Submitted:
18 April 2025
Posted:
22 April 2025
You are already at the latest version
Abstract
Phytate, an antinutritional molecule in poultry feed, can be degraded by applying phytase, but its use in developing countries is often limited due to importation instead of local production. Here, inexpensive raw materials have been used to optimize the production of a thermostable phytase from an indigenous strain of Bacillus subtilis SP11 that was isolated from a broiler farm in Dhaka. SP11 was identified using 16s rDNA and fermentation of phytase was optimized using Plack-ett-Burman design and response surface methodology, revealing that three substrates including the raw material mustard meal (2.21 % w/v) cause maximum phytase production of 436 U/L at 37oC and 120 rpm for 72 hours, resulting in a 3.7-fold increase compared to unoptimized media. The crude enzyme showed thermostability up to 80oC (may withstand the feed pelleting process) with an optimum pH 6 (near crop and small intestine pH), while retaining 96% activity at 41oC (body temperature of the chicken). In vitro dephytinization demonstrated its applicability, releasing 1956 µg of inorganic phosphate per g of wheat bran per hour. This phytase has the potential to reduce the burden of phytase importation in Bangladesh by making local production and application capable, contributing to sustainable poultry nutrition.
Keywords:
phytase
; mustard meal
; plackett-burman design
; response surface methodology
; Bacillus subtilis
1. Introduction
Phytate makes up more than 85% of total phosphorus in oilseeds and cereal grains and is the main organic phosphorus storage form in plants[1]. Non-ruminant animals like pigs and poultry eat diets high in phytate-phosphorus, which is poorly digested as they do not produce the enzyme phytase that degrades phytic acid[2]. Phytate can complex with protein at low pH (stomach) and bind with divalent metal cations at high pH (intestine) which makes these nutrients less available to the animal[3]. Besides being an antinutritional molecule for monogastric animals, it can also cause eutrophication[4].
Phytase can catalyze the removal of inorganic phosphate from phytate along with the bound nutrients[5]. As non-ruminants lack the enzyme, adding exogenous phytase to the feed can enhance nutrition availability, thus enhancing growth and less eutrophication[6]. Exogenous phytase addition in feed can also exert a positive effect on the immune system, intestinal microbiota, and antioxidant status[7]. There are several types of phytases from several sources and they have some advantages over each other[8,9,10,11,12,13]. Phytases from Bacillus species are Beta-propeller phytase (BPP) as opposed to the usual commercial phytase from fungal or other bacterial sources which are mostly histidine acid phosphatases (HAP). Although BPP has in general less activity than HAP, they have the advantage of having strict substrate specificity for phytate only (thus unlike HAP, it avoids ATP, GTP and NADH) and being inherently thermostable (so it can withstand the necessary industrial feed pelleting process)[14,15]. Nevertheless, partly due to inefficient enzyme prodction methods, Bacillus phytases have not been applied on a large scale [16].
The global market size for phytase is valued at 547.74 million in 2022 and is predicted to reach 949.96 million by the year 2031[17]. In Bangladesh however, phytases are not locally produced, but rather imported[18] which increases the cost and causes less use of phytase in poultry feed. Also, the long transportation hinders the enzyme’s shelf life and activity. And if these are not thermostable enough they lose significant activity when going through the industrial feed pelleting processes. So, thermostable phytase production from an indigenous strain inside the country can address these issues in Bangladesh.
So, this study focused on 1. Isolating and identifying phytase-producing indigenous bacteria 2. Production optimization of its phytase using both statistical (placket-burman and central composite design) and classical (change one factor at a time) approaches and 3. characterization of the crude enzyme.
2. Materials and Methods
2.1. Sample Collection
Soil samples were collected from three different poultry farms in Dhaka city, Bangladesh. Samples were immediately taken to the enzyme and fermentation biotechnology laboratory, at the University of Dhaka, and preserved in dry and cool conditions until processing.
2.2. Isolation of Phytase-Producing Bacteria
To isolate the phytase-producing bacteria from these samples, approximately 1g of soil was suspended in 0.85% saline up to 10 mL. Diluted samples (10-4 or 10-5) were then spread onto the Phytate Specific Medium (PSM) Plate adapted from Howsan et al.[19] (glucose 15.0 g/l, NH4NO3 5.0 g/l , Na-phytate 5.0g/l, CaCl2 0.3g/l, MgSO4 0.5g/l, MnSO4 0.01g/l, FeSO4 0.01g/l and 1.5% agar, pH adjusted to 6) and incubated at 37oC for 3 days. PSM plates were later observed for the clear zones of hydrolysis around the colonies which indicates the production of extracellular phytase. A clear zone around the colony was evaluated by the plate detection method described by Bea et al.[20] using cobalt chloride and ammonium molybdate/ammonium vanadate solution.
The zone ratio was calculated to determine the efficiency of phytate degradation in PSM plates[21] using the formula:
Z= zone diameter/colony diameter
2.3. Quantitative Screening of Phytase-Producing Bacteria
The bacteria showing clear zone on the PSM were selected for further evaluation of their capacity to produce enzyme in liquid medium. Submerged fermentation was carried out in the 250 ml flask containing 50 ml of Phytase Production Medium (PPM- same composition as PSM without agar) with 5% inoculum size for 3 days at 37ºC and 120 rpm. The inoculum was prepared by taking a loopful of colonies from the plates and inoculating them into 5 mL TSB broth. At 24-hour intervals the phytase activity was measured.
2.4. Phytase Assay
For phytase assay, the method used in this experiment was adopted from Bea et al. [20], which is based on the detection of inorganic phosphate. The cell-free supernatant (CFS) was collected by centrifugation of the fermented broth at 10,000g for 10 minutes. Phytase activity was determined by incubating 300µl of the enzyme solution (CFS) with 1.2 ml of substrate solution [0.2%(w/v) sodium phytate (Sigma, St Louis, MO, USA) in 0.1 M sodium acetate buffer, pH 5.0] for 30 min at 37oC. The reaction was stopped by adding 1.5 ml of 10% (w/v) trichloroacetic acid. From this mixture, 1.5 ml was transferred to a new tube and mixed with 1.5 ml ammonium molybdate ferrous sulfate mixture, which had been prepared by mixing 4 volumes of 1.5% (w/v) ammonium molybdate (Merck, Germany) in 5.5% sulfuric acid (Merck, Germany) with 1 volume of 2.7% (w/v) ferrous sulfate solution. The production of phosphomolybdate was measured spectrophotometrically at 700 nm. From the phosphate standard curve, the amount of liberated Phosphate per ml was found and the enzyme activity was calculated (U/mL). One U is defined as the activity that releases 1 μmol of inorganic phosphate from 0.2% (w/v) sodium phytate per minute at pH 5 and 37oC.
2.5. Identification of SP11 Isolate
Morphological, Microscopic, and various Biochemical tests [Kligler Iron Agar (KIA), citrate utilization , Motility Indole Urease (MIU), Methyl Red- Voges Proskauer (MR-VP), Starch hydrolysis] were determined for the presumptive identification of the SP 11 strain.
To confirm the identification, 16s rDNA sequencing was performed. Total DNA was prepared from isolates using the technique described by Bravo et al.[22]. 16s rDNA (1465 bp) was PCR amplified using Universal primers, 27F (Forward Primer: 5’AGAGTTTGATCMTGGCTCAG3’) and 1492R (Reverse Primer: 5’CGGTTACCTTGTTACGACTT3’), using a thermocycler (annealing Temperature 50oC). PCR products were verified by agarose gel electrophoresis and purified using a purification kit (ATPTM Gel/PCR Fragment DNA Extraction Kit). Purified PCR products were sequenced by chain termination method from DNA Solution Lab, Dhaka, Bangladesh. Bio Edit Sequence Alignment Editor and Mega 7.0 software were used for sequence alignments and the construction of phylogenetic trees respectively.
2.6. Detection of Full-Length Gene Encoding Phytase
DNA was isolated by the method described previously. Related sequences were collected from NCBI and primers with overhangs (for later cloning) were designed using MEGA to amplify a 1200 bp phytase gene. The primers were analyzed with OligoAnalyzer. Phyt.CD S-F: 5'-GGATCCATGAAGGTTCCAAAAACAATGCTGC-3' (Tm¬=61oC) and Phyt.CD S-R: 5'-CTCGAGCTAGCCGTCAGAACGGTCTTTCA-3' (Tm=63.5oC). PCR was performed (each step had 30 seconds for 35 cycles) to obtain the amplicon having an annealing temperature of 58oC.
2.7. Single Factor Analysis
The effect of inoculum size, incubation temperature, pH, and shaking speed on phytase production by Bacillus subtilis SP11 was evaluated by changing a single factor at a time. Fermentation was done using various inoculum sizes (1%, 5%, 10%), temperatures (30oC, 37oC and 42oC), media pH (3, 4, 5, 6, control, 7, 8, 9, 10), and shaking speed (0, 60, 120, 180)
2.8. Plackett-Burman Design (PBD)
PBD was used to evaluate the effect of different media components on phytase production. 19 factors. These are: 7 cheap raw materials for carbon, nitrogen, and mainly for phytate sources (Rice bran, Wheat bran, Soybean meal, Corn meal, mustard meal, Linseed Meal and Sesame Meal), 4 carbon sources [ raw (cane molasses) and organic (glucose as rapid metabolizing and sucrose and citrate as slow metabolizing)], 3 nitrogen sources [inorganic (ammonium nitrate) and organic (Tryptone and yeast extract)], 1 phosphate sources (K2HPO4), and 4 mineral sources (CaCl2, MgCl2, FeSO4, MnSO4). Each factor had 2 levels: high level (+ 1) and low level (− 1), as shown in Table.
24 experimental runs were done and the responses were expressed by the first-order model as shown in Equation:
where Y represents the response variable, ß0 is the interception coefficient and ßi is the coefficient of the linear effects of the 19 independent variables (X1 – X19)
2.9. Central Composite Design (CCD)
CCD was used to determine the optimum levels of three positive significant factors identified in the placket-Burman Design (Mustard Meal, tryptone, and Yeast Extract) and to study the interactions among them for maximum phytase production. Each variable had five levels: two axial (-α, +α), two cube points (-1,+1), and a center point (0) where α is 1.68. 20 experimental runs were conducted with center points repeated six times as per the design. The following quadratic polynomial equation depicts the statistical relationship between the dependent variable Phytase activity (Y) and the selected independent variables:
where Y is the predicted response (Phytase activity U/L); β0 is the model intercept; Xi and Xj are the independent variables, βi is linear coefficients; βij is the cross-product coefficients; βii is the quadratic coefficients. For optimizing and determining interaction coefficients across many parameters the analysis of contour and surface plots achieved was utilized Rstudio.
2.10. Characterization of Crude Phytase
Thermostability of the enzyme was evaluated by treating the crude enzyme at 20oC, 30oC, 40oC, 50oC, 60oC,70oC, 80oC, and 90oC in a water bath for 1 hour. After heat treatment, the enzyme assay is done at standard assay conditions. A control is set which is not subject to any heat treatment. To investigate the optimum temperature for the enzyme, the enzyme assay is done at varying assay temperatures which are 25oC, 30oC, 37oC, 41oC, 50oC, 56oC, 65oC, and 70oC. Other conditions were as per the standard assay conditions. To determine the optimum pH for phytase activity enzyme assay is done in various pH buffer (3,4,5,6,7, and 8). All the standard protocol for phytase assay was followed except the use of specific buffers for different pH and individual standard curves were used for the buffers. The effect of metal ions on the enzyme’s catalytic behavior was studied by pre-incubating phytase enzyme at room temperature in a specified ion (5mM final concentration) containing buffer solution The metals ions were Ca2+, Cu2+, Mg2+, Fe2+, Zn2+, Co2+, Mn2+, Fe3+, and EDTA. After 1 hour of incubation, substrate (0.2% Na-phytate) was added and the relative activity of the enzyme was measured under standard assay conditions (untreated enzyme was taken as control).
2.11. In-Vitro Dephytinization
A modified method from Suresh S and Radha K[23] was adapted to test the applicability of the crude enzyme in dephytinization. 1g of wheat bran or mustard meal was placed into 18 mL of 0.1M pH 5 acetate buffer in 100 mL Erlenmeyer flask. 2 mL of crude phytase was added and incubated at 120 rpm and 37oC. After incubation for 30, 60, 90, and 120 minutes, the suspension was centrifuged at 10000g for 10 min and the amount of inorganic phosphorus was detected by the method described previously.
2.12. Statistical Analysis
For statistical analyses, experiment designs, and data visualization Minitab 21.4.2, Graphpad prism 9 and Rstudio 4.1 were used. Statistical significance among means was assumed at 5% significance level
3. Results
3.1 Isolation and Quantitative Screening of Phytase-Producing Bacteria
After 72 hours of incubation in the phytate-specific medium, twelve isolates were found positive indicated by clear zone around the colonies. Here, SP11 showed the largest zone ratio (2.3) while SP13 showed the smallest zone ratio (1.3) shown in Table S1 and Figure 1a. The highest activities were seen after 72 hours for all 12 isolates (Figure 1b). Among the isolates, SP11 showed the highest activity (118 U/L) while DM1 showed the least activity (60 U/L) in the broth.
3.2. Identification of the SP11 Isolate and Detection of Full-Length Gene Encoding Phytase
The colonies (Figure 2a) were opaque and creamy-white, flat circular, and large. Microscopic observation showed that SP11 is a gram-positive rod-shaped bacterium (Figure 3b). Biochemical test results are provided in Table S2. Based on these observations, the presumptive identification was Bacillus spp. Amplification of the 1200 bp long gene encoding phytase was done by polymerase chain reaction which is shown by the agarose gel electrophoresis in the second lane (Figure 2c) which confirms the presence of the gene in the strain. Blast analysis of the 16S rDNA sequence showed that the similarity of the 16S rDNA sequence of the strain was as high as 99.85% with one of the Bacillus subtilis strains. Using MEGA 7.0 for cluster analysis, we found that the strain could be clustered with Bacillus subtilis IAM 12118. (Figure 1d). Therefore, the strain was named Bacillus subtilis SP11.
3.3 Single Factor Analysis
The effect of different incubation times, inoculum size, incubation temperature and media pH has been evaluated for phytase production. Phytase production is maximum for 5% inoculum size at 72 hours (Figure 3a) while the optimum pH and temperature are pH 6.5 and 37oC respectively (Figure 3b and 3c). Within the tested periods, the effect of 10% inoculum size is not very sharp and 1% inoculum size didn’t reach its peak, but the time to reach its peak can be safely assumed to be not optimum for the fermentation.
3.4. Determining Significant Variable for Phytase Production Using Plackett-Burman Design:
In order to screen the most significant variables that affect the phytase production by Bacillus subtilis SP11, Plackket-Burman Design (PBD) was utilized with 19 independent variables shown in Table 1.
The experimental design having 24 experimental runs is shown in Table 2 along with the results. Phytase production ranged from 11.05 U/L to 394.57 U/L, a wide range indicating the importance of optimization and finding significant variables.
The coefficients, p-value, and t-statistics obtained by statistical analysis are shown in Table 3 where smaller p-values (at 5% significance level) and larger absolute values of t-statistics (critical value 2.776) indicate the significant variables; positive and negative signs of the coefficients or t-value indicate positive and negative effects respectively.
By analyzing the results, five variables found to be significant where three were positively significant (Mustard Meal, Tryptone, and Yeast Extract) and two were negatively significant (NH4NO3 and sucrose), as depicted in Figure 4a.
The remaining fourteen variables were nonsignificant where some had positive effects (rice bran, wheat bran, soybean meal, linseed meal, cane molasses, glucose, K2HPO4, CaCl2, MgCl2, FeSO4, and MnSO4) while others had negative effect (corn meal, sesame meal and sodium citrate). Maximum production is seen in 4th run where positive significant variables had high values and negative significant variables had low values. The first-order polynomial regression equation representing the production of phytase activity is as follows:
| Y phytase activity | = | 169.09 + 6.56 X1 + 7.73 X2 - 4.55 X3 + 1.83 X4 - 2.45 X5 + 38.22 X6 + 7.01 X7 + 0.57 X8 + 19.14 X9 - 57.08 X10 + 40.87 X11 + 25.40 X12 + 20.15 X13 - 22.77 X14 + 14.04 X15 + 21.80 X16 + 11.50 X17 + 19.41 X18 - 14.24 X19 |
3.5. Evaluation of the Model’s Fitness
ANOVA results (Table 3) show that the model has a p-value of 0.03, indicating that changes in the variables are significantly associated with changes in phytase production. A high R2 value (97.38%) indicates a good fit for the model as it is unable to explain only 2.62% variation in the results. The normal probability plot (Figure 4b) shows that the residuals are near the diagonal line which indicates that they are distributed normally, verified with a p-value of 0.768. Figure 4c presents the plot of predicted vs. observed values of phytase production. Data points that gathered around the diagonal line suggest a good correlation between the predicted and observed values. This shows that predicted phytase production fits well with the observed results. In Figure 4d, predicted values are plotted against the residuals, showing the random but equal distribution of residuals below and above the x-axis, suggesting a lack of any recognizable pattern; thus, it supports the adequacy of the appropriate model. These overall indicate that the model is well-fitted.
3.6. Central Composite Design to Determine the Optimum Amount
Based on the t-values from PBD, three positive significant variables were found to be mustard meal, tryptone, and yeast extract. These 3 components were used in the response surface methodology model to determine their optimum level and interaction. Twenty experimental runs were conducted using the Central composite design, having six central points, fourteen factorial, and six axial points (shown in Table 4). Phytase production varied depending on the composition and ranged from 153.9 U/L to 427 U/L, both occurred in factorial points. The highest activity is achieved when each of the three factors are at a high level and the lowest activity is seen when mustard meal and tryptone are at a low level but yeast extract is at a high level.
3.7. Statistical Analysis of the Central Composite Design
Multiple regression analysis (Table 5) shows that yeast extract (P-value: 0.003) has a critical role in optimization. It also suggests a significant positive interaction between mustard meal and yeast extract (P-value: 0.007), and Tryptone and yeast extract (p value: 0.013). To determine the relationship between phytase production (response) and mustard meal (X6), tryptone (X11), and yeast extract (X12), a second-order polynomial equation is deducted to predict the response (Y) in terms of the independent variables X6, X11, and X12.
The ANOVA (Table 5) suggests that the model is very significant having a high F-value (9.26) and low P-value (0.001). Overall, this CCD's linear, square, and two-way interaction effects are significant, having a p-value less than 0.05. The model has a good fit of data suggested by a ‘lack of fit’ value of 0.061, which indicates the probability of concluding that the model does not explain the data well when it actually does is 6.1%. So, it is not evident that the model doesn’t fit the data considering the 5% significance level. An R2 value of 89.28% suggests that it can explain the 89.28% variability in the results and cannot account for the rest of the 10.72% error.
3.8. Response Surface Plots and Contour Plots
To investigate the interactive effect of the three variables on phytase production, response surface plots and contour plots (Figure 5.) were drawn against any two variables while having another variable fixed at its central level. In response surface plots, the Z-axis refers to the phytase activity (response variable). Figure 5a and 5b demonstrates the interactive effect of mustard meal (X6) and tryptone (X11) on phytase production. Their mutual interaction is antagonistic (negative coefficient), but non-significant (p-value: 0.063) considering a 5% significance level. While yeast extract is held at the central level, it can be seen that phytase production maximizes when the level of mustard meal and tryptone is between moderate to high levels. There is low phytase activity when both are at the lowest level, but when one variable is at the highest level and the other one is at the lowest level, it shows a modest amount of phytase activity. However, when both factors are at the highest level, the production decreases. This suggests that the antagonism between these two factors when yeast extract is moderate, is least evident when both are in the middle of moderate to high levels, but more evident when they are at low levels.
Figure 5c and 5d shows the phytase activity as a function of mustard meal and yeast extract. These two components have significant synergistic effects (p-value: 0.007) on phytase production and there is still a modest amount of phytase activity even when both variables had the lowest level. When tryptone is fixed at the central level, phytase production decreases when mustard meal and yeast extract levels are below moderate levels, while maximum production occurs around the high levels. A similar case is seen in Figure 5e and 5f which indicates a significant synergistic effect of tryptone and yeast extract on phytase production. These graphs show that there is increased phytase activity when yeast extract is present, indicating the critical role of yeast extract in the production of phytase from B. subtilis SP11, also evident from regression analysis (p-value: 0.003, t-value: 3.95). During experimental observations, we got the highest activity at the factorial points where each of the three variables was at a high level; and this can be justified by the plots. The antagonism of mustard meal and tryptone at a high level might have been slightly masked by the synergistic effect of high levels of yeast extract with both of them.
3.9. Verification of the Model
Response optimizer in Minitab was used to find the optimum composition of the three factors for maximum phytase production. The predicted value was 401 U/L for 2.21 % mustard meal, 1.95% tryptone, and 1.8% yeast extract, with 0.91 composite desirability and a 95% confidence level of 337 to 465.3 U/L activity. Experimental runs were conducted to verify the prediction and we obtained 436 U/L which falls within the 95% confidence level. The verification experiment demonstrated that the model efficiently predicted the ideal media composition for phytase production by B. subtilis SP11 with approximately 92% accuracy. Although one of the experimental results (3rd run) in CCD had higher activity than what was predicted to be the maximum, the verification run confirmed the acceptability of the prediction. The reason for this discrepancy is that the regression equation predicts a lower activity for the third run than the observed result.

Table 7.
Predicted optimum media composition for phytase production by CCD
| Mustard Meal | Tryptone | Yeast Extract | Predicted | 95% Cl | Desirability | Observed | Accuracy |
| 2.21 % w/v | 1.95 % w/v | 1.85 % w/v | 401.121 (U/L) | (337 U/L, 465.3 U/L) | 0.91 | 436 U/L | 92% |
3.10. Time Course Profile
Figure 7 demonstrates the kinetics of shake flask fermentation of phytase. The bacterial growth reaches the stationary phase within 24 hours and the death phase at 60 hours as indicated by cell density (absorbance at O.D600). Carbohydrates are rapidly utilized within 12 hours from 12.6 g/L to 2.6 g/L and then maintained at an almost stationary level with a very slight decrease to 1.9 g/L. However, protein content initially increased from 1.5 g/L to 1.67 g/L at 6 hours and slowly decreased afterward to 1 g/L. Both Phytase and protease have peaks (90 U/mL and 435 U/L respectively) during the death phase, at 72 hours and 84 hours respectively.
3.11. Characterization of Crude Phytase
Crude phytase in the cell-free supernatant was characterized for its thermal stability, optimum temperature, and effect of metal ions and EDTA. Figure 7a shows that crude phytase retains more than 70% activity even after being treated at 80o C for 1hour, indicating that it is thermostable. It also shows that crude phytase has an optimum temperature of 50o C while retaining more than 95% of its activity at 41o C. As shown in Figure 7b, the optimum pH of the enzyme is pH 6 and activity greatly decreases below pH 5 and above pH 7. Figure 7c suggests that metal ions like Mg2+, Ca2+, Mn2+, Zn2+ do not affect phytase activity significantly. Cu2+ and Fe2+ on the other hand show an inhibitory effect on phytase. EDTA severely reduces phytase activity, making it less than half of its maximum activity. This indicates that this enzyme requires metal ions as co-factors to function as EDTA chelates metal ions.
3.12. In Vitro Dephytinization of Wheat Bran
The crude phytase released inorganic phosphate from the raw materials in a time-dependent manner. Mustard meal showed the highest phosphate release rate and corn meal showed the least. Within its two hours of incubation, the average velocities of inorganic phosphate release were 896, 725, 870, and 1057 μg/g/h for wheat bran, corn meal, soybean meal, and mustard meal respectively.
4. Discussion
The goal of this study was to isolate a thermostable phytase-producing bacteria from broiler farms and optimize media composition using agro-industrial by-products to maximize phytase production. Twelve Isolates were selected based on the zone ratio in the PSM plate ranging from 1.3 to 2.3. A quantitative screening with the phytase production medium and the earlier zone ratio helped conclude that the SP11 strain is the best phytase producer among these isolated strains. Bacterial identification with microscopic, biochemical, and 16s rDNA characterization revealed that the species is Bacillus subtilis and the presence of the phytase gene was confirmed by PCR amplification. Mussa et al. screened for phytase-producing bacteria from and around poultry farms but found no Bacillus species as a notable producer[24].
To optimize the fermentation protocol including both conditions and media composition, we initially analyzed by changing single fermentation condition per experiment, followed by statistical optimization using Plackett-Burman design (PBD) to identify significant media components and Response Surface Methodology (RSM) to determine the optimum amount. Single factor analysis revealed that phytase production maximally occurs at pH 6.4 and 37oC for 72 hours of fermentation, conditions that were applied later in the experiments. A similar case is seen for another Bacillus species[25] but the optimum temperature (30oC) and incubation time (48h) differed for B. subtilis US417[16] As B. subtilis is a mesophilic bacterium, it is not unusual for the ideal temperature to be 37oC.
Plackett-Burman design (PBD) showed that mustard meal, tryptone, and yeast extract enhance but ammonium nitrate and sucrose inhibit phytase production significantly. The substantial positive effect of yeast extract[16] and tryptone[26] and the negative effect of ammonium nitrate[27] on Bacillus spp. have been previously reported which agrees with our findings. This indicates the critical role of nitrogen sources in phytase production. Mustard meal might have a multifunctional role here, serving as the main source of carbon (crude fiber:12-13%) and phytate (2-3%) and also providing nitrogen (crude protein: 30-32%) and required minerals[29,30]. Our study finds mustard meal as a fermentation media component for the first time as no previous report could be retrieved. However, our findings with sucrose as a significant inhibitor of phytase synthesis contradict previous report of B. subtilis MJA where sucrose was the best carbon source for phytase production[31].
Three positive factors from PBD were used in response surface methodology (RSM) to find the ideal amount. Central composite design (CCD) analysis suggested the optimum amount to be 2.21% mustard meal, 1.95% tryptone, and 1.8% yeast extract for a maximum phytase activity of 436 U/L with a 92% accuracy. Kammoun R. et al.[16] used 5% w/v wheat bran for B. subtilis US417 and found 0.75g yeast extract per gram of wheat bran to be the optimum amount. Our study found a similar ratio (0.81g yeast extract per gram of mustard meal), but the amount is half. However, 0.5% w/v tryptone was optimum for Bacillus sp. HCYL03[26] which is almost one-fourth of our finding with tryptone. This creates a significant limitation of the study as it will increase the cost of production. CCD obtained a 3.7 fold increase in the phytase production compared to initial unoptimized media. The high amount of required tryptone, yeast extract, and mustard meal along with the inhibition by ammonium nitrate and no dependence on simple carbon sources like cane molasses, glucose, or sucrose suggests that this strain’s phytase production is highly associated with utilization of polysaccharides as well as protein and peptide.
Monitoring the cell growth, nutrient utilization, and enzyme production were done at various time points up to 96 hours. Choi et al. reported that phytase production in Bacillus spp. is increased in the stationary phase[28]. However, Figure 7c shows that B. subtilis SP11 has the highest phytase activity at the death phase. Their study also suggested that phytase production may be related to nutrient limitation, which gives an idea about the inhibition of ammonium nitrate and sucrose on the phytase production by B. subtilis SP11 while yeast extract, tryptone, and complex substrates like mustard meal enhance it. The initial increase of protein concentration may be due to the release of protein from mustard meal and then with the rise of protease activity it started getting utilized.
McCapes et al. suggested a combination of 85.7oC conditioning temperature and 4.1 min heating time for effective feed pelleting processes[32] due to which the feed enzymes need to be thermostable. Our crude phytase being thermostable, as shown in Figure 7a, is thus compatible with the industrial feed pelleting process. The optimum temperature for the crude phytase of this strain is 50oC which is close to various previous reports stating 45oC[24] to 55oC[33]. It retains more than 95% of its activity at the body temperature of a chicken, which is near 41oC, thus ideal for poultry application[34]. Crude phytase from B. subtilis SP11 shows optimum pH of 6 which agrees with the study with B. subtilis B.S.46[35]. Nonetheless, their study showed high phytase activity even at pH 10 and no activity below pH 5 whereas our crude phytase is active in pH range 3-8. The pH of the crop and duodenum of different poultry birds is near 6[36,37], making this enzyme suitable for poultry. Figure 7c suggests that the enzyme requires metal ions as cofactors because EDTA, which chelates metal ions, causes a massive decrease in phytase activity. This supports the mechanism of action of beta-propeller phytase, found in B. subtilis, requiring calcium ions for its action[38] . However, Mg2+, Zn2+and Mn2+ do not affect phytase activity significantly which opposed the result shown by Rocky-Salimi et al. where metal ions inhibited phytase[34]. Fe2+ and Cu2+ inhibiting crude phytase may be due to the complex formation of phytate with these ions that leads to insufficient binding to the active site and poor substrate availability[39].
Although, the final optimized production was still lower than many of the reports of other bacteria and fungi, Bangladesh however, like many other countries, does not locally produce phytase and relies on imports from other countries[18] which is why this holds an opportunity for the local poultry application and food security. To test the applicability of crude crude phtase, In vitro dephytinization of different raw materials such as corn meal, soybean meal, wheat bran and mustard meal were conducted. It shows that the crude phytase released higher phytate phosphates per gram of substrate per hour (896 to 1057 μg) than some of the previous reports from Bacillus subtilis (759 μg), indicating its applicability in poultry feed dephytinization[40]. The highest release seen in mustard meal may explain why it was the best substrate for phytase production.
5. Conclusions
Bacillus subtilis SP11 produces thermostable phytase for which media formulations are optimized utilizing agro-industrial by-product mustard meal. The strain’s ability to retain enzyme activity at industrial as well as poultry physiological conditions and its higher dephytinization activity holds industrial potential for application in poultry feed. These findings can help mitigate feed enzyme import reliance in Bangladesh and address sustainable poultry farms.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Zone ratio (Z) of various isolates in Phytate Specific Medium (PSM), qualitatively indicating the production of phytase; Table S2: Biochemical test results of SP11 isolate
Author Contributions
Conceptualization, S.N.K, M.M. and M.A.M.K., ; methodology, S., M.M.M., M.A.M.K and T.A.; software, M.A.M.K, M.M; validation, S.N.K., M.A.M.K.; formal analysis, M.M.K; investigation, S.N.K., M.M.K., M.A.A.M. ; resources, S.N.K., M.M.K., M.A.A.M. ; data curation, M.A.M.K. ; writing—original draft preparation, M.A.M.K.; writing—review and editing, S.N.K., M.M.K., and M.A.A.M., ; visualization, M.A.M.K; supervision, S.N.K; project administration, S.N.K.; All authors have read and agreed to the published version of the manuscript.”
Funding
This research received no external funding
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflicts of interest
References
- Haefner, S.; Knietsch, A.; Scholten, E.; Braun, J.; Lohscheidt, M.; Zelder, O. Biotechnological Production and Applications of Phytases. Appl Microbiol Biotechnol 2005, 68, 588–597. [CrossRef]
- Greiner, R.; Egli, I. Determination of the Activity of Acidic Phytate-Degrading Enzymes in Cereal Seeds. J Agric Food Chem 2003, 51, 847–850. [CrossRef]
- Charles Bai, S.; Hardy, R.W.; Hamidoghli, A. Diet Analysis and Evaluation. In Fish Nutrition; Hardy, R.W., Kaushik, S.J., Eds.; Elsevier, 2022; pp. 709–743.
- Kumar, A.; Singh, B.; Raigond, P.; Sahu, C.; Mishra, U.N.; Sharma, S.; Lal, M.K. Phytic Acid: Blessing in Disguise, a Prime Compound Required for Both Plant and Human Nutrition. Food Research International 2021, 142, 110193. [CrossRef]
- Dersjant-Li, Y.; Awati, A.; Schulze, H.; Partridge, G. Phytase in Non-ruminant Animal Nutrition: A Critical Review on Phytase Activities in the Gastrointestinal Tract and Influencing Factors. J Sci Food Agric 2015, 95, 878–896. [CrossRef]
- Joudaki, H.; Aria, N.; Moravej, R.; Rezaei Yazdi, M.; Emami-Karvani, Z.; Hamblin, M.R. Microbial Phytases: Properties and Applications in the Food Industry. Curr Microbiol 2023, 80, 374. [CrossRef]
- Valente Junior, D.T.; Genova, J.L.; Kim, S.W.; Saraiva, A.; Rocha, G.C. Carbohydrases and Phytase in Poultry and Pig Nutrition: A Review beyond the Nutrients and Energy Matrix. Animals 2024, 14, 226. [CrossRef]
- Mullaney, E.J.; Ullah, A.H.J. The Term Phytase Comprises Several Different Classes of Enzymes. Biochem Biophys Res Commun 2003, 312, 179–184. [CrossRef]
- Mullaney, E.J.; Daly, C.B.; Ullah, A.H.J. Advances in Phytase Research. In; 2000; pp. 157–199.
- Balaban, N.P.; Suleimanova, A.D.; Shakirov, E. V.; Sharipova, M.R. Histidine Acid Phytases of Microbial Origin. Microbiology (N Y) 2018, 87, 745–756. [CrossRef]
- Kumar, V.; Yadav, A.N.; Verma, P.; Sangwan, P.; Saxena, A.; Kumar, K.; Singh, B. β-Propeller Phytases: Diversity, Catalytic Attributes, Current Developments and Potential Biotechnological Applications. Int J Biol Macromol 2017, 98, 595–609. [CrossRef]
- Hegeman, C.E.; Grabau, E.A. A Novel Phytase with Sequence Similarity to Purple Acid Phosphatases Is Expressed in Cotyledons of Germinating Soybean Seedlings. Plant Physiol 2001, 126, 1598–1608. [CrossRef]
- Puhl, A.A.; Gruninger, R.J.; Greiner, R.; Janzen, T.W.; Mosimann, S.C.; Selinger, L.B. Kinetic and Structural Analysis of a Bacterial Protein Tyrosine Phosphatase-like Myo -inositol Polyphosphatase. Protein Science 2007, 16, 1368–1378. [CrossRef]
- Konietzny, U.; Greiner, R. Molecular and Catalytic Properties of Phytate-degrading Enzymes (Phytases). Int J Food Sci Technol 2002, 37, 791–812. [CrossRef]
- Chen, C.; Cheng, K.; Ko, T.; Guo, R. Current Progresses in Phytase Research: Three-Dimensional Structure and Protein Engineering. ChemBioEng Reviews 2015, 2, 76–86. [CrossRef]
- Kammoun, R.; Farhat, A.; Chouayekh, H.; Bouchaala, K.; Bejar, S. Phytase Production by Bacillus Subtilis US417 in Submerged and Solid State Fermentations. Ann Microbiol 2012, 62, 155–164. [CrossRef]
- Animal Feed Phytase Market Size, Share & Trends Analysis Report By Form (Liquid, Granules or Powder), By Class (3-Phytase, 6-Phytase, Others), By Source (Plants, Microorganisms), By Livestock, By Region, And by Segment Forecasts, 2023-2031.
- Phytase Import Data of Bangladesh - 191 Import Shipments.
- Howson, S.J.; Davis, R.P. Production of Phytate-Hydrolysing Enzyme by Some Fungi. Enzyme Microb Technol 1983, 5, 377–382. [CrossRef]
- Bae, H.D.; Yanke, L.J.; Cheng, K.-J.; Selinger, L.B. A Novel Staining Method for Detecting Phytase Activity. J Microbiol Methods 1999, 39, 17–22. [CrossRef]
- Li, G.-E.; Wu, X.-Q.; Ye, J.-R.; Hou, L.; Zhou, A.-D.; Zhao, L. Isolation and Identification of Phytate-Degrading Rhizobacteria with Activity of Improving Growth of Poplar and Masson Pine. World J Microbiol Biotechnol 2013, 29, 2181–2193. [CrossRef]
- Bravo, A.; Sarabia, S.; Lopez, L.; Ontiveros, H.; Abarca, C.; Ortiz, A.; Ortiz, M.; Lina, L.; Villalobos, F.J.; Peña, G.; et al. Characterization of Cry Genes in a Mexican Bacillus Thuringiensis Strain Collection. Appl Environ Microbiol 1998, 64, 4965–4972. [CrossRef]
- Suresh, S.; Radha, K. Effect of a Mixed Substrate on Phytase Production by Rhizopus Oligosporus MTCC 556 Using Solid State Fermentation and Determination of Dephytinization Activities in Food Grains. Food Sci Biotechnol 2015, 24, 551–559. [CrossRef]
- Mussa, L.A.; Yadetie, D.M.; Temesgen, E.A.; Tefera, A.T.; Gemeda, M.T. Isolation and In-Vitro Characterization of Extracellular Phytase Producing Bacterial Isolates for Potential Application in Poultry Feed. BMC Microbiol 2023, 23, 296. [CrossRef]
- Suliasih, S.; Widawati, S.; Ikhwani, A.; Suyadi, S.; Sudiana, I.M. Phytase Activity of Phytase-Producing Bacteria Isolated from Mangrove Sediment. IOP Conf Ser Earth Environ Sci 2022, 976, 12041. [CrossRef]
- Sardar, R.; Asad, M.; Ahmad, M.; Ahmad, T. Optimization of Phytase Production by Bacillus Sp. (HCYL03) under Solid-State Fermentation by Using Box-Behnken Design. Brazilian Archives of Biology and Technology 2022, 65. [CrossRef]
- Rocky-Salimi, K.; Hashemi, M.; Safari, M.; Mousivand, M. Valorisation of Untreated Cane Molasses for Enhanced Phytase Production by Bacillus Subtilis K46b and Its Potential Role in Dephytinisation. J Sci Food Agric 2017, 97, 222–229. [CrossRef]
- Choi, Y.M.; Suh, H.J.; Kim, J.M. Purification and Properties of Extracellular Phytase from Bacillus Sp. KHU-10. J Protein Chem 2001, 20, 287–292. [CrossRef]
- Cheva-Isarakul, B.; Tangtaweewipat, S.; Sangsrijun, P.; Yamauchi, K. Chemical Composition and Metabolizable Energy of Mustard Meal. J Poult Sci 2003, 40, 221–225. [CrossRef]
- Sarker, A.K.; Saha, D.; Begum, H.; Zaman, A.; Rahman, M.M. Comparison of Cake Compositions, Pepsin Digestibility and Amino Acids Concentration of Proteins Isolated from Black Mustard and Yellow Mustard Cakes. AMB Express 2015, 5, 22. [CrossRef]
- El-Toukhy, N.M.K.; Youssef, A.S.; Mikhail, M. Isolation, Purification and Characterization of Phytase from Bacillus Subtilis MJA. Afr J Biotechnol 2013, 12.
- McCapes, R.H.; Ekperigin, H.E.; Cameron, W.J.; Ritchie, W.L.; Slagter, J.; Stangeland, V.; Nagaraja, K. V Effect of a New Pelleting Process on the Level of Contamination of Poultry Mash by Escherichia Coli and Salmonella. Avian Dis 1989, 33, 103–111.
- Kerovuo, J.; Lauraeus, M.; Nurminen, P.; Kalkkinen, N.; Apajalahti, J. Isolation, Characterization, Molecular Gene Cloning, and Sequencing of a Novel Phytase from Bacillus Subtilis. Appl Environ Microbiol 1998, 64, 2079–2085. [CrossRef]
- Troxell, B.; Petri, N.; Daron, C.; Pereira, R.; Mendoza, M.; Hassan, H.M.; Koci, M.D. Poultry Body Temperature Contributes to Invasion Control through Reduced Expression of Salmonella Pathogenicity Island 1 Genes in Salmonella Enterica Serovars Typhimurium and Enteritidis. Appl Environ Microbiol 2015, 81, 8192–8201. [CrossRef]
- Rocky-Salimi, K.; Hashemi, M.; Safari, M.; Mousivand, M. A Novel Phytase Characterized by Thermostability and High PH Tolerance from Rice Phyllosphere Isolated Bacillus Subtilis B.S.46. J Adv Res 2016, 7, 381–390. [CrossRef]
- Kierończyk, B.; Rawski, M.; Długosz, J.; Swiatkiewicz, S.; Józefiak, D. Avian Crop Function – A Review. Annals of Animal Science 2016, 16, 653–678. [CrossRef]
- Farner, D.S. The Hydrogen Ion Concentration in Avian Digestive Tracts. Poult Sci 1942, 21, 445–450. [CrossRef]
- Shin, S.; Ha, N.-C.; Oh, B.-C.; Oh, T.-K.; Oh, B.-H. Enzyme Mechanism and Catalytic Property of β Propeller Phytase. Structure 2001, 9, 851–858. [CrossRef]
- Zhao, T.; Yong, X.; Zhao, Z.; Dolce, V.; Li, Y.; Curcio, R. Research Status of Bacillus Phytase. 3 Biotech 2021, 11, 415. [CrossRef]
- Jain, J.; Singh, B. Phytase Production and Development of an Ideal Dephytinization Process for Amelioration of Food Nutrition Using Microbial Phytases. Appl Biochem Biotechnol 2017, 181, 1485–1495. [CrossRef]
Figure 1.
Isolation and screening of phytase-producing bacteria. (a) Isolation of phytase producing bacteria in phytate specific medium (PSM) plate. The halo zones indicate zone of phytate degradation. (b) Phytase production by the bacteria in the phytase production medium (broth) was measured at 24-hour intervals for 3 days
Figure 1.
Isolation and screening of phytase-producing bacteria. (a) Isolation of phytase producing bacteria in phytate specific medium (PSM) plate. The halo zones indicate zone of phytate degradation. (b) Phytase production by the bacteria in the phytase production medium (broth) was measured at 24-hour intervals for 3 days
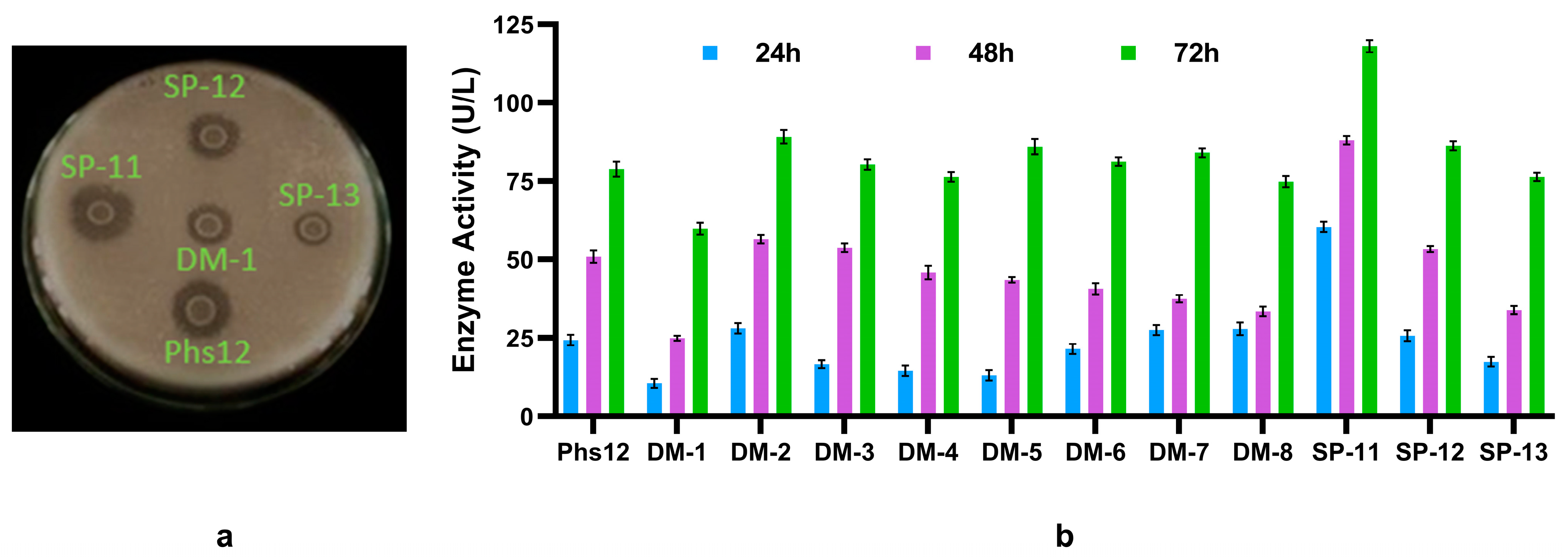
Figure 2.
Identification of the phytase-producing SP11 isolate. (a) Colony morphology of SP11 isolate (b) 100x microscopic observation of SP11 (c) PCR confirmation of phytase gene (d) 16s rDNA-based phylogenetic tree showing SP11 as a Bacillus subtilis species
Figure 2.
Identification of the phytase-producing SP11 isolate. (a) Colony morphology of SP11 isolate (b) 100x microscopic observation of SP11 (c) PCR confirmation of phytase gene (d) 16s rDNA-based phylogenetic tree showing SP11 as a Bacillus subtilis species
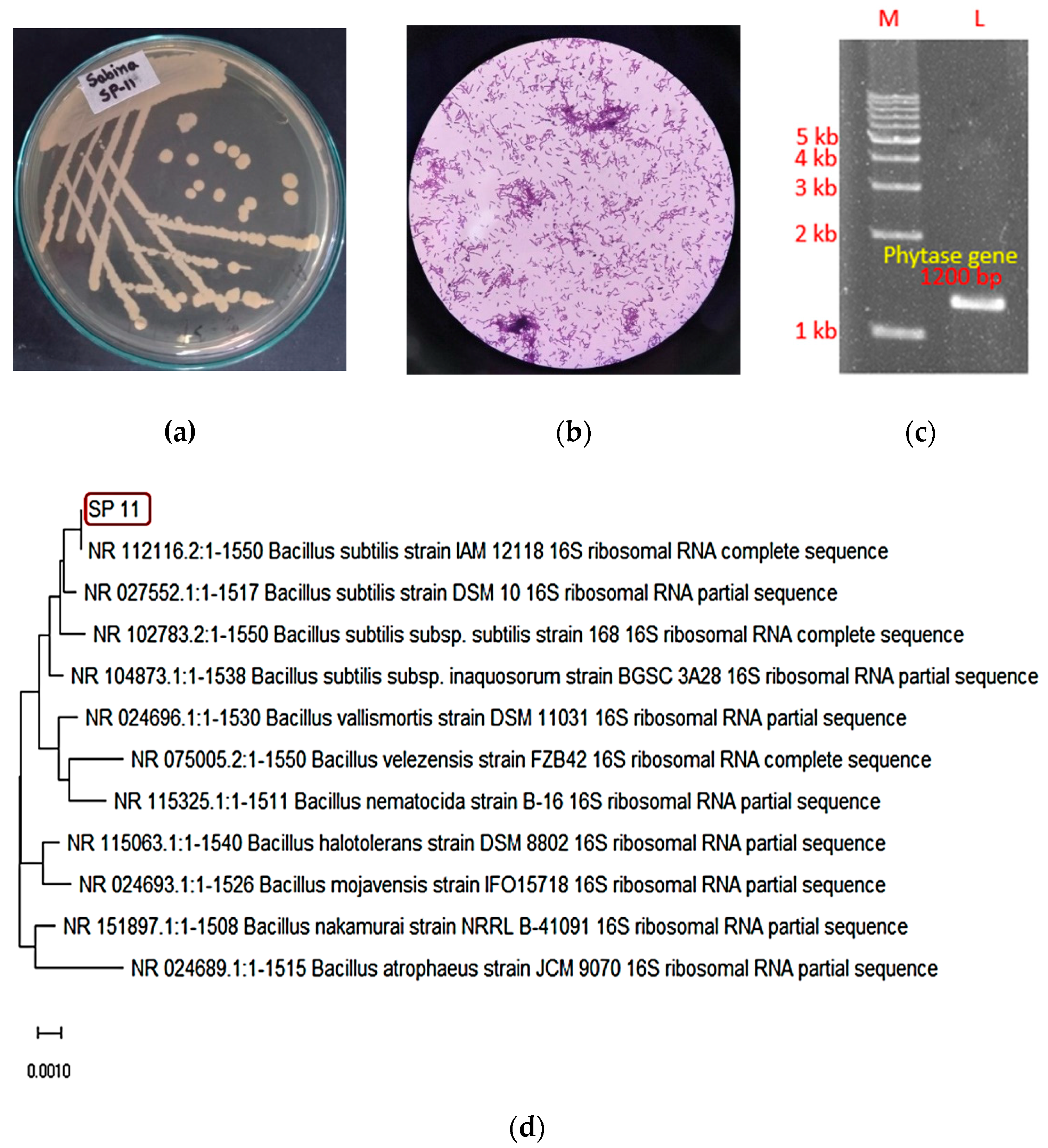
Figure 3.
Factors influencing the production of phytase. Effect of (a) incubation time and inoculum size (b) various media pH and (c) different temperatures. Relative activity has been measured compared to the highest observed value for a particular experiment.
Figure 3.
Factors influencing the production of phytase. Effect of (a) incubation time and inoculum size (b) various media pH and (c) different temperatures. Relative activity has been measured compared to the highest observed value for a particular experiment.
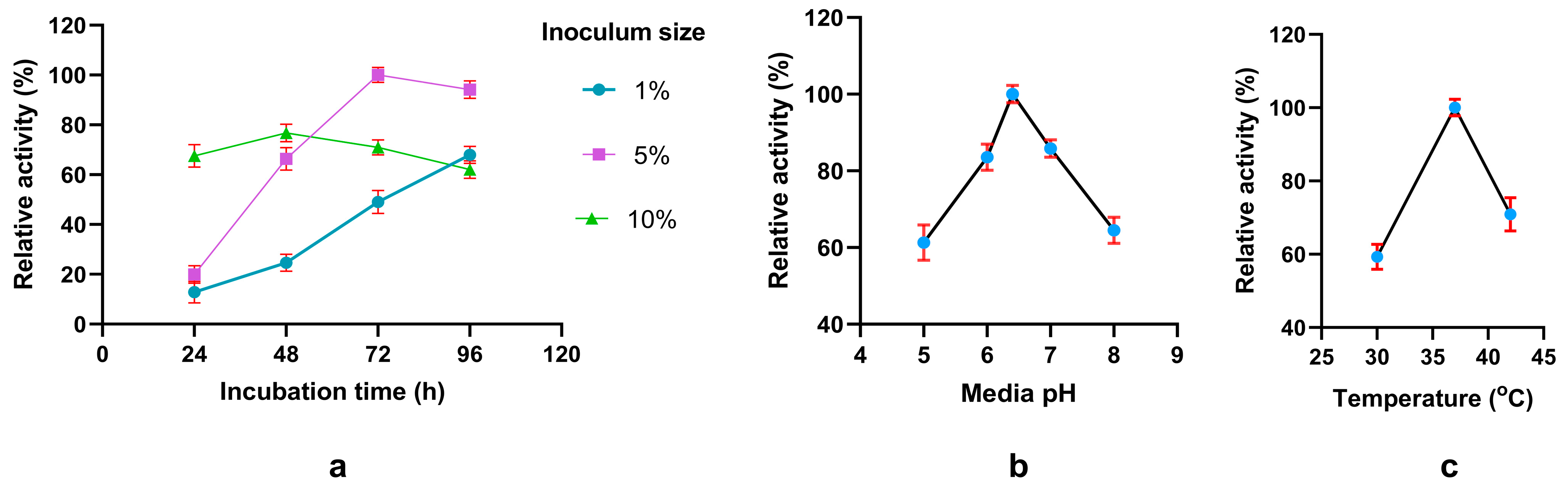
Figure 4.
Analysis of results obtained by placket-burman design. (a) Pareto chart of standardized effect (absolute values of t-statistics) draws a reference line (α) on the chart. Any effect greater than α is statistically significant). Evaluation of the model’s fitness (b, c, and d) where the red region indicates 95% confidence bands. (b) Normal probability plot of residuals for phytase production. (c) Correlation between observed value and predicted values, (d) Residual versus predicted values show the equal distribution of residuals along both sides of the y-axis with no recognizable pattern.
Figure 4.
Analysis of results obtained by placket-burman design. (a) Pareto chart of standardized effect (absolute values of t-statistics) draws a reference line (α) on the chart. Any effect greater than α is statistically significant). Evaluation of the model’s fitness (b, c, and d) where the red region indicates 95% confidence bands. (b) Normal probability plot of residuals for phytase production. (c) Correlation between observed value and predicted values, (d) Residual versus predicted values show the equal distribution of residuals along both sides of the y-axis with no recognizable pattern.
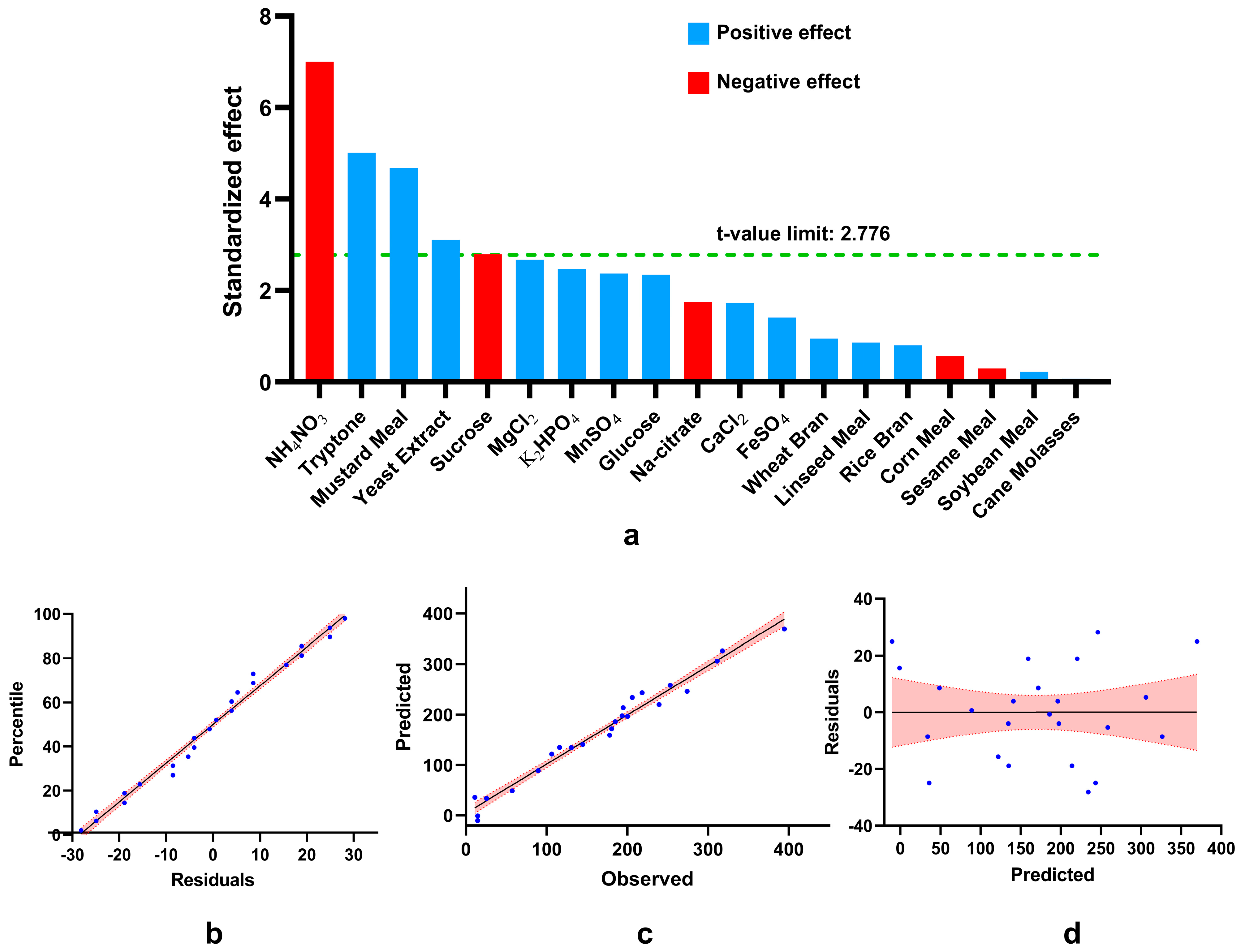
Figure 5.
Response surface plots and contour plots showing how the tryptone, mustard meal, and yeast extract composition affect the production of phytase. (a) Surface plot and (b) contour plot of mustard meal vs tryptone (yeast extract held at 1.1 %), (c) Surface plot and (d) contour plot of mustard meal vs yeast extract (tryptone held at 1.4 %), (e) Surface plot and (f) contour plot of tryptone vs yeast extract (mustard meal held at 1.6%).
Figure 5.
Response surface plots and contour plots showing how the tryptone, mustard meal, and yeast extract composition affect the production of phytase. (a) Surface plot and (b) contour plot of mustard meal vs tryptone (yeast extract held at 1.1 %), (c) Surface plot and (d) contour plot of mustard meal vs yeast extract (tryptone held at 1.4 %), (e) Surface plot and (f) contour plot of tryptone vs yeast extract (mustard meal held at 1.6%).
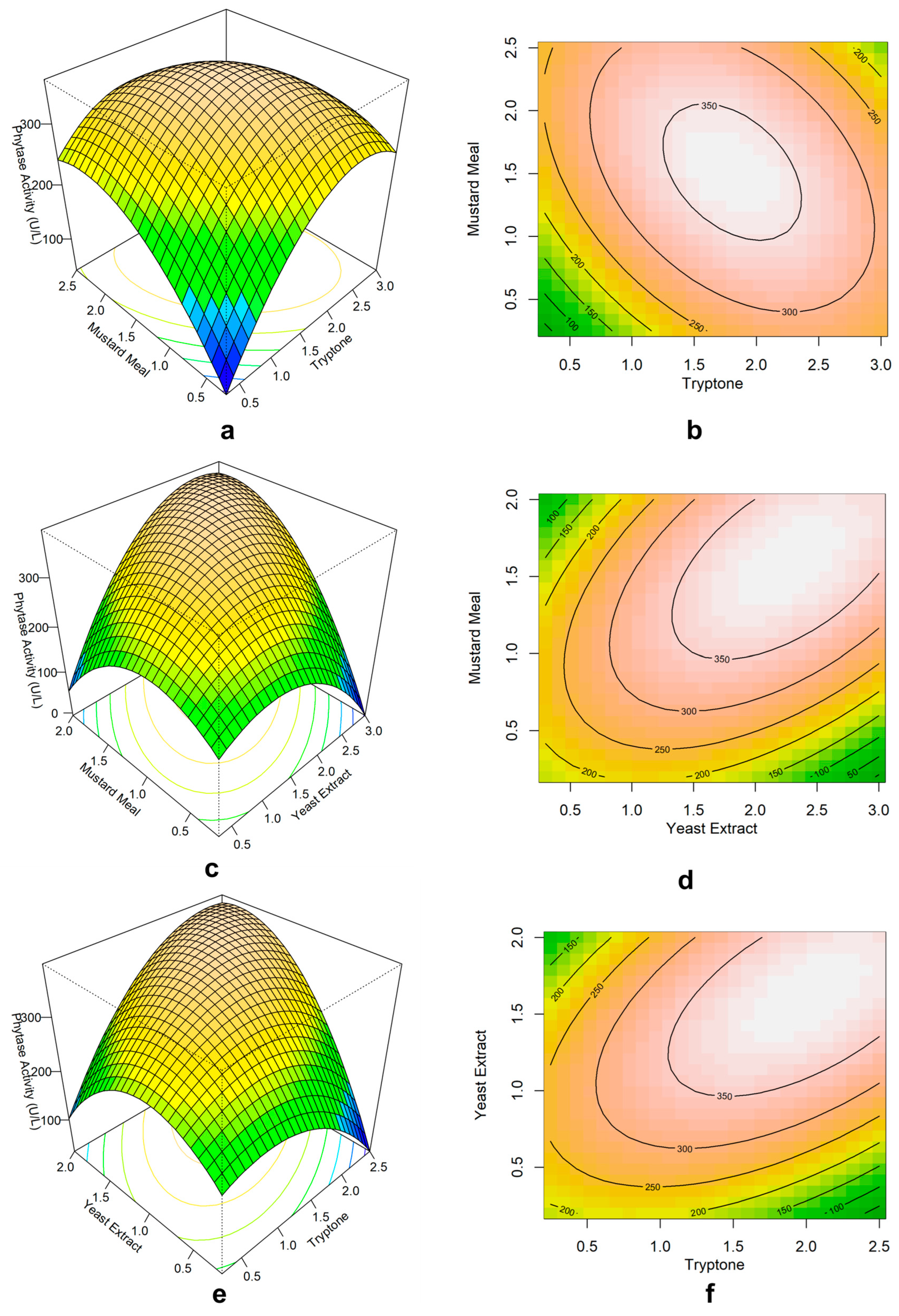
Figure 6.
Time course profile of Cell growth and enzyme production by B. subtilis SP11.
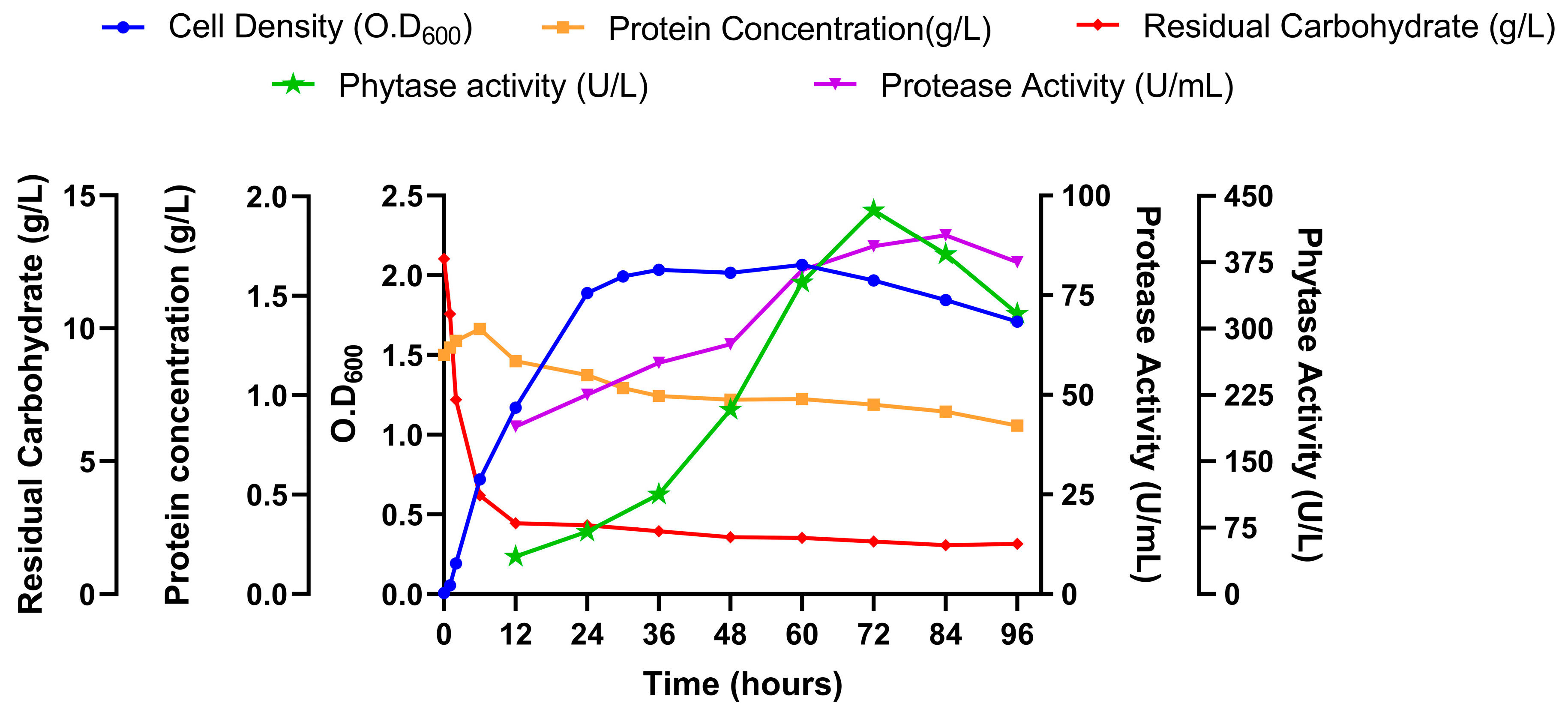
Figure 7.
Characterization of the crude phytase. (a) Thermal stability (activity relative to untreated control) and temperature optima. (b) pH optima and (c) Effect of metal ions and EDTA (activity relative to untreated control). Different letters differ at 5% significance level at turkey’s test
Figure 7.
Characterization of the crude phytase. (a) Thermal stability (activity relative to untreated control) and temperature optima. (b) pH optima and (c) Effect of metal ions and EDTA (activity relative to untreated control). Different letters differ at 5% significance level at turkey’s test

Figure 8.
In vitro dephytinization of wheat bran, corn meal, soybean meal and mustard meal using crude phytase.
Figure 8.
In vitro dephytinization of wheat bran, corn meal, soybean meal and mustard meal using crude phytase.
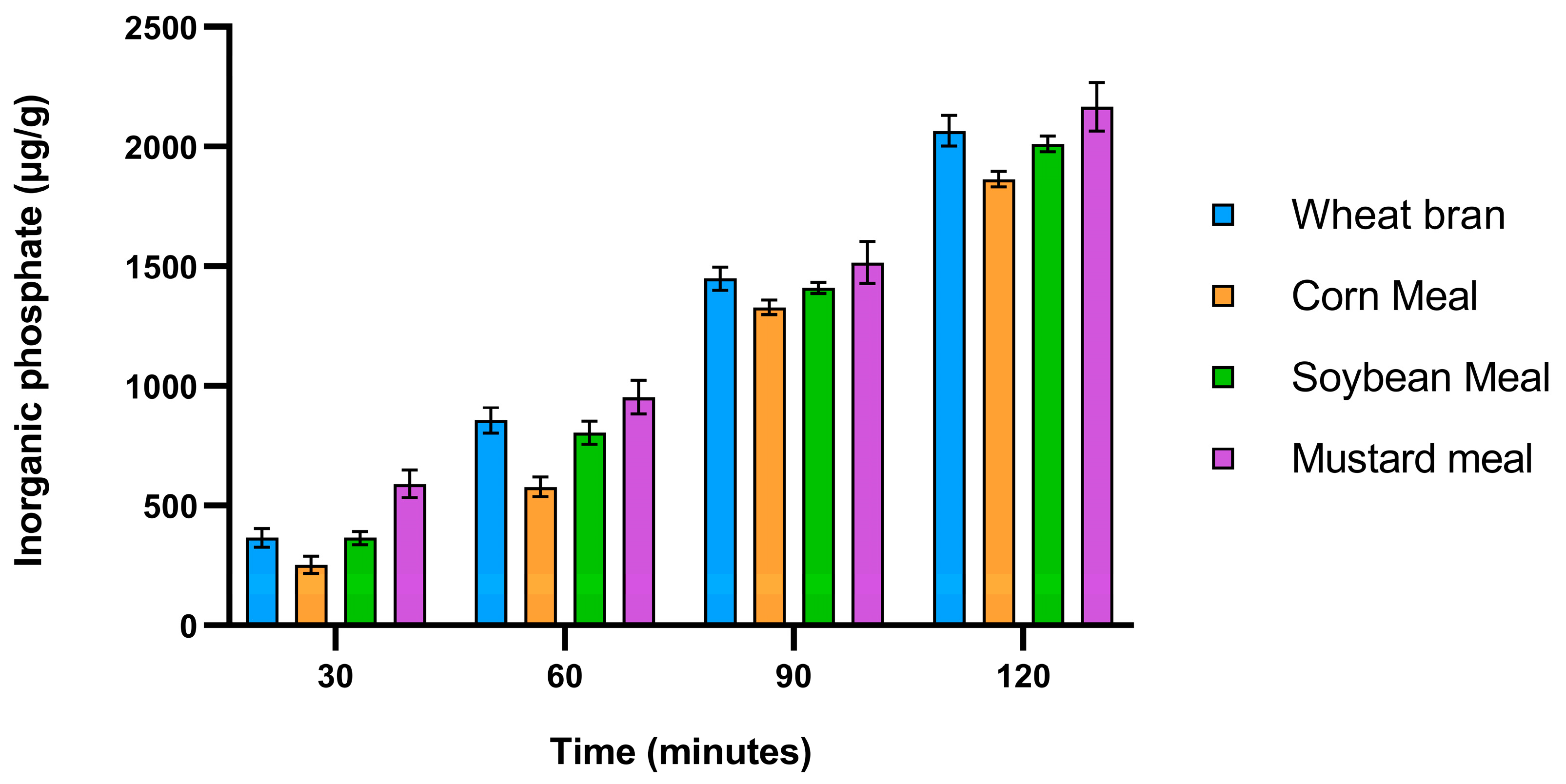
Table 1.
Nineteen Independent variables used for Plackett-Burman Design for the optimization of phytase production by Bacillus subtilis SP11 having two levels: High (+1) and Low (-1).
Table 1.
Nineteen Independent variables used for Plackett-Burman Design for the optimization of phytase production by Bacillus subtilis SP11 having two levels: High (+1) and Low (-1).
| Code | Variable | Low Value (-1) (%w/v) | High Value (+1) (% w/v) |
| X1 | Rice Bran | 0 | 1 |
| X2 | Wheat Bran | 0 | 1 |
| X3 | Corn Meal | 0 | 1 |
| X4 | Soybean Meal | 0 | 1 |
| X5 | Sesame Meal | 0 | 1 |
| X6 | Mustard Meal | 0 | 1 |
| X7 | Linseed Meal | 0 | 1 |
| X8 | Cane Molasses | 0 | 2 |
| X9 | Glucose | 0.4% | 1 |
| X10 | NH4NO3 | 0.2% | 0.5 |
| X11 | Tryptone | 0 | 1 |
| X12 | Yeast Extract | 0 | 0.5 |
| X13 | K2HPO4 | 0.02% | 0.1 |
| X14 | Sucrose | 0 | 1 |
| X15 | CaCl2 | 0.002% | 0.02 |
| X16 | MgCl2 | 0.002% | 0.02 |
| X17 | FeSO4 | 0.002% | 0.02 |
| X18 | MnSO4 | 0.002% | 0.02 |
| X19 | Sodium Citrate | 0 | 1 |
Table 2.
Twenty-four experimental runs for Plackett-Burman Design with nineteen different variables with Low and High Values and their phytase activity. Red and green colored boxes indicate low value (-1) and high value (+1) respectively
Table 2.
Twenty-four experimental runs for Plackett-Burman Design with nineteen different variables with Low and High Values and their phytase activity. Red and green colored boxes indicate low value (-1) and high value (+1) respectively
| Run | A | B | C | D | E | F | G | H | J | K | L | M | N | O | P | Q | R | S | T | Phytase activity (U/L) | ||
| Observed | Predicted | Residuals | ||||||||||||||||||||
| 1 | 89.59 | 88.868 | 0.7225 | |||||||||||||||||||
| 2 | 194.99 | 213.874 | -18.8842 | |||||||||||||||||||
| 3 | 180.54 | 171.998 | 8.5425 | |||||||||||||||||||
| 4 | 394.57 | 369.679 | 24.8908 | |||||||||||||||||||
| 5 | 185.13 | 185.852 | -0.7225 | |||||||||||||||||||
| 6 | 57.29 | 48.748 | 8.5425 | |||||||||||||||||||
| 7 | 253.13 | 258.414 | -5.2842 | |||||||||||||||||||
| 8 | 317.90 | 326.443 | -8.5425 | |||||||||||||||||||
| 9 | 193.63 | 197.611 | -3.9808 | |||||||||||||||||||
| 10 | 11.05 | 35.941 | -24.8908 | |||||||||||||||||||
| 11 | 218.45 | 243.341 | -24.8908 | |||||||||||||||||||
| 12 | 144.84 | 140.859 | 3.9808 | |||||||||||||||||||
| 13 | 14.62 | -1.006 | 15.6258 | |||||||||||||||||||
| 14 | 130.56 | 134.541 | -3.9808 | |||||||||||||||||||
| 15 | 106.25 | 121.876 | -15.6258 | |||||||||||||||||||
| 16 | 178.16 | 159.276 | 18.8842 | |||||||||||||||||||
| 17 | 25.50 | 34.043 | -8.5425 | |||||||||||||||||||
| 18 | 116.11 | 134.994 | -18.8842 | |||||||||||||||||||
| 19 | 239.36 | 220.476 | 18.8842 | |||||||||||||||||||
| 20 | 206.04 | 234.189 | -28.1492 | |||||||||||||||||||
| 21 | 274.21 | 246.061 | 28.1492 | |||||||||||||||||||
| 22 | 311.44 | 306.156 | 5.2842 | |||||||||||||||||||
| 23 | 14.62 | -10.271 | 24.8908 | |||||||||||||||||||
| 24 | 200.26 | 196.279 | 3.9808 | |||||||||||||||||||
Table 3.
Statistical analysis of Plackett–Burman design experimental results
| Term | Coefficients | t-statistics | F-value | p-value | Contribution |
| Constant | 169.09 | 20.73 | 0.000 | ||
| Rice Bran | 6.56 | 0.8 | 0.65 | 0.466 | 0.42% |
| Wheat Bran | 7.73 | 0.95 | 0.9 | 0.397 | 0.59% |
| Corn Meal | -4.55 | -0.56 | 0.31 | 0.607 | 0.20% |
| Soybean Meal | 1.83 | 0.22 | 0.05 | 0.834 | 0.03% |
| Sesame Meal | -2.45 | -0.3 | 0.09 | 0.779 | 0.06% |
| Mustard Meal | 38.22 | 4.68 | 21.95 | 0.009 | 14.38% |
| Linseed Meal | 7.01 | 0.86 | 0.74 | 0.439 | 0.48% |
| Cane Molasses | 0.57 | 0.07 | 0 | 0.948 | 0.00% |
| Glucose | 19.14 | 2.35 | 5.5 | 0.079 | 3.61% |
| NH4NO3 | -57.08 | -7 | 48.94 | 0.002 | 32.07% |
| Tryptone | 40.87 | 5.01 | 25.1 | 0.007 | 16.45% |
| Yeast Extract | 25.4 | 3.11 | 9.69 | 0.036 | 6.35% |
| K2HPO4 | 20.15 | 2.47 | 6.1 | 0.069 | 4.00% |
| Sucrose | -22.77 | -2.79 | 7.79 | 0.049 | 5.10% |
| CaCl2 | 14.04 | 1.72 | 2.96 | 0.16 | 1.94% |
| MgCl2 | 21.8 | 2.67 | 7.14 | 0.056 | 4.68% |
| FeSO4 | 11.5 | 1.41 | 1.99 | 0.231 | 1.30% |
| MnSO4 | 19.41 | 2.38 | 5.66 | 0.076 | 3.71% |
| Na-citrate | -14.24 | -1.75 | 3.05 | 0.156 | 2.00% |
| Analysis of Variance (ANOVA) | |||||
| Source | DF | Adj SS | Adj MS | F-Value | P-Value |
| Model | 19 | 237392 | 12494 | 7.82 | 0.030 |
| Residuals | 4 | 8840 | 2210 | ||
| R2 = 97.38% | |||||
Table 4.
Central Composite Design for phytase production by Bacillus subtilis SP11 along with results and residuals. For axial points α =1.68, Low value=-1, Medium=0 and High value=+1
Table 4.
Central Composite Design for phytase production by Bacillus subtilis SP11 along with results and residuals. For axial points α =1.68, Low value=-1, Medium=0 and High value=+1
| Run | Mustard Meal (X6) | Tryptone (X11) | Yeast Extract (X12) | Phytase Activity (U/L) | |||
| Observed | Predicted | Residual | |||||
| 1 | 0 | 0 | 0 | 388.8 | 361.9 | 26.8 | |
| 2 | 1 | -1 | -1 | 257.2 | 244.1 | 13.1 | |
| 3 | 1 | 1 | 1 | 427.0 | 386.5 | 40.5 | |
| 4 | -1 | -1 | -1 | 219.2 | 236.4 | -17.2 | |
| 5 | 0 | 0 | 0 | 374.7 | 361.9 | 12.8 | |
| 6 | 0 | 0 | 0 | 378.7 | 361.9 | 16.8 | |
| 7 | -1 | 1 | 1 | 323.6 | 313.4 | 10.2 | |
| 8 | 0 | 0 | -1.68 | 199.4 | 189.3 | 10.1 | |
| 9 | -1 | 1 | -1 | 231.8 | 246.4 | -14.6 | |
| 10 | 0 | 0 | 1.68 | 274.6 | 317.7 | -43.1 | |
| 11 | 0 | 0 | 0 | 353.8 | 361.9 | -8.2 | |
| 12 | 0 | 0 | 0 | 349.7 | 361.9 | -12.3 | |
| 13 | 1.68 | 0 | 0 | 232.0 | 287.4 | -55.4 | |
| 14 | -1 | -1 | 1 | 153.9 | 151.2 | 2.7 | |
| 15 | 1 | -1 | 1 | 367.7 | 329.8 | 37.9 | |
| 16 | 1 | 1 | -1 | 169.2 | 148.6 | 20.6 | |
| 17 | 0 | 1.68 | 0 | 287.7 | 310.2 | -22.5 | |
| 18 | -1.68 | 0 | 0 | 242.0 | 219.5 | 22.5 | |
| 19 | 0 | 0 | 0 | 331.6 | 361.9 | -30.3 | |
| 20 | 0 | -1.68 | 0 | 243.7 | 254.1 | -10.5 | |
| Level | Mustard meal (% w/v) |
Tryptone (% w/v) |
Yeast extract (% w/v) |
||||
| - α | 0.3 | 0.25 | 0.2 | ||||
| -1 | 0.85 | 0.71 | 0.57 | ||||
| 0 | 1.65 | 1.38 | 1.1 | ||||
| +1 | 2.45 | 2.04 | 1.64 | ||||
| + α | 3 | 2.5 | 2 | ||||
Table 5.
Multiple regression analysis and ANOVA of the central composite design.
| Term | Coefficient | t-statistics | p-value |
|---|---|---|---|
| Constant | 361.9 | 24.86 | 0.000 |
| Mustard Meal | 20.19 | 2.09 | 0.063 |
| Tryptone | 16.66 | 1.72 | 0.115 |
| Yeast Extract | 38.16 | 3.95 | 0.003 |
| Mustard Meal*Mustard Meal | -38.35 | -4.08 | 0.002 |
| Tryptone*Tryptone | -28.2 | -3 | 0.013 |
| Yeast Extract*Yeast Extract | -38.33 | -4.08 | 0.002 |
| Mustard Meal*Tryptone | -26.4 | -2.09 | 0.063 |
| Mustard Meal*Yeast Extract | 42.7 | 3.38 | 0.007 |
| Tryptone*Yeast Extract | 38.1 | 3.02 | 0.013 |
| Source | DF | F-value | p-value |
| Model | 9 | 9.26 | 0.001 |
| Linear | 3 | 7.65 | 0.006 |
| Square | 3 | 11.82 | 0.001 |
| 2-Way Interaction | 3 | 8.3 | 0.005 |
| Lack-of-Fit | 5 | 4.55 | 0.061 |
| R2 = 89.28% | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



