
Submitted:
21 April 2025
Posted:
22 April 2025
You are already at the latest version
Abstract
Yellow leaf disease (YLD), caused by areca palm yellow leaf phytoplasma (AYLP), is a devastating disease that severely impacts the sustainable development of the areca palm industry. Efficient and accurate detection methods are considered crucial for the diagnosis and management of YLD. To address issues like false positive results using universal nested PCR primers, this study designed specific outer and internal primers based on the conserved regions of the 16S rDNA sequence of phytoplasmas, resulting in a nested PCR primer set, HNP-1F/HNP-1R and HNP-2F/HNP-2R. This set consistently amplified a single specific target band of 429 bp from 16SrI and 16SrII groups of AYLP. The sensitivity threshold for the 16SrI group of AYLP was 7.5×10-7 ng/μL, while that for the 16SrII group of AYLP was 4 ×10-3 ng/μL. To verify and evaluate the efficiency, the developed nested PCR system was used to detect leaf samples collected from trees showing leaf yellowing symptoms in areca palm plantations, and the results showed that the primer set could specifically detect AYLP. This primer set is characterized by rapid and accurate detection, high specificity, and high sensitivity compared to the traditional universal primer set of P1/P7 and R16mF2/R16mR1 or R16mF2/R16mR1 and R16mF2n/R2, providing a scientific basis for the specific diagnosis and early control of yellow leaf disease of areca palm.
Keywords:
areca palm (Areca catechu L.)
; yellow leaf disease of areca palm
; areca palm yellow leaf phytoplasma
; nested PCR
; molecular detection
1. Introduction
Areca palm (Areca catechu L.) is a perennial evergreen tree belonging to the genus Areca in the family Palmaceae. It is native to Malaysia and now widely distributed across South and Southeast Asia including 16 countries and regions such as India and Malaysia [1]. In China, areca palm is mainly cultivated in Hainan and Taiwan, with smaller amounts grown in Yunnan, Guangdong, and Guangxi [2]. The Areca palm holds significant dietary and medicinal value, ranking the first among the "Four Main Southern Medical Plants" (areca palm Areca catechu L., Alpinia oxyphylla Miq., Amomum villosum Lour., and Morinda officinalis How.). Its flowers and fruits are rich in various essential nutrients and its fruits, seeds, peels, and flowers can be used medicinally to treat conditions such as beriberi, swelling, and digestive issues [3]. The fruit of the areca palm, arecanut, is economically valuable and, when chewed by customers, promotes salivation, generates heat, and relieves fatigue, making it a popular chewing product. Areca palm is one of Hainan’s economically important "Six Trees" crops (coconut Cocos nucifera L., areca palm, oil palm Elaeis guineensis Jacq., rubber tree Hevea brasiliensis Muell. Arg., Dalbergia odorifera T. Chen, and agaru Aquilaria agallocha (Lour.) Roxb) in agriculture, accounting for over 95% of the national planting area [4]. With rising consumer demand and prices in China, the planting area and output value of areca palm in Hainan have increased annually. Currently, areca palm has become one of the key economic sources for more than 2.3 million farmers in Hainan [5], playing a crucial role in the provincial agricultural and rural development and in increasing farmers' income and wealth.
Yellow leaf disease of areca palm (YLD) caused by areca palm yellow leaf phytoplasma (AYLP) is a lethal disease first reported in the central region of Kerala, India in 1914 [6]. This disease was also observed in China in 1981 [7] and Sri Lanka in 2015 [8]. Since its initial discovery in Tunchang County, Hainan Province, China, in 1981, YLD has spread to nearly all areca palm growing areas in Hainan, causing severe damage to the local areca palm industry [5,9]. Due to the lack of effective control measures in production, pathogen detection remains the primary method for managing this disease.
Three phytoplasmas groups or subgroups responsible for YLD have been identified in India: 16SrI-B, 16SrXI-B, and 16SrXVI [10,12]. In China, earlier researches had indicated that the phytoplasmas causing YLD belong to the 16SrI-B and 16SrI-G subgroups [13,14]. Recently, our research team discovered new phytoplasmas, specifically from the 16SrII and 16SrXXXII groups, in YLD samples collected from Hainan [15,16].
To date, various detection methods have been developed for the YLD-causing phytoplasmas, including electron microscopy [18], enzyme-linked immunosorbent assays (ELISA) [19], nested PCR [14,20], loop-mediated isothermal amplification (LAMP) [21], droplet digital PCR (ddPCR) [22], and quantitative real-time PCR (qPCR) [10,23]. However, each of these methods has its advantages and limitations. For instance, electron microscopy assays require complex sample preparation, are time-consuming, and costly. Serological assays may face challenges in preparing antisera and risk of cross-reactivity with host antigens. Those molecular methods developed recently were used to detect AYLP in 16SrI group. Meanwhile, the LAMP technique, while capable of detection at a constant temperature, has a high primer design complexity [24] and is prone to contamination, leading to false positives. Compared to conventional PCR, qPCR and ddPCR involve higher reagent costs, more stringent laboratory conditions, and expensive equipment. Additionally, products from these methods (ELISA, LAMP, and qPCR) cannot be sequenced. In contrast, amplified products of nested PCR using universal phytoplasma primers can be sequenced, making it widely used for phytoplasma detection and classification research. However, nested PCR using universal phytoplasma primers may sometimes result in false-positive outcomes in the amplification of 16S rDNA sequences, significantly affecting detection efficiency [25]. In recent research, our team developed nested PCR primer sets F4/R1 and F2/R2 [20] to address detection challenges posed by the low titer and uneven distribution of AYLP in areca palm plants [4,26]. Though this primer set significantly improved the detection efficiency than universal primer sets like P1/P7 and R16mF2/R16mR1 or P1/P7 and R16mF2n/R2; subsequent studies showed that it, just like the universal primers, sometimes amplified non-specific fragments too (our lab’s unpublished data). In addition, research indicate that the 16SrI group is widely distributed in Hainan Province [17] and that there are many of cases where the 16SrII group has been identified in Wenchang City and Danzhou City (our unpublished data) , while only once case of the 16SrXXXII groups was observed [16]. Currently, there is no diagnostic technique available that can simultaneously detect both 16SrI and 16SrII groups of AYLP. To resolve this issue of non-specific amplification and to meet the needs for the detection of 16SrI group AYLP and novel identified 16SrII group AYLP, this study developed a nested PCR detection system comprising two specific primer pairs of four candidates, based on the 16S rDNA sequences of 16SrI and 16SrII group AYLPs and other areca palm-related pathogens. The goal is to establish a new specific nested PCR primers combination for accurate detection of AYLPs (16SrI and 16SrII groups) in Hainan, providing technical support for AYLP detection, identification, and early prevention of YLD in areca palm seedlings and infected plants in the field.
2. Results
2.1. Application of Universal Nested PCR for Phytoplasma Detection
A total of 335 areca palm genomic DNA samples were collectively tested using the universal nested PCR primer set targeting phytoplasmas. Among these samples, 50 exhibited amplification bands of approximately 1400 bp (Figure 1). The resulting PCR products from these 50 positive samples were subsequently sequenced by Sangon Biotech (Shanghai, China). The sequencing data revealed the following distribution: among the 50 samples, 16 samples showed amplification specific to areca chloroplast, 20 exhibited amplification bands corresponding to bacteria, only 10 displayed specific amplification related to phytoplasmas, and the remaining four samples yielded no detectable sequencing results (Figure 2).
2.2. Nested PCR Primer Test and Combination
By performing multiple sequence alignments of 16S rDNA from areca palm yellow leaf phytoplasma, areca chloroplasts, and endophytic bacteria, and following the principles of PCR primer design, we ultimately designed one pair of outer primer HNP-1F/R and three pairs of internal primers HNP-2F/2R, HNP-3F/3R, and HNP-4F/4R (Table 2).
These nested PCR primers were tested using genomic DNA samples from areca palm leaves infected with phytoplasma groups 16SrI and 16SrII, as well as genomic DNA samples from areca palm bacterial leaf blight pathogen and pineapple phytoplasma as references. Both HNP-2F/2R and HNP-3F/3R amplified target bands only in positive control and areca palm samples infected with phytoplasma groups 16SrI and 16SrII (Figure 3A and Figure 3B), indicating good performance. However, HNP-4F/4R also amplified a band of approximately 1500 bp in areca palm bacterial leaf blight pathogen DNA (Figure 3C). The combinations of HNP-1F/1R with either HNP-2F/2R or HNP-3F/3R were used to form a nested PCR in subsequent experiments.
2.3. Specificity Validation of Nested PCR
Using the outer primers HNP-1F/1R and internal primers HNP-2F/2R and HNP-3F/3R, the specificity of detection of areca palm DNA infected with phytoplasmas from the 16SrI and 16SrII groups, as well as DNA from five other pathogens was conducted. The results indicated that HNP-2F/2R only amplified a target band of approximately 429 bp in positive control and in areca palm samples infected with phytoplasmas from the 16SrI and 16SrII groups (Figure 4A). However, HNP-3F/3R, in addition to the three mentioned samples, also amplified a 652bp band in healthy areca palm samples and samples infected with phytoplasmas from the 16SrXXXII group (Figure 4B). Consequently, HNP-2F/2R was selected as the internal primer set for subsequent experiments.
2.4. Optimization of PCR Annealing Temperatures
In the nested PCR reaction, the annealing temperatures for both the outer and internal primers were systematically optimized within the range of 40°C to 60°C. For the outer primers targeting HNP-1F/1R, there was no significant difference in band intensity between annealing temperatures of 40.0°C to 53.6°C. However, as the annealing temperature increased from 56.0°C to 60.0°C, the band intensity decreased (Figure 5A). Therefore, we selected 53.6°C as the optimal annealing temperature.
Regarding the internal primers targeting HNP-2F/2R, a gradient annealing temperature approach was employed. Bands corresponding to the target were consistently visible within the temperature range of 46.4°C to 51.3°C, while no amplification occurred at 60.0°C (Figure 5B). Consequently, we determined the annealing temperature for HNP-2F/2R to be 51.3°C.
2.5. Sensitivity Determination of Nested PCR
To assess the sensitivity of this method for detecting the two phytoplasma groups, nested PCR using the outer primers HNP-1F/1R and internal primers HNP-2F/2R was conducted on recombinant plasmids containing 16S rDNA segments from the 16SrI and 16SrII phytoplasma groups.
Stable amplification of a single specific target band was achieved for the 16SrI group across plasmid concentrations ranging from 7.5 ng/μL to 7.5 × 10-7 ng/μL. However, as the concentration decreased, the amplification band gradually became fainter and disappeared at 7.5 × 10-7 ng/μL (Figure 6A). Similarly, for the 16SrII group, a single specific target band (429 bp) was consistently yielded across plasmid concentrations ranging from 4 ng/μL to 4 × 10-3 ng/μL. The amplification band intensity decreased with decreasing plasmid concentration and disappeared at 4 × 10-3 ng/μL (Figure 6B).
These results indicate that the nested PCR method has a minimum detection limit of 7.5 × 10-7 ng/μL for the 16SrI phytoplasma group and 4 × 10-3 ng/μL for the 16SrII phytoplasma group.
2.6. Comparison of The Newly Developed Nested PCR with Universal Nested PCR
To compare the detection efficiency of the nested PCR method developed in this study with that of the universal nested PCR, 30 field-collected areca samples were tested.
The universal nested PCR method failed to detect phytoplasma, except in the positive control (Figure 7A). In contrast, our newly developed nested PCR method successfully detected phytoplasma in 10 of the tested areca samples (Figure 7B). Sequencing confirmed that all 10 PCR products corresponded to the 16SrI group phytoplasma. This study highlights the effectiveness of our specific nested PCR approach for detecting AYLP and provides valuable insights for routine diagnosis of yellow leaf disease of areca palm research.
2.7. Construction and Analysis of Phylogenetic Trees
A phylogenetic tree was constructed using sequences of representative strains from the phytoplasma groups/subgroups, based on the respective amplified fragments of R16mF2/R1 and HNP-2F/2R, using MEGA 11 software. In both phylogenetic trees, phytoplasma of the 16SrI and 16SrII groups were each clustered into a separate branch. Additionally, the phylogenetic tree constructed from the sequence fragments from HNP-2F/2R showed differences in the subdivision of 16SrI subgroups compared to the tree from the R16mF2/R1 sequence fragments. This indicates that the nested PCR method established in this study can identify and group the phytoplasmas associated with YLD in the 16SrI and 16SrII groups but cannot differentiate at the subgroup level within the 16SrI group (Figures 8A and Figure 8B). The multiple sequence alignment also indicates that the fragments amplified by HNP-2F/2R exhibit high similarity to the 16SrI group AYLP (Figure 8C).
3. Discussion
Phytoplasmas, originally known as mycoplasma-like organisms (MLOs) [27], belong to the prokaryote group without cell walls [28,29]. Phytoplasmas are prokaryotic microorganisms that specifically parasitize the sieve tube cells in the phloem of plants. Researches have shown that their titer within plants is uneven and varies with temperature [30,31]. Unfortunately, phytoplasmas are difficult to cultivate artificially [32], which hinders their systematic identification and classification using traditional isolation and culture methods commonly employed for other prokaryotic and eukaryotic organisms (such as plant pathogenic bacteria and fungi) [33,34]. Therefore, the establishment of specific and sensitive detection methods is crucial for the study of aetiology, epidemiology, and control of phytoplasma diseases, including areca palm yellow leaf disease. In this study, internal and outer primers based on the 16S rDNA specific regions of AYLP were designed. A primer set, HNP-1F/HNP-1R and HNP-2F/HNP-2R, was ultimately screened for specific amplification of AYLP from the novel designed primers. The annealing temperature was optimized, and then the optimal nested PCR system for detecting areca palm yellow leaf phytoplasma was established.
The universal primers P1/P7 and R16mF2/R1 are commonly used for detecting phytoplasmas from different hosts. However, their non-specific amplification is quite notable. In this study, these universal primers were used for the amplification of 16S rDNA of 335 areca palm leaf samples. The result showed that among the 50 samples that amplified with the target size bands (about 1400 bp), only 10 (20%) were ultimately confirmed as areca palm yellow leaf phytoplasma (AYLP) according to the results of sequences alignment; 16 (32%) were identified as areca palm chloroplast, and 20 (40%) were other bacteria, with the remaining samples showing no signal. This result indicated that the specificity of the universal primers P1/P7 and R16mF2/R1 for the amplification of AYLP is relatively poor, resulting in a high false-positive rate (72% to 80%). A similar result was found in the detection of the pathogenic phytoplasma in sisal purple leafroll disease [35]. In our preliminary studies during 2018 and 2019, 100 leaf samples were detected using universal primer sets P1/P7 and R16mF2/R16mR1, only one sample was successfully amplified, its sequence was verified as AYLP. To solve the problem of the low detection efficiency of AYLP, a nested PCR method had been developed with novel primer sets F4/R1 and F2/R2 [20] with a target band of 525 bp had been designed based on 16S rDNA gene sequences of AYLP. Though this primer set significantly improved detection efficiency (80 out of 482, 17%) than universal primer set P1/P7 and R16mF2/R16mR1 (1 out of 100, 1%), subsequent studies showed that this primer set also led to non-specific amplification of sequences (for example, Bacillus sp.) in the areca leaf and suspected vector insects (our lab unpublished data). The newly nested PCR detection method was developed by performing multiple sequence alignments of the 16S rDNA sequences of the reported areca palm yellow leaf phytoplasma and other bacteria. This approach was used to select specific regions of the phytoplasma for primer design and screening, ensuring high specificity. Specificity tests demonstrated that this method could detect the 16SrI and 16SrII groups of phytoplasma currently found in areca palms, while other groups of phytoplasma and other bacteria were not detected as positive.
In terms of detection sensitivity, the newly developed nested PCR detection method has a minimum detection limit of 16 copies/μL (7.5×10-7 ng/μL) for the 16SrI group of AYLP. In comparison, the existing TaqMan qPCR detection method has a limit of 1.16 copies/μL [23], and ddPCR has a limit of 0.07 copies/μL [22]. However, the nested PCR detection method developed in this study can also simultaneously detect the 16SrII group of phytoplasmas, with a minimum detection limit of 1.3×10³ copies/μL (4 ×10-3 ng/μL).
Regarding cost and detection efficacy, this newly established method is more cost-effective and less prone to contamination compared to qPCR and ddPCR. Additionally, all amplified fragments from the samples sequenced thus far have been identified as phytoplasmas. This method also allows for the construction of phylogenetic trees from the sequencing data to identify positive samples, a capability that current qPCR and ddPCR methods lack.
The nested PCR detection method developed in this study can be used for the detection and identification of areca palm yellow leaf phytoplasma (16SrI and 16SrII groups). It provides technical support for subsequent AYLP detection, identification, and early prevention and control of areca palm yellow leaf disease.
4. Materials and Methods
4.1. Materials
Areca leaf samples showing yellow symptoms collected from the field and stored in our laboratory were used in this study (Table 1).
4.2. Extraction of Total DNA from Areca Plam Leaf Samples Showing Yellow
Leaf samples of areca palm weighing approxima was 0.1 g were taken and cut into 1 mm pieces. DNA was extracted following the instructions provided by TianGen Biotech Co., Ltd. (Beijing, China), using the plant genomic DNA extraction kit, quantified using Nanodrop 2000, and stored at -20 °C for subsequent use.
4.3. Construction of Recombinant Plasmids for 16S rDNA of Areca Palm Yellow Leaf Phytoplasma
Yellow leaf disease (YLD) is a major disease affecting areca palm plants. Rapid detection of the pathogen responsible for YLD is crucial for effective management. In this study, universal nested PCR primers P1/P7 and R16mF2/R16mR1 (Table 2) targeting the 16S rDNA of phytoplasmas were employed to amplify the total DNA extracted from Areca leaf showing YLD. The reaction mixture consisted of 2 μL of DNA template, 12.5 μL of 2 × Taq PCR Master Mix (Aidlab Biotechnologies Co., Ltd, Beijing, China, PC0902), and 1 μL of each primer (10 μM), with ddH2O added to a final volume of 25 μL. The PCR procedure included an initial denaturation at 94 °C for 3 minutes, followed by 35 cycles of denaturation at 94 °C for 30 seconds, annealing at 55 °C for 30 seconds, extension at 72 °C for 1 minute, and a final extension at 72 °C for 5 minutes.
After the first round of PCR using external primers P1/P7, the product was diluted 20-fold, and a second round of amplification was performed using internal primers R16mF2/R16mR1. The procedure was like the first round, except for an annealing temperature of 50 °C. The amplified products were analyzed by electrophoresis on a 1% agarose gel (120 V, 35 minutes) and visualized using a gel imaging system (Synoptics Ltd, USA, GBOX-F3-LFB).
To construct recombinant plasmids, the PCR products were purified using a gel extraction kit (Tiangen Biotech, Beijing, China, DP209-02). Subsequently, a rapid ligation kit (Sangon Biotech, Shanghai, China, B522214-0020) was used to ligate the purified products into the T-vector. The resulting plasmids were transformed into DH5α competent cells (Sangon Biotech, Shanghai, China, B528413-0010). Positive clones were selected in LB sterile liquid medium supplemented with 50 ng/μL ampicillin, and plasmid DNA was extracted using a plasmid mini-prep kit (Tiangen Biotech, Beijing, China, DP103-02) and sequenced by Sangon Biotech (Shanghai, China).
4.4. Design and Primary Screening of Specific Primers
In this study, outer and internal primers were designed and screened for detecting the 16S rDNA of phytoplasmas associated with yellow leaf disease of areca palm (YLD). For this, previously reported 16S rDNA sequences from the 16SrI-B subgroup and the 16SrI-G subgroup of YLD phytoplasmas were utilized, along with the 16S rDNA sequence of phytoplasmas from the 16SrII subgroup, which was previously characterized in our laboratory. To enhance specificity, these 16S rDNA sequences were also aligned and compared with those from Areca chloroplasts, areca bacterial leaf spot pathogens, and other bacterial species (Table 3). Based on sequence variations, several nested PCR primers were designed and synthesized in Sangon Biotech (Shanghai, China).
In the first round of amplification, genomic DNA samples from areca palm leaves infected with phytoplasma groups 16SrI and 16SrII, as well as DNA of areca bacterial leaf blight pathogen (Burkholderia andropogonis) and Pantoea ananatis, were amplified using the primers HNP-1F/1R. Chrysanthemum genomic DNA infected with phytoplasma group 16SrI was used as a positive control, and H2O served as the negative control. The first-round PCR products were diluted 20 times, and three pairs of nested primers (HNP-2F/2R, HNP-3F/3R, and HNP-4F/4R) were used to amplify the first-round PCR products, aiming to select the better nested PCR primer combination.
4.5. Specificity Verification of Nested PCR
The specificity of the method developed in the current study was validated by using external primers HNP-1F/1R to amplify genomic DNA from various sources, including healthy areca palm leaves, areca palm bacterial leaf spot pathogens, pineapple pan-genus bacteria, YLD phytoplasmas (16SrI and 16SrII), and kenaf plants infected with 16SrXXXII phytoplasmas. Additionally, the complementary DNA was included from areca yellow leaf virus 1 (Table 1). After diluting the first round PCR product 20-fold and using it as a template, a second-round amplification was performed with the internal primers pairs mentioned above. The nested PCR reaction system and cycling conditions were identical to those described in section 1.3. The amplified products were analyzed by electrophoresis on a 1% agarose gel (120 V, 35 minutes) and visualized using a gel imaging system (Synoptics Ltd, USA, GBOX-F3-LFB).
4.6. Optimization of Annealing Temperatures for Nested PCR with Internal Primers
To optimize the annealing temperatures for the nested PCR using internal primers in the detection of phytoplasmas, the HNP-1F/1R + HNP-2F/2R primer set was utilized, targeting the positive Chrysanthemum phytoplasma genomic DNA samples (Table 1).
The first round of amplification using primers HNP-1F/1R, the reaction mixture was same as in section 1.3. The PCR cycling conditions included an initial denaturation at 94 °C for 3 minutes, followed by 35 cycles of denaturation at 94 °C for 30 seconds, annealing with a temperature gradient ranging from 40 °C to 60 °C (in 12 steps), annealing for 30 seconds, extension at 72 °C for 1 minute, and a final extension at 72 °C for 5 minutes.
After the completion of the first PCR round, the product was diluted 20-fold, and the second round of amplification using primers HNP-2F/2R was performed. The reaction mixture for the second round was the same as the first, with a temperature gradient ranging from 40 °C to 60 °C (also in 12 steps). The cycling conditions remained consistent, and the amplified products were analyzed by electrophoresis on a 1% agarose gel (120 V, 35 minutes) and visualized using a gel imaging system.
4.7. Establishment of Nested PCR Reaction System
In this study, a nested PCR reaction system using external primers HNP-1F/1R was established for the first round of amplification. After completing the first PCR round, the resulting product was diluted 20-fold, and the second round of amplification was performed using internal primers HNP-2F/2R. The mixture and PCR cycling condition for the two rounds of reactions were the same as in section 1.6, but the annealing temperature was determined to be 42.5 °C (while for the second PCR round, the annealing temperature was 46 °C).
4.8. Sensitivity Testing of Newly Developed Nested PCR
Sensitivity testing was conducted for the nested PCR developed in this study, using recombinant plasmids S97-1 and W4-1, each containing 16S rDNA segments from 16SrI and 16SrII of AYLP, respectively. These plasmids were subjected to a 10-fold serial dilution. S97-1 was diluted from 7.5 ng/μL to 7.5×10-8 ng/μL. W4-1 was diluted from 4 ng/μL to 400 zg/μL. The HNP-1F/1R + HNP-2F/2R nested PCR primer set was employed to amplify the diluted S97-1 and W4-1 recombinant plasmids. The nested PCR reaction system and cycling conditions were consistent with those described in section 1.6.
4.9. Comparison of Newly Developed Nested PCR Method with Universal Nested PCR Method
In this study, 30 samples of areca palm showing yellow symptoms were simultaneously tested to evaluate the performance of two detection methods using universal primers and the newly developed nested PCR primer set.
The universal nested PCR method followed the reaction steps described in section 1.3. The newly established nested PCR method followed the reaction steps outlined in section 1.6.
4.10. Phylogenetic Analysis of the Fragment Amplified from Primers HNP1F/1R and HNP-2F/2R
The sequence fragments obtained from the aforementioned sequencing and assembly process, along with 16S rDNA sequences of different groups of plant pathogens from GenBank, were used to construct a phylogenetic tree. The Neighbor-Joining (NJ) method in MEGA 11 was employed with a bootstrap value of 1000 [35].
Author Contributions
Conceptualization, Q.H.T. and X.Q.Z.; methodology, H.Y.G., Q.H.T. and X.Q.Z.; validation, H.Y.G.; formal analysis, H.Y.G.; in-vestigation, X.L.M, Z.W.L., W.W.S., S.J., and W.Q.Q.; data curation, H.Y.G., Q.H.T. and X.Q.Z.; writing—original draft preparation, H.Y.G.; writing—review and editing, H.Y.G., Q.H.T. and X.Q.Z.; review, Q.H.T. and X.Q.Z.; supervision, Q.H.T. and X.Q.Z.; and project administration, Q.H.T. and X.Q.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Hainan Key Project (ZDYF2025XDNY118 and ZDYF2022XDNY208) and the Project of Yazhouwan Scientific and Technological Administration of Sanya (SYND-2022-36).
Data Availability Statement
DNA sequences are available in the GenBank database, with the accession numbers listed in the Results. All other relevant data are within the paper and Supplementary Materials.
Acknowledgments
We sincerely thank Graduate student Wang Yuhang and Liu Yueyue, undergraduate interns Wang Huiqing, Xu Caide, Deng Ting and Zheng Xingxing participated in patial sample collections and DNA extraction from 2020 to 2022, and scientific research as-sistants Ma Luping, Ma Pei, Guo Chunping and Long Shida participated in patial DNA extraction during 2019 and 2024. 16SrI Group DNA sample of PT-1 from Paulownia sp. and 16SrIV Group DNA sample of ZF-1 from Jujube Ziziphus jujuba were generously provided by Dr. Gao Rui in Shandong Institute of Pomology of Shandong Academy of Agricultural Sciences. The oringinal manuscript was grammar proofreading by Dr Syed Majid of Rasheed Department of Agriculture Entomology Section, Bacha Khan University, Charsadda, Pakistan. We would like to express our heartfelt thanks to them. We also thank our laboratory partners for their technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| YLD | Yellow leaf disease of areca palm |
| AYLP | areca palm yellow leaf phytoplasma |
| LAMP | loop-mediated isothermal amplification |
| qPCR | quantitative real-time PCR |
| ddPCR | droplet digital PCR |
Appendix

Table A1.
Areca palm leaf samples used in the detection assays using novel nested PCR in this study.
Table A1.
Areca palm leaf samples used in the detection assays using novel nested PCR in this study.
| No | Sample | Result | City/County | Time | No | Sample | Result | City/County | Time |
|---|---|---|---|---|---|---|---|---|---|
| 1 | J069 | Positive | Wenchang | 2022 | 169 | ZB1-2 | Negative | Wenchang | 2022 |
| 2 | A056 | Positive | Wenchang | 2022 | 170 | MQR4 | Negative | Ledong | 2021 |
| 3 | T40 | Positive | Wenchang | 2022 | 171 | HL3-4 | Negative | Ledong | 2021 |
| 4 | W4 | Positive | Wenchang | 2022 | 172 | HL3-1 | Negative | Ledong | 2021 |
| 5 | S97 | Positive | Wenchang | 2022 | 173 | HL1-2 | Negative | Ledong | 2021 |
| 6 | M25 | Positive | Wenchang | 2023 | 174 | HZ3-2 | Negative | Ledong | 2021 |
| 7 | H61 | Positive | Wenchang | 2023 | 175 | HZ5-2 | Negative | Ledong | 2021 |
| 8 | H64 | Positive | Wenchang | 2023 | 176 | T52 | Negative | Ledong | 2021 |
| 9 | R71 | Positive | Waning | 2022 | 177 | NY1-2 | Negative | Wenchang | 2022 |
| 10 | R72 | Positive | Waning | 2022 | 178 | ZB2-2 | Negative | Wenchang | 2022 |
| 11 | R61 | Negative | Waning | 2022 | 179 | M22 | Negative | Wenchang | 2022 |
| 12 | R62 | Negative | Waning | 2022 | 180 | M23 | Negative | Wenchang | 2022 |
| 13 | R63 | Negative | Waning | 2022 | 181 | M25 | Negative | Wenchang | 2022 |
| 14 | R64 | Negative | Waning | 2022 | 182 | M26 | Negative | Wenchang | 2022 |
| 15 | R65 | Negative | Waning | 2022 | 183 | M27 | Negative | Wenchang | 2022 |
| 16 | R66 | Negative | Waning | 2022 | 184 | M30 | Negative | Wenchang | 2022 |
| 17 | R67 | Negative | Waning | 2022 | 185 | M31 | Negative | Wenchang | 2022 |
| 18 | R68 | Negative | Waning | 2022 | 186 | L30 | Negative | Wenchang | 2021 |
| 19 | R69 | Negative | Waning | 2022 | 187 | L32 | Negative | Wenchang | 2021 |
| 20 | R70 | Negative | Waning | 2022 | 188 | L33 | Negative | Wenchang | 2021 |
| 21 | R73 | Negative | Waning | 2022 | 189 | S40 | Negative | Wenchang | 2023 |
| 22 | R74 | Negative | Waning | 2022 | 190 | S41 | Negative | Wenchang | 2023 |
| 23 | R75 | Negative | Waning | 2022 | 191 | S42 | Negative | Wenchang | 2023 |
| 24 | R76 | Negative | Waning | 2022 | 192 | S43 | Negative | Wenchang | 2023 |
| 25 | R77 | Negative | Waning | 2022 | 193 | S44 | Negative | Wenchang | 2023 |
| 26 | R78 | Negative | Waning | 2022 | 194 | S45 | Negative | Wenchang | 2023 |
| 27 | R79 | Negative | Waning | 2022 | 195 | S46 | Negative | Wenchang | 2023 |
| 28 | R80 | Negative | Waning | 2022 | 196 | S47 | Negative | Wenchang | 2023 |
| 29 | R81 | Negative | Waning | 2022 | 197 | S50 | Negative | Wenchang | 2023 |
| 30 | R82 | Negative | Waning | 2022 | 198 | W11 | Negative | Wenchang | 2023 |
| 31 | R83 | Negative | Waning | 2022 | 199 | W12 | Negative | Wenchang | 2023 |
| 32 | R84 | Negative | Waning | 2022 | 200 | W13 | Negative | Wenchang | 2023 |
| 33 | R85 | Negative | Waning | 2022 | 201 | W14 | Negative | Wenchang | 2023 |
| 34 | R86 | Negative | Waning | 2022 | 202 | W15 | Negative | Wenchang | 2023 |
| 35 | R87 | Negative | Waning | 2022 | 203 | W16 | Negative | Wenchang | 2023 |
| 36 | R88 | Negative | Waning | 2022 | 204 | W17 | Negative | Wenchang | 2023 |
| 37 | R89 | Negative | Waning | 2022 | 205 | W18 | Negative | Wenchang | 2023 |
| 38 | R90 | Negative | Waning | 2022 | 206 | W19 | Negative | Wenchang | 2023 |
| 39 | W1 | Negative | Waning | 2022 | 207 | W20 | Negative | Wenchang | 2023 |
| 40 | W2 | Negative | Wenchang | 2022 | 208 | W21 | Negative | Wenchang | 2023 |
| 41 | W3 | Negative | Wenchang | 2022 | 209 | W22 | Negative | Wenchang | 2023 |
| 42 | W3-4 | Negative | Wenchang | 2022 | 210 | W23 | Negative | Wenchang | 2023 |
| 43 | A09 | Negative | Wenchang | 2022 | 211 | W24 | Negative | Wenchang | 2023 |
| 44 | A12 | Negative | Wenchang | 2022 | 212 | W25 | Negative | Wenchang | 2023 |
| 45 | A13 | Negative | Wenchang | 2022 | 213 | W26 | Negative | Wenchang | 2023 |
| 46 | A14 | Negative | Wenchang | 2022 | 214 | W27 | Negative | Wenchang | 2023 |
| 47 | A16 | Negative | Wenchang | 2022 | 215 | W28 | Negative | Wenchang | 2023 |
| 48 | A19 | Negative | Wenchang | 2022 | 216 | W29 | Negative | Wenchang | 2023 |
| 49 | A22 | Negative | Wenchang | 2022 | 217 | W30 | Negative | Wenchang | 2023 |
| 50 | A24 | Negative | Wenchang | 2022 | 218 | W31 | Negative | Wenchang | 2023 |
| 51 | A27 | Negative | Wenchang | 2022 | 219 | W32 | Negative | Wenchang | 2023 |
| 52 | A30 | Negative | Wenchang | 2022 | 220 | W33 | Negative | Wenchang | 2023 |
| 53 | A31 | Negative | Wenchang | 2022 | 221 | W34 | Negative | Wenchang | 2023 |
| 54 | A32 | Negative | Wenchang | 2022 | 222 | W35 | Negative | Wenchang | 2023 |
| 55 | A34 | Negative | Wenchang | 2022 | 223 | W36 | Negative | Wenchang | 2023 |
| 56 | A36 | Negative | Wenchang | 2022 | 224 | W37 | Negative | Wenchang | 2023 |
| 57 | A37 | Negative | Wenchang | 2022 | 225 | W38 | Negative | Wenchang | 2023 |
| 58 | A39 | Negative | Wenchang | 2022 | 226 | W39 | Negative | Wenchang | 2023 |
| 59 | A49 | Negative | Wenchang | 2022 | 227 | B2-1 | Negative | Baoting | 2021 |
| 60 | A50 | Negative | Wenchang | 2022 | 228 | B2-2 | Negative | Baoting | 2021 |
| 61 | A51 | Negative | Wenchang | 2022 | 229 | B2-4 | Negative | Baoting | 2021 |
| 62 | A55 | Negative | Wenchang | 2022 | 230 | B2-5 | Negative | Baoting | 2021 |
| 63 | A57 | Negative | Wenchang | 2022 | 231 | B2-6 | Negative | Baoting | 2021 |
| 64 | A59 | Negative | Wenchang | 2022 | 232 | B2-7 | Negative | Baoting | 2021 |
| 65 | A60 | Negative | Wenchang | 2022 | 233 | B2-9 | Negative | Baoting | 2021 |
| 66 | B13 | Negative | Baoting | 2020 | 234 | B4-1 | Negative | Baoting | 2021 |
| 67 | B4 | Negative | Baoting | 2020 | 235 | B4-2 | Negative | Baoting | 2021 |
| 68 | B17 | Negative | Baoting | 2020 | 236 | B4-3 | Negative | Baoting | 2021 |
| 69 | B20 | Negative | Baoting | 2020 | 237 | B4-4 | Negative | Baoting | 2021 |
| 70 | B21 | Negative | Baoting | 2020 | 238 | B4-5 | Negative | Baoting | 2021 |
| 71 | B23 | Negative | Baoting | 2020 | 239 | B4-6 | Negative | Baoting | 2021 |
| 72 | B24 | Negative | Baoting | 2020 | 240 | B4-7 | Negative | Baoting | 2021 |
| 73 | B25 | Negative | Baoting | 2020 | 241 | B4-8 | Negative | Baoting | 2021 |
| 74 | B27 | Negative | Baoting | 2020 | 242 | B4-9 | Negative | Baoting | 2021 |
| 75 | B30 | Negative | Baoting | 2020 | 243 | B4-10 | Negative | Baoting | 2021 |
| 76 | B31 | Negative | Baoting | 2020 | 244 | B4-11 | Negative | Baoting | 2021 |
| 77 | B32 | Negative | Baoting | 2020 | 245 | B4-12 | Negative | Baoting | 2021 |
| 78 | B33 | Negative | Baoting | 2020 | 246 | B4-13 | Negative | Baoting | 2021 |
| 79 | B37 | Negative | Baoting | 2020 | 247 | B4-14 | Negative | Baoting | 2021 |
| 80 | B41 | Negative | Baoting | 2020 | 248 | B4-15 | Negative | Baoting | 2021 |
| 81 | B44 | Negative | Baoting | 2020 | 249 | B4-16 | Negative | Baoting | 2021 |
| 82 | B46 | Negative | Baoting | 2020 | 250 | B4-17 | Negative | Baoting | 2021 |
| 83 | B47 | Negative | Baoting | 2020 | 251 | B4-18 | Negative | Baoting | 2021 |
| 84 | B49 | Negative | Baoting | 2020 | 252 | B4-19 | Negative | Baoting | 2021 |
| 85 | B53 | Negative | Baoting | 2020 | 253 | B5-4 | Negative | Baoting | 2021 |
| 86 | B54 | Negative | Baoting | 2020 | 254 | B5-5 | Negative | Baoting | 2021 |
| 87 | B56 | Negative | Baoting | 2020 | 255 | B5-6 | Negative | Baoting | 2021 |
| 88 | B57 | Negative | Baoting | 2020 | 256 | B5-10 | Negative | Baoting | 2021 |
| 89 | B61 | Negative | Baoting | 2020 | 257 | B6-1 | Negative | Baoting | 2021 |
| 90 | B63 | Negative | Baoting | 2020 | 258 | B6-2 | Negative | Baoting | 2021 |
| 91 | B76 | Negative | Baoting | 2020 | 259 | B6-4 | Negative | Baoting | 2021 |
| 92 | C21 | Negative | Qionghai | 2020 | 260 | B6-5 | Negative | Baoting | 2021 |
| 93 | C26 | Negative | Qionghai | 2020 | 261 | B6-6 | Negative | Baoting | 2021 |
| 94 | C27 | Negative | Qionghai | 2020 | 262 | B6-8 | Negative | Baoting | 2021 |
| 95 | C41 | Negative | Qionghai | 2020 | 263 | B6-10 | Negative | Baoting | 2021 |
| 96 | C47 | Negative | Qionghai | 2020 | 264 | B6-11 | Negative | Baoting | 2021 |
| 97 | C61 | Negative | Qionghai | 2020 | 265 | F04-2 | Negative | Tunchang | 2020 |
| 98 | C68 | Negative | Qionghai | 2020 | 266 | F05-2 | Negative | Tunchang | 2020 |
| 99 | C69 | Negative | Qionghai | 2020 | 267 | F05-1 | Negative | Tunchang | 2020 |
| 100 | C70 | Negative | Qionghai | 2020 | 268 | F06-1 | Negative | Tunchang | 2020 |
| 101 | C73 | Negative | Qionghai | 2020 | 269 | F07-1 | Negative | Tunchang | 2020 |
| 102 | C85 | Negative | Qionghai | 2020 | 270 | F022-1 | Negative | Tunchang | 2020 |
| 103 | C87 | Negative | Qionghai | 2020 | 271 | F028-2 | Negative | Tunchang | 2020 |
| 104 | C89 | Negative | Qionghai | 2020 | 272 | F023-2 | Negative | Tunchang | 2020 |
| 105 | C90 | Negative | Qionghai | 2020 | 273 | F035-1 | Negative | Tunchang | 2020 |
| 106 | C91 | Negative | Qionghai | 2020 | 274 | F036-1 | Negative | Tunchang | 2020 |
| 107 | C95 | Negative | Qionghai | 2020 | 275 | F036-2 | Negative | Tunchang | 2020 |
| 108 | C99 | Negative | Qionghai | 2020 | 276 | F037-1 | Negative | Tunchang | 2020 |
| 109 | C100 | Negative | Qionghai | 2020 | 277 | F037-2 | Negative | Tunchang | 2020 |
| 110 | BSL-1 | Negative | Ding'an | 2023 | 278 | F038-1 | Negative | Tunchang | 2020 |
| 111 | BSL-2 | Negative | Ding'an | 2023 | 279 | F038-2 | Negative | Tunchang | 2020 |
| 112 | BSL-3 | Negative | Ding'an | 2023 | 280 | F039-1 | Negative | Tunchang | 2020 |
| 113 | BSL-4 | Negative | Ding'an | 2023 | 281 | F040-1 | Negative | Tunchang | 2020 |
| 114 | BSL-5 | Negative | Ding'an | 2023 | 282 | F040-2 | Negative | Tunchang | 2020 |
| 115 | BSL-6 | Negative | Ding'an | 2023 | 283 | F041-2 | Negative | Tunchang | 2020 |
| 116 | BSL-7 | Negative | Ding'an | 2023 | 284 | F044-1 | Negative | Tunchang | 2020 |
| 117 | BSL-8 | Negative | Ding'an | 2023 | 285 | F044-2 | Negative | Tunchang | 2020 |
| 118 | BSL-9 | Negative | Ding'an | 2023 | 286 | F045-1 | Negative | Tunchang | 2020 |
| 119 | BSL-10 | Negative | Ding'an | 2023 | 287 | F045-2 | Negative | Tunchang | 2020 |
| 120 | BSL-11 | Negative | Ding'an | 2023 | 288 | F049-2 | Negative | Tunchang | 2020 |
| 121 | BSL-12 | Negative | Ding'an | 2023 | 289 | F055-2 | Negative | Tunchang | 2020 |
| 122 | BSL-13 | Negative | Ding'an | 2023 | 290 | F099-1 | Negative | Tunchang | 2020 |
| 123 | BSL-14 | Negative | Ding'an | 2023 | 291 | C002-1 | Negative | Qionghai | 2020 |
| 124 | BSL-15 | Negative | Ding'an | 2023 | 292 | C007-2 | Negative | Qionghai | 2020 |
| 125 | BSL-16 | Negative | Ding'an | 2023 | 293 | C013-2 | Negative | Qionghai | 2020 |
| 126 | BSL-17 | Negative | Ding'an | 2023 | 294 | C018Y-1 | Negative | Qionghai | 2020 |
| 127 | BSL-18 | Negative | Ding'an | 2023 | 295 | C018Y-2 | Negative | Qionghai | 2020 |
| 128 | BSL-19 | Negative | Ding'an | 2023 | 296 | C021-1 | Negative | Qionghai | 2020 |
| 129 | BSL-20 | Negative | Ding'an | 2023 | 297 | C023-1 | Negative | Qionghai | 2020 |
| 130 | XS-1 | Negative | Ding'an | 2023 | 298 | C026-1 | Negative | Qionghai | 2020 |
| 131 | XS-2 | Negative | Ding'an | 2023 | 299 | C026-2 | Negative | Qionghai | 2020 |
| 132 | XS-3 | Negative | Ding'an | 2023 | 300 | C031-2 | Negative | Qionghai | 2020 |
| 133 | XS-4 | Negative | Ding'an | 2023 | 301 | C048-1 | Negative | Qionghai | 2020 |
| 134 | XS-5 | Negative | Ding'an | 2023 | 302 | C064-1 | Negative | Qionghai | 2020 |
| 135 | XS-6 | Negative | Ding'an | 2023 | 303 | C065-2 | Negative | Qionghai | 2020 |
| 136 | XS-7 | Negative | Ding'an | 2023 | 304 | C071-2 | Negative | Qionghai | 2020 |
| 137 | XS-8 | Negative | Ding'an | 2023 | 305 | C072-2 | Negative | Qionghai | 2020 |
| 138 | XS-9 | Negative | Ding'an | 2023 | 306 | C076-2 | Negative | Qionghai | 2020 |
| 139 | XS-10 | Negative | Ding'an | 2023 | 307 | C077-2 | Negative | Qionghai | 2020 |
| 140 | XS-11 | Negative | Ding'an | 2023 | 308 | C078-1 | Negative | Qionghai | 2020 |
| 141 | LK-1 | Negative | Ding'an | 2023 | 309 | C080-2 | Negative | Qionghai | 2020 |
| 142 | LK-2 | Negative | Ding'an | 2023 | 310 | C056-2 | Negative | Qionghai | 2020 |
| 143 | LK-3 | Negative | Ding'an | 2023 | 311 | C065-2 | Negative | Qionghai | 2020 |
| 144 | LK-4 | Negative | Ding'an | 2023 | 312 | D008-2 | Negative | Ding’an | 2020 |
| 145 | LK-5 | Negative | Ding'an | 2023 | 313 | D039-1 | Negative | Ding’an | 2020 |
| 146 | LK-6 | Negative | Ding'an | 2023 | 314 | D062-2 | Negative | Ding’an | 2020 |
| 147 | LK-7 | Negative | Ding'an | 2023 | 315 | D093-2 | Negative | Ding’an | 2020 |
| 148 | LK-8 | Negative | Ding'an | 2023 | 316 | ZY2-1 | Negative | Wenchang | 2020 |
| 149 | LK-9 | Negative | Ding'an | 2023 | 317 | 7-4-2 | Negative | Wenchang | 2020 |
| 150 | LK-10 | Negative | Ding'an | 2023 | 318 | 003-2 | Negative | Wenchang | 2020 |
| 151 | LK-11 | Negative | Ding'an | 2023 | 319 | B021-2 | Negative | Baoting | 2020 |
| 152 | LK-12 | Negative | Ding'an | 2023 | 320 | B027-2 | Negative | Baoting | 2020 |
| 153 | LK-13 | Negative | Ding'an | 2023 | 321 | B029-2 | Negative | Baoting | 2020 |
| 154 | LK-14 | Negative | Ding'an | 2023 | 322 | B030-1 | Negative | Baoting | 2020 |
| 155 | LK-15 | Negative | Ding'an | 2023 | 323 | B032-1 | Negative | Baoting | 2020 |
| 156 | LK-16 | Negative | Ding'an | 2023 | 324 | B037-2 | Negative | Baoting | 2020 |
| 157 | LK-17 | Negative | Ding'an | 2023 | 325 | B038-2 | Negative | Baoting | 2020 |
| 158 | LK-18 | Negative | Ding'an | 2023 | 326 | B042-1 | Negative | Baoting | 2020 |
| 159 | LK-19 | Negative | Ding'an | 2023 | 327 | B042-2 | Negative | Baoting | 2020 |
| 160 | LK-20 | Negative | Ding'an | 2023 | 328 | B055-2 | Negative | Baoting | 2020 |
| 161 | I027 | Negative | Qionghai | 2022 | 329 | B059-1 | Negative | Baoting | 2020 |
| 162 | N01 | Negative | Lingshui | 2022 | 330 | B059-2 | Negative | Baoting | 2020 |
| 163 | N06 | Negative | Lingshui | 2022 | 331 | B056-1 | Negative | Baoting | 2020 |
| 164 | N07 | Negative | Lingshui | 2022 | 332 | B056-2 | Negative | Baoting | 2020 |
| 165 | N012 | Negative | Lingshui | 2022 | 333 | B061-1 | Negative | Baoting | 2020 |
| 166 | N013 | Negative | Lingshui | 2022 | 334 | B069-1 | Negative | Baoting | 2020 |
| 167 | ZH-40 | Negative | Lingshui | 2022 | 335 | B069-2 | Negative | Baoting | 2020 |
| 168 | NLT | Negative | Lingshui | 2022 | |||||
Note: a total of 10 leaf samples were positive in the detection using our novel nest PCR.
References
- Kumar, S.N.; Bai, K.V.K.; Rajagopal, V.; Aggarwal, P.K. Simulating coconut growth, development and yield with the infocrop-coconut model. Tree Physiology 2008, 28. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.J.; Gong, M. Current Development status and countermeasures of Arecanut planting and processing industry in Hainan. Chinese Journal of Tropical Agriculture 2019, 39, 91–94. [Google Scholar]
- Chen, F.; Liu, T.; L, J.J.; Chen, J.X. The medical value of areca nut. China Tropical Medicine 2014, 14, 243–245. [Google Scholar]
- Tang, Q.H.; Meng, X.L.; Yu, S.S.; Lin, Z.W.; Niu, X.Q.; Song, W.W.; Qin, W.Q. Forty years of research on “yellow leaf disease of areca palm” in China: New progress of the causal agent and the management. Chinese Journal of Tropical Crops 2022, 43, 1010–1022. [Google Scholar]
- Tang, Q.H.; Song, W.W.; Yu, S.S.; Niu, X.Q.; Qin, W.Q. Questions and foresight on the causal agent of Arecanut yellow leaf disease researches and it’s management Plant Protection 2021, 47, 6–11.
- Abeysinghe, S.; Abeysinghe, P.D.; Kanatiwela-de Silva, C.; Udagama, P.; Warawichanee, K.; Aljafar, N.; Kawicha, P.; Dickinson, M. Refinement of the taxonomic structure of 16SrXI and 16SrXIV phytoplasmas of gramineous plants using multilocus sequence typing. Plant Disease 2016, 100, 2001–2010. [Google Scholar] [CrossRef]
- Jin, K.X.; Sun, F.S.; Chen, M.R.; Luo, D.Q.; Cai, X.Z. Yellows disease of betel nut palm in Hainan, China. Scientia Silvae Sinicae 1995, 31, 556–558. [Google Scholar]
- Kanatiwela-de Silva, C.; Damayanthi, M.; de Silva, R.; Dickinson, M.; de Silva, N.; Udagama, P. Molecular and scanning electron microscopic proof of phytoplasma associated with areca palm yellow leaf disease in Sri Lanka. Plant Disease 2015, 99, 1641–1641. [Google Scholar] [CrossRef]
- Lin, Z.W.; Tang, Q.H.; Meng, X.L.; Song, W.W.; Yu, F.Y.; Huang, S.C.; Niu, X.Q.; Qin, W.Q. Occurrence and distribution of areca pathological yellowing in Hainan, China and Its phytoplasma detection analysis. Chinese Journal of Tropical Crops 2022, 43, 2106–2113. [Google Scholar]
- Nair, S.; Roshna, O.M.; Soumya, V.P.; Hegde, V.; Suresh Kumar, M.; Manimekalai, R.; Thomas, G.V. Real-Time PCR technique for detection of Arecanut yellow leaf disease phytoplasma. Australasian Plant Pathology 2014, 43, 527–529. [Google Scholar] [CrossRef]
- Chaithra, M.; Priya, M.; Kumar, S.; Manimekalai, R.; Rao, G. Detection and characterization of 16SrI-B phytoplasmas associated with yellow leaf disease of Arecanut palm in India. Phytopathogenic Mollicutes 2015, 4, 77–82. [Google Scholar]
- Sumi, K.; Priya, M.; Kumar, S.; Rao, G.; Rao, K. Molecular confirmation and interrelationship of phytoplasmas associated with diseases of palms in South India. Phytopathogenic Mollicutes 2015, 4, 41–52. [Google Scholar] [CrossRef]
- Zhou, Y.K.; Gan, B.C.; Zhang, Z.; Sui, C.; Wei, J.H.; Yang, Y.; Yang, X.Q. Detection of the phytoplasmas associated with yellow leaf disease of Areca catechu L. in Hainan province of China by Nested PCR. Chinese Agricultural Science Bulletin 2010, 26, 381–384. [Google Scholar]
- Che, H.Y.; Wu, C.T.; Fu, R.Y.; Wen, Y.S.; Ye, S.B.; Luo, D.Q. Molecular identification of pathogens from Areca nut yellow leaf disease in Hainan. Chinese Journal of Tropical Crops 2010, 31, 83–87. [Google Scholar]
- Lin, Z.; Song, W.; Tang, Q.; Meng, X. First Report of 16SrII group-related phytoplasma associated with areca palm yellow leaf disease on Areca catechu in China. Plant Disease 2023, 107, 3275. [Google Scholar] [CrossRef]
- Yu, S.S.; Zhu, A.N.; Che, H.Y.; Song, W.W. Molecular identification of ‘Candidatus phytoplasma Malaysianum’-related strains associated with Areca Catechu palm yellow leaf disease and phylogenetic diversity of the phytoplasmas within the 16SrXXXII group. Plant Disease 2024, 108, 1331–1343. [Google Scholar] [CrossRef]
- Lin, Z.W.; Tang, Q.H.; Long, S.D.; Niu, X.Q.; Liu, F.; Song, W.W. Establishment of Real-time fluorescent quantitative PCR method for detection of areca yellows phytoplasma 16SrI group. Molecular Plant Breeding 2024, 1–15. [Google Scholar]
- Nayar, R.; Seliskar, C.E. Mycoplasma like organisms associated with yellow leaf disease of Areca Catechu L. Forest Pathology 1978, 8, 125–128. [Google Scholar] [CrossRef]
- Rajeev, G.; Vijayamma Ramakrishnan Nair, P.; Vaganan, M.; Sasikala, M.; Solomon, J.; Nair, G. Microscopic and polyclonal antibody-based detection of yellow leaf disease of Arecanut (Areca Catechu L.). Archives of Phytopathology and Plant Protection 2011, 44, 1093–1104. [Google Scholar] [CrossRef]
- Meng, X.L.; Tang, Q.H.; Lin, Z.W.; Niu, X.Q.; Liu, B.; Song, W.W. Developing efficient primers to detect phytoplasmas in areca palms infected with yellow leaf disease. Molecular Plant Breeding 2022, 20, 4624–4633. [Google Scholar]
- Yu, S.S.; Che, H.Y.; Wang, S.J.; Lin, C.L.; Lin, M.X.; Song, W.W.; Tang, Q.H.; Yan, W.; Qin, W.-Q. Rapid and efficient detection of 16SrI group areca palm yellow leaf phytoplasma in China by Loop-Mediated Isothermal Amplification. Journal of Plant Pathology 2020, 36, 459–467. [Google Scholar] [CrossRef]
- Yu, S.; Zhang, X.; Song, W.; Qin, W. Accurate and sensitive detection of areca palm yellow leaf phytoplasma in China by Droplet Digital PCR targeting tuf gene sequence. Annals of Applied Biology 2022, 181, 152–159. [Google Scholar] [CrossRef]
- Lin, Z.W.; Meng, X.L.; Tang, Q.H.; Niu, X.Q.; Song, W.W. Establishment of TaqMan probe real-time fluorescent quantitative PCR detection method for areca palm yellow leaf phytoplasma. Chinese Journal of Tropical Crops 2023, 1–8. [Google Scholar]
- Fan, X.X.; Zhao, Y.G.; Li, L.; Wu, X.D.; Wang, Z.L. Research progress of recombinant Enzyme Polymerase Amplification (RPA) in rapid detection of diseases. China Animal Health Inspection 2016, 33, 72–77. [Google Scholar]
- Lu, P.P.; Wu, W.H.; Zheng, J.L.; Wang, G.H.; He, C.P.; Lin, P.Q.; Huang, X.; Liang, Y.Q.; Yi, K.X. Establishment and optimization of single-tube Nested PCR detection technique for phytoplasma related to sisal purple leafroll disease. Journal of Agricultural Biotechnology 2021, 29(7), 1426–1434. [Google Scholar]
- Lin, Z.W.; Long, S.D.; Meng, X.L.; Tang, Q.H.; Niu, X.Q.; Wang, Y.N.; Song, W.W. Cloning and sequence analysis of tuf, secA and rp genes of areca yellows phytoplasma. Molecular Plant Breeding 2024, 1–15. [Google Scholar]
- Mou, H.Q.; Zhu, S.F.; Xu, X.; Zhao, W.J. An overview of research on phytoplasma-induced diseases. Plant Protection 2011, 37, 17–22. [Google Scholar]
- Bertaccini, A. Plants and phytoplasmas: when bacteria modify plants. Plants (Basel) 2022, 11, 1425. [Google Scholar] [CrossRef]
- Wei, W.; Zhao, Y. Phytoplasma taxonomy: nomenclature, classification, and identification. Biology (Basel) 2022, 11, 1119. [Google Scholar] [CrossRef]
- Dai, Q.; Liu, B.S.; He, F.T.; Chen, Z.W. Relationship between the seasonal temperature variation and the phytoplasmal amounts in mulberry trees. Scientia Silvae Sinicae 1998, 34, 76–80. [Google Scholar]
- Duan, Y.W.; Yang, Y.; Lu, Q.L.; Xiao, Z.L.; Li, D.Y.; Zhang, W.Z.; Zhao, W.J. Molecular identification of beggarweed witches′-broom phytoplasma. Plant Quarantine 2019, 33, 29–32. [Google Scholar]
- Song, X.B.; Huang, F.; Tang, Y.F.; Cui, Y.P.; Ling, J.F.; Chen, X. First report of; little leaf disease caused by phytoplasma on Breynia disticha in China. Plant Protection 2024, 50, 272 277+292. [Google Scholar]
- Yang, Y.; Jiang, L.; Li, S.F. Research progress on phytoplasma classification and identification. Plant Quarantine 2020, 34, 13–20. [Google Scholar]
- Lai, F.; Li, Y.; Xu, Q.C.; Tian, G.Z. The present status on classification of phytoplasmas. Microbiology China 2008, 291–295. [Google Scholar]
- Wang, G.; Wu, W.; Tan, S.; Liang, Y.; He, C.; Chen, H.; Huang, X.; Yi, K. Development of a specific Nested PCR assay for the detection of 16SrI group phytoplasmas associated with sisal purple leafroll disease in sisal plants and mealybugs. Plants 2022, 11, 2817. [Google Scholar] [CrossRef]
- Deng, S.; Hiruki, C. Amplification of 16S rRNA genes from culturable and nonculturable mollicutes. Journal of Microbiological Methods 1991, 14, 53–61. [Google Scholar] [CrossRef]
- Gundersen, D.E.; Lee, I.M.; Gundersen, D.E.; Lee, I.M. Ultrasensitive detection of phytoplasmas by Nested-PCR assays using two universal primer pairs. Phytopathologia Mediterranea 1996, 35, 144–151. [Google Scholar]
- Zhu, A.N.; Yu, S.S.; Su, L.H.; Liu, L.; Song, W.W.; Yan, W. Molecular detection and genetic variation of phytoplasmas from eight plants in garden of areca palm with yellow leaf disease in China. Chinese Journal of Tropical Crops 2023, 44, 1190–1202. [Google Scholar]
Figure 1.
Profile of 16S rDNA sequences amplified by nested PCR with universal primer P1/P7 followed by R16mF2/R16mR1, from areca samples. M: DL2000 DNA Marker; 1-19: areca samples; NC1: blank control of the first round of PCR; NC2: blank control of the second round of PCR; PC: positive control, periwinkle DNA, the same for the following figures.
Figure 1.
Profile of 16S rDNA sequences amplified by nested PCR with universal primer P1/P7 followed by R16mF2/R16mR1, from areca samples. M: DL2000 DNA Marker; 1-19: areca samples; NC1: blank control of the first round of PCR; NC2: blank control of the second round of PCR; PC: positive control, periwinkle DNA, the same for the following figures.
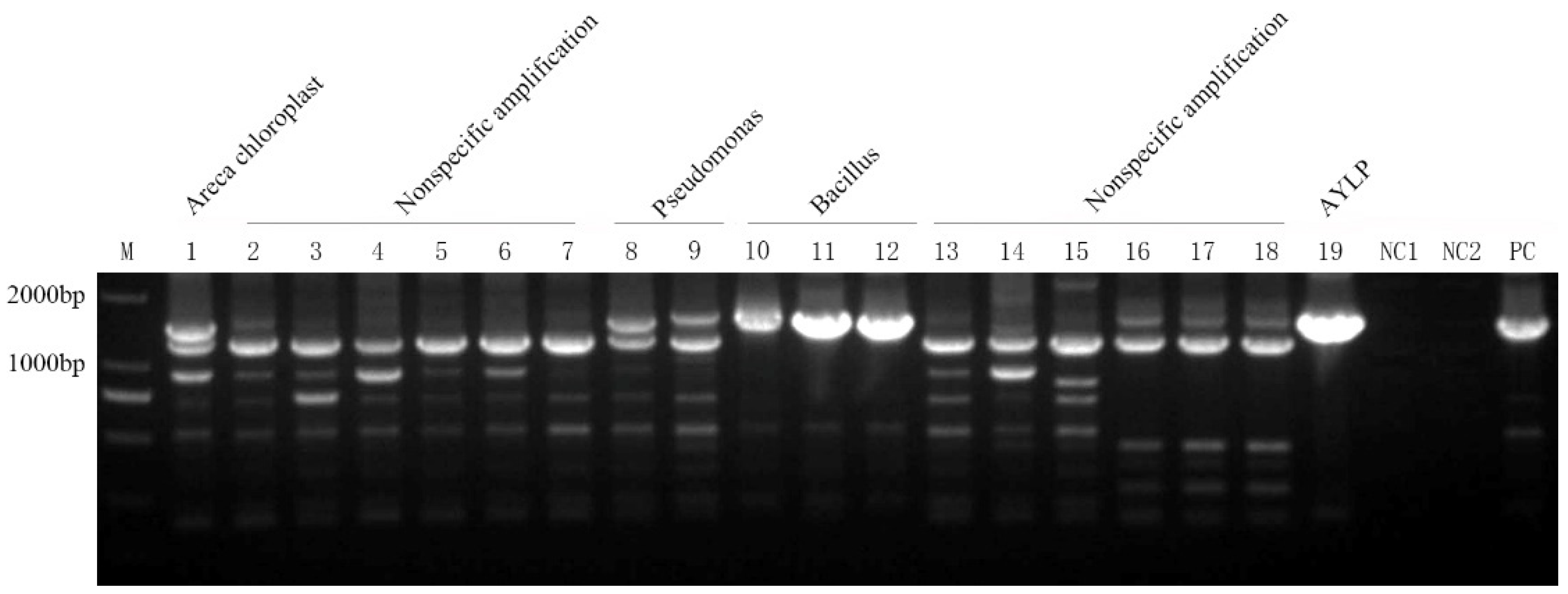
Figure 2.
Ratios of different sequences in 50 samples.
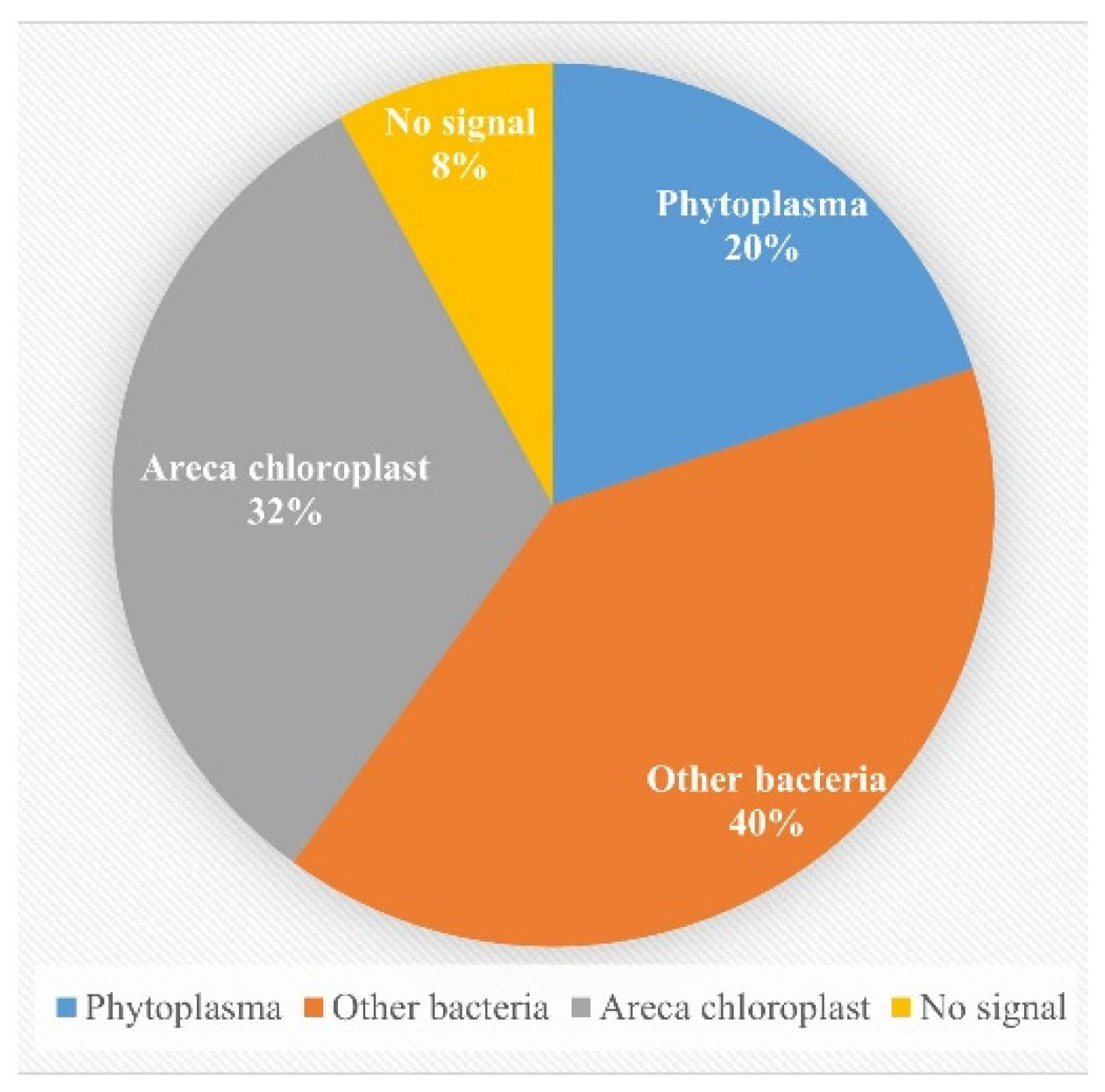
Figure 3.
The preliminary screening of three pairs of internal primers designed in this study. A: Profile of 16S rDNA sequences amplified by nested PCR with primer HNP-1F/1R followed by HNP-2F/2R; B: Profile of 16S rDNA sequences amplified by nested PCR with primer HNP-1F/1R followed by HNP-3F/3R; C: Profile of 16S rDNA sequences amplified by nested PCR with primer HNP-1F/1R followed by HNP-4F/4R. M: marker DL2000; NC1: blank control for the first round of PCR; NC2: blank control for the second round of PCR; PC: positive control.
Figure 3.
The preliminary screening of three pairs of internal primers designed in this study. A: Profile of 16S rDNA sequences amplified by nested PCR with primer HNP-1F/1R followed by HNP-2F/2R; B: Profile of 16S rDNA sequences amplified by nested PCR with primer HNP-1F/1R followed by HNP-3F/3R; C: Profile of 16S rDNA sequences amplified by nested PCR with primer HNP-1F/1R followed by HNP-4F/4R. M: marker DL2000; NC1: blank control for the first round of PCR; NC2: blank control for the second round of PCR; PC: positive control.
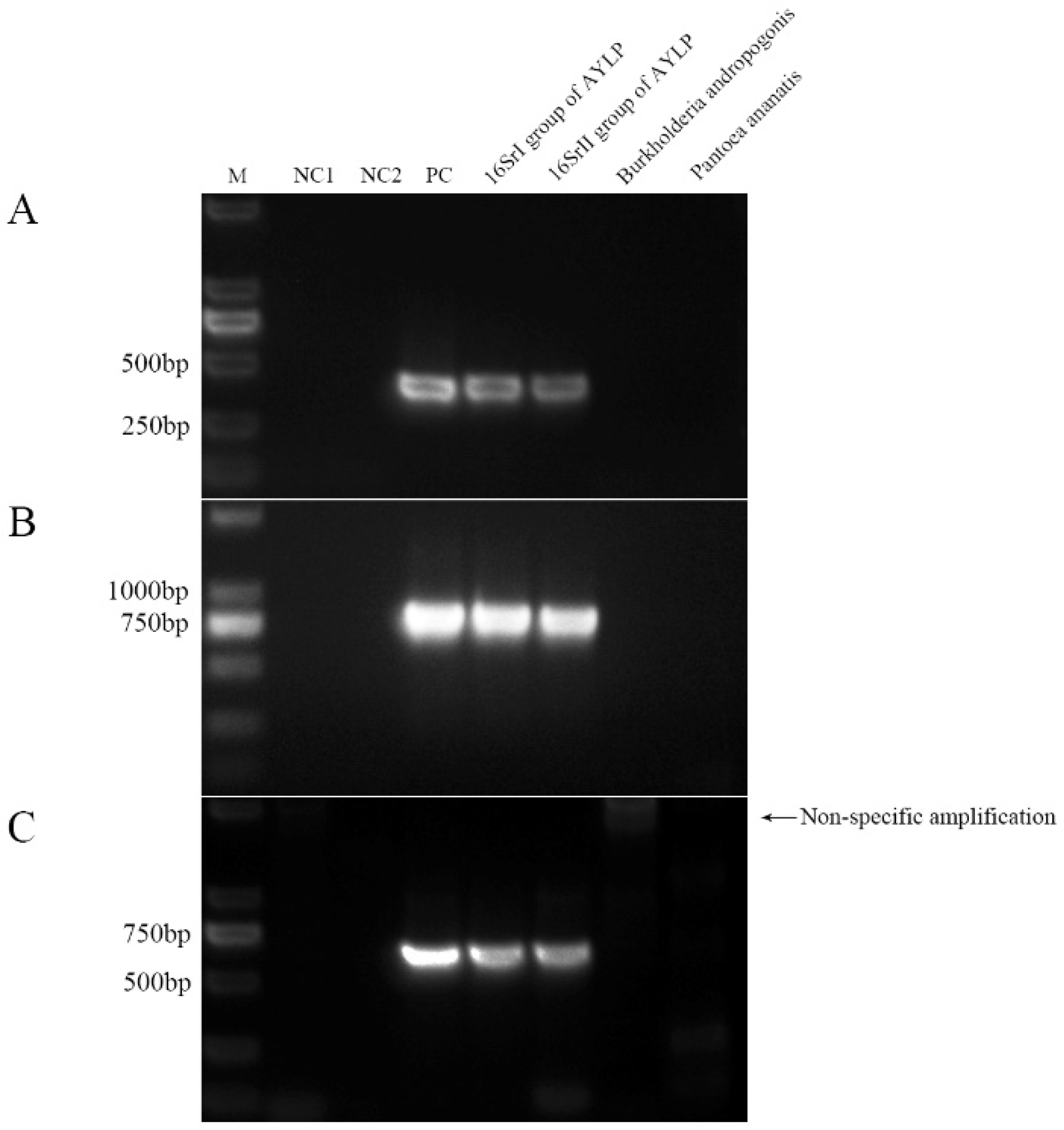
Figure 4.
Profile of 16S rDNA sequences amplified by nested PCR with primer HNP-1F/1R followed by HNP-2F/2R and HNP-3F/3R respective for the specificity validation. A: HNP-2F/2R, B: HNP-3F/3R. M: marker DL2000; NC1: blank control for the first round of PCR; NC2: blank control for the second round of PCR; PC: positive control; 1: healthy areca leaf sample; 2: Burkholderia andropogonis (Robbsia andropogonis); 3: Pantoea ananatis; 4: Jujube witches-broom phytoplasma; 5: AYLP of 16SrI group; 6: AYLP of 16SrII group; 7: Trema tomentosa witches'-broom phytoplasma; 8: leaf sample infected by areca palm velarivirus 1.
Figure 4.
Profile of 16S rDNA sequences amplified by nested PCR with primer HNP-1F/1R followed by HNP-2F/2R and HNP-3F/3R respective for the specificity validation. A: HNP-2F/2R, B: HNP-3F/3R. M: marker DL2000; NC1: blank control for the first round of PCR; NC2: blank control for the second round of PCR; PC: positive control; 1: healthy areca leaf sample; 2: Burkholderia andropogonis (Robbsia andropogonis); 3: Pantoea ananatis; 4: Jujube witches-broom phytoplasma; 5: AYLP of 16SrI group; 6: AYLP of 16SrII group; 7: Trema tomentosa witches'-broom phytoplasma; 8: leaf sample infected by areca palm velarivirus 1.
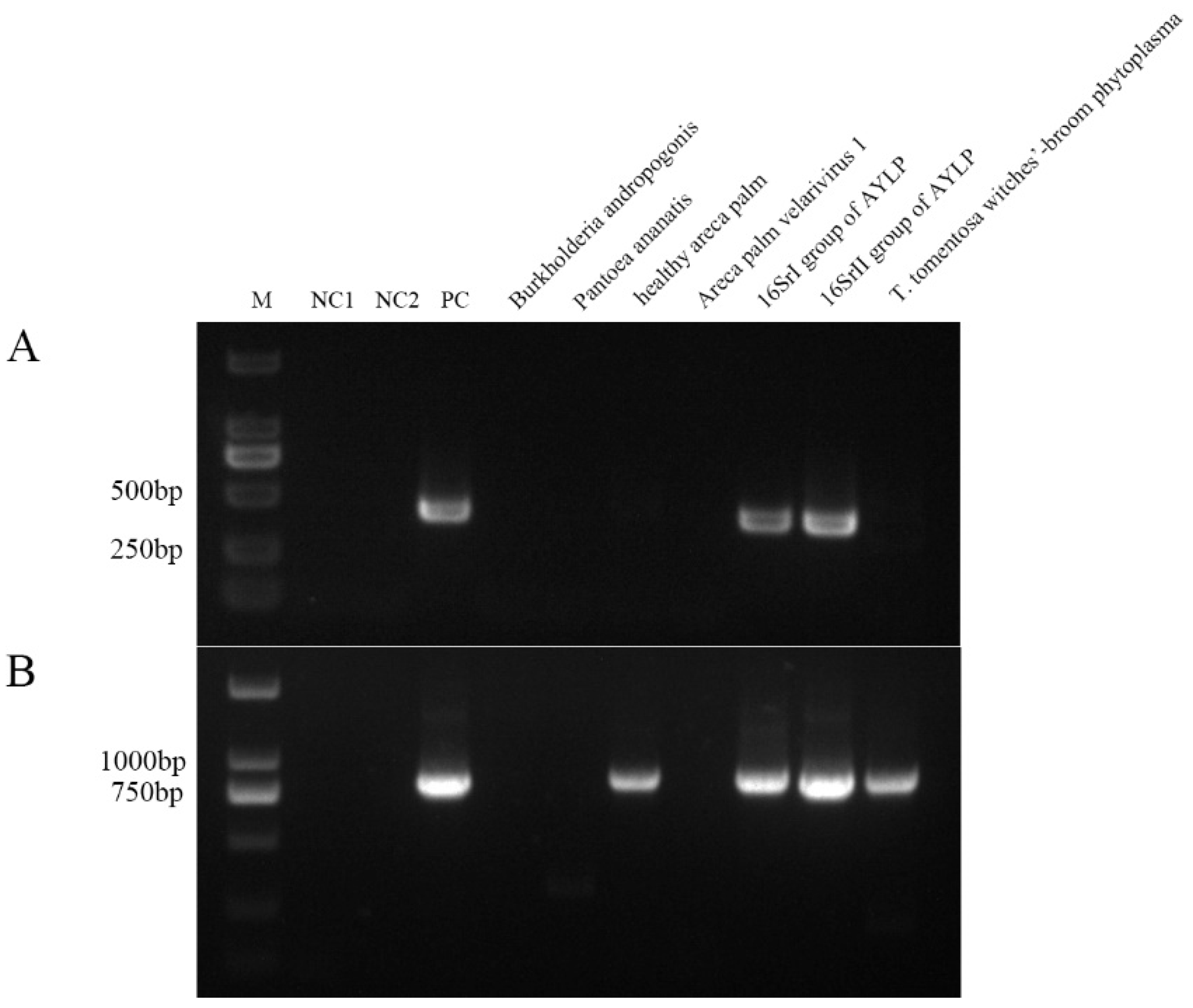
Figure 5.
A: Annealing temperature optimized for the outer primer HNP-1F/1R. M: marker DL2000; NC: blank control. M: marker DL2000; NC: blank control. B: Annealing temperature optimized for the inner primer HNP-2F/2R. M: marker DL2000; NC: blank control.
Figure 5.
A: Annealing temperature optimized for the outer primer HNP-1F/1R. M: marker DL2000; NC: blank control. M: marker DL2000; NC: blank control. B: Annealing temperature optimized for the inner primer HNP-2F/2R. M: marker DL2000; NC: blank control.
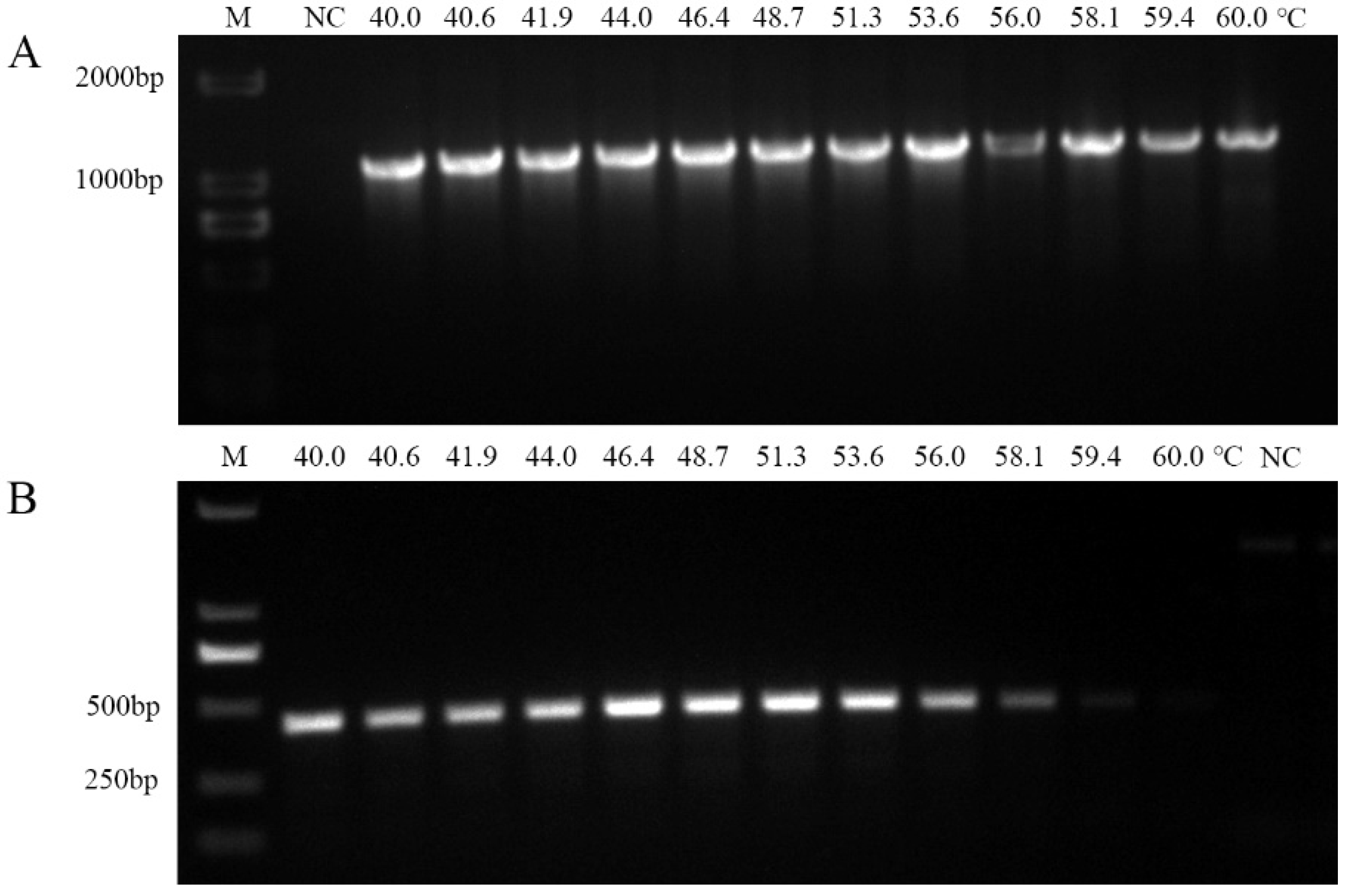
Figure 6.
Sensitivity test for nested PCR primer set of HNP-1F/1R and HNP-2F/2R. A: Gel electrophoresis for sensitivity of 16SrI phytoplasma using novel nested PCR. M: marker DL2000; NC1: blank control for the first round of PCR; NC2: blank control for the second round of PCR. B: Gel electrophoresis results for the sensitivity of 16SrII phytoplasma using novel nested PCR M: marker DL2000; NC: blank control.
Figure 6.
Sensitivity test for nested PCR primer set of HNP-1F/1R and HNP-2F/2R. A: Gel electrophoresis for sensitivity of 16SrI phytoplasma using novel nested PCR. M: marker DL2000; NC1: blank control for the first round of PCR; NC2: blank control for the second round of PCR. B: Gel electrophoresis results for the sensitivity of 16SrII phytoplasma using novel nested PCR M: marker DL2000; NC: blank control.
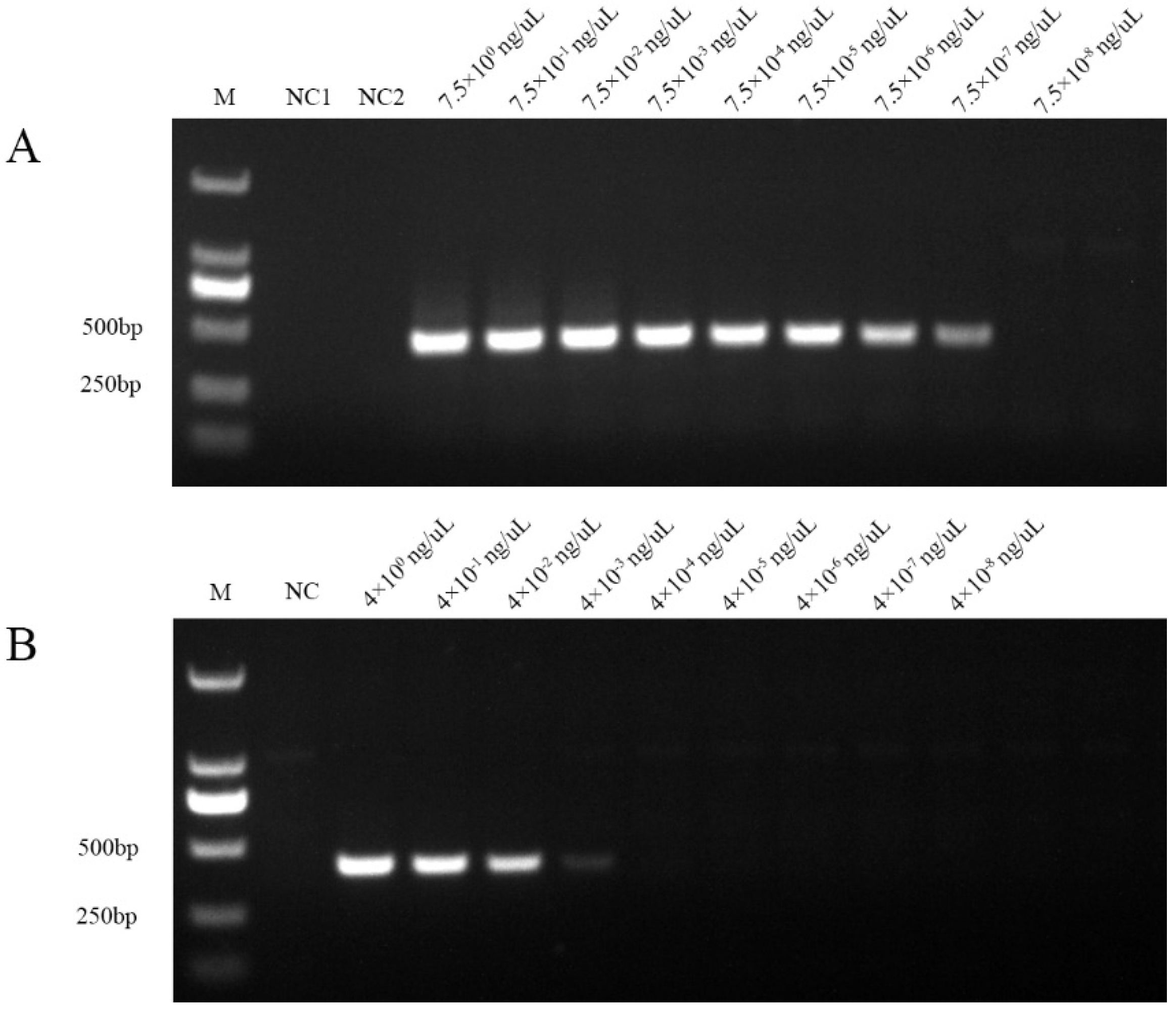
Figure 7.
Comparison of Profile of 16S rDNA sequences amplified using universal primers with the novel primers developed in this study. M: marker DL2000; 1: blank control for the first round of PCR; 2: blank control for the second round of PCR; 3: healthy areca leaf sample; 4: positive control; 5-34: areca leaf samples collected in the fields.
Figure 7.
Comparison of Profile of 16S rDNA sequences amplified using universal primers with the novel primers developed in this study. M: marker DL2000; 1: blank control for the first round of PCR; 2: blank control for the second round of PCR; 3: healthy areca leaf sample; 4: positive control; 5-34: areca leaf samples collected in the fields.
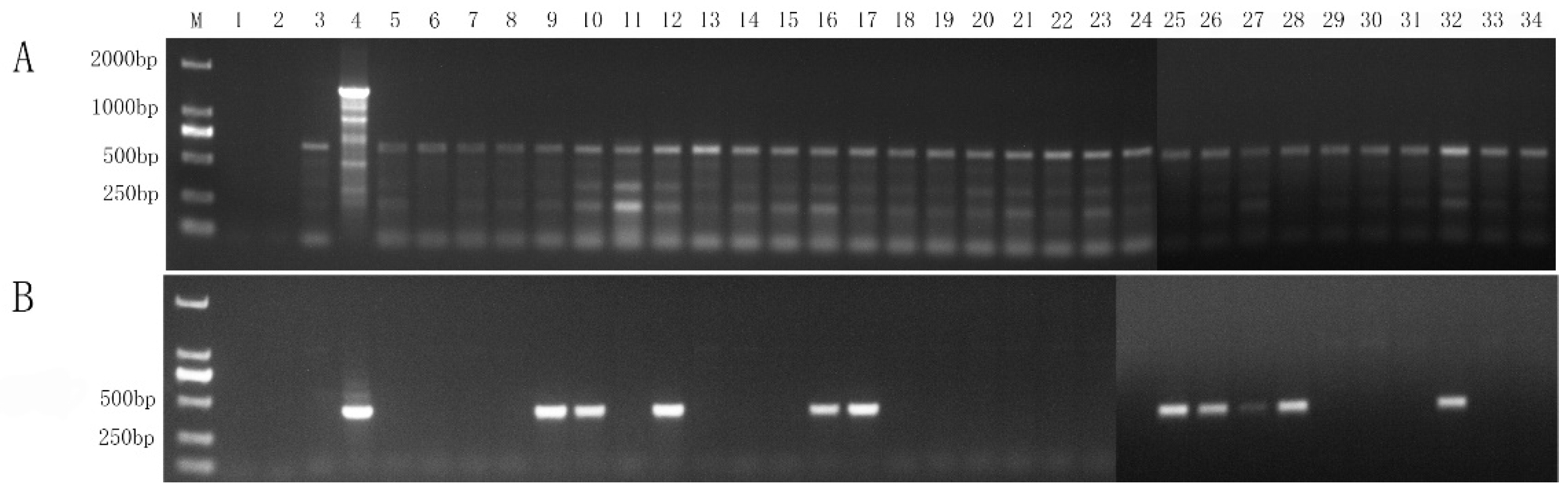
Figure 8.
Phylogenetic tree constructed based on 16S rDNA gene sequences of phytoplasmas. A: Phylogenetic tree constructed from the amplified fragments of universal primers targeting the 16S rDNA gene in phytoplasmas. B: Phylogenetic tree constructed from the amplified fragments of nested primers established in this study for the 16S rDNA gene in phytoplasmas. C: Multiple sequence alignment of 10 positive PCR product sequences and the T40 sequence used for constructing the phylogenetic tree with 16SrI-B subgroup AYLP.
Figure 8.
Phylogenetic tree constructed based on 16S rDNA gene sequences of phytoplasmas. A: Phylogenetic tree constructed from the amplified fragments of universal primers targeting the 16S rDNA gene in phytoplasmas. B: Phylogenetic tree constructed from the amplified fragments of nested primers established in this study for the 16S rDNA gene in phytoplasmas. C: Multiple sequence alignment of 10 positive PCR product sequences and the T40 sequence used for constructing the phylogenetic tree with 16SrI-B subgroup AYLP.
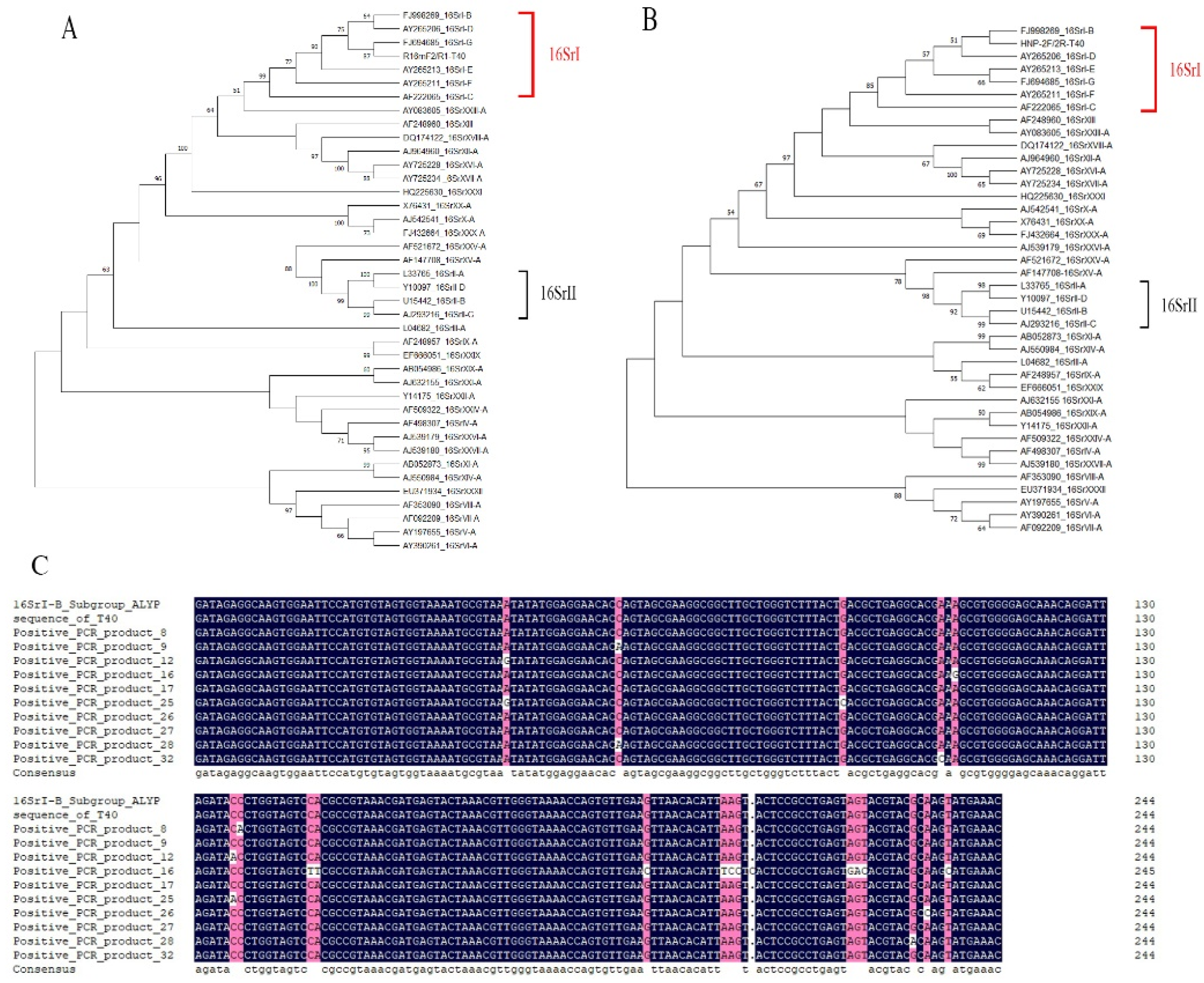
Table 1.
Areca leaf samples used in this study.
| Samples | Taxonomy | Host plant | Location |
|---|---|---|---|
| J069 | 16SrI Group | Areca palm Areca catechu | Wenchang, Hainan |
| S97 | 16SrI Group | Areca palm A. catechu | Wenchang, Hainan |
| T40 | 16SrI Group | Areca palm A. catechu | Wenchang, Hainan |
| H-9 | 16SrI Group | Periwinkle Catharanthus roseus | Wenchang, Hainan |
| A056 | 16SrII Group | Areca palm A. catechu | Wenchang, Hainan |
| W4 | 16SrII Group | Areca palm A. catechu | Wenchang, Hainan |
| MW1-1 | Burkholderia andropogonis | Areca palm A. catechu | Wenchang, Hainan |
| I027 | Areca palm velarivirus 1 | Areca palm A. catechu | Wenchang, Hainan |
| TC-1 | Pantoea ananatis | Areca palm A. catechu | Tunchang, Hainan |
| SHM-1 | 16SrXXXII Group | Trema tomentosa | Qionghai, Hainan |
| PT-1 | 16SrI Group | Paulownia sp. | Taian, Shandong |
| ZF-1 | 16SrIV Group | Jujube Ziziphus jujuba | Taian, Shandong |
Table 2.
Primers for amplification of 16S rDNA gene using nested-PCR.
| Primer | Primer sequence (5’- 3’) | Annealing temperature (℃) | Target fragment length (bp) | References |
|---|---|---|---|---|
| P1 | AAGAATTTGATCCTGGCTCAGGATT | 55 | 1800 | [36] |
| P7 | CGTCCTTCATCGGCTCTT | |||
| R16mF2 | CATGCAAGTCGAACGGA | 50 | 1500 | [37] |
| R16mR1 | CTTAACCCCAATCATCGA | |||
| HNP-1F | TTCTTGTTTTTAAAAGACCT | 44 | 1072 | This study |
| HNP-1R | AAACTTGCGCTTCAGCT | |||
| HNP-2F | TGTGGTCTAAGTGCAAT | 48 | 429 | This study |
| HNP-2R | CTGATAACCTCCACTGTGTT | |||
| HNP-3F | TTCTTGTTTTTAAAAGACCT | 50 | 837 | This study |
| HNP-3R | ATAACCTCCACTGTGTTTCT | |||
| HNP-4F | AATGCTCAACATTGTGATGCT | 48 | 652 | This study |
| HNP-4R | AAACTTGCGCTTCAGCT |
Table 3.
Reference sequences used in this study.
| NO. | Samples | GenBank accession number | Application |
|---|---|---|---|
| 1 | 16Sr I-B SubGroup AYLP | FJ998269 | Primer Design/Phylogenetic analysis |
| 2 | 16Sr I-G SubGroup AYLP | FJ694685 | Primer Design/Phylogenetic analysis |
| 3 | 16Sr II-A SubGroup AYLP | OQ586085 | Primer Design |
| 4 | Areca Catechu Chloroplast | NC_050163 | Primer Design |
| 5 | Burkholderia andropogonis | NR_104960 | Primer Design |
| 6 | Pantoea ananatis | MW174802 | Primer Design |
| 7 | Chrysophyllum albidum | LC110196 | Primer Design |
| 8 | Curtobacterium citreum | MF319766 | Primer Design |
| 9 | Curtobacterium luteum | JX437941 | Primer Design |
| 10 | Sphingomonas yantingensis | MF101149 | Primer Design |
| 11 |
Bacillus cereus (Robbsia andropogonis) |
HQ833025 | Primer Design |
| 12 | Staphylococcus epidermidis | CP040883 | Primer Design |
| 13 | Xanthomonas sacchari | MN889285 | Primer Design |
| 14 | Xanthomonas campestris | JX415480 | Primer Design |
| 15 | |||
| 16 | 16SrI-C SubGroup | AF222065 | Phylogenetic analysis |
| 17 | 16SrI-D SubGroup | AY265206 | Phylogenetic analysis |
| 18 | 16SrI-E SubGroup | AY265213 | Phylogenetic analysis |
| 19 | 16SrI-F SubGroup | AY265211 | Phylogenetic analysis |
| 20 | |||
| 21 | 16SrII-A SubGroup | L33765 | Phylogenetic analysis |
| 22 | 16SrII-B SubGroup | U15442 | Phylogenetic analysis |
| 23 | 16SrII-C SubGroup | AJ293216 | Phylogenetic analysis |
| 24 | 16SrII-D SubGroup | Y10097 | Phylogenetic analysis |
| 25 | 16SrIII-A SubGroup | L04682 | Phylogenetic analysis |
| 26 | 16SrIV-A SubGroup | AF498307 | Phylogenetic analysis |
| 27 | 16SrV-A SubGroup | AY197655 | Phylogenetic analysis |
| 28 | 16SrVI-A SubGroup | AY390261 | Phylogenetic analysis |
| 29 | 16SrVII-A SubGroup | AF092209 | Phylogenetic analysis |
| 30 | 16SrVIII-A SubGroup | AF353090 | Phylogenetic analysis |
| 21 | 16SrIX-A SubGroup | AF248957 | Phylogenetic analysis |
| 32 | 16SrX-A SubGroup | AJ542541 | Phylogenetic analysis |
| 33 | 16SrXI-A SubGroup | AB052873 | Phylogenetic analysis |
| 34 | 16SrXII-A SubGroup | AJ964960 | Phylogenetic analysis |
| 35 | 16SrXIII Group | AF248960 | Phylogenetic analysis |
| 36 | 16SrXIV-A SubGroup | AJ550984 | Phylogenetic analysis |
| 37 | 16SrXV-A SubGroup | AF147708 | Phylogenetic analysis |
| 38 | 16SrXVI-A SubGroup | AY725228 | Phylogenetic analysis |
| 39 | 16SrXVII-A SubGroup | AY725234 | Phylogenetic analysis |
| 40 | 16SrXVIII-A SubGroup | DQ174122 | Phylogenetic analysis |
| 41 | 16SrXIX-A SubGroup | AB054986 | Phylogenetic analysis |
| 42 | 16SrXX-A SubGroup | X76431 | Phylogenetic analysis |
| 43 | 16SrXXI-A SubGroup | AJ632155 | Phylogenetic analysis |
| 44 | 16SrXXII-A SubGroup | Y14175 | Phylogenetic analysis |
| 45 | 16SrXXIII-A SubGroup | AY083605 | Phylogenetic analysis |
| 46 | 16SrXXIV-A SubGroup | AF509322 | Phylogenetic analysis |
| 47 | 16SrXXV-A SubGroup | AF521672 | Phylogenetic analysis |
| 48 | 16SrXXVI-A SubGroup | AJ539179 | Phylogenetic analysis |
| 49 | 16SrXXVII-A SubGroup | AJ539180 | Phylogenetic analysis |
| 50 | 16SrXXIX Group | EF666051 | Phylogenetic analysis |
| 51 | 16SrXXX-A SubGroup | FJ432664 | Phylogenetic analysis |
| 52 | 16SrXXXI Group | HQ225630 | Phylogenetic analysis |
| 53 | 16SrXXXII Group | EU371934 | Phylogenetic analysis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



