
Submitted:
24 April 2025
Posted:
24 April 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
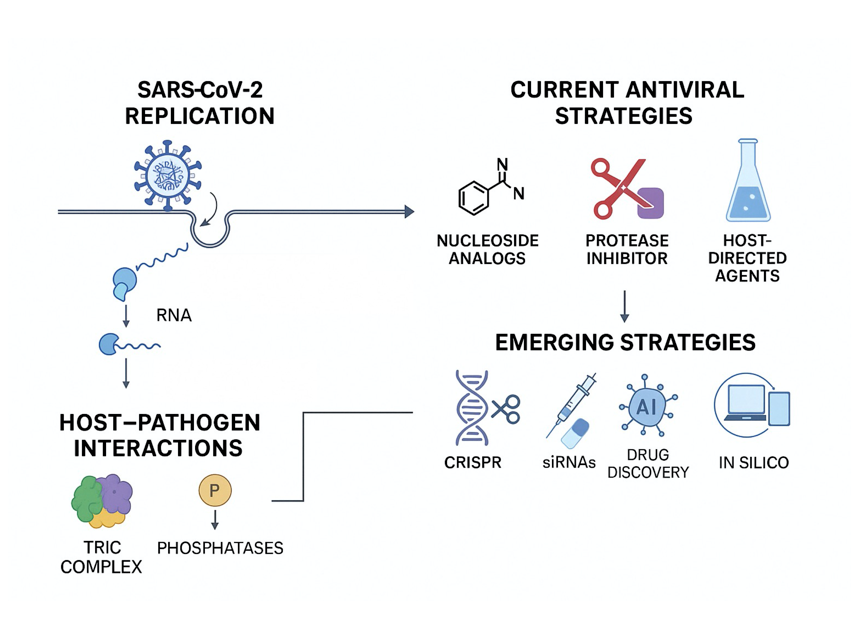
SARS-CoV-2 replication remains a critical and main target for therapeutic interventions. The current review synthesizes existing knowledge to provide an in-depth analysis of molecular insights and both current and emerging therapy methods, moving past reviews centered on antivirals and general replication processes. We examined antiviral tactics aimed at these replication pathways, including direct-acting nucleoside analogs (remdesivir, molnupiravir), protease inhibitors (nirmatrelvir), and host-directed agents influencing viral entry and RNA synthesis. Emphasizing therapeutic constraints and evolutionary escape, this study also investigates synergistic drug combinations and resistance mechanisms. Discussed for their capacity to efficiently handle next coronavirus dangers are emerging methods—from CRISPR-based gene-silencing, nanoparticle-delivered siRNAs, and AI-driven drug discovery. Highlighted as new antiviral targets are host-pathogen interactions including adaptation via the TRiC complex and phosphatase pathways. This paper offers a road map for improving therapeutic tactics against SARS-CoV-2 and related developing viruses with molecular virology, pharmacology, and computational biology.
Keywords:
SARS-CoV-2
; RNA-dependent RNA polymerase
; viral replication
; COVID-19
; host–pathogen interactions
; drug discovery and development
; antivirals
; AI drug discovery
1. Introduction
The global outbreak of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2 in late 2019 has resulted in a pandemic called coronavirus illness 2019 (COVID-19) that has massively affected both the health and economic sectors. On March 21, 2023, an estimated 761 million people were reportedly infected by the said virus. With this number, approximately 6.8 million fatalities were reported [1].
This has led for COVID-19 to be one of the most catastrophic global health concerns of the new millennium. In a short span of time, the scientific community made extraordinary progress in understanding the virus and finding countermeasures. These include elucidation of the life cycle of the virus and the viral protein structures in the first year of the global outbreak onset. Over 20 high-resolution structures of the viral RNA polymerase complex were reported. This has resulted in an in-depth knowledge of the replication mechanism and methods to inhibit it [2,3,4].
Building upon the initial groundwork studies on the molecular basis of SARS-CoV-2 replication, it has ultimately opened a new research avenue in designing future therapeutic interventions. Based on its viral structure, SARS-CoV-2 is similar to other coronaviruses. It is an enveloped positive-sense RNA virus that has a quite large ~30 kb genomic size, one of the largest of any RNA virus. In comparison, planarian secretory cell nidovirus (PSCNV) has a genome size of 41.1 kb [5], rotaviruses has about 26–30 kb [6], ebolavirus has about ~19 kb [7], and the influenza virus has about ~13.5 kb [8,9].
The viral replication process involves a multi-protein replication/transcription complex that allows the replication of the RNA genome within the infected host cell. The virus interacts with host cellular pathways, which in turn can elude immune defenses. The virus uses host resources for its replication, which leads to dysregulation of the host’s normal host cell functions [8].
In the past five years, antiviral strategies targeting the viral replication cycle have been studied as either developed or repurposed drugs. In particular, direct-acting antivirals such as remdesivir and molnupiravir [10], and protease inhibitors (nirmatrelvir in Paxlovid) that indirectly affect replication through the inhibition of the viral replicase protein processing [11].
The present review offers a summary of the SARS-CoV-2 replication molecular mechanism and host cell interactions, while assessing the potential impact of this information in the development of future therapeutic strategies. The review examines both wet laboratory experimental findings and computational approaches that have expanded our present understanding of the viral replication mechanism. In brief, the review includes the molecular mechanism of viral RNA replication, viral-host interactions, and current and emerging antiviral strategies. The paper also looks at the future directions in this field by highlighting potential novel targets and possible innovative antiviral strategies. In whole, this integrative approach aims to synthesize the current advancements in SARS-CoV-2 replication as a guide for future emerging research undertakings.
2. Molecular Mechanisms of SARS-CoV-2 Replication
SARS-CoV-2 shares a similar genomic organization and replication method as those of other coronaviruses. Upon entrance in the host cell, the single-stranded positive-sense RNA genome of the virus directly works as an mRNA and has a 5′ cap and 3′ poly(A) tail [8,12,13]. Sixteen non-structural proteins (nsp; nsp1-16) comprise the viral replication and transcription machinery. These proteins are autocatalytically cleaved by the viral proteases coming from two large polyprotein (pp1a and pp1ab) that are encoded by the 5′ two-thirds of the genome (open reading frames ORF1a and ORF1ab)[14,15].
Meanwhile, the spike (S), Envelope (E), Membrane (M), and nucleocapsid (N) proteins serve as auxiliary and structural proteins. These proteins are encoded by the remaining 3′ region of the genome. Furthermore, these proteins are expressed from the sub-genomic mRNAs generated during the infection process (Figure 1) [15,16,17].
2.1. Replication Complex and Enzymatic Machinery
The RNA-dependent RNA polymerase (RdRp) complex is a multimeric protein complex that is central to coronavirus replication. The minimum functional components for this multimeric complex are comprised by the nsp12-7-8 super-complex [20,21].
Dissecting the structure and function oof nsp12, the protein has a catalytic polymerase subunit that functions with a canonical “right-hand” polymerase domain containing, fingers, palms and thumb subdomains that allow RNA synthesis [8,22]. In addition, nsp12 has an N-terminal nidovirus-specific RNA-dependent nucleotidyltransferase domain (NiRAN). This plays an important function in viral replication and possibly plays a role in priming or capping [22,23].
The two co-factors, nsp7 and nsp8, form the active polymerase by binding with nsp12. Nsp12 has been reported to have little or marginal activity by itself. Two subunits of nsp8 and one nsp7 subunit form a complex with nsp12 to form a functional RdRp holoenzyme. This complex serves as a processivity factor that aids in the gripping of the RNA template and in stabilizing the polymerase during the viral replication process (Figure 2) [20,21].
Several other viral nsps play essential functions in genome replication and RNA processing. The replication-transcription complex (RTC) functions in copying the viral RNA. This includes the 5′-to-3′ helicase/translocase nsp13 that is responsible for the unwinding of the RNA secondary structure and moves along the template during RNA synthesis [24,25]. The nsp14 protein functions as a 3′–5′ exoribonuclease (ExoN) that removes misincorporate nucleotides. Thus, contributing to increased replication fidelity via the RTC’s proofreading mechanism [26].
This unusual proofreading capability of RNA viruses most likely helps coronaviruses retain their enormous genomes. Moreover, the N7-methyltransferase domain of nsp14 operates in conjunction with nsp10 [27]. nsp16 functions as a 2′-O-methyltransferase with nsp10 as a co-factor. This allows the 5′ end of the viral RNA to establish a cap structure [28]. In turn, this guarantees the efficient translation and stability of viral mRNAs. Other nsps, in particular nsp9 and nsp10, support the RTC through RNA binding and regulation [29].
2.2. Genome Replication Cycle
The multi-step replication of SARS-CoV-2 occurs in the cytoplasm of infected cells [30]. After viral entry and uncoating, the translation of the genomic RNA generates the replicase polyprotein (pp1a/pp1ab) [31]. These are then cleaved to produce nsps. These nsps rapidly form the RTC in the cellular membranes. A characteristic of coronavirus replication is the remodeling of the host intracellular membranes. This generates specific microenvironments for RNA reproduction [32].
A double-membrane vesicles (DMV) that are derived from the endoplasmic reticulum and work as a protected replication organelle is formed upon the induction of SARS-CoV-2 [33,34]. Nsp3 and nsp4, viral transmembrane proteins, bind to the endoplasmic reticulum and curate these vesicles. The formed nsp3/nsp4 complex bends the membranes into vesicles that recruit replication proteins [35,36].
The RdRp complex is tethered to the cytosolic face of DMVs and is concentrated in these compartments [29]. This is due to the N-terminal domain of nsp3, which can directly bind to nsp12 polymerase [8].
The RTC generates a complementary negative-sense RNA from the genomic RNA template within the DMV [13,37]. This negative strand functions as a template for the synthesis of new positive-sense genomic RNA, a series of sub-genomic RNAs that align with the 3′ regions of the genome, and supports the expression of structural and accessory proteins [13,38]. The polymerases engage in a distinct form of discontinuous transcription during sub-genomic mRNA replication. The RdRp intermittently moves from one portion of the negative-strand template to connect with the leader sequence at the 5′ end. This template-switching mechanism leads to the generation of sub-genomic RNAs that possess a 5′ leader while encoding distinct downstream open reading frames (ORFs) such as S, E, M, and N [38].
Although proofreading provides high fidelity, the virus continues to accumulate mutations over time [39,40]. This, along with RNA recombination events, has resulted in the emergence of various SARS-CoV-2 variants [39]. Following the synthesis of new genomic RNAs, these molecules are encapsulated within virions [41]. The nucleocapsid (N) protein associates with genomic RNA to create a ribonucleoprotein complex [42], which is subsequently enveloped by a lipid membrane that incorporates the S, E, and M proteins at assembly sites within the ER–Golgi intermediate compartment [43]. Progeny virions are transported within vesicles and subsequently released from the cell through exocytosis. An infected cell can release thousands of new virions, which go on to infect neighboring cells and, in an organism, can disseminate to other tissues [41,43].
2.3. Host Factors in Viral Replication
To fulfill its reproduction cycle, SARS-CoV-2 depends on the host proteins and cellular processes. The virus utilizes the host ribosomes for protein synthesis and relies on the host cell’s provision of nucleotides, amino acids, and energy.
Recent proteomic studies have indicated connections between SARS-CoV-2 non-structural proteins and the host proteins. In particular, nsp12 interacts with the host chaperonin complex, often referred to as TRiC (or CCT) [44].
TRiC supports the correct folding or assembly of the polymerase complex. This, in turn, allows reliance on the host for effective replication [45]. The P323L mutation of the nsp12 has been associated to influence this interaction. This nsp12 protein variant alongside the D614G spike mutant was found to have reduced dependence on the host phosphatase complex. This indicated a possible adaptive modification for enhanced replication in the host human cell [46]. These findings show that SARS-CoV-2 can adapt to its host cellular environment, which can lead to improving its replicative fitness [47]. In such cases, host pathways (including chaperones and phosphatases) might be targeted as host-directed antivirals [48,49]. Such findings underscore how SARS-CoV-2 can adapt to the host cellular environment and potentially improve its replicative fitness. They also point to host pathways (chaperones and phosphatases) that might be targeted by host-directed antivirals [44,46].
3. Drugs Targeting SARS-CoV-2 Replication
Efforts to create therapeutics that inhibit the replication of SARS-CoV-2 has been unprecedented. These drugs encompass direct-acting antivirals that target viral enzymes and host-targeting medications that indirectly impede viral propagation. Here, we discuss four groups of antiviral drugs that disrupt the SARS-CoV-2 replication cycle, along with aspects such as resistance and efficacy.
Therapeutics geared toward the inhibition of the replication of SARS-CoV-2 have been unprecedented in recent times. These drugs can be classified as direct-acting antivirals that target the viral replication complex and those that target host-target medications that indirectly impede viral replication and propagation.
3.1. Nucleoside Analogue Polymerase Inhibitors
Targeting the RdRp using nucleoside or nucleotide analogues has been a key tactic to stop viral replication [50]. Remdesivir was the inaugural antiviral medication authorized for COVID-19. It is a monophosphoramidate prodrug of an adenosine analogue; upon intracellular metabolism to the active remdesivir triphosphate (RTP), it competes with ATP and is integrated into viral RNA by nsp12 [44]. The integration of remdesivir into an elongating RNA strand leads to delayed chain termination, causing RNA synthesis to cease after the addition of many additional nucleotides [4] (Figure 3).
Structural studies have shown that remdesivir causes the RdRp to stall by preventing further translocation of the RNA template. In clinical trials, intravenous remdesivir moderately accelerated recovery in hospitalized COVID-19 patients and continues to be a routine treatment for severe cases [52,53]. Significantly, the virus’s proofreading function (nsp14) does not completely eliminate remdesivir, enabling it to maintain its antiviral effectiveness [54]. However, resistance may develop under selective pressure: in vitro studies have revealed mutations in nsp12 (notably at residues V166 and E802) that provide partial resistance to remdesivir [55]. More importantly, such mutants have not dominated in clinical isolates, possibly due to fitness costs and the relatively short treatment courses used.
Another nucleoside analogue that has been approved as an oral antiviral for COVID-19 is molnupiravir [56] .It is a prodrug of β-D-N4-hydroxycytidine that behaves as a mutagenic agent. Unlike remdesivir, molnupiravir does not result in chain termination. Rather, its active form is incorporated into viral RNA in lieu of cytidine or uridine, causing the RdRp to make an erroneous base-pairing (it can pair with guanine or adenine) [57,58]. This leads to the accumulation of mutations in the viral genome, a process known as lethal mutagenesis (Figure 4).
Molnupiravir fundamentally induces errors in the virus, leading to the death of the viral population due to genomic instability [60]. Clinical trials with patients with mild-to-moderate COVID-19 demonstrated that a 5-day oral regimen of molnupiravir markedly decreased the likelihood of hospitalization or mortality compared to placebo [61]. Because molnupiravir targets the conserved polymerase, it is still effective against various SARS-CoV-2 mutations [58,62]. Nonetheless, theoretical apprehensions exist over the potential consequences of its mutagenic action, such as its impact on host DNA or the emergence of novel viral mutations [60]. However, current research suggests it is harmless, and the viral ExoN proofreading mechanism only partially mitigates its effects [57,60].
Favipiravir (T-705), a purine base analogue, was another nucleoside analogue evaluated early in the pandemic [63,64]. Favipiravir exhibits extensive efficacy against RNA viruses; however, SARS-CoV-2 has demonstrated notable resistance to it [65]. In COVID-19, clinical trials of favipiravir have yielded inconsistent outcomes, and it is not considered a frontline therapy [66,67]. Despite its limited effectiveness when used alone against SARS-CoV-2, ribavirin, a riboside analogue with known activity against some RNA viruses, has been explored in combination regimens [68,69]. The oral derivative of remdesivir’s parent nucleotide (GS-441524), VV116 (deuviridine), is a new nucleoside analogue that has demonstrated potential. The oral bioavailability of VV116 is significantly higher (between 80-90%) [70,71] and in a clinical trial it demonstrated non-inferiority to Paxlovid in early COVID-19, making it a potentially useful oral polymerase inhibitor [72].
3.2. Protease Inhibitors
Two proteases that are necessary for breaking down the replicase polyprotein are encoded by SARS-CoV-2: nsp5 major protease and nsp3 papain-like protease. By inhibiting these proteases, the viral RTC is unable to mature [73,74]. Nirmatrelvir is a peptidomimetic inhibitor of the primary protease (3CLpro) that was developed and, when combined with the pharmacokinetic enhancer ritonavir (which slows down nirmatrelvir’s metabolism), produced the oral drug Paxlovid [73,75]. Nirmatrelvir indirectly halts RNA replication by inhibiting 3CLpro, as the polymerase and other nsps are unable to be released from the polyprotein [74]. In the EPIC-HR Phase 2/3 trial, Paxlovid demonstrated remarkable efficacy by decreasing the likelihood of hospitalization or death by ~89% in high-risk outpatients when administered within a few days of symptom onset [73]. For high-risk patients, it has been a game-changer for early intervention. Although mutations in the 3CL-pro substrate-binding pocket can result in resistance to nirmatrelvir (a number of these mutations have been identified in vitro [76,77], substantial resistance has not yet been extensively disseminated in circulating strains. With the continued usage of Paxlovid, it is crucial to continuously monitor 3CL-pro variations [78].
Additional protease inhibitors have been investigated. The HIV protease inhibitor lopinavir, when tested in combination with ritonavir during early clinical trials, demonstrated no significant benefit for treating COVID-19 [79]. Ensitrelvir, an investigational 3CLpro inhibitor, has demonstrated efficacy in clinical trials conducted in Japan, indicating that protease inhibition represents a validated target for SARS-CoV-2 beyond nirmatrelvir [80]. The papain-like protease (PLpro) represents a potential antiviral target. Inhibitors of PLpro, including GRL-0617, have been identified in preclinical studies. Despite this, none are currently utilized in clinical settings [81].
3.3. Host-Directed Antivirals
One strategy currently explored for inhibiting SARS-CoV-2 replication is through host-target antiviral mechanisms. This process targets the host cell processes hijacked by the viral during its replication cycle. This entails the repurposing of existing approved drugs that are used beyond their conventional indications. Studies on various compounds that might significantly inhibit SARS-CoV-2 replication and significantly affect therapeutic outcomes via host-directed processes have been reported.
Host-target antiviral strategies for SARS-CoV-2 are increasingly becoming a focal point in the quest to manage COVID-19 [82,83]. These approaches leverage the study of host cell processes that the virus exploits during its replication cycle, alongside repurposing existing medications that have been approved for other indications [82]. An array of studies has demonstrated various compounds that may significantly inhibit SARS-CoV-2 replication and enhance therapeutic outcomes through host-directed mechanisms.
Fluoxetine, a selective serotonin reuptake inhibitor (SSRI), along with GS-441524 has been found to synergistically affect SARS-CoV-2 variants in vitro. Findings show that viral replication was reduced through targeting the cellular pathways used by the virus [84]. Compounds that inhibit virus entry into the host cell through the ACE2 receptor and TMPRSS2 protease have been receiving increasing attention. TMPRSS2 inhibitors and ACE2 receptor blockers have limited viral access to host cells. [85,86]. Investigational compounds that inhibit these receptors may enhance the effectiveness of existing antivirals and mitigate viral entry.
The small-molecule inhibitors designed for modulating intracellular signaling pathways that support viral replication have been explored as a route for possible therapeutic intervention. Inhibition of the pathways involved in cellular stress responses or inflation might indirectly suppress the ability of the virus to replicate efficiently [87].
3.4. Combination Therapies and Drug Synergy
The management of COVID19 has gradually utilized combination techniques, especially in serious cases or to combat resistance. The application of antivirals that target various stages of the replication cycle may yield synergistic effects [88,89]. The observed synergy between remdesivir and nirmatrelvir is an excellent example. In vitro, the combination of these two medications demonstrated synergistic antiviral action, suppressing SARS-CoV-2, including Omicron variant strains, more effectively than either drug alone [90,91,92]. By blocking RNA chain elongation and blocking polyprotein processing, respectively, this combination targets replication through two complementary methods.
Remdesivir has been investigated in conjunction with ribavirin, yielding noteworthy results. Ribavirin, while ineffective as a standalone treatment, functions as a base analogue that promotes mutations, resembling the process of lethal mutagenesis. The combination of remdesivir with the other drug demonstrated a synergistic effect, resulting in the complete eradication of SARS-CoV-2 in the cell culture at relatively low concentrations [93]. The virus’s capacity to reproduce was overwhelmed by the combination, which drastically increased the polymerase’s mistake rate while also impeding nucleotide addition. This implies a potential therapeutic strategy for “lethal mutagenesis” that involves the combination of a mutagenic agent and a chain terminator [93,94]. Triple combinations including ribavirin or favipiravir with other antivirals have also been proposed to further boost the efficacy and prevent escape mutants [93,95,96].
Molnupiravir has also been evaluated in pharmacological combinations. Molnupiravir synergizes with various other agents, such as nirmatrelvir and specific host-targeting medicines, according to a recent study [90,91,97]. For example, the combination of molnupiravir with the TMPRSS2 inhibitor camostat, which obstructs viral entrance, or with the DHODH inhibitor brequinar, which diminishes the nucleotide pools essential for viral RNA synthesis, resulted in enhanced suppression of SARS-CoV-2 in lung cell cultures [97,98]. The combination of molnupiravir and nirmatrelvir demonstrated significant synergy, highlighting the advantage of simultaneously targeting polymerase and protease activities [97,99]. In addition, triple therapy using molnupiravir, camostat, and brequinar suppressed viral replication much more in vitro, suggesting that these three medications could be used as a cocktail to treat resistant infections or immunocompromised people who have persistent virus shedding [97,98,100].
Alongside the new antivirals, numerous authorized medications have been modified for possible efficacy against SARS-CoV-2. Several of these repurposed candidates focused on host factors, such as the antiparasitic ivermectin or different kinase inhibitors, whereas a select handful exhibited direct or indirect antiviral properties. An illustrative instance is the antidepressant fluoxetine and the antifungal itraconazole. These medicines, each with unique primary targets (serotonin reuptake and fungal sterol synthesis, respectively), were shown to augment the antiviral efficacy of remdesivir in cell culture [101,102]. The combination of fluoxetine with remdesivir or itraconazole with remdesivir diminished infectious viral generation by over 90% compared to remdesivir administered alone [103]. The precise antiviral contributions of fluoxetine and itraconazole are still being investigated, but fluoxetine may have lysosomotropic effects that impede viral egress, while itraconazole may disrupt cholesterol trafficking for viral replication. The use of safe, off-patent medications to increase the effectiveness of direct antivirals is nevertheless made possible by these findings [104].
In addition to antivirals that directly inhibit replication, immunomodulators have been utilized as adjunctive therapies in COVID-19 treatment to manage hyperinflammation. Although not classified as antivirals, medications such as the corticosteroid dexamethasone and the JAK inhibitor baricitinib (which received emergency use authorization in conjunction with remdesivir) contribute to improved outcomes by alleviating the harmful inflammatory response [103,105]. Interestingly, baricitinib may also have minor antiviral effects by blocking clathrin-mediated endocytosis, which is necessary for viral entry. This makes it a host-targeted treatment that can be used in conjunction with direct antivirals [103,106,107].
A comprehensive survey of antiviral therapies directly or indirectly targeting SARS-CoV-2 replication is summarized in Table 1. The list includes molecules assigned by their molecular targets, such as the RNA-dependent RNA polymerase (nsp12), main protease (3CLpro), papain-like protease (PLpro), and host cellular factors. Moreover, host-targeting chemicals that change cellular pathways for viral replication or entrance were included. Combination therapies that are proving enhanced and effective viral efficacy offer lower resistance probability. Overall, this emphasizes the need for multi-target strategies in COVID-19 control and underscores the current available treatments.
3.5. Antiviral Resistance Considerations
The rapid development of antiviral medication raises questions about resistance. SARS-CoV-2 is an excellent example of an RNA virus’s high rate of mutation, even though its proofreading capacity slows the rate of change [94,108]. Clinical resistance to the main antivirals is not yet a major concern. Patients treated with remdesivir have been observed to occasionally shed variants containing nsp12 mutations, and in vitro studies indicate that the virus can acquire these alterations [109,110]. Molnupiravir’s mechanism complicates the virus’s ability to develop resistance, as any single mutation it may acquire is equally probably harmful as beneficial. This is because the drug drives the virus into an evolutionary dead end via an error catastrophe [111,112]. Based on substantial experience with HIV and HCV therapy, resistance to protease inhibitors is a serious problem. With the continued usage of Paxlovid, it will be essential to continuously monitor the usage of Paxlovid, and it will be essential to continuously monitor 3CL-pro sequences in clinical isolates [113]. Because the virus needs to acquire numerous independent mutations at the same time to evade multiple medications, combination therapy dramatically reduces the possibility of resistance. This notion serves as the foundation of current approaches to COVID 1-19 treatments and future pandemic preparedness [114,115].
4. Future Trends and Emerging Antiviral Strategies
The effectiveness of vaccines and first-generation antivirals in addressing the COVID-19 pandemic is evident; however, the ongoing emergence of new variants and the risk of novel coronaviruses transferring from animals to humans highlight the need for innovative antiviral strategies [116]. Future research will concentrate on broad-spectrum inhibitors, host-targeted therapies, and advanced drug discovery technologies to surpass the virus [117].
4.1. Novel Targets in the Viral Replication Cycle
Beyond the polymerase and protease, researchers are looking into other viral targets that may be used therapeutically. The proofreading exonuclease (nsp14) represents a significant target; inhibiting nsp14’s ExoN activity may enhance the efficacy of nucleotide analogue drugs by obstructing the virus’s ability to correct drug-induced errors [8].
Attempts are being made to identify nsp14 small-molecule inhibitors that could either cause deadly mutagenesis on their own or successfully “sensitize” the virus to currently available polymerase inhibitors. Similar to this, the distinct NiRAN domain of nsp12, which is necessary for capping viral RNA, is distinctive to nidoviruses and has no analogue in human cells; as such, it is a prospective target for medication [8,118]. A compound that could bind the NiRAN domain and prevent its activity could potentially impair the virus’s capacity to initiate new RNA strands or properly terminate them, resulting in nonfunctional RNA outputs. Furthermore, the multifunctional nsp3, especially the papain-like protease domain and macrodomains that counteract host defenses, offers additional antiviral targets. A potent PLpro inhibitor could disrupt polyprotein processing and reduce immune suppression by the virus, providing a dual advantage [118,119].
4.2. Emerging and Novel Host-Directed Antivirals
To attain broad-spectrum efficiency against the current and future coronaviruses, it is advantageous to target host components essential for viral replication [88]. The stability of the host genome reduces the likelihood of mutations, hence decreasing the establishment of resistance via alterations in the target; however, viral adaptations to lessen reliance on that component may occasionally occur. A previously mentioned host-targeted approach involves the use of TMPRSS2 inhibitors, specifically camostat or nafamostat, to obstruct viral entry [120]. Although these medications do not function as direct replication inhibitors, they obstruct the virus from integrating its DNA into the host cell. The cellular pyrimidine biosynthesis pathway represents another target for intervention; inhibitors such as brequinar and teriflunomide diminish the availability of UTP/CTP for viral RNA synthesis. When used alongside direct polymerase inhibitors, such host-targeted agents can induce a more profound replication crisis in the virus [97]. Interest exists in targeting host chaperones, such as Hsp90 or the TRiC complex mentioned earlier, which facilitate the folding of viral enzymes. Disrupting these chaperones may selectively destabilize viral proteins such as the polymerase or protease [121,122].
Intracellular trafficking modulators, such as vesicle formation inhibitors and autophagy disruptors, may interfere with the development of replication organelles [123]. A concern related to host-directed agents is the potential for toxicity, as they interfere with normal cellular functioning. Numerous host-targeting pharmaceuticals, including JAK kinase inhibitors, IL-1β inhibitors, and other agents aimed at inflammatory pathways, are now under evaluation, have received approval for diverse conditions, and exhibit well-established safety guidelines [124]. Identifying host factors that the virus relies on and can temporarily block in patients without significant harm is crucial. In the context of coronaviruses, the interferon system may be utilized; the administration of interferon, specifically type III interferons for localized activity in the lungs, early in the infection can augment the host’s antiviral response. Clinical trials are examining inhaled interferon-λ as a treatment strategy to augment the host’s antiviral response and therefore inhibit viral replication [125,126].
4.3. AI and Computational Drug Discovery
Using artificial intelligence (AI) and computational techniques during the COVID-19 pandemic has significantly accelerated the drug discovery process. To effectively identify possible inhibitors of SARS-CoV-2 replication proteins, researchers used machine learning algorithms in conjunction with structure-based virtual screening. Various novel compounds were proposed by AI systems for evaluation over the course of several months [127]. Using crowdsourcing to generate designs for 3CLpro inhibitors and machine learning to reduce millions of candidates to a few hundred synthesized compounds, the COVID Moonshot project demonstrated low-nanomolar potency sometimes [128]. Research efforts have focused on the polymerase, with one study assessing more than one hundred flavonoid molecules through in silico techniques. This research identified several compounds, including myricetin derivatives, that bind to a conserved allosteric pocket on nsp12, separate from the active site [129]. Despite the development of active-site resistance to analogues, predicted allosteric inhibitors, which are not impacted by active-site mutations, offer a viable means of blocking the polymerase. Analysis of combination therapy data driven by AI has been used to predict synergistic drug pairs, guiding researchers to focus on the most promising combinations among various options [130]. The integration of big data from viral genomics, chemogenomics, and clinical outcomes via machine learning models offers the potential to customize antiviral strategies for specific variants or patient populations in the future [131]. The emergence of a new coronavirus may lead AI models, which have been trained on SARS-CoV-2 and other viral data, to rapidly identify repurposed drugs or novel compound scaffolds. that may be effective, thereby reducing the essential time in the pandemic response [132,133].
4.4. Gene Silencing and RNA-Targeting Therapies
Directly targeting the viral RNA genome or its transcripts with nucleic acid-based treatments is a distinct approach [134]. Small interfering RNAs (siRNAs) can be designed to target conserved regions of the SARS-CoV-2 genome, thereby activating the RNA-induced silencing complex to degrade viral RNAs. Experiments have demonstrated that siRNAs targeting the RdRp or other regions can markedly decrease viral replication in the cell culture. Delivering siRNA to target cells in patients, such as respiratory epithelial cells, presents a challenge; however, nanoparticle delivery systems are under investigation [135].
CRISPR-based antivirals have also been proposed: the RNA-guided RNase Cas13 can be programmed to cleave SARS-CoV-2 RNA genomes inside cells [136]. Cas13 given by lipid nanoparticles has been shown in vitro to inhibit the replication of SARS-CoV-2 and other RNA viruses by preferentially deleting viral RNA, according to proof-of-concept experiments. However, these technologies are still far from clinical usage, and represent a new paradigm of antivirals that, by merely reprogramming the guide sequences, might be efficiently modified to new viral threats [137,138].
4.5. Pan-Coronavirus and Broad-Spectrum Antivirals
Finally, developing antivirals that act against various coronaviruses and potentially all RNA viruses, in addition to SARS-CoV-2,is an important goal for the future [139,140]. Although ribavirin and more recent substances like NHC are examples of broad-spectrum nucleoside analogues that target the polymerase, their clinical use may be limited by toxicity or ineffectiveness [134,141]. The goal of the current research is to find compounds that precisely target the conserved structures found in viral proteins. The interface between nsp12 and nsp7/nsp8, together with the zinc-binding domains of the nsp13 helicase, is conserved among coronaviruses and may function as targets for engineered inhibitors [142]. Another technique entails targeting the host components that are critical for various viruses. One instance is DHODH because many RNA viruses require increased nucleotide synthesis [143]. Another strategy involves targeting innate immune regulators to broadly augment the antiviral state whole minimizing harmful inflammation [144].
In preparation for future pandemics, researchers are screening libraries of compounds against panels of animal coronaviruses and SARS-CoV-2 [145]. These agents may be stockpiled and deployed quickly in response to a new coronavirus transmission to humans. The COVID-19 pandemic has underscored the necessity of having accessible antivirals to optimize the response time during health emergencies. The development of broad-spectrum antivirals may reduce their specificity; nevertheless, they provide protection against unexpected threats [146].
Emerging antiviral strategies against SARS-CoV-2 are summarized in Table 2. These emerging strategies include, but are not limited to, using artificial intelligence to speed compound discovery and optimize combination therapies, investigating host-directed antivirals that block vital cellular processes, and targeting new viral replication components including the NiRAN domain and PLpro. Programable platforms for direct RNA degradation are provided by gene-silencing tools, including siRNAs and CRISPR-based Cas13 systems. Ultimately, broad-spectrum antivirals and pan-coronavirus seek to ensure pandemic preparedness with compounds targeting conserved viral or host elements, thus tackling cross-species transmission risks.
5. Conclusion
The COVID-19 pandemic has advanced our understanding of coronavirus replication and renewed interest in antiviral drugs. SARS-CoV-2 replication involves complicated viral protein activities and dynamic host-virus interactions. Over the past few years, researchers have discovered the polymerase complex’s atomic structure, RNA manufacturing mechanism ,and how the virus alters cellular membranes and avoids immune detection. This comprehension has directly impacted the development of targeted therapeutics. RNA polymerase inhibitors like remdesivir and molnupiravir and protease inhibitors like nirmatrelvir were developed to limit viral polyprotein processing. These interventions have shown their effectiveness in reducing disease severity and saving lives.
Current therapies, such as early administration and intravenous remdesivir delivery, are limited by the virus’s development and require caution. While not yet a clinical issue for SARS-CoV-2, antiviral resistance remains a concern. The advent of variants with changed replication or immune evasion features highlights the need to expand antiviral therapies. Researchers are exploring next-generation antivirals. These include cutting-edge tactics like CRISPR-based antivirals, host-targeted treatments that may provide broad protection against several coronaviruses, and inhibitors of several viral enzymes (helicases, exonucleases, etc.) AI and computer approaches speed up drug discovery
To summarize, the development of successful therapeutic strategies for COVID-19 depends on an understanding of the molecular mechanisms underlying SARS-CoV-2 replication. To contain the pandemic and prepare for future outbreaks, researchers are discovering novel viral inhibitory targets and techniques. Addressing SARS-CoV-2 and other deadly viruses requires a synergistic approach that combines molecular virology, pharmacology, and new technology. The insights and antivirals developed during the COVID-19 pandemic will shape future viral disease management.
Author Contributions
Conceptualization: B.J.J.S. and I.L.F.; writing: original draft preparation: B.J.J.S. and I.L.F.; writing review and editing: B.J.J.S. and I.L.F. All authors have read and agreed to the version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
B.J.J.S. would like to acknowledge the support of the Ministry of Education, Culture, Sports, Science and Technology (MEXT).
Conflicts of Interest
All authors declare no conflicts of interest.
References
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.-L. Characteristics of SARS-CoV-2 and COVID-19. Nat Rev Microbiol 2021, 19, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, A.; Preiss, A.J.; Xiao, X.; Brannock, M.D.; Alexander, G.C.; Chew, R.F.; Fitzgerald, M.; Hill, E.; Kelly, E.P.; Mehta, H.B.; et al. Effect of Nirmatrelvir/Ritonavir (Paxlovid) on Hospitalization among Adults with COVID-19: An EHR-Based Target Trial Emulation from N3C. medRxiv 2023, 2023.05.03.23289084. [CrossRef]
- Faisal, H.M.N.; Katti, K.S.; Katti, D.R. Differences in Interactions Within Viral Replication Complexes of SARS-CoV-2 (COVID-19) and SARS-CoV Coronaviruses Control RNA Replication Ability. JOM (1989) 2021, 73, 1684–1695. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Mao, C.; Luan, X.; Shen, D.-D.; Shen, Q.; Su, H.; Wang, X.; Zhou, F.; Zhao, W.; Gao, M.; et al. Structural Basis for Inhibition of the RNA-Dependent RNA Polymerase from SARS-CoV-2 by Remdesivir. Science 2020, 368, 1499–1504. [Google Scholar] [CrossRef]
- Saberi, A.; Gulyaeva, A.A.; Brubacher, J.L.; Newmark, P.A.; Gorbalenya, A.E. A Planarian Nidovirus Expands the Limits of RNA Genome Size. PLoS Pathog 2018, 14, e1007314. [Google Scholar] [CrossRef]
- Krell, P.J. An Introduction to Viruses of Invertebrates. In Encyclopedia of Virology; Elsevier, 2021; pp. 699–723 ISBN 978-0-12-814516-6.
- Bettini, A.; Lapa, D.; Garbuglia, A.R. Diagnostics of Ebola Virus. Front. Public Health 2023, 11, 1123024. [Google Scholar] [CrossRef]
- Hillen, H.S. Structure and Function of SARS-CoV-2 Polymerase. Current Opinion in Virology 2021, 48, 82–90. [Google Scholar] [CrossRef]
- Terrier, O.; Si-Tahar, M.; Ducatez, M.; Chevalier, C.; Pizzorno, A.; Le Goffic, R.; Crépin, T.; Simon, G.; Naffakh, N. Influenza Viruses and Coronaviruses: Knowns, Unknowns, and Common Research Challenges. PLoS Pathog 2021, 17, e1010106. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, A.; Singh, R.; Misra, A. Molnupiravir in COVID-19: A Systematic Review of Literature. Diabetes and Metabolic Syndrome: Clinical Research & Reviews 2021, 15, 102329. [Google Scholar] [CrossRef]
- Qiu, Y.; Wen, H.; Wang, H.; Sun, W.; Li, G.; Li, S.; Wang, Y.; Zhai, J.; Zhan, Y.; Su, Y.; et al. Real-World Effectiveness and Safety of Nirmatrelvir-Ritonavir (Paxlovid)-Treated for COVID-19 Patients with Onset of More than 5 Days: A Retrospective Cohort Study. Front. Pharmacol. 2024, 15, 1401658. [Google Scholar] [CrossRef]
- Naqvi, A.A.T.; Fatima, K.; Mohammad, T.; Fatima, U.; Singh, I.K.; Singh, A.; Atif, S.M.; Hariprasad, G.; Hasan, G.M.; Hassan, Md. I. Insights into the SARS-CoV-2 Genome, Structure, Evolution, Pathogenesis and Therapies: Structural Genomics Approach. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease 2020, 1866, 165878. [Google Scholar] [CrossRef]
- Brant, A.C.; Tian, W.; Majerciak, V.; Yang, W.; Zheng, Z.-M. SARS-CoV-2: From Its Discovery to Genome Structure, Transcription, and Replication. Cell Biosci 2021, 11, 136. [Google Scholar] [CrossRef] [PubMed]
- Hillen, H.S. Structure and Function of SARS-CoV-2 Polymerase. Current Opinion in Virology 2021, 48, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Polatoğlu, I.; Oncu-Oner, T.; Dalman, I.; Ozdogan, S. COVID-19 in Early 2023: Structure, Replication Mechanism, Variants of SARS-CoV-2, Diagnostic Tests, and Vaccine & Drug Development Studies. MedComm 2023, 4, e228. [Google Scholar] [CrossRef] [PubMed]
- Baggen, J.; Vanstreels, E.; Jansen, S.; Daelemans, D. Cellular Host Factors for SARS-CoV-2 Infection. Nat Microbiol 2021, 6, 1219–1232. [Google Scholar] [CrossRef]
- Yan, W.; Zheng, Y.; Zeng, X.; He, B.; Cheng, W. Structural Biology of SARS-CoV-2: Opening the Door for Novel Therapies. Sig Transduct Target Ther 2022, 7, 26. [Google Scholar] [CrossRef]
- Guo, Y.-R.; Cao, Q.-D.; Hong, Z.-S.; Tan, Y.-Y.; Chen, S.-D.; Jin, H.-J.; Tan, K.-S.; Wang, D.-Y.; Yan, Y. The Origin, Transmission and Clinical Therapies on Coronavirus Disease 2019 (COVID-19) Outbreak-an Update on the Status. Mil Med Res 2020, 7, 11. [Google Scholar] [CrossRef]
- Fehr, A.R.; Perlman, S. Coronaviruses: An Overview of Their Replication and Pathogenesis. Methods Mol Biol 2015, 1282, 1–23. [Google Scholar] [CrossRef]
- Subong, B.J.J.; Ozawa, T. Bio-Chemoinformatics-Driven Analysis of Nsp7 and Nsp8 Mutations and Their Effects on Viral Replication Protein Complex Stability. CIMB 2024, 46, 2598–2619. [Google Scholar] [CrossRef]
- Reshamwala, S.M.S.; Likhite, V.; Degani, M.S.; Deb, S.S.; Noronha, S.B. Mutations in SARS-CoV-2 Nsp7 and Nsp8 Proteins and Their Predicted Impact on the Replication/Transcription Complex Structure. Journal of Medical Virology 2021, 93, 4616–4619. [Google Scholar] [CrossRef]
- Gao, Y.; Yan, L.; Huang, Y.; Liu, F.; Zhao, Y.; Cao, L.; Wang, T.; Sun, Q.; Ming, Z.; Zhang, L.; et al. Structure of the RNA-Dependent RNA Polymerase from COVID-19 Virus. Science 2020, 368, 779–782. [Google Scholar] [CrossRef]
- Slanina, H.; Madhugiri, R.; Bylapudi, G.; Schultheiß, K.; Karl, N.; Gulyaeva, A.; Gorbalenya, A.E.; Linne, U.; Ziebuhr, J. Coronavirus Replication–Transcription Complex: Vital and Selective NMPylation of a Conserved Site in Nsp9 by the NiRAN-RdRp Subunit. Proc. Natl. Acad. Sci. U.S.A. 2021, 118, e2022310118. [Google Scholar] [CrossRef] [PubMed]
- Mickolajczyk, K.J.; Shelton, P.M.M.; Grasso, M.; Cao, X.; Warrington, S.E.; Aher, A.; Liu, S.; Kapoor, T.M. Force-Dependent Stimulation of RNA Unwinding by SARS-CoV-2 Nsp13 Helicase. Biophysical Journal 2021, 120, 1020–1030. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, Q.; Malone, B.; Llewellyn, E.; Pechersky, Y.; Maruthi, K.; Eng, E.T.; Perry, J.K.; Campbell, E.A.; Shaw, D.E.; et al. Ensemble Cryo-EM Reveals the Conformational States of the Nsp13 Helicase in the SARS-CoV-2 Helicase Replication–Transcription Complex. Nat Struct Mol Biol 2022, 29, 250–260. [Google Scholar] [CrossRef]
- Chinthapatla, R.; Sotoudegan, M.; Srivastava, P.; Anderson, T.K.; Moustafa, I.M.; Passow, K.T.; Kennelly, S.A.; Moorthy, R.; Dulin, D.; Feng, J.Y.; et al. Interfering with Nucleotide Excision by the Coronavirus 3′-to-5′ Exoribonuclease. Nucleic Acids Research 2023, 51, 315–336. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Kong, F.; Hou, Y.J.; Wang, Q. Crucial Mutation in the Exoribonuclease Domain of Nsp14 of PEDV Leads to High Genetic Instability during Viral Replication. Cell Biosci 2021, 11, 106. [Google Scholar] [CrossRef]
- Bouvet, M.; Lugari, A.; Posthuma, C.C.; Zevenhoven, J.C.; Bernard, S.; Betzi, S.; Imbert, I.; Canard, B.; Guillemot, J.-C.; Lécine, P.; et al. Coronavirus Nsp10, a Critical Co-Factor for the Activation of Multiple Replicative Enzymes. J Biol Chem 2014, 289, 25783–25796. [Google Scholar] [CrossRef]
- Yang, H.; Rao, Z. Structural Biology of SARS-CoV-2 and its Implications for Therapeutic Development. Nat Rev Microbiol 2021, 19, 685–700. [Google Scholar] [CrossRef]
- Klein, S.; Cortese, M.; Winter, S.L.; Wachsmuth-Melm, M.; Neufeldt, C.J.; Cerikan, B.; Stanifer, M.L.; Boulant, S.; Bartenschlager, R.; Chlanda, P. SARS-CoV-2 Structure and Replication Characterized by in Situ Cryo-Electron Tomography. Nat Commun 2020, 11, 5885. [Google Scholar] [CrossRef]
- Malone, B.; Urakova, N.; Snijder, E.J.; Campbell, E.A. Structures and Functions of Coronavirus Replication–Transcription Complexes and Their Relevance for SARS-CoV-2 Drug Design. Nat Rev Mol Cell Biol 2022, 23, 21–39. [Google Scholar] [CrossRef]
- Wong, L.H.; Edgar, J.R.; Martello, A.; Ferguson, B.J.; Eden, E.R. Exploiting Connections for Viral Replication. Front. Cell Dev. Biol. 2021, 9, 640456. [Google Scholar] [CrossRef]
- Roingeard, P.; Eymieux, S.; Burlaud-Gaillard, J.; Hourioux, C.; Patient, R.; Blanchard, E. The Double-Membrane Vesicle (DMV): A Virus-Induced Organelle Dedicated to the Replication of SARS-CoV-2 and Other Positive-Sense Single-Stranded RNA Viruses. Cell. Mol. Life Sci. 2022, 79, 425. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, T.; Zhong, L.; Zhang, W.; Zhang, Y.; Yu, X.; Yuan, S.; Ni, T. Molecular Architecture of the Coronavirus Double-Membrane Vesicle Pore Complex. Nature 2024, 633, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.M.; Chen, Y.-J.; Cho, W.J.; Tai, A.W.; Tsai, B. Reticulons Promote the Formation of ER-Derived Double-Membrane Vesicles That Facilitate SARS-CoV-2 Replication. Journal of Cell Biology 2023, 222, e202203060. [Google Scholar] [CrossRef]
- Hagemeijer, M.C.; Monastyrska, I.; Griffith, J.; Van Der Sluijs, P.; Voortman, J.; Van Bergen En Henegouwen, P.M.; Vonk, A.M.; Rottier, P.J.M.; Reggiori, F.; De Haan, C.A.M. Membrane Rearrangements Mediated by Coronavirus Nonstructural Proteins 3 and 4. Virology 2014, 458–459, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Alexandersen, S.; Chamings, A.; Bhatta, T.R. SARS-CoV-2 Genomic and Subgenomic RNAs in Diagnostic Samples Are Not an Indicator of Active Replication. Nat Commun 2020, 11, 6059. [Google Scholar] [CrossRef]
- Telwatte, S.; Martin, H.A.; Marczak, R.; Fozouni, P.; Vallejo-Gracia, A.; Kumar, G.R.; Murray, V.; Lee, S.; Ott, M.; Wong, J.K.; et al. Novel RT-ddPCR Assays for Measuring the Levels of Subgenomic and Genomic SARS-CoV-2 Transcripts. Methods 2022, 201, 15–25. [Google Scholar] [CrossRef]
- Markov, P.V.; Ghafari, M.; Beer, M.; Lythgoe, K.; Simmonds, P.; Stilianakis, N.I.; Katzourakis, A. The Evolution of SARS-CoV-2. Nat Rev Microbiol 2023, 21, 361–379. [Google Scholar] [CrossRef]
- Mack, A.H.; Menzies, G.; Southgate, A.; Jones, D.D.; Connor, T.R. A Proofreading Mutation with an Allosteric Effect Allows a Cluster of SARS-CoV-2 Viruses to Rapidly Evolve. Molecular Biology and Evolution 2023, 40, msad209. [Google Scholar] [CrossRef]
- Prydz, K.; Saraste, J. The Life Cycle and Enigmatic Egress of Coronaviruses. Molecular Microbiology 2022, 117, 1308–1316. [Google Scholar] [CrossRef]
- Perdikari, T.M.; Murthy, A.C.; Ryan, V.H.; Watters, S.; Naik, M.T.; Fawzi, N.L. SARS-CoV-2 Nucleocapsid Protein Phase-separates with RNA and with Human hnRNPs. The EMBO Journal 2020, 39, e106478. [Google Scholar] [CrossRef]
- Pearson, G.J.; Mears, H.V.; Broncel, M.; Snijders, A.P.; Bauer, D.L.V.; Carlton, J.G. ER-Export and ARFRP1/AP-1–Dependent Delivery of SARS-CoV-2 Envelope to Lysosomes Controls Late Stages of Viral Replication. Sci. Adv. 2024, 10, eadl5012. [Google Scholar] [CrossRef]
- Gordon, D.E.; Hiatt, J.; Bouhaddou, M.; Rezelj, V.V.; Ulferts, S.; Braberg, H.; Jureka, A.S.; Obernier, K.; Guo, J.Z.; Batra, J.; et al. Comparative Host-Coronavirus Protein Interaction Networks Reveal Pan-Viral Disease Mechanisms. Science 2020, 370, eabe9403. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.; Wang, S.; Kwon, Y.; Reid, A.A.; Robison, R.; Shen, P.; Willardson, B. Folding of the SARS-CoV-2 RNA Polymerase by the Cytosolic Chaperonin CCT. The FASEB Journal 2022, 36, fasebj.2022–36.S1. [Google Scholar] [CrossRef]
- Alruwaili, M.; Armstrong, S.; Prince, T.; Erdmann, M.; Matthews, D.A.; Davidson, A.; Aljabr, W.; Hiscox, J.A. SARS-CoV-2 NSP12 Associates with the TRiC Complex and the P323L Substitution Is a Host Adaption 2023.
- Liu, Q.; Zhao, S.; Hou, Y.; Ye, S.; Sha, T.; Su, Y.; Zhao, W.; Bao, Y.; Xue, Y.; Chen, H. Ongoing Natural Selection Drives the Evolution of the SARS-CoV-2 Genomes 2020.
- Shi, J.; Du, T.; Wang, J.; Tang, C.; Lei, M.; Yu, W.; Yang, Y.; Ma, Y.; Huang, P.; Chen, H.; et al. Aryl Hydrocarbon Receptor Is a Proviral Host Factor and a Candidate Pan-SARS-CoV-2 Therapeutic Target. Sci. Adv. 2023, 9, eadf0211. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Pu, Y.; Yuan, S.; Pache, L.; Churas, C.; Weston, S.; Riva, L.; Simons, L.M.; Cisneros, W.; Clausen, T.; et al. Global siRNA Screen Reveals Critical Human Host Factors of SARS-CoV-2 Multicycle Replication 2024.
- Jockusch, S.; Tao, C.; Li, X.; Anderson, T.K.; Chien, M.; Kumar, S.; Russo, J.J.; Kirchdoerfer, R.N.; Ju, J. A Library of Nucleotide Analogues Terminate RNA Synthesis Catalyzed by the Polymerases of Coronaviruses That Cause SARS and COVID-19. Antiviral Research 2020, 180, 104857. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; De Clercq, E. Therapeutic Options for the 2019 Novel Coronavirus (2019-nCoV). Nat Rev Drug Discov 2020, 19, 149–150. [Google Scholar] [CrossRef]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for Treating COVID-19 — Final Report. N Engl J Med 2020, 383, 1813–1826. [Google Scholar] [CrossRef]
- Olender, S.A.; Perez, K.K.; Go, A.S.; Balani, B.; Price-Haywood, E.G.; Shah, N.S.; Wang, S.; Walunas, T.L.; Swaminathan, S.; Slim, J.; et al. Remdesivir for Severe Coronavirus Disease 2019 (COVID-19) Versus a Cohort Receiving Standard of Care. Clinical Infectious Diseases 2021, 73, e4166–e4174. [Google Scholar] [CrossRef]
- Ogando, N.S.; Zevenhoven-Dobbe, J.C.; Van Der Meer, Y.; Bredenbeek, P.J.; Posthuma, C.C.; Snijder, E.J. The Enzymatic Activity of the Nsp14 Exoribonuclease Is Critical for the Replication of MERS-CoV and SARS-CoV-2. J Virol 2020, 94, e01246–20. [Google Scholar] [CrossRef]
- Peng, J.-Y.; Lahser, F.; Warren, C.; He, X.; Murray, E.; Wang, D. The Impact of SARS-CoV-2 Nsp14 Proofreading on Nucleoside Antiviral Activity: Insights from Genetic and Pharmacological Investigations 2024.
- Lee, C.-C.; Hsieh, C.-C.; Ko, W.-C. Molnupiravir—A Novel Oral Anti-SARS-CoV-2 Agent. Antibiotics 2021, 10, 1294. [Google Scholar] [CrossRef]
- Kabinger, F.; Stiller, C.; Schmitzová, J.; Dienemann, C.; Kokic, G.; Hillen, H.S.; Höbartner, C.; Cramer, P. Mechanism of Molnupiravir-Induced SARS-CoV-2 Mutagenesis. Nat Struct Mol Biol 2021, 28, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Pang, Z.; Li, M.; Lou, F.; An, X.; Zhu, S.; Song, L.; Tong, Y.; Fan, H.; Fan, J. Molnupiravir and Its Antiviral Activity Against COVID-19. Front. Immunol. 2022, 13, 855496. [Google Scholar] [CrossRef]
- Malone, B.; Campbell, E.A. Molnupiravir: Coding for Catastrophe. Nat Struct Mol Biol 2021, 28, 706–708. [Google Scholar] [CrossRef] [PubMed]
- Masyeni, S.; Iqhrammullah, M.; Frediansyah, A.; Nainu, F.; Tallei, T.; Emran, T.B.; Ophinni, Y.; Dhama, K.; Harapan, H. Molnupiravir: A Lethal Mutagenic Drug against Rapidly Mutating Severe Acute Respiratory Syndrome Coronavirus 2—A Narrative Review. 4.4 Journal of Medical Virology 2022, 94, 3006–3016. [Google Scholar] [CrossRef]
- Caraco, Y.; Crofoot, G.E.; Moncada, P.A.; Galustyan, A.N.; Musungaie, D.B.; Payne, B.; Kovalchuk, E.; Gonzalez, A.; Brown, M.L.; Williams-Diaz, A.; et al. Phase 2/3 Trial of Molnupiravir for Treatment of COVID-19 in Nonhospitalized Adults. NEJM Evidence 2022, 1. [Google Scholar] [CrossRef]
- Strizki, J.M.; Gaspar, J.M.; Howe, J.A.; Hutchins, B.; Mohri, H.; Nair, M.S.; Kinek, K.C.; McKenna, P.; Goh, S.L.; Murgolo, N. Molnupiravir Maintains Antiviral Activity against SARS-CoV-2 Variants and Exhibits a High Barrier to the Development of Resistance. Antimicrob Agents Chemother 2024, 68, e00953–23. [Google Scholar] [CrossRef]
- Erdem, H.A.; Korkmaz Ekren, P.; Çağlayan, D.; Işikgöz Taşbakan, M.; Yamazhan, T.; Taşbakan, M.S.; Sayiner, A.; Gökengi̇N, D. Treatment of SARS-CoV-2 Pneumonia with Favipiravir: Early Results from the Ege University Cohort, Turkey. Turk J Med Sci 2021, 51, 912–920. [Google Scholar] [CrossRef]
- Ghasemnejad-Berenji, M.; Pashapour, S. Favipiravir and COVID-19: A Simplified Summary. Drug Res (Stuttg) 2021, 71, 166–170. [Google Scholar] [CrossRef]
- Manabe, T.; Kambayashi, D.; Akatsu, H.; Kudo, K. Favipiravir for the Treatment of Patients with COVID-19: A Systematic Review and Meta-Analysis. BMC Infect Dis 2021, 21, 489. [Google Scholar] [CrossRef]
- Juul, S.; Nielsen, E.E.; Feinberg, J.; Siddiqui, F.; Jørgensen, C.K.; Barot, E.; Holgersson, J.; Nielsen, N.; Bentzer, P.; Veroniki, A.A.; et al. Interventions for the Treatment of COVID-19: Second Edition of a Living Systematic Review with Meta-Analyses and Trial Sequential Analyses (The LIVING Project). PLoS ONE 2021, 16, e0248132. [Google Scholar] [CrossRef]
- Shrestha, D.B.; Budhathoki, P.; Khadka, S.; Shah, P.B.; Pokharel, N.; Rashmi, P. Favipiravir versus Other Antiviral or Standard of Care for COVID-19 Treatment: A Rapid Systematic Review and Meta-Analysis. Virol J 2020, 17, 141. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Su, Y.; Yu, Y.; Wu, C.; Chen, J.; Wang, S.; Jiang, J. Ribavirin Therapy for Severe COVID-19: A Retrospective Cohort Study. International Journal of Antimicrobial Agents 2020, 56, 106114. [Google Scholar] [CrossRef] [PubMed]
- Messina, E.; Danise, A.; Ferrari, G.; Andolina, A.; Chiurlo, M.; Razanakolona, M.; Barakat, M.; Israel, R.J.; Castagna, A. Ribavirin Aerosol for treating SARS-CoV-2: A Case Series. Infect Dis Ther 2021, 10, 2791–2804. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Wang, Y.; Zhang, M.; Xie, Y.; Wu, Q.; Liang, L.; Cao, Y.; Duan, H.; Tian, G.; Ma, J.; et al. Safety, Tolerability, and Pharmacokinetics of VV116, an Oral Nucleoside Analog against SARS-CoV-2, in Chinese Healthy Subjects. Acta Pharmacol Sin 2022, 43, 3130–3138. [Google Scholar] [CrossRef]
- Xiao, N.; Huang, X.; Kang, X.; Zang, W.; Li, B.; Kiselev, S. The Safety and Efficacy of the Oral Antiviral Drug VV116 for the Treatment of COVID-19: A Systematic Review. Medicine 2023, 102, e34105. [Google Scholar] [CrossRef]
- McCarthy, M.W. VV116 as a Potential Treatment for COVID-19. Expert Opinion on Pharmacotherapy 2023, 24, 675–678. [Google Scholar] [CrossRef]
- Hammond, J.; Leister-Tebbe, H.; Gardner, A.; Abreu, P.; Bao, W.; Wisemandle, W.; Baniecki, M.; Hendrick, V.M.; Damle, B.; Simón-Campos, A.; et al. Oral Nirmatrelvir for High-Risk, Nonhospitalized Adults with COVID-19. N Engl J Med 2022, 386, 1397–1408. [Google Scholar] [CrossRef]
- Marzi, M.; Vakil, M.K.; Bahmanyar, M.; Zarenezhad, E. Paxlovid: Mechanism of Action, Synthesis, and In Silico Study. BioMed Research International 2022, 2022, 7341493. [Google Scholar] [CrossRef]
- Marzolini, C.; Kuritzkes, D.R.; Marra, F.; Boyle, A.; Gibbons, S.; Flexner, C.; Pozniak, A.; Boffito, M.; Waters, L.; Burger, D.; et al. Recommendations for the Management of Drug–Drug Interactions Between the COVID -19 Antiviral Nirmatrelvir/Ritonavir (Paxlovid) and Comedications. Clin Pharma and Therapeutics 2022, 112, 1191–1200. [Google Scholar] [CrossRef]
- Zhou, Y.; Gammeltoft, K.A.; Ryberg, L.A.; Pham, L.V.; Fahnøe, U.; Binderup, A.; Hernandez, C.R.D.; Offersgaard, A.; Fernandez-Antunez, C.; Peters, G.H.J.; et al. Nirmatrelvir-resistant SARS-CoV-2 Variants with High Fitness in Vitro 2022.
- Iketani, S.; Mohri, H.; Culbertson, B.; Hong, S.J.; Duan, Y.; Luck, M.I.; Annavajhala, M.K.; Guo, Y.; Sheng, Z.; Uhlemann, A.-C.; et al. Multiple Pathways for SARS-CoV-2 Resistance to Nirmatrelvir 2022.
- Costacurta, F.; Dodaro, A.; Bante, D.; Schöppe, H.; Peng, J.-Y.; Sprenger, B.; He, X.; Moghadasi, S.A.; Egger, L.M.; Fleischmann, J.; et al. A Comprehensive Study of SARS-CoV-2 Main Protease (Mpro) Inhibitor-Resistant Mutants Selected in a VSV-Based System. PLoS Pathog 2024, 20, e1012522. [Google Scholar] [CrossRef]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. A Trial of Lopinavir–Ritonavir in Adults Hospitalized with Severe COVID-19. N Engl J Med 2020, 382, 1787–1799. [Google Scholar] [CrossRef]
- Kuroda, T.; Nobori, H.; Fukao, K.; Baba, K.; Matsumoto, K.; Yoshida, S.; Tanaka, Y.; Watari, R.; Oka, R.; Kasai, Y.; et al. Efficacy Comparison of the 3CL Protease Inhibitors Ensitrelvir and Nirmatrelvir against SARS-CoV-2 in Vitro and in Vivo. Journal of Antimicrobial Chemotherapy 2023, 78, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Garnsey, M.R.; Robinson, M.C.; Nguyen, L.T.; Cardin, R.; Tillotson, J.; Mashalidis, E.; Yu, A.; Aschenbrenner, L.; Balesano, A.; Behzadi, A.; et al. Discovery of SARS-CoV-2 Papain-like Protease (PLpro ) Inhibitors with Efficacy in a Murine Infection Model. Sci. Adv. 2024, 10, eado4288. [Google Scholar] [CrossRef] [PubMed]
- Săndulescu, O.; Apostolescu, C.G.; Preoțescu, L.L.; Streinu-Cercel, A.; Săndulescu, M. Therapeutic Developments for SARS-CoV-2 Infection—Molecular Mechanisms of Action of Antivirals and Strategies for Mitigating Resistance in Emerging Variants in Clinical Practice. Front. Microbiol. 2023, 14, 1132501. [Google Scholar] [CrossRef]
- Saul, S.; Einav, S. Old Drugs for a New Virus: Repurposed Approaches for Combating COVID-19. ACS Infect. Dis. 2020, 6, 2304–2318. [Google Scholar] [CrossRef] [PubMed]
- Brunotte, L.; Zheng, S.; Mecate-Zambrano, A.; Tang, J.; Ludwig, S.; Rescher, U.; Schloer, S. Combination Therapy with Fluoxetine and the Nucleoside Analog GS-441524 Exerts Synergistic Antiviral Effects against Different SARS-CoV-2 Variants In Vitro. Pharmaceutics 2021, 13, 1400. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Xia, S.; Liu, M.; Wang, C.; Xu, W.; Lan, Q.; Feng, S.; Qi, F.; Bao, L.; Du, L.; Liu, S.; et al. Inhibition of SARS-CoV-2 (Previously 2019-nCoV) Infection by a Highly Potent Pan-Coronavirus Fusion Inhibitor Targeting Its Spike Protein That Harbors a High Capacity to Mediate Membrane Fusion. Cell Res 2020, 30, 343–355. [Google Scholar] [CrossRef]
- Lei, S.; Chen, X.; Wu, J.; Duan, X.; Men, K. Small Molecules for treating COVID-19. Sig Transduct Target Ther 2022, 7, 387. [Google Scholar] [CrossRef]
- Batool, S.; Chokkakula, S.; Jeong, J.H.; Baek, Y.H.; Song, M.-S. SARS-CoV-2 Drug Resistance and Therapeutic Approaches. Heliyon 2025, 11, e41980. [Google Scholar] [CrossRef]
- Rahmah, L.; Abarikwu, S.O.; Arero, A.G.; Essouma, M.; Jibril, A.T.; Fal, A.; Flisiak, R.; Makuku, R.; Marquez, L.; Mohamed, K.; et al. Oral Antiviral Treatments for COVID-19: Opportunities and Challenges. Pharmacol. Rep 2022, 74, 1255–1278. [Google Scholar] [CrossRef] [PubMed]
- Gidari, A.; Sabbatini, S.; Schiaroli, E.; Bastianelli, S.; Pierucci, S.; Busti, C.; Saraca, L.M.; Capogrossi, L.; Pasticci, M.B.; Francisci, D. Synergistic Activity of the Remdesivir–Nirmatrelvir Combination in a SARS-CoV-2 In Vitro Model and a Case Report. Viruses 2023, 15, 1577. [Google Scholar] [CrossRef] [PubMed]
- Gidari, A.; Sabbatini, S.; Schiaroli, E.; Bastianelli, S.; Pierucci, S.; Busti, C.; Comez, L.; Libera, V.; Macchiarulo, A.; Paciaroni, A.; et al. The Combination of Molnupiravir with Nirmatrelvir or GC376 Has a Synergic Role in the Inhibition of SARS-CoV-2 Replication In Vitro. Microorganisms 2022, 10, 1475. [Google Scholar] [CrossRef] [PubMed]
- Woodall, M.; Ellis, S.; Zhang, S.; Kembou-Ringert, J.; Kite, K.-A.; Buggiotti, L.; Jacobs, A.I.; Agyeman, A.A.; Masonou, T.; Palor, M.; et al. Efficient in Vitro Assay for Evaluating Drug Efficacy and Synergy against Emerging SARS-CoV-2 Strains. Antimicrob Agents Chemother 2025, 69, e01233–24. [Google Scholar] [CrossRef]
- García-Crespo, C.; De Ávila, A.I.; Gallego, I.; Soria, M.E.; Durán-Pastor, A.; Somovilla, P.; Martínez-González, B.; Muñoz-Flores, J.; Mínguez, P.; Salar-Vidal, L.; et al. Synergism between Remdesivir and Ribavirin Leads to SARS-CoV-2 Extinction in Cell Culture. British J Pharmacology 2024, 181, 2636–2654. [Google Scholar] [CrossRef]
- Robson, F.; Khan, K.S.; Le, T.K.; Paris, C.; Demirbag, S.; Barfuss, P.; Rocchi, P.; Ng, W.-L. Coronavirus RNA Proofreading: Molecular Basis and Therapeutic Targeting. Molecular Cell 2020, 79, 710–727. [Google Scholar] [CrossRef]
- Do, T.N.D.; Abdelnabi, R.; Boda, B.; Constant, S.; Neyts, J.; Jochmans, D. The Triple Combination of Remdesivir (GS-441524), Molnupiravir, and Ribavirin Is Highly Efficient in Inhibiting Coronavirus Replication in Human Nasal Airway Epithelial Cell Cultures and in a Hamster Infection Model. Antiviral Research 2024, 231, 105994. [Google Scholar] [CrossRef]
- Mayor, J.; Engler, O.; Rothenberger, S. Antiviral Efficacy of Ribavirin and Favipiravir against Hantaan Virus. Microorganisms 2021, 9, 1306. [Google Scholar] [CrossRef]
- Wagoner, J.; Herring, S.; Hsiang, T.-Y.; Ianevski, A.; Biering, S.B.; Xu, S.; Hoffmann, M.; Pöhlmann, S.; Gale, M.; Aittokallio, T.; et al. Combinations of Host- and Virus-Targeting Antiviral Drugs Confer Synergistic Suppression of SARS-CoV-2. Microbiol Spectr 2022, 10, e03331–22. [Google Scholar] [CrossRef]
- White, J.M.; Schiffer, J.T.; Bender Ignacio, R.A.; Xu, S.; Kainov, D.; Ianevski, A.; Aittokallio, T.; Frieman, M.; Olinger, G.G.; Polyak, S.J. Drug Combinations as a First Line of Defense against Coronaviruses and Other Emerging Viruses. mBio 2021, 12, e03347–21. [Google Scholar] [CrossRef]
- Jeong, J.H.; Chokkakula, S.; Min, S. C.; Kim, B.K.; Choi, W.-S.; Oh, S.; Yun, Y.S.; Kang, D.H.; Lee, O. J.; Kim, E.-G.; et al. Combination Therapy with Nirmatrelvir and Molnupiravir Improves the Survival of SARS-CoV-2 Infected Mice. Antiviral Research 2022, 208, 105430. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, T.R.; Cynthia, B.; Thevendran, R.; Maheswaran, S. Prospects of Innovative Therapeutics in Combating the COVID-19 Pandemic. Mol Biotechnol 2024. [Google Scholar] [CrossRef]
- Abdulaziz, L.; Elhadi, E.; Abdallah, E.A.; Alnoor, F.A.; Yousef, B.A. Antiviral Activity of Approved Antibacterial, Antifungal, Antiprotozoal and Anthelmintic Drugs: Chances for Drug Repurposing for Antiviral Drug Discovery. JEP 2022, Volume 14, 97–115. [Google Scholar] [CrossRef]
- Hamid, A.; Mäser, P.; Mahmoud, A.B. Drug Repurposing in the Chemotherapy of Infectious Diseases. Molecules 2024, 29, 635. [Google Scholar] [CrossRef]
- Chatterjee, B.; Thakur, S.S. Remdesivir and Its Combination With Repurposed Drugs as COVID-19 Therapeutics. Front. Immunol. 2022, 13, 830990. [Google Scholar] [CrossRef]
- Mahdi, M.; Hermán, L.; Réthelyi, J.M.; Bálint, B.L. Potential Role of the Antidepressants Fluoxetine and Fluvoxamine for treating COVID-19. IJMS 2022, 23, 3812. [Google Scholar] [CrossRef]
- Al-Hajeri, H.; Baroun, F.; Abutiban, F.; Al-Mutairi, M.; Ali, Y.; Alawadhi, A.; Albasri, A.; Aldei, A.; AlEnizi, A.; Alhadhood, N.; et al. Therapeutic Role of Immunomodulators during the COVID-19 Pandemic: a Narrative Review. Postgraduate Medicine 2022, 134, 160–179. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, S.C.J.; Tse, C.L.Y.; Burry, L.; Dresser, L.D. Baricitinib: A Review of Pharmacology, Safety, and Emerging Clinical Experience in COVID-19. Pharmacotherapy 2020, 40, 843–856. [Google Scholar] [CrossRef]
- Frediansyah, A.; Tiwari, R.; Sharun, K.; Dhama, K.; Harapan, H. Antivirals for COVID-19: A Critical Review. Clinical Epidemiology and Global Health 2021, 9, 90–98. [Google Scholar] [CrossRef]
- Villa, T.G.; Abril, A.G.; Sánchez, S.; De Miguel, T.; Sánchez-Pérez, A. Animal and Human RNA Viruses: Genetic Variability and the Ability to Overcome Vaccines. Arch Microbiol 2021, 203, 443–464. [Google Scholar] [CrossRef]
- Checkmahomed, L.; Carbonneau, J.; Du Pont, V.; Riola, N.C.; Perry, J.K.; Li, J.; Paré, B.; Simpson, S.M.; Smith, M.A.; Porter, D.P.; et al. In Vitro Selection of Remdesivir-Resistant SARS-CoV-2 Demonstrates High Barrier to Resistance. Antimicrob Agents Chemother 2022, 66, e00198–22. [Google Scholar] [CrossRef] [PubMed]
- Szemiel, A.M.; Merits, A.; Orton, R.J.; MacLean, O.A.; Pinto, R.M.; Wickenhagen, A.; Lieber, G.; Turnbull, M.L.; Wang, S.; Furnon, W.; et al. In Vitro Selection of Remdesivir Resistance Suggests the Evolutionary Predictability of SARS-CoV-2. PLoS Pathog 2021, 17, e1009929. [Google Scholar] [CrossRef]
- Sanderson, T.; Hisner, R.; Donovan-Banfield, I.; Hartman, H.; Løchen, A.; Peacock, T.P.; Ruis, C. A Molnupiravir-Associated Mutational Signature in the Global SARS-CoV-2 Genomes. Nature 2023, 623, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; He, G.; Huang, W. A Novel Model of Molnupiravir against SARS-CoV-2 Replication: Accumulated RNA Mutations to Induce Error Catastrophe. Sig Transduct Target Ther 2021, 6, 410. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.T.; Yang, Q.; Gribenko, A.; Perrin, B.S.; Zhu, Y.; Cardin, R.; Liberator, P.A.; Anderson, A.S.; Hao, L. Genetic Surveillance of SARS-CoV-2 Mpro Reveals High Sequence and Structural Conservation before the Introduction of Protease Inhibitor Paxlovid. mBio 2022, 13, e00869–22. [Google Scholar] [CrossRef]
- Peluso, M.J.; Deeks, S.G. Mechanisms of Long COVID and the Path toward Therapeutics. Cell 2024, 187, 5500–5529. [Google Scholar] [CrossRef]
- Lupașcu (Moisi), R.E.; Ilie, M.I.; Velescu, B. Ștefan; Udeanu, D.I.; Sultana, C.; Ruță, S.; Arsene, A.L. COVID-19-Current Therapeutical Approaches and Future Perspectives. Processes 2022, 10, 1053. [Google Scholar] [CrossRef]
- Chala, B.; Tilaye, T.; Waktole, G. Re-Emerging COVID-19: Controversy of Its Zoonotic Origin, Risks of Severity of Reinfection and Management. IJGM 2023, Volume 16, 4307–4319. [Google Scholar] [CrossRef]
- Gu, X.; Zheng, M.; Gao, Y.; Lin, S.; Zhang, X.; Chen, C.; Zhu, H.; Sun, W.; Zhang, Y. Overview of Host-Directed Antiviral Targets for Future Research and Drug Development. Acta Pharmaceutica Sinica B 2025, S2211383525001431. [Google Scholar] [CrossRef]
- Schuller, M.; Zarganes-Tzitzikas, T.; Bennett, J.; De Cesco, S.; Fearon, D.; Von Delft, F.; Fedorov, O.; Brennan, P.E.; Ahel, I. Discovery and Development Strategies for SARS-CoV-2 NSP3 Macrodomain Inhibitors. Pathogens 2023, 12, 324. [Google Scholar] [CrossRef]
- Petushkova, A.I.; Zamyatnin, A.A. Papain-Like Proteases as Coronaviral Drug Targets: Current Inhibitors, Opportunities, and Limitations. Pharmaceuticals 2020, 13, 277. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, M.; Damalanka, V.C.; Tartell, M.A.; Chung, D.H.; Lourenço, A.L.; Pwee, D.; Mayer Bridwell, A.E.; Hoffmann, M.; Voss, J.; Karmakar, P.; et al. A Novel Class of TMPRSS2 Inhibitors that Potently Block SARS-CoV-2 and MERS-CoV Viral Entry and Protect Human Epithelial Lung Cells. Proc. Natl. Acad. Sci. U.S.A. 2021, 118, e2108728118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yu, W. Heat Shock Proteins and Viral Infection. Front. Immunol. 2022, 13, 947789. [Google Scholar] [CrossRef]
- Wickramaratne, A.C.; Wickner, S.; Kravats, A.N. Hsp90, a Team Player in Protein Quality Control and the Stress Response in Bacteria. Microbiol Mol Biol Rev 2024, 88, e00176–22. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, T.; Song, M.; Jin, M.; Liu, S.; Guo, K.; Zhang, Y. Rab1b-GBF1-ARFs Mediated Intracellular Trafficking Is Required for Classical Swine Fever Virus Replication in Swine Umbilical Vein Endothelial Cells. Veterinary Microbiology 2020, 246, 108743. [Google Scholar] [CrossRef]
- Chikhoune, L.; Poggi, C.; Moreau, J.; Dubucquoi, S.; Hachulla, E.; Collet, A.; Launay, D. JAK Inhibitors (JAKi): Mechanisms of Action and Perspectives in Systemic and Autoimmune Diseases. La Revue de Médecine Interne 2025, 46, 89–106. [Google Scholar] [CrossRef]
- Sodeifian, F.; Nikfarjam, M.; Kian, N.; Mohamed, K.; Rezaei, N. The Role of Type I Interferon for treating COVID-19. Journal of Medical Virology 2022, 94, 63–81. [Google Scholar] [CrossRef] [PubMed]
- Savan, R.; Gale, M. Innate Immunity and Interferon in the SARS-CoV-2 Infection Outcome. Immunity 2023, 56, 1443–1450. [Google Scholar] [CrossRef]
- Floresta, G.; Zagni, C.; Gentile, D.; Patamia, V.; Rescifina, A. Artificial Intelligence Technologies for COVID-19 De Novo Drug Design. IJMS 2022, 23, 3261. [Google Scholar] [CrossRef]
- Chodera, J.; Lee, A.A.; London, N.; Von Delft, F. Crowdsourcing Drug Discovery for Pandemics. Nat. Chem. 2020, 12, 581–581. [Google Scholar] [CrossRef]
- Faisal, S.; Badshah, S.L.; Kubra, B.; Sharaf, M.; Emwas, A.-H.; Jaremko, M.; Abdalla, M. Computational Study of SARS-CoV-2 RNA Dependent RNA Polymerase Allosteric Site Inhibition. Molecules 2021, 27, 223. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Yang, S.; He, S.; Li, F. AI Drug Discovery Tools and Analysis Technology: New Methods Aid in Studying the Compatibility of Traditional Chinese Medicine. Pharmacological Research-Modern Chinese Medicine 2025, 14, 100566. [Google Scholar] [CrossRef]
- Maghsoudi, S.; Taghavi Shahraki, B.; Rameh, F.; Nazarabi, M.; Fatahi, Y.; Akhavan, O.; Rabiee, M.; Mostafavi, E.; Lima, E.C.; Saeb, M.R.; et al. A Review on Computer-aided Chemogenomics and Drug Repositioning for Rational COVID -19 Drug Discovery. Chem Biol Drug Des 2022, 100, 699–721. [Google Scholar] [CrossRef] [PubMed]
- Mottaqi, M.S.; Mohammadipanah, F.; Sajedi, H. Contribution of Machine Learning Approaches in Response to SARS-CoV-2 Infection. Informatics in Medicine Unlocked 2021, 23, 100526. [Google Scholar] [CrossRef] [PubMed]
- Rajput, A.; Thakur, A.; Mukhopadhyay, A.; Kamboj, S.; Rastogi, A.; Gautam, S.; Jassal, H.; Kumar, M. Prediction of Repurposed Drugs for Coronaviruses Using Artificial Intelligence and Machine Learning. Computational and Structural Biotechnology Journal 2021, 19, 3133–3148. [Google Scholar] [CrossRef]
- Huchting, J. Targeting Viral Genome Synthesis as a Broad-Spectrum Approach against RNA Virus Infections. Antivir Chem Chemother 2020, 28, 204020662097678. [Google Scholar] [CrossRef]
- Tolksdorf, B.; Heinze, J.; Niemeyer, D.; Röhrs, V.; Berg, J.; Drosten, C.; Kurreck, J. Development of a Highly Stable, Active Small Interfering RNA with Broad Activity against SARS-CoV Viruses. Antiviral Research 2024, 226, 105879. [Google Scholar] [CrossRef]
- Abbott, T.R.; Dhamdhere, G.; Liu, Y.; Lin, X.; Goudy, L.; Zeng, L.; Chemparathy, A.; Chmura, S.; Heaton, N.S.; Debs, R.; et al. Development of CRISPR as an Antiviral Strategy to Combat SARS-CoV-2 and Influenza. Cell 2020, 181, 865–876.e12. [Google Scholar] [CrossRef]
- Li, T.; Yang, Y.; Qi, H.; Cui, W.; Zhang, L.; Fu, X.; He, X.; Liu, M.; Li, P.; Yu, T. CRISPR/Cas9 Therapeutics: Progress and Prospects. Sig Transduct Target Ther 2023, 8, 36. [Google Scholar] [CrossRef]
- Najafi, S.; Tan, S.C.; Aghamiri, S.; Raee, P.; Ebrahimi, Z.; Jahromi, Z.K.; Rahmati, Y.; Sadri Nahand, J.; Piroozmand, A.; Jajarmi, V.; et al. Therapeutic Potentials of the CRISPR-Cas Genome Editing Technology in Human Viral Infections. Biomedicine and Pharmacotherapy 2022, 148, 112743. [Google Scholar] [CrossRef]
- Von Delft, A.; Hall, M.D.; Kwong, A.D.; Purcell, L.A.; Saikatendu, K.S.; Schmitz, U.; Tallarico, J.A.; Lee, A.A. Accelerating Antiviral Drug Discovery: Lessons from COVID-19. Nat Rev Drug Discov 2023, 22, 585–603. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, A.; Ludwig, S. Host-Targeted Antivirals against SARS-CoV-2 in Clinical Development-Prospect or Disappointment? Antiviral Research 2025, 235, 106101. [Google Scholar] [CrossRef] [PubMed]
- Seley-Radtke, K.L.; Thames, J.E.; Waters, C.D. Broad Spectrum Antiviral Nucleosides—Our Best Hope for the Future. In Annual Reports in Medicinal Chemistry; Elsevier, 2021; Vol. 57, pp. 109–132 ISBN 978-0-323-91511-3.
- Kirchdoerfer, R.N.; Ward, A.B. Structure of the SARS-CoV Nsp12 Polymerase Bound to the Nsp7 and Nsp8 Co-Factors. Nat Commun 2019, 10, 2342. [Google Scholar] [CrossRef] [PubMed]
- Xiong, R.; Zhang, L.; Li, S.; Sun, Y.; Ding, M.; Wang, Y.; Zhao, Y.; Wu, Y.; Shang, W.; Jiang, X.; et al. Novel and Potent Inhibitors Targeting DHODH Are Broad-Spectrum Antivirals against RNA Viruses, Including the Newly-Emerged Coronavirus SARS-CoV-2. Protein Cell 2020, 11, 723–739. [Google Scholar] [CrossRef]
- Sievers, B.L.; Cheng, M.T.K.; Csiba, K.; Meng, B.; Gupta, R.K. SARS-CoV-2 and Innate Immunity: The Good, the Bad, and the “Goldilocks. ” Cell Mol Immunol 2023, 21, 171–183. [Google Scholar] [CrossRef]
- Italiya, J.; Bhavsar, T.; Černý, J. Assessment and Strategy Development for SARS-CoV-2 Screening in Wildlife: A Review. Vet World 2023, 1193–1200. [Google Scholar] [CrossRef]
- Karim, M.; Lo, C.-W.; Einav, S. Preparing for the next Viral Threat with Broad-Spectrum Antivirals. Journal of Clinical Investigation 2023, 133, e170236. [Google Scholar] [CrossRef]
Figure 1.
SARS-CoV-2 replication cycle and key molecular events targeted by antivirals. The SARS-CoV-2 replication cycle starts with (1) binding and viral entry, in which the virus attaches to the host cell receptor (such as ACE2) and is internalized by endocytosis or membrane fusion. The viral genome is then released into the cytoplasm. For (3) translation of viral polyprotein including the RNA-dependent RNA polymerase (RdRp) and other non-structural proteins (nsps), the positive-sense RNA genome is directly employed as mRNA. To generate both full-length genomic RNA and subgenomic RNAs, these proteins form the replication-transcription complex (RTC), therefore mediating (4) RNA replication and transcription. At the endoplasmic reticulum, the subgenomic RNA act as templates for (5) translation of the structural proteins S, E, M, and N. These proteins travel to assembly sites where (6) structural proteins mix with the nucleocapsid, which encapsidate the fresh genomic RNA. Finally, (7) virion assembly occurs and (8) exocytosis releases new virions. Figure created with BioRender.com, adapted from Guo, 2020 [18] and Fehr, et al. 2015 [19], licensed under CC BY 4.0.
Figure 1.
SARS-CoV-2 replication cycle and key molecular events targeted by antivirals. The SARS-CoV-2 replication cycle starts with (1) binding and viral entry, in which the virus attaches to the host cell receptor (such as ACE2) and is internalized by endocytosis or membrane fusion. The viral genome is then released into the cytoplasm. For (3) translation of viral polyprotein including the RNA-dependent RNA polymerase (RdRp) and other non-structural proteins (nsps), the positive-sense RNA genome is directly employed as mRNA. To generate both full-length genomic RNA and subgenomic RNAs, these proteins form the replication-transcription complex (RTC), therefore mediating (4) RNA replication and transcription. At the endoplasmic reticulum, the subgenomic RNA act as templates for (5) translation of the structural proteins S, E, M, and N. These proteins travel to assembly sites where (6) structural proteins mix with the nucleocapsid, which encapsidate the fresh genomic RNA. Finally, (7) virion assembly occurs and (8) exocytosis releases new virions. Figure created with BioRender.com, adapted from Guo, 2020 [18] and Fehr, et al. 2015 [19], licensed under CC BY 4.0.
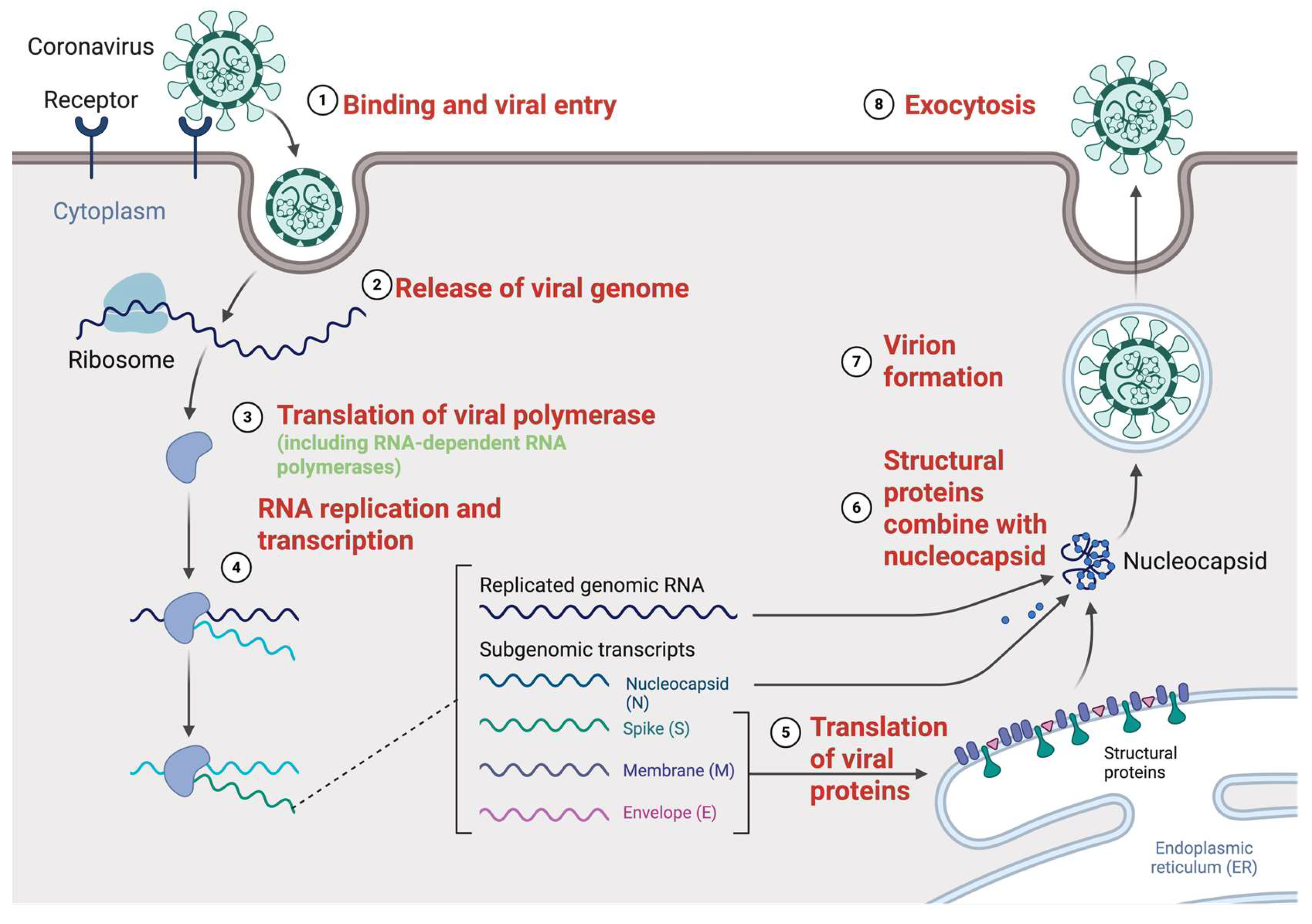
Figure 2.
SARS-CoV-2’s core viral replication machinery is made up of the nsp12–nsp7–nsp8 supercomplex, which reflects the minimal functional unit needed for RNA synthesis. The nsp12 subunit (A-chain, red) is the main catalytic polymerase. The nsp7 cofactor (C-chain, depicted in green) binds to nsp12 and stabilizes the system. Acting as cofactors and extensions of the template RNA-binding platform, the nsp8 subunits (B-chain in cyan and D-chain in purple) increase the processivity. Using Robetta, comparative modeling produced this structural model; the template structure was PDB ID: 8GWE. Reproduced from Subong, B.J.J., & Ozawa, T. (2024) [20]. Licensed under CC BY 4.0.
Figure 2.
SARS-CoV-2’s core viral replication machinery is made up of the nsp12–nsp7–nsp8 supercomplex, which reflects the minimal functional unit needed for RNA synthesis. The nsp12 subunit (A-chain, red) is the main catalytic polymerase. The nsp7 cofactor (C-chain, depicted in green) binds to nsp12 and stabilizes the system. Acting as cofactors and extensions of the template RNA-binding platform, the nsp8 subunits (B-chain in cyan and D-chain in purple) increase the processivity. Using Robetta, comparative modeling produced this structural model; the template structure was PDB ID: 8GWE. Reproduced from Subong, B.J.J., & Ozawa, T. (2024) [20]. Licensed under CC BY 4.0.
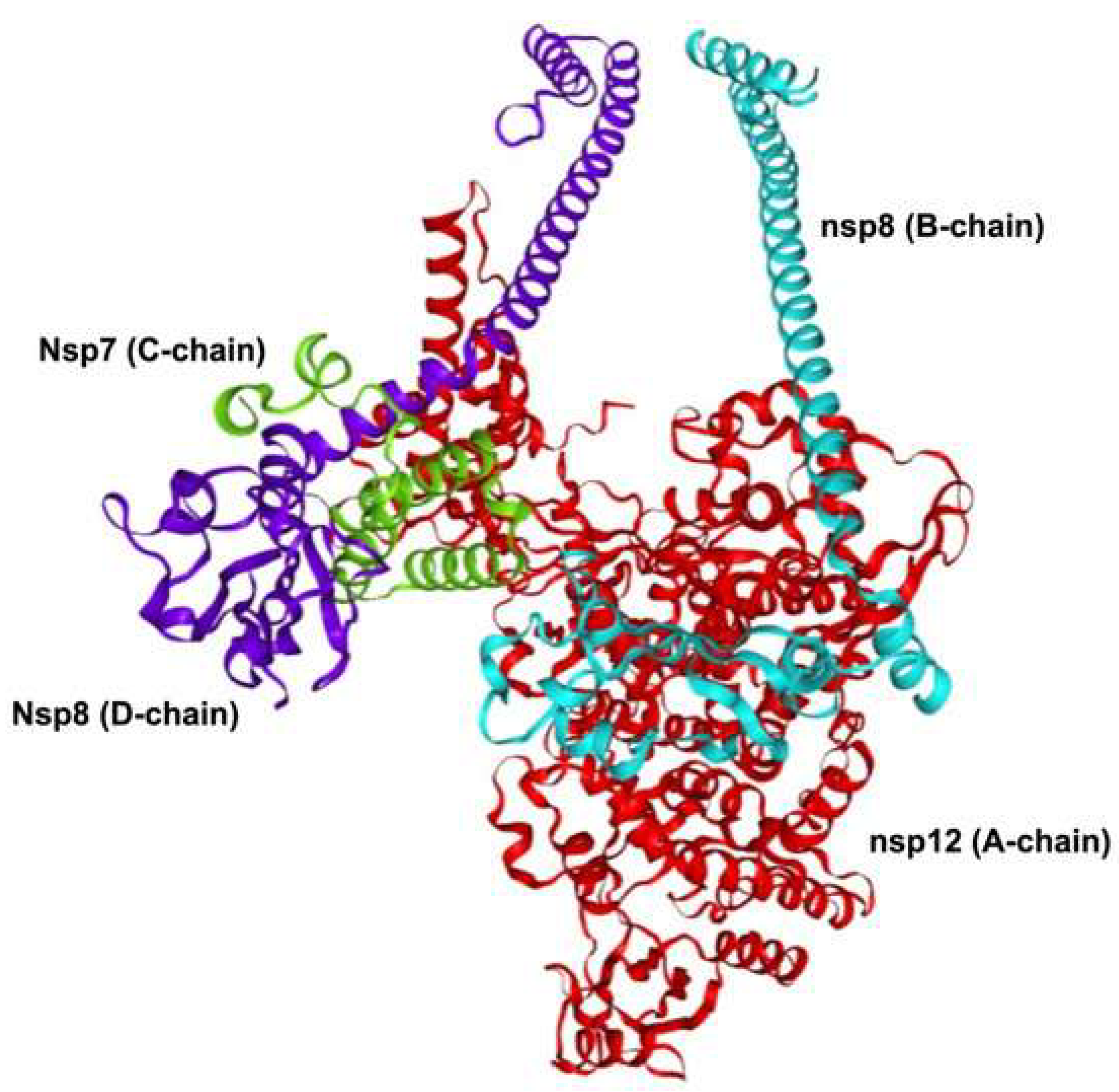
Figure 3.
Action mechanism of remdesivir and its active metabolite GS-441524 against SARS-CoV-2 replication. SARS-CoV-2, upon entering the host cell, uses the host ribosome to convert the RNA-dependent RNA polymerase (RdRp), a main enzyme encoded inside ORF1b. The monophosphoramidate prodrug remdesivir is metabolized intracellularly to its active nucleoside analogue, GS-441524. RdRp includes this molecule in nascent viral RNA, where it mimics adenosine. Once included, it causes postponed chain termination, therefore stopping more elongation of the RNA strand and thus stopping viral replication. Highlighting the binding pocket of GS-441524, the bottom right inset reveals the RdRp holoenzyme structure (PDB ID: 6M71). Figure created with BioRender.com, adapted from Li, G. & De Clercq, E. (2020) [51], licensed under CC BY 4.0.
Figure 3.
Action mechanism of remdesivir and its active metabolite GS-441524 against SARS-CoV-2 replication. SARS-CoV-2, upon entering the host cell, uses the host ribosome to convert the RNA-dependent RNA polymerase (RdRp), a main enzyme encoded inside ORF1b. The monophosphoramidate prodrug remdesivir is metabolized intracellularly to its active nucleoside analogue, GS-441524. RdRp includes this molecule in nascent viral RNA, where it mimics adenosine. Once included, it causes postponed chain termination, therefore stopping more elongation of the RNA strand and thus stopping viral replication. Highlighting the binding pocket of GS-441524, the bottom right inset reveals the RdRp holoenzyme structure (PDB ID: 6M71). Figure created with BioRender.com, adapted from Li, G. & De Clercq, E. (2020) [51], licensed under CC BY 4.0.
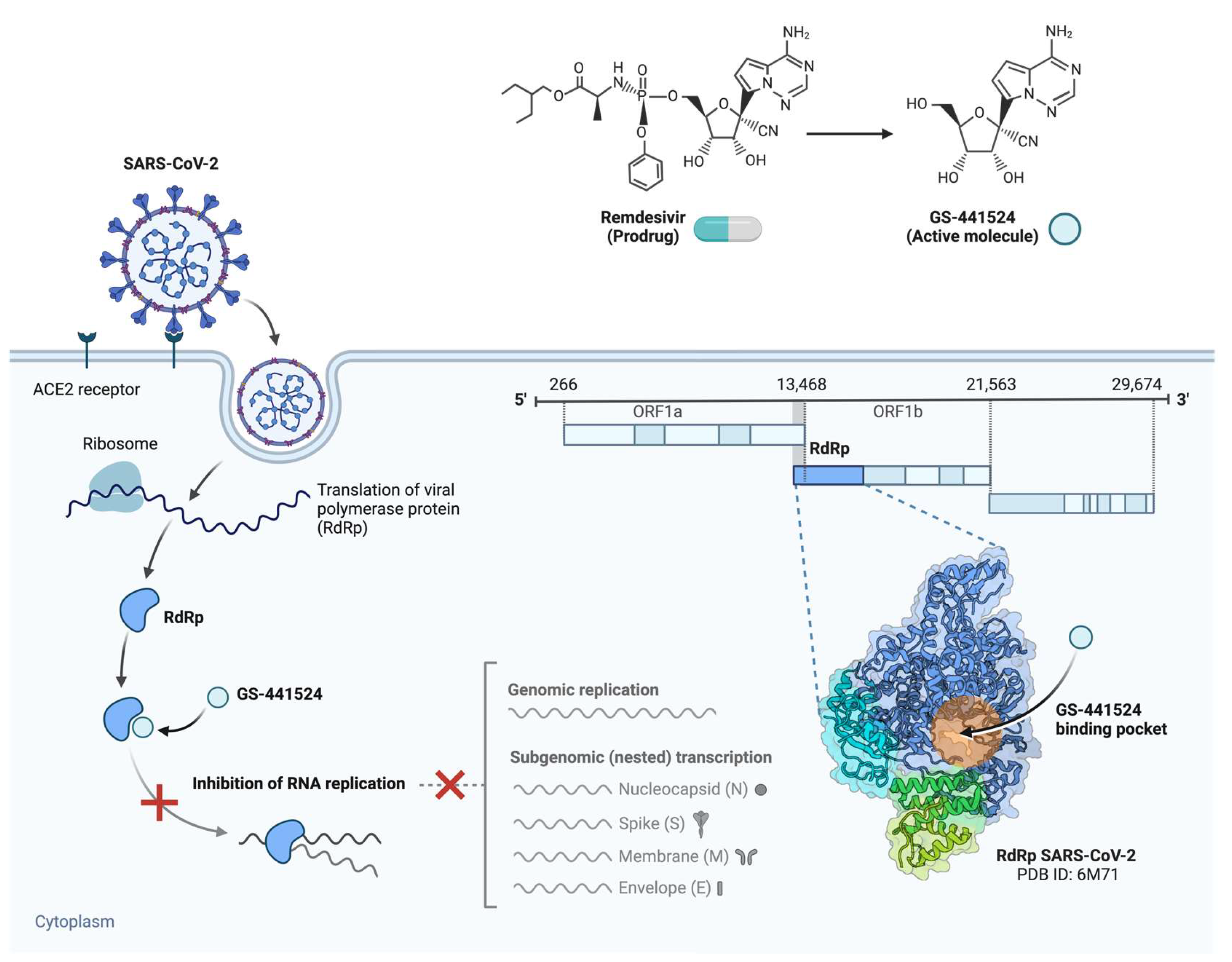
Figure 4.
Action mechanism of mutagenic nucleoside analogues (molnupiravir) in SARS-CoV-2 replication. Viral entry causes the host cell to translate the viral RNA-dependent RNA polymerase (RdRp), which drives RNA synthesis. Competing with natural cytidine triphosphate (CTP), mutagenic nucleoside analogues like the active triphosphate form of molnupiravir (MTP) are misincorporate into viral RNA. Base-pair mismatches and the buildup of mutations follow from this, causing viral inactivation—a process called lethal mutagenesis. This approach enhances molnupiravir’s antiviral effectiveness against several SARS-CoV-2 variants.Figure created with BioRender.com, adapted from Malone, B. & Campbell, E.A. (2021)[59], licensed under CC BY 4.0.
Figure 4.
Action mechanism of mutagenic nucleoside analogues (molnupiravir) in SARS-CoV-2 replication. Viral entry causes the host cell to translate the viral RNA-dependent RNA polymerase (RdRp), which drives RNA synthesis. Competing with natural cytidine triphosphate (CTP), mutagenic nucleoside analogues like the active triphosphate form of molnupiravir (MTP) are misincorporate into viral RNA. Base-pair mismatches and the buildup of mutations follow from this, causing viral inactivation—a process called lethal mutagenesis. This approach enhances molnupiravir’s antiviral effectiveness against several SARS-CoV-2 variants.Figure created with BioRender.com, adapted from Malone, B. & Campbell, E.A. (2021)[59], licensed under CC BY 4.0.
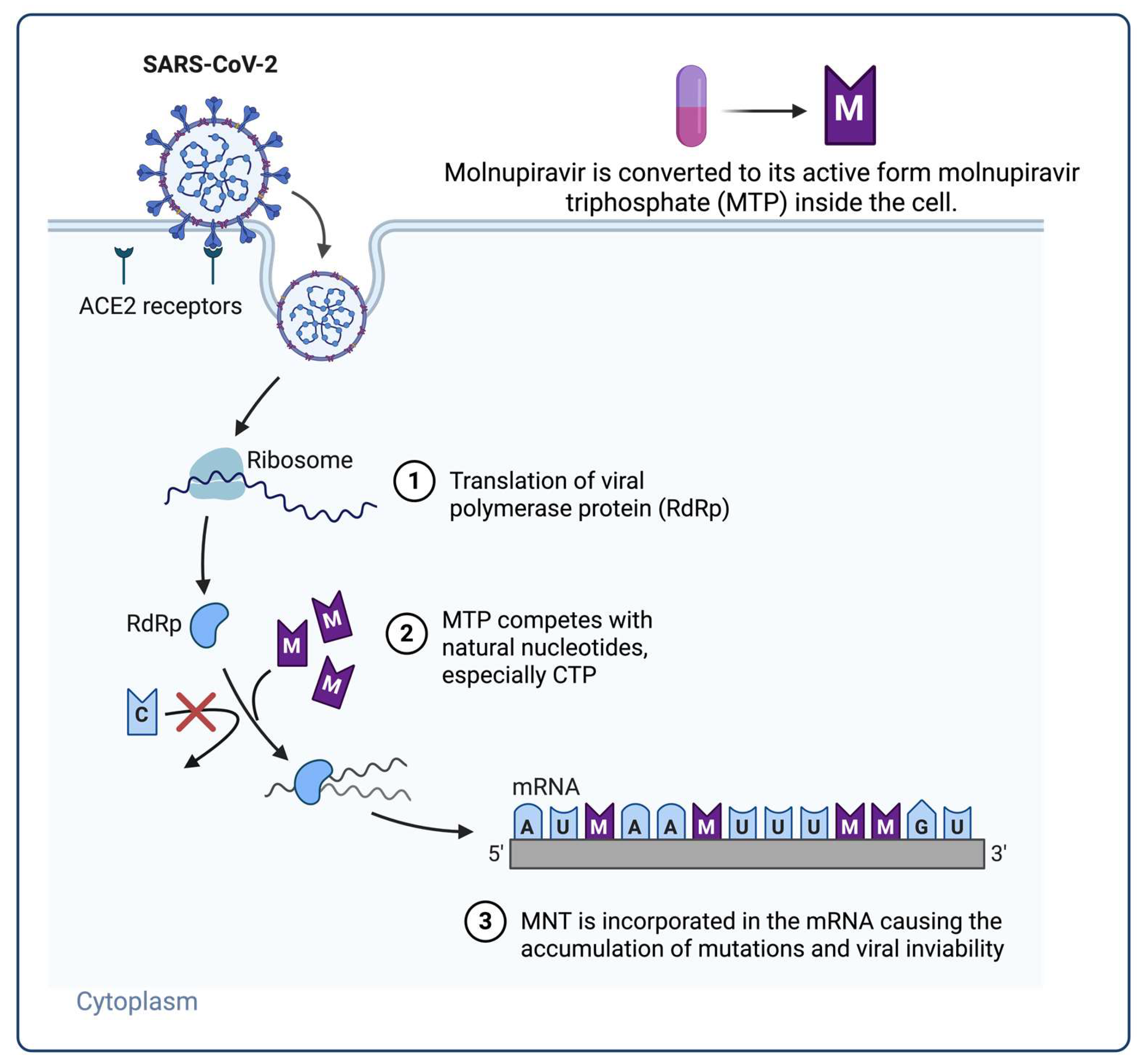

Table 1.
Summary of Antiviral Therapeutics Targeting SARS-CoV-2.
| Drug/Compound | Target/Mechanism | Drug Class | Mode of Action | Clinical Use/Efficacy | Resistance/Limitations |
| Remdesivir | RdRp (nsp12) | Nucleoside analogue | Adenosine analogue; chain termination via delayed RNA translocation | IV use; moderately accelerates recovery in hospitalized patients | Partial resistance via nsp12 mutations (e.g., V166, E802); fitness cost limits spread |
| Molnupiravir | RdRp | Nucleoside analogue | Cytidine/uridine analogue; induces lethal mutagenesis | Oral use; reduces hospitalization in mild-to-moderate cases | Theoretical mutagenic risks; not fully neutralized by proofreading |
| Favipiravir | RdRp | Nucleoside analogue | Purine analogue; weak RNA synthesis inhibition | Limited effect; not a frontline drug | Requires high concentrations; inconsistent trial results |
| Ribavirin | RdRp | Nucleoside analogue | Guanosine analogue; promotes mutagenesis | Alone: limited effect; synergistic in combinations | High toxicity; synergizes with remdesivir |
| VV116 (Deuviridine) | RdRp | Nucleoside analogue | Oral prodrug of GS-441524 | Comparable to Paxlovid in trials; high bioavailability | Not widely approved |
| Nirmatrelvir + Ritonavir (Paxlovid) | 3CLpro | Protease inhibitor | Blocks polyprotein cleavage; halts replication complex maturation | Oral use; ~89% reduction in hospitalization if early | Resistance via 3CLpro mutations (in vitro); clinical relevance not yet clear |
| Ensitrelvir | 3CLpro | Protease inhibitor | Similar to nirmatrelvir | Effective in Japan trials | Investigational |
| Lopinavir + Ritonavir | 3CLpro | Protease inhibitor | HIV protease inhibitor; weak SARS-CoV-2 activity | No benefit in early COVID-19 trials | Largely abandoned |
| GRL-0617 | PLpro | Protease inhibitor | Preclinical inhibitor of papain-like protease | Preclinical stage | Not clinically available |
| Fluoxetine | Host (cell pathways) | SSRI / host-directed | Synergistic with GS-441524; may block viral egress | In vitro synergy with remdesivir | Lysosomotropic mechanism; repurposed drug |
| Itraconazole | Host (cholesterol trafficking) | Antifungal / host-directed | Disrupts sterol trafficking needed for replication | Enhances remdesivir efficacy in vitro | Repurposed, not virus-specific |
| Baricitinib | Host (JAK/STAT + endocytosis) | JAK inhibitor/immunomodulator | Blocks cytokine storm; may block viral entry | EUA with remdesivir; reduces inflammation | Minor antiviral activity; adjunctive |
| Camostat | TMPRSS2 | Host entry inhibitor | Blocks spike protein priming | In vitro efficacy; used in combinations | Investigational |
| Brequinar | DHODH (pyrimidine synthesis) | Host-targeting | Reduces the nucleotide pools needed for RNA synthesis | Strong synergy with molnupiravir | Host toxicity concerns |
| Remdesivir + Nirmatrelvir | RdRp + 3CLpro | Antiviral synergy | Blocks RNA synthesis and protein processing | Superior in vitro and in case reports | Dual targeting reduces the resistance potential |
| Remdesivir + Ribavirin | RdRp | Chain terminator + mutagen | Complete viral extinction in vitro | Enhances the mutational burden and replication block | Not in clinical use |
| Molnupiravir + Nirmatrelvir | RdRp + 3CLpro | Dual-action antiviral | Polymerase + protease inhibition | Enhanced synergy; potential for resistant strains | Preclinical and early trial stages |
| Molnupiravir + Camostat + Brequinar | Multi-target | Polymerase + entry + nucleotide synthesis | Maximal suppression in vitro | For persistent/resistant infections | Not tested clinically |
Table 2.
Future Trends and Emerging Antiviral Strategies Against SARS-CoV-2.
| Strategy | Targets/Approaches | Key Compounds/Tools | Potential Impact |
| Novel Viral Targets | nsp14 exonuclease (ExoN), the NiRAN domain of nsp12, PLpro, and macrodomains | PLpro inhibitors (e.g., GRL-0617), and experimental NiRAN inhibitors | Enhances polymerase inhibitor efficacy; reduces immune suppression; introduces new viral drug targets |
| Host-Directed Antivirals | TMPRSS2, pyrimidine biosynthesis (e.g., DHODH), TRiC, Hsp90, JAK-STAT pathways, IFN-λ system | Camostat, nafamostat, brequinar, baricitinib, teriflunomide, interferon lambda (IFN-λ) | Reduced viral resistance potential; broad-spectrum activity; may pose host toxicity risks |
| AI and Computational Drug Discovery | Structure-based virtual screening, machine learning prediction, synergy modeling, allosteric pockets on RdRp (nsp12) | COVID Moonshot, flavonoid derivatives (e.g., myricetin), ML-based compound ranking | Accelerates drug discovery; identifies novel scaffolds; enables variant-specific antiviral tailoring |
| Gene silencing and RNA-Targeting Therapies | siRNA targeting conserved genome regions (e.g., RdRp); CRISPR-Cas13-mediated cleavage of viral RNA | siRNA-nanoparticle delivery systems and Cas13-based antiviral platforms | Programable and rapidly deployable antivirals; adaptable to emerging viruses |
| Pan-Coronavirus and Broad-Spectrum Antivirals | Conserved viral domains (nsp12-nsp7/nsp8 interface, nsp13 helicase); shared host dependency factors (e.g., DHODH, IFN) | Ribavirin, NHC, DHODH inhibitors, innate immune modulators | Enables pandemic preparedness; broad viral coverage; strategic stockpiling for future zoonotic outbreaks |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



