphoid tissue in B, C and D panels is marked with an oval. (E) With 40X objective, the presence of lymphocytic cells and some macrophages in the submucosal area of the tissue challenged with L3 of H. contortus, and (F) infiltrate in the crypts of the tissue stimulated with L. acidophilus MVs from field strain.
Figure 4.
Sections of ovine abomasal tissue stimulated with MV of Lactobacillus acidophilus stained with H&E. Abomasal tissue from free grazing ovine. (A) Unstimulated tissue (Ctrl -), (B) Tissue challenged with 1500 L3 of H. contortus (Ctrl +), (C) Tissue stimulated with MVS of L. acidophilus field strain and ATCC, (D) Tissue stimulated with MVS of L. acidophilus field strain and ATCC. Mucosal-associated lymphoid tissue in B, C and D panels is marked with an oval. (E) With 40X objective, the presence of lymphocytic cells and some macrophages in the submucosal area of the tissue challenged with L3 of H. contortus, and (F) infiltrate in the crypts of the tissue stimulated with L. acidophilus MVs from field strain.
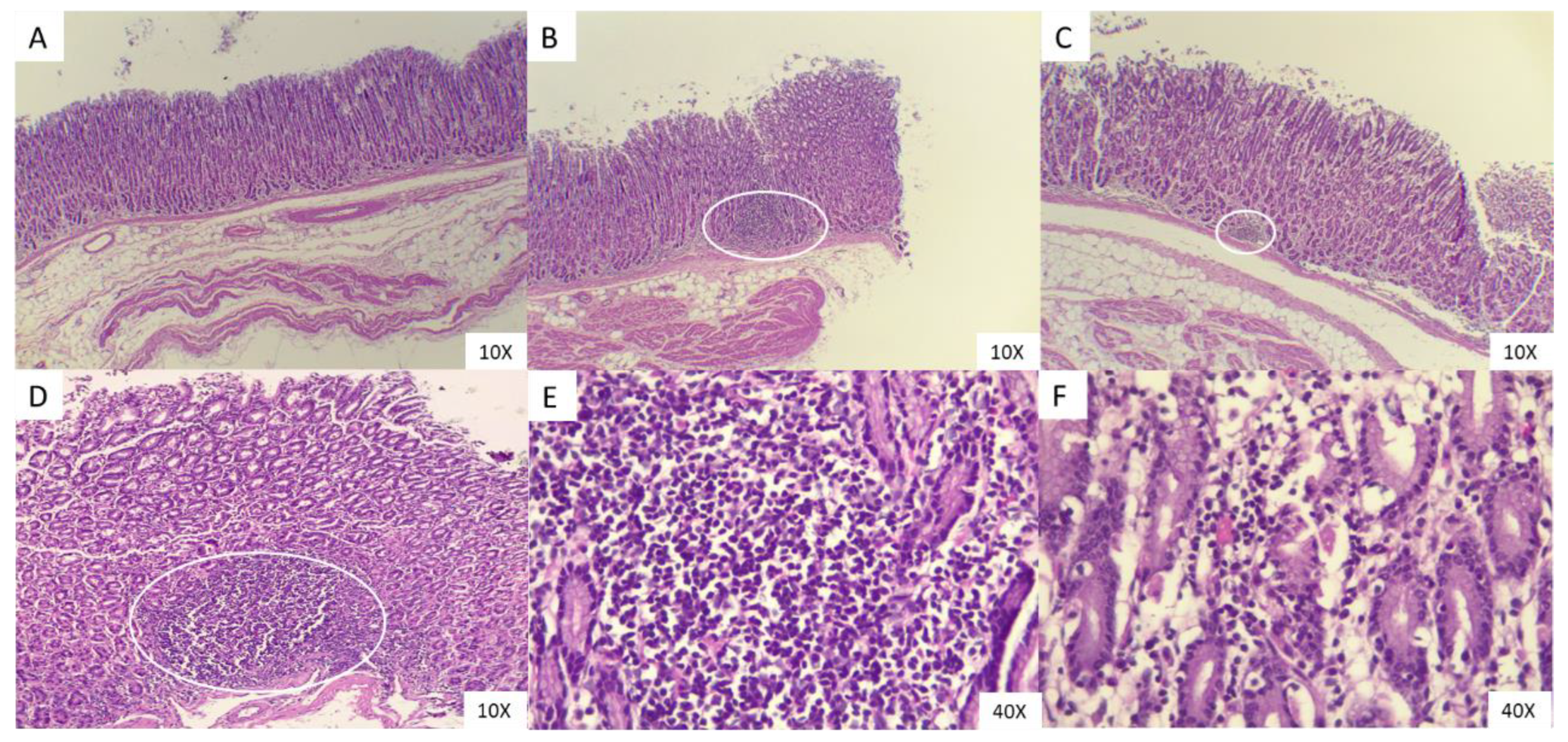
Figure 5.
Percentage mortality of Haemonchus contortus L3. Larvae mortality assay of H. contortus L3 incubated with 30 μg of MVs of L. acidophilus field strain (FI) and ATCC strain. As a positive control, 0.05% ivermectin (IVM) was used and showed a 98% mortality of the larvae, reaching a significant difference with the negative control (water) and the treatments with both L. acidophilus MVs treatments. One-way ANOVA p=0.001(****).
Figure 5.
Percentage mortality of Haemonchus contortus L3. Larvae mortality assay of H. contortus L3 incubated with 30 μg of MVs of L. acidophilus field strain (FI) and ATCC strain. As a positive control, 0.05% ivermectin (IVM) was used and showed a 98% mortality of the larvae, reaching a significant difference with the negative control (water) and the treatments with both L. acidophilus MVs treatments. One-way ANOVA p=0.001(****).
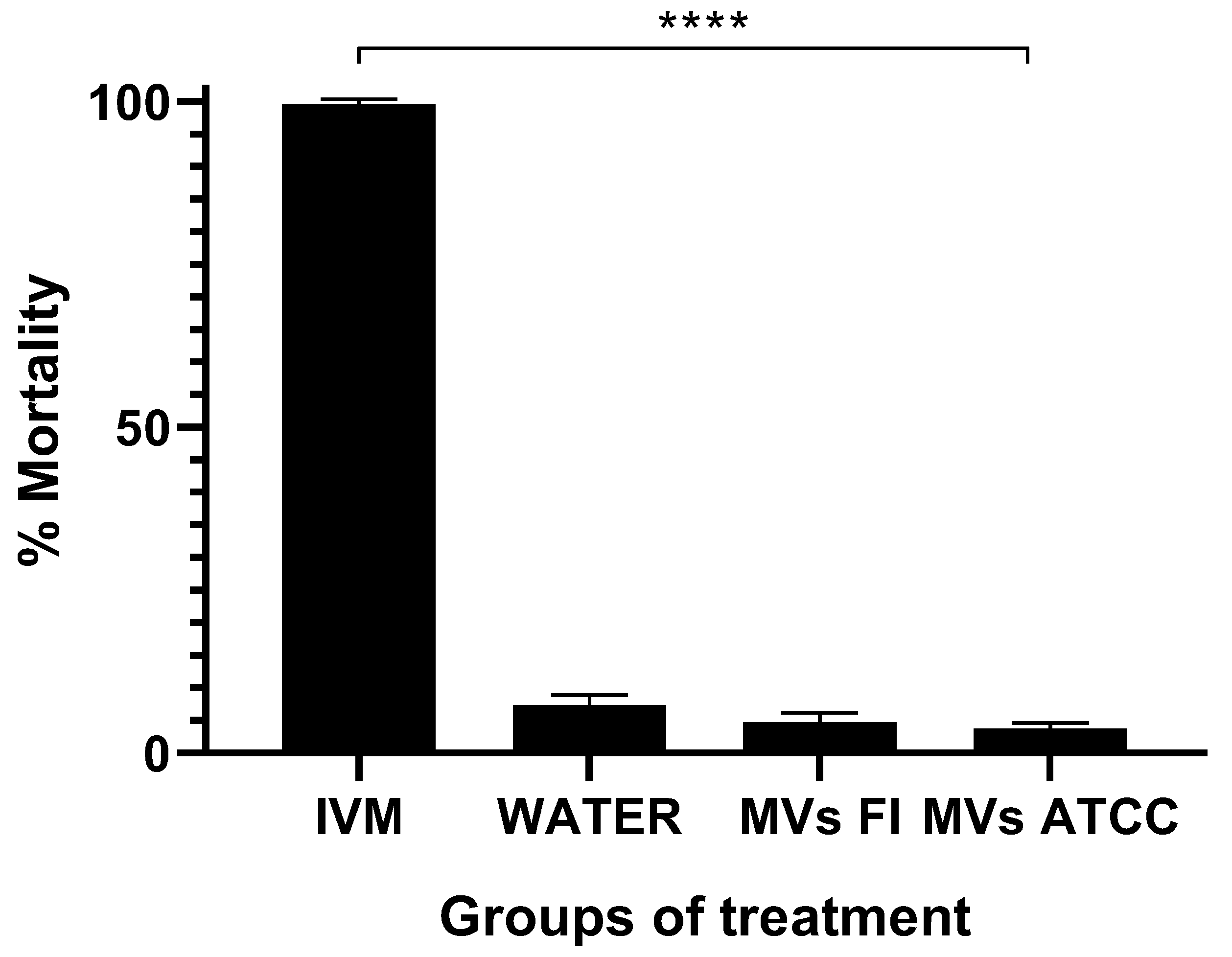
Figure 6.
RAW 264.7 cells stimulated with Lactobacillus acidophilus MVs and relative expression of IL-1β and TNF-α mRNA. RAW 264.7 cells (A) untreated (negative control), (B) stimulated with 10 μg of MVs obtained from a field strain (MVs FI). In activated macrophages more pseudopods are observed (black arrow), (C) Relative mRNA expression of TNF-α and (D) Relative mRNA expression of IL-1β. Stimulation with MVs increases the relative messenger expression of both cytokines respect to the negative control. Tukey’s multiple comparisons test p= 0.0044 (**). Messenger RNA (mRNA), Interleukin 1 beta (IL-1β), Tumor necrosis factor alpha (TNF-α).
Figure 6.
RAW 264.7 cells stimulated with Lactobacillus acidophilus MVs and relative expression of IL-1β and TNF-α mRNA. RAW 264.7 cells (A) untreated (negative control), (B) stimulated with 10 μg of MVs obtained from a field strain (MVs FI). In activated macrophages more pseudopods are observed (black arrow), (C) Relative mRNA expression of TNF-α and (D) Relative mRNA expression of IL-1β. Stimulation with MVs increases the relative messenger expression of both cytokines respect to the negative control. Tukey’s multiple comparisons test p= 0.0044 (**). Messenger RNA (mRNA), Interleukin 1 beta (IL-1β), Tumor necrosis factor alpha (TNF-α).
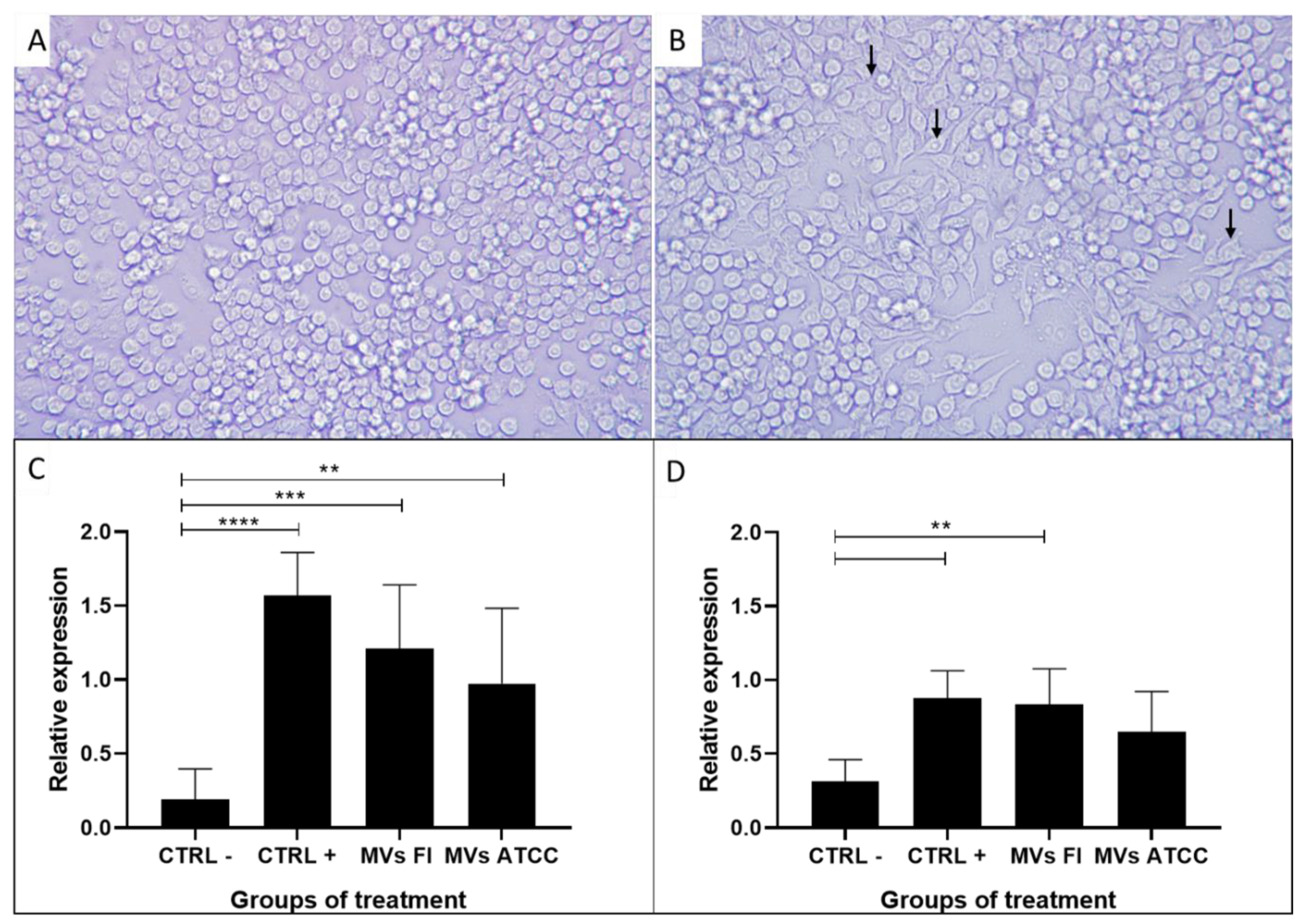

Table 1.
Primer sequence for cells RAW264.7 genes.
| Primer gene | Sequence [5’- 3’] | Alignment temperature | Length [bp] |
|---|---|---|---|
| IL-1β | FwGTGTGTGACGTTCCCATTA Rv CGTTGCTTGGTTCTCCTTGT | 62°C | 170 |
| TNF-α | FwTATGGCTCAGGGTCCAACTC Rv CTCCCTTTGCAGAACTCAGG | 62°C | 174 |
| HPRT1 | FwATTCCCAACAGACAGACAGACAGAA Rv TTAGGTCGGAAGGCATCAT | 50° C | 224 |
Fw – forward primer; Rv – reverse primer; IL 1β– interleukin one beta; TNFα – tumor necrosis factor alpha; HPRT1 – hypoxanthine-guanine phosphoribosyltransferase enzyme; bp – base pairs.
Table 2.
Proteins shared by MVs, whole bacteria, and cell-free supernatants of field and ATCC Lactobacillus acidophilus.
Table 2.
Proteins shared by MVs, whole bacteria, and cell-free supernatants of field and ATCC Lactobacillus acidophilus.
| Proteins | Fractions | ||||||
|---|---|---|---|---|---|---|---|
| Description | PM/ kDa |
FI MVs | FI WB | FI CFS | ATCC MVS | ATCC WB | ATCC CFS |
| p75 | 75 | ✓ | ✓ | - | ✓ | ✓ | - |
| FtsZ protein | 64 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ |
| ABC transporter ATP-binding and membrane spanning protein | 59 | ✓ | - | - | - | - | - |
| Glutamine ABC transporter permease protein glnP | 54 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ |
| p40 | 40 | ✓ | - | - | ✓ | - | - |
| Cell division protein DivIB | 32 | ✓ | - | - | ✓ | - | - |
| Bacteriocin like proteins BLP | 10 | ✓ | ✓ | - | ✓ | ✓ | - |
| 9 | ✓ | ✓ | - | ✓ | ✓ | - | |
| 6 | ✓ | - | - | ✓ | - | - | |
The table shows the proteins of both L. acidophilus strains, field strain [FI] and ATCC present [✓] or absent [-] obtained from the fractions: MVs [FI MVS and ATCC MVS], whole bacteria [FI WB and ATCC WB] and cell-free supernatant [FI CFS and ATCC CFS]. By analyzing the molecular weights reported by the KODAK Gel-Logic 100 system software, the proteins were compared with those reported in the literature.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



