
Submitted:
23 April 2025
Posted:
24 April 2025
You are already at the latest version
Abstract
Saline-alkali soils are characterized by high salt content and low organic matter, which hinders plant growth and limits their carbon sequestration capacity. Using of biofertilizer has been recognized as a promising strategy for saline-alkali soil improvement. This study evaluated the potential of biofertilizer to improve the saline-alkali soil properties and enhance carbon sequestration through field experiments, while also exploring the underlying mechanisms. The soil supplemented with 30 and 60 t·ha-1 of biofertilizer resulted into an increase in per-hectare carbon sequestration by 102 t CO2e and 144 t CO2e, respectively, in comparison to soil without biofertilizer (control). The improvement in soil properties showed an increase in Suaeda salsa biomass, enhancements in soil enzymes activities and elevation in total organic content, as evident by correlation analysis and Mantel tests. Furthermore, the metabolism of organic matter promotes an increase in total inorganic carbon. The results indicates that the use of biofertilizer is an effective strategy for improving saline-alkali soil and enhancing carbon sequestration.
Keywords:
aline-alkali soil
; biofertilizer
; carbon sequestration
; Suaeda salsa
; soil enzyme activity
1. Introduction
Terrestrial ecosystems store large amounts of soil organic carbon (SOC) and soil inorganic carbon (SIC), which are twice the size of the atmospheric carbon pools and four times that of the biotic carbon pools [1]. However, widespread soil salinization and alkalinization severely inhibit the ability of plants to absorb and fix carbon dioxide, thereby reducing soil terrestrial ecosystems carbon pool [2]. With the over-exploitation of land resources, the degree of soil salinization and alkalinization is increasing [3]. In China, there are up to 3.69×107 ha of saline-alkali soil, of which 9.21×106 ha is cultivated land, which represents 6.6% of the country's total cultivated land area [4]. Excessive alkalinity and salinity in soil lead to the degradation of soil structure and reduction in its fertility. Furthermore, these conditions cause ionic imbalance and stress in plant cells, which negatively affect crop growth and threaten grain security [5,6]. Therefore, developing effective strategies for improving saline-alkali soil is essential for achieving sustainable agricultural development, enhancing soil carbon sequestration, and mitigating climate change.
The organic carbon pool is a crucial component of the global carbon cycle. The processes of organic carbon production and degradation can significantly influence the absorption and emission of carbon dioxide [7]. Plants absorb, store, and utilize atmospheric carbon dioxide through photosynthesis, converting it into organic matter [8,9]. This process increases plant biomass and SOC content [10]. Additionally, plant biomass itself acts as an organic carbon pool, which can further increase the carbon sequestration [11]. Recent studies show that SIC is also an important part of soil carbon pool. Soil in saline-alkali regions contain large amount of SIC. The SIC is an inert carbon, and its residence time in the soil is much longer than that of SOC, by a factor of 10 to 10,000 [12]. Therefore, both SOC and SIC in saline-alkali soil should be included in the soil carbon sequestration accounting system.
Applying organic amendments is a common method for improving saline-alkali soil. Previous studies have shown that long-term application of organic fertilizers improves the properties of saline-alkali soil and increases its total organic carbon (TOC) content [5]. Li et al. found that adding organic materials to saline-alkali soil significantly increases plant biomass and SOC [13]. Zhang et al. found that returning crop residues to the field significantly reduces soil salinity [14]. This reduction enhances aggregate stability, thereby increasing SOC content. The inoculation of saline-alkali tolerant plant growth-promoting bacteria is an effective method to increase plant biomass in saline-alkali soil [15]. Besides, organic fertilizers can promote soil fertility and microbial community activity, establishing optimal environment for plant growth [16]. The application of organic fertilizers and the inoculation of plant growth-promoting rhizobacteria (PGPR) are both effective methods of soil improvement. To promote crop growth and increase carbon sequestration, the synergistic effects of organic fertilizers and PGPR should be emphasized [17]. Additionally, soil enzymes play a crucial role in soil carbon cycling by directly influencing the mineralization and decomposition of organic carbon, while also releasing inorganic nutrients that plants can absorb [18,19]. Studies have reported that applying biofertilizer can reduce nitrogen loss from farmland, decrease NH3 volatilization, and lower emissions of N2O and other greenhouse gases [20,21,22]. However, there is still limited research on assessing the response of soil carbon sequestration to the application of biofertilizer.
Saline-alkali soil is regarded as a complex system, where the formation of soil carbon may involve multiple mechanisms [23]. In the early stage, we did an experiment about biofertilizer improving saline-alkali land and promoting the growth of crop [24]. In order to further explore its influence on carbon sequestration in agricultural ecosystem, a field experiment was conducted in the region. Our objectives were to: (1) examine the effects of biofertilizer on the improvement of Saline-alkali soil and promote the growth of Suaeda salsa; (2) assess the carbon sequestration capacity in the investigated field; (3) explore the mechanism for increase in carbon sequestration, S. salsa biomass, and soil enzymes activities.
2. Materials and Methods
2.1. Experimental Design
The field experiment was conducted in the saline botanical garden (45°26′8′′N, 85°0′6′′E) in Karamay City, Xinjiang Uygur Autonomous Region, China (Figure 1). The region is located in a temperate continental arid climate zone, with an average annual precipitation of 105 mm and an average annual evaporation of 3009 mm. Additionally, the temperature fluctuates significantly with the seasons, reaching a maximum of 43.6 °C in summer and a minimum of -40.5 °C in winter. The main vegetation in the region is xerophytes with salt tolerance. This area has a high degree of soil salinization, low soil fertility and weak carbon sequestration capacity [25,26,27].
2.2. Field Experiment Design
Table S1 presents the physicochemical properties of soil and characteristics of the biofertilizer for this experiment. The biofertilizer contains Bacillus subtilis at a concentration exceeding 2.0×108 CFU·g-1, in accordance with the Chinese standard NY 884-2012. S. salsa is a common salt-tolerant crop and it can thrive in a drip irrigation system without fertilization [28]. In the current study, a field experiment was conducted to investigate the effect of biofertilizer on S. salsa and its impact on the carbon sequestration process. The field experiment included three treatments: CK (with no biofertilizer), A (60 t·ha-1 biofertilizer), and B (30 t·ha-1 biofertilizer). Each treatment was replicated three times and randomly distributed across 6 m × 10 m plots. All treatments involved the application of only the biofertilizer, with no additional supplementation. Prior to addition of biofertilizer, the plots were leveled and cleared for stones and weed roots. The biofertilizer was spread evenly on the surface of the plots using a 35 cm rotary tiller through multiple passes. After adding biofertilizer, drip irrigation tapes were installed for all treatments, with all other conditions kept consistent across treatments.
2.3. Soil Sampling and Measurement
Baseline soil samples were collected before the start of the experiment. Subsequently, soil samples were collected after adding biofertilizer (May 2021, T1), during the seedling stage of S. salsa (July 2021, T2), and during the fruiting stage of S. salsa (September 2021, T3). The additional soil samples were collected eight months after harvest in May 2022 (labelled as T4) in order to investigate long-term effect of biofertilizer application on soil. Nine soil samples were collected from different locations within each treatment at a depth of 0-35 cm. The collected samples were mixed into a composite sample, with three such composite samples for each treatment. In case of T2 and T3, rhizosphere soil samples were collected by shaking the soil adhering to the roots into sampling bags. Nine samples were collected, mixed into a composite sample, and this process was repeated three times. The collected soil samples were either refrigerated at 4 °C or air-dried for subsequent soil parameter analysis. Soil pH was measured using a pH meter (INESA, Shanghai, China) in a 1:2.5 (w/v) soil-water extract. Ammonium nitrogen (NH4+-N), nitrate nitrogen (NO3--N), and nitrite nitrogen (NO2--N) were determined using potassium chloride extraction and spectrophotometric methods (China Standard HJ 634-2012). Total carbon (TC), TOC, total inorganic carbon (TIC), total nitrogen (TN), and carbon-to-nitrogen ratio (C/N) were measured using an elemental analyzer (Vario MAX cube). Available phosphorus (AP) was determined using sodium bicarbonate extraction and molybdenum-antimony colorimetric method according to China Standard HJ 704-2014. Total soluble salt (TSS) content was measured according to China Standard NY 1121.16-2006. SOM was determined using the loss-on-ignition method according to China Standard HJ 761-2015. The soil water content (SWC) was determined by gravimetric method according to China standard HJ 613-2011. The concentration of different elements (Na, K, Mg, Ca, Cl) were determined using an Inductively Coupled Plasma Optical Emission Spectrometer (ICP-OES) (Varian, Vista Pro), and the anion SO42- content was determined using ion chromatography (Thermo Fisher, Aquion), referring to the Chinese standard HJ 832-2017.
2.4. Plant Sampling and Measurement
Plant samples were collected at the T2 and T3 stages. In each treatment, a 0.6 m × 0.6 m quadrat was randomly set up. After harvesting the plants within the quadrat, their height and fresh weight were measured. The plants were dried in an oven at 75 °C and cumulative biomass was obtained by summing the dry weight of all plants per unit area. The dried samples were then crushed, and sieved through a 200-mesh screen. Subsequently, the plant samples were analyzed to determine the TC, TIC, TOC, TN, C/N, and readily oxidizable carbon (ROC) using an elemental analyzer (Elementar S-TOC cube). Ash content was measured using the ignition method: the sieved samples were burnt in a muffle furnace at 600 °C for 3 hours, then cooled to room temperature and weighed. The concentration of different elements (Na, K, Mg, Ca, Cl) was determined using an Inductively Coupled Plasma Optical Emission Spectrometer (ICP-OES) (Varian, Vista Pro), referring to the Chinese standard HJ 832-2017 for metal detection methods.
2.5. Carbon Sequestration Accounting
Due to the uneven distribution of crop growth in different quadrats, we used cumulative biomass (the sum of the biomass of all crops per unit area) for carbon sequestration accounting. The plant carbon content ratio is the proportion of carbon in the plant’s total mass. The conversion factor between CO2 and C is 44/12. Referring to the Chinese standard DB31/T 1234-2020, the carbon sequestration per hectare of plants is calculated using the following formula:
In the formula, Cp represents the plant carbon sequestration amount (t CO2e), Bp represents the plant biomass (t·ha-1), CF represents the plant carbon content ratio (%), A represents the unit planting area (ha), and 3.67 is the conversion factor between CO2 and C.
Considering the lower solubility of carbonates in saline-alkali soil, their TIC content cannot be ignored. We used TC as an indicator to assess the carbon sequestration capacity of saline-alkali soil. Referring to the Chinese standard NY/T 4300-2023, the carbon sequestration per hectare of soil is calculated using the following formula:
In this formula, Cs represents the soil carbon sequestration amount (t CO2e), TC represents the total soil carbon content (mg·g⁻¹), D represents the soil bulk density (g·cm⁻³), E represents the soil depth (cm), A represents the unit area (ha), 3.67 is the conversion factor between CO2 and C, and 0.1 is the unit conversion factor.
2.6. Soil Enzymes Activities Measurement
The dried soil was cleared from stones, plant residues, and other debris, and then ground using a mortar and pestle, Finally, the soil was sieved it through a 100-mesh sieve and analyzed for various enzymes such as urease (UE), alkaline phosphatase (ALP), catalase (CAT), cellulase (CL) and sucrase (SC) by measuring their activity using enzyme assay kits (Solarbio Life Science, Beijing, China).
2.7. Statistical Analysis
The average values and standard deviations of the physicochemical properties of soil and biofertilizer carrier were compiled using Excel and Origin 2024 software. The significant difference in soil properties, plant biomass, plant physicochemical properties, and soil enzymes activities were tested using univariate analysis of variance (ANOVA) and Tukey test. Difference between rhizosphere and non-rhizosphere was tested by paired sample T test. Wilcoxon rank sum test was carried out with R v4.2.0, and 0.05 value was considered as the statistical significance threshold to test the difference in carbon and other indicators in different treatments. Linear regression analysis was conducted to examine the relationships between plant biomass and plant carbon-nitrogen content, plant salinity, soil properties, and enzymes activities. The Spearman correlation coefficient between plant carbon-nitrogen and salinity was calculated using the R “corrplot” package. The “linkET” package was used for Mantel tests to determine the impact of different environmental factors on the contents of TOC, TIC, and TC [29].
3. Results
3.1. Soil Physicochemical Properties
The physicochemical properties of soil were determined in order to explore the effect of biofertilizer on saline-alkali soil. A significant difference in physicochemical properties of soil was observed among different treatments (Table S2). In comparison to CK, the application of biofertilizer increased SOM content by 1 to 1.9 times. The SOM content was highest during the T3 period and significantly decreased after crop harvest (p < 0.05). The biofertilizer reduced soil TSS content by 16.3% to 22.6% as compared to CK, with significant differences between treatments A, B, and CK in all periods after sowing (T2, T3, T4). During the crop growing period (T2, T3), the SWC of treatment A was significantly higher than treatments B and CK. The application of biofertilizer significantly increased soil AP, with treatment A having the highest AP content. In comparison to CK, soil TN increased by 65.4% and 22.6% for treatments A and B, respectively. During the crop growing period (T2, T3), the soil NH4+-N and NO3 --N levels was also increased in comparison to that of CK. In general, the application of biofertilizer reduced soil TSS, increased SOM content and other nutrients, hence effectively improved saline-alkali soil.
In comparison to CK, soil Cl content in treatments A and B was decreased by 31.7% and 23.4%, respectively, whereas Na content was decreased by 23.9% and 26.2%, respectively. The application of biofertilizer resulted into a more significant reduction in Cl content, while effect on Na content was non-significant. No significant difference was observed between treatments for other elements and ions besides Na and Cl (Figure 2). In the original soil samples, the salt elements were primarily Na and Cl, which together accounted for over 60%. Biofertilizer altered the composition of soil salt, significantly reducing the proportions of Cl and Na in soil (Figure S1).
3.2. Biomass and Physiological Characteristics of S. salsa
The biomass and physiological characteristics of plant S. salsa were measured in order to determine the effect of biofertilizer on its growth. The results showed that the biofertilizer significantly improved the plant height, dry weight, fresh weight, and plant biomass (Figure 3). At T3, the dry weight of treatment A and B was approximately 4 and 2 times higher than that of CK. The maximum plant height was found in treatment A, which was about 41.9% and 20.9% higher than that of CK and B, respectively. The cumulative biomass in treatments A and B was increased by 54.2 t·ha-1 and 38.5 t·ha-1 as compared to CK, respectively. At T3, the plant TC in treatment A and B increased by 5.4 and 2.2%, respectively as compared to CK (Figure S2a). At T3, the plant TN in treatment A and B increased by 58.3% and 36.7%, respectively as compared to CK (Figure S2c). The Na content in treatments A and B increased by 76.3% and 33.9%, respectively as compared to CK, while the Cl content increased by 49.9% and 22.2%, respectively (Figure S3).
Linear regression analysis showed that as plant biomass increased; TOC and TC showed an increasing while ROC and TIC showed a significant decreasing trend (Figure S4). Linear regression analysis of plant biomass and plant salt elements indicated that Na and Cl significantly increased with increasing plant biomass (Figure S5). Additionally, Spearman correlation analysis revealed a significant positive correlation between TOC and both Cl and Na contents in plants, with TC showing a similar trend (Figure S6).
3.3. Calculation of Carbon Sequestration
The effect of biofertilizer on carbon sequestration by plant and soil was determined in treatments A, B and CK. The cumulative biomass for A, B, and CK was recorded as 74.3 t·ha-1, 58.5 t·ha-1, and 20.1 t·ha-1, respectively (Figure 3d), whereas plant carbon content was 31.2%, 30.2%, and 29.5%, respectively (Figure S2a). According to formula 1, the plant carbon sequestration for A, B, and CK per hectare was 84.9 t CO2e, 64.9 t CO2e, and 21.7 t CO2e, respectively (Figure 4a). In comparison to CK, the application of biofertilizer in treatments A and B increased plant carbon sequestration per-hectare by 63.2 t CO2e and 43.1 t CO2e, respectively.
The application of biofertilizer significantly increased the soil TOC, by 2.9 times and 2 times compared to CK. Soil TOC reached at its peak at T3 and decreased significantly after crop harvest (Figure S7a). Soil TIC exhibited an overall increasing trend, and was opposite to soil TOC after T2 (Figure S7b). In comparison to CK, soil TC content in treatments A and B was increased by 35.9% and 22.9%, respectively with a gradually increasing tread from T1 to T4 (p < 0.05). Soil TC in treatments A and B was significantly higher than CK throughout the experiment (Figure S7c). TC, TOC, and TIC content in rhizosphere soil was significantly higher than in non-rhizosphere soil in all treatments at T3 (Figure S8). In the experimental area, soil bulk density was 1.5 g·cm-3, and both soil tillage and sampling depths were 35 cm. Based on Formula 2, the soil carbon sequestration per-hectare for treatments A, B, and CK was 498 t CO2e, 450 t CO2e, and 366 t CO2e, respectively. The carbon content of the biofertilizer used in this experiment was 23.2%. Treatments A and B added 50.8 t CO2e and 25.4 t CO2e of carbon to the field, respectively (Figure 4b). Compared to CK, The treatments A and B increased soil carbon sequestration per-hectare by 80.7 t CO2e and 58.6 t CO2e, respectively.
The application of biofertilizer significantly increased total carbon sequestration in treatment A and B about 583 t CO2e and 515 t CO2e, respectively, per hectare in comparison to CK (388 t CO2e) (Figure 4c). In comparison to CK, the application of 60 and 30 t·ha-1 of biofertilizer resulted into an increase in carbon sequestration by 144 and 102 t CO2e per hectare
3.4. Soil Enzymes Activities
Soil enzymes activity was measured in order to determine the effect of biofertilizer. The enzymes activity was significantly increased after application of biofertilizer as shown in Figure 5. The enzyme activity was increased with the increase in concentration of biofertilizer as evident by the higher activity in treatment A as compared to B. Additionally, the activity of all enzymes in rhizosphere soil, except for CAT, was significantly higher than in non-rhizosphere soil (Figure S9).
3.5. Interaction of Environmental Factors and Their Relationship with Soil Carbon Components
The results of linear regression analysis showed that plant biomass increased significantly with the increase of SOM and nutrients, the decrease of soil TSS, and the increase of soil enzymes activities in saline-alkali soil (Figure 6). Pearson correlation analysis showed that SOM, TOC, TC, SWC, AP, TN, and NH4+-N were positively correlated with the soil enzymes activities, while soil enzymes activities were negatively correlated with TSS and C/N (Figure 7).
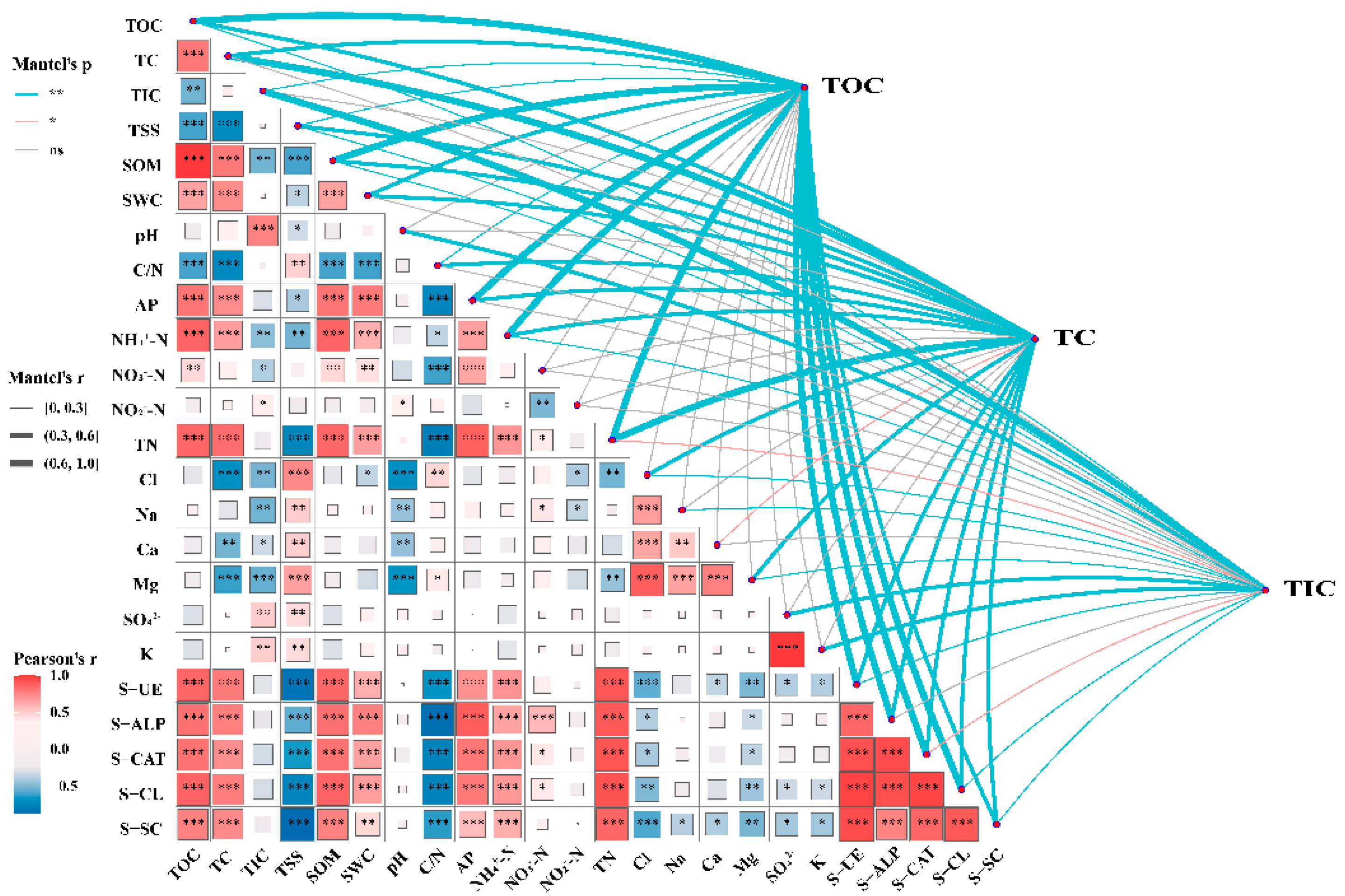
With the increase of plant biomass, TOC content significantly increased, while TIC content showed an opposite trend (Figure 6). Mantel tests of soil carbon components and environmental factors showed that soil factors and enzymes activities significantly impacted soil carbon components (Figure 7). SOM content had the greatest impact on TOC. The environmental factor with the greatest impact on TIC is pH, while TN is the environmental factor that has the greatest impact on TC. From the values of Mantel’s r, it can be observed that environmental factors have the greater impact on TOC compared to TC and TIC. Specifically, eight environmental factors have Mantel’s r values exceeding 0.6, indicating their strong impact on TOC.
4. Discussion
4.1. Improvement of Saline-Alkali Soil by Biofertilizer
In this study, biofertilizer effectively improved the physicochemical properties and nutrient content of saline-alkali soil. During the four-month crop growing period, the application of biofertilizer significantly increased SOM content. The results of the T4 period indicate that although SOM content significantly decreased after crop harvest, the soil amended with biofertilizer still had higher SOM compared to CK. This indicates that biofertilizer can effectively increase SOM over the long term. The increase of SOM is mainly due to the input of organic matter in biofertilizer, as well as increase of organic carbon sequestration. Rhizodeposition and root exudates can input more effective organic matter into soil [30]. Additionally, plant roots contribute more to organic carbon, which is one of the most effective organic carbon inputs in soil [31], which might explain the significant decrease of SOM content in T4 period.
Nitrogen and phosphorus are vital nutrients for plant growth and are integral to soil nutrient cycling processes [32]. The application of biofertilizer significantly increased soil nutrient content during the crop growing period. The increase in soil nutrient content may be due to the increase in nutrient input of biofertilizer and plant roots [33]. The SWC in treatment A during the T2 and T3 periods was significantly higher than in treatments B and CK, but it decreased significantly after crop harvest. Plant growth increases soil water retention and prevents soil erosion [34]. This may be the reason for the decrease in soil SWC after the harvest of S. salsa.
The increase in soil salinity adversely affects soil nutrients, enzymes activities, and crops. Severe soil salt stress can significantly reduce crop yield and the carbon absorbed through photosynthesis [35,36]. TSS in soil are an important criterion for assessing soil salinization level [37]. Previous studies have demonstrated that S. salsa is a typical halophyte, which can absorb salts from the soil and accumulate them within itself [38,39]. The same results were found in this study, where soil TSS content in all three treatments at the T4 stage was significantly lower than at the T1 stage, with the reduction being more significant in the biofertilizer treatment. This demonstrates that S. salsa can reduce soil TSS, and the application of biofertilizer enhances this process. Linear regression analysis of plant biomass and soil TSS content also confirmed this result, showing a significant decrease in soil TSS content with an increase in plant biomass. It is mentioned in section 3.1 that the reduction in soil salinity is primarily reflected in low Cl and Na content. The Na and Cl content in S. salsa in treatments A and B was significantly higher than in CK. These results imply that biofertilizer primarily reduces soil salinity stress by increasing the absorption of Cl and Na by S. salsa.
4.2. Growth Promotion of S. salsa by Biofertilizer
In our study, the application of biofertilizer significantly promoted growth of S. salsa. The plant height, dry weight, fresh weight, and plant biomass increased significantly. Salt stress elevates the osmotic pressure of the soil solution, restricting normal water absorption by roots and causing physiological dehydration in plants, ultimately hindering their growth [40]. Therefore, the reduction in soil salinity may be the primary reason for the increase in plant biomass. Besides, the mineralization of organic nitrogen components plays a crucial role in providing nitrogen for plants [33,41]. Under high nitrogen conditions, plant demand for AP also increases [42]. In this study, the application of biofertilizer increased soil TN and AP content, providing the essential nutrients needed for S. salsa’s growth. The significant positive correlation between plant biomass and TN and AP further supports our hypothesis (Figure 6).
Additionally, soil enzymes can convert nutrients into forms available for plant uptake and protect plants from abiotic stress [43,44]. The increase in soil enzymes activities has a positive effect on plant biomass accumulation. In this study, soil enzymes activities were positively correlated with plant biomass (Figure 6), suggesting that biofertilizer may also promote plant growth by increasing enzymes activities in the soil.
4.3. Enhancement of Soil Enzymes Activities by Biofertilizer
Soil enzymes are essential for the decomposition of organic matter and the cycling of nutrients, and their activities are influenced by various factors [45]. S-UE and S-ALP are important enzymes affecting the nitrogen and phosphorus cycles in the soil, converting organic nitrogen and phosphorus into forms available to plants. Their activities are considered reliable indicators for assessing changes in soil quality [43]. S-CAT is an important enzyme for promoting plant growth and development, breaking down hydrogen peroxide in the soil and protecting plants from oxidative stress [44]. S-SC and S-CL play significant roles in soil carbon transformation and accumulation. S-SC can break down sucrose into fructose and glucose, increasing the soluble nutrients in the soil. S-CL can catalyze the decomposition of cellulose into glucose, providing a usable carbon source for microorganisms [46].
In this study, soil enzymes activities in all treatments were initially increased followed by a gradual decrease. Compared to CK, the application of biofertilizer significantly increased soil enzymes activities, and this increase was biofertilizer concentration dependent, the higher the amount of biofertilizer in soil, the higher the enzymes activities. This may be because S. salsa root exudates provide an energy source for microorganisms, promoting increased soil enzymes activities [43]. The application of biofertilizer significantly increased plant biomass, further enhancing soil enzymes activities. After crop harvesting, the reduction in energy sources led to an overall decrease in soil enzymes activities. Studies have found that the activities of S-SC and S-CL enzymes are usually positively correlated with SOM content, and increase in enzymes activities can promote the decomposition and transformation of SOM [46,47]. In this study, SOM, SWC, AP, TN, and NH4+-N showed a positive correlation with the enzyme activities (Figure 7), which leads to an improvement in saline-alkali soil properties. We observed that the enzymes activities in rhizosphere soils were significantly higher than in non-rhizosphere soils. This may be because soil enzymes primarily originate from plant root exudates and microbial cell secretions in the soil [48]. Additionally, roots are hotspots of microbial activity, which leads to higher enzymes activities in rhizosphere soils [49].
4.4. Interaction of Biotic and Abiotic Factors and Its Impact on Carbon Sequestration
Carbon sequestration in vegetation is of great significance for material cycling and energy flow [50]. However, it has long been an overlooked method for reducing atmospheric carbon [51]. In this study, the application of biofertilizer significantly increased the carbon sequestration of plants. The plant carbon is primarily stored as plant biomass after being converted from CO2 into organic carbon through photosynthesis [52]. Therefore, the increase in plant carbon sequestration is likely due to the increase in plant biomass. Correlation analysis in this study also confirmed that plant biomass is positively associated with plant TC content. Additionally, the improvement in soil properties and the increase in enzymes activities significantly increased the plant biomass, indirectly increasing the plant carbon sequestration [53]. Our study indicates that carbon sequestration in plants plays a significant role in the carbon sequestration process, and its carbon sequestration potential should not be underestimated.
The soil carbon pool is an important component of the Earth's carbon cycle and plays a crucial role in regulating the global climate. In this study, biofertilizer significantly increased the soil TC content, and it increases with the application rate. Soil TC includes both SOC from organisms and SIC present as carbonate minerals [54]. The humus introduced by biofertilizer accumulates in the soil, which can facilitate SOC sequestration [5,55]. From the temporal variation graph of soil TOC content, it was observed that the soil TOC content was highest during the growth period of S. salsa and significantly decreased in T4 period (Figure S7a). This trend is consistent with the changes in SOM, indicating that SOM content may be the primary factor influencing SOC content [56]. Additionally, Mantel test results further confirmed this viewpoint. Previous studies have shown that the availability of soil N and P elements significantly affects SOC sequestration, and the increase of SOC is accompanied by a higher level of N and P [57]. The accumulation of TSS in the soil can disrupt soil structure and decrease soil carbon sequestration capacity [27]. Soil C/N can indicate organic matter decomposition rates and nitrogen mineralization potential [57], and the processes of SOM formation and decomposition are closely associated with soil microorganisms [7]. Previous research has shown that under high C/N conditions, microorganisms may slow down their carbon use efficiency (CUE), which leads to a preferential consumption of dissolved organic matter [58]. In our study, C/N was negatively correlated with soil TC, which may be due to higher CUE under low C/N conditions. A higher CUE indicates greater growth efficiency, with relatively less carbon released to the atmosphere and more carbon stored in the soil [59]. However, microbial CUE was not measured in this study, and its potential impact on soil carbon sinks could be substantial. Future research could further investigate the microbial mechanisms behind changes in carbon. These perspectives suggest that the improvement of saline-alkali soil is conducive to the increase of SOC content.
The reduction in plant biomass may decrease the total amount of rhizodeposition entering the soil, further reducing the input of organic carbon from roots [31,60]. This indicates that an increase in plant biomass may lead to an increase in SOC content. The increase in SOM can enhance SOC accumulation, as both its formation and decomposition are influenced by soil enzymes [58,61]. In our study, soil enzymes activities showed a significant positive correlation with both SOM and SOC. Therefore, we believe that the increase in soil enzymes activities contributes to the rise in SOC.
Historically, research on soil carbon components has primarily focused on SOC, with insufficient attention given to SIC [62]. However, soils in arid and semi-arid regions contain substantial inorganic carbon pools, and even minor changes can significantly impact the global carbon cycle and climate change [63]. Our results show that soil TIC content generally exhibits an increasing trend, with a notable decrease during the T3 period. This decrease may be due to a reduction in soil pH during T3, which accelerated the dissolution of SIC [64]. Previous studies have shown that chemical fertilizer application may lead to soil acidification and significant SIC loss [65]. In this study, the application of biofertilizer had no significant effect on soil pH, and the alkaline conditions in the soil promoted carbonate accumulation [66]. After crop harvest, soil TIC content exhibits an increasing trend, which is opposite to the change in TOC. The type of carbonate contributing to SIC sinks is pedogenic carbonate, which primarily originates from CO2 produced through various pathways [55,67]. Therefore, we hypothesize that this may be due to the CO2 produced from SOC decomposition promoting the formation of SIC [68]. On the other hand, CO2 released from SIC dissolution enhances crop photosynthesis, which in turn promotes SOC formation [69]. This process supports the biochemical cycle between SIC and SOC [1]. The changes in soil TC further support our hypothesis.
The rhizosphere is the most active component in soil and is closely related to the soil carbon cycle [19]. The interactions among soil, microorganisms, and plant roots form a complex relationship that results in differences in carbon content between rhizosphere and non-rhizosphere soils [70]. Our study shows that TC, TOC, and TIC content in rhizosphere soils of all treatments were significantly higher than in non-rhizosphere soils. This may be due to the ability of plant roots to release photosynthetically fixed carbon into the rhizosphere environment, which then enters the rhizosphere soil through microbial activity [71].
4. Conclusions
This study assessed the impact of applying biofertilizer on carbon sequestration of saline-alkali soil ecosystem through field experiments. This study indicated that the application of biofertilizer significantly improves the properties of saline-alkali soil, increases soil enzymes activities, and promotes the growth of S. salsa. Meanwhile, these changes significantly affect the carbon sequestration of both plants and soil. It is noteworthy that the impact of biofertilizer on soil carbon sequestration is primarily through its effect on SOC content, while SIC is mainly influenced by pH and SOC content. Our results indicate that using biofertilizer to improve saline-alkali soil and increase its carbon sequestration is a promising strategy.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Properties of the saline-alkali soil and the biofertilizer; Table S2: Soil physicochemical properties; Figure S1: Soil salt composition in different treatments; Figure S2: Carbon and nitrogen indexes of S. salsa; Figure S3: Changes of salt content in S. salsa; Figure S4: Linear regression analysis of plant biomass and carbon and nitrogen indexes in S. salsa; Figure S5: Linear regression analysis of plant biomass and salt elements in S. salsa; Figure S6: Spearman correlation coefficients between the carbon and nitrogen indexes and salinity in S. salsa; Figure S7: Changes of soil carbon components; Figure S8: Contents of different carbon components in rhizosphere(R) and non-rhizosphere(N-r) soil; Figure S9: Enzyme activities in rhizosphere(R) and non-rhizosphere(N-r) soil.
Author Contributions
Conceptualization, Z.B.; Methodology, Z.B., J.W. and R.L.; Investigation, J.W. and R.L.; Data curation, J.W. and R.L.; Visualization, J.W. and R.L.; Writing - original draft, J.W. and R.L.; Writing - review and editing, S.A., J.W., X.P. and X.Z.; Project administration, Z.B. and X.P.; Funding acquisition, Z.B.; Supervision, Z.B., X.Z. and J.W. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Key Research & Development Program of China (No. 2023YFF1304101), the Research Project of Binzhou Institute of Technology (Nos. GYY-NYHJ-2023-WT-005 and GYY-DTFZ-2022-004) and the Open Project of Key Laboratory of Environmental Biotechnology, CAS (No. kf2020015).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available on request.
Acknowledgments
The authors are thankful to Binzhou Institute of Technology for providing consumables for the experimentation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ma, Y.; Yu, Y.; Nan, S.; Chai, Y.; Xu, W.; Qin, Y.; Li, X.; Bodner, G. Conversion of SIC to SOC Enhances Soil Carbon Sequestration and Soil Structural Stability in Alpine Ecosystems of the Qinghai-Tibet Plateau. Soil Biol. Biochem. 2024, 195, 109452. [Google Scholar] [CrossRef]
- Metternicht, G.I.; Zinck, J.A. Remote Sensing of Soil Salinity: Potentials and Constraints. Remote Sens. Environ. 2003, 85, 1–20. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y. Land Exploitation Resulting in Soil Salinization in a Desert–Oasis Ecotone. Catena 2013, 100, 50–56. [Google Scholar] [CrossRef]
- Zhang, S.; Xue, L.; Liu, J.; Xia, L.; Jia, P.; Feng, Y.; Hao, X.; Zhao, X. Biochar Application Reduced Carbon Footprint of Maize Production in the Saline−alkali Soils. Agric. Ecosyst. Environ. 2024, 368, 109001. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhang, P.; Wu, Y.; Wu, X.; Ni, H.; Lu, Q.; Zang, S. Long-Term Surface Composts Application Enhances Saline-Alkali Soil Carbon Sequestration and Increases Bacterial Community Stability and Complexity. Environ. Res. 2024, 240, 117425. [Google Scholar] [CrossRef]
- Xing, J.; Li, X.; Li, Z.; Wang, X.; Hou, N.; Li, D. Remediation of Soda-Saline-Alkali Soil through Soil Amendments: Microbially Mediated Carbon and Nitrogen Cycles and Remediation Mechanisms. Sci. Total Environ. 2024, 924, 171641. [Google Scholar] [CrossRef]
- Wu, H.; Cui, H.; Fu, C.; Li, R.; Qi, F.; Liu, Z.; Yang, G.; Xiao, K.; Qiao, M. Unveiling the Crucial Role of Soil Microorganisms in Carbon Cycling: A Review. Sci. Total Environ. 2024, 909, 168627. [Google Scholar] [CrossRef]
- Deng, L.; Yuan, H.; Xie, J.; Ge, L.; Chen, Y. Herbaceous Plants Are Better than Woody Plants for Carbon Sequestration. Resour. Conserv. Recycl. 2022, 184, 106431. [Google Scholar] [CrossRef]
- Yang, K.; Zhang, Q.; Zhu, J.; Wang, Q.; Gao, T.; Wang, G.G. Mycorrhizal Type Regulates Trade-Offs between Plant and Soil Carbon in Forests. Nat. Clim. Change 2024, 14, 91–97. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, C.; Li, S.; Bramston, D. A Planting Optimization Strategy to Improve the Carbon Sink Benefit for Urban Green-Taking the Communal Green of Nanjing Forestry University as an Example. Ecol. Indic. 2024, 159, 111619. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, D.; Yuan, J.; Li, Y.; Li, J.; Miao, Y.; Chen, Z.; He, T.; Ding, W. Wetland Restoration after Agricultural Abandonment Enhances Soil Organic Carbon Efficiently by Stimulating Plant- Rather than Microbial-Derived Carbon Accumulation in Northeast China. Catena 2024, 241, 108077. [Google Scholar] [CrossRef]
- Song, J.; Zhang, H.; Zamanian, K.; Chang, F.; Yu, R.; Wang, J.; Zhou, J.; Li, Y. Inorganic Carbon Accumulation in Saline Soils via Modification Effects of Organic Amendments on Dissolved Ions and Enzymes Activities. Catena 2024, 241, 108039. [Google Scholar] [CrossRef]
- Li, S.; Zhao, L.; Wang, C.; Huang, H.; Zhuang, M. Synergistic Improvement of Carbon Sequestration and Crop Yield by Organic Material Addition in Saline Soil: A Global Meta-Analysis. Sci. Total Environ. 2023, 891, 164530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Su, X.; Meng, H.; Wang, H.; Yan, X.; Qin, D.; Liu, C.; Men, Y.; Zhang, X.; Song, X.; et al. Long-Term Cotton Stubble Return and Subsoiling Improve Soil Organic Carbon by Changing the Stability and Organic Carbon of Soil Aggregates in Coastal Saline Fields. Soil Tillage Res. 2024, 241, 106127. [Google Scholar] [CrossRef]
- Jana, G.A.; Glick, B.R.; Yaish, M.W. Salt Tolerance in Plants: Using OMICS to Assess the Impact of Plant Growth-Promoting Bacteria (PGPB). In Mitigation of Plant Abiotic Stress by Microorganisms; Elsevier, 2022; pp. 299–320 ISBN 978-0-323-90568-8.
- Cai, J.-F.; Fan Jiang; Liu, X. -S.; Sun, K.; Wang, W.; Zhang, M.-X.; Li, H.-L.; Xu, H.-F.; Kong, W.-J.; Yu, F.-H. Biochar-Amended Coastal Wetland Soil Enhances Growth of Suaeda Salsa and Alters Rhizosphere Soil Nutrients and Microbial Communities. Sci. Total Environ. 2021, 788, 147707. [Google Scholar] [CrossRef]
- Liu, T.; Wang, S.; Chen, Y.; Luo, J.; Hao, B.; Zhang, Z.; Yang, B.; Guo, W. Bio-Organic Fertilizer Promoted Phytoremediation Using Native Plant Leymus Chinensis in Heavy Metal(Loid)s Contaminated Saline Soil. Environ. Pollut. 2023, 327, 121599. [Google Scholar] [CrossRef]
- Li, M.; Li, X.; Shi, Y.; Jiang, Y.; Xue, R.; Zhang, Q. Soil Enzyme Activity Mediated Organic Carbon Mineralization Due to Soil Erosion in Long Gentle Sloping Farmland in the Black Soil Region. Sci. Total Environ. 2024, 929, 172417. [Google Scholar] [CrossRef]
- Qu, Y.; Tang, J.; Liu, B.; Lyu, H.; Duan, Y.; Yang, Y.; Wang, S.; Li, Z. Rhizosphere Enzyme Activities and Microorganisms Drive the Transformation of Organic and Inorganic Carbon in Saline–Alkali Soil Region. Sci. Rep. 2022, 12, 1314. [Google Scholar] [CrossRef]
- Sun, B.; Gu, L.; Bao, L.; Zhang, S.; Wei, Y.; Bai, Z.; Zhuang, G.; Zhuang, X. Application of Biofertilizer Containing Bacillus Subtilis Reduced the Nitrogen Loss in Agricultural Soil. Soil Biol. Biochem. 2020, 148, 107911. [Google Scholar] [CrossRef]
- Sun, B.; Bai, Z.; Bao, L.; Xue, L.; Zhang, S.; Wei, Y.; Zhang, Z.; Zhuang, G.; Zhuang, X. Bacillus Subtilis Biofertilizer Mitigating Agricultural Ammonia Emission and Shifting Soil Nitrogen Cycling Microbiomes. Environ. Int. 2020, 144, 105989. [Google Scholar] [CrossRef]
- Wu, S.; Zhuang, G.; Bai, Z.; Cen, Y.; Xu, S.; Sun, H.; Han, X.; Zhuang, X. Mitigation of Nitrous Oxide Emissions from Acidic Soils by Bacillus Amyloliquefaciens, a Plant Growth-promoting Bacterium. Glob. Change Biol. 2018, 24, 2352–2365. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, J.; Zhang, Y.; Gong, H.; Hou, R.; Sun, Z.; Ouyang, Z. Soil Microbes from Saline–Alkali Farmland Can Form Carbonate Precipitates. Agronomy 2023, 13, 372. [Google Scholar] [CrossRef]
- Li, R.; Sun, B.; Song, M.; Yan, G.; Hu, Q.; Bai, Z.; Wang, J.; Zhuang, X. Improvement of Saline Soil Properties and Brassica Rapa L. Growth Using Biofertilizers. Sustainability 2024, 16, 2196. [Google Scholar] [CrossRef]
- Li, Z.; Tian, C.; Zhang, R.; Mohamed, I.; Liu, Y.; Zhang, G.; Pan, J.; Chen, F. Plastic Mulching with Drip Irrigation Increases Soil Carbon Stocks of Natrargid Soils in Arid Areas of Northwestern China. Catena 2015, 133, 179–185. [Google Scholar] [CrossRef]
- Luo, K.; Wang, H.; Ma, C.; Wu, C.; Zheng, X.; Xie, L. Carbon Sinks and Carbon Emissions Balance of Land Use Transition in Xinjiang, China: Differences and Compensation. Sci. Rep. 2022, 12, 22456. [Google Scholar] [CrossRef]
- Yang, S.; Hao, X.; Xu, Y.; Yang, J.; Su, D. Meta-Analysis of the Effect of Saline-Alkali Land Improvement and Utilization on Soil Organic Carbon. 2022.
- Li, C.Y.; He, R.; Tian, C.Y.; Song, J. Utilization of Halophytes in Saline Agriculture and Restoration of Contaminated Salinized Soils from Genes to Ecosystem: Suaeda Salsa as an Example. Mar. Pollut. Bull. 2023, 197, 115728. [Google Scholar] [CrossRef]
- Huang, H.; LinkET: Everything Is Linkable. R Package Version 0.0.3 2021. Available online: https://github.com/Hy4m/linkET (accessed on 4 June 2024).
- Kuzyakov, Y. Priming Effects: Interactions between Living and Dead Organic Matter. Soil Biol. Biochem. 2010, 42, 1363–1371. [Google Scholar] [CrossRef]
- Peter, P.C. Soil Organic Matter/Carbon Dynamics in Contrasting Tillage and Land Management Systems: A Case for Smallholder Farmers with Degraded and Marginal Soils. Commun. Soil Sci. Plant Anal. 2017, 48, 2013–2031. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, C.; Guo, J.; Zhang, L.; Xuan, J.; Chen, A.; You, C. Short-Term Phosphorus Addition Augments the Effects of Nitrogen Addition on Soil Respiration in a Typical Steppe. Sci. Total Environ. 2021, 761, 143211. [Google Scholar] [CrossRef]
- Sharma, T.; Arya, V.M.; Sharma, V.; Sharma, S.; Popescu, S.M.; Thakur, N.; M. Iqbal, J.; El-Sheikh, M.A.; Baath, G.S. Impact of Cropping Intensity on Soil Nitrogen and Phosphorus for Sustainable Agricultural Management. J. King Saud Univ. - Sci. 2024, 36, 103244. [Google Scholar] [CrossRef]
- Hu, L.; Zhou, Q.; Peng, D.; Shi, C.; Sun, Q.; Hu, Y.; Li, Y. Effects of Vegetation Restoration on the Temporal Variability of Soil Moisture in the Humid Karst Region of Southwest China. J. Hydrol.: Reg. Stud. 2024, 53, 101852. [Google Scholar] [CrossRef]
- Zhang, J.; Cai, J.; Xu, D.; Wu, B.; Chang, H.; Zhang, B.; Wei, Z. Soil Salinization Poses Greater Effects than Soil Moisture on Field Crop Growth and Yield in Arid Farming Areas with Intense Irrigation. J. Clean. Prod. 2024, 451, 142007. [Google Scholar] [CrossRef]
- Zhang, S.; Rasool, G.; Wang, S.; Zhang, Y.; Guo, X.; Wei, Z.; Zhang, X.; Yang, X.; Wang, T. Biochar and Chlorella Increase Rice Yield by Improving Saline-Alkali Soil Physicochemical Properties and Regulating Bacteria under Aquaculture Wastewater Irrigation. Chemosphere 2023, 340, 139850. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Feng, Q.; Zhou, Y.; Mao, X.; Chen, Y.; Xu, H. Dynamic Changes in Water and Salinity in Saline-Alkali Soils after Simulated Irrigation and Leaching. PLOS ONE 2017, 12, e0187536. [Google Scholar] [CrossRef]
- Liu, S.; Wang, N.; Li, D.; Tian, C.; Zhang, K.; Hu, M.; Niu, H.; Zhao, Z. Effects of Halophyte Suaeda Salsa Continuous Cropping on Physical and Chemical Properties of Saline Soil under Drip Irrigation in Arid Regions. Agric. Ecosyst. Environ. 2024, 371, 109076. [Google Scholar] [CrossRef]
- Xiong, Y.; Wang, S.; Cui, C.; Wu, X.; Zhu, J. Suaeda Salsa NRT1.1 Is Involved in the Regulation of Tolerance to Salt Stress in Transgenic Arabidopsis. Int. J. Mol. Sci. 2023, 24, 12761. [Google Scholar] [CrossRef]
- He, K.; He, G.; Wang, C.; Zhang, H.; Xu, Y.; Wang, S.; Kong, Y.; Zhou, G.; Hu, R. Biochar Amendment Ameliorates Soil Properties and Promotes Miscanthus Growth in a Coastal Saline-Alkali Soil. Appl. Soil Ecol. 2020, 155, 103674. [Google Scholar] [CrossRef]
- Allen, C.R.; Burr, M.D.; Camper, A.K.; Moss, J.J.; Stein, O.R. Seasonality, C:N Ratio and Plant Species Influence on Denitrification and Plant Nitrogen Uptake in Treatment Wetlands. Ecological Engineering 2023, 191, 106946. [Google Scholar] [CrossRef]
- Han, B.; He, Y.; Chen, J.; Wang, Y.; Shi, L.; Lin, Z.; Yu, L.; Wei, X.; Zhang, W.; Geng, Y.; et al. Different Microbial Functional Traits Drive Bulk and Rhizosphere Soil Phosphorus Mobilization in an Alpine Meadow after Nitrogen Input. Sci. Total Environ. 2024, 931, 172904. [Google Scholar] [CrossRef]
- Adetunji, A.T.; Ncube, B.; Meyer, A.H.; Olatunji, O.S.; Mulidzi, R.; Lewu, F.B. Soil pH, Nitrogen, Phosphatase and Urease Activities in Response to Cover Crop Species, Termination Stage and Termination Method. Heliyon 2021, 7, e05980. [Google Scholar] [CrossRef]
- Mollavali, M.; Bolandnazar, S.A.; Schwarz, D.; Rohn, S.; Riehle, P.; Zaare Nahandi, F. Flavonol Glucoside and Antioxidant Enzyme Biosynthesis Affected by Mycorrhizal Fungi in Various Cultivars of Onion ( Allium Cepa L.). J. Agric. Food Chem. 2016, 64, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-M.; Li, Q.; Liang, W.-J.; Jiang, Y. Distribution of Soil Enzyme Activities and Microbial Biomass Along a Latitudinal Gradient in Farmlands of Songliao Plain, Northeast China. Pedosphere 2008, 18, 431–440. [Google Scholar] [CrossRef]
- Miao, S.; Wu, X.; Guo, Y.; Zhao, H.; Qiao, Y. Different Responses of Enzyme Activities to 6-year Warming after Transplant of the 12 Types of Soils. J. Plant Nutr. Soil Sci. 2023, 186, 487–494. [Google Scholar] [CrossRef]
- Feng, Q.; Wang, B.; Chen, M.; Zhang, J.; Zhang, X.; Wu, P. Calcium Alginate−biochar Composite Promotes Nutrient Retention, Enzyme Activity, and Plant Growth in Lime Soil. Environ. Technol. Innov. 2024, 35, 103670. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Feng, Y. The Effects of Biochar Addition on Soil Physicochemical Properties: A Review. Catena 2021, 202, 105284. [Google Scholar] [CrossRef]
- Wen, L.; Peng, Y.; Zhou, Y.; Cai, G.; Lin, Y.; Li, B. Effects of Conservation Tillage on Soil Enzyme Activities of Global Cultivated Land: A Meta-Analysis. Journal of Environmental Management 2023, 345, 118904. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, C.; Chen, S.; Zhang, Y.; Yu, T.; Xue, X.; Wu, L.; Zhou, W.; Yun, X.; Yan, R.; et al. Moderate Grazing Increased Carbon, Nitrogen and Phosphorus Storage in Plants and Soil in the Eurasian Meadow Steppe Ecosystem. Sci. Total Environ. 2024, 914, 169864. [Google Scholar] [CrossRef]
- Yu, C.; Mao, J.-H.; Huang, X.; Zhang, Y. Carbon Capture by Phytomass Storage and Trading to Mitigate Climate Change and Preserve Natural Resources. Environ. Sustain. Indic. 2024, 22, 100358. [Google Scholar] [CrossRef]
- Wang, Y.; Chang, Q.; Li, X. Promoting Sustainable Carbon Sequestration of Plants in Urban Greenspace by Planting Design: A Case Study in Parks of Beijing. Urban For. Urban Green. 2021, 64, 127291. [Google Scholar] [CrossRef]
- Scholz, F.; Hasse, U. Permanent Wood Sequestration: The Solution to the Global Carbon Dioxide Problem. ChemSusChem 2008, 1, 381–384. [Google Scholar] [CrossRef]
- Lin, Z.; Price, G.W.; Liang, C.; Burton, D.L.; Lynch, D.H. Effects on Soil Carbon Storage from Municipal Biosolids Application to Agricultural Fields. Journal of Environmental Management 2024, 361, 121249. [Google Scholar] [CrossRef] [PubMed]
- Bughio, M.A.; Wang, P.; Meng, F.; Qing, C.; Kuzyakov, Y.; Wang, X.; Junejo, S.A. Neoformation of Pedogenic Carbonates by Irrigation and Fertilization and Their Contribution to Carbon Sequestration in Soil. Geoderma 2016, 262, 12–19. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Liu, C.; Zhu, E.; Jia, J.; Feng, X. Shifting Relationships between SOC and Molecular Diversity in Soils of Varied Carbon Concentrations: Evidence from Drained Wetlands. Geoderma 2023, 433, 116459. [Google Scholar] [CrossRef]
- Gong, H.; Li, Y.; Li, S. Effects of the Interaction between Biochar and Nutrients on Soil Organic Carbon Sequestration in Soda Saline-Alkali Grassland: A Review. Glob. Ecol. Conserv. 2021, 26, e01449. [Google Scholar] [CrossRef]
- Huang, W.; Wu, J.; Pan, X.; Tan, X.; Zeng, Y.; Shi, Q.; Liu, T.; Zeng, Y. Effects of Long-Term Straw Return on Soil Organic Carbon Fractions and Enzyme Activities in a Double-Cropped Rice Paddy in South China. J. Integr. Agric. 2021, 20, 236–247. [Google Scholar] [CrossRef]
- Manzoni, S.; Taylor, P.; Richter, A.; Porporato, A.; Ågren, G.I. Environmental and Stoichiometric Controls on Microbial Carbon-use Efficiency in Soils. New Phytol. 2012, 196, 79–91. [Google Scholar] [CrossRef]
- Preece, C.; Peñuelas, J. Rhizodeposition under Drought and Consequences for Soil Communities and Ecosystem Resilience. Plant Soil 2016, 409, 1–17. [Google Scholar] [CrossRef]
- Korkanç, S.Y. Effects of Afforestation on Soil Organic Carbon and Other Soil Properties. Catena 2014, 123, 62–69. [Google Scholar] [CrossRef]
- Raza, S.; Irshad, A.; Margenot, A.; Zamanian, K.; Li, N.; Ullah, S.; Mehmood, K.; Ajmal Khan, M.; Siddique, N.; Zhou, J.; et al. Inorganic Carbon Is Overlooked in Global Soil Carbon Research: A Bibliometric Analysis. Geoderma 2024, 443, 116831. [Google Scholar] [CrossRef]
- Liang, J.; Zhao, Y.; Chen, L.; Liu, J. Soil Inorganic Carbon Storage and Spatial Distribution in Irrigated Farmland on the North China Plain. Geoderma 2024, 445, 116887. [Google Scholar] [CrossRef]
- Raza, S.; Zamanian, K.; Ullah, S.; Kuzyakov, Y.; Virto, I.; Zhou, J. Inorganic Carbon Losses by Soil Acidification Jeopardize Global Efforts on Carbon Sequestration and Climate Change Mitigation. J. Clean. Prod. 2021, 315, 128036. [Google Scholar] [CrossRef]
- Chen, L.; Zhou, G.; Feng, B.; Wang, C.; Luo, Y.; Li, F.; Shen, C.; Ma, D.; Zhang, C.; Zhang, J. Saline–Alkali Land Reclamation Boosts Topsoil Carbon Storage by Preferentially Accumulating Plant-Derived Carbon. Sci. Bull. 2024, S2095-9273(24)00357-8, doi:10.1016/j.scib.2024.03.063. [CrossRef]
- Xu, T.; Zhang, M.; Ding, S.; Liu, B.; Chang, Q.; Zhao, X.; Wang, Y.; Wang, J.; Wang, L. Grassland Degradation with Saline-Alkaline Reduces More Soil Inorganic Carbon than Soil Organic Carbon Storage. Ecol. Indic. 2021, 131, 108194. [Google Scholar] [CrossRef]
- Huber, D.P.; Lohse, K.A.; Commendador, A.; Joy, S.; Aho, K.; Finney, B.; Germino, M.J. Vegetation and Precipitation Shifts Interact to Alter Organic and Inorganic Carbon Storage in Cold Desert Soils. Ecosphere 2019, 10, e02655. [Google Scholar] [CrossRef]
- Bai, T.; Qiu, Y.; Hu, S. Nitrogen Availability Mediates the Effects of Roots and Mycorrhizal Fungi on Soil Organic Carbon Decomposition: A Meta-Analysis. Pedosphere 2024, 34, 289–296. [Google Scholar] [CrossRef]
- Lin, Z.-B.; Zhang, R.-D. Dynamics of Soil Organic Carbon Under Uncertain Climate Change and Elevated Atmospheric CO2. Pedosphere 2012, 22, 489–496. [Google Scholar] [CrossRef]
- Shao, G.; Xu, Y.; Zhou, J.; Tian, P.; Ai, J.; Yang, Y.; Zamanian, K.; Zeng, Z.; Zang, H. Enhanced Soil Organic Carbon Stability in Rhizosphere through Manure Application. Soil Tillage Res. 2024, 244, 106223. [Google Scholar] [CrossRef]
- Su, X.; Zhang, L.; Meng, H.; Wang, H.; Zhao, J.; Sun, X.; Song, X.; Zhang, X.; Mao, L. Long-Term Conservation Tillage Increase Cotton Rhizosphere Sequestration of Soil Organic Carbon by Changing Specific Microbial CO2 Fixation Pathways in Coastal Saline Soil. J. Environ. Manag. 2024, 358, 120743. [Google Scholar] [CrossRef]
Figure 1.
Photo of experimental field and plot distribution map of different treatments.

Figure 2.
Changes of soil salt content. K (a) Na(b), Mg(c), Cl(d), Ca(e), SO42-(f). CK (with no biofertilizer); A (60 t·ha-1 biofertilizer); B (30 t·ha-1 biofertilizer). Different letters indicate that there is significant difference (P < 0.05) between different biofertilizers applications in the same period. T1, T2, T3, and T4 represent May 2021, July 2021, September 2021 and May 2022, respectively.
Figure 2.
Changes of soil salt content. K (a) Na(b), Mg(c), Cl(d), Ca(e), SO42-(f). CK (with no biofertilizer); A (60 t·ha-1 biofertilizer); B (30 t·ha-1 biofertilizer). Different letters indicate that there is significant difference (P < 0.05) between different biofertilizers applications in the same period. T1, T2, T3, and T4 represent May 2021, July 2021, September 2021 and May 2022, respectively.
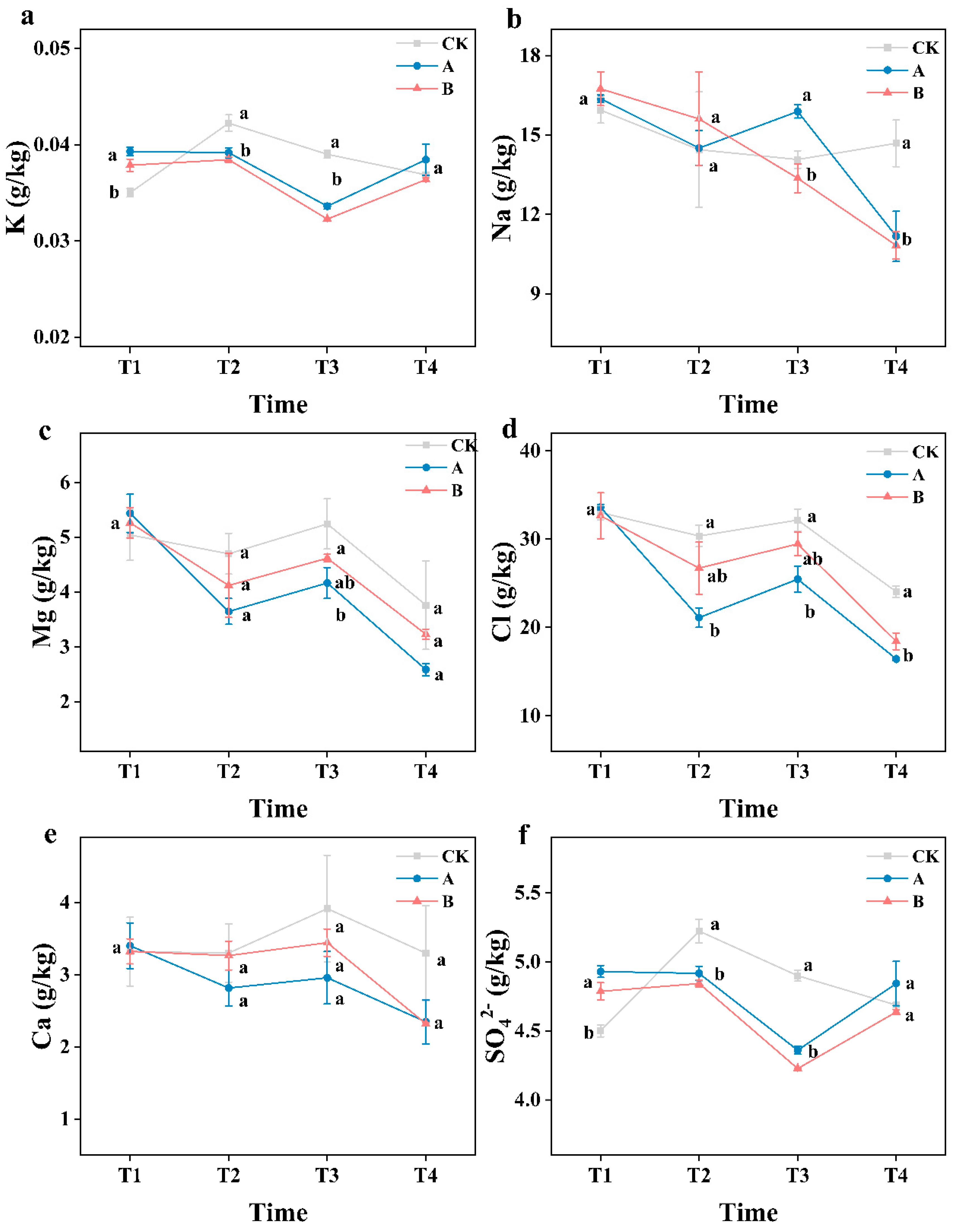
Figure 3.
Growth index of S. salsa in different periods. plant height (a), dry weight (b), fresh weight (c), plant biomass (d). The values in (a), (b) and (c) are the average values of plant samples, and the values in (d) are the sum of all plant biomass. CK (with no biofertilizer), A (60 t·ha-1 biofertilizer), B (30 t·ha-1 biofertilizer). Different letters indicate that there are significant differences (p < 0.05) between different biofertilizers applications in the same period. T2 and T3 represent July 2021 and September 2021, respectively.
Figure 3.
Growth index of S. salsa in different periods. plant height (a), dry weight (b), fresh weight (c), plant biomass (d). The values in (a), (b) and (c) are the average values of plant samples, and the values in (d) are the sum of all plant biomass. CK (with no biofertilizer), A (60 t·ha-1 biofertilizer), B (30 t·ha-1 biofertilizer). Different letters indicate that there are significant differences (p < 0.05) between different biofertilizers applications in the same period. T2 and T3 represent July 2021 and September 2021, respectively.
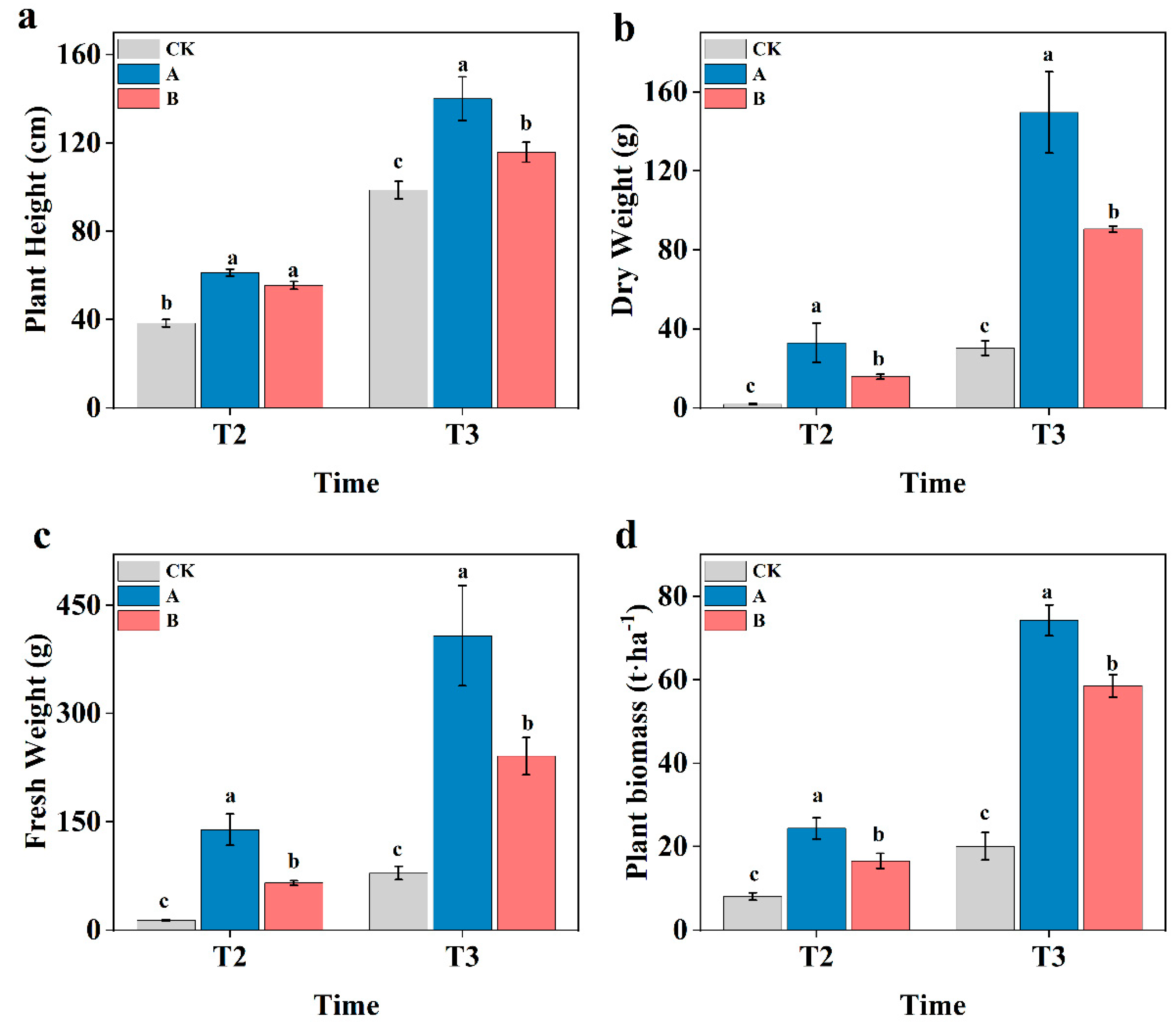
Figure 4.
The per-hectare carbon sequestration. plant carbon sequestration (a), soil carbon sequestration (b), total carbon sequestration (c). CK (with no biofertilizer), A (60 t·ha-1 biofertilizer), B (30 t·ha-1 biofertilizer). Different letters indicate that there are significant differences (p < 0.05) between different biofertilizers applications.
Figure 4.
The per-hectare carbon sequestration. plant carbon sequestration (a), soil carbon sequestration (b), total carbon sequestration (c). CK (with no biofertilizer), A (60 t·ha-1 biofertilizer), B (30 t·ha-1 biofertilizer). Different letters indicate that there are significant differences (p < 0.05) between different biofertilizers applications.
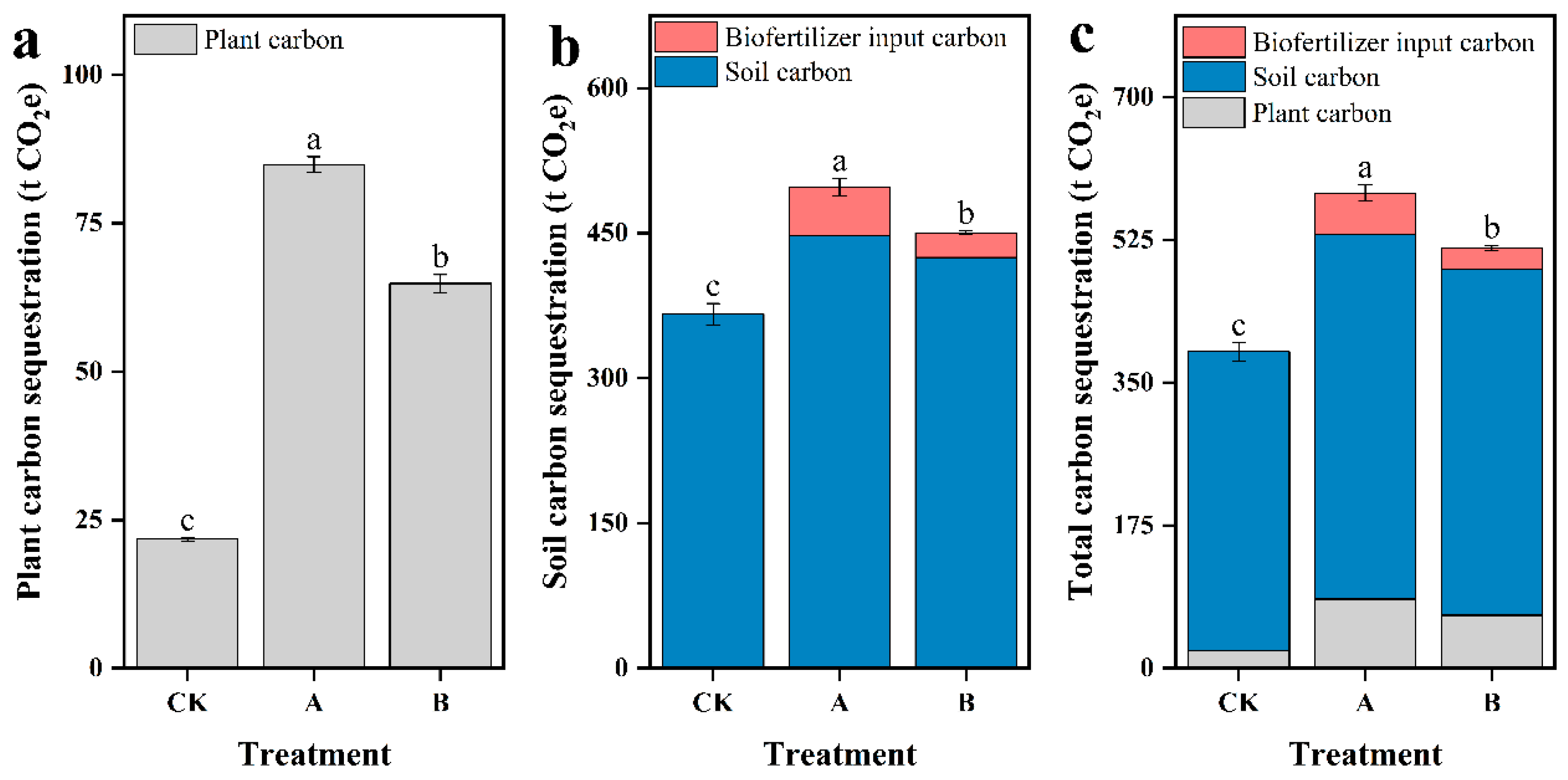
Figure 5.
Changes of soil enzymes activities. S-UE (a), S-CL (b), S-SC (c), S-ALP (d), S-CAT (e). CK (with no biofertilizer), A (60 t·ha-1 biofertilizer), B (30 t·ha-1 biofertilizer). Different letters indicate that there are significant differences (p < 0.05) between different biofertilizers applications in the same period. T1, T2, T3, and T4 represent May 2021, July 2021, September 2021 and May 2022, respectively.
Figure 5.
Changes of soil enzymes activities. S-UE (a), S-CL (b), S-SC (c), S-ALP (d), S-CAT (e). CK (with no biofertilizer), A (60 t·ha-1 biofertilizer), B (30 t·ha-1 biofertilizer). Different letters indicate that there are significant differences (p < 0.05) between different biofertilizers applications in the same period. T1, T2, T3, and T4 represent May 2021, July 2021, September 2021 and May 2022, respectively.
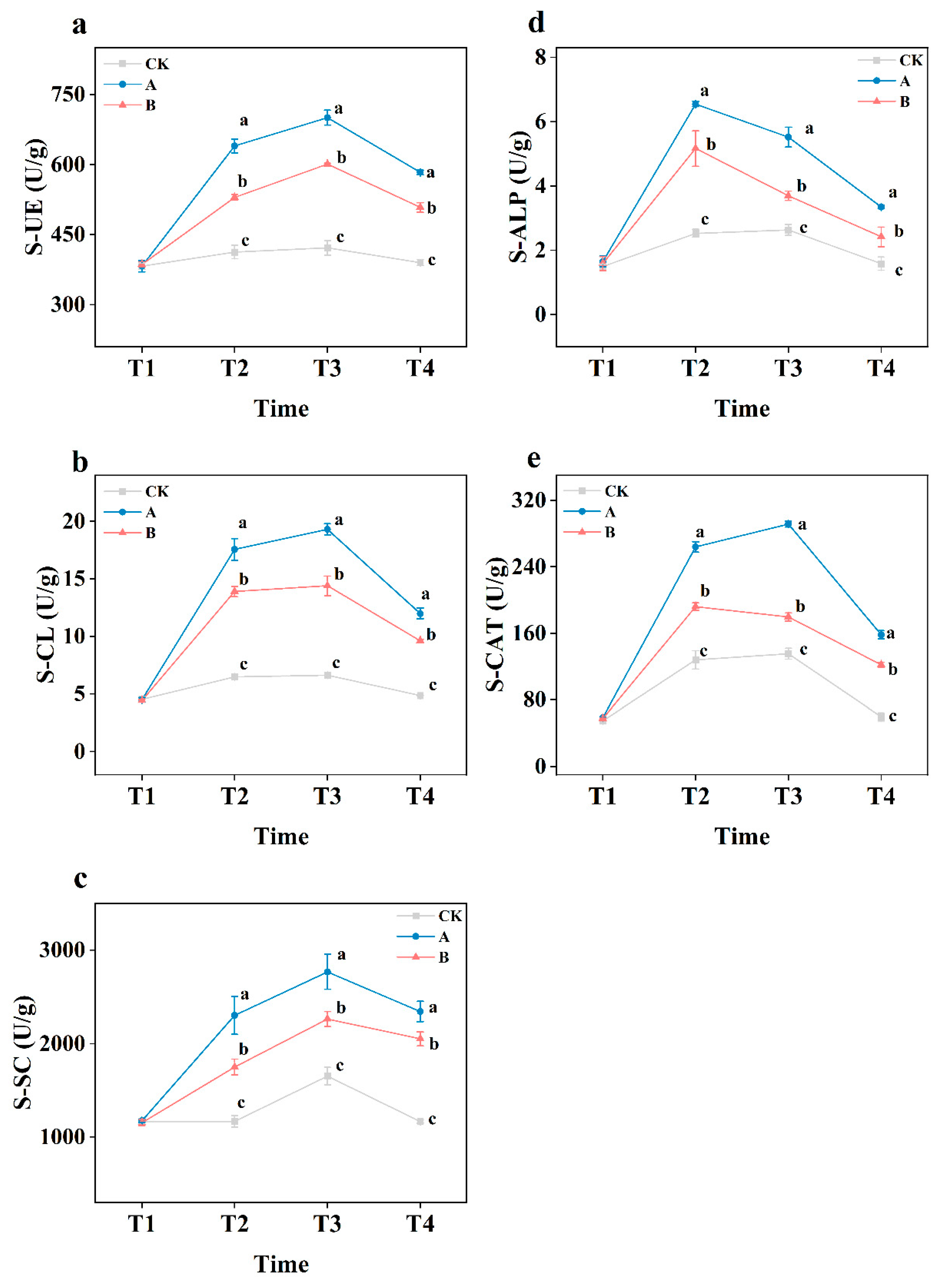
Figure 6.
Linear regression analysis of plant biomass with soil properties and enzymes activities. Different balls represent different samples.
Figure 6.
Linear regression analysis of plant biomass with soil properties and enzymes activities. Different balls represent different samples.
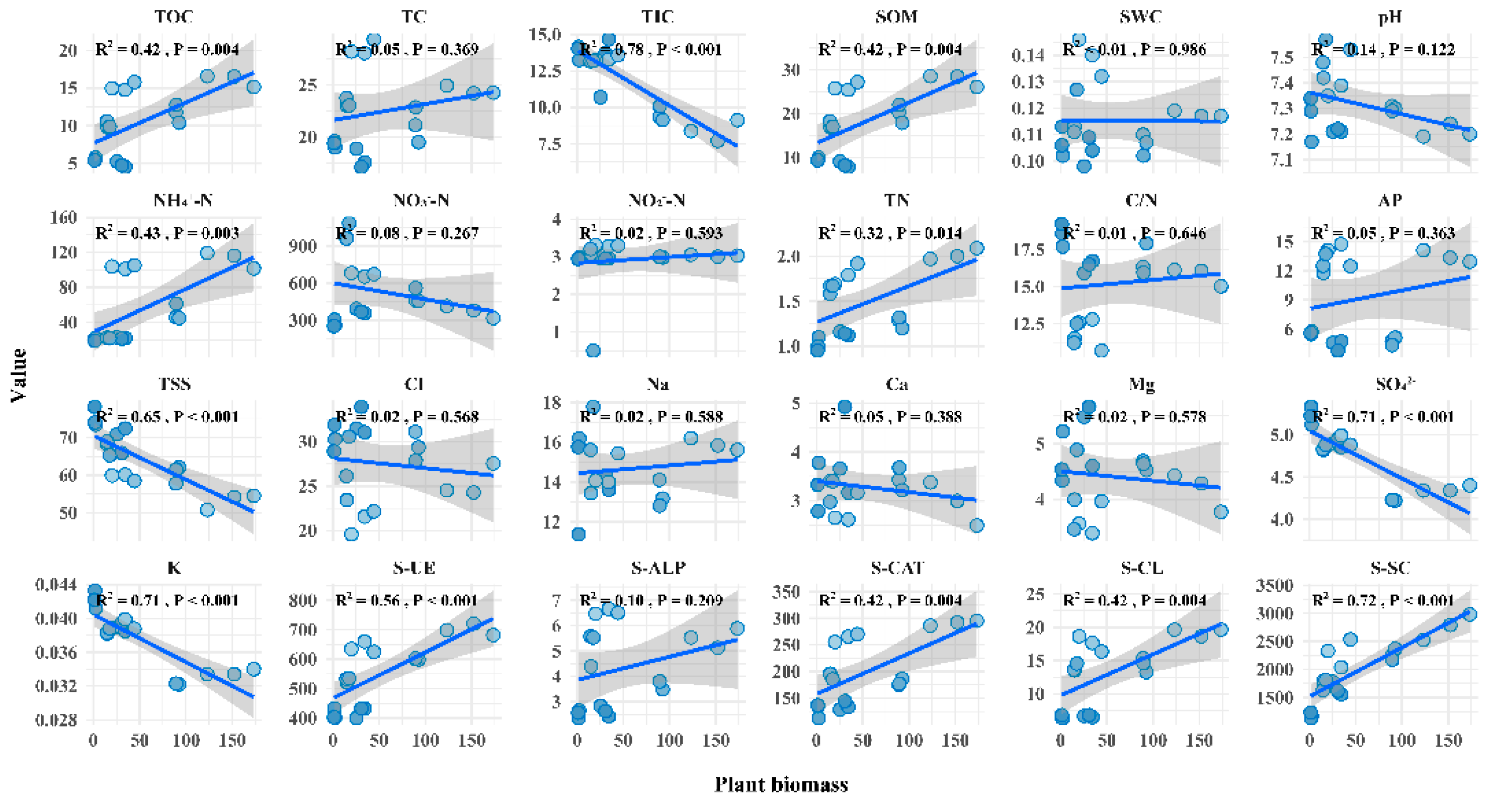
Figure 7.
Mantel test of soil carbon components and environmental factors. The color and thickness of lines represent the significance (Mantel′s p) and correlation coefficient (Mantel′s r), respectively. The color gradient and the size of the square represent Pearson correlation coefficient (Pearson′s r). The level of significance is denoted by * (p <0.05), ** (p <0.01), and *** (p < 0.001).
Figure 7.
Mantel test of soil carbon components and environmental factors. The color and thickness of lines represent the significance (Mantel′s p) and correlation coefficient (Mantel′s r), respectively. The color gradient and the size of the square represent Pearson correlation coefficient (Pearson′s r). The level of significance is denoted by * (p <0.05), ** (p <0.01), and *** (p < 0.001).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



