Submitted:
23 April 2025
Posted:
23 April 2025
You are already at the latest version
Abstract
The ontological categorization of the cellular elements of the brain was proposed over a century ago by Santiago Ramón y Cajal (neurons, astroglia) and Pío del Río-Hortega (oligodendroglia, microglia). It combines histochemical observations of morphology with allied inferences about the specialized functions and origins (ectoderm or mesoderm) of each cellular element. This ontology shapes modern neuroscience, with the main non-neuronal cells – astroglia, oligodendroglia and microglia – viewed as having distinct primary roles relating to metabolic support, myelination and protection of neurons, the information signaling cells. Yet contemporary techniques, ranging from electrophysiology to single-cell transcriptomics and ultrahigh resolution spectroscopy, are revealing intersecting molecular profiles and functional capacities of these cell groups, for example metabolic support, neuroimmune and signaling functions in oligodendroglia. Here we identify discrepancies in current glial paradigms from empirical, evolutionary and pragmatic perspectives. We suggest that a subset of cells often viewed as oligodendroglia instead have iron-related primary functions that are central to all aspects of brain activity, rather than myelin-related primary functions. We discuss implications for pathogenesis across the spectrum of neuropsychiatric and neurological disorders, including neurodegenerative conditions such as Alzheimer’s disease and other less common cognitive, movement and neurobehavioral disorders, stroke and cerebrovascular disease, glioblastoma and other brain cancers and neuroimmune conditions. We also briefly address the question of where these cells reside within existing glial compartments and lineages, implications for the ontological classification of other glial cells and research limitations that must be overcome going forward.
Keywords:
iron
; glia
; oligodendrocyte
; myelin
; evolution
; mental health
; Alzheimer’s disease
; stroke
; cancer
; DNAzyme sensors
“I am a firm believer that, without speculation there is no good & original observation.” Charles Darwin, letter to Alfred Wallace [1,2].
1. Introduction
The accepted categorization of the main cellular elements of the central nervous system (CNS) as neurons, macroglia (astrocytes, oligodendrocytes) and microglia arose from histochemical studies over a century ago [3,4,5,6,7,8,9,10,11,12,13]. The primary function of each element was inferred from circumstantial structural and contextual evidence gleaned from histological observations. Information exchange by neurons was deduced from their interconnected dendritic networks, metabolic support by astrocytes in locations bridging neurons and vessels, immune responses by microglia around pathological lesions and myelination by oligodendrocytes wrapping their processes around axons.
We are entering a new era of understanding as modern biotechnologies, including novel chimerization models, single molecule arrays and multiomics profiling of individual CNS cells [14], provide new feature sets for increasingly precise ontological classification. In the past decade, this has led to proposals for new subgroupings within each of the astroglial (e.g., [15,16,17]), microglial (e.g., [18,19,20]) and oligodendroglial (e.g., [21,22]) classes.
As many as nine or more different subtypes of astroglia may exist, distinguished by spatial localization, molecular profiles and responses to inflammatory stimuli that may differ between species, sexes, brain regions and health or disease states [15]. Similarly, there may also be multiple phenotypically distinct microglial subtypes with distinct properties and functions that respond differently to stimuli, including so-called rod microglia and ‘dark microglia’ [18,19,20,23,24]. These new findings entail fundamental departures from past views of microglia and astroglia, with some microglia subgroups proposed to have different inherent phenotypes and functionalities, rather than being functionally different facets of a single inherent highly plastic phenotype that alters in response to environmental factors, as once believed [18,19,20]. Yet a crucial distinction needs to be made between subtype classification based on innate features that reflect inherent subtypes, as opposed to plasticity reflecting reactive states [19,25,26]. It is important to determine whether different astroglial or microglial subgroups observed in different studies are indeed innately different subtypes, rather than different reactive states.
Separation between different cell types of essential inherent primary functions that may otherwise come into conflict is likely to have evolutionary advantages. For example, having one group of cells with the primary function of migratory defensive responses to life-threatening pathogens and another group with the primary function of delivering essential nutrients to neurons helps prevent cells becoming trapped in a stasis of mutually exclusive, conflicting responses. Conversely, there are likely evolutionary benefits in redundancy of vital functions across different glial types. The primary, definitive, functional feature of microglia has been their neuroimmune responses, including phagocytosis, yet CNS defense is now also considered an important function of astroglia, supplementing their recognized roles in CNS metabolic homeostasis, with many immune response functions shared with microglia, including phagocytosis, and with overlapping targets and molecular profiles [11,27,28]. These redundant functions may override primary astroglial functions in pathological or other unusual circumstances where microglia responses are inadequate. Similarly, oligodendroglia (OLG) can exhibit metabolic or neuroimmune functions [29,30,31,32,33].
It is unknown if function-based classification alone can distinguish glial species effectively without also considering other features lacking clear involvement in vital activities e.g., morphology or structural markers such as glial fibrillary acidic protein (GFAP). Increasingly precise subclassifications, functional or otherwise, are of limited use, and may even be invalid, if the ontology itself is not an accurate representation of the biological entities being classified. For example, consider two cell groups arising from a common ancestor during evolution that have since diverged and perform very different, possibly even mutually exclusive, functions but retain some shared phenotypic features. Classifications focusing on shared phenotypic features, blind to divergent primary functions, will not usefully distinguish subgroups. We believe there are important blind spots of this kind in the present glial ontology.
Here we challenge the accuracy of the existing ontology and suggest some cells in recognized glial categories have essential primary functions beyond the triad of immune response, general metabolic support and production of myelin, the electrical insulating material ensheathing some CNS axons. This is exemplified by the OLG, even though this lineage might perhaps appear to have the clearest grounds for function-based classification of the three main glial types. Oligodendrocytes (myelination-competent cells) and their committed progenitors have near unique, non-redundant, direct or indirect functions in myelin production that do not appear to be readily subsumed by cells considered to be microglia or astroglia, except in artificially engineered scenarios [34,35].
Yet despite this apparently unequivocal function, we propose some cells often viewed as OLG instead have essential primary functions relating to maintenance of brain iron homeostasis, that are independent of myelin-related functions. These cells are able to load, store and transport relatively large amounts of iron and often exhibit strong labeling for iron by ,3’ diaminobenzidine-enhanced Perls’ stain (DAB+Perls’) for ferric (Fe3+) iron [36]. As it is not currently clear where these cells reside in existing glial compartments and lineages or instead have separate compartments, lineage/s or both, we call these cells ferriglia (Fe+glia), to distinguish them from other glia (notably astrocytes, microglia) that are not primarily specialized for iron handling and do not usually accumulate comparable amounts of iron in healthy brain but may acquire iron in pathological circumstances.
We hypothesize Fe+glia provide a safe and efficient ‘iron supply chain’ that confers powerful survival advantages, protecting against toxicity or iron-scavenging pathogens. From evolutionary and pragmatic perspectives, essential primary roles in iron homeostasis such as transferring iron between blood vessels, neurons, astrocytes and other glia, and myelin, are unlikely to be compatible with competing primary roles in myelination that tether cells to myelin for long periods. We believe these considerations necessitate reassessment of the validity and utility of classical OLG ontology, and deeper contemplation of how this might require rethinking the ontological classification of CNS cells in general.
2. History
The non-neuronal component of the nervous system was first described by 18th century European scientists and named neuroglia or simply ‘glia’, from the Greek word for glue, by Rudolph Virchow in 1856 [23]. This reflected the prevailing belief that the ‘glia’ consisted of a connective structure binding together the neural tissue [10,23,37], though other scientists such as Carl Bergmann contemporaneously published descriptions of non-neuronal cellular entities, reviewed [38]. We use ‘neural’ to encompass nervous system components in general, including glia, and ‘neuronal’ specifically for neurons.
2.1. Cajal’s Classification of Neurons and Astroglia
The first systematic ontology of CNS cells, and forerunner of the current ontology, is credited to Santiago Ramón y Cajal [4,5,37,39], and included the concept of neuroglial cells. Cajal proposed three anatomically and functionally distinct groups of CNS cellular entities that he termed elements [6,7,12,39]. He hypothesized that information-related nervous system functions are performed by cells we now call neurons, which he categorized as the first cellular element of the CNS. In the early 20th century, the non-neuronal ‘glue’ component of the CNS was still considered by some prominent early histologists, including Camillo Golgi, to be an extended, interconnected, reticular network or syncytium [40]. Cajal instead proposed star-like cellular entities (astrocytes) comprised a second ‘neuroglial’ element providing nutritional and metabolic support to neurons [7,41,42].
2.2. Hortega’s Classification of Microglia and Oligodendroglia
Cajal incorrectly proposed a third CNS element of small, apolar cells without detectable processes by Golgi staining [6]. Pío del Río Hortega developed an improved silver carbonate stain [43] revealing fine ‘elongations’ (appendages or processes) and proposed the present classification of glia based on phenotypic features, postulated embryonic source (ectoderm or mesoderm), location and inferences about function [33,44].
Some cells described by Hortega, which he named microglia, for their small size, were commonly seen in gray matter [45,46]. He proposed microglia were the true ‘third element’, originating from the mesoderm outside the CNS, unlike the first two elements, both of ectodermal neuroepithelial origin, neurons and macroglia, then comprising protoplasmic and fibrous astroglia [33,45,46,47]. Hortega correctly deduced migratory, surveillance and phagocytic functions of microglia from histochemical observations across varied physiological and pathological contexts [9,12]. His classification of microglia as the third element was thus multifaceted, based on inferences about their non-neuroglial nature (originating outside the CNS), as well as their morphology (small size, amoeboid or ramified shape), functions and greater density in gray compared to white matter.
Together, these features distinguished microglia from oligodendroglia (OLG), the other new cell group defined by Hortega [8,9,10,12,33,44,45]. Hortega’s descriptions of OLG initially focused on apolar ‘interfascicular’ cells grouped in bundles or columns between nerve fibers in white matter, then expanded to a wider range of morphologies and locations, including gray matter (Section 3; [8,9,10,12,33,44]). He later named these cells ‘oligodendroglia’, from Greek terms signifying scarce or few (‘oligo’) and processes (‘dendro’). Hortega proposed oligodendrocytes were the myelinating cells of the brain, describing OLG with fine processes spiraling around myelinated nerve fibers [8]. Hortega’s findings were validated by Wilder Penfield in 2024 [48]. Examples of OLG and other glial morphologies are shown in Figure 1.
Hortega categorized OLG as macroglia i.e., ‘large glia’, the second element that already contained the astroglia, thereby establishing the accepted ontological classification of the three major recognized glial species that prevails a century later [6,8,9,10,11,12,13,14,23,33,38,44,45,46]. The second element also came to include other glia such as ependymal cells, recognized as being cellular in nature in the late 1800s [49,50], before Cajal’s classification.
Figure 1.
Promiscuous relationships of iron-rich glia that may masquerade as OLG. While glia sometimes exhibit distinctive morphologies, inferring species or functions from location and morphology alone is unreliable, as exemplified by small iron-rich glial cells (‘Fe+glia’) usually having few processes and eccentric nuclei, features often ascribed to OLG, that may be an unrecognized cell group. Some Fe+glia engage in proliferative activities in iron-rich neighborhoods, including myelin-ensheathed fiber tracts, or nestle beside vessels, neurons or other cells [51]. Fe+glia may participate in iron-transfer mechanisms via iron-laden extensions, possibly involving transferrin, clathrin-mediated endocytosis or both or kiss and run’ or other exchanges of iron-containing vesicles [52,53,54,55].
Figure 1.
Promiscuous relationships of iron-rich glia that may masquerade as OLG. While glia sometimes exhibit distinctive morphologies, inferring species or functions from location and morphology alone is unreliable, as exemplified by small iron-rich glial cells (‘Fe+glia’) usually having few processes and eccentric nuclei, features often ascribed to OLG, that may be an unrecognized cell group. Some Fe+glia engage in proliferative activities in iron-rich neighborhoods, including myelin-ensheathed fiber tracts, or nestle beside vessels, neurons or other cells [51]. Fe+glia may participate in iron-transfer mechanisms via iron-laden extensions, possibly involving transferrin, clathrin-mediated endocytosis or both or kiss and run’ or other exchanges of iron-containing vesicles [52,53,54,55].
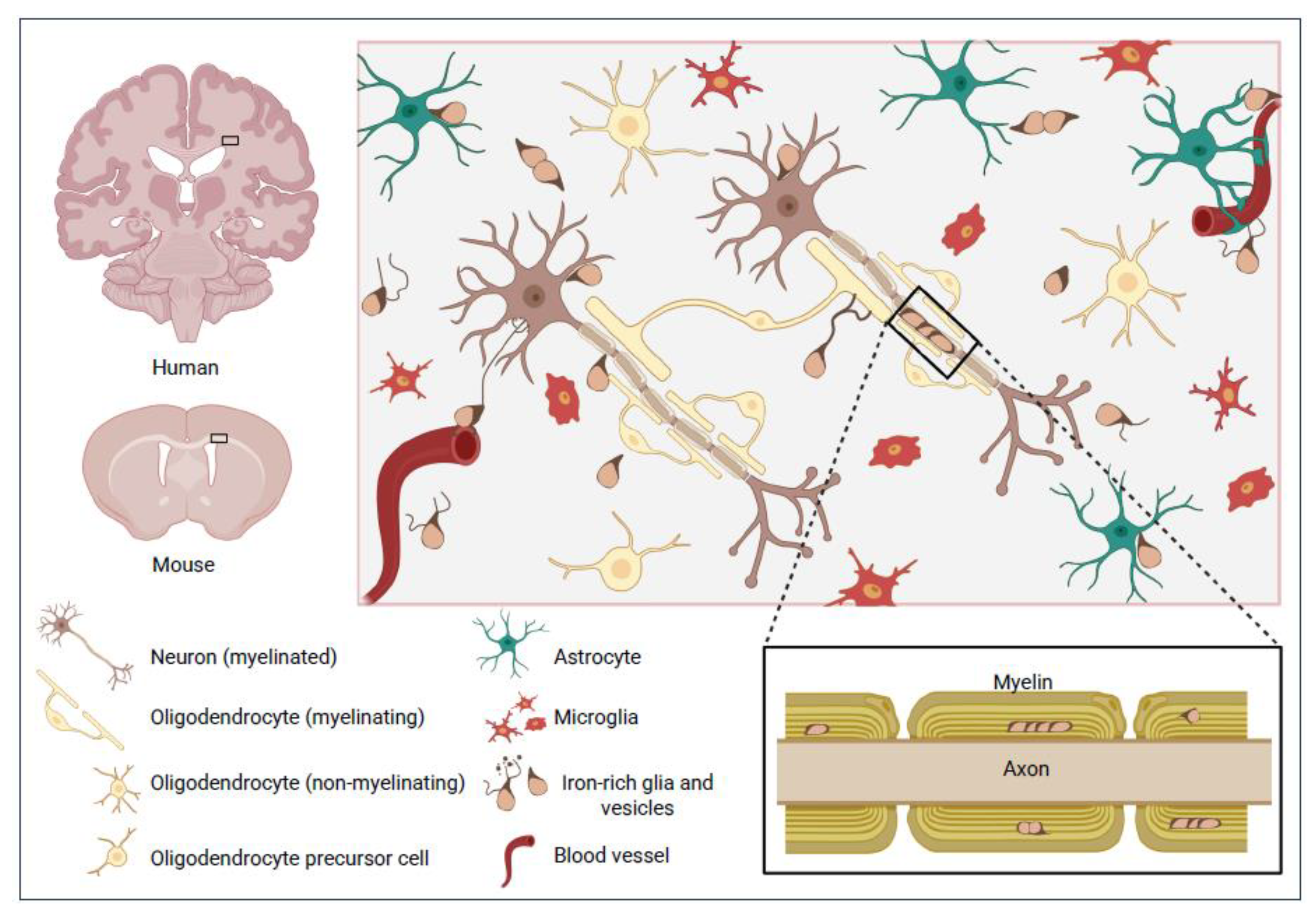
3. Paradoxical Properties of Cells Currently Classified as OLG
The cellular ontology of CNS neurons and glia developed by Cajal, Hortega and other pioneering histopathologists was an extraordinary achievement, given the complexity of the brain, the paucity of foundational knowledge and the technical constraints on the restricted set of features able to be interrogated at the time. Yet even in Hortega’s original descriptions, the cell group classified as OLG was recognized to have paradoxical properties not inevitably consistent with primary functions in myelination [8,33,44]. Hortega studied OLG in diverse regions of brain or spinal cord from several vertebrate species, including monkey and newborn or adult human, under physiological or pathological conditions. Over time, as his observations expanded across different physiological and pathological contexts, his perceptions of what constituted and defined OLG as separate from other elements also shifted.
3.1. Invalid Presumptions Arising from Inferring Myelination or OLG Identity from Cell Size
Many early histologists, including Hortega, already recognized the problems of categorizing glial types by morphology alone. Morphology is plastic (e.g., Figure 1 and Figure 2) and changes with many factors, including cell cycle phase and interactions with other cells, activation state, aging, nutrients, toxins and other environmental variables. Yet it is still widely used to identify OLG, either alone or with location, unsupported by cell marker labeling.
3.1.1. Size
One perplexing discrepancy in descriptions of OLG by Hortega and later researchers involves their size. While usually grouped with astroglia as macroglia, rather than ‘microglia’, some OLG are depicted as considerably smaller than astroglia by Hortega (reproduced [12]) and others [56]. Hortega himself noted many cells he considered to be OLG were similar in size to microglia and easily confused without staining of cell processes… ‘as the microglial elements, and many of the oligodendroglia are small, and all of them with prolongations, confusion is easy when dealing with rounded nucleus cells and incompletely stained protoplasmic appendages and without their real features’ (translated Iglesias-Rozas and Garrosa [57]). In this passage, Hortega considered OLG to be distinguished by their close relationship with nerve fibers yet he elsewhere described cells he considered to be OLG in interstitial or juxta-vascular locations without clear relationships with nerve fibers. (The prefix juxta- signifies ‘near or adjacent to’ and peri- signifies ‘surrounding or around’ e.g., juxta-vascular oligodendrocyte; peri-axonal myelin sheath.)
3.1.2. Other Morphological Features – Eccentric Nucleus, Shape and Fine, Sparse Processes
In Hortega’s diagrams, some OLG have a central nucleus and processes spiraling round multiple axons, yet other OLG have an eccentric nucleus and, consistent with their name, few (‘oligo’) processes (‘dendro’) [8,12,33,44], illustrated in Figure 1. Glia with similar oval or ‘raindrop’ shaped morphologies, with eccentric nuclei and only a few processes, are still often assumed to be OLG, even without marker studies or other evidence.
Later research widely assumes myelination is the primary function of cells with this distinctive morphology, often inferring OLG identity and myelinating capabilities from morphology alone, without marker validation. Yet this group of glia is recognized to sometimes exhibit paradoxical properties not clearly consistent with primary myelinating functions, including locations not near myelin [8,33,44,58,59] and mitotic figures [60].
3.2. Invalid Presumptions Arising from Inferring Myelination or OLG Identity from Location
Although Hortega distinguished OLG as being more common in white matter than microglia, he also described abundant OLG in gray matter. Besides four myelinating oligodendrocyte subtypes [33,44], he also described cells with OLG-like morphologies in apparent non-myelinating contexts in juxta-vascular, juxta-neuronal and interstitial locations in white and gray matter. Though electron microscopy shows some juxta-neuronal OLG myelinate neuronal cell bodies [61], other OLG-like cells appear to be neither myelination-competent oligodendrocytes nor committed progenitors of oligodendrocytes.
3.2.1. Nature and Roles of Myelin at Juxta-Neuronal or Other Non-Axonal Sites
Even when OLG are engaged in peri-neuronal myelination at non-axonal sites, this ‘myelin’ may differ in wrapping structure or other salient features from peri-axonal myelin, which serves to accelerate electrical signaling via saltatory conduction, in addition to providing metabolic support (e.g., nutrient delivery, waste clearance) and protecting nerve fibers from chemical or physical damage. Myelin-like substances at other sites may likewise provide physical protection and metabolic support to the neuronal cell body yet may differ in the nature of these functions or effects on neurotransmission. For example, nutrients required by the soma may differ from those needed along long axons. This is relevant when considering the relative importance and competing requirements of myelination versus other vital functions in which OLG or OLG-like cells might be engaged, such as transporting iron or other essential nutrients, or waste, between blood vessels and myelin.
3.2.2. Assumptions About the Nature and Roles of OLG-like Cells at Juxta-Myelin Sites
Conversely, even OLG near myelin may not have primary myelin-related functions, as noted by Wood and Bunge: ‘demonstration of the connections between oligodendrocyte somas and myelin sheaths is inherently very difficult, however, and it is possible that there are substantial numbers of cells in white matter, resident among the myelinating oligodendrocytes, that do not directly husband myelin segments’ [62].
We note that even processes clearly extending to myelin sheaths may not be myelinating. Fine processes from OLG-like cells to myelin across interstitial expanses several times the length of the soma are depicted by Hortega [8,9,10,12,33,44,47] and others [56] in classical diagrams. These appear to be an inefficient and vulnerable strategy for long-term myelination of axons, but a feasible, readily relocatable, ‘pop-up’ pipeline for transfer of iron or other materials between myelin and blood vessels, neurons or other cells. Figure 2 shows iron-rich cells (Fe+glia) with OLG-like morphologies extending iron-rich processes across interstitial expanses (2A-C) or into vessels (F-H) or undergoing mitosis (D,E).
3.3. Non-Myelin Related Functions of Subsets of Cells Usually Assumed to Be OLG
It is often assumed non-myelinating cells with OLG-type morphologies described by Hortega and others [8,9] are quiescent oligodendrocytes that can myelinate in appropriate circumstances [62,63], or oligodendrocytes with other functions supplementing primary functions related to myelination [62,63,64,65,66]. This is largely unexplored, perhaps because such functions are usually deemed subsidiary to the primary function of myelination. Below we consider features of OLG and related cells in juxta-neuronal, juxta-vascular or interstitial locations, focusing on criteria used to identify these cells as OLG and proposed non-myelin related functions.
3.3.1. Juxta-Neuronal OLG
The literature includes reports of both non-myelinating and myelinating juxta-neuronal OLG, possibly reflecting different subgroups of these cells that manifest differently in different contexts. Juxta-neuronal OLG immunoreactive for 2’,3’-cyclic nucleotide-3’phosphodiesterase (CNPase) are found throughout cerebral gray matter, at greater density in deep cortical compared to superficial layers [66,67]. This marker labels oligodendrocyte precursor cells (OPC) and committed oligodendrocyte precursors (COP), as well as mature oligodendrocytes, and may be a marker to commitment to eventual myelin formation [68].
While CNPase labeling does not necessarily imply myelination, electrophysiology, electron microscopy and other approaches show some juxta-neuronal oligodendrocytes in layer 5 of mouse neocortex myelinate surrounding axons [61]. This study also showed these oligodendrocytes can integrate with astrocytes in a glial syncytium that buffers K+ and constrains high-frequency neuronal action potential firing. This revitalizes early concepts of glial reticular networks championed by Golgi and others [40] (Section 2.1, 3.3.2), equivalent to ‘functional syncytia’ and suggests descriptions of astroglial networks (e.g., [69,70]) should incorporate details of interconnections with OLG.
In contrast, another study examining cell markers and gene/protein expression profiles concluded some juxta-neuronal OLG are non-myelinating cells with a unique phenotype, expressing oligodendrocyte transcription factor 2 (OLIG2) but also other transcription factors and platelet-derived growth factor (PDGF) receptor-αβ and its ligand PDGF-CC [71]. Whether myelinating or not, juxta-neuronal oligodendrocytes may provide metabolic and functional support to neurons. For example, glutamine synthetase produced by some juxta-neuronal OLG is needed for normal glutamatergic neuronal transmission of nearby neurons, independent of their myelinating functions [72,73]. Changes in juxta-neuronal oligodendrocyte numbers, distribution or both are also observed in a number of neurological disorders [67]. It is unclear if there are distinct subsets of juxta-neuronal OLG with different inherent properties or if observed differences in health or disease are instead plastic responses to environmental factors, analogous to the debate on microglia mentioned above [74]. Research is needed on whether juxta-neuronal OLG or OLG-like cells supply iron or other metals to neurons and clinical implications.
3.3.2. Juxta-Vascular OLG.
Relationships of OLG with brain vasculature are unclear. Some OPCs may use vessels as a scaffold for migration, detaching when differentiating into mature oligodendrocytes [75], but mature oligodendrocytes may also associate with vessels. Hortega detailed the complex expansions of plexuses formed by OLG (reproduced [8,9,10,12,33,44,47]), including apparent non-myelin juxta-vascular plexuses of ‘crisscrossing’ OLG processes. Hortega acknowledged these could be interpreted as neuroglial reticular systems, similar to extended single entities with fused cytoplasm proposed by early pathologists such as Virchow and Golgi [40], early versions of modern ‘functional syncytia’. However his observations suggested the cytoplasm of individual OLG does not anastomose with that of other OLG, astrocytes or microglia in a single, interconnected structure.
This area is rarely studied, but comprehensive examination of OLG in brains from young adult mice (age 6-12 weeks) estimated about one in six CNPase-immunoreactive OLG are in close proximity (<1 μm) to the brain vasculature, in particular capillaries. Plexus formation was not mentioned, but while the nature of functional interactions between OLG and vessels remains unclear, there was evidence of direct contact with the vascular basement membrane, suggesting exchange of information (signaling networks) or metabolites with endothelial cells [75].
Relationships between juxta-vascular OLG and iron regulation are poorly understood but research by the Connor and Moos groups, Morris and others has shown juxta-vascular cells with morphologies similar to OLG contain radiochemically and histochemically detectable iron and iron-related proteins in human brain, consistent with roles in iron transport and uptake from endothelial cells into the brain [58,76,77], although these functions remain to be confirmed. Figure 2 shows complex nets of fine, iron-rich Fe+glial processes near vessels and in interstitial spaces, that appear more consistent with primary roles in iron transfer than in myelination.
3.3.3. Interstitial OLG
Cells with OLG-like morphologies have been observed scattered throughout the brain in humans, primate, rodents and other mammals [78], singly or in doublets (including mitotic figures) or small clusters, without apparent contact with myelin, neurons or vessels. The functions of these cells are unknown but may include immune responses with secretion or uptake of cytokines and chemokines. The observation that primary OPCs exhibit phagocytic capabilities, and when exposed to IFNγ can cross-present antigens to cytotoxic CD8 T cells in vitro, also demonstrates immune-like functions of OLG cells [79,80]. Such findings suggest that cells currently considered to be OLG can adopt an altered phenotype resembling that of immune cells under both normal and disease conditions. Again, it remains to be seen whether such OLG have different inherent properties or if observed differences in health or disease are instead plastic changes, as discussed above.
3.3.4. Progenitor Cells of the OLG Lineage
Observations of mitotic figures in some cells with OLG-like morphology in juxta-axonal and other locations are not consistent with these cells being oligodendrocytes, which are post-mitotic. Such cells are instead often assumed to be OPC. As mitotic figures are often seen in Fe+glia with OLG-like morphologies, these cells could also be OPC. However OLG precursor/progenitor cells labeled by markers such as nerve/glial antigen 2 (NG2; chondroitin sulfate proteoglycan 4) or PDGFR-α [81,82,83] typically display very different morphologies and are considerably larger than Fe+glia and, to our knowledge, there has not been clear demonstration of ferric iron in cells co-labeled for these markers.
3.3.5. Olfactory Ensheathing Glia
Olfactory information is sent from peripheral sensory neurons in olfactory epithelium to terminal synapses with neurons in the olfactory glomeruli. The olfactory nerve has non-myelinated axons, surrounded by a special type of glia, olfactory ensheathing cells (OEC), with features of both astrocytes and Schwann cells. These cells support neural transmission and olfactory sensory neuronal regeneration and participate in innate immune responses [84,85,86]. OEC can myelinate in some experimental settings and express CNPase [68]. Based on MRI and ferritin levels, the OECs develop high levels of iron at the glia limitans in the olfactory bulb by age 12 weeks in wildtype mice [87] and may constitute a form of Fe+glia. Olfactory lobes arose early in brain evolution. The existence of early iron-rich ensheathing cells that do not myelinate under normal conditions is consistent with other functions of both these cells and myelin (Section 5.3.1, Section 5.4, Section 5.5.).
The olfactory bulb also contains oligodendrocytes or other OLG [86,88], thought to be generated locally from progenitors that may express proteolipid protein (PLP) but not platelet derived growth factor receptor (PDGFR) [89]. These OLG interact with OEC and may myelinate dendritic segments and neuron bodies surrounding olfactory glomeruli [86,88]. There is little information on their iron content and relationships to OEC, if any.
3.4. Limitations of Past Research
When exploring non-myelin related OLG features to identify novel functions, as above, it is important to distinguish changes in cell species or type (i.e., the inherent identity of the cell) from plastic changes in cell state, sometimes termed reactivity, which can have functional or clinical implications [18,19,90]. For example, functions involving neuroimmune factors such as cytokines or chemokines might fluctuate reversibly in OLG in responses to stressors while the broader phenotype defining cell type is unchanged.
Another limitation is that researchers usually seek non-canonical OLG functions by first selecting for OLGs using markers specific for oligodendrocytes or OPCs that detect cells specialized for myelin production or precursor species that can generate myelin-producing cells, then investigating other functions of selected cells. Examples of commonly used markers include OPC markers such as NG2 or PDGFR-α, myelin-related markers such as CNPase, neurite outgrowth inhibitor (NogoA), anti-adenomatous polyposis coli clone (CC1) or pan-OLG markers such as OLIG2.
While restricting analyses to cells committed to myelin-related functions is appropriate for investigating additional functions of these cells, this overlooks the possibility that some cells assumed from morphology or location to be OLG do not have specialized myelin-related capabilities and instead may have other, potentially even more important, functions, such as iron regulation. We earlier reported most iron-rich glia with ‘OLG-like’ morphology (in terms of small size, eccentric nuclei and few processes) do not show myelin markers such as myelin basic protein or PLP in mice age 12 weeks [51]. This appeared independent of location (juxta-myelin, juxta-neuronal, juxta-vascular or interstitial).
‘Ferriglia’ are common and widely distributed in healthy and pathogenic mouse CNS, and are present in cortex and deep gray matter as well as white matter. Yet they are unlikely to have been detected in many past studies using markers such as those above, specific to cells committed to eventual myelin formation, specifically OPC, COPs or oligodendrocytes. In healthy mouse [65] or human brain [59,60,91,92], their small size may cause them to be overlooked and their oval or ‘raindrop’ shape, eccentric nuclei and few fine processes of variable length/branching (Figure 2) can be mistaken for OLG or other glia. Ferriglial iron levels in brains with healthy iron balance can also be near or below sensitivity thresholds of histochemical stains, especially in leached post-mortem tissue.
We propose these cells are not members of known oligodendroglial or other glial compartments. It is possible that OLG and Fe+glia lineages arose by divergence from a common early progenitor cell or that self-renewing mitotic Fe+glia reside upstream of known OLG lineage compartments generate OLG progenitors in asymmetric divisions in response to myelination-related signals. Alternatively, there may be one or more separate ferriglial lineages not including OLG or other glia. More research into relationships between Fe+glia and OLGs is required, including whether or not there are common evolutionary or developmental ancestors. More research is needed on Fe+glia and their origins.
4. Rationales for Separation of Iron Regulation from, and Precedence over, Myelination
Some, though not all, of the issues discussed above are recognized in attempts to define the nature of OLG by a core gene regulatory network, e.g., [93], although as above, approaches focusing on regulatory genes identified in studies of cells with myelin-related function or their precursors will not capture essential core regulatory features of cells with non-myelin related primary functions in iron regulation or other activities.
The high levels of iron in iron-rich Fe+glia assumed to be OLG and myelin-containing structures are usually attributed teleologically to high requirements of myelination for iron (Section 4.2). As above, most Fe+glia in healthy brain are usually considered to be oligodendrocytes or other OLG or sometime astrocytes or microglia, yet we earlier showed that in mice most Fe+glia do not express the pan-OLG marker OLIG2 and, conversely, most OLIG2-expressing cells have little detectable iron even in mice with high brain iron [51]. The morphology of many Fe+glia, notably their small size, is also not consistent with those of oligodendrocytes or known oligodendrocyte precursors (Figure 2).
Below we consider in more depth evidence for the possibility that at least some Fe+glia have primary roles in iron regulation rather than myelination.
4.1. Evolutionary Advantages of Separating Iron Regulation and Myelination
Early studies by Connor, Morris and others suggested some OLG might have functions in brain iron regulation and supply to other cells [59,62,63,64,65,92] but this has not been developed in the literature. It is widely and often implicitly assumed that most iron travels extracellularly through the brain bound to molecular carriers such as transferrin, ferritin or citrate, analogous to the blood. Instead, we propose some iron-rich cells with OLG morphology (Fe+glia) have primary iron-regulatory functions that include distribution of iron to other CNS cells within their cell processes, and not primary myelin-related functions. Consequently, whether these cells should be classified as OLG needs to be reconsidered.
Some juxta-myelin glia with high iron, ferritin or transferrin content might indeed be OLG, with primary roles in myelination or generation of myelinating cells, and subsidiary roles in iron transfer directly to the axon or soma of myelinated neurons by unknown mechanisms, perhaps via invagination of OLG membranes into the peri-axonal space [94]. However this appears unlikely to be the only cellular regulation of iron in the brain.
From a pragmatic perspective, there are likely to be strong evolutionary advantages of separating primary iron regulation and primary myelination functions. Having one cell type responsible for both may threaten survival in situations where these functions directly conflict. For example, myelinating cells could not easily relocate to sites of hemolytic bleeding or pathogen invasion or even simply to obtain iron from nearby vessels or provide iron to cells in regions without nearby vessels or myelin when required.
4.2. Rethinking Relationships of Iron, Myelin and Iron-Rich, OLG-like Ferriglia
High levels of iron in some cells with OLG-like morphologies, assumed to be oligodendrocytes or OPCs, has been proposed to reflect high iron requirements for lipid and myelin formation, e.g., [92,95] but there may also be other explanations [96]. While intense myelin formation during development or remyelination likely involves high iron usage, it is unclear why myelinating oligodendrocytes in healthy adult brain, with slow myelin turnover [97], would require more iron than neurons, which usually have higher mitochondrial and metabolic activity than myelinating oligodendrocytes in healthy adult brain [98], yet lower iron levels than other brain cells [51,99].
We have suggested hydrophobic, lipid-rich, myelinated or myelin-like structures may provide safe reservoirs for brain iron storage [51]. Ferriglia may deposit surplus iron in myelin for later retrieval and delivery to neurons and other brain cells not specialized for iron storage and vulnerable to iron toxicity. We suggest myelinated or myelin-like structures and Fe+glia may have important primary functions in iron sequestration which confer strong survival advantages that precede in evolution, and are independent of, advantages from saltatory conduction of electrical signals.
This is not addressed effectively in the literature, which considers myelin-related functions the primary definitive feature of cells classed as OLG. Many studies are intentionally designed, or unconsciously biased, to investigate only cells committed to myelination or generating myelination-competent progeny [30,64,66,72,100]. This circularity precludes testing the concepts proposed here, i.e., that some cells classified as OLGs by morphology or location are primarily specialized for other functions such as iron regulation, not myelin-related functions, and that such cells may not reside in OLG lineage compartments. Similarly, both histochemical and single cell transcriptomic studies may use markers for OLG such as the iron transporter protein transferrin, that may also detect Fe+glia, causing Fe+glia to be assigned OLG identities. Numbers of easily visualized iron-rich Fe+glia are low in healthy brain, in the absence of iron dyshomeostasis, which may also cause these cells to be overlooked in small groups of unassigned cells not labeling for markers used to assign different cell IDs.
We hope this review inspires research directly examining this new paradigm. Here we instead consider indirect evidence from evolution research that nervous system cells specialized for iron regulation preceded cells specialized for myelin generation.
5. Evolutionary Emergence of Molecular and Cellular Mechanisms for Iron Regulation
Iron is the action hero in the story of life, a powerful catalytic enabler for a multitude of diverse bioactivities that have shaped all terrestrial organisms. Figure 3 gives a broad perspective of the story of iron and terrestrial life. Here we review evidence for iron as a primary driver in the emergence of life, independent of other early world views such as Iron-Sulfur world, RNA world, Metabolism-first or Genes-first. Irrespective of which world view prevails, we contend that iron is likely to have had central roles. In particular, there is strong evidence for our main theme - the emergence of iron-regulating systems and cells before the emergence of nervous systems and myelin.
Literature on evolution is continuously evolving. Aspects of the discussion below, such as dating, orders of emergence and properties of organisms and their iron-regulation and nervous systems, will in turn evolve as perspectives change, new information emerges and conflicting findings are resolved. Orders and timing of evolutionary events were based on recently revised reconciliation of molecular clock and fossil record estimates [101], OneZoom Tree of Life Explorer (https://www.onezoom.org/) or other sources cited elsewhere, recognizing these estimates may alter as more information emerges.
5.1. Iron as a Powerful Driver of the Emergence and Evolution of Early Terrestrial Life
Iron was a prominent constituent of the early planetesimals that formed after the Sun was created ~4.6 billion years ago (Ga), being abundant in the inner core of the newborn Earth, which formed ~4.5 Ga, and dispersing to the outer layers via convection currents, volcanic activity and hydrothermal vents [102,103,104,105,106,107].
5.1.1. Iron and the Emergence of Early Macromolecules and Membranes
Iron is an avid electron trader. It can exchange electrons with hydrogen, sulfate and other important early elements and has been cast as the primeval driver of early biochemistry[106,108,109,110]. From prevailing geochemical and biochemical perspectives, iron catalysis and iron-containing compounds with catalytic capabilities, such as Fe-S clusters, likely preceded emergence of larger molecules. Iron can catalyze generation of amino acids [111], peptides [112] and nucleobases [113], under conditions similar to those likely to have existed in early sediments and hydrothermal vents [114]. Iron is also able to catalyze simple bilayer membrane formation. Sequestration of iron in phospholipid bilayers, vesicles, endomembranes or other organelles and, ultimately, specialized cells, can help drive biochemical reactions by enabling high local concentrations of iron and other reactive species, provide safe storage reservoirs for iron and protect against iron theft by competing life forms [102,103,115,116,117,118,119].
It is hypothesized [120] that enzymes containing Fe-S clusters were already present 4 billion years ago in the Last Universal Common Ancestor (LUCA; Figure 3), a primitive prokaryote [121], and iron continues to have essential roles in diverse vital bioprocesses, spanning generation of energy, proteins, RNA, DNA and other macromolecules, oxygen respiration and cell replication, in all known cellular species from Bacteria, Archaea, Eukaryotes and plants through to Animalia.
As detailed below, we hypothesize that nervous system glial cells specialized for iron regulation and storage, i.e., Fe+glia, originally evolved from cells with primary functions in iron-regulation already present in multicellular organisms before the emergence of the nervous system, and that the iron-regulating cells in animals in turn arose initially from iron-regulating, non-animal unicellular ancestors.
5.1.2. Iron in Non-Animal Organisms
Iron-regulating systems were central to the existence of essentially all terrestrial lifeforms for over three billion years before multicellular Animalia and the simplest nervous systems arose. This in turn was over a hundred thousand years before myelination emerged (Figure 3). Early organisms evolved many systems for acquiring, using and storing iron [122]. Complex regulatory systems featuring homologs of most of the proteins central to human iron regulation are present in bacteria and other uni- and multi-cellular non-animal organisms such as fungi and plants [123,124,125,126]. Examples include proteins resembling human proteins such as the iron storage protein ferritin [127,128], iron regulatory protein 1, frataxin (FXN), which can serve as an iron sensor, Nramp 1 and 2 (divalent metal transport DMT1) and ferric chelate reductases [129,130]. For example, phytoplankton have highly sophisticated machinery for acquisition of iron, the main factor limiting their reproduction [131].
5.2. Iron and the Evolution of the Nervous System in Animals (Animalia)
The ubiquity and complexity of iron-regulating molecules in early metazoan animals such as sponges, jellyfish, corals and placozoa testify that vital iron regulatory systems important for unicellular and multicellular non-animal life were present in the hypothetical last common ancestor of all animals (Metazoa), a multicellular organism living over ~600 million years ago (Ma) [101,132,133].
Order of emergence of early animals such as sponges (Porifera), comb jellyfish (Ctenophora) and other early phyla (e.g., Placozoa, Cnidaria) is contentious [101,134,135,136], in part reflecting parallax of data from different sources e.g., fossil records, genomics [137,138]. It is also hard to map nervous system evolution accurately, partly due to lack of consensus on what constitutes a nervous system, a question beyond the scope of this review debated elsewhere [139,140,141].
Nervous system evolution is often framed as proceeding from simple nerve nets via segmentation, ganglia and spinal cord formation to the complex brain. To explore glial evolution with function in mind, we instead define stages in nervous system evolution by increasing integration and enclosure: i. proto-neural cells, with some features of both early neurons and early glia, ii. nerve networks comprising communicating nets of neurons without clear integrated control, iii. integrated (‘centralized’) unenclosed nervous systems, including nerve rings, iv. integrated, partially enclosed systems without meninges, and v. integrated systems fully enclosed in meninges or meninge-like structures [142,143,144,145,146].
We cover Porifera and Placozoa, without neurons or nervous systems [147,148], before Cnidaria and Ctenophora, which have diffuse neural nets and sometimes even centralized nervous systems (Section 5.2.3). While order of emergence of these phyla is debated, this accords with recent reconciliation of fossil records, molecular clocks and other analyses that puts Porifera first [101,136]. Regardless of order, the key point here is that molecules and cells with specialized iron-related functions are present in all these early phyla.
5.2.1. Iron in Sponges and Other Early Animals Without Neurons or Nervous Systems
Transcript or protein homologs of various important human iron-regulatory molecules have been observed in sponges, among the earliest metazoa e.g., ferritin heavy chain (FTH; confirmed at the protein level [149]), and the iron carrier transferrin, ferroportin, iron-regulatory protein 1 and heme synthesis components [148,149]. Iron-responsive, self-renewing, pluripotent stem-like cells may also be present [150,151,152]. Though usually considered to lack neurons, some sponges possess proto-neural forerunners of nervous system cells [150,152], termed ‘neuroids’ [153], and early forms of iron-dependent human neurotransmitters, e.g., GABA, glutamate [154,155,156,157].
Specialized iron-regulating cells in may also be present in another very early phylum, Placozoa, comprising small, disk-shaped marine creatures with few specialized cell types. Placozoa can preferentially acquire iron in gastrodermal cells by uptake of ferritin [158], suggesting specialized functions in iron handling, and some placozoa express the iron transporter melanotransferrin or non-response to iron deficiency 1 (NRF1) protein, a transferrin homolog coordinating iron balance [159,160,161]. Although no neurons have been identified, a few specialized cells expressing neural transcription factors such as NEUROD and OLIGs 1-3 may be proto-neural forerunners of nervous system cells [161].
It is unknown how early proto-neural or proto-neuronal precursors evolved to neurons, defined here as electrically excitable cells with specialized projections having primary functions in secretory or electrical signaling (adapted from [162]). Neurons and support cells (‘glia’) probably arose more than once in evolution and may have devolved in some lineages, contributing to uncertainty of phyla emergence order (Section 5.2). Greater understanding of iron regulation may come from detailed lineage characterization in early species, as underway for Placazoa, with neural and iron-related molecular profiling [161].
5.2.2. Iron Regulation in Jellyfish and Other Early Animals with Early Nervous Systems
Like placozoans, ctenophorans (comb jellyfish) can take up and digest ferritin in specialized cells [163] and some cnidarians (e.g., other jellyfish, sea anemones, corals, hydra) may also have complex iron-regulatory systems, including proteins such as ferritins [164], ferrochelatase and FXN, important in iron-sulfur (Fe-S) complex synthesis [165,166,167]. In humans, FXN mutations can cause neurosensory or motor impairment (Section 7.1.2.), suggesting FXN may have provided Fe-S clusters essential for mitochondrial activity in early prototypes of sensory and motor neurons.
Nervous systems comprising neurons connected by networks of inter-communicating cell processes that form diffuse nets, partially condensed plexuses or nerve rings are present in extant members of these early emerging phyla [140,168,169]. Key iron-dependent neurotransmitters are again observed (e.g., glutamate, GABA, dopamine [168,170,171,172]), suggesting iron was already central to many activities in the earliest nervous systems. Glia are not usually thought to be present in these early systems, [168,170,171,172], being considered to have evolved later in Bilateria (bilaterally symmetric organisms), with, or after, emergence of the central nervous system (CNS), e.g., [173].
Organisms in Ctenophora and Cnidaria typically feature radial symmetry and are not classed as Bilateria. Yet some exhibit bilateral symmetry and nerve rings in these phyla may constitute a single, integrated, ‘centralized’ command center for high level control of functions of ‘peripheral’ neurons in tentacles or ‘arms’ [140,168,169]. These deceptively simple systems can perform neurosensory and motor control functions, overriding local nerve control of individual arms for coordinated functions such as predation or jet propulsion and complex behaviors such as navigating by sun compass or visual cues and using individual tentacles to deliver captured prey to the mouth [171,172,174,175].
‘Division of labor’ between ‘proto-neurons’ and ‘proto-glia’ probably began in early metazoan phyla. Separation of iron-regulation from information handling is likely to confer strong evolutionary advantages, due to high toxicity risks from storing iron in cells specialized for signaling where iron regulation is not the primary function. Substrates and enzymes for synthesizing glutamate and other neurotransmitters are restricted to non-neuronal cells in some cnidaria [168,170,171,172,176,177,178]. Electrically active epithelial cells ensheathing neuron bundles in jellyfish and other early species are also considered to provide dedicated neuronal support that can be deemed to be a ‘glial’ function [140,179,180].
We propose cells specializing primarily for iron-related support functions (i.e., Fe+glia) were present in some species from very early in nervous system development, throughout the evolutionary of proto-neurons and neurons.
While this is not yet clearly demonstrated for the CNS, analogous phenomena occur in evolution of the eye, where iron is stored in melanin pigment that is often visualizable without staining. ‘Proto-eyes’ of larvae of the small cnidarian box jellyfish Tripedalia comprise a single pigmented photoreceptor cell expressing both opsin light receptive molecules and iron-containing dark melanin pigment that helps determine light direction and protect against excessive light exposure [181,182,183]. Melanin can also provide safe storage for intracellular iron. This ‘proto-eye’ is thought to have subsequently undergone asymmetric division into a specialized sensory neuron (photoreceptor) and a specialized iron-regulating pigmented glial cell providing metabolic support to the neuron [183]. Such proto-eyes may have been forerunners of the human eye, with Dunaief and colleagues reporting iron regulating genes involved in melanogenesis in vertebrate retina [184].
5.3. The Transition from Non-Myelinating to Myelinating Vertebrate Species
The examples above confirm cellular iron regulation, probably including early iron-rich proto-glial cells, was present in very early stages of animal evolution. In this Section we fast-forward to much later in evolution and consider the complex iron regulatory systems, probably including Fe+glia, that were present when myelin with key structural and functional features homologous to human myelin emerged, with the evolution of jawed vertebrates (rays and sharks) from jawless vertebrates (lamprey and hagfish). We will then back-track from this well-established ‘endpoint’ to develop a new paradigm for understanding the factors driving preceding evolution from iron-rich protoglia to iron-rich glia, i.e., Fe+glia, and the early ensheathment of neurons by ‘proto-myelin’ (Section 5.4).
5.3.1. Iron Regulation in Ensheathing but Non-Myelinating Jawless Vertebrates (Agnatha)
The switch from a non-myelinated nervous system without oligodendrocytes to myelin produced by oligodendrocytes is thought to have coincided with divergence of jawed (Gnathostomata) from jawless (Agnatha) vertebrates early in vertebrate evolution [185,186,187]. There are only two known extant Agnatha. The lampreys, primitive ‘vampires of the deep’, have sucker-like mouths lined with rings of sharp teeth that latch onto prey and siphon out blood and flesh. Hagfish lack oral suction disks and have evolved other ways of feeding without a jaw [188], including burrowing within dead or dying prey and absorbing nutrients, including iron, through their skin and gut epithelia [189].
Research has focused on lampreys, which have unique iron metabolizing organs, specialized bipolar iron-loading cells, melanin-containing macrophages in regions with iron-related activities, e.g., absorption, phagocytosis [190,191] and iron storage within adipocytes in fat tissue [192]. Pigmented cells likely to contain iron, occasionally with morphologies consistent with Fe+glia, appear around the periphery of the CNS by prolarval stage S15 [193]. Ferric iron in ferritin and small amounts of ferrous iron are seen in round, glycogen- and lipid-rich meningeal cells in larvae and juvenile brain and spinal cord [194].
Lamprey homologs of key iron regulatory genes include transferrin, ferritin heavy chain 1, prohibitin 2, which may control ferritin synthesis, the iron response element/iron regulatory protein (IRE/IRP) system and iron-regulating bone morphogenetic proteins (BMPs) [191,195,196,197,198,199,200,201,202,203]. The BMP receptor neogenin is expressed in ependymo-radial glial cells [204], possibly analogies to mammalian tanycytes (Section 5.4.1.1.).
Lamprey nervous system have similarities to ours [205,206], with iron-dependent neuronal systems ([205,207]) and glial-related transcription or growth factors e.g., Sox10 ortholog SoxE2, Nkx2.2, which regulates OLG precursors, and PDGFRα [185,205]. These may represent a glial gene-regulatory network in the vertebrate common ancestor [185]. While lacking myelin and myelinating cells, lamprey have ensheathing glia with some features of mammalian olfactory ensheathing cells (OEC; Section 3.3.5.), combining axon wrapping with metabolic support [186,208,209]. Whether these glia have iron-regulatory roles and their relationship, if any, to the iron-rich cells described above, is unknown.
5.3.2. Evidence for Myelin in Prehistoric Shark Fossils - the Oldest Known Jawed Vertebrate
Mammalian myelin and myelinating glia arose relatively early in vertebrate evolution [210,211]. The earliest surviving jawed vertebrate species (gnathostomes) in which myelin resembling human myelin has been reported are cartilaginous fishes, including rays and sharks (chondrichthyes), with myelination present in all chondrichthyes studied [187]. No known living species exist between the non-myelinated jawless lampreys and well-myelinated jawed rays and sharks.
This has led to suggestions that the myelin sheath and the hinged jaw were acquired together in evolution, with both arising from the neural crest, which gives rise to the jaw and most of the peripheral nervous system [187,212]. Zalc and colleagues have reported evidence for myelinated axons in a fossilized placoderm from the Devonian period, the oldest jawed fish known, and propose this is consistent with myelin similar to our own first emerging at this time [213].
Yet although most research has focused on oligodendroglia and myelin in vertebrates, beginning with sharks and other cartilaginous fish and moving forward, different types of glial support cells [93,214] and myelin-like substances [215,216,217] arose independently multiple times in evolution. Consideration of these other forms of myelin-related substances provides important insights into myelinating glia and other ensheathing glia and the functions of the substances they produce.
5.4. Nervous System Enclosure and the Emergence of Ferriglia and Ensheathing Glia
Glial support cells that cover portions of neuronal bodies or axons with non-myelin protolipidaceous substances are present in various species closely related to vertebrates within the bilaterian superphylum Deuterostomia. This superphylum eventually gave rise to homo sapiens and includes the major clade Chordata of which vertebrates are a subphylum. Myelin-like substances are also present in some species in the other main bilaterian superphylum Protostomia, which diverged from Deuterostomia ~570 to 560 Ma, and gave rise to arthropods, insects and many other species. Glia are proposed to have been present in the last common ancestor of these groups [218] and so are also found in most descendant species.
Here we will discuss non-myelinating ensheathing glia in early deuterostomes, with dorsal, primarily tube-like centralized nervous systems and peripheral ganglia, before moving on to protostomes, with ventral, primarily ganglionic, nervous systems (Section 5.5). Although glial evolution is usually considered to relate to ‘centralization’, we instead hypothesize that the main evolutionary drivers of glial cells with dedicated functions in the metabolic nurture or physical protection of neurons were factors that limited the direct access of nervous systems to external nutrients or restricted their ability to regenerate.
Examples include changes in organism and nervous system size, complexity or enclosure, and transitions to non-marine environments. Such factors can vary considerably even in closely related species and, as further discussed below, may provide more plausible explanations for the diverse evolutionary pathways by which glia emerged (and were sometimes lost) than nervous system centralization per se, as is usually assumed.
Regardless of emergence trajectories, storage of surplus iron and iron supply to neurons were probably early functions of glia, which are more expendable and easily replaced than post-mitotic neurons in complex, established communication networks, making them better suited for storing essential but potentially toxic substances such as iron.
Glia with primary iron-regulating functions (Fe+glia) may have arisen by repurposing of existing generic iron-regulatory cells that eventually became sequestered with neurons from the rest of the organism, and not by de novo development of iron trafficking by glial cells already solely serving the nervous system, as sometimes suggested (e.g., [85]). We propose glial support roles co-evolved with segregation of nervous systems from the rest of the organism by brain barriers restricting exposure to the blood or other circulatory systems or cerebrospinal fluid and enclosure within protective coverings, e.g., cartilaginous capsules or meninges.
5.4.1. Brain Barrier Systems
Although brain barriers now mainly comprise specialized endothelia (blood-brain barrier) or epithelia (choroid plexus; arachnoid epithelia), prototypal brain barriers in early organisms were predominantly glial [219,220,221,222]. This is exemplified by the early Bilateria subphylum Echinodermata, arising ~509 Ma, which has radial arm nerves and centralized nerve rings similar to Cnidaria and Ctenophora and a circulatory system open to the surrounding sea water. Echinodermata are non-chordate deuterostomes, and have a partly enclosed CNS with non-ensheathing glia forming the lining of the epineural canal, an early forerunner of the cerebral ventricles in the human brain [223,224,225].
Modern analogs of these glia in humans and other vertebrates include tanycytes, which in rat brain stain strongly for iron and ferritin, as well as other ependymoglial cells contributing to brain barrier formation in the cerebroventricular lining [59,226]. Although research is needed to determine if analogous glia in echinodermata also have specialized functions in regulating CNS iron supply, this suggests there may have been Fe+glia in nervous systems of multicellular organisms over 100 Ma before OLG cells with primary functions in myelination arose in early jawed vertebrates.
Evidence for glial-like neuronal support cells in early evolution is not restricted to Echinodermata. Other early bilaterians with glia-like cells supporting neurons in simple nets include Xenacoelomorpha, early invertebrates arising ~540 Ma, where glia-like cells may have convergently evolved at least five times [227]. Little is known about iron-regulating cells in these species [228].
5.4.2. Brain Enclosure by Meninges and Related Structures
There is also little information on evolution of meninges-like structures enclosing the brain. The other two chordate deuterostome subgroups, which preceded Vertebrata, are the Lancelets (cephalochordates; also called amphioxus), small eel-like animals which emerged within ~10 million years of Echinodermata; Figure 3) and the Tunicata (urochordates), which include Ascidiacea, sea squirts that have a tadpole-like larval form. In ~1899-1902, Italian anatomist Giuseppe Sterzi reported a spinal meninx comprising a single mesenchymal leaflet in Lancelets and its evolution in later species to the external dura mater and an internal ‘secondary meninx leaflet’ which then gave rise to the arachnoid and pia mater [229], but there has been almost no research in this area since then.
This makes Lancelets the first phylum known to have a primitive meninges and an early prototype of CNS enclosure, with a primitive neural tube and rudimentary ‘brain’. Perhaps not coincidentally, they are also the earliest known examples in deuterostomes of glial ensheathment of neurons, though ‘true’ multilayered myelin is not present [230,231,232,233,234,235,236].
We hypothesize proteolipidaceous sheathing provides Lancelet neurons with readily accessible stores of lipids and, importantly, iron (needed for mitochondrial energy generation), along with other essential nutrients such as fat-soluble vitamins. We conjecture these are early prototypes of mechanisms that supply energy via lipid metabolism or nutrients to axons from oligodendroglia or myelin in the mammalian brain [180,237,238,239,240,241,242]. Meninges protect nervous systems against pathogens competing for host iron and other nutrients, as well as from physical and chemical trauma. Meninx may have co-evolved with local storage of lipids, iron and other nutrients within nervous systems, though whether initially driving or secondary to accumulation of internal CNS nutrient stores is unclear.
There is also evidence for cells involved in iron regulation both within and outside the nervous system and for non-sheathing glial cells in the urochordate deuterostomes Ascidiacea, sea squirts with primitive neural tubes that evolved within ~10 million years of Echinodermata (Figure 3). The CNS of the tadpole-like ascidian larva also has neurons and glia, with two large pigmented sensory cells containing melanin [243,244,245]. This may be synthesized at greater rates when dietary iron is abundant [246], consistent with regulated iron storage functions. Iron does not appear to have been investigated in ascidian neural cells using sensitive, enhanced histochemical iron detection methods (Section 6).
It is beyond our scope to consider the many routes by which early nervous systems evolved, the uncertainty around orders and timing of emergence, often complicated by large differences between larval, juvenile and adult nervous systems even within a single species, as well as difficulties distinguishing convergent evolution from devolution/loss scenarios, and the possibility that sometimes less complex systems arising from ‘devolutionary’ loss of function may have evolutionary advantages in some environments.
However, while it may not be feasible to trace precise evolutionary sequences, the examples above support the hypothesis that specialized iron-regulating cells arose early in animal evolution and corollary that some of these cells evolved to roles in storing and transporting iron and probably also other nutrients within nervous systems.
5.5. Evolution of Myelin-like Ensheathing Structures in Invertebrate Nervous Systems
We now turn to the protostome invertebrates, with ventral, mostly ganglionic nervous systems, and examine the emergence and persistence of proteolipidaceous myelin-like ensheathing structures around neuronal soma and axons. From evolutionary perspectives, ‘true myelin’ usually referring to compact human or vertebrate myelin that confers biologically significant increases in axonal conduction rates, is widely considered superior to invertebrate myelin and other proteolipidaceous structures that often (but not always) lack this property, despite similarities in composition, structure or other functions.
As above, faster nerve conduction is widely considered the main driver for myelin evolution, but this may be of less fundamental importance than is often assumed [247]. Concordant with the roles proposed for iron-regulatory glia above, we hypothesize that the essential primary function served by myelin and other neuronal ensheathing structures, and driving their evolution, is the supply of iron and, secondarily, other nutrients to neurons and not saltatory conduction. We propose, in corollary, that saltatory conduction was instead a serendipitous downstream consequence of early incomplete ensheathment that became a secondary supplemental evolutionary driver.
5.5.1. Evolution of Invertebrate Myelin-like Structures
Like glia [93,214], myelin-like substances arose independently multiple times in protostome evolution [215,216,217]. In considering myelin evolution, it is important to recognize that while some researchers view ensheathing structures in the invertebrate nervous system as inferior to ‘true’ human or vertebrate myelin, some invertebrate ensheathing structures support very high conduction rates, comparable to vertebrates. Conversely, while the driver for myelin evolution is widely considered to be the benefits of rapid nerve conduction, especially in larger species, these may be less essential than often assumed [247].
For example, the phylum Cephalopoda, which arose ~330 Ma and includes octopus, cuttlefish and squid, contains some of the largest protostomes, with the largest brains. Yet although glia are present and nerve fibers are sometimes thinly sheathed by myelin-like structures, cephalopods lack compact, conduction-enhancing myelin structures comparable to vertebrate myelin, with rapid conduction instead supported by large diameter axons [248,249,250]. The Cephalopod brain is relatively complex and partly enclosed in a cartilaginous cranium, with a centralized component resembling the nerve rings that control tentacular nerves in cnidarians and some other species, rather than chordate/vertebrate brain structures. Most nutrients are transferred from the cephalopod circulation (hemolymph) across a glial barrier analogous to barriers described above for Echinodermata and below for Drosophila. This is again consistent with the possibility that local storage of iron and other nutrients in rudimentary ensheathing structures may have conferred early evolutionary advantages rather than saltatory conduction, even in species with long axons.
Structures with increasing similarities to vertebrate myelin are present in various later arising invertebrates, e.g., some Crustacea such as crayfish and crabs [217,251]. Notably some small copepod zooplankton ~1-10mm long, arising ~300 Ma [250], have well-organized, multilayered myelin-like structures. While differing from vertebrate myelin in wrapping modes, insulation properties or substructures such as nodes of Ranvier, some of these structures, notably fenestrated shrimp ‘myelin’, support very rapid conduction rates [252], but this does not necessarily translate to faster escape reactions [247,253].
However many protostome myelin-like structures do not appear to confer any increase in conduction rate, again suggesting the multiple instances of independent emergence and persistence of these structures reflects other, even more compelling, evolutionary advantages. Intriguingly, electron microscopy suggests some myelin-like ‘ensheathing’ structures in these early species are not generated by glia and instead arise inside healthy neuronal axolemma or soma, sometimes even extruding out to contact other cells [215,254,255].
This phenomena is not restricted to invertebrates or to neural cells. Lipid structures resembling myelin by electron microscopy, termed myeloid, myelinoid or myelin figures or bodies, can arise in non-neural cells in various vertebrates, including cells in lamprey undergoing metamorphosis [256] and rat [257] or human hepatocytes or kidney cells [258]. Membranous whorls of laminated phospholipids are also observed within human HeLa cells exposed to iron or other amphiphilic cations [257,259], though this is usually only observed in pathological contexts, often with degeneration of lysosomes, endoplasmic reticulum or other membranous organelles.
Phospholipid bilayers can also form spontaneously in some conditions, and this may be accelerated by iron (Section 5.1.1.), suggesting such structures may arise in response to high iron levels in the absence of specialized cellular synthesis machinery. Alternatively, some cells may retain ‘generic’ ancestral cellular machinery from early metazoan or pre-metazoan organisms for sequestering iron in vesicles or other membranous structures. Non-specialized cells may have only restricted capacity to store surplus iron safely, even by co-opting lysosomes or other membranous organelles, with excessive loading causing pathological organelle damage. Emergence of OLG-like cells primarily specialized for regulated generation of iron-sequestering membranous material could confer strong evolutionary advantages, although such cells may not themselves necessarily have initially had, or subsequently retained, primary specialized functions in iron storage or transport.
From anthropocentric perspectives, such structures may likewise sometimes be discounted as irrelevant to evolution of ‘true’ (i.e., human) myelin but we believe it is illuminating to recognize the existence of a spectrum of lipid-rich nervous system structures of differing origins and complexity that might confer other important benefits aside from insulation and accelerated signal conduction [253]. We propose the primary evolutionary advantage of such structures is their ability to function as safe ‘bulk storage’ reservoirs for iron and other nutrients, as well as for lipids themselves.
The evolution of advanced iron regulatory systems in invertebrates with non-myelin ensheathing structures is illustrated by Drosophila fruit flies, arising ~12 Ma. Drosophila brain is enclosed in a water-resistant protective cuticle formed predominantly of chitin, hydrocarbons and other lipids [260,261], with a blood-brain barrier comprised of glial cells [262]. Drosophila do not produce myelin but have ensheathing glia which, although proposed to resemble OLG more closely than other glia, despite not myelinating [263], do not produce wrappings that contain Ranvier node equivalents [264] or appear to increase nerve conduction velocities [265,266,267,268].
Drosophila larval neural stem cells – ‘neuroblasts’ – appear to depend on exogenous iron provided by glia for survival. This is stored in ferritin, which is produced in all subtypes of Drosophila glia, including astrocyte-like glia and perineurial glia, as well as ensheathing glia [269]. Extracellular ferritin in Drosophila is high in iron, unlike serum ferritin in humans [270], and appears to be transferred from the glia to the neuroblasts in vesicles [269]. The Drosophila genome encodes multiple ferritin and transferrin homologs, which may be expressed differentially in healthy brain or in response to infection or other stressors [271]. Importantly, outside the nervous system, a Drosophila transferrin homolog transfers iron to fat bodies for sequestration during pathogen infections, as iron is less readily accessible within these structures [272]. It is feasible that iron is contained within ensheathing structures produced by Drosophila glia and that these provide a safe local reservoir of iron, lipids and other nutrients for neurons, although little research appears to have addressed this directly.
In sum, we have presented a new iron-centric paradigm for understanding the evolution of nervous system iron regulation that has generated testable, fundamental hypotheses about Fe+glia and the primary functions of lipidaceous ensheathing structures homologous to myelin. To test these concepts, more research is needed to characterize cellular iron expression in key milestone species in the evolution of nervous systems, iron-rich glia, myelin and other ensheathing substances. Below we consider techniques that may help overcome some of the problems which have limited past research in this area.
6. Technical Considerations
It is clear from the literature that there has been little appreciation of the fundamental importance of deeper understanding of the cellular regulation of iron in the human brain and other nervous systems across evolution. One big factor contributing to the blind spots in the literature and the lack of research in this area has been the technical difficulties studying iron, which can be redistributed or lost during tissue processing with organic solvents. Had Hortega had access to the more sensitive iron labeling techniques now available, it is likely that neuroscience and evolutionary research would evolved very differently. Even today, few studies have used enhanced iron histochemical staining or other sensitive techniques to investigate the distribution of iron in nervous system cells and their ancestors in non-mammalian species. Below we briefly review historical and contemporary labeling or label-free methods of imaging iron at cellular and subcellular levels.
6.1. Challenges in Studying Iron and Other Metallic Elements in Brain Glial Cells
Iron is the most abundant transition metal in the human brain, but to gain full understanding of how iron in cells and tissues is compartmentalized and bound requires exceptional analytical sensitivity and specificity [274]. Viewing iron homeostasis as a system at the whole tissue level is key to determining its role in brain function in health and disease, and immunohistochemical analysis of proteins responsible for iron homeostasis can provide valuable insights [275]. Yet preserving iron chemistry in post-mortem tissue samples presents a key analytical challenge; conventional formalin fixation can incur substantive, unpredictable and compartment-specific effects on the levels of iron and other elements in tissue [276,277].
Conventional histological staining is also problematic for brain metals analysis. The Prussian or Berlin blue stain, developed in the 1860s [278], targets ferric iron but also detects ferrous iron, which is highly labile and rapidly converts to the ferric form [36]. Although Hortega trialed Prussian blue iron staining of human brain sections, he and colleagues achieved only pale staining of OLG by this method, perhaps explaining why he did not give due recognition to the possibility of OLG-like cells having iron regulatory roles. Many decades elapsed before this was revisited by James Connor, Torben Moos, ourselves and others [51,59,76,96,279,280] using modern techniques that included immunolabeling of iron-related proteins with enhanced Perls’ immunostaining using diaminobenzidine [36] or other agents.
Whilst suitable for conditions associated with iron over-accumulation, even with modern enhancements [36,76], stains such as Perls’ Prussian Blue may lack the requisite sensitivity for representative visualization of normal iron levels in brain tissue [281]. In addition, the dyes used are engaged in a competitive exchange equilibrium with endogenous ligands and are therefore unsuitable for multi-metal analysis [281]. This is significant when considering that metal transporters are commonly shared by different metals, so dysregulation of iron, for example, may cause knock-on effects for other essential metals and vice versa. Staining can also disrupt the native tissue chemistry and thereby limit accurate speciation of metal ions with regard to properties such as oxidation state [282]. For redox metals such as iron, this may be important in the context of oxidative damage mechanisms proposed to be among the factors implicating iron dysregulation in neurodegenerative disorders [275,283,284].
6.2. Small Molecule Fluorescence Probes and DNAzyme-Based Fluorescent Sensors of Labile Iron
New labeling approaches are now coming through that may reduce such problems and provide additional insights into brain iron homeostasis. In recent years, synthetic small-molecule probes have been developed by employing a tandem activity-based sensing and labelling strategy [285,286]. These fluorescence probes allow selective detection of Fe2+ that is loosely bound to surrounding molecules.
Another example is the use of DNAzyme-based fluorescent sensors to detect labile iron in different valence states. A DNAzyme-based sensor is a special type of biosensor that uses a DNA molecule with enzyme-like activity to detect specific substances. These sensors work by combining a DNAzyme with a detection system, such as a color change or fluorescence, to signal the presence of a target, such as metal ions, small molecules or genetic material.
Iron sensing DNAzymes utilize RNA-cleaving DNAzyme, which consists of an enzyme strand and a complementary substrate strand containing a single RNA linkage at the cleavage site. A fluorophore is conjugated to the enzyme strand, while two quenchers are positioned at opposite termini of the enzyme and substrate strands to suppress fluorescence in the intact state. Upon Fe ion binding, the DNAzyme catalyzes substrate cleavage, lowering the melting temperature at which the two DNA strands separate. This separation physically distances the fluorophore from the quencher, resulting in a fluorescence turn-on signal proportional to ion concentration. Such DNAzyme-based Fe sensors offer a sensitive, selective platform that is a promising alternative to traditional sensors for detecting different Fe forms with high selectivity, specificity and tunability. Importantly, this design decouples the target recognition core from the signal output, which allows multiplexing by incorporating fluorophores with distinct emission wavelengths. This property allows simultaneous imaging of the same metal ions with different valence states.
Two of us (YW, YL) have reported the construction and initial characterization of highly selective DNAzyme-based fluorescent sensors for selective, simultaneous imaging of each of Fe²⁺ and Fe³⁺ by using in vitro selection from a large DNA library of up to 1015 different sequences and converting to fluorescent sensors by the ‘catalytic beacon’ approach [287]. Although Fe+glia were not described specifically in this initial characterization, small iron-rich cells, some with visibly eccentric nuclei, consistent with features of Fe+glia described in this review, were picked up by the sensors in eleven-month-old 5xFAD mice and age, strain and sex-matched WT B6SJLF1/J) mice (Figure 4A in [287]). The sensors may provide a valuable platform for studying labile iron species, as opposed to iron bound within protein structures. By differentiating iron states and distribution in brain tissue, DNAzyme-based sensors also have the potential to enhance understanding of iron valence states in Fe+glia and other cells under different conditions and their contributions to iron homeostasis in health and pathogenesis, offering additional insights that may inform targeted therapeutic strategies.
6.3. Label-Free Imaging of Iron and Other Metallic Elements
Nowadays, the capacity to generate image contrast in a label-free manner using increasingly sophisticated chemical imaging techniques such as x-ray spectromicroscopy can also provide important alternative perspectives. Depending on the precise method applied, this class of techniques offers potential for multi-metal analysis of tissue samples at micro/nanometre length scales, with outstanding sensitivity and specificity [288], the option of collecting complementary information on metal chemical state and the potential to map, in the same experiment, corresponding distributions of certain organic components such as neuromelanin or amyloid beta [282,288,289,290,291].
Although modern multi-omics is now being widely applied in evolutionary research, to test the ideas put forward here, multi-omics needs to be performed in conjunction with established, as well as new, techniques for analyzing iron. This will be pivotal in advancing knowledge of nervous system iron regulation at the cellular level and understanding the past and present neurobiology of iron. It will also be important for gaining insights into the roles of Fe+glia in nervous system disease.
7. Involvement of Iron-Rich Cells Resembling OLG in Neurological Conditions
Iron and oligodendroglia have been widely studied in myelin disorders. Yet if Fe+glia help maintain healthy iron balance of neurons and other brain cells, Fe+glia dysfunction is likely to be implicated in iron imbalances in other brain conditions not primarily involving myelin. The many roles of iron in fundamental brain processes likely to be important for mental health are too numerous to address here, however we have briefly reviewed some of the neuropsychiatric conditions involving iron imbalance elsewhere [51]. Below we consider various other brain conditions likely to involve Fe+glia.
7.1. Iron-Rich Glia in Brain Diseases Involving Mutations in Iron-Related Proteins
Pathology of iron-rich brain cells with OLG-like morphologies – putative Fe+glia – is seen in some conditions involving mutations in proteins with primary or secondary roles in iron homeostasis. Two examples are discussed below.
7.1.1. Hereditary Neuroferritinopathy
Rare mutations in ferritin light chain 1 (FTL1), important in iron storage, cause hereditary neuroferritinopathy, a member of the ‘neurodegeneration with brain iron accumulation family (NBIA)’ [292,293,294,295]. It usually presents with chorea, dystonia or, less commonly, parkinsonism, progressing to severe motor disability, speech and swallowing deficits. It features cytoplasmic and nuclear ferritin inclusion bodies and increased levels of intracellular labile iron, with coalescence of abnormal intracellular iron and ferritin aggregates [296,297,298]. There is patent pathology of OLG-like cells containing intracellular iron or ferritin aggregates, with astroglia and neurons also affected, and extensive cell death throughout grey matter as well as white matter and myelin loss [296,297,298,299], but if, as appears likely, Fe+glia are affected, as well as recognized OLG species, and the nature of Fe+glial involvement remain to be determined.
7.1.2. Friedreich’s Ataxia
Friedreich’s ataxia (FRDA) is a rare genetic disease with characteristic symptoms of progressive cerebellar and sensory ataxia and limb dysmetria, primarily involving neuronal atrophy and synaptic degeneration, with myelin pathology considered secondary [300]. It is caused by FXN gene mutations reducing frataxin expression and function [301]. Frataxin regulates synthesis of mitochondrial iron-sulphur (Fe-S) clusters needed to make ATP [302] and is highly conserved in most prokaryotic and eukaryotic cells [303,304] (see also Section 4.2.2.). While not known to regulate iron directly, it may do so indirectly by altering heme synthesis through perturbation of mitochondrial respiratory chain function or other mechanisms.
Iron accumulation and related pathology only occur in specific regions in FRDA [302], consistent with heterogeneous mechanisms of iron regulation. The cerebellar dentate nucleus exhibits neuronal atrophy and synaptic terminal abnormalities that may reflect mitochondrial iron dysmetabolism [305]. Re-distribution of iron or ferritin from cells morphologically resembling OLG to other glia [305] could reflect astrocytes or microglia subsuming iron-regulatory functions or phagocytosing iron-containing materials due to dysfunction or death of specialized iron-regulating cells (Fe+glia). Effective treatment of FRDA may need to address the heterogeneous nature of iron homeostasis in different regions and cellular elements.
7.2. Iron-Rich Glia in Brain Diseases Involving Amyloids
Pathology that may involve Fe+glia is also seen in some common and rare brain diseases involving amyloid, including Alzheimer’s disease (AD), certain α synucleionopathies and transthyretin (TTR)-related siderosis.
7.2.1. Alzheimer’s Disease
Iron-rich cells with OLG-like morphology have been observed in AD, which features cognitive impairment and deposition of plaque structures containing Aβ amyloid aggregates. As in normal human brain, iron in post-mortem AD brain is observed in cells with OLG-like morphology, with eccentric nuclei positioned towards one side of the cell – i.e., Fe+glia. Transferrin or ferritin, individually or together, have been reported in these cells in normal brain and, for transferrin, also in AD brain, AD brain ferritin in [59,60,91]. Iron-rich cells with OLG-like morphology are often seen surrounding plaque structures containing Aβ amyloid aggregates in post-mortem brain from AD patients [65,91,306].
While literature on OLG-like cells in AD largely focuses on myelin and white matter loss [307,308], co-localization of Aβ aggregates with Fe+glia might interfere with other potentially important, functions of Fe+glia. This is also relevant when considering other conditions involving Aβ or other forms of amyloid. Notably, abnormal brain iron deposition (siderosis) has attracted recent attention in the context of severe, sometimes fatal responses to new immunotherapies targeting the Alzheimer’s Aβ peptide, sometimes also involving cerebrovascular amyloid angiopathy [309,310,311]. Understanding the biology of iron-rich glia may provide unexpected insights into these phenomena.
7.2.2. Superficial Siderosis with Amyloidosis
Superficial siderosis, featuring build-up of hemosiderin in the subarachnoid space of the brain or spinal cord, usually arises from iron deposition secondary to cerebrovascular hemorrhage. However this can also be caused by other factors, including co-aggregation of iron and amyloid, as illustrated by meningovascular amyloidosis of the thyroid hormone transporter transthyretin (TTR) [312]. Amyloid derived from TTR differs in protein composition from Aβ amyloid but can still cause abnormal glial iron accumulation, as illustrated by the autopsy of a patient with dementia, ataxia and leptomeningovascular, subpial and subependymal TTR amyloidosis, associated with siderosis, superficial cortical reactive gliosis and abnormal cortical glial iron accumulation, with little evidence of microvascular hemorrhage or myelin pallor [313]. One of us (SR) has shown pluripotential stem cell fate may be shifted towards OLG myelinating phenotypes in TTR null mice [314,315,316,317], with increased ‘oligodendrogenic to neurogenic’ ratios [318]. This suggests TTR normally limits cell fate commitment to the OLG lineage. More knowledge of . As glial iron homeostasis can be disrupted in patients with some TTR mutations, as above, it is possible TTR has direct or indirect roles in brain iron regulation involving Fe+glia that may affect normal brain functioning.
7.2.3. α-Synucleinopathies
The α-synucleinopathies are characterized by intracellular protein inclusions in neurons (e.g., Parkinson’s disease; PD), or glia (e.g., multiple system atrophy; MSA). While OLG changes can occur in PD or related diseases such as progressive supranuclear palsy that sometimes exhibit synuclein abnormalities [319,320], research on synucleinopathy of OLG lineage cells has mainly focused on MSA, a severe, fast-progressing, adult onset degenerative disease featuring impairment of the autonomic nervous system and movement [321]. The hallmark of MSA is cytoplasmic inclusions containing α-synuclein in glia usually considered to be oligodendrocytes, correlating with measures of pathogenesis [322]. Iron homeostasis may be dysregulated in MSA post-mortem brain [323], with regional or cellular redistribution, e.g., accumulation at some sites and functional deficiency at others [275,324]. It is unknown if Fe+glia have counterparts among iron-rich cells in MSA brain (e.g., [275]), if dysregulated glial iron homeostasis affects OLG maturation in MSA or, conversely, if perturbation of differentiation or maturation of OLG lineage cells and/or Fe+glia drives iron dyshomeostasis. It is also unknown how, if at all, any of these phenomena relate to α-synuclein aggregation.
OLG lineage specification and function appear dependent on tightly regulated gene expression changes determined by DNA methylation and other epigenetic mechanisms [325]. Epigenetic studies confirm dysregulation of oligodendrocyte-related genes in MSA e.g., myelin-associated oligodendrocyte basic protein (MOBP) [323,326,327]. It is unknown if decreased levels of MOBP accompanying increased DNA methylation in MSA affect the differentiation capacity of OPC or, conversely, if decreased MOBP expression reflects a shift from OLG differentiation towards Fe+glia proliferation.
We saw decreases in transcripts for MOBP and other oligodendrocyte-related genes in human post-mortem brain from two cases with neurodegeneration and brain iron accumulation without confirmed genetic etiology and mouse models with elevated brain iron loading [328,329]. In mouse brain, iron remained mostly distributed between myelin and glia, with only a small number of Fe+glia co-labeling for the pan-OLG marker OLIG2 and few, if any, for the astrocyte marker GFAP or the microglia marker IBA1 [51]. It will be illuminating to elucidate the nature and directionality of relationships between alterations in DNA methylation or other epigenomic factors and expression of proteins with proliferation or differentiation of Fe+glia or OPC.Relationships between α-synuclein glial cytoplasmic inclusions, iron dyshomeostasis and perturbation of iron-related proteins, and OLG fate and dysfunction in MSA warrant further investigation.
7.3. Cerebrovascular Disease and Stroke
Surprisingly little is known about OLG responses in stroke. Cells with OLG morphologies and functions in iron regulation in healthy brain might participate in clearing iron after hemorrhagic stroke. However cells with OLG morphologies may be more vulnerable than other glia to damage from ischemic insults [330,331]. For example, rat OLGs in the infarct core, identified by morphology after hemotoxylin/eosin staining and electron microscopy, were observed to swell as early as 30 minutes post-stroke and die within 2 hours [332]. This vulnerability could reflect high levels of iron, oxidative metabolic activity or glutamate receptor permeability, features consistent with non-myelin related primary functions, yet identities and functions of affected cells are rarely studied.
Sun and colleagues identified proliferation of OLG progenitors in the hyper acute phase (<24hours) as a rapid compensatory response to ischemic stroke in rats, consistent with functions in salvaging penumbral tissue, based on co-labeling for Ki67 and NG2+ (OPC and pericyte marker) [333]. Changes may also occur at later stages. Kishida and colleagues reported altered locations and phenotypes of OPC identified by PDGFRα from interstitial (away from microvessels) to intermediate- or juxta-vascular (near or fully wrapped around microvessels) with ischemic stroke in mice [334]. It is unknown if affected cells in either scenario have high iron contents or other factors that might contribute to increased susceptibility to ischemic damage. More research is required to assess involvement of non-myelinating cells with OLG-like morphologies and iron-regulating functions (i.e., candidate Fe+glia) in cerebrovascular disease and stroke responses.
7.4. Brain Cancers
Gliomas, the most prevalent and aggressive primary brain tumors, arise from malignant transformation of glial cells or their progenitors. Nutritional influences, including iron, feature in the multifaceted interactions of environmental risk factors with inherent genetic alterations that are proposed to modify glioma risk and progression from low-grade to high-grade gliomas [335,336]. Dysregulation of iron homeostatic systems is characteristic of gliomas, particularly glioblastomas, the most common and aggressive subtype of glioma, with aberrant expression of proteins such as homeostatic iron regulatory protein [337,338,339]. Iron has emerged as a promising target for glioma therapy due to its pivotal role in glioblastoma tumorigenesis. For example, long non-coding RNA (lncRNA) RP1-86C11.7 interacts with the microRNA hsa-miR-144-3p to suppresses transferrin receptor 1 levels, thereby inhibiting malignant properties of glioblastoma cells, including their proliferation and survival [340].
Yet many current and emerging treatments fail to consider the complex cellularity of gliomas and surrounding tissue [337]. With specific regard to how proliferation-competent iron rich cells with OLG-like morphologies may contribute to glioma, glioma stem cell-like cells (GSCs) with self-renewal capacity have high iron uptake compared to matched glioma non-stem cells [341,342,343] and are capable of self-renewal [337]. These cells may share some features with Fe+glia observed in healthy brain. Targeting such cells by iron chelation or other strategies might be effective in reducing tumor proliferation but might also impact normal iron-rich glial cells and, potentially, iron supply to neurons. Deeper understanding of proliferative Fe+glia is essential for developing safe, effective cancer therapeutics targeting brain iron.
7.5. CNS Infections and Neuroimmune Disorders
Some cells with OLG-like morphologies or expressing OLG-related markers in human brain or mouse models can manifest molecular profiles related to immune responses (e.g., cytokine receptors or transcripts for genes involved in antigen processing and presentation such as major histocompatibility complex (MHC) class I and II), suggesting these cells can undergo phenotypic changes to immune functions that can supplement or even take precedence over myelin-related functions and permit active participation in immune-related processes in healthy or disease-related contexts [21,80].
Iron has rarely been investigated and roles of Fe+glia in infection are unclear. Sequestration in cells or other membranous structures helps protect host iron from pathogens, as illustrated by iron-laden hemoglobin in red blood cells and containment of iron within other pigments such as hemerythrin or melanin (Section 4). As the main iron-sequestering cells in the healthy brain, Fe+glia are likely to be both frontline defenders against pathogens infiltrating brain tissue and, conversely, potential targets, yet iron is rarely studied in these contexts. With as many as 4 in 5 people who acquire SARS-CoV-2 infections developing neurological symptoms sometimes persisting long-term [344,345,346] and new pathogens continuing to emerge, greater understanding of neuroimmune roles of iron-regulating glia is needed, whatever their ontological classification.
8. Considerations for Iron Chelation and Other Therapies
Iron chelators have been trialed for some of the diseases above, notably glioblastoma [347,348], AD, PD, NBIA and FRDA, but while some trials returned mixed or promising results (e.g., [349,350]), others were unsuccessful and sometimes even exacerbated disease, notably recent AD and PD trials [351,352,353,354]. Patients with these conditions often do not display whole body iron overload [355] and chelator effects may be very different for brain cells with different iron loading properties. For example, high chelator doses might be needed to reduce iron in Fe+glia specialized for iron storage but may cause iron deficiency in neurons, already low in iron, directly or indirectly by interference with neuronal iron supply by Fe+glia. Developing selective therapeutic approaches for specific cellular targets to restore iron homeostasis in different disease contexts requires deeper understanding of the heterogeneous cellular elements of iron regulation.
9. Conclusions
We have presented strong evidence across multiple fronts for the existence of a novel fundamental neural cell group with primary functions relating to CNS iron regulation. Confirming the existence of these cells and understanding their properties and the implications for contemporary neuroscience, neurology and neuropsychiatry requires deeper knowledge of ferriglial subtypes in humans and other species, their evolution and where they reside in relation to recognized glial lineages and compartments.
If valid, the core ideas presented here will fundamentally reframe our understanding of the brain, its main cellular components, how iron has contributed to the emergence of life, Animalia and the shaping of nervous systems throughout animal evolution, and how our brains control iron in health and disease. We opened with the importance of speculation for original observation and will close with acknowledgement of the importance of other functions that are also served by speculation [356]. While contemporary knowledge does not speak to the validity of all the ideas above, we have tried to use the fresh perspectives opened by our conjectures to interrogate and integrate existing knowledge in insightful and illuminating ways. We hope this informs ongoing debates, lays foundations for testing the hypotheses and stimulates new ideas-in this area, even if ours are wrong.
Data Availability Statement
No new data were generated or analyzed in support of this research.
Acknowledgments
We thank Anjila Dongol for assisting with an earlier version of Figure 1. RH is supported by the Neurosurgical Research Foundation; CB is supported by Alzheimer’s Research UK and the Multiple System Atrophy Trust; JFC is supported by the Engineering and Physical Sciences Research Council [EP/X031179/1]; JB is supported by Race Against Dementia (ARUK-RADF2022B-008) funded by the Joseph and Lilian Sully Foundation in collaboration with Alzheimer’s Research UK; YK is supported by the Japan Society for the Promotion of Science (No. 23K10953) and Takeda Science Foundation 2022; YW and YL were supported by the National Institutes of Health (R35GM141931), the Alzheimer’s Association (AARG-NTF-22-926483), and the Robert A. Welch Foundation (grant F-0020).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Darwin, C.; Burkhardt, F.; Smith, S. The Correspondence of Charles Darwin: 1821-1836/[editors Frederick Burkhardt, Sydney Smith]; Cambridge University Press: 1985.
- Keynes, R.; Darwin, C. Charles Darwin’s zoology notes & specimen lists from HMS Beagle. 2001.
- Rozo, J.A.; Martínez-Gallego, I.; Rodríguez-Moreno, A. Cajal, the neuronal theory and the idea of brain plasticity. Front. Neuroanat. 2024, 18, 1331666. [CrossRef]
- García-Marín, V.; García-López, P.; Freire, M. Cajal's contributions to glia research. Trends Neurosci. 2007, 30, 479–487. [CrossRef]
- Tremblay, M.; Lecours, C.; Samson, L.; Sánchez-Zafra, V.; Sierra, A. From the Cajal alumni Achúcarro and Río-Hortega to the rediscovery of never-resting microglia. Front. Neuroanat. 2015, 9, 45–45. [CrossRef]
- y Cajal, R. Contribución al conocimiento de la neuroglia del cerebro humano. Trab Lab Invest Biol 1913, 11, 255.
- y Cajal, S.R. Sobre un nuevo proceder de impregnacion de la neuroglia y sus resultados en los centros nerviosos del hombre y animales; 1913.
- del Río-Hortega, P. La glía de escasas radiaciones (oligodendroglía) boletín de la real sociedad española de historia natural. Estud Neurol 1921, 21, 63-92.
- del Río-Hortega, P. El “tercer elemento” de los centros nerviosos [The “third element” of the nerve centers]. Bol Soc Esp Biol 1919, 9, 68-166.
- Kettenmann, H.; Verkhratsky, A. Neuroglia: the 150 years after. Trends Neurosci. 2008, 31, 653–659. [CrossRef]
- Verkhratsky, A.; Nedergaard, M. Physiology of Astroglia. Physiol. Rev. 2018, 98, 239–389. [CrossRef]
- Boullerne, A.I.; Feinstein, D.L. History of Neuroscience I. Pío del Río-Hortega (1882–1945): The Discoverer of Microglia and Oligodendroglia. ASN Neuro 2020, 12. [CrossRef]
- Delgado-García, J.M. Cajal and the Conceptual Weakness of Neural Sciences. Front. Neuroanat. 2015, 9. [CrossRef]
- James, O.G.; Mehta, A.R.; Behari, M.; Chandran, S. Centenary of the oligodendrocyte. The Lancet Neurology 2021, 20, 422.
- Hasel, P.; Rose, I.V.L.; Sadick, J.S.; Kim, R.D.; Liddelow, S.A. Neuroinflammatory astrocyte subtypes in the mouse brain. Nat. Neurosci. 2021, 24, 1475–1487. [CrossRef]
- Batiuk, M.Y.; Martirosyan, A.; Wahis, J.; de Vin, F.; Marneffe, C.; Kusserow, C.; Koeppen, J.; Viana, J.F.; Oliveira, J.F.; Voet, T.; et al. Identification of region-specific astrocyte subtypes at single cell resolution. Nat. Commun. 2020, 11, 1–15. [CrossRef]
- Tabata, H. Diverse subtypes of astrocytes and their development during corticogenesis. Front. Neurosci. 2015, 9, 114–114. [CrossRef]
- Stratoulias, V.; Ruiz, R.; Kanatani, S.; Osman, A.M.; Keane, L.; Armengol, J.A.; Rodríguez-Moreno, A.; Murgoci, A.-N.; García-Domínguez, I.; Alonso-Bellido, I.; et al. ARG1-expressing microglia show a distinct molecular signature and modulate postnatal development and function of the mouse brain. Nat. Neurosci. 2023, 26, 1008–1020. [CrossRef]
- Stratoulias, V.; Venero, J.L.; Tremblay, M.-È.; Joseph, B. Microglial subtypes: diversity within the microglial community. EMBO J. 2019, 38, e101997. [CrossRef]
- Tremblay, M. Microglial functional alteration and increased diversity in the challenged brain: Insights into novel targets for intervention. Brain, Behav. Immun. - Heal. 2021, 16, 100301. [CrossRef]
- Jäkel, S.; Agirre, E.; Falcão, A.M.; van Bruggen, D.; Lee, K.W.; Knuesel, I.; Malhotra, D.; Ffrench-Constant, C.; Williams, A.; Castelo-Branco, G. Altered human oligodendrocyte heterogeneity in multiple sclerosis. Nature 2019, 566, 543–547. [CrossRef]
- Marques, S.; Zeisel, A.; Codeluppi, S.; van Bruggen, D.; Falcão, A.M.; Xiao, L.; Li, H.; Häring, M.; Hochgerner, H.; Romanov, R.A.; et al. Oligodendrocyte heterogeneity in the mouse juvenile and adult central nervous system. Science 2016, 352, 1326–1329. [CrossRef]
- Prinz, M.; Jung, S.; Priller, J. Microglia Biology: One Century of Evolving Concepts. Cell 2019, 179, 292–311. [CrossRef]
- Giordano, K.R.; Denman, C.R.; Dubisch, P.S.; Akhter, M.; Lifshitz, J. An update on the rod microglia variant in experimental and clinical brain injury and disease. Brain Commun. 2021, 3, fcaa227. [CrossRef]
- Paolicelli, R.C.; Sierra, A.; Stevens, B.; Tremblay, M.-E.; Aguzzi, A.; Ajami, B.; Amit, I.; Audinat, E.; Bechmann, I.; Bennett, M.; et al. Microglia states and nomenclature: A field at its crossroads. Neuron 2022, 110, 3458–3483. [CrossRef]
- Kurtz, A.; Seltmann, S.; Bairoch, A.; Bittner, M.-S.; Bruce, K.; Capes-Davis, A.; Clarke, L.; Crook, J.M.; Daheron, L.; Dewender, J.; et al. A Standard Nomenclature for Referencing and Authentication of Pluripotent Stem Cells. Stem Cell Rep. 2018, 10, 1–6. [CrossRef]
- Konishi, H.; Koizumi, S.; Kiyama, H. Phagocytic astrocytes: Emerging from the shadows of microglia. Glia 2022, 70, 1009–1026. [CrossRef]
- Geyer, S.; Jacobs, M.; Hsu, N.-J. Immunity Against Bacterial Infection of the Central Nervous System: An Astrocyte Perspective. Front. Mol. Neurosci. 2019, 12, 57. [CrossRef]
- Akay, L.A.; Effenberger, A.H.; Tsai, L.-H. Cell of all trades: oligodendrocyte precursor cells in synaptic, vascular, and immune function. Genes Dev. 2021, 35, 180–198. [CrossRef]
- Asadollahi, E.; Trevisiol, A.; Saab, A.S.; Looser, Z.J.; Dibaj, P.; Ebrahimi, R.; Kusch, K.; Ruhwedel, T.; Möbius, W.; Jahn, O.; et al. Oligodendroglial fatty acid metabolism as a central nervous system energy reserve. Nat. Neurosci. 2024, 27, 1934–1944. [CrossRef]
- Omari, K.M.; John, G.; Lango, R.; Raine, C.S. Role for CXCR2 and CXCL1 on glia in multiple sclerosis. Glia 2005, 53, 24–31. [CrossRef]
- Omari, K.M.; John, G.R.; Sealfon, S.C.; Raine, C.S. CXC chemokine receptors on human oligodendrocytes: implications for multiple sclerosis. Brain 2005, 128, 1003–1015. [CrossRef]
- Pérez-Cerdá, F.; Sánchez-Gómez, M.V.; Matute, C. Pío del Río Hortega and the discovery of the oligodendrocytes. Front Neuroanat 2015, 9, 92.
- Ghasemi-Kasman, M.; Zare, L.; Baharvand, H.; Javan, M. In vivo conversion of astrocytes to myelinating cells by miR-302/367 and valproate to enhance myelin repair. J Tissue Eng Regen Med 2018, 12, e462-e472.
- Khanghahi, A.M.; Satarian, L.; Deng, W.; Baharvand, H.; Javan, M. In vivo conversion of astrocytes into oligodendrocyte lineage cells with transcription factor Sox10; Promise for myelin repair in multiple sclerosis. PLOS ONE 2018, 13, e0203785. [CrossRef]
- Meguro, R.; Asano, Y.; Odagiri, S.; Li, C.; Iwatsuki, H.; Shoumura, K. Nonheme-iron histochemistry for light and electron microscopy: a historical, theoretical and technical review. Arch. Histol. Cytol. 2007, 70, 1–19. [CrossRef]
- Bentivoglio, M.; Cotrufo, T.; Ferrari, S.; Tesoriero, C.; Mariotto, S.; Bertini, G.; Berzero, A.; Mazzarello, P. The Original Histological Slides of Camillo Golgi and His Discoveries on Neuronal Structure. Front. Neuroanat. 2019, 13, 3. [CrossRef]
- Chvátal, A.; Verkhratsky, A. An Early History of Neuroglial Research: Personalities. Neuroglia 2018, 1, 245–257. [CrossRef]
- y Cajal, S.R. Estructura de los centros nerviosos de las aves; 1888.
- Saceleanu, V.M.; Covache-Busuioc, R.-A.; Costin, H.-P.; Glavan, L.-A.; Ciurea, A.V. An Important Step in Neuroscience: Camillo Golgi and His Discoveries. Cells 2022, 11, 4112. [CrossRef]
- Parpura, V.; Verkhratsky, A. Astrocytes revisited: concise historic outlook on glutamate homeostasis and signaling. Croat. Med J. 2012, 53, 518–528. [CrossRef]
- Ramos, A.J. Astroglial heterogeneity: merely a neurobiological question? Or an opportunity for neuroprotection and regeneration after brain injury? Neural Regen Res 2016, 11, 1739-1741.
- del Rio-Hortega, P. Noticia de un nuevo y fácil método para la coloración de la neuroglia y del tejido conjuntivo. Trab Lab Invest Biol 1918, 15, 367-378.
- del Río-Hortega, P. Tercera aportación al conocimiento morfológico e interpretación funcional de la oligodendroglía; 1928.
- del Rio-Hortega, P. Cytology & cellular pathology of the nervous system; Hoeber: 1932.
- del Rio-Hortega, P. The microglia. 1939.
- del Rio-Hortega, P.; De Estudios, P.A. La microglia y su transformacion en células en bastoncito y cuerpos granulo-adiposos. Arch Neurobiol (Madr) 1920, 1, 171.
- Penfield, W. Oligodendroglia and its relation to classical neuroglia. Brain 1924, 47, 430–452. [CrossRef]
- Del Bigio, M.R. History of research concerning the ependyma: a view from inside the human brain. Front. Cell. Neurosci. 2024, 17, 1320369. [CrossRef]
- Butt, A.; Verkhratsky, A. Neuroglia: Realising their true potential. Brain Neurosci. Adv. 2018, 2. [CrossRef]
- for the UK Brain Expression Consortium (UKBEC); Heidari, M.; Johnstone, D.M.; Bassett, B.; Graham, R.M.; Chua, A.C.G.; House, M.J.; Collingwood, J.F.; Bettencourt, C.; Houlden, H.; et al. Brain iron accumulation affects myelin-related molecular systems implicated in a rare neurogenetic disease family with neuropsychiatric features. Mol. Psychiatry 2016, 21, 1599–1607. [CrossRef]
- Schnatz, A.; Müller, C.; Brahmer, A.; Krämer-Albers, E. Extracellular Vesicles in neural cell interaction and CNS homeostasis. FASEB BioAdvances 2021, 3, 577–592. [CrossRef]
- Pistono, C.; Bister, N.; Stanová, I.; Malm, T. Glia-Derived Extracellular Vesicles: Role in Central Nervous System Communication in Health and Disease. Front. Cell Dev. Biol. 2021, 8. [CrossRef]
- Granseth, B.; Odermatt, B.; Royle, S.J.; Lagnado, L. Clathrin-Mediated Endocytosis Is the Dominant Mechanism of Vesicle Retrieval at Hippocampal Synapses. Neuron 2006, 51, 773–786. [CrossRef]
- LoGiudice, L.; Matthews, G. The synaptic vesicle cycle: is kissing overrated? Neuron 2006, 51, 676-677.
- Ishikawa, K. On the Genesis of Hortega Cells.. Folia Anat. Jpn. 1932, 10, 229–248_1. [CrossRef]
- Iglesias-Rozas, J.R.; Garrosa, M. Río-Hortega’s third contribution to the morphological knowledge and functional interpretation of the oligodendroglia; Newnes: 2013.
- Connor, J.R.; Menzies, S.L. Relationship of iron to oligodendrocytes and myelination. Glia 1996, 17, 83-93.
- Connor, J.R.; Benkovic, S.A. Iron regulation in the brain: histochemical, biochemical, and molecular considerations. Ann Neurol 1992, 32, S51-S61.
- Connor, J.R.; Menzies, S.L.; Martin, S.M.S.; Mufson, E.J. Cellular distribution of transferrin, ferritin, and iron in normal and aged human brains. J. Neurosci. Res. 1990, 27, 595–611. [CrossRef]
- Battefeld, A.; Klooster, J.; Kole, M.H.P. Myelinating satellite oligodendrocytes are integrated in a glial syncytium constraining neuronal high-frequency activity. Nat. Commun. 2016, 7, 11298–11298. [CrossRef]
- Wood, P.; Bunge, R.P. The biology of the oligodendrocyte. In Oligodendroglia; Springer: 1984; pp. 1-46.
- Ludwin, S. The perineuronal satellite oligodendrocyte: A role in remyelination. Acta Neuropathol 1979, 47, 49-53.
- Belachew, S.; Yuan, X.; Gallo, V. Unraveling Oligodendrocyte Origin and Function by Cell-Specific Transgenesis. Dev. Neurosci. 2001, 23, 287–298. [CrossRef]
- Connor, J.R.; Menzies, S.L. Cellular management of iron in the brain. J. Neurol. Sci. 1995, 134, 33–44. [CrossRef]
- Takasaki, C.; Yamasaki, M.; Uchigashima, M.; Konno, K.; Yanagawa, Y.; Watanabe, M. Cytochemical and cytological properties of perineuronal oligodendrocytes in the mouse cortex. Eur. J. Neurosci. 2010, 32, 1326–1336. [CrossRef]
- Bernstein, H.-G.; Keilhoff, G.; Dobrowolny, H.; Guest, P.C.; Steiner, J. Perineuronal oligodendrocytes in health and disease: the journey so far. Prog. Neurobiol. 2019, 31, 89–99. [CrossRef]
- Radtke, C.; Sasaki, M.; Lankford, K.L.; Gallo, V.; Kocsis, J.D. CNPase Expression in Olfactory Ensheathing Cells. BioMed Res. Int. 2011, 2011, 608496. [CrossRef]
- Zhou, M.; Kiyoshi, C.M. Astrocyte syncytium: a functional reticular system in the brain. Neural Regen. Res. 2019, 14, 595–596. [CrossRef]
- Pacholko, A.G.; Wotton, C.A.; Bekar, L.K. Astrocytes - the ultimate effectors of long-range neuromodulatory networks? Front Cell Neurosci 2020, 14, 581075.
- Szuchet, S.; Nielsen, J.A.; Lovas, G.; Domowicz, M.S.; de Velasco, J.M.; Maric, D.; Hudson, L.D. The genetic signature of perineuronal oligodendrocytes reveals their unique phenotype. Eur. J. Neurosci. 2011, 34, 1906–1922. [CrossRef]
- van Landeghem, F.K.; Weiss, T.; von Deimling, A. Expression of PACAP and glutamate transporter proteins in satellite oligodendrocytes of the human CNS. Regul Pept 2007, 142, 52-59.
- Xin, W.; Mironova, Y.A.; Shen, H.; Marino, R.A.; Waisman, A.; Lamers, W.H.; Bergles, D.E.; Bonci, A. Oligodendrocytes Support Neuronal Glutamatergic Transmission via Expression of Glutamine Synthetase. Cell Rep. 2019, 27, 2262–2271.e5. [CrossRef]
- Wellman, S.M.; Cambi, F.; Kozai, T.D. The role of oligodendrocytes and their progenitors on neural interface technology: A novel perspective on tissue regeneration and repair. Biomaterials 2018, 183, 200–217. [CrossRef]
- Palhol, J.S.C.; Balia, M.; Terán, F.S.-R.; Labarchède, M.; Gontier, E.; Battefeld, A. Direct association with the vascular basement membrane is a frequent feature of myelinating oligodendrocytes in the neocortex. Fluids Barriers CNS 2023, 20, 1–17. [CrossRef]
- Moos, T.; Møllgård, K. A sensitive post-DAB enhancement technique for demonstration of iron in the central nervous system. Histochem. 1993, 99, 471–475. [CrossRef]
- Morris, C.M.; Candy, J.M.; Keith, A.B.; Oakley, A.E.; Taylor, G.A.; Edwardson, J.A.; Bloxham, C.A.; Pullen, R.G.; Gocht, A. Brain iron homeostasis. J. Inorg. Biochem. 1992, 47, 257–265. [CrossRef]
- Wawrzyniak, A.; Balawender, K.; Lalak, R.; Staszkiewicz, R.; Boroń, D.; Grabarek, B.O. Oligodendrocytes in the periaqueductal gray matter and the corpus callosum in adult male and female domestic sheep. Brain Res. 2022, 1792, 148036. [CrossRef]
- Kirby, L.; Jin, J.; Cardona, J.G.; Smith, M.D.; Martin, K.A.; Wang, J.; Strasburger, H.; Herbst, L.; Alexis, M.; Karnell, J.; et al. Oligodendrocyte precursor cells present antigen and are cytotoxic targets in inflammatory demyelination. Nat. Commun. 2019, 10, 1–20. [CrossRef]
- Falcão, A.M.; van Bruggen, D.; Marques, S.; Meijer, M.; Jäkel, S.; Agirre, E.; Samudyata; Floriddia, E.M.; Vanichkina, D.P.; Ffrench-Constant, C.; et al. Disease-specific oligodendrocyte lineage cells arise in multiple sclerosis. Nat. Med. 2018, 24, 1837–1844. [CrossRef]
- Richardson, W.D.; Young, K.M.; Tripathi, R.B.; McKenzie, I. NG2-glia as Multipotent Neural Stem Cells: Fact or Fantasy?. Neuron 2011, 70, 661–673. [CrossRef]
- Belachew, S.; Chittajallu, R.; Aguirre, A.A.; Yuan, X.; Kirby, M.; Anderson, S.; Gallo, V. Postnatal NG2 proteoglycan–expressing progenitor cells are intrinsically multipotent and generate functional neurons. J. Cell Biol. 2003, 161, 169–186. [CrossRef]
- Belachew, S.; Gallo, V. Synaptic and extrasynaptic neurotransmitter receptors in glial precursors' quest for identity. Glia 2004, 48, 185–196. [CrossRef]
- Ramón-Cueto, A.; Avila, J. Olfactory ensheathing glia: properties and function. Brain Res. Bull. 1998, 46, 175–187. [CrossRef]
- Mietto, B.S.; Jhelum, P.; Schulz, K.; David, S. Schwann Cells Provide Iron to Axonal Mitochondria and Its Role in Nerve Regeneration. J. Neurosci. 2021, 41, 7300–7313. [CrossRef]
- Valverde, F.; Lopez-Mascaraque, L. Neuroglial arrangements in the olfactory glomeruli of the hedgehog. J. Comp. Neurol. 1991, 307, 658–674. [CrossRef]
- Liu, L.; Tao-Cheng, J.-H.; Rallapalli, H.; Dodd, S.; Bouraoud, N.; Koretsky, A.P. High iron in mouse olfactory ensheathing cells at the glia limitans in the olfactory bulb underlies MRI T2* hypointensity. Imaging Neurosci. 2024, 2, 1–15. [CrossRef]
- Zhao, D.; Hu, M.; Liu, S. Glial cells in the mammalian olfactory bulb. Front. Cell. Neurosci. 2024, 18, 1426094. [CrossRef]
- Spassky, N.; Heydon, K.; Mangatal, A.; Jankovski, A.; Olivier, C.; Queraud-Lesaux, F.; Goujet-Zalc, C.; Thomas, J.L.; Zalc, B. Sonic hedgehog-dependent emergence of oligodendrocytes in the telencephalon: evidence for a source of oligodendrocytes in the olfactory bulb that is independent of PDGFRα signaling. Development 2001, 128, 4993–5004. [CrossRef]
- Wong, F.K.; Favuzzi, E. The brain's polymath: Emerging roles of microglia throughout brain development. Curr. Opin. Neurobiol. 2023, 79, 102700. [CrossRef]
- Connor, J.; Menzies, S.; St. Martin, S.; Mufson, E. A histochemical study of iron, transferrin, and ferritin in Alzheimer’s diseased brains. J Neurosci Res 1992, 31, 75-83.
- Morris, C.; Candy, J.; Oakley, A.; Bloxham, C.; Edwardson, J. Histochemical Distribution of Non-Haem Iron in the Human Brain. Acta Anat (Basel) 1992, 144, 235–257. [CrossRef]
- Hines, J.H. Evolutionary Origins of the Oligodendrocyte Cell Type and Adaptive Myelination. Front. Neurosci. 2021, 15. [CrossRef]
- Zhang, P.; Land, W.; Lee, S.; Juliani, J.; Lefman, J.; Smith, S.R.; Germain, D.; Kessel, M.; Leapman, R.; Rouault, T.A.; et al. Electron tomography of degenerating neurons in mice with abnormal regulation of iron metabolism. J. Struct. Biol. 2005, 150, 144–153. [CrossRef]
- Cheli, V.T.; Correale, J.; Paez, P.M.; Pasquini, J.M. Iron Metabolism in Oligodendrocytes and Astrocytes, Implications for Myelination and Remyelination. ASN Neuro 2020, 12. [CrossRef]
- Gerber, M.R.; Connor, J.R. Do oligodendrocytes mediate iron regulation in the human brain? Ann Neurol 1989, 26, 95-98.
- Werner, H.; Buscham, T.; Eichel, M.; Siems, S. Turning to myelin turnover. Neural Regen. Res. 2019, 14, 2063–2066. [CrossRef]
- Meyer, N.; Rinholm, J.E. Mitochondria in myelinating oligodendrocytes: slow and out of breath? Metabolites 2021, 11, 359.
- Reinert, A.; Morawski, M.; Seeger, J.; Arendt, T.; Reinert, T. Iron concentrations in neurons and glial cells with estimates on ferritin concentrations. BMC Neurosci. 2019, 20, 1–14. [CrossRef]
- Fünfschilling, U.; Supplie, L.M.; Mahad, D.; Boretius, S.; Saab, A.S.; Edgar, J.; Brinkmann, B.G.; Kassmann, C.M.; Tzvetanova, I.D.; Möbius, W.; et al. Glycolytic oligodendrocytes maintain myelin and long-term axonal integrity. Nature 2012, 485, 517–521. [CrossRef]
- Carlisle, E.; Yin, Z.; Pisani, D.; Donoghue, P.C.J. Ediacaran origin and Ediacaran-Cambrian diversification of Metazoa. Sci. Adv. 2024, 10, eadp7161. [CrossRef]
- Wade, J.; Byrne, D.J.; Ballentine, C.J.; Drakesmith, H. Temporal variation of planetary iron as a driver of evolution. Proc. Natl. Acad. Sci. USA 2021, 118. [CrossRef]
- Nairz, M.; Schroll, A.; Sonnweber, T.; Weiss, G. The struggle for iron - a metal at the host-pathogen interface. Cell. Microbiol. 2010, 12, 1691–1702. [CrossRef]
- Deamer, D. The Role of Lipid Membranes in Life’s Origin. Life 2017, 7, 5. [CrossRef]
- Burcar, B.T.; Barge, L.M.; Trail, D.; Watson, E.B.; Russell, M.J.; McGown, L.B. RNA Oligomerization in Laboratory Analogues of Alkaline Hydrothermal Vent Systems. Astrobiology 2015, 15, 509–522. [CrossRef]
- Camprubi, E.; Jordan, S.F.; Vasiliadou, R.; Lane, N. Iron catalysis at the origin of life. IUBMB Life 2017, 69, 373–381. [CrossRef]
- Heard, A.W.; Bekker, A.; Kovalick, A.; Tsikos, H.; Ireland, T.; Dauphas, N. Oxygen production and rapid iron oxidation in stromatolites immediately predating the Great Oxidation Event. Earth Planet. Sci. Lett. 2022, 582. [CrossRef]
- Johnson, J.E.; Present, T.M.; Valentine, J.S. Iron: Life’s primeval transition metal. Proc Natl Acad Sci U S A 2024, 121, e2318692121.
- Wächtershäuser, G. From volcanic origins of chemoautotrophic life to Bacteria, Archaea and Eukarya. Philos. Trans. R. Soc. B: Biol. Sci. 2006, 361, 1787–1806; discussion 1806-1788. [CrossRef]
- Muchowska, K.B.; Varma, S.J.; Moran, J. Synthesis and breakdown of universal metabolic precursors promoted by iron. Nature 2019, 569, 104–107. [CrossRef]
- Barge, L.M.; Flores, E.; Baum, M.M.; VanderVelde, D.G.; Russell, M.J. Redox and pH gradients drive amino acid synthesis in iron oxyhydroxide mineral systems. Proc. Natl. Acad. Sci. 2019, 116, 4828–4833. [CrossRef]
- Sydow, C.; Sauer, F.; Siegle, A.F.; Trapp, O. Iron-Mediated Peptide Formation in Water and Liquid Sulfur Dioxide under Prebiotically Plausible Conditions**. ChemSystemsChem 2022, 5. [CrossRef]
- Shanker, U.; Bhushan, B.; Bhattacharjee, G.; Kamaluddin Formation of Nucleobases from Formamide in the Presence of Iron Oxides: Implication in Chemical Evolution and Origin of Life. Astrobiology 2011, 11, 225–233. [CrossRef]
- Georgieva, M.N.; Little, C.T.; Maslennikov, V.V.; Glover, A.G.; Ayupova, N.R.; Herrington, R.J. The history of life at hydrothermal vents. Earth-Science Rev. 2021, 217. [CrossRef]
- Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K., Walter, P. The lipid bilayer. In Molecular Biology of the Cell, 4th ed.; Garland Science: New York, 2002.
- Suwalsky, M.; Martínez, F.; Cárdenas, H.; Grzyb, J.; Strzałka, K. Iron affects the structure of cell membrane molecular models. Chem. Phys. Lipids 2005, 134, 69–77. [CrossRef]
- Płaczkiewicz, J.; Gieczewska, K.; Musiałowski, M.; Adamczyk-Popławska, M.; Bącal, P.; Kwiatek, A. Availability of iron ions impacts physicochemical properties and proteome of outer membrane vesicles released by Neisseria gonorrhoeae. Sci. Rep. 2023, 13, 18733. [CrossRef]
- Gould, S.B.; Garg, S.G.; Martin, W.F. Bacterial Vesicle Secretion and the Evolutionary Origin of the Eukaryotic Endomembrane System. Trends Microbiol. 2016, 24, 525–534. [CrossRef]
- Cohen, Z.R.; Todd, Z.R.; Wogan, N.; Black, R.A.; Keller, S.L.; Catling, D.C. Plausible Sources of Membrane-Forming Fatty Acids on the Early Earth: A Review of the Literature and an Estimation of Amounts. ACS Earth Space Chem. 2022, 7, 11–27. [CrossRef]
- Weiss, M.C.; Sousa, F.L.; Mrnjavac, N.; Neukirchen, S.; Roettger, M.; Nelson-Sathi, S.; Martin, W.F. The physiology and habitat of the last universal common ancestor. Nat. Microbiol. 2016, 1, 16116. [CrossRef]
- Moody, E.R.R.; Álvarez-Carretero, S.; Mahendrarajah, T.A.; Clark, J.W.; Betts, H.C.; Dombrowski, N.; Szánthó, L.L.; Boyle, R.A.; Daines, S.; Chen, X.; et al. The nature of the last universal common ancestor and its impact on the early Earth system. Nat. Ecol. Evol. 2024, 8, 1654–1666. [CrossRef]
- Bradley, J.M.; Svistunenko, D.A.; Wilson, M.T.; Hemmings, A.M.; Moore, G.R.; Le Brun, N.E. Bacterial iron detoxification at the molecular level. J. Biol. Chem. 2020, 295, 17602–17623. [CrossRef]
- Philpott, C.C. Iron uptake in fungi: A system for every source. Biochim. et Biophys. Acta (BBA) - Mol. Cell Res. 2006, 1763, 636–645. [CrossRef]
- Bairwa, G.; Jung, W.H.; Kronstad, J.W. Iron acquisition in fungal pathogens of humans. Metallomics 2017, 9, 215–227. [CrossRef]
- Liu, Y.; Xiong, Z.; Wu, W.; Ling, H.-Q.; Kong, D. Iron in the Symbiosis of Plants and Microorganisms. Plants 2023, 12, 1958. [CrossRef]
- Sun, J.; Xiao, S.; Xue, C. The tug-of-war on iron between plant and pathogen. Phytopathol. Res. 2023, 5, 1–14. [CrossRef]
- Arosio, P.; Cairo, G.; Bou-Abdallah, F. A Brief History of Ferritin, an Ancient and Versatile Protein. Int. J. Mol. Sci. 2024, 26, 206. [CrossRef]
- Grant, R.; Filman, D.; Finkel, S.; Kolter, R.; Hogle, J. The crystal structure of Dps, a ferritin homolog that binds and protects DNA. Nat. Struct. Mol. Biol. 1998, 5, 294–303. [CrossRef]
- Peracino, B.; Buracco, S.; Bozzaro, S. The Nramp (Slc11) proteins regulate development, resistance to pathogenic bacteria and iron homeostasis in Dictyostelium discoideum. J. Cell Sci. 2013, 126, 301–311. [CrossRef]
- Peracino, B.; Monica, V.; Primo, L.; Bracco, E.; Bozzaro, S. Iron metabolism in the social amoeba Dictyostelium discoideum: A role for ferric chelate reductases. Eur. J. Cell Biol. 2022, 101, 151230. [CrossRef]
- Schoffman, H.; Lis, H.; Shaked, Y.; Keren, N. Iron–Nutrient Interactions within Phytoplankton. Front. Plant Sci. 2016, 7, 1223–1223. [CrossRef]
- Ros-Rocher, N.; Pérez-Posada, A.; Leger, M.M.; Ruiz-Trillo, I. The origin of animals: an ancestral reconstruction of the unicellular-to-multicellular transition. Open Biol. 2021, 11, 200359. [CrossRef]
- Müller, W.E. Review: How was metazoan threshold crossed? The hypothetical Urmetazoa. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2001, 129, 433–460. [CrossRef]
- Delsuc, F.; Philippe, H.; Tsagkogeorga, G.; Simion, P.; Tilak, M.-K.; Turon, X.; López-Legentil, S.; Piette, J.; Lemaire, P.; Douzery, E.J.P. A phylogenomic framework and timescale for comparative studies of tunicates. BMC Biol. 2018, 16, 39. [CrossRef]
- Moroz, L.L.; Kohn, A.B. Independent origins of neurons and synapses: insights from ctenophores. Philos. Trans. R. Soc. B: Biol. Sci. 2016, 371, 20150041. [CrossRef]
- Kapli, P.; Telford, M.J. Topology-dependent asymmetry in systematic errors affects phylogenetic placement of Ctenophora and Xenacoelomorpha. Sci. Adv. 2020, 6, eabc5162. [CrossRef]
- Holland, L.Z. Evolution of basal deuterostome nervous systems. J. Exp. Biol. 2015, 218, 637–645. [CrossRef]
- Satoh, N.; Rokhsar, D.; Nishikawa, T. Chordate evolution and the three-phylum system. Proc. R. Soc. B: Biol. Sci. 2014, 281, 20141729. [CrossRef]
- Miguel-Tomé, S.; Llinás, R.R. Broadening the definition of a nervous system to better understand the evolution of plants and animals. Plant Signal. Behav. 2021, 16, 1927562. [CrossRef]
- Mackie, G.O.; Meech, R.W. Central Circuitry in the Jellyfish Aglantha Digitale: I. The Relay System. J. Exp. Biol. 1995, 198, 2261–2270. [CrossRef]
- Watanabe, H.; Fujisawa, T.; Holstein, T.W. Cnidarians and the evolutionary origin of the nervous system. Dev. Growth Differ. 2009, 51, 167–183. [CrossRef]
- Evans, S.D.; Droser, M.L.; Erwin, D.H. Developmental processes in Ediacara macrofossils. Proc. R. Soc. B: Biol. Sci. 2021, 288, 20203055. [CrossRef]
- Gavilán, B.; Perea-Atienza, E.; Martínez, P. Xenacoelomorpha: a case of independent nervous system centralization?. Philos. Trans. R. Soc. B: Biol. Sci. 2016, 371, 20150039. [CrossRef]
- Arendt, D.; Bertucci, P.Y.; Achim, K.; Musser, J.M. Evolution of neuronal types and families. Curr. Opin. Neurobiol. 2019, 56, 144–152. [CrossRef]
- Liebeskind, B.J.; Hillis, D.M.; Zakon, H.H.; Hofmann, H.A. Complex Homology and the Evolution of Nervous Systems. Trends Ecol. Evol. 2016, 31, 127–135. [CrossRef]
- Hejnol, A.; Rentzsch, F. Neural nets. Curr. Biol. 2015, 25, R782–R786. [CrossRef]
- Moroz, L.L.; Romanova, D.Y. Alternative neural systems: What is a neuron? (Ctenophores, sponges and placozoans). Front. Cell Dev. Biol. 2022, 10, 1071961. [CrossRef]
- Natalio, F.; Wiese, S.; Friedrich, N.; Werner, P.; Tahir, M.N. Localization and Characterization of Ferritin in Demospongiae: A Possible Role on Spiculogenesis. Mar. Drugs 2014, 12, 4659–4676. [CrossRef]
- Finoshin, A.D.; Adameyko, K.I.; Mikhailov, K.V.; Kravchuk, O.I.; Georgiev, A.A.; Gornostaev, N.G.; Kosevich, I.A.; Mikhailov, V.S.; Gazizova, G.R.; Shagimardanova, E.I.; et al. Iron metabolic pathways in the processes of sponge plasticity. PLOS ONE 2020, 15, e0228722. [CrossRef]
- Müller, W.E.; Korzhev, M.; Le Pennec, G.; Müller, I.M.; Schröder, H.C. Origin of metazoan stem cell system in sponges: first approach to establish the model (Suberites domuncula). Biomol. Eng. 2003, 20, 369–379. [CrossRef]
- Krasko, A.; Schröder, H.C.; Batel, R.; Grebenjuk, V.A.; Steffen, R.; Müller, I.M.; Müller, W.E. Iron Induces Proliferation and Morphogenesis in Primmorphs from the Marine SpongeSuberites domuncula. DNA Cell Biol. 2002, 21, 67–80. [CrossRef]
- Müller, W.E. The stem cell concept in sponges (Porifera): Metazoan traits. Semin. Cell Dev. Biol. 2006, 17, 481–491. [CrossRef]
- Lechauve, C.; Jager, M.; Laguerre, L.; Kiger, L.; Correc, G.; Leroux, C.; Vinogradov, S.; Czjzek, M.; Marden, M.C.; Bailly, X. Neuroglobins, Pivotal Proteins Associated with Emerging Neural Systems and Precursors of Metazoan Globin Diversity. J. Biol. Chem. 2013, 288, 6957–6967. [CrossRef]
- Ho, V.R.; Goss, G.G.; Leys, S.P. ATP and glutamate coordinate contractions in the freshwater sponge Ephydatia muelleri. J. Exp. Biol. 2025, 228. [CrossRef]
- Adameyko, K.I.; Burakov, A.V.; Finoshin, A.D.; Mikhailov, K.V.; Kravchuk, O.I.; Kozlova, O.S.; Gornostaev, N.G.; Cherkasov, A.V.; Erokhov, P.A.; Indeykina, M.I.; et al. Conservative and Atypical Ferritins of Sponges. Int. J. Mol. Sci. 2021, 22, 8635. [CrossRef]
- Riesgo, A.; Farrar, N.; Windsor, P.J.; Giribet, G.; Leys, S.P. The Analysis of Eight Transcriptomes from All Poriferan Classes Reveals Surprising Genetic Complexity in Sponges. Mol. Biol. Evol. 2014, 31, 1102–1120. [CrossRef]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E.A.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon queenslandica genome and the evolution of animal complexity. Nature 2010, 466, 720–726. [CrossRef]
- Mayorova, T.D.; Koch, T.L.; Kachar, B.; Jung, J.H.; Reese, T.S.; Smith, C.L. Placozoan secretory cell types implicated in feeding, innate immunity and regulation of behavior. PLOS ONE 2025, 20, e0311271. [CrossRef]
- Romanova, D.Y.; Povernov, A.A.; Nikitin, M.A.; Borman, S.I.; Frank, Y.A.; Moroz, L.L. Long-term dynamics of placozoan culture: emerging models for population and space biology. Front. Cell Dev. Biol. 2025, 12, 1514553. [CrossRef]
- Najera, D.G.; Dittmer, N.T.; Weber, J.J.; Kanost, M.R.; Gorman, M.J. Phylogenetic and sequence analyses of insect transferrins suggest that only transferrin 1 has a role in iron homeostasis. Insect Sci. 2020, 28, 495–508. [CrossRef]
- Najle, S.R.; Grau-Bové, X.; Elek, A.; Navarrete, C.; Cianferoni, D.; Chiva, C.; Cañas-Armenteros, D.; Mallabiabarrena, A.; Kamm, K.; Sabidó, E.; et al. Stepwise emergence of the neuronal gene expression program in early animal evolution. Cell 2023, 186, 4676–4693.e29. [CrossRef]
- Jékely, G.; Keijzer, F.; Godfrey-Smith, P. An option space for early neural evolution. Philos. Trans. R. Soc. B: Biol. Sci. 2015, 370, 20150181. [CrossRef]
- Steinmetz, P.R.H. A non-bilaterian perspective on the development and evolution of animal digestive systems. Cell Tissue Res. 2019, 377, 321–339. [CrossRef]
- A Schwarz, J.; Brokstein, P.B.; Voolstra, C.; Terry, A.Y.; Manohar, C.F.; Miller, D.J.; Szmant, A.M.; Coffroth, M.A.; Medina, M. Coral life history and symbiosis: Functional genomic resources for two reef building Caribbean corals, Acropora palmata and Montastraea faveolata. BMC Genom. 2008, 9, 97–97. [CrossRef]
- Zhou, Y.; Liu, H.; Feng, C.; Lu, Z.; Liu, J.; Huang, Y.; Tang, H.; Xu, Z.; Pu, Y.; Zhang, H. Genetic adaptations of sea anemone to hydrothermal environment. Sci. Adv. 2023, 9, eadh0474. [CrossRef]
- Jain, A.; Connolly, E.L. Mitochondrial iron transport and homeostasis in plants. Front. Plant Sci. 2013, 4, 348. [CrossRef]
- Smith, H.L.; Pavasovic, A.; Surm, J.M.; Phillips, M.J.; Prentis, P.J. Evidence for a Large Expansion and Subfunctionalization of Globin Genes in Sea Anemones. Genome Biol. Evol. 2018, 10, 1892–1901. [CrossRef]
- Sprecher, S.G. Neural Cell Type Diversity in Cnidaria. Front. Neurosci. 2022, 16, 909400. [CrossRef]
- Koizumi, O.; Hamada, S.; Minobe, S.; Hamaguchi-Hamada, K.; Kurumata-Shigeto, M.; Nakamura, M.; Namikawa, H. The nerve ring in cnidarians: its presence and structure in hydrozoan medusae. Zoology 2014, 118, 79–88. [CrossRef]
- Moroz, L.L. Convergent evolution of neural systems in ctenophores. J. Exp. Biol. 2015, 218, 598–611. [CrossRef]
- Kass-Simon, G.; Pierobon, P. Cnidarian chemical neurotransmission, an updated overview. Comp Biochem Physiol A Mol Integr Physiol 2007, 146, 9-25.
- Carlberg, M.; Anctil, M. Biogenic amines in coelenterates. Comp Biochem Physiol C Comp Pharmacol Toxicol 1993, 106, 1-9.
- De Robertis, E.M.; Tejeda-Muñoz, N. Evo-Devo of Urbilateria and its larval forms. Dev. Biol. 2022, 487, 10–20. [CrossRef]
- Katsuki, T.; Greenspan, R.J. Jellyfish nervous systems. Curr. Biol. 2013, 23, R592–R594. [CrossRef]
- Satterlie, R.A. The search for ancestral nervous systems: an integrative and comparative approach. J. Exp. Biol. 2015, 218, 612–617. [CrossRef]
- Chari, T.; Weissbourd, B.; Gehring, J.; Ferraioli, A.; Leclère, L.; Herl, M.; Gao, F.; Chevalier, S.; Copley, R.R.; Houliston, E.; et al. Whole-animal multiplexed single-cell RNA-seq reveals transcriptional shifts acrossClytiamedusa cell types. Sci. Adv. 2021, 7, eabh1683. [CrossRef]
- Oren, M.; Brikner, I.; Appelbaum, L.; Levy, O. Fast Neurotransmission Related Genes Are Expressed in Non Nervous Endoderm in the Sea Anemone Nematostella vectensis. PLOS ONE 2014, 9, e93832. [CrossRef]
- Sheloukhova, L.; Watanabe, H. Evolution of glial cells: a non-bilaterian perspective. Neural Dev. 2024, 19, 10. [CrossRef]
- Garm, A.; Poussart, Y.; Parkefelt, L.; Ekström, P.; Nilsson, D.-E. The ring nerve of the box jellyfish Tripedalia cystophora. Cell Tissue Res. 2007, 329, 147–157. [CrossRef]
- Rey, S.; Zalc, B.; Klämbt, C. Evolution of glial wrapping: A new hypothesis. Dev. Neurobiol. 2020, 81, 453–463. [CrossRef]
- Gehring, W.J. New Perspectives on Eye Development and the Evolution of Eyes and Photoreceptors. J. Hered. 2005, 96, 171–184. [CrossRef]
- O'Connor, M.; Garm, A.; Marshall, J.N.; Hart, N.S.; Ekström, P.; Skogh, C.; Nilsson, D.-E.; Megan, O.; Anders, G.; N., M.J.; et al. Visual pigment in the lens eyes of the box jellyfish Chiropsella bronzie. Proc. R. Soc. B: Biol. Sci. 2010, 277, 1843–1848. [CrossRef]
- Gehring, W.; Seimiya, M. Eye evolution and the origin of Darwin's eye prototype. Ital. J. Zoöl. 2010, 77, 124–136. [CrossRef]
- Wolkow, N.; Li, Y.; Maminishkis, A.; Song, Y.; Alekseev, O.; Iacovelli, J.; Song, D.; Lee, J.C.; Dunaief, J.L. Iron upregulates melanogenesis in cultured retinal pigment epithelial cells. Exp. Eye Res. 2014, 128, 92–101. [CrossRef]
- Yuan, T.; York, J.R.; McCauley, D.W. Gliogenesis in lampreys shares gene regulatory interactions with oligodendrocyte development in jawed vertebrates. Dev. Biol. 2018, 441, 176–190. [CrossRef]
- Bullock, T.H.; Moore, J.K.; Fields, R. Evolution of myelin sheaths: Both lamprey and hagfish lack myelin. Neurosci. Lett. 1984, 48, 145–148. [CrossRef]
- Zalc, B. The acquisition of myelin: An evolutionary perspective. Brain Res. 2016, 1641, 4–10. [CrossRef]
- Clark, A.J.; Uyeno, T.A. Feeding in jawless fishes. In Feeding in Vertebrates: Evolution, Morphology, Behavior, Biomechanics, Bels, V., Whishaw, I.Q., Eds.; Springer International Publishing: Cham, 2019; pp. 189-230.
- Glover, C.N.; Niyogi, S.; Blewett, T.A.; Wood, C.M. Iron transport across the skin and gut epithelia of Pacific hagfish: Kinetic characterisation and effect of hypoxia. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2016, 199, 1–7. [CrossRef]
- Tsioros, K.; Youson, J. Intracellular distribution of iron (and associated elements) in various cell types of larvae and juveniles of the sea lamprey (Petromyzon marinus). Tissue Cell 1997, 29, 137–162. [CrossRef]
- Hall, S.J.; Youson, J.H. Distribution of ferric iron in larval lampreys, Petromyzon marinus L. Histol Histopathol. 1988, 3, 7–20.
- Macey, D.J.; Youson, J.H. Occurrence and Structure of Iron Inclusions in Adipocytes of Larval Lampreys. Acta Zoöl. 1990, 71, 69–76. [CrossRef]
- González-Llera, L.; Shifman, M.I.; Barreiro-Iglesias, A. Repulsive guidance molecule A (RGMA) is widely expressed in the CNS of the sea lamprey Petromyzon marinus during early development. MicroPubl Biol. 2024, 2024. [CrossRef]
- Youson, J.H.; Sargent, P.A.; Pearce, G.W. Iron and aluminum deposition in the meninges of the lamprey: Identification of an aluminum-ferritin inclusion body. Anat. Rec. 1989, 223, 13–20. [CrossRef]
- Barreiro-Iglesias, A.; Laramore, C.; Shifman, M.; Anadón, R.; Selzer, M.; Rodicio, M. The sea lamprey tyrosine hydroxylase: cDNA cloning and in situ hybridization study in the brain. Neuroscience 2010, 168, 659–669. [CrossRef]
- Andersen, Ø.; Pantopoulos, K.; Kao, H.; Muckenthaler, M.; Youson, J.H.; Pieribone, V. Regulation of iron metabolism in the sanguivore lamprey Lampetra fluviatilis --molecular cloning of two ferritin subunits and two iron-regulatory proteins (IRP) reveals evolutionary conservation of the iron-regulatory element (IRE)/IRP regulatory system. Eur. J. Biochem. 1998, 254, 223–229. [CrossRef]
- Macey, D.J.; Cake, M.H.; Potter, I.C. Exceptional iron concentrations in larval lampreys (Geotria australis) and the activities of superoxide radical detoxifying enzymes. Biochem. J. 1988, 252, 167–172. [CrossRef]
- Webster, R.; Pollara, B. Isolation and partial characterization of transferrin in the sea lamprey, Petromyzon marinus. Comp. Biochem. Physiol. 1969, 30, 509–527. [CrossRef]
- Smalley, S.R.; Macey, D.J.; Potter, I.C. Changes in the amount of nonhaem iron in the plasma, whole body, and selected organs during the postlarval life of the lampreyGeotria australis. J. Exp. Zoöl. 1986, 237, 149–157. [CrossRef]
- Yang, W.; Mu, B.; You, J.; Tian, C.; Bin, H.; Xu, Z.; Zhang, L.; Ma, R.; Wu, M.; Zhang, G.; et al. Non-classical ferroptosis inhibition by a small molecule targeting PHB2. Nat. Commun. 2022, 13, 7473. [CrossRef]
- Guo, J.; Lyu, S.; Qi, Y.; Chen, X.; Yang, L.; Zhao, C.; Wang, H. Molecular evolution and gene expression of ferritin family involved in immune defense of lampreys. Dev. Comp. Immunol. 2023, 146, 104729. [CrossRef]
- Birceanu, O.; Ferreira, P.; Neal, J.; Sunga, J.; Anthony, S.; Davidson, S.M.; Edwards, S.L.; Wilson, J.M.; Youson, J.H.; Vijayan, M.M.; et al. Divergent Pathways of Ammonia and Urea Production and Excretion during the Life Cycle of the Sea Lamprey. Physiol. Biochem. Zoöl. 2022, 95, 551–567. [CrossRef]
- McCauley, D.W.; Bronner-Fraser, M. Conservation and divergence of BMP2/4 genes in the lamprey: expression and phylogenetic analysis suggest a single ancestral vertebrate gene. Evol. Dev. 2004, 6, 411–422. [CrossRef]
- González-Llera, L.; Shifman, M.I.; Barreiro-Iglesias, A. Neogenin expression in ependymo-radial glia of the larval sea lamprey Petromyzon marinus spinal cord. MicroPubl Biol 2023, 2023. [CrossRef]
- Lamanna, F.; Hervas-Sotomayor, F.; Oel, A.P.; Jandzik, D.; Sobrido-Cameán, D.; Santos-Durán, G.N.; Martik, M.L.; Stundl, J.; Green, S.A.; Brüning, T.; et al. A lamprey neural cell type atlas illuminates the origins of the vertebrate brain. Nat. Ecol. Evol. 2023, 7, 1714–1728. [CrossRef]
- Robertson, B.; Kardamakis, A.; Capantini, L.; Pérez-Fernández, J.; Suryanarayana, S.M.; Wallén, P.; Stephenson-Jones, M.; Grillner, S. The lamprey blueprint of the mammalian nervous system. Prog Brain Res 2014, 212, 337-349. [CrossRef]
- Ryczko, D.; Cone, J.J.; Alpert, M.H.; Goetz, L.; Auclair, F.; Dubé, C.; Parent, M.; Roitman, M.F.; Alford, S.; Dubuc, R. A descending dopamine pathway conserved from basal vertebrates to mammals. Proc. Natl. Acad. Sci. 2016, 113, E2440–E2449. [CrossRef]
- Foo, L.C.; Dougherty, J.D. Aldh1L1 is expressed by postnatal neural stem cells in vivo. Glia 2013, 61, 1533–1541. [CrossRef]
- Weil, M.-T.; Heibeck, S.; Töpperwien, M.; Dieck, S.T.; Ruhwedel, T.; Salditt, T.; Rodicio, M.C.; Morgan, J.R.; Nave, K.-A.; Möbius, W.; et al. Axonal Ensheathment in the Nervous System of Lamprey: Implications for the Evolution of Myelinating Glia. J. Neurosci. 2018, 38, 6586–6596. [CrossRef]
- Möbius, W.; Patzig, J.; Nave, K.-A.; Werner, H.B. Phylogeny of proteolipid proteins: divergence, constraints, and the evolution of novel functions in myelination and neuroprotection. Neuron Glia Biol. 2008, 4, 111–127. [CrossRef]
- Kitagawa, K.; Sinoway, M.; Yang, C.; Gould, R.; Colman, D. A proteolipid protein gene family: Expression in sharks and rays and possible evolution from an ancestral gene encoding a pore-forming polypeptide. Neuron 1993, 11, 433–448. [CrossRef]
- Zalc, B.; Colman, D.R. Origins of Vertebrate Success. Science 2000, 288, 271–271. [CrossRef]
- Grassé, P. Le système nerveux des insectes. Traité de Zoologie. Anatomi, Systematique, Biologie. T. VIII, fasc. III 1975, 321-510.
- Hartline, D.K. The evolutionary origins of glia. Glia 2011, 59, 1215–1236. [CrossRef]
- Tomassy, G.S.; Dershowitz, L.B.; Arlotta, P. Diversity Matters: A Revised Guide to Myelination. Trends Cell Biol. 2016, 26, 135–147. [CrossRef]
- Knowles, L. The Evolution of Myelin: Theories and Application to Human Disease. J. Evol. Med. 2017, 5, 1–23. [CrossRef]
- Roots, B.I. The phylogeny of invertebrates and the evolution of myelin. Neuron Glia Biol. 2008, 4, 101–109. [CrossRef]
- Helm, C.; Karl, A.; Beckers, P.; Kaul-Strehlow, S.; Ulbricht, E.; Kourtesis, I.; Kuhrt, H.; Hausen, H.; Bartolomaeus, T.; Reichenbach, A.; et al. Early evolution of radial glial cells in Bilateria. Proc. R. Soc. B: Biol. Sci. 2017, 284, 20170743. [CrossRef]
- Abbott, N.J. Dynamics of CNS Barriers: Evolution, Differentiation, and Modulation. Cell. Mol. Neurobiol. 2005, 25, 5–23. [CrossRef]
- Bundgaard, M.; Abbott, N.J. All vertebrates started out with a glial blood-brain barrier 4–500 million years ago. Glia 2008, 56, 699–708. [CrossRef]
- Abbott, N.J.; Bundgaard, M. Electron-dense tracer evidence for a blood?brain barrier in the cuttlefishSepia officinalis. J. Neurocytol. 1992, 21, 276–294. [CrossRef]
- Bundgaard, M.; Abbott, N.J. Fine structure of the blood?brain interface in the cuttlefishSepia officinalis (Mollusca, Cephalopoda). J. Neurocytol. 1992, 21, 260–275. [CrossRef]
- Zueva, O.; Khoury, M.; Heinzeller, T.; Mashanova, D.; Mashanov, V. The complex simplicity of the brittle star nervous system. Front. Zoöl. 2018, 15, 1–26. [CrossRef]
- Märkel, K.; Röser, U. Ultrastructure and organization of the epineural canal and the nerve cord in sea urchins (Echinodermata, Echinoida). Zoomorphology 1991, 110, 267–279. [CrossRef]
- Mashanov, V.; Zueva, O. Radial glia in echinoderms. Developmental neurobiology 2019, 79, 396-405.
- Langlet, F. Tanycyte Gene Expression Dynamics in the Regulation of Energy Homeostasis. Front. Endocrinol. 2019, 10, 286. [CrossRef]
- Quiroga, S.Y.; Bonilla, E.C.; Bolaños, D.M.; Carbayo, F.; Litvaitis, M.K.; Brown, F.D. Evolution of flatworm central nervous systems: Insights from polyclads. Genet. Mol. Biol. 2015, 38, 233–248. [CrossRef]
- Bailly, X.; Reichert, H.; Hartenstein, V. The urbilaterian brain revisited: novel insights into old questions from new flatworm clades. Dev. Genes Evol. 2012, 223, 149–157. [CrossRef]
- Riva, A.; Orrù, B.; Riva, F.T. Giuseppe Sterzi (1876–1919) of the University of Cagliari: A brilliant neuroanatomist and medical historian. The Anatomical Record 2000, 261, 105-110. [CrossRef]
- Benito-Gutiérrez, È. A gene catalogue of the amphioxus nervous system. Int. J. Biol. Sci. 2006, 2, 149–160. [CrossRef]
- Holland, N.D.; Somorjai, I.M.L. The sensory peripheral nervous system in the tail of a cephalochordate studied by serial blockface scanning electron microscopy. J. Comp. Neurol. 2020, 528, 2569–2582. [CrossRef]
- Bozzo, M.; Lacalli, T.C.; Obino, V.; Caicci, F.; Marcenaro, E.; Bachetti, T.; Manni, L.; Pestarino, M.; Schubert, M.; Candiani, S. Amphioxus neuroglia: Molecular characterization and evidence for early compartmentalization of the developing nerve cord. Glia 2021, 69, 1654–1678. [CrossRef]
- Gattoni, G.; Andrews, T.G.R.; Benito-Gutiérrez, È. Restricted Proliferation During Neurogenesis Contributes to Regionalisation of the Amphioxus Nervous System. Front. Neurosci. 2022, 16, 812223. [CrossRef]
- He, C.; Han, T.; Liao, X.; Zhou, Y.; Wang, X.; Guan, R.; Tian, T.; Li, Y.; Bi, C.; Lu, N.; et al. Phagocytic intracellular digestion in amphioxus ( Branchiostoma ). Proc. R. Soc. B: Biol. Sci. 2018, 285, 20180438. [CrossRef]
- He, C.; Han, T.; Liao, X.; Zhou, Y.; Wang, X.; Guan, R.; Tian, T.; Li, Y.; Bi, C.; Lu, N.; et al. Correction to ‘Phagocytic intracellular digestion in amphioxus ( Branchiostoma )’. Proc. R. Soc. B: Biol. Sci. 2018, 285, 20181277. [CrossRef]
- Bosch-Queralt, M.; Fledrich, R.; Stassart, R.M. Schwann cell functions in peripheral nerve development and repair. Neurobiol. Dis. 2022, 176, 105952. [CrossRef]
- Clausen, J. The Effect of Vitamin A Deficiency on Myelination in the Central Nervous System of the Rat. Eur. J. Biochem. 1969, 7, 575–582. [CrossRef]
- Popescu, D.C.; Huang, H.; Singhal, N.K.; Shriver, L.; McDonough, J.; Clements, R.J.; Freeman, E.J. Vitamin K enhances the production of brain sulfatides during remyelination. PLOS ONE 2018, 13, e0203057. [CrossRef]
- Gomez-Pinedo, U.; Cuevas, J.A.; Benito-Martín, M.S.; Moreno-Jiménez, L.; Esteban-Garcia, N.; Torre-Fuentes, L.; Matías-Guiu, J.A.; Pytel, V.; Montero, P.; Matias-Guiu, J. Vitamin D increases remyelination by promoting oligodendrocyte lineage differentiation. Brain Behav. 2019, 10, e01498. [CrossRef]
- Goudarzvand, M.; Javan, M.; Mirnajafi-Zadeh, J.; Mozafari, S.; Tiraihi, T. Vitamins E and D3 Attenuate Demyelination and Potentiate Remyelination Processes of Hippocampal Formation of Rats Following Local Injection of Ethidium Bromide. Cell. Mol. Neurobiol. 2009, 30, 289–299. [CrossRef]
- Stadelmann, C.; Timmler, S.; Barrantes-Freer, A.; Simons, M. Myelin in the Central Nervous System: Structure, Function, and Pathology. Physiol. Rev. 2019, 99, 1381–1431. [CrossRef]
- Asadollahi, E.; Trevisiol, A.; Saab, A.S.; Looser, Z.J.; Dibaj, P.; Ebrahimi, R.; Kusch, K.; Ruhwedel, T.; Möbius, W.; Jahn, O.; et al. Oligodendroglial fatty acid metabolism as a central nervous system energy reserve. Nat. Neurosci. 2024, 27, 1934–1944. [CrossRef]
- Jeffery, W.R. Closing the wounds: One hundred and twenty five years of regenerative biology in the ascidian Ciona intestinalis. Genesis 2014, 53, 48–65. [CrossRef]
- Osugi, T.; Sasakura, Y.; Satake, H. The nervous system of the adult ascidian Ciona intestinalis Type A (Ciona robusta): Insights from transgenic animal models. PLOS ONE 2017, 12, e0180227–e0180227. [CrossRef]
- de Abreu, I.S.; Wajsenzon, I.J.R.; Dias, J.C.; Allodi, S.; Monteiro-De-Barros, C. Central nervous system regeneration in ascidians: cell migration and differentiation. Cell Tissue Res. 2022, 390, 335–354. [CrossRef]
- Stacey, J.; Driedzic, W. Temporal variability in, and impact of food availability on vanadium and iron concentrations in Ciona intestinalis tissues (Tunicata, Ascidiacea). J. Exp. Mar. Biol. Ecol. 2010, 386, 11–18. [CrossRef]
- Waggett, R.J.; Buskey, E.J. Escape reaction performance of myelinated and non-myelinated calanoid copepods. J. Exp. Mar. Biol. Ecol. 2008, 361, 111–118. [CrossRef]
- Hama, K. Some observations on the fine structure of the giant synapse in the stellate ganglk of the squid, Doryteuphis bleekeri. Z Zellforsch Mikrosk Anat 1962, 56, 437–444. [CrossRef]
- Bear, R.S.; Schmitt, F.O.; Young, J.Z. The sheath components of the giant nerve fibres of the squid. Proc. R. Soc. London. Ser. B. Biol. Sci. 1937, 123, 496–504. [CrossRef]
- Pumphrey, R.J.; Young, J.Z. The Rates of Conduction of Nerve Fibres of Various Diameters in Cephalopods. J. Exp. Biol. 1938, 15, 453–466. [CrossRef]
- Peracchia, C. Direct communication between axons and sheath glial cells in crayfish. Nature 1981, 290, 597–598. [CrossRef]
- Hsu), K.X.(.; Terakawa, S. Fenestration nodes and the wide submyelinic space form the basis for the unusually fast impulse conduction of shrimp myelinated axons. J. Exp. Biol. 1999, 202, 1979–1989. [CrossRef]
- Wang, S.S.-H.; Shultz, J.R.; Burish, M.J.; Harrison, K.H.; Hof, P.R.; Towns, L.C.; Wagers, M.W.; Wyatt, K.D. Functional Trade-Offs in White Matter Axonal Scaling. J. Neurosci. 2008, 28, 4047–4056. [CrossRef]
- Wilson, C.H.; Hartline, D.K. Novel organization and development of copepod myelin. ii. nonglial origin. J. Comp. Neurol. 2011, 519, 3281–3305. [CrossRef]
- Wilson, C.H.; Hartline, D.K. Novel organization and development of copepod myelin. i. ontogeny. J. Comp. Neurol. 2011, 519, 3259–3280. [CrossRef]
- Sidon, E.W.; Youson, J.H. Morphological changes in the liver of the sea lamprey, Petromyzon marinus L., during metamorphosis: I. Atresia of the bile ducts. J. Morphol. 1983, 177, 109–124. [CrossRef]
- Herdson, P.B.; Kaltenbach, J.P. Electron microscope studies on enzyme activity and the isolation of thiohydantoin-induced myelin figures in rat liver. J. Cell Biol. 1965, 25, 485–493. [CrossRef]
- Choung, H.Y.G.; Jean-Gilles, J.; Goldman, B. Myeloid bodies is not an uncommon ultrastructural finding. Ultrastruct. Pathol. 2022, 46, 130–138. [CrossRef]
- Jauregui, H.; Bradford, W.D.; Arstila, A.U.; Kinney, T.D.; Trump, B.F. Iron metabolism and cell membranes. III. Iron-induced alterations in HeLa cells. Am J Pathol. 1975, 80, 33–52.
- Ostrowski, S.; A Dierick, H.; Bejsovec, A. Genetic Control of Cuticle Formation During Embryonic Development ofDrosophila melanogaster. Genetics 2002, 161, 171–182. [CrossRef]
- Wang, Y.; Farine, J.-P.; Yang, Y.; Yang, J.; Tang, W.; Gehring, N.; Ferveur, J.-F.; Moussian, B. Transcriptional Control of Quality Differences in the Lipid-Based Cuticle Barrier in Drosophila suzukii and Drosophila melanogaster. Front. Genet. 2020, 11, 887. [CrossRef]
- Limmer, S.; Weiler, A.; Volkenhoff, A.; Babatz, F.; Klã¤Mbt, C. The Drosophila blood-brain barrier: development and function of a glial endothelium. Front. Neurosci. 2014, 8, 365–365. [CrossRef]
- Yildirim, K.; Petri, J.; Kottmeier, R.; Klämbt, C. Drosophila glia: Few cell types and many conserved functions. Glia 2018, 67, 5–26. [CrossRef]
- Banerjee, S.; Bhat, M.A. Glial ensheathment of peripheral axons in Drosophila. J. Neurosci. Res. 2007, 86, 1189–1198. [CrossRef]
- Beckers, P.; Helm, C.; Purschke, G.; Worsaae, K.; Hutchings, P.; Bartolomaeus, T. The central nervous system of Oweniidae (Annelida) and its implications for the structure of the ancestral annelid brain. Front. Zoöl. 2019, 16, 6. [CrossRef]
- Fernández, I.; Pardos, F.; Benito, J.; Roldán, C. Ultrastructural observations on the phoronid nervous system. J. Morphol. 1996, 230, 265–281. [CrossRef]
- Temereva, E.; Wanninger, A. Development of the nervous system in Phoronopsis harmeri (Lophotrochozoa, Phoronida) reveals both deuterostome- and trochozoan-like features. BMC Evol. Biol. 2012, 12, 121–121. [CrossRef]
- Garlick, R.L.; Williams, B.J.; Riggs, A.F. The hemoglobins of Phoronopsis viridis, of the primitive invertebrate phylum phoronida: Characterization and subunit structure. Arch. Biochem. Biophys. 1979, 194, 13–23. [CrossRef]
- Ma, Z.; Wang, W.; Yang, X.; Rui, M.; Wang, S. Glial ferritin maintains neural stem cells via transporting iron required for self-renewal in Drosophila. eLife 2024, 13. [CrossRef]
- Tang, X.; Zhou, B. Ferritin is the key to dietary iron absorption and tissue iron detoxification inDrosophila melanogaster. FASEB J. 2013, 27, 288–298. [CrossRef]
- Wan, B.; Belghazi, M.; Lemauf, S.; Poirié, M.; Gatti, J.-L. Proteomics of purified lamellocytes from Drosophila melanogaster HopT identifies new membrane proteins and networks involved in their functions. Insect Biochem. Mol. Biol. 2021, 134, 103584. [CrossRef]
- Iatsenko, I.; Marra, A.; Boquete, J.-P.; Peña, J.; Lemaitre, B. Iron sequestration by transferrin 1 mediates nutritional immunity inDrosophila melanogaster. Proc. Natl. Acad. Sci. USA 2020, 117, 7317–7325. [CrossRef]
- Verkhratsky, A.; Arranz, A.M.; Ciuba, K.; Pękowska, A. Evolution of neuroglia. Ann. New York Acad. Sci. 2022, 1518, 120–130. [CrossRef]
- Collingwood, J.F.; Davidson, M.R. The role of iron in neurodegenerative disorders: insights and opportunities with synchrotron light. Front. Pharmacol. 2014, 5, 191. [CrossRef]
- Visanji, N.P.; Collingwood, J.F.; Finnegan, M.E.; Tandon, A.; House, E.; Hazrati, L.-N. Iron Deficiency in Parkinsonism: Region-Specific Iron Dysregulation in Parkinson's Disease and Multiple System Atrophy. J. Park. Dis. 2013, 3, 523–537. [CrossRef]
- Hackett, M.J.; McQuillan, J.A.; El-Assaad, F.; Aitken, J.B.; Levina, A.; Cohen, D.D.; Siegele, R.; Carter, E.A.; Grau, G.E.; Hunt, N.H.; et al. Chemical alterations to murine brain tissue induced by formalin fixation: implications for biospectroscopic imaging and mapping studies of disease pathogenesis. Anal. 2011, 136, 2941–2952. [CrossRef]
- Schrag, M.; Dickson, A.; Jiffry, A.; Kirsch, D.; Vinters, H.V.; Kirsch, W. The effect of formalin fixation on the levels of brain transition metals in archived samples. BioMetals 2010, 23, 1123–1127. [CrossRef]
- Perls, M. Nachweis von Eisenoxyd in gewissen Pigmenten. Archiv F Pathol Anat 1867, 39, 42–48. [CrossRef]
- Moos, T. Immunohistochemical localization of intraneuronal transferrin receptor immunoreactivity in the adult mouse central nervous system. J Comp Neurol 1996, 375, 675-692. [CrossRef]
- Moos, T.; Nielsen, T.R.; Skjørringe, T.; Morgan, E.H. Iron trafficking inside the brain. J. Neurochem. 2007, 103, 1730–1740. [CrossRef]
- McRae, R.; Bagchi, P.; Sumalekshmy, S.; Fahrni, C.J. In Situ Imaging of Metals in Cells and Tissues. Chem. Rev. 2009, 109, 4780–4827. [CrossRef]
- Brooks, J.; Everett, J.; Lermyte, F.; Tjhin, V.T.; Banerjee, S.; O'Connor, P.B.; Morris, C.M.; Sadler, P.J.; Telling, N.D.; Collingwood, J.F. Label-Free Nanoimaging of Neuromelanin in the Brain by Soft X-ray Spectromicroscopy. Angew. Chem. Int. Ed. Engl. 2020, 59, 11984–11991. [CrossRef]
- Crichton, R.R.; Dexter, D.T.; Ward, R.J. Brain iron metabolism and its perturbation in neurological diseases; Springer: 2012.
- Everett, J.; Lermyte, F.; Brooks, J.; Tjendana-Tjhin, V.; Plascencia-Villa, G.; Hands-Portman, I.; Donnelly, J.M.; Billimoria, K.; Perry, G.; Zhu, X. Biogenic metallic elements in the human brain? Sci Adv 2021, 7, eabf6707.
- Pezacki, A.T.; Gao, J.; Chang, C.J. Designing small-molecule and macromolecule sensors for imaging redox-active transition metal signaling. Curr. Opin. Chem. Biol. 2024, 83, 102541. [CrossRef]
- Pezacki, A.T.; Gonciarz, R.L.; Okamura, T.; Matier, C.D.; Torrente, L.; Cheng, K.; Miller, S.G.; Ralle, M.; Ward, N.P.; DeNicola, G.M.; et al. A tandem activity-based sensing and labeling strategy reveals antioxidant response element regulation of labile iron pools. Proc. Natl. Acad. Sci.USA 2024, 121, e2401579121. [CrossRef]
- Wu, Y.; Torabi, S.-F.; Lake, R.J.; Hong, S.; Yu, Z.; Wu, P.; Yang, Z.; Nelson, K.; Guo, W.; Pawel, G.T.; et al. Simultaneous Fe 2+ /Fe 3+ imaging shows Fe 3+ over Fe 2+ enrichment in Alzheimer’s disease mouse brain. Sci. Adv. 2023, 9, eade7622. [CrossRef]
- Collingwood, J.F.; Adams, F. Chemical imaging analysis of the brain with X-ray methods. Spectrochim. Acta Part B: At. Spectrosc. 2017, 130, 101–118. [CrossRef]
- Brooks, J.; Everett, J.; Hill, E.; Billimoria, K.; Morris, C.M.; Sadler, P.J.; Telling, N.; Collingwood, J.F. Nanoscale synchrotron x-ray analysis of intranuclear iron in melanised neurons of Parkinson’s substantia nigra. Commun. Biol. 2024, 7, 1–7. [CrossRef]
- Telling, N.D.; Everett, J.; Collingwood, J.F.; Dobson, J.; van der Laan, G.; Gallagher, J.J.; Wang, J.; Hitchcock, A.P. Iron Biochemistry is Correlated with Amyloid Plaque Morphology in an Established Mouse Model of Alzheimer's Disease. Cell Chem. Biol. 2017, 24, 1205–1215.e3. [CrossRef]
- Lermyte, F.; Everett, J.; Brooks, J.; Bellingeri, F.; Billimoria, K.; Sadler, P.J.; O’connor, P.B.; Telling, N.D.; Collingwood, J.F. Emerging Approaches to Investigate the Influence of Transition Metals in the Proteinopathies. Cells 2019, 8, 1231. [CrossRef]
- Hayflick, S.J.; Kurian, M.A.; Hogarth, P. Neurodegeneration with brain iron accumulation. Handb Clin Neurol 2018, 147, 293-305.
- Meyer, E.; Kurian, M.A.; Hayflick, S.J. Neurodegeneration with Brain Iron Accumulation: Genetic Diversity and Pathophysiological Mechanisms. Annu. Rev. Genom. Hum. Genet. 2015, 16, 257–279. [CrossRef]
- Kolarova, H.; Tan, J.; Strom, T.M.; Meitinger, T.; Wagner, M.; Klopstock, T. Lifetime risk of autosomal recessive neurodegeneration with brain iron accumulation (NBIA) disorders calculated from genetic databases. EBioMedicine 2022, 77, 103869. [CrossRef]
- Svetel, M.; Dragasevic, N.; Petrovic, I.; Novakovic, I.; Tomic, A.; Kresojevic, N.; Stankovic, I.; Kostic, V. NBIA syndromes: a step forward from the previous knowledge. Neurol India 2021, 69, 1380-1388.
- Kumar, N.; Rizek, P.; Jog, M. Neuroferritinopathy: pathophysiology, presentation, differential diagnoses and management. Tremor Other Hyperkinet Mov (N Y) 2016, 6, 355.
- Levi, S.; Rovida, E. Neuroferritinopathy: From ferritin structure modification to pathogenetic mechanism. Neurobiol. Dis. 2015, 81, 134–143. [CrossRef]
- Chinnery, P.F.; Crompton, D.E.; Birchall, D.; Jackson, M.J.; Coulthard, A.; Lombès, A.; Quinn, N.; Wills, A.; Fletcher, N.; Mottershead, J.P.; et al. Clinical features and natural history of neuroferritinopathy caused by the FTL1 460InsA mutation. Brain 2006, 130, 110–119. [CrossRef]
- Kurzawa-Akanbi, M.; Keogh, M.; Tsefou, E.; Ramsay, L.; Johnson, M.; Keers, S.; Ochieng, L.W.; McNair, A.; Singh, P.; Khan, A.; et al. Neuropathological and biochemical investigation of Hereditary Ferritinopathy cases with ferritin light chain mutation: Prominent protein aggregation in the absence of major mitochondrial or oxidative stress. Neuropathol. Appl. Neurobiol. 2020, 47, 26–42. [CrossRef]
- Keita, M.; McIntyre, K.; Rodden, L.N.; Schadt, K.; Lynch, D.R. Friedreich Ataxia: Clinical Features and New Developments. Neurodegener. Dis. Manag. 2022, 12, 267–283. [CrossRef]
- Campuzano, V.; Montermini, L.; Moltò, M.D.; Pianese, L.; Cossée, M.; Cavalcanti, F.; Monros, E.; Rodius, F.; Duclos, F.; Monticelli, A.; et al. Friedreich’s Ataxia: Autosomal Recessive Disease Caused by an Intronic GAA Triplet Repeat Expansion. Science 1996, 271, 1423–1427. [CrossRef]
- Monfort, B.; Want, K.; Gervason, S.; D’autréaux, B. Recent Advances in the Elucidation of Frataxin Biochemical Function Open Novel Perspectives for the Treatment of Friedreich’s Ataxia. Front. Neurosci. 2022, 16, 838335. [CrossRef]
- Koutnikova, H.; Campuzano, V.; Foury, F.; Dollé, P.; Cazzalini, O.; Koenig, M. Studies of human, mouse and yeast homologues indicate a mitochondrial function for frataxin. Nat. Genet. 1997, 16, 345–351. [CrossRef]
- Nair, M.; Adinolfi, S.; Pastore, C.; Kelly, G.; Temussi, P.; Pastore, A. Solution Structure of the Bacterial Frataxin Ortholog, CyaY: Mapping the iron binding sites. Structure 2004, 12, 2037–2048. [CrossRef]
- Koeppen, A.H.; Michael, S.C.; Knutson, M.D.; Haile, D.J.; Qian, J.; Levi, S.; Santambrogio, P.; Garrick, M.D.; Lamarche, J.B. The dentate nucleus in Friedreich’s ataxia: the role of iron-responsive proteins. Acta Neuropathol. 2007, 114, 163–173. [CrossRef]
- Todorich, B.; Zhang, X.; Connor, J.R. H-ferritin is the major source of iron for oligodendrocytes. Glia 2011, 59, 927–935. [CrossRef]
- Bartzokis, G. Age-related myelin breakdown: a developmental model of cognitive decline and Alzheimer’s disease. Neurobiol. Aging 2004, 25, 5–18. [CrossRef]
- Nasrabady, S.E.; Rizvi, B.; Goldman, J.E.; Brickman, A.M. White matter changes in Alzheimer’s disease: a focus on myelin and oligodendrocytes. Acta Neuropathol. Commun. 2018, 6, 1–10. [CrossRef]
- Honig, L.S.; Sun, Y.; Irizarry, M.C.; Swanson, C.J.; Dhadda, S.; Charil, A.; E Hart, D.; Noble, J.M.; Huey, E.D.; Teich, A.F. Neuropathological autopsy findings in an individual with Alzheimer’s disease who received long-term treatment with lecanemab (BAN2401). Alzheimer's Dement. 2022, 18. [CrossRef]
- Solopova, E.; Romero-Fernandez, W.; Harmsen, H.; Ventura-Antunes, L.; Wang, E.; Shostak, A.; Maldonado, J.; Donahue, M.J.; Schultz, D.; Coyne, T.M.; et al. Fatal iatrogenic cerebral β-amyloid-related arteritis in a woman treated with lecanemab for Alzheimer’s disease. Nat. Commun. 2023, 14, 8220. [CrossRef]
- Reish, N.J.; Jamshidi, P.; Stamm, B.; Flanagan, M.E.; Sugg, E.; Tang, M.; Donohue, K.L.; McCord, M.; Krumpelman, C.; Mesulam, M.-M.; et al. Multiple Cerebral Hemorrhages in a Patient Receiving Lecanemab and Treated with t-PA for Stroke. New Engl. J. Med. 2023, 388, 478–479. [CrossRef]
- Mascalchi, M.; Salvi, F.; Pirini, M.; D’errico, A.; Ferlini, A.; Lolli, F.; Plasmati, R.; Tessa, C.; Villari, N.; Tassinari, C. Transthyretin amyloidosis and superficial siderosis of the CNS. Neurology 1999, 53, 1498–1503. [CrossRef]
- Ziskin, J.L.; Greicius, M.D.; Zhu, W.; Okumu, A.N.; Adams, C.M.; Plowey, E.D. Neuropathologic analysis of Tyr69His TTR variant meningovascular amyloidosis with dementia. Acta Neuropathol. Commun. 2015, 3, 1–7. [CrossRef]
- Episkopou, V.; Maeda, S.; Nishiguchi, S.; Shimada, K.; A Gaitanaris, G.; E Gottesman, M.; Robertson, E.J. Disruption of the transthyretin gene results in mice with depressed levels of plasma retinol and thyroid hormone.. Proc. Natl. Acad. Sci. 1993, 90, 2375–2379. [CrossRef]
- Monk, J.A.; Sims, N.A.; Dziegielewska, K.M.; Weiss, R.E.; Ramsay, R.G.; Richardson, S.J. Delayed development of specific thyroid hormone-regulated events in transthyretin null mice. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E23–E31. [CrossRef]
- Richardson, S.J. Cell and molecular biology of transthyretin and thyroid hormones. Int Rev Cytol 2007, 258, 137-193. [CrossRef]
- Alshehri, B.; Pagnin, M.; Lee, J.Y.; Petratos, S.; Richardson, S.J. The Role of Transthyretin in Oligodendrocyte Development. Sci. Rep. 2020, 10, 4189. [CrossRef]
- Vancamp, P.; Gothié, J.-D.; Luongo, C.; Sébillot, A.; Le Blay, K.; Butruille, L.; Pagnin, M.; Richardson, S.J.; Demeneix, B.A.; Remaud, S. Gender-specific effects of transthyretin on neural stem cell fate in the subventricular zone of the adult mouse. Sci. Rep. 2019, 9, 19689. [CrossRef]
- Ahmed, Z.; Asi, Y.T.; Lees, A.J.; Revesz, T.; Holton, J.L. Identification and Quantification of Oligodendrocyte Precursor Cells in Multiple System Atrophy, Progressive Supranuclear Palsy and Parkinson's Disease. Brain Pathol. 2012, 23, 263–273. [CrossRef]
- Murthy, M.; Fodder, K.; Miki, Y.; Rambarack, N.; De Pablo Fernandez, E.; Pihlstrøm, L.; Mill, J.; Warner, T.T.; Lashley, T.; Bettencourt, C. DNA methylation patterns in the frontal lobe white matter of multiple system atrophy, Parkinson’s disease, and progressive supranuclear palsy: a cross-comparative investigation. Acta Neuropathol. 2024, 148, 1–20. [CrossRef]
- Miki, Y.; Tsushima, E.; Foti, S.C.; Strand, K.M.; Asi, Y.T.; Yamamoto, A.K.; Bettencourt, C.; Oliveira, M.C.B.; De Pablo-Fernández, E.; Jaunmuktane, Z.; et al. Identification of multiple system atrophy mimicking Parkinson’s disease or progressive supranuclear palsy. Brain 2021, 144, 1138–1151. [CrossRef]
- Ozawa, T.; Paviour, D.; Quinn, N.P.; Josephs, K.A.; Sangha, H.; Kilford, L.; Healy, D.G.; Wood, N.W.; Lees, A.J.; Holton, J.L.; et al. The spectrum of pathological involvement of the striatonigral and olivopontocerebellar systems in multiple system atrophy: clinicopathological correlations. Brain 2004, 127, 2657–2671. [CrossRef]
- Piras, I.S.; Bleul, C.; Schrauwen, I.; Talboom, J.; Llaci, L.; De Both, M.D.; Naymik, M.A.; Halliday, G.; Bettencourt, C.; Holton, J.L.; et al. Transcriptional profiling of multiple system atrophy cerebellar tissue highlights differences between the parkinsonian and cerebellar sub-types of the disease. Acta Neuropathol. Commun. 2020, 8, 1–20. [CrossRef]
- Kaindlstorfer, C.; Jellinger, K.A.; Eschlböck, S.; Stefanova, N.; Weiss, G.; Wenning, G.K. The Relevance of Iron in the Pathogenesis of Multiple System Atrophy: A Viewpoint. J. Alzheimer's Dis. 2018, 61, 1253–1273. [CrossRef]
- Fodder, K.; de Silva, R.; Warner, T.T.; Bettencourt, C. The contribution of DNA methylation to the (dys)function of oligodendroglia in neurodegeneration. Acta Neuropathol. Commun. 2023, 11, 1–15. [CrossRef]
- Bettencourt, C.; Foti, S.C.; Miki, Y.; Botia, J.; Chatterjee, A.; Warner, T.T.; Revesz, T.; Lashley, T.; Balazs, R.; Viré, E.; et al. White matter DNA methylation profiling reveals deregulation of HIP1, LMAN2, MOBP, and other loci in multiple system atrophy. Acta Neuropathol. 2019, 139, 135–156. [CrossRef]
- Bettencourt, C.; Miki, Y.; Piras, I.S.; de Silva, R.; Foti, S.C.; Talboom, J.S.; Revesz, T.; Lashley, T.; Balazs, R.; Viré, E.; et al. MOBP and HIP1 in multiple system atrophy: New α-synuclein partners in glial cytoplasmic inclusions implicated in the disease pathogenesis. Neuropathol. Appl. Neurobiol. 2021, 47, 640–652. [CrossRef]
- Heidari, M.; Gerami, S.H.; Bassett, B.; Graham, R.M.; Chua, A.C.; Aryal, R.; House, M.J.; Collingwood, J.F.; Bettencourt, C.; Houlden, H.; et al. Pathological relationships involving iron and myelin may constitute a shared mechanism linking various rare and common brain diseases. Rare Dis. 2016, 4, e1198458. [CrossRef]
- Bettencourt, C.; Forabosco, P.; Wiethoff, S.; Heidari, M.; Johnstone, D.M.; Botía, J.A.; Collingwood, J.F.; Hardy, J.; Milward, E.A.; Ryten, M.; et al. Gene co-expression networks shed light into diseases of brain iron accumulation. Neurobiol. Dis. 2016, 87, 59–68. [CrossRef]
- Mifsud, G.; Zammit, C.; Muscat, R.; Di Giovanni, G.; Valentino, M. Oligodendrocyte Pathophysiology and Treatment Strategies in Cerebral Ischemia. CNS Neurosci. Ther. 2014, 20, 603–612. [CrossRef]
- Huang, S.; Ren, C.; Luo, Y.; Ding, Y.; Ji, X.; Li, S. New insights into the roles of oligodendrocytes regulation in ischemic stroke recovery. Neurobiol. Dis. 2023, 184, 106200. [CrossRef]
- Pantoni, L.; Garcia, J.H.; Gutierrez, J.A. Cerebral White Matter Is Highly Vulnerable to Ischemia. Stroke 1996, 27, 1641-1646; discussion 1647. [CrossRef]
- Sun, J.; Fang, Y.-Q.; Ren, H.; Chen, T.; Guo, J.-J.; Yan, J.; Song, S.; Zhang, L.-Y.; Liao, H. WIN55,212-2 protects oligodendrocyte precursor cells in stroke penumbra following permanent focal cerebral ischemia in rats. Acta Pharmacol. Sin. 2013, 34, 119–128. [CrossRef]
- Kishida, N.; Maki, T.; Takagi, Y.; Yasuda, K.; Kinoshita, H.; Ayaki, T.; Noro, T.; Kinoshita, Y.; Ono, Y.; Kataoka, H.; et al. Role of Perivascular Oligodendrocyte Precursor Cells in Angiogenesis After Brain Ischemia. J. Am. Hear. Assoc. 2019, 8, e011824. [CrossRef]
- Kyritsis, A.P.; Bondy, M.L.; Levin, V.A. Modulation of Glioma Risk and Progression by Dietary Nutrients and Antiinflammatory Agents. Nutr. Cancer 2011, 63, 174–184. [CrossRef]
- Torti, S.V.; Torti, F.M. Iron and cancer: more ore to be mined. Nat Rev Cancer 2013, 13, 342-355.
- Shenoy, G.; Connor, J.R. A closer look at the role of iron in glioblastoma. Neuro-Oncology 2023, 25, 2136–2149. [CrossRef]
- Troike, K.M.; Wang, S.Z.; Silver, D.J.; Lee, J.; E Mulkearns-Hubert, E.; Hajdari, N.; Ghosh, P.K.; E Kay, K.; Beilis, J.L.; E Mitchell, S.; et al. Homeostatic iron regulatory protein drives glioblastoma growth via tumor cell-intrinsic and sex-specific responses. Neuro-Oncology Adv. 2023, 6, vdad154. [CrossRef]
- Kerins, M.J.; Ooi, A. The Roles of NRF2 in Modulating Cellular Iron Homeostasis. Antioxid. Redox Signal. 2018, 29, 1756–1773. [CrossRef]
- Ma, Q.; Wang, X.; Li, J. LncRNA RP1-86C11.7 exacerbates the glioma progression and oncogenicity by hsa-miR-144-3p/TFRC signaling.. Transl. Oncol. 2021, 14, 101215. [CrossRef]
- Neilsen, B.K.; Sleightholm, R.; McComb, R.; Ramkissoon, S.H.; Ross, J.S.; Corona, R.J.; Miller, V.A.; Cooke, M.; Aizenberg, M.R. Comprehensive genetic alteration profiling in primary and recurrent glioblastoma. J Neurooncol 2019, 142, 111-118.
- Schonberg, D.L.; Lubelski, D.; Miller, T.E.; Rich, J.N. Brain tumor stem cells: Molecular characteristics and their impact on therapy. Mol. Asp. Med. 2014, 39, 82–101. [CrossRef]
- Schonberg, D.L.; Miller, T.E.; Wu, Q.; Flavahan, W.A.; Das, N.K.; Hale, J.S.; Hubert, C.G.; Mack, S.C.; Jarrar, A.M.; Karl, R.T.; et al. Preferential Iron Trafficking Characterizes Glioblastoma Stem-like Cells. Cancer Cell 2015, 28, 441–455. [CrossRef]
- Granholm, A.-C. Long-Term Effects of SARS-CoV-2 in the Brain: Clinical Consequences and Molecular Mechanisms. J. Clin. Med. 2023, 12, 3190. [CrossRef]
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: major findings, mechanisms and recommendations. Nat. Rev. Microbiol. 2023, 21, 133–146. [CrossRef]
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Author Correction: Long COVID: major findings, mechanisms and recommendations. Nat. Rev. Microbiol. 2023, 21, 408–408. [CrossRef]
- Reisbeck, L.; Linder, B.; Tascher, G.; Bozkurt, S.; Weber, K.J.; Herold-Mende, C.; van Wijk, S.J.L.; Marschalek, R.; Schaefer, L.; Münch, C.; et al. The iron chelator and OXPHOS inhibitor VLX600 induces mitophagy and an autophagy-dependent type of cell death in glioblastoma cells. Am. J. Physiol. Physiol. 2023, 325, C1451–C1469. [CrossRef]
- Jakobsson, A.W.; Kundu, S.; Guo, J.; Chowdhury, A.; Zhao, M.; Lindell, E.; Bergsten, P.; Swartling, F.J.; Sjöblom, T.; Zhang, X. Iron Chelator VLX600 Inhibits Mitochondrial Respiration and Promotes Sensitization of Neuroblastoma Cells in Nutrition-Restricted Conditions. Cancers 2022, 14, 3225. [CrossRef]
- Elincx-Benizri, S.; Glik, A.; Merkel, D.; Arad, M.; Freimark, D.; Kozlova, E.; Cabantchik, I.; Hassin-Baer, S. Clinical Experience With Deferiprone Treatment for Friedreich Ataxia. J. Child Neurol. 2016, 31, 1036–1040. [CrossRef]
- Pandolfo, M.; Arpa, J.; Delatycki, M.B.; Le Quan Sang, K.H.; Mariotti, C.; Munnich, A.; Sanz-Gallego, I.; Tai, G.; Tarnopolsky, M.A.; Taroni, F.; et al. Deferiprone in Friedreich ataxia: A 6-Month randomized controlled trial. Ann. Neurol. 2014, 76, 509–521. [CrossRef]
- Ayton, S.; Barton, D.; Brew, B.; Brodtmann, A.; Clarnette, R.; Desmond, P.; Devos, D.; Ellis, K.A.; Fazlollahi, A.; Fradette, C.; et al. Deferiprone in Alzheimer disease: a randomized clinical trial. JAMA Neurol 2025, 82, 11-18.
- Devos, D.; Labreuche, J.; Rascol, O.; Corvol, J.C.; Duhamel, A.; Guyon Delannoy, P.; Poewe, W.; Compta, Y.; Pavese, N.; Ruzicka, E.; et al. Trial of deferiprone in Parkinson’s disease. N Engl J Med 2022, 387, 2045-2055.
- Iankova, V.; Karin, I.; Klopstock, T.; Schneider, S.A. Emerging Disease-Modifying Therapies in Neurodegeneration With Brain Iron Accumulation (NBIA) Disorders. Front. Neurol. 2021, 12. [CrossRef]
- Klopstock, T.; Tricta, F.; Neumayr, L.; Karin, I.; Zorzi, G.; Fradette, C.; Kmieć, T.; Büchner, B.; E Steele, H.; Horvath, R.; et al. Safety and efficacy of deferiprone for pantothenate kinase-associated neurodegeneration: a randomised, double-blind, controlled trial and an open-label extension study. Lancet Neurol. 2019, 18, 631–642. [CrossRef]
- Wilson, R.B.; Lynch, D.R.; Fischbeck, K.H. Normal serum iron and ferritin concentrations in patients with Friedreich's ataxia. Ann. Neurol. 1998, 44, 132–134. [CrossRef]
- Currie, A. Science & Speculation. Erkenntnis 2021, 88, 597–619. [CrossRef]
Figure 2.
Iron-rich glia (Fe+glia) in wildtype AKR/J mouse brain at age 9 months. Small cells (diameter often <10μm) with morphological features often attributed to OLG (eccentric nuclei, few processes) often show intense cytoplasmic labeling with 3,3’ diaminobenzidine-enhanced Perls’ stain for iron [36], with little nuclei labeling. Photomicrography (Zeiss Axioscan) shows fine plexuses of criss-crossing fibers (A,B,D,E,G), as described by Hortega for OLG (Section 3.3.2). Interstitial Fe+glia (A-C) often make apparent contact with iron-rich, fibrous, myelin-like structures some distance away via very long fine processes that appear poorly suited for stable myelination and may instead be transient extensions for uni- or bi-directional transfer of iron or other substances between myelinated reservoirs and neurons or other cells. Some Fe+glia display mitotic figures consistent with proliferation (D,E). Juxta-vascular Fe+glia (F-H) are also seen, sometimes adjoining iron-rich myelin-containing structures (D). These occasionally appear to extend fine processes into the vessel lumen, consistent with uni- or bi-directional transfer of iron or other substances to or from blood (G,H).
Figure 2.
Iron-rich glia (Fe+glia) in wildtype AKR/J mouse brain at age 9 months. Small cells (diameter often <10μm) with morphological features often attributed to OLG (eccentric nuclei, few processes) often show intense cytoplasmic labeling with 3,3’ diaminobenzidine-enhanced Perls’ stain for iron [36], with little nuclei labeling. Photomicrography (Zeiss Axioscan) shows fine plexuses of criss-crossing fibers (A,B,D,E,G), as described by Hortega for OLG (Section 3.3.2). Interstitial Fe+glia (A-C) often make apparent contact with iron-rich, fibrous, myelin-like structures some distance away via very long fine processes that appear poorly suited for stable myelination and may instead be transient extensions for uni- or bi-directional transfer of iron or other substances between myelinated reservoirs and neurons or other cells. Some Fe+glia display mitotic figures consistent with proliferation (D,E). Juxta-vascular Fe+glia (F-H) are also seen, sometimes adjoining iron-rich myelin-containing structures (D). These occasionally appear to extend fine processes into the vessel lumen, consistent with uni- or bi-directional transfer of iron or other substances to or from blood (G,H).
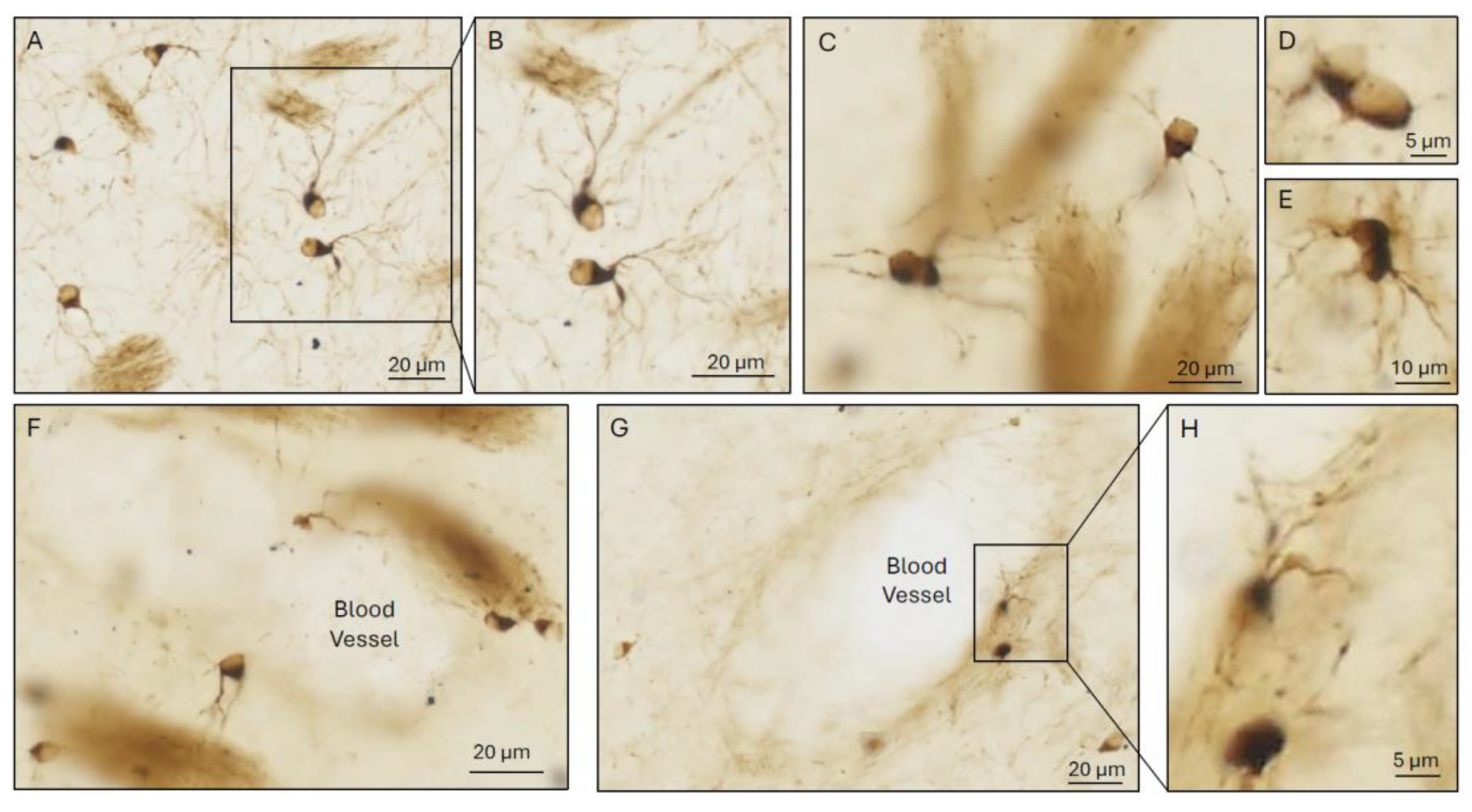
Figure 3.
The life story of iron. Top timeline traverses events after solar system formation ~4.6 billion years ago (Ga), including: emergence of key biomolecular building blocks of life (lipids, proteins, RNA, DNA) expedited by iron catalysis; last universal common ancestor (LUCA) – hypothetical single-celled organism with Fe-S cluster enzymes (Section 5.1.1.); Great Oxidation event (Section 4); conjectured LCA at key evolutionary divergence points e.g., emergence of multicellular Animalia ~700-800 million years ago (Ma) and Biliateria, coinciding with emergence of early neural systems, and other important milestones on the road to the human brain (Section 5.1.2 and Section 5.1.3). Time estimates are rounded to nearest 5 million years. Center timeline summarizes iron environments shaping terrestrial life, from iron-rich environment with superabundant reactive ferrous iron (Fe2+) that kickstarted vital bioreactions in early single-celled lifeforms, including bacteria, transitioning to iron-poor environment after mass oxidation of Fe2+ to insoluble ferric ion (Fe3+), with evolution of diverse systems for effective iron acquisition and utilization in organisms already intrinsically dependent on iron (Section 5.1.). Schema below shows milestones in iron management solutions and nervous system development, including: ferritin homologs - prototypes of modern human iron storage systems enabling iron stockpiling, some already under IRE control - and early Animalia (e.g., Porifera) with nascent nervous system-related transcriptomes; membranous vesicles; specialized cells initially regulating iron at the whole organism level (‘ferricytes’; Section 5.2.) and later regulating iron specifically within nervous systems (‘ferriglia’), parallel with emergence of ensheathing glia and preceding the advent of myelin (Section 5.3.). Time estimates from [101], OneZoom Tree of Life Explorer https://www.onezoom.org/ or as cited elsewhere. Images in figure are our own, public domain or icons created in https://BioRender.com.
Figure 3.
The life story of iron. Top timeline traverses events after solar system formation ~4.6 billion years ago (Ga), including: emergence of key biomolecular building blocks of life (lipids, proteins, RNA, DNA) expedited by iron catalysis; last universal common ancestor (LUCA) – hypothetical single-celled organism with Fe-S cluster enzymes (Section 5.1.1.); Great Oxidation event (Section 4); conjectured LCA at key evolutionary divergence points e.g., emergence of multicellular Animalia ~700-800 million years ago (Ma) and Biliateria, coinciding with emergence of early neural systems, and other important milestones on the road to the human brain (Section 5.1.2 and Section 5.1.3). Time estimates are rounded to nearest 5 million years. Center timeline summarizes iron environments shaping terrestrial life, from iron-rich environment with superabundant reactive ferrous iron (Fe2+) that kickstarted vital bioreactions in early single-celled lifeforms, including bacteria, transitioning to iron-poor environment after mass oxidation of Fe2+ to insoluble ferric ion (Fe3+), with evolution of diverse systems for effective iron acquisition and utilization in organisms already intrinsically dependent on iron (Section 5.1.). Schema below shows milestones in iron management solutions and nervous system development, including: ferritin homologs - prototypes of modern human iron storage systems enabling iron stockpiling, some already under IRE control - and early Animalia (e.g., Porifera) with nascent nervous system-related transcriptomes; membranous vesicles; specialized cells initially regulating iron at the whole organism level (‘ferricytes’; Section 5.2.) and later regulating iron specifically within nervous systems (‘ferriglia’), parallel with emergence of ensheathing glia and preceding the advent of myelin (Section 5.3.). Time estimates from [101], OneZoom Tree of Life Explorer https://www.onezoom.org/ or as cited elsewhere. Images in figure are our own, public domain or icons created in https://BioRender.com.
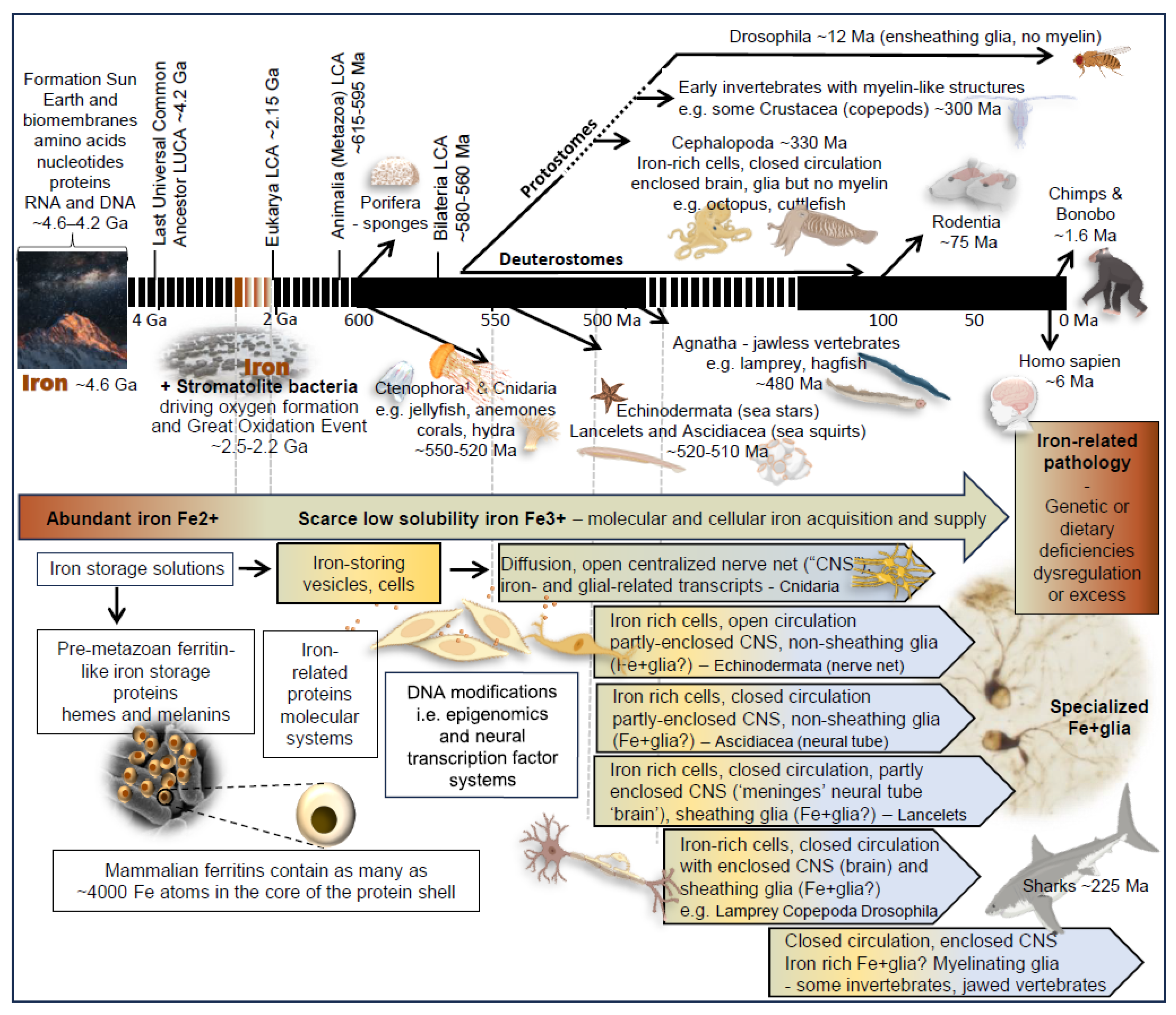
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



