
Submitted:
11 July 2025
Posted:
15 July 2025
You are already at the latest version
Abstract
Pest management has entered a new era with the emergence of three innovative antisense technologies: RNA interference (RNAi), contact unmodified antisense DNA biotechnology (CUADb), and CRISPR/Cas systems. These approaches function through sequence-specific nucleic acid duplex formation and guided nuclease activity, offering unprecedented precision for targeted pest control. While RNA-guided systems such as RNAi and CRISPR/Cas were originally discovered in non-insect models as fundamental biological defense mechanisms (primarily against viruses), the DNA-guided CUADb system was first identified in insect pests as a practical pest control tool, with its broader role in ribosomal RNA (rRNA) biogenesis recognized later. These discoveries have revealed an entirely new dimension of gene regulation, with profound implications for sustainable pest management. Although RNAi, CUADb, and CRISPR/Cas share some mechanistic similarities, they differ in their mode of action, specificity, and applicability. No single approach provides a universal solution for all insect pests; instead, each is likely to be most effective against specific pest groups. Moreover, these technologies allow for rapid adaptation of control strategies to overcome target-site resistance, ensuring long-term efficacy. This review summarizes the core functional characteristics, potential applications, and current limitations of each antisense technology, emphasizing their complementary roles in advancing environmentally sustainable pest control. By bridging foundational biological discoveries with applied innovations, this work offers new perspectives on integrating these tools into modern pest management frameworks.
Keywords:
RNAi
; CUADb
; CRISPR/Cas
; antisense technologies
; insect pest control
1. Introduction
Nucleic acids, DNA and RNA, orchestrate cellular processes through precise complementary interactions [1]. The fundamental principles of Watson–Crick base pairing, coupled with the action of specific enzymes, govern essential biological mechanisms such as replication, transcription, translation, and gene expression regulation [2]. The specificity and fidelity of these processes arise from the unique combinations of nitrogenous bases, which form the molecular basis of genetic control. Three innovative antisense technologies, RNA interference (RNAi) [3], contact unmodified antisense DNA biotechnology (CUADb) [4-6], and clustered regularly interspaced short palindromic repeats and CRISPR-associated proteins (CRISPR/Cas) [7-9], have harnessed these nucleic acid interactions to develop targeted genetic effects. These technologies rely on the formation of sequence-specific duplexes: RNAi (guide RNA–messenger RNA (mRNA)) [10], CUADb (guide DNA-ribosomal RNA (rRNA)) [5, 11-14], and CRISPR/Cas (guide RNA-genomic DNA) [9,15,16], which then recruit specialized nucleases such as Argonaute (Ago) [17], rRNase [5,6], and CRISPR-associated protein [9].
The emergence of these technologies was contingent on the accumulation of critical data in bioinformatics, in vitro nucleic acid synthesis, and the development of laboratory tools. Typically, a pivotal insight or a successful experiment reveals a pattern that can subsequently be reliably applied in practice. In the case of RNAi, the key finding was the use of an antisense fragment within double-stranded RNA (dsRNA), for СUADb it was the use of rRNA as a target for antisense oligonucleotides, and for CRISPR/Cas it was the understanding that the target molecule for the antisense effect is genomic DNA. These innovations have set new standards in molecular genetics and are now being widely explored for their applications in insect pest control. Each of these technologies emerged from fundamental research, evolving from uncertain beginnings into powerful tools with transformative potential. Since their core mechanism relies on the complementary binding of antisense molecule, either DNA or RNA, to a target nucleic acid, they are collectively referred to as antisense technologies (Table 1). While RNA- and DNA-guided nucleases have been extensively studied, a targeted DNA-cleaving mechanism utilizing guide DNA to cleave target DNA of pests via a specific nuclease has not yet been developed, representing a potential direction for future research.
Chemical insecticides remain a cornerstone of insect pest management [18]. There are several key factors that drive the development of new classes of insecticides and the most important of these is economic cost of insect pest damage to agriculture and insecticide resistance which has dramatically and relentlessly increased since the mid-20th century [18-20]. The general mechanism underlying insecticide resistance is natural selection, which leads to an increase in frequency of specific alleles formed as a result of random mutations in insect pest population [21]. Antisense technologies (RNAi, CUADb, and CRISPR/Cas) are able to counteract insecticide resistance by targeting conserved genes or conserved gene regions and by facilitating the rapid development of effective pest control agents when resistance to existing insecticides emerges. While CUADb [6] and RNAi [3] demonstrate promising potential as next-generation bioinsecticides, due to their fast biodegradability, selectivity, and low carbon footprint [12], CRISPR/Cas is primarily used to genetically attenuate insect pest populations through genetic engineering [7]. Nevertheless, these innovative antisense technologies and their combinations, offer an expansive repertoire for controlling insect pests, and the central challenge lies in selecting the optimal pest management strategy for each specific case.
The development of antisense technologies was predicted on the discovery of the DNA double helix, followed by a formative era marked by pioneering studies such as Grineva and co-workers’ site-specific modification of valine tRNA [22] and the work of Zamecnik and Stephenson involving modified DNA against Rous sarcoma virus [23]. However, it became critically important to develop methods employing unmodified (natural) nucleic acids and to transform them into cost-effective, environmentally safe antisense technologies. While the RNAi was discovered by a research group in the USA in 1998 [3], CUADb was introduced in 2008 and further developed in Crimea by another team [4,6]. Multiple other research groups, primarily from Lithuania, Sweden and USA contributed to the development of the CRISPR/Cas9 approach in 2012 [7,24,25], culminating in the establishment of the three main antisense technologies for pest control at the turn of the 21st century. Although CUADb offers a relatively simple algorithm for pesticide design, RNAi and CRISPR/Cas are still progressing toward streamlined, reliable strategies for creating selective and effective pest control agents. The primary objective of this review is to provide a concise historical overview of the emergence of antisense technologies and to assess their potential in insect pest control. Overall, we provide an evaluation of the current capabilities and limitations of antisense technologies in this field, and propose directions for future development.
2. RNAi
2.1. History of discovery
The discovery of RNAi was inspired by the pioneering work of Zamecnik and Stephenson (1978), who demonstrated that a short antisense sequence of modified nucleic acid could inhibit the replication of the Rous sarcoma virus [23]. In 1998, Mello and Fire investigated the effect of antisense and sense RNA fragments on the development of the nematode Caenorhabditis elegans. They aimed to elucidate the mechanisms underlying the efficacy of sense RNA fragments synthesized by bacteriophage RNA polymerase (specifically, DNA-dependent RNA polymerases) and employed dsRNA fragments as a controls. Bacteriophage polymerases, although specific, occasionally generate ectopic transcripts, and DNA transgene arrays (also known as extrachromosomal arrays) are known to produce aberrant RNA products. Mello and Fire hypothesized that the interfering RNA population might include molecules with double-stranded features. Unexpectedly, it was the dsRNA fragment that triggered a potent silencing effect on the endogenous mex-3 mRNA transcript, which is highly expressed in the gonads and early embryos of the nematode. The groundbreaking discovery, published in Nature in 1998 [3], established the foundational concept of RNAi: dsRNA molecules initiate gene silencing pathways by targeting homologous transcripts [26,27]. Subsequent studies revealed that short antisense RNA fragments (21–23 nucleotides in length) are produced during RNAi and mediate the degradation of target mRNAs [28] (Figure 1).
2.2. How RNAi works in insect pests
RNAi has been widely studied for its potential in agricultural pest management, particularly through the silencing of essential insect genes to suppress pest populations. The small interfering RNA (siRNA) pathway, the most extensively characterized RNAi mechanism, consists of two branches: the exogenous siRNA (exo-siRNA) pathway, which mediates antiviral defense, and the endogenous siRNA (endo-siRNA) pathway, responsible for transposon suppression within the genome [29]. The exo-siRNA pathway is commonly exploited in experimental and applied contexts by introducing dsRNA to silence specific target genes [30,31]. For effective gene silencing, long dsRNA fragments (>200 bp) are typically used to trigger the production of short antisense siRNAs [26,27]. The RNAi process begins with cellular uptake of dsRNA, followed by cleavage into siRNAs (21–25 nucleotides) by the enzyme Dicer. These siRNAs are then incorporated into the RNA-induced silencing complex (RISC), where Argonaute (Ago) proteins serve as the catalytic core [32]. One strand of the siRNA duplex, known as the guide strand, directs RISC to the complementary mRNA target, leading to transcript degradation or translational repression [33].
Although much of the research has focused on the siRNA machinery, the RNAi pathway in insects is more complex and includes three sub-pathways: siRNA, micro (miRNA), and piwi-interacting RNA (piRNAs). miRNAs (18-24 nucleotides) are involved in diverse regulatory processes and interact with Ago proteins, while piRNAs (26–31 nucleotides) are primarily responsible for silencing transposable elements, particularly in germline cells, through interactions with PIWI proteins. The application of artificially synthesized dsRNA can influence not only the siRNA pathway but also genes associated with the miRNA pathway, suggesting the existence of an interactive regulatory network [34].
RNAi is particularly robust and systemic in Coleopteran insects [29]. However, systemic RNAi, defined as the uptake of dsRNA from the environment followed by its intercellular transport, is limited in Diptera, Lepidoptera [35-37], and sap-feeding Hemiptera [38,39], thereby constraining its application in protection. Nonetheless, RNAi has demonstrated considerable potential for pest control, especially in Coleoptera species, by targeting key genes involved in development, detoxification, and reproduction (Table 2).
2.3. Perspectives and limitations of RNAi for insect pest control
Despite the remarkable progress in RNAi research over the past 25 years, its practical application in insect pest and disease vector management remains limited. To date, only one commercial dsRNA-based biopesticide Calantha® has been developed for foliar application, while a few others are currently in the development pipeline. Additionally, three transgenic crops expressing dsRNA are commercially available.
Calantha® is the first sprayable dsRNA-based biopesticide targeting the Colorado potato beetle (CPB, Leptinotarsa decemlineata), was commercially released by GreenLight Biosciences in 2023 [65,66]. However, as with dsRNA traits, sprayable dsRNAs are likely to be used in combination with other insect control tools. Sprayable dsRNAs degrade quickly in the environment [67], and careful consideration needs to be given to spray intervals and the insect life cycle so that only a single generation is exposed to the dsRNA product, after which insecticide treatments should be rotated to a different MoA [68,69].
Several challenges have inhibited commercialization of RNAi-based products, including variable RNAi efficacy across insect species, competition from transgenic Bt crops, and inability to effectively control some major sucking pests. Another concern is the potential evolution of RNAi resistance. For example, studies have shown that western corn rootworm (Diabrotica virgifera virgifera) [70], willow leaf beetle (Plagiodera versicolora) [71], and CPB [72,73] can develop resistance to dsRNA-based control. Of note, Calantha® can be deployed in pesticide rotation programmes to mitigate selection for resistance in CPB [74]. Nonetheless, RNAi, when combined with precision agriculture and integrated pest management (IPM) strategies, could significantly enhance sustainability. Opportunities for synergy exist with advancing the technologies such as CUADb, CRISPR/Cas technology, microbial-based RNAi production, and nanoformulations for improved dsRNA delivery. Of note, RNA molecules are prone to degradation in the environment. To enhance their effectiveness, researchers are developing more stable RNA formulations and innovative delivery methods.
Understanding dsRNA structure is essential for RNAi applications in pest control and further developments because it dictates how dsRNA is processed into small interfering RNAs (siRNAs), which then target specific genes for silencing. This knowledge is vital for optimizing dsRNA sequences, delivery methods, and target selection to achieve effective pest control [75,76]. Although the effects of dsRNA-mediated interference are highly potent and specific, there are several concerns that should be taken into account when designing RNAi-based experiments. First, if a sequence is shared among multiple closely related genes, RNAi may unintentionally silence several members of the gene family or homologous genes in related species [77]. Second, genes with low expression levels may exhibit resistance to RNAi, at least partially. For example, if the target protein is very stable, its depletion occurs much more slowly despite transcript degradation. Moreover, the function of a target protein can be compensated for by related proteins, which may even be upregulated by the cell to counteract the loss. Additionally, the transcription of the target gene itself could be upregulated by a regulatory gene network in response to RNAi mediated knock-down [78].
Importantly, by understanding structure and how Ago proteins bind to guide RNAs and target RNAs, scientists can design more effective and specific RNAi tools for pest management. Realizing the structural interactions between Ago proteins and these RNAs allows for precise design of small RNAs that will specifically target genes essential for the pest's survival or reproduction [79,80]. Also structural insights into the active site of Ago proteins and how they interact with target RNAs help optimize the design of siRNAs for efficient RNAi-mediated gene silencing [79,80]. Additionally, understanding the structural interactions within RISC can help optimize RNAi delivery and enhance silencing efficacy [81]. Finally, by understanding how Ago proteins function, scientists can develop RNAi-based pest control strategies that are specific, effective, and environmentally friendly. This includes designing RNAi tools for various pests and crops, as well as optimizing delivery methods [82].
The implementation of RNAi-based pest control strategies is currently hindered by regulatory hurdles and public apprehension. Gaining regulatory approval and public acceptance will depend on transparent scientific communication, rigorous safety evaluations, and evidence-based demonstrations of efficacy. A major practical limitation remains the high cost of dsRNA production; publicly accessible in vitro synthesis methods are still prohibitively expensive, exceeding $50 per milligram [83-87].
Despite these challenges, RNAi presents a promising avenue for insect pest management due to its capacity to selectively silence genes vital for insect development and survival. This specificity provides an opportunity to develop environmentally sustainable pest control solutions. However, the successful deployment of dsRNA-based insecticides will require coordinated efforts from researchers, policymakers, and industry stakeholders worldwide. Future advancements, particularly in dsRNA formulation, delivery systems, and cost-effective production, are expected to drive the broader adoption of RNAi technologies over the next 25 years, potentially transforming pest management in agriculture and beyond [88].
3. CUADb
3.1. History of discovery
Unmodified DNA, as a programmable molecule and polymer of natural origin, has always attracted researchers. Unfortunately, for a long time it was believed that unmodified oligonucleotides are toxic to cells and degraded quickly in all eukaryotic cells under the action of nucleases [89], including insects [11,90]. Some articles literally stated that unmodified (phosphodiester) antisense oligonucleotides should not be used for these experiments [91]. In addition, it was assumed that rRNA is resistant to degradation in the presence of antisense DNA oligonucleotides [92,93]. Generally, at the beginning of the current century, there was no clue in scientific literature that DNA can serve as an insecticide.
DNA insecticidal activity was discovered out of curiosity in the spongy moth Lymantria dispar. In 2007, Oberemok began research in the field of transovarial transmission of L. dispar multiple nucleopolyhedrovirus (LdMNPV) as a part of his doctoral studies. Two specific primers were selected within the anti-apoptotic gene (IAP-3) of LdMNPV: a forward primer from the sense strand (5’-GCCGGCGGAACTGGCCCA-3; oligoBIR fragment) and a reverse primer from the antisense strand (5’-CGACGTGGTGGCACGGCG-3’; oligoRING fragment) [94]. These primers initiate the formation of 317 bp long amplicon during PCR in the presence of the LdMNPV DNA. On purified virus preparations, the primers formed the expected 317 bp long amplicon [95]. However, when searching for the virus in the tissues of the virus-free L. dispar, primers initiated the formation of several amplicons of different lengths, indicating that they were not specific enough to detect LdMNPV in host tissues. Consequently, obtained data indicated that the L. dispar genome contained regions homologous to fragments of the IAP-3 gene of the LdMNPV, a phenomenon previously reported for other viruses [96]. Thus, the developed primers were not suitable for accurate detection of the LdMNPV in insect tissues. Hypothetically, oligoBIR and oligoRING fragments could target the gene expression of homologous L. dispar IAP genes to induce apoptosis in insect cells. In April 2008, Oberemok decided to test the primers in an unusual way: he applied small drops of an aqueous primer solution to the surface of spongy moth larvae [4,97]. To his surprise, after 3-5 days, the larvae began to die in significant numbers due to the applied DNA fragments of the virus genome. This pioneering experiment marked the beginning of research into the development of a previously unknown class of contact DNA insecticides (oligonucleotide insecticides, or briefly olinscides) and the CUADb platform. The first publications of these results appeared in the Ukrainian patent (№36445) in 2008, followed by articles in Pesticide Biochemistry and Physiology [98,99]. The earliest 18–20 nt long oligonucleotide insecticides based on anti-apoptotic genes demonstrated their effectiveness in LdMNPV-free larvae and were even more potent in LdMNPV-infected spongy moth larvae [94]. Unique antisense DNA sequences of 11-20 nt can provide high selectivity in action; however, their effectiveness heavily depends on the concentration of the target RNA. As a result, CUADb has been developed and now shows the best results on pests targeting their rRNA (which constitutes 80% of cellular RNA), utilizing the DNA containment mechanism (DNAc) to create a powerful algorithm of insect pest control (Figure 2).
The discovery of oligonucleotide insecticides introduced a novel dimension to insect pest management by utilizing nucleic acids as contact insecticides. This concept was further explored in RNAi research, and three years later, Wang et al. [100] successfully demonstrated, for the first time, the use of dsRNA fragments as contact insecticides in pest control.
3.2. How it works on insect pests
In 2019, Oberemok and colleagues introduced three major advancements to improve the CUADb platform. First, insect ribosomal RNA (rRNA) was selected as the primary target for oligonucleotide insecticides [11]. Targeting pre-rRNA and rRNA enhances efficacy because these RNA types constitute approximately 80% of the total cellular RNA [101]. In contrast, mRNAs, despite their diversity, make up only about 5% of the total RNA. As a result, targeting rRNA significantly increases the signal-to-noise ratio, reaching approximately 100,000:1 when compared to random mRNA sequences. In insects, rRNA is present in both nuclear and mitochondrial forms. Nuclear rRNA includes 28S (~3900 nt), 18S (~1920 nt), 5.8S (~160 nt), and 5S (~120 nt), while mitochondrial rRNA includes 16S (~1140 nt) and 12S (~600 nt). Together, they represent the majority of cellular RNA, making them ideal targets for antisense oligonucleotides.
Second, the length of oligonucleotide insecticides was reduced to 10–12 nucleotides (nt) in order to decrease production costs. Shorter sequences result in higher yields during DNA synthesis using the phosphoramidite method, thus providing a greater mass of product. An oligonucleotide insecticide of 11 nt can maintain sufficient sequence specificity, with a uniqueness frequency of approximately 1 in 4.19 million sequences—adequate for most agricultural systems [102]. For more diverse environments, such as forest ecosystems, longer sequences (15–20 nt) may be used to enhance selectivity [12].
Third, it was discovered insects belonging to the suborder Sternorrhyncha (Hemiptera) are susceptible to unmodified antisense oligonucleotides [6]. Since then, oligonucleotide insecticides have been successfully applied against a range of sap-feeding insect pests targeting 28S rRNA in Unaspis euonymi, Dynaspidiotus britannicus, Icerya purchasi, Ceroplastes japonicus, Aonidia lauri, Coccus hesperidum [5,12,13, 103-106]; 18S rRNA in Pseudococcus viburni [107]; and the internal transcribed spacer 2 (ITS2) of pre-rRNA in Macrosiphoniella sanborni, Schizolachnus pineti [14,108], and Trioza alacris [109]. ITS2-targeting oligonucleotides have also demonstrated acaricidal potential against the spider mite Tetranychus urticae [110,111]. Typically, a single contact treatment with oligonucleotide insecticides at 100 ng/μl results in mortality rates of approximately 80% within 3 to 14 days [12,106]. The first successful application of these insecticides within Sternorrhyncha was conducted on U. euonymi in 2019 [104,112] (Table 3).
It is important to note that studies on Sternorrhynchan insects demonstrated that unmodified oligodeoxyribonucleotides (also known as oligonucleotide insecticides or olinscides) can induce both upregulation and downregulation of target genes through a mechanism known as DNA containment. This mechanism operates in two sequential phases: the first involves the arrest of target rRNA, leading to a functional block of ribosomes and triggering a compensatory overexpression of rRNA via rDNA transcription. The second phase involves degradation of the arrested rRNA by ribonuclease enzymes (rRNases) [5,6,114]. During this process, the olinscide binds to its complementary target rRNA sequence, forming a DNA–RNA duplex. In the presence of rRNase, this duplex is processed through a mechanism analogous to a zipper, referred to as the ‘genetic zipper’ [109]. The genetic zipper method is not only an innovative tool but also serves as a predictive algorithm capable of estimating the efficacy of a specific olinscide against the intended target species and closely related organisms possessing identical rRNA target sequences. [109].
Oligonucleotide insecticides can be designed using DNAInsector program (available at dnainsector.com) or developed manually based on pre-rRNA and rRNA sequences retrieved from the GenBank database. Practically, this means that anyone with basic sequence knowledge can design an olinscide complementary to the pre-rRNA or rRNA of a Sternorrhynchan insect with a high likelihood of success. However, to ensure species selectivity, it is essential to compare homologous sites in non-target organisms to avoid off-target effects. Synthesis of oligonucleotide insecticides is typically performed using the phosphoramidite method, either in liquid-phase or solid-phase formats. Available solid-phase synthesizers include the ASM-800 (BIOSSET, Russia), OligoPilotTM (Cytiva, Sweden), and the 10-Column DNA Synthesizer (PolyGen, Germany) [105]. The standard preparation involves dissolving the olinscide in nuclease-free water at a concentration of 1 mg per 10 mL and applying it to approximately 1 m² of infested plant foliage.
3.2. Perspectives and limitations of CUADb for insect pest control
Currently, DNA-based technologies are revolutionizing plant protection by enabling the development of novel control agents with advanced characteristics. DNA, as a molecule of natural origin, is inherently more compatible with ecosystem balance, including agroecosystems. Its programmability allows the design of highly specific treatments, such as insecticidal and acaricidal agents. Moreover, in the event of resistance development, adaptive strategies can be readily implemented, such as redesigning oligonucleotide insecticides (olinscides) by shifting the target site upstream or downstream of the resistance-conferring region in pre-rRNA and rRNA sequences [5]. Oligonucleotide insecticides offer several distinct advantages: they exhibit a low carbon footprint, high specificity to target pests, rapid biodegradability in ecosystems, and minimal risk of target-site resistance. Furthermore, their efficacy can now be predicted across different pest species, particularly in phylogenetically related taxa with conserved target sequences [109]. Given these benefits, it is likely only a matter of time before such DNA-based products become mainstream tools in crop protection. The mode of action, termed the DNA containment mechanism, is well understood. The synthesis of DNA fragments via the phosphoramidite method has been optimized, and delivery routes (primarily contact-based; oral delivery is less promising) have been established. Although the cost of DNA insecticides remains a concern, this challenge is expected to diminish in the near future. Indeed, DNA insecticides for specific pest species are already becoming cost-competitive with conventional chemical insecticides [12].
For example, in the control of conifer aphids, CUADb has significantly reduced costs by utilizing liquid-phase DNA synthesis [13,105]. Sumitomo Chemical Co., Ltd. (Tokyo, Japan), a leading company in liquid-phase synthesis, reportedly offers 1 kg of unmodified 11-nucleotide oligonucleotides at 25,000 USD (personal communication). In contrast, the cost of synthesizing the same quantity using standard solid-phase synthesis, which is more widely accessible in research laboratories, can reach approximately $1 million. At a recommended application rate of 200 L per hectare with an active ingredient concentration of 0.1 mg/L (0.1 ng/mL), the treatment cost per hectare using liquid-phase synthesis is approximately $0.50 per hectare when produced via liquid-phase DNA synthesis [13]. This affordability allows for frequent applications under field conditions. However, for many Sternorrhyncha pests, effective control requires a significantly higher concentration, around 0.05 g/L, which elevates the cost to approximately $250 per hectare, even when using the more economical liquid-phase synthesis method. Consequently, achieving an optimal balance between cost-effectiveness and formulation efficiency will be critical for the broader adoption of nucleic acid-based insecticides. Current estimates suggest that CUADb-based approaches could potentially manage 10–15% of all insect pest species.
While oligonucleotide insecticides have demonstrated strong efficacy against hemipteran and lepidopteran pests, they have shown much lower activity against coleopterans such as L. decemlineata [115]. The incorporation of adjuvants (e.g., spreaders, adhesives, penetrants, and UV protectants) into formulations may enhance their effectiveness; however, the environmental safety of such additives must be thoroughly evaluated beforehand. Another critical factor in olinscide design is the consideration of non-canonical base pairing, such as A:C (C:A) and G:U (T:G) interactions [116,117], which may occur between olinscides and imperfectly matched rRNA target sites. These interactions could compromise specificity and pose risks to non-target organisms. Therefore, these possibilities must be carefully considered during the design phase [15,13].
Although the CUAD-based ‘genetic zipper’ method presents a promising platform for cost-effective DNA insecticide production, further optimization of manufacturing processes and cost reduction strategies are required for large-scale implementation. Research efforts are currently focused on enhancing specificity by targeting unique regions of pest rRNA to minimize off-target effects. Additionally, a deeper understanding of the mechanisms governing olinscide uptake by insect cells is crucial for improving treatment efficacy. In summary, the "genetic zipper" method offers substantial promise for insect pest control. However, its successful scaling-up will depend on addressing several challenges, including specificity, delivery mechanisms, production efficiency, resistance management, and environmental impact. Overcoming these barriers will enable CUAD-based oligonucleotide insecticides to emerge as a viable, sustainable, and widely adoptable strategy for modern pest management.
4. CRISPR/Cas
4.1. History of discovery
While the development of RNA interference (RNAi) technology and the CUADb database followed a relatively linear and well-documented trajectory, the emergence of CRISPR/Cas technology resulted from the collective efforts of numerous research groups, making it difficult to attribute its discovery to a single individual or team. This section provides a brief overview of the widely accepted scientific consensus on the key milestones in the development of the CRISPR/Cas system. We acknowledge the contributions of many researchers whose names may not be individually mentioned.
In 1987, Ishino and colleagues discovered a previously unknown repeat sequence in Escherichia coli, although its significance was not fully recognized at the time [118]. In 1989, Spanish scientist Mojica, while studying the archaeal microbe Haloferax mediterranei, observed a similar pattern, repeated palindromic sequences of approximately 30 base pairs, separated by spacer sequences of roughly 36 base pairs. These sequences did not resemble any known repeat families in prokaryotes at that time [119]. Later Mojica and Jansen introduced the term CRISPR (clustered regularly interspaced short palindromic repeats) to describe these structures [25]. In 2002, Cas (CRISPR-associated) genes were identified adjacent to CRISPR loci by Jansen et al. [120], suggesting a functional relationship. In 2007, Horvath and colleagues provided the first experimental evidence for the role of the CRISPR-Cas system in adaptive immunity. They demonstrated that Streptococcus thermophilus required the Cas7 protein to acquire resistance to phages, although those strains already containing a phage-derived spacer maintained resistance without Cas7. This suggested Cas7's involvement in spacer acquisition rather than the immunity process itself [121]. In contrast, Cas9 (previously referred to as Cas5, Csn1, or Csx12) which contains two nuclease motifs (HNH and RuvC), was shown to be essential for phage resistance, indicating its direct role in nucleic acid cleavage and adaptive immunity [122-124]. In 2011, Charpentier identified a small RNA in Streptococcus pyogenes known as trans-activating CRISPR RNA (tracrRNA), which is essential for the processing of CRISPR RNAs (crRNAs) and for guiding the Cas9 complex to cleave target DNA [7]. In 2012, Šikšnys et al. published a seminal study in Proceedings of the National Academy of Sciences, demonstrating that the Cas9 protein could be programmed with custom-designed spacers within the CRISPR array to cleave specific DNA targets in vitro [24]. Independently, Charpentier and Jennifer Doudna, in a paper published in Science, showed that Cas9 could be directed by synthetic crRNAs and tracrRNAs to cleave double-stranded DNA at targeted sites, with each of the nuclease domains cleaving opposite strands [25]. These collective findings established the foundational principles for genome editing using the CRISPR/Cas9 system. Although Charpentier, Doudna, and Šikšnys are widely credited with demonstrating the genome-editing potential of the CRISPR/Cas system in 2012, the overall development of the technology reflects a cumulative effort involving many researchers and significant scientific advancements over several decades.
4.2. How it works on insect pests
CRISPR/Cas9 system is the predominant tool in gene editing used for insect pest control, characterized by its simplicity and efficiency. The system consists of two essential components: sgRNA (single-guide RNA) and Cas9 nuclease. sgRNA is an engineered fusion of CRISPR RNA (crRNA, defining the target DNA sequence) and trans-activating CRISPR RNA (tracrRNA, facilitating Cas9 binding), guiding the Cas9 enzyme to specific genomic loci via base pairing. Cas9, a multi-domain DNA endonuclease from Streptococcus pyogenes, recognizes a protospacer adjacent motif (PAM) sequence (typically 5’-NGG-3’) near the target site. Upon binding, it creates a double-stranded break (DSB) three base pairs upstream of PAM using its RuvC and HNH catalytic domains, with HNH cleaving the target strand and RuvC cleaving the opposite strand [125-129]. Following DSB formation, cellular repair mechanisms engage through either NHEJ (non-homologous end joining, often resulting in small deletions and insertions) or HDR (homology-directed repair, enabling precise genetic modifications with a repair template) [129,130]. When the coding sequence (CDS) of the desired gene is targeted by CRISPR/Cas9 and repaired via NHEJ, which is an error-prone process, the insertions or deletions of random base pairs may result in the disruption of the CDS and, thus, nonfunctional protein or no protein is formed at all. Meanwhile, HDR can generate mutations (gene correction or insertions) through integrating the exogenous genes of interest, which is called gene knock-in [130,131] (Figure 3).
Since its initial adoption in Drosophila, CRISPR/Cas9 has been adopted in a broad array of insect species. The aim of genome editing in insect pest management it is to introduce engineered traits into wild pest populations, offering a promising strategy to manage invasive species [132,133].
In the context of insect pests, CRISPR/Cas can be applied in several keyways (Table 4). First, disrupting genes involved in insecticide resistance. A comprehensive and objective review of the progress made using CRISPR/Cas9 in insecticide resistance studies has been generalized according to species from different insect orders and the types of gene editing in the review article by Xu et al. [134]. Secondly, targeting genes that regulate reproduction or sex determination. For example, CRISPR-Cas9 can be used to target genes like Astra-2 (a sex determination gene) in insects like the Anastrepha ludens, which are also known as the Mexfl, leading to sterility in males and intersexual phenotypes in females [135]. Also, Doublesex (Dsx) gene is a key regulator of sexual differentiation in insects. CRISPR-Cas9 can be employed to target the Dsx gene, either by disrupting its normal expression or by manipulating its isoforms (variants). For instance, in diamondback moths, altering the Dsx gene has resulted in gender-specific sterility, meaning males exhibit female-specific transcripts and females exhibit male-specific transcripts [136]. Thirdly, suppressing vector by specifically targeting genes competence in disease-transmitting insects. For example, by modifying these genes, researchers can disrupt the mosquito's ability to transmit diseases like malaria, dengue fever, and Zika [137-139]. Fourthly, developing gene drives to spread deleterious traits through populations, such as cause sterility or reduced fitness in pest species, leading to population suppression or even eradication [129, 140]. These mechanisms bypass Mendelian inheritance using mechanisms such as homing endonuclease genes and transposable elements. CRISPR-based gene drives rely on the Cas9 enzyme and a guide RNA to cut the wild-type allele and copy the engineered allele during DNA repair, thereby ensuring the trait spreads in subsequent generations [129,141-143]. Recent results by Meccariello et al. (2024) show that the Medfly Ceratitis capitata is highly susceptible to homing-based gene drive strategies. By targeting the Medfly transformer gene, it was also demonstrated how the CRISPR-Cas9 gene drive can be linked to sexual conversion, resulting in genetic females being transformed into fertile and harmless XX males [144], beetles, moths and grasshoppers [145-147]. Fifthly, CRISPR-based genome editing in some cases proved to be more efficient and stable in comparison to other techniques. For example, in insect species such as silkworms, CRISPR/Cas-mediated knockdown of the homeobox gene Scr led to developmental abnormalities and malformed adult structures, demonstrating its potential as an alternative to RNAi in Lepidoptera [148]. Recent work demonstrated that chloroplast-engineered dsRNA targeting Frankliniella occidentalis can be combined with CRISPR/Cas9 technology to efficiently control this pest [149,150]. Finally, this technology has improved our understanding of insect physiology, development, morphology, and behavior. For example, the advances in genome editing technology have led to an increased usage of CRISPR/Cas9 technology for the functional study of different genes, particular in S. frugiperda, opening new avenues for exploring the genetic underpinnings of biology of this serious pest [130].
4.3. Perspectives and limitations of CRISPR/Cas for insect pest control
CRISPR/Cas technology offers a powerful perspective for insect pest control, enabling gene editing and potentially leading to sustainable pest management strategies, such as gene drives and insect-resistant plants. The employment of molecular genetic engineering and insect transformation through CRISPR/Cas9 in multiple species has overcome many previously intractable problems using traditional methods which mainly relied on naturally occurring genetic mutations or elements. CRISPR/Cas genome editing is widely used for its effectiveness and simplicity. Different Cas proteins like Cas3, Cas12a, and Cas13a are now used to expand gene editing, natural and engineered, for precise editing, base editing, prime editing, and gene regulation. Recent advances enabled DNA-free editing, allowing genome modification without inducing double-stranded breaks [175-177].
Unfortunately, today it is almost impossible to predict the outcome from gene editing of a particular insect pest as well as which gene is better to target by this approach to gain maximum effect. CRISPR/Cas systems can sometimes make unintended edits to the genome at locations other than the target site, potentially leading to unforeseen consequences. Delivering the CRISPR/Cas system into insects can be challenging, and the efficiency of delivery can vary. There are environmental concerns about the potential impact of gene drives and insect-resistant plants on non-target species and ecosystems, as well as the long-term consequences of these interventions. Gene drives rely on successful mating between modified and wild insects, but mating barriers (e.g., pre- and postzygotic barriers) can limit gene flow and prevent the spread of the gene drive. Genetically modified insects may experience fitness costs, which can affect their ability to compete with wild populations and reduce the effectiveness of the gene drive. Precise gene knock-in using HDR can be challenging due to the difficulty in supplying sufficient donor DNA templates [178]. Achieving complex genome modifications can be difficult with CRISPR/Cas, and mosaicism (variation in the genome of different cells) can be a problem in some cases. While population replacement can be effective, ensuring the long-term sustainability of gene drive systems and preventing their unintended spread into non-target species requires careful consideration [130,179]. The long-term effects of CRISPR-Cas-mediated modifications on insect development, behavior, and interactions with the environment are still being investigated [134,180].
As the field of insect molecular biology progresses, CRISPR/Cas technology has the potential to transform pest management by offering a safe and highly efficient solution. The use of CRISPR/Cas technology extends beyond traditional methods, enabling researchers to explore various pest management strategies at the genetic level and offering a more refined and sustainable approach for future applications in the control of insect pests. This may also facilitate further development of the IPM by incorporating principles of synthetic biology in diverse agricultural settings [130].
5. Conclusion
Antisense technologies are based on natural mechanisms that regulate life processes. Complementary interactions between nucleic acids underpin fundamental biological functions such as cell division, metabolism and defense. The development of practical tools leveraging these antisense mechanisms represents a highly relevant and promising area of scientific research. We believe that sustained investment over decades in this field is justified to realize effective and environmentally friendly solutions for pest management. RNAi, CUADb, and CRISPR-Cas systems are powerful gene silencing tools that operate at distinct molecular levels and offer varying degrees of permanence. RNAi and CUADb transiently reduce gene expression by targeting mRNA and rRNA of pests, respectively, whereas CRISPR-Cas induces permanent gene silencing by altering genomic DNA. The CUADb system is unique among these technologies due to its DNA-guided mechanism and its role in rRNA biogenesis, although it shares certain functional similarities with RNAi and CRISPR/Cas. Together, these systems hold considerable promise in combating insecticide resistance by targeting conserved genes and facilitating the development of effective pest control agents.
Application of these technologies in insect pest control has already demonstrated significant potential. Notably, RNAi, CUADb and CRISPR/Cas exhibit optimal efficacy against specific insect pest groups, highlighting the potential for combining these approaches to enhance their effectiveness across broader pest populations. Although some competition exists among these modern antisense technologies, the physiological and genetic diversity of pest species suggests that no single approach will serve as a universal solution. Currently, antisense-based insect pest control technologies remain in the development phase. As data on their successes and limitations continue to accumulate, major companies are actively pursuing commercialization. Meanwhile, legislative framework governing the implementation of antisense technologies are gradually evolving in various countries. Although rapid breakthroughs may not be imminent, steady progress is inevitable, and will ultimately contribute to improvements in both human health and environmental sustainability.
In summary, RNAi, CUADb, and CRISPR/Cas represent three complementary "worlds" of antisense technologies with promising future applications in diverse fields. These technologies hold potential for improving agriculture and developing new pest control strategies. CRISPR/Cas can create mutations for long-term changes, while RNAi and CUADb provide temporary gene silencing to block certain gene function or quickly adapt to resistance of insect pests. This combination of antisense tools offers versatility in addressing various aspects of pest control, including resistance management, population suppression, and developing novel and eco-friendly pest management strategies. However, challenges related to delivery, affordability, and environmental considerations need to be addressed for these technologies to reach their full potential.
Author Contributions
Conceptualization, V.O.; software, H.K., N.G. and V.O.; validation, V.O., and N.G.; formal analysis, V.O.; resources, V.O.; writing—original draft preparation, H.K., N.G., V.S., N.N., R.F., A.C., J.A., V.O. and K.L.; writing—review and editing, N.G., A.J. and V.O.; visualization, H.K., N.G. and V.O.; supervision, N.G. and V.O.; project administration, N.G. and V.O.; funding acquisition, V.O. All authors have read and agreed to the published version of the manuscript.
Funding
The research was funded by a grant from the Russian Science Foundation № 25-16-20070, https://rscf.ru/project/25-16-20070/.
Data Availability Statement
No new data were created.
Acknowledgments
We thank our many colleagues, too numerous to name, for the technical advances and lively discussions that prompted us to write this brief research report. We apologize to the many colleagues whose work has not been cited. Experiments were carried out at the Molecular Genetics and Biotechnologies Lab created within the framework of a state assignment V.I. Vernadsky Crimean Federal University for 2024 and the planning period of 2024–2026 No. FZEG-2024–0001. We are very much indebted to all reviewers and our colleagues from the lab on DNA technologies, PCR analysis, and creation of DNA insecticides (V.I. Vernadsky Crimean Federal University, Department of General Biology and Genetics), and OLINSCIDE BIOTECH LLC for valuable comments on our manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Minchin, S.; Lodge, J. Understanding biochemistry: structure and function of nucleic acids. Essays Biochem. 2019, 63, 433–456. [Google Scholar] [CrossRef] [PubMed]
- Westhof, E.; Yusupov, M.; Yusupova, G. Recognition of Watson-Crick base pairs: constraints and limits due to geometric selection and tautomerism. F1000Prime Rep. 2014, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V. Method of Elimination of Phyllophagous Insects from Order Lepidoptera. Ukraine Patent UA. 2008, 445, 27. [Google Scholar]
- Gal'chinsky, N.V.; Yatskova, E.V.; Novikov, I.A.; Sharmagiy, A.K.; Plugatar, Y.V.; Oberemok, V.V. Mixed insect pest populations of Diaspididae species under control of oligonucleotide insecticides: 3′-end nucleotide matters. Pesticide Biochem. Physiol. 2024, 200, 105838. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V.; Laikova, K.V.; Gal’chinsky, N.V. Contact unmodified antisense DNA (CUAD) biotechnology: list of pest species successfully targeted by oligonucleotide insecticides. Front. Agron. 2024, 6, 1415314. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E.A. Programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Gasiunas, G.; Barrangou, R.; Horvath, P.; Siksnys, V. Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc. Natl. Acad. Sci. U S A. 2012, 109, E2579–E2586. [Google Scholar] [CrossRef] [PubMed]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Rana, T.M. Molecular mechanisms of RNA-triggered gene silencing machineries. Acc Chem Res. 2012, 45, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V.; Laikova, K.V.; Gal'chinsky, N.V.; Useinov, R.Z.; Novikov, I.A.; Temirova, Z.Z.; Shumskykh, M.N.; Krasnodubets, A.M.; Repetskaya, A.I.; Dyadichev, V.V.; et al. DNA insecticide developed from the Lymantria dispar 5.8S ribosomal RNA gene provides a novel biotechnology for plant protection. Sci Rep. 2019, 9, 6197. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V.; Laikova, K.V.; Andreeva, O.A.; Gal'chinsky, N.V. Oligonucleotide insecticides and RNA-based insecticides: 16 years of experience in contact using of the next generation pest control agents. J. Plant. Dis. Prot. 2024, 131, 1837–1852. [Google Scholar] [CrossRef]
- Oberemok, V.V.; Puzanova, Y.V.; Gal’chinsky, N.V. The 'genetic zipper' method offers a cost-effective solution for aphid control. Front. Insect Sci. 2024, 4, 1467221. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V.; Gal’chinsky, N.V.; Novikov, I.A.; Sharmagiy, A.K.; Yatskova, E.V.; Laikova, E.V.; Plugatar, Yu. rRNA-specific antisense DNA and dsDNA trigger rRNA biogenesis and cause potent insecticidal effect on insect pest Coccus hesperidum L. biorXiv 2024. biorXiv 2024. [Google Scholar]
- Wiles, M.V.; Qin, W.; Cheng, A.W.; Wang, H. CRISPR-Cas9-mediated genome editing and guide RNA design. Mamm Genome. [CrossRef]
- Li, T.; Yang, Y.; Qi, H.; Cui, W.; Zhang, L.; Fu, X.; He, X.; Liu, M.; Li, P-f. ; Yu T. CRISPR/Cas9 therapeutics: progress and prospects. Sig Transduct Target Ther. 2023, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zuo, Z.; Shao, W; Jin, Y. ; Meng, Y. The expanding roles of Argonautes: RNA interference, splicing and beyond. Brief Funct Genomics. 2018, 17, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.F.; Castanheira, E.M.S.; Sousa, S.F. The Buzz on Insecticides: A Review of Uses, Molecular Structures, Targets, Adverse Effects, and Alternatives. Molecules 2023, 28, 3641. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, J.A.; Fan, R.; Naz, H.; Bamisile, B.S.; Hafeez, M.; Ghani, M.I.; Wei, Y.; Xu, Y.; Chen, X. Insights into insecticide-resistance mechanisms in invasive species: Challenges and control strategies. Front. Physiol. 2023, 13, 1112278. [Google Scholar] [CrossRef] [PubMed]
- Gul, H.; Gadratagi, B.G.; Güncan, A.; Tyagi, S.; Ullah, F.; Desneux, N.; Liu, X. Fitness costs of resistance to insecticides in insects. Front. Physiol. 14, 1238111. https. [CrossRef]
- Hawkins, N.J.; Bass, C.; Dixon, A.; Neve, P. The evolutionary origins of pesticide resistance. Biol. Rev. Camb. Philos. Soc. 2019, 94, 135–155. [Google Scholar] [CrossRef] [PubMed]
- Belikova, A.; Zarytova, V.; Grineva, N. Synthesis of ribonucleosides and diribonucleoside phosphates containing 2-chloroethylamine and nitrogen mustard residues. Tetrahedron Lett. 1967, 37, 3557–3562. [Google Scholar] [CrossRef] [PubMed]
- Zamecnik, P.C.; Stephenson, M.L. Inhibition of Rous sarcoma virus replication and cell transformation by a specific oligodeoxynucleotide. Proc. Natl. Acad. Sci. USA. 1978, 75, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S. The Heroes of CRISPR. Cell 2016, 164(1-2), 18-28. [CrossRef]
- Shmakova, A.A.; Shmakova, O.P.; Karpukhina, A.A.; Vassetzky, Y.S. CRISPR/Cas: History and Perspectives. Russ. J. Dev. Bio. 2022, 53, 272–282. [Google Scholar] [CrossRef]
- Tomoyasu, Y.; Miller, S.C.; Tomita, S.; Schoppmeier, M.; Grossmann, D.; Bucher, G. Exploring systemic RNA interference in insects: a genome-wide survey for RNAi genes in Tribolium. Genome Biol. 2008, 9, R10. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, P. Key mechanistic principles and considerations concerning RNA interference. Front. Plant Sci. 2020, 11, 1237. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Shu, R.; Liu, J. The development and improvement of ribonucleic acid therapy strategies. Mol. Ther. Nucleic Acids. 2021, 26, 997–1013. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.Y.; Palli, S.R. Mechanisms, applications, and challenges of insect RNA interference. Annual review of entomology 2020, 65, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.M.; Silver, K.; Zhang, J.; Park, Y.; Zhu, K.Y. Molecular mechanisms influencing efficiency of RNA interference in insects. Pest. Manag. Sci. 2019, 75, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.M.; Song, H.; Yu, Z.; Biondi, M.; Bai, J.; Shi, X.; Weerasekara, S.M.; Hua, D.H.; Silver, K.; Zhang, J.; et al. Comparison of strategies for enhancing RNA interference efficiency in Ostrinia nubilalis. Pest. Manag. Sci. 2021, 77, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Vogel, E.; Santos, D.; Mingels, L.; Verdonckt, T; Broeck, J. V. RNA Interference in Insects: Protecting Beneficials and Controlling Pests. Front. Physiol. 2019, 9, 434563. [Google Scholar] [CrossRef] [PubMed]
- Girard, C.; Budin, K.; Boisnard, S.; Zhang, L.; Debuchy, R.; Zickler, D.; Espagne, E. RNAi-Related Dicer and Argonaute Proteins Play Critical Roles for Meiocyte Formation, Chromosome-Axes Lengths and Crossover Patterning in the Fungus Sordaria macrospora. Front. Cell Dev. Biol. 2021, 9, 684108. [Google Scholar] [CrossRef] [PubMed]
- Silver, K.; Cooper, A.M.; Zhu, K.Y. Strategies for enhancing the efficiency of RNA interference in insects. Pest. Manag. Sci. 2021, 77, 2645–2658. [Google Scholar] [CrossRef] [PubMed]
- Terenius, O.; Papanicolaou, A.; Garbutt, J.S.; Eleftherianos, I.; Huvenne, H.; Kanginakudru, S.; Albrechtsen, M.; An, C.; Aymeric, J.L.; Barthel, A.; et al. RNA interference in Lepidoptera: an overview of successful and unsuccessful studies and implications for experimental design. J. Insect Physiol. 2011, 57, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Lucena-Leandro, V.S.; Abreu, E.F.A.; Vidal, L.A.; Torres, C.R.; Junqueira, C.I.C.V.F.; Dantas, J.; Albuquerque, É. V.S. Current Scenario of Exogenously Induced RNAi for Lepidopteran Agricultural Pest Control: From dsRNA Design to Topical Application. Int. J. Mol. Sci. 2022, 23, 15836. [Google Scholar] [CrossRef] [PubMed]
- Christiaens, O.; Tardajos, M.G.; Martinez Reyna, Z.L.; Dash, M.; Dubruel, P.; Smagghe, G. Increased RNAi Efficacy in via the Formulation of dsRNA With Guanylated Polymers. Front. Physiol. 2018, 9, 316. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.G.; Robinson, K.E.; Asgari, S.; Mitter, N. Current scenario of RNAi-based hemipteran control. Pest. Manag. Sci. 2021, 77, 2188–2196. [Google Scholar] [CrossRef] [PubMed]
- Kaplanoglu, E.; Kolotilin, I.; Menassa, R.; Donly, C. Plastid Transformation of Micro-Tom Tomato with a Hemipteran Double-Stranded RNA Results in RNA Interference in Multiple Insect Species. Int. J. Mol. Sci. 2022, 23, 3918. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Peng, H.; Yao, Q.; Chen, H.; Xie, Q.; Tang, B.; Zhang, W. Developmental control of a lepidopteran pest Spodoptera exigua by ingestion of bacteria expressing dsRNA of a non-midgut gene. PloS One 2009, 4, e6225. [Google Scholar] [CrossRef] [PubMed]
- Zha, W.; Peng, X.; Chen, R.; Du, B.; Zhu, L.; He, G. Knockdown of midgut genes by dsRNA-transgenic plant-mediated RNA interference in the hemipteran insect Nilaparvata lugens. PloS One 2011, 6, e20504. [Google Scholar] [CrossRef] [PubMed]
- Prentice, K.; Christiaens, O.; Pertry, I.; Bailey, A.; Niblett, C.; Ghislain, M.; Gheysen, G.; Smagghe, G. RNAi-based gene silencing through dsRNA injection or ingestion against the African sweet potato weevil Cylas puncticollis (Coleoptera: Brentidae). Pest. Manag. Sci. 2017, 73, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Camargo, R.A.; Barbosa, G.O.; Possignolo, I.P.; Peres, L.E.; Lam, E.; Lima, J.E.; Figueira, A.; Marques-Souza, H. RNA interference as a gene silencing tool to control Tuta absoluta in tomato (Solanum lycopersicum). PeerJ. 2016, 4, e2673. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Liu, J.; Chen, Y.; Zhang, J.; Zhong, G. RNAi-mediated knockdown of α-Spectrin depresses reproductive performance in female Bactrocera dorsalis. Pesticide Biochem. Physiol. 2023, 196, 105611. [Google Scholar] [CrossRef] [PubMed]
- Arya, S.K.; Singh, S.; Upadhyay, S.K.; Tiwari, V.; Saxena, G.; Verma, P.C. RNAi-based gene silencing in Phenacoccus solenopsis and its validation by in planta expression of a double-stranded RNA. Pest. Manag. Sci. 2021, 77, 1796–1805. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Z.; Jiang, Y.X.; Li, M.W.; Li, J.W.; Zha, B.H.; Yang, G. Double-stranded RNA-degrading enzymes reduce the efficiency of RNA interference in Plutella xylostella. Insects 2021, 12, 712. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Xu, X.; Li, J.; Hull, J.J.; Chen, L.; Liang, G. A spray-induced gene silencing strategy for Spodoptera frugiperda oviposition inhibition using nanomaterial-encapsulated dsEcR. International Journal of Biological Macromolecules 2024, 281, 136503. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.F.; Liu, T.T.; Zhao, Y.Q.; Luo, J.; Feng, H.Y.; Zhou Gong, L.L.; Zhang, M.Q.; He, Y.Y.; Hull, J.J.; Dewer, Y.; et al. RNA interference-screening of potentially lethal gene targets in the white-backed planthopper Sogatella furcifera via a spray-induced and nanocarrier-delivered gene silencing system. Journal of Agricultural and Food Chemistry 2024, 72, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, H.; Wang, Y.; Wei, L.; Lyu, J.; Shan, Z.; Zhang, X.; Fan, D. RNA Interference Reveals the Impacts of CYP6CY7 on Imidacloprid Resistance in Aphis glycines. Insects 2024, 15, 188. [Google Scholar] [CrossRef] [PubMed]
- Saberi, E.; Mondal, M.; Paredes-Montero, J.R.; Nawaz, K.; Brown, J.K.; Qureshi, J.A. Optimal dsRNA Concentration for RNA Interference in Asian Citrus Psyllid. Insects 2024, 15, 58. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.X.; Bao, H.Q.; Yan, Z.C.; Wang, J.; Wang, S.; Li, Y.X. Knockdown of vitellogenin receptor based on minute insect RNA interference methods affects the initial mature egg load in the pest natural enemy Trichogramma dendrolimi. Insect Science 2024. [CrossRef] [PubMed]
- Liu, J.; Yang, Y.; Yang, Q.; Lin, X.; Liu, Y.; Li, Z.; Swevers, L. Successful oral RNA interference efficiency in the silkworm Bombyx mori through nanoparticle-shielded dsRNA delivery. Journal of Insect Physiology 2025, 104749. [Google Scholar] [CrossRef] [PubMed]
- Darweesh, A.F.; Fahmy, I.; Ali, M.; Elwahy, A. RNA interference of cysteine protease genes for the management of whitefly (Bemisia tabaci) by oral route. Egyptian Journal of Botany 2025, 65, 43–56. [Google Scholar] [CrossRef]
- Shimomura, K.; Sakita, K.; Terajima, T.; Tomizawa, M. Gene Silencing of Olfactory Receptor Coreceptor by Systemic RNA Interference in Callosobruchus maculatus. Journal of Chemical Ecology 2025, 51, 5. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.P.; Yang, X.B.; Yang, H.; Zhou, C.; Long, G.Y.; Jin, D.C. Knockdown of the β-N-acetylhexosaminidase genes by RNA interference inhibited the molting and increased the mortality of the white-backed planthopper, Sogatella furcifera. Pesticide Biochem. Physiol. 2025, 207, 106216. [Google Scholar]
- Bera, P.; Suby, S.B.; Dixit, S.; Vijayan, V.; Kumar, N.; Sekhar, J.C.; Vadassery, J. Identification of novel target genes for RNAi mediated management of the pest, Fall Armyworm (Spodoptera frugiperda, JE Smith). Crop Protection 2025, 187, 106972. [Google Scholar] [CrossRef]
- Schellens, S.; Lenaerts, C.; Pérez Baca, M.D.R.; Cools, D.; Peeters, P.; Marchal, E.; Vanden Broeck, J. Knockdown of the Halloween genes Spook, Shadow and Shade influences oocyte development, egg shape, oviposition and hatching in the desert locust. Int. J. Mol. Sci. 2022, 23, 9232. [Google Scholar] [CrossRef] [PubMed]
- Mun, S.; Young Noh, M.; Dittmer, N.T.; Muthukrishnan, S.; Kramer, K.J.; Kanost, M.R.; Arakane, Y. Cuticular protein with a low complexity sequence becomes cross-linked during insect cuticle sclerotization and is required for the adult molt. Sci. Rep. 2015, 5, 10484. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.; Dai, L.; Gao, H.; Sun, Y.; Liu, B.; Chen, H. Identification, expression patterns and RNA interference of aquaporins in Dendroctonus armandi (Coleoptera: Scolytinae) larvae during overwintering. Front. Physiol. 2019, 10, 967. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.M.; Ashfaq, M.; Khan, A.A.; Naseem, M.T.; Mansoor, S. Evaluation of potential RNA-interference-target genes to control cotton mealybug, Phenacoccus solenopsis (Hemiptera: Pseudococcuidae). Insect science 2018, 25, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Araujo, R.N.; Santos, A.; Pinto, F.S.; Gontijo, N.F.; Lehane, M.J.; Pereira, M.H. RNA interference of the salivary gland nitrophorin 2 in the triatomine bug Rhodnius prolixus (Hemiptera: Reduviidae) by dsRNA ingestion or injection. Insect biochemistry and molecular biology 2006, 36, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, D.; Yao, Q.; Zhang, J.; Dong, X.; Tian, H.; Chen, J.; Zhang, W. Feeding-based RNA interference of a trehalose phosphate synthase gene in the brown planthopper, Nilaparvata lugens. Insect Molecular Biology 2010, 19, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Shang, F.; Xiong, Y.; Xia, W.K.; Wei, D.D.; Wei, D.; Wang, J.J. Identification, characterization and functional analysis of a chitin synthase gene in the brown citrus aphid, Toxoptera citricida (Hemiptera, Aphididae). Insect Molecular Biology 2016, 25, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Shen, J. Target genes for RNAi in pest control: A comprehensive overview. Entomologia Generalis 2024, 44. [Google Scholar] [CrossRef]
- Pallis, S.; Alyokhin, A.; Manley, B.; Rodrigues, T.; Barnes, E.; Narva, K. Effects of Low Doses of a Novel dsRNA-based Biopesticide (Calantha) on the Colorado Potato Beetle. J. Econ. Entomol. 2023, 116, 456–461. [Google Scholar] [CrossRef] [PubMed]
- GreenLight Biosciences. Available online: https://www.greenlightbiosciences.com/international-crop-network-validates-ledprona-as-a-new-mode-of-action-group/ (accessed on 20 January 2025).
- Bachman, P.; Fischer, J.; Song, Z.; Urbanczyk-Wochniak, E.; Watson, G. Environmental fate and dissipation of applied dsRNA in soil, aquatic systems, and plants. Front. Plant Sci. 2020, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Nauen, R. IRAC: mode of action classification and insecticide resistance management. Pesticide Biochem. Physiol. 2015, 121, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Storer, N.; Porter, A.; Slater, R.; Nauen, R. Insecticide resistance management and industry: the origins and evolution of the insecticide resistance action committee (IRAC) and the mode of action classification scheme. Pest. Manag. Sci. 2021, 77, 2609–2619. [Google Scholar] [CrossRef] [PubMed]
- Khajuria, C.; Ivashuta, S.; Wiggins, E.; Flagel, L.; Moar, W.; Pleau, M.; Miller, K.; Zhang, Y.; Ramaseshadri, P.; Jiang, C. Development and characterization of the first dsRNA-resistant insect population from western corn rootworm, Diabrotica virgifera virgifera LeConte. PLoS One 2018, 13, e0197059. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Zhang, M.; Zhang, J. Characterization and potential mechanism of resistance to double-stranded RNA in willow leaf beetle, Plagiodera versicolora. Journal of Pest Science 2024, 97, 2217–2226. [Google Scholar] [CrossRef]
- Mishra, S.; Dee, J.; Moar, W.; Dufner-Beattie, J.; Baum, J.; Dias, N.P.; Alyokhin, A.; Buzza, A.; Rondon, S.I.; Clough, M.; et al. Selection for high levels of resistance to double-stranded RNA (dsRNA) in Colorado potato beetle (Leptinotarsa decemlineata Say) using non-transgenic foliar delivery. Scientific Reports 2021, 11, 6523. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Moar, W.; Jurat-Fuentes, J.L. Larvae of Colorado potato beetle (Leptinotarsa decemlineata Say) resistant to double stranded RNA (dsRNA) remain susceptible to small-molecule pesticides. Pest. Manag. Sci. 2024, 80, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Narva, K.; Toprak, U.; Alyokhin, A.; Groves, R.; Jurat-Fuentes, J.L.; Moar, W.; Nauen, R. Whipple S, Head G. Insecticide resistance management scenarios differ for RNA-based sprays and traits. Insect Mol Biol. [CrossRef]
- Christiaens, O.; Whyard, S.; Vélez, A.M.; Smagghe, G. Double-Stranded RNA Technology to Control Insect Pests: Current Status and Challenges. Front. Plant Sci. 2020, 11, 451. [Google Scholar] [CrossRef] [PubMed]
- Cedden, D.; Güney, G.; Rostás, M.; Bucher, G. Optimizing dsRNA sequences for RNAi in pest control and research with the dsRIP web platform. BMC Biol. 2025, 23, 114. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Peng, Y.; Zhang, H.; Wang, K.; Zhao, C.; Zhu, G.; Reddy Palli, S.; Han, Z. Off-target effects of RNAi correlate with the mismatch rate between dsRNA and non-target mRNA. RNA Biol. 2021, 18, 1747–1759. [Google Scholar] [CrossRef] [PubMed]
- Cedden, D.; Bucher, G. The quest for the best target genes for RNAi-mediated pest control. Insect Mol Biol. 2024. [CrossRef] [PubMed]
- Yang, J.; Cho, W.C.; Zheng, Y. Argonaute proteins: Structural features, functions and emerging roles. Journal of Advanced Research 2020, 24, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Höck, J.; Meister, G. The Argonaute protein family. Genome Biol. 2008, 9, 210. [Google Scholar] [CrossRef] [PubMed]
- Okamura, K.; Ishizuka, A.; Siomi, H.; Siomi, M.C. Distinct roles for Argonaute proteins in small RNA-directed RNA cleavage pathways. Genes Dev. 2004, 18, 1655–1666. [Google Scholar] [CrossRef] [PubMed]
- Hoang BTL, Fletcher SJ, Brosnan CA, Ghodke AB, Manzie N, Mitter N. RNAi as a Foliar Spray: Efficiency and Challenges to Field Applications. Int. J. Mol. Sci. 2022, 23, 6639. [Google Scholar] [CrossRef] [PubMed]
- Socha, W.; Kwasnik, M.; Larska, M.; Rola, J.; Rozek, W. Vector-borne viral diseases as a current threat for human and animal health—One Health perspective. Journal of Clinical Medicine 2022, 11, 3026. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Wang, B.; Lei, G.; Chen, G.; Liu, D. Advances in nanocarriers to improve the stability of dsRNA in the environment. Front. Bioeng. Biotechnol. 2022, 10, 974646. [Google Scholar] [CrossRef] [PubMed]
- Nitnavare, R.B.; Bhattacharya, J.; Singh, S. : Kour, A.; Hawkesford, M.J.; Arora, N. Next generation dsRNA-based insect control: Success so far and challenges. Front. Plant Sci. 2021, 12, 673576. [Google Scholar] [CrossRef] [PubMed]
- Narva, K.; Toprak, U.; Alyokhin, A.; Groves, R.; Jurat-Fuentes, J.L.; Moar, W.; Nauen, R.; Whipple, S.; Head, G. Insecticide resistance management scenarios differ for RNA-based sprays and traits. Insect Mol Biol. 2025. [Google Scholar] [CrossRef] [PubMed]
- Verdonckt, T.W.; Vanden Broeck, J. Methods for the Cost-Effective Production of Bacteria-Derived Double-Stranded RNA for in vitro Knockdown Studies. Front. Physiol. 2022, 13, 836106. [Google Scholar] [CrossRef] [PubMed]
- Palli, S.R. RNAi turns 25: contributions and challenges in insect science. Front. Insect Sci. 2023, 3, 1209478. [Google Scholar] [CrossRef] [PubMed]
- Egli, M.; Manoharan, M. Chemistry, structure and function of approved oligonucleotide therapeutics. Nucleic Acids Res. 2023, 51, 2529–2573. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Wang, K.; Fu, W.; Sheng, C.; Han, Z. Biochemical comparison of dsRNA degrading nucleases in four different insects. Front Physiol. 2018, 9, 624–10.3389. [Google Scholar] [CrossRef] [PubMed]
- Dias, N.; Stein, C.A. Antisense oligonucleotides: basic concepts and mechanisms. Mol Cancer Ther. 2002, 1, 347–355. [Google Scholar] [PubMed]
- Bachellerie, J.P.; Cavaillé, J.; Hüttenhofer, A. The expanding snoRNA world. Biochimie. 2002. [Google Scholar] [CrossRef] [PubMed]
- Will, C.L. , Lührmann, R., 2001. Spliceosomal UsnRNP biogenesis, structure and function. Curr Opin Cell Biol. [CrossRef]
- Oberemok, V.V.; Laikova, K.V.; Zaitsev, A.S.; Shumskykh, M.N.; Kasich, I.N.; Gal'chinsky, N.V.; Bekirova, V.V.; Makarov, V.V.; Agranovsky, A.A.; Gushchin, V.A.; et al. Molecular Alliance of Lymantria dispar Multiple Nucleopolyhedrovirus and a Short Unmodified Antisense Oligonucleotide of Its Anti-Apoptotic IAP-3 Gene: A Novel Approach for Gypsy Moth Control. Int. J. Mol. Sci. 2017, 18, 2446. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V. DNA markers in investigation of interaction between Lymantria dispar multiple nucleopolyhedrovirus and its host gypsy moth. Dissertation, Taurida National VI Vernadsky University, 2011.
- Cerio, R.J.; Vandergaast, R.; Friesen, P.D. Host insect inhibitor-of-apoptosis SfIAP functionally replaces baculovirus IAP but is differentially regulated by its N-terminal leader. J. Virol. 2010, 84, 11448–11460. [Google Scholar] [CrossRef] [PubMed]
- Manju, M.; Nirosha, V.; Tullika, T.; Mankhanniang, G. DNA insecticides: an emerging tool in pest management. AGRIALLIS. 2022, 4. [Google Scholar]
- Oberemok, V.V.; Skorokhod, O.A. Single-stranded DNA fragments of insect-specific nuclear polyhedrosis virus act as selective DNA insecticides for gypsy moth control. Pestic. Biochem. Physiol. 2014, 113, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V.; Laikova, K.V.; Zaitsev, A.S.; Gushchin, V.A.; Skorokhod, O.A. The RING for gypsy moth control: Topical application of fragment of its nuclear polyhedrosis virus anti-apoptosis gene as insecticide. Pestic Biochem Physiol. 2016, 131, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H.; Li, H.; Miao, X. Second-generation sequencing supply an effective way to screen RNAi targets in large scale for potential application in pest insect control. PloS One 2011, 6, e18644. [Google Scholar] [CrossRef] [PubMed]
- Warner, J.R. The economics of ribosome biosynthesis in yeast. Trends Biochem. Sci. 1999, 24, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V.; Useinov, R.Z.; Skorokhod, O.A.; Gal’chinsky, N.V.; Novikov, I.A.; Makalish, T.P.; Yatskova, E.V.; Sharmagiy, A.K.; Golovkin, I.O.; Gninenko, Y.I.; et al. Oligonucleotide insecticides for green agriculture: regulatory role of contact DNA in plant- insect interactions. Int. J. Mol. Sci. 2022, 23, 15681. [Google Scholar] [CrossRef] [PubMed]
- Useinov, R.Z.; Galchinsky, N.; Yatskova, E.; Novikov, I.; Puzanova, Y.; Trikoz, N.; Sharmagiy, A.; Plugatar, Y; Laikova, K. ; Oberemok, V. To bee or not to bee: creating DNA insecticides to replace non-selective organophosphate insecticides for use against the soft scale insect Ceroplastes Japonicus green. J. Plant. Prot. Res. 2020, 60, 406–409. [Google Scholar]
- Gal’chinsky, N.; Useinov, R.; Yatskova, E.; Laikova, K.; Novikov, I.; Gorlov, M.; Trikoz, N.; Sharmagiy, A.; Plugatar, Y.; Oberemok, V. A breakthrough in the efficiency of contact DNA insecticides: rapid high mortality rates in the sap-sucking insects Dynaspidiotus britannicus Comstock and Unaspis euonymi Newstead. J. Plant. Prot. Res. 2020, 60, 220–223. [Google Scholar] [CrossRef]
- Gal’chinsky, N.V.; Yatskova, E.V.; Novikov, I.A.; Useinov, R.Z.; Kouakou, N.J.; Kouame, K.F.; Kra, K.D.; Sharmagiy, A.K.; Plugatar, Y.V.; Laikova, K.V.; et al. Icerya purchasi Maskell (Hemiptera: Monophlebidae) control using low carbon footprint oligonucleotide insecticides. Int. J. Mol. Sci. 2023, 24, 11650. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V.; Gal'chinsky, N.V.; Useinov, R.Z.; Novikov, I.A.; Puzanova, Y.V.; Filatov, R.I.; Kouakou, N.J.; Kouame, K.F.; Kra, K.D.; Laikova, K.V. Four Most Pathogenic Superfamilies of Insect Pests of Suborder Sternorrhyncha: Invisible Superplunderers of Plant Vitality. Insects 2023, 14, 462. [Google Scholar] [CrossRef] [PubMed]
- Novikov, A.; Yatskova, E.; Bily, A.; Puzanova, Y.; Sharmagiy, A.; Oberemok, V. Efficient control of the obscure mealybug Pseudococcus viburni with DNA insecticides. In vitro cellular & developmental biology-animal. Springer, New York, USA.
- Puzanova, Y.V.; Novikov, I.A.; Bilyk, A.I.; Sharmagiy, A.K.; Plugatar, Y.V.; Oberemok, V.V. Perfect complementarity mechanism for aphid control: oligonucleotide insecticide macsan-11 selectively causes high mortality rate for Macrosiphoniella sanborni Gillette. Int. J. Mol. Sci. 2023, 24, 11690. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V.; Novikov, I.A.; Yatskova, E.V.; Bilyk, A.I.; Sharmagiy, A.K.; Gal'chinsky, N.V. Potent and selective ‘genetic zipper’ method for DNA-programmable plant protection: innovative oligonucleotide insecticides against Trioza alacris Flor. Chem. Biol. Technol. Agric. 2024, 11, 144. [Google Scholar] [CrossRef]
- Gavrilova, D.; Grizanova, E.; Novikov, I.; Laikova, E.; Zenkova, A.; Oberemok, V.; Dubovskiy, I. Antisense DNA acaricide targeting pre-rRNA of two-spotted spider mite Tetranychus urticae as efficacy-enhancing agent of fungus Metarhizium robertsii. Journal of Invertebrate Pathology. 2025, 108297. [Google Scholar] [CrossRef] [PubMed]
- IZ. Available online: https://en.iz.ru/en/1870413/maria-neduk-denis-gricenko/agrarian-evolution-dna-drug-will-destroy-main-enemy-greenhouse-crops (accessed on 16 April 2025).
- Oberemok, V.V.; Laikova, K.V.; Useinov, R.Z.; Gal’chinsky, N.V.; Novikov, I.A.; Gorlov, M.V.; Balykina, E.V.; Trikoz, N.N.; Yatskova, E.V.; Sharmagiy, A.K. High mortality of sap-sucking insects one week after topical application of DNA insecticides. In Vitro Cell. Dev. Biol. Anim. 2020, 56, 39. [Google Scholar]
- Plugatar, Y.V.; Chichkanova, E.S.; Yatskova, E.V.; Sharmagii, A.K.; Oberemok, V.V. An innovative method of Diaspis echinocacti Bouche control using DNA insecticide on Opuntia ficus-indica (L.) Mill. in the Nikitsky Botanical Garden, Crimea. South Russia: ecology Dev. 2021, 16, 119–128. [Google Scholar] [CrossRef]
- Oberemok, V.V.; Gal’chinsky, N.V. Oligonucleotide insecticides (contact unmodified antisense DNA biotechnology) and RNA biocontrols (double-stranded RNA technology): newly born fraternal twins in plant protection. bioRxiv 2024, 584797. [Google Scholar] [CrossRef]
- Oberemok, V.; Laikova, K.; Shumskykh, M.; Kenyo, I.; Kasich, I.; Deri, K.; Seidosmanova, E.; Krasnodubets, A.; Bekirova, V.; Gal'chinsky, N. A primary attempt of Leptinotarsa decemlineata control using contact DNA insecticide based on short antisense oligonucleotide of its CYP6B gene. J. Plant Prot. Res. 2018, 58, 106–108. [Google Scholar] [CrossRef]
- Du, Q.; Thonberg, H.; Wang, J.; Wahlestedt, C.; Liang, Z.A. A systematic analysis of the silencing effects of an active siRNA at all single-nucleotide mismatched target sites. Nucleic Acids Res. 2005, 33, 1671–1677. [Google Scholar] [CrossRef] [PubMed]
- Luige, O.; Karalė, K.; Bose, P.P.; Bollmark, M.; Tedebark, U.; Murtola, M.; Strömberg, R. Influence of sequence variation on the RNA cleavage activity of Zn2+-dimethyl-dppz-PNA-based artificial enzymes. RSC Adv. 2022, 12, 5398–5406. [Google Scholar] [CrossRef] [PubMed]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef] [PubMed]
- Mojica, M.J.; Juez, G.; Rodríguez-Valera, F. Transcription at different salinities of Haloferax mediterranei sequences adjacent to partially modified PstI sites. Mol. Microbiol. 1993, 9, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.; Embden, J.D.A.; Gaastra, W.; Schouls, L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 2002, 43, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Bolotin, A.; Quinquis, B.; Sorokin, A.; Ehrlich, S.D. Clustered regularly interspaced short palindrome repeats (CRISPRs) have spacers of extrachromosomal origin. Microbiology 2005, 151, 2551–2561. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Grishin, N.V.; Shabalina, S.A.; Wolf, Y.I.; Koonin, E.V. A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action. Biol. Direct. 2006, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Aljabali, A.A.A.; El-Tanani, M.; Tambuwala, М.М. Principles of CRISPR-Cas9 technology: Advancements in genome editing and emerging trends in drug delivery. Journal of Drug Delivery Science and Technology 2024, 92, 105338. [Google Scholar] [CrossRef]
- Asokan, R.; Rai, A.; Dash, S.; Manamohan, M.; Ashok, K.; Bhargava, C.N.; Wishard, R.; Pradhan, S.K.; Parvathy, M.S. Application of genome editing in entomology. Indian Journal of Entomology 2022, 96–103. [Google Scholar]
- Khan, M.D.; Ahmad, B.; Ahmed, S.F.; Ijaz, M.; Abdin, Z.U.; Ghafoor, I.; Siddiqui, G.M.; Adrees, T.; Ali, S.; Shahzadi, N.; Rafa, H.U. CRISPR-Cas genome editing in crops: A promising frontier for climate-smart agriculture. Phytopathogenomics and Disease Control 2023, 2, 71–77. [Google Scholar] [CrossRef]
- Wang, J.Y.; Doudna, J.A. CRISPR technology: A decade of genome editing is only the beginning. Science 2023, 379, eadd8643. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Pausch, P.; Doudna, J.A. Structural biology of CRISPR–Cas immunity and genome editing enzymes. Nature Reviews Microbiology 2022, 20, 641–656. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F. Development of CRISPR-Cas systems for genome editing and beyond. Quarterly Reviews of Biophysics. 2019, 52, e6. [Google Scholar] [CrossRef]
- Salum, Y.M.; Yin, A.; Zaheer, U.; Liu, Y.; Guo, Y.; He, W. CRISPR/Cas9-Based Genome Editing of Fall Armyworm (Spodoptera frugiperda): Progress and Prospects. Biomolecules 2024, 14, 1074. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Aumann, R.A.; Häcker, I.; Schetelig, M.F. CRISPR-based genetic control strategies for insect pests. J. Integr. Agric. 2023, 22, 651–668. [Google Scholar] [CrossRef]
- Ranian, K.; Zahoor, M.K.; Zulhussnain, M.; Ahmad, A. CRISPR/Cas9 mediated sex-ratio distortion by sex specific gene editing in Aedes aegypti. Saudi Journal of Biological Sciences 2022, 29, 3015–3022. [Google Scholar] [CrossRef] [PubMed]
- Zulhussnain, M.; Zahoor, M.K.; Ranian, K.; Ahmad, A.; Jabeen, F. CRISPR Cas9 mediated knockout of sex determination pathway genes in Aedes aegypti. Bulletin of Entomological Research. 2023, 113, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Wang, M.; Zeng, J.; Sun, H.; Wei, X.; Jiang, H.; Shentu, X.; Sun, D. CRISPR/Cas Technology in Insect Insecticide Resistance. Insects 2025, 16, 345. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Handler, A.M. CRISPR/Cas9-mediated gene editing in an exogenous transgene and an endogenous sex determination gene in the Caribbean fruit fly, Anastrepha suspensa. Gene 2019, 691, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.e.; Liu, Z.; Xu, J.; Li, X.; Bi, H.; Andongma, A.A.; Niu, C.; Huang, Y. Mutation of doublesex induces sex-specific sterility of the diamondback moth Plutella xylostella. Insect Biochem. Mol. Biol. 2019, 112, 103180. [Google Scholar] [CrossRef] [PubMed]
- Nourani, L.; Mehrizi, A.A.; Pirahmadi, S.; Pourhashem, Z.; Asadollahi, E.; Jahangiri, B. CRISPR/Cas advancements for genome editing, diagnosis, therapeutics, and vaccine development for Plasmodium parasites, and genetic engineering of Anopheles mosquito vector. Infect Genet Evol. 2023, 109, 105419. [Google Scholar] [CrossRef] [PubMed]
- Zamanian, M.; Andersen, E.C. Prospects and challenges of CRISPR/Cas genome editing for the study and control of neglected vector-borne nematode diseases. FEBS J. 2016, 283, 3204–3221. [Google Scholar] [CrossRef] [PubMed]
- Bimal Kumar Sahoo, B.M.; Mahesh Pathak, M.; Sushruta Boruah, S.; Kasturi Sarmah, K. BIO-ENGINEERED DEFENDERS: CRISPR-CAS9 UNLEASHING DISEASE-RESISTANT INSECTS TO CURB VECTOR-BORNE DISEASES", Futuristic Trends in Agriculture Engineering & Food Sciences; IIP Series; 2024; Volume 3, Book 6, Volume 3, pp. 42–52. [CrossRef]
- Xu, J.; Xu, X.; Zhan, S.; Huang, Y. Genome editing in insects: current status and challenges. National Science Review 2019, 6, 399–401. [Google Scholar] [CrossRef] [PubMed]
- Courtier-Orgogozo, V.; Morizot, B.; Boëte, C. Using CRISPR-based gene drive for agriculture pest control. EMBO reports. 2017, 18, 1481–1481. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.H.; Du, J.; Chu, C.Y.; Madhav, M.; Hughes, G.L.; Champer, J. Symbionts and gene drive: two strategies to combat vector-borne disease. Trends in Genetics 2022, 38, 708–723. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, L.; Wei, L.; Wang, Y.; Han, Z. Advancements and Future Prospects of CRISPR-Cas-Based Population Replacement Strategies in Insect Pest Management. Insects 2024, 15, 653. [Google Scholar] [CrossRef] [PubMed]
- Meccariello, A.; Hou, S.; Davydova, S.; Fawcett, J.D.; Siddall, A.; Leftwich, P.T.; Krsticevic, F.; Papathanos, P.A.; Windbichler, N. Gene drive and genetic sex conversion in the global agricultural pest Ceratitis capitata. Nature Communications 2024, 15, 372. [Google Scholar] [CrossRef] [PubMed]
- Oye, K.A.; Esvelt, K.; Appleton, E.; Catteruccia, F.; Church, G.; Kuiken, T.; Lightfoot, S.B.Y.; McNamara, J.; Smidler, A.; Collins, J.P. Regulating gene drives. Science 2014, 345, 626–628. [Google Scholar] [CrossRef] [PubMed]
- Shukla, J.N.; Palli, S.R. Tribolium castaneum Transformer-2 regulates sex determination and development in both males and females. Insect biochemistry and molecular biology 2013, 43, 1125–1132. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.J.; Gould, F.; Lorenzen, M.; Grubbs, N.; Edwards, O.; O’Brochta, D. Agricultural production: assessment of the potential use of Cas9-mediated gene drive systems for agricultural pest control. Journal of Responsible Innovation 2018, 5(sup1), S98–S120. [Google Scholar] [CrossRef]
- Tang, Y.H.; Zhang, X.; Dai, Z.C.; Li, H.; Yang, Y.; Zhao, T.J.; Yuan, D.Q.; Qian, W.L.; Cheng, D.J. CRISPR-Cas13-mediated RNA editing in the silkworm Bombyx mori. Zool Res. 2024, 45, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Bulle, M.; Sheri, V.; Aileni, M.; Zhang, B. Chloroplast Genome Engineering: A Plausible Approach to Combat Chili Thrips and Other Agronomic Insect Pests of Crops. Plants 2023, 12, 3448. [Google Scholar] [CrossRef] [PubMed]
- Han, W.K.; Yang, Y.L.; Si, Y.X.; Wei, Z.Q.; Liu, S.R.; Liu, X.L.; Yan, Q.; Dong, S.L. Involvement of GOBP2 in the perception of a sex pheromone component in both larval and adult Spodoptera litura revealed using CRISPR/Cas9 mutagenesis. Insect Biochemistry and Molecular Biology 2022, 141, 103719. [Google Scholar] [CrossRef] [PubMed]
- Gantz, V.M.; Jasinskiene, N.; Tatarenkova, O.; Fazekas, A.; Macias, V.M.; Bier, E.; James, A.A. Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. Proceedings of the National Academy of Sciences 2015, 112, E6736–E6743. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.H.; Chereddy, S.C.; Howell, J.L.; Palli, S.R. Genome editing in the fall armyworm, Spodoptera frugiperda: Multiple sgRNA/Cas9 method for identification of knockouts in one generation. Insect biochemistry and molecular biology 2020, 122, 103373. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Wang, J.; Bi, H.; Li, X.; Merchant, A.; Zhang, P.; Zhang, Q.; Zhou, X. CRISPR/Cas9-mediated mutagenesis of sex-specific doublesex splicing variants leads to sterility in Spodoptera frugiperda, a global invasive pest. Cells 2022, 11, 3557. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhao, T.; Liu, X.; Li, T.; He, L.; Wang, Q.; Wang, L.; Zhou, L. CRISPR/Cas9-Mediated Mutagenesis of Antennapedia in Spodoptera frugiperda. Insects 2023, 15, 16. [Google Scholar] [CrossRef] [PubMed]
- Anu, C.N.; Ashok, K.; Bhargava, C.N.; Dhawane, Y.; Manamohan, M.; Jha, G.K.; Asokan, R. CRISPR/Cas9 mediated validation of spermatogenesis-related gene, tssk2 as a component of genetic pest management of fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae). Archives of Insect Biochemistry and Physiology 2024, 116, e22121. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ma, Y.; Wang, F.; Yang, Y.; Wu, S.; Wu, Y. Disruption of nicotinic acetylcholine receptor α6 mediated by CRISPR/Cas9 confers resistance to spinosyns in Plutella xylostella. Pest. Manag. Sci. 2020, 76, 1618–1625. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Harvey-Samuel, T.; Yang, J.; You, M.; Alphey, L. CRISPR/Cas9-based functional characterization of the pigmentation gene ebony in Plutella xylostella. Insect Molecular Biology 2021, 30, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.Y.; Liang, L.Q.; Li, Q.F.; Zhu, Y.N.; Guo, Y.K.; Zheng, Q.L.; Lin, Y.; Yang, D.L.; Li, Z.G.; Su, S.K. CRISPR/Cas9 mediated knockout of Amyellow-y gene results in melanization defect of the cuticle in adult Apis mellifera. Journal of insect physiology 2021, 132, 104264. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Roy, M.C.; Al Baki, M.A.; Jung, J.K.; Lee, D.; Kim, Y. CRISPR/Cas9 mutagenesis against sex pheromone biosynthesis leads to loss of female attractiveness in Spodoptera exigua, an insect pest. PLoS One 2021, 16, e0259322. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.X.; Li, W.X.; Lyu, J.; Hu, Y.T.; Huang, G.; Zhang, W.Q. CRISPR/Cas9-mediated knockout of the NlCSAD gene results in darker cuticle pigmentation and a reduction in female fecundity in Nilaparvata lugens (Hemiptera: Delphacidae). Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 2021, 256, 110921. [Google Scholar]
- Fu, X.; Li, R.; Qiu, Q.; Wang, M.; Zhao, T.; Zhou, L. Study on the function of Helicoverpa armigera Wnt1 gene using CRISPR/Cas9 system. Journal of Asia-Pacific Entomology 2022, 25, 101869. [Google Scholar] [CrossRef]
- Bi, H.; Merchant, A.; Gu, J.; Li, X.; Zhou, X.; Zhang, Q. CRISPR/Cas9-mediated mutagenesis of abdominal-A and ultrabithorax in the Asian corn borer, Ostrinia furnacalis. Insects 2022, 13, 384. [Google Scholar] [CrossRef] [PubMed]
- Konu, M.; Kulmuni, J.; Viljakainen, L. Genetic modification of the ant Lasius niger using CRISPR-Cas9 technology. Insect Molecular Biology 2023, 32, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Bi, H.; Xu, X.; Li, X.; Wang, Y.; Zhou, S.; Huang, Y. CRISPR/Cas9-mediated Serine protease 2 disruption induces male sterility in Spodoptera litura. Front. Physiol. 2022, 13, 931824. [Google Scholar] [CrossRef] [PubMed]
- Shirk, B.D.; Shirk, P.D.; Furlong, R.B.; Scully, E.D.; Wu, K.; Siegfried, B.D. Gene editing of the ABC Transporter/White locus using CRISPR/Cas9-mediated mutagenesis in the Indian Meal Moth. Journal of Insect Physiology 2023, 145, 104471. [Google Scholar] [CrossRef] [PubMed]
- Ashok, K.; Bhargava, C.N.; Babu, K.P.; Rohan, W.; Manamohan, M.; Rai, A.; Sanjay, K.P.; Parvathy, M.S.; Kennedy, J.S.; Asokan, R. First report on CRISPR/Cas9 mediated editing of the eye colour gene, tryptophan 2, 3-dioxygenase in egg plant shoot and fruit borer Leucinodes orbonalis Guenée (Lepidoptera: Crambidae). Journal of Asia-Pacific Entomology 2023, 26, 102031. [Google Scholar] [CrossRef]
- Ashok, K.; Bhargava, C.N.; Venkatesh, R.; Balasubramani, V.; Murugan, M.; Geethalakshmi, V.; Manamohan, M.; Jha, G.K.; Asokan, R. Molecular characterization and CRISPR/Cas9 validation of the precursor of egg yolk protein gene, vitellogenin of Leucinodes orbonalis Guenée (Lepidoptera: Crambidae). Gene 2025, 933, 148925. [Google Scholar] [CrossRef] [PubMed]
- Ashok, K.; Bhargava, C.N.; Asokan, R.; Pradeep, C.; Kennedy, J.S.; Rai, A.; Manamohan, M. First report on the utility of pupal case for early determination of CRISPR/Cas9 ribonucleoprotein mediated genomic edits in the oriental fruit fly, Bactrocera dorsalis (Hendel) (Tephritidae: Diptera). Archives of Insect Biochemistry and Physiology 2023, 113, e22024. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, C.N.; Ashok, K.; Asokan, R.; Prasad Babu, K.; Parvathy, M.S.; Yogi, D.; Shashikala, T.; Chiranth, R.K.; Ashok, U.; Harsha, C.G. CRISPR/Cas9 Mediated Editing of the white (wh) locus Affects Body Size and Reproduction of the Oriental Fruit Fly, Bactocera dorsalis (Hendel). Agricultural Research 2024, 1–8. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Karuppannasamy, A.; Sujatha, P.M.; Nagaraja, B.C.; Narayanappa, A.C.; Chalapathi, P.; Dhawane, Y.; Bynakal, S.; Riegler, M.; Maligeppagol, M.; et al. Embryonic microinjection of ribonucleoprotein complex (Cas9+ sgRNA) of white gene in melon fly, Zeugodacus cucurbitae (Coquillett) (Diptera: Tephritidae) produced white eye phenotype. Archives of Insect Biochemistry and Physiology 2023, 114, e22059. [Google Scholar] [CrossRef] [PubMed]
- Sujatha, P.M.; Karuppannasamy, A.; Chalapathi, P.; Dhawane, Y.; Narasimhappa, N.S.; Narayanappa, A.C.; Munikrishnappa, V.K.T.; Chikmagalur Nagaraja, B.; Thalooru, S.; Kesavan, S.; et al. CRISPR/Cas9 Ribo nucleoprotein complex-mediated editing of the OBP13 gene affected the response of male Bactrocera dorsalis (Diptera: Tephritidae) to methyl eugenol. Journal of Applied Entomology 2024. [CrossRef]
- Yadav, A.K.; Butler, C.; Yamamoto, A.; Patil, A.A.; Lloyd, A.L.; Scott, M.J. CRISPR/Cas9-based split homing gene drive targeting doublesex for population suppression of the global fruit pest Drosophila suzukii. Proceedings of the National Academy of Sciences 2023, 120, e2301525120. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; James, B.; Williams, M.; Bachler, A.; Tay, W.T.; Walsh, T.; Frese, M. Cry1 resistance in a CRISPR/Cas9-mediated HaCad1 gene knockout strain of the Australian cotton bollworm Helicoverpa armigera conferta (Lepidoptera: Noctuidae). Pest. Manag. Sci. 2025, 81, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, Y.; Nakamura, T.; Watanabe, T.; Barnett, A.A.; Tomonari, S.; Ylla, G.; Whittle, C.A.; Noji, S.; Mito, T.; Extavour, C.G. Establishment of CRISPR/Cas9-based knock-in in a hemimetabolous insect: targeted gene tagging in the cricket Gryllus bimaculatus. Development 2025, 152. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Bauer, D.E.; Chiarle, R. Assessing and advancing the safety of CRISPR-Cas tools: from DNA to RNA editing. Nat Commun. 2023, 14, 212. [Google Scholar] [CrossRef] [PubMed]
- Richardson, C.; Kelsh, R.N.; J Richardson, R. New advances in CRISPR/Cas-mediated precise gene-editing techniques. Dis. Model. Mech. 2023, 16, dmm049874. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Munawar, N.; Khan, Z.; Qusmani, A.T.; Khan, S.H.; Jamil, A.; Ashraf, S.; Ghouri, M.Z.; Aslam, S.; Mubarik, M.S.; et al. An Outlook on Global Regulatory Landscape for Genome-Edited Crops. Int. J. Mol. Sci. 2021, 22, 11753. [Google Scholar] [CrossRef] [PubMed]
- Asad, M.; Chang, Y.; Liao, J.; Yang, G. CRISPR/Cas9 Genome Editing in the Diamondback Moth: Current Progress, Challenges, and Prospects. Int. J. Mol. Sci. 2025, 26, 1515. [Google Scholar] [CrossRef] [PubMed]
- Komal, J.; Desai, H.R.; Samal, I.; Mastinu, A.; Patel, R.D.; Kumar, P.V.D.; Majhi, P.K.; Mahanta, D.K.; Bhoi, T.K. Unveiling the Genetic Symphony: Harnessing CRISPR-Cas Genome Editing for Effective Insect Pest Management. Plants (Basel) 2023, 12, 3961. [Google Scholar] [CrossRef] [PubMed]
- Moon, T.T.; Maliha, I.J.; Khan, A.A.M.; Chakraborty, M.; Uddin, M.S.; Amin, M.R.; Islam, T. CRISPR-Cas Genome Editing for Insect Pest Stress Management in Crop Plants. Stresses 2022, 2, 493–514. [Google Scholar] [CrossRef]
Figure 1.
Main route of RNAi used for creation of dsRNA insecticides (exogenous siRNA pathway): 1 – dsRNA cleavage by Dicer; 2 – silencing complex formation and silencing complex activation; 3 – target mRNA degradation by Ago.
Figure 1.
Main route of RNAi used for creation of dsRNA insecticides (exogenous siRNA pathway): 1 – dsRNA cleavage by Dicer; 2 – silencing complex formation and silencing complex activation; 3 – target mRNA degradation by Ago.
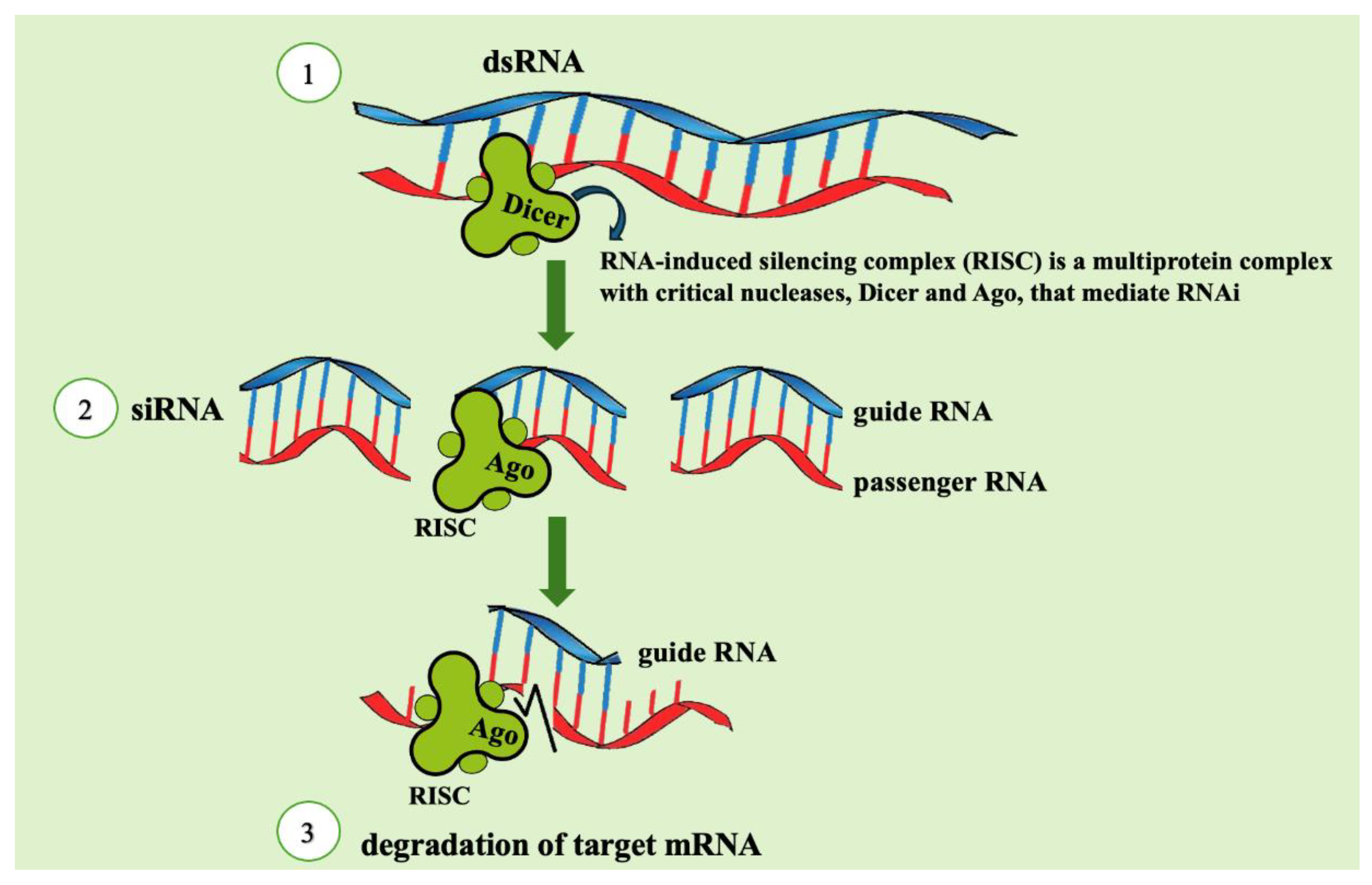
Figure 2.
Advanced features of CUADb and DNA containment (DNAc) as the main mechanism of action of oligonucleotide insecticides (olinscides): 1 – antisense DNA sequences complementary to rRNA and/or pre-rRNA of pests are used as oligonucleotide insecticides; 2 – formation of duplex between DNA (oligonucleotide insecticide) and RNA (rRNA and/or pre-rRNA) of the pests; 3 – at first step of DNAc, antisense DNA oligonucleotide (oligonucleotide insecticide) ‘arrests’ target rRNA and/or pre-rRNA) and interferes with normal functioning and biogenesis of ribosomes (‘arrested’ ribosomes); 4 – at second step of DNAc, rRNase cleaves target rRNA and substantial decrease in its concentration occurs.
Figure 2.
Advanced features of CUADb and DNA containment (DNAc) as the main mechanism of action of oligonucleotide insecticides (olinscides): 1 – antisense DNA sequences complementary to rRNA and/or pre-rRNA of pests are used as oligonucleotide insecticides; 2 – formation of duplex between DNA (oligonucleotide insecticide) and RNA (rRNA and/or pre-rRNA) of the pests; 3 – at first step of DNAc, antisense DNA oligonucleotide (oligonucleotide insecticide) ‘arrests’ target rRNA and/or pre-rRNA) and interferes with normal functioning and biogenesis of ribosomes (‘arrested’ ribosomes); 4 – at second step of DNAc, rRNase cleaves target rRNA and substantial decrease in its concentration occurs.
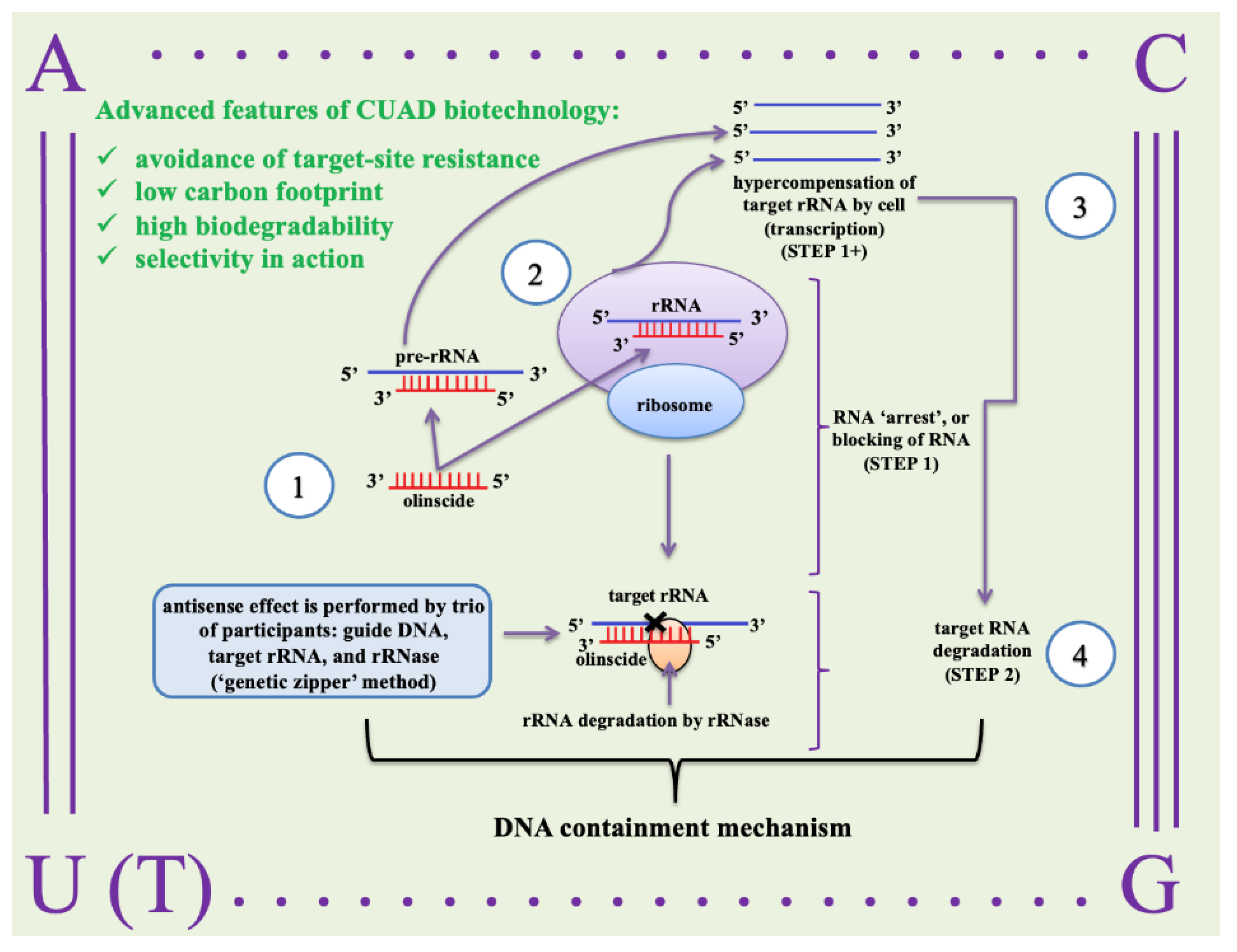
Figure 3.
CRISPR/Cas9 system used for insect pest control: 1 – sgRNA-Cas9 complex formation; 2 – formation of RNA-DNA duplex with target DNA; 3 – cleavage of target double-stranded DNA by Cas9; 4 – DNA repair through non-homologous end joining (NHEJ) or homologous-dependent repair (HDR); * crRNA – CRISPR RNA; tracrRNA – trans-activating crRNA.
Figure 3.
CRISPR/Cas9 system used for insect pest control: 1 – sgRNA-Cas9 complex formation; 2 – formation of RNA-DNA duplex with target DNA; 3 – cleavage of target double-stranded DNA by Cas9; 4 – DNA repair through non-homologous end joining (NHEJ) or homologous-dependent repair (HDR); * crRNA – CRISPR RNA; tracrRNA – trans-activating crRNA.
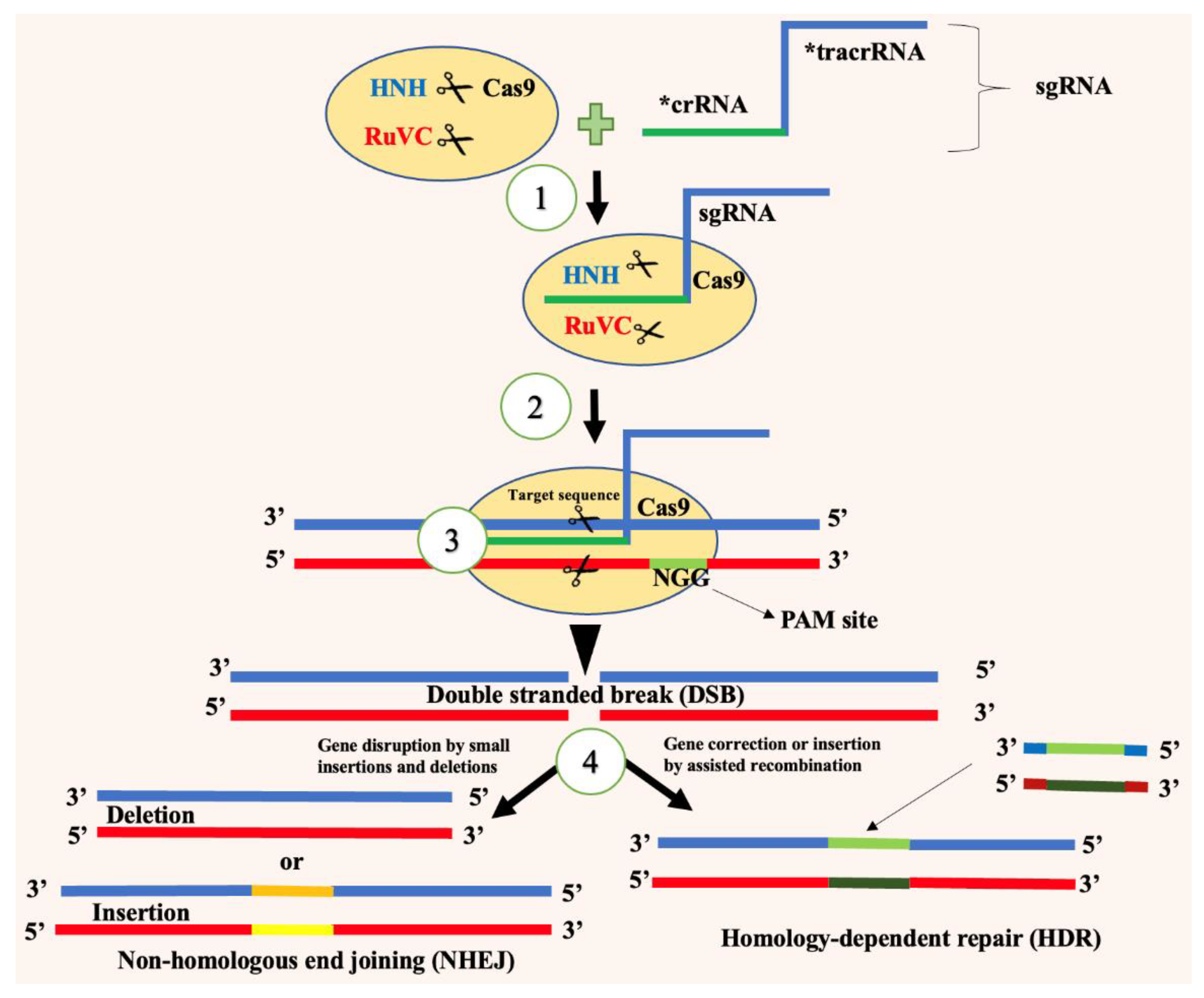

Table 1.
Antisense technologies based on unmodified nucleic acids and used for insect pest control: guide nucleic acid (RNA or DNA) forms duplex with target nucleic acid (RNA or DNA) and the target nucleic acid is cleaved by specific nuclease.
Table 1.
Antisense technologies based on unmodified nucleic acids and used for insect pest control: guide nucleic acid (RNA or DNA) forms duplex with target nucleic acid (RNA or DNA) and the target nucleic acid is cleaved by specific nuclease.

Table 2.
List of pest species successfully targeted by RNAi.
| Sl. No. | Names of model insects | Targeted gene(s) | Affected processes | References |
|---|---|---|---|---|
| 1 | Beet armyworm, Spodoptera exigua (Lepidoptera) | Chitin synthase gene A | Chitin synthesis | [40] |
| 2 | Brown planthopper, Nilaparvata lugens (Hemiptera) | NlHT1, Nlcar, Nltry | Digestive system | [41] |
| 3 | African sweet potato weevil, Cylas puncticollis (Coleoptera) | Snf7 | Digestive system | [42] |
| 4 | Tomato pinworm, Tuta absoluta (Lepidoptera) | Vacuolar ATPase-A and Arginine kinase | High mortality | [43] |
| 5 | Oriental fruit fly, Bactrocera dorsalis (Diptera) | α-Spectrin | Oviposition and ovary size | [44] |
| 6 | Cotton mealybug, Phenacoccus solenopsis (Hemiptera) | Krüppel homologue-1, ADP-ATP/Translocase, IDGF-1 | Not specified | [45] |
| 7 | Diamondback moth, Plutella xylostella (Lepidoptera) | PxCht | Chitin synthesis | [46] |
| 8 | Fall armyworm, Spodoptera frugiperda (Lepidoptera) | Met, EcR, USP | Reproductive system, fertility | [47] |
| 9 | White-backed planthopper, Sogatella furcifera (Hemiptera) | hsc70-3, PP-α | Insect metamorphosis | [48] |
| 10 | Soybean aphid, Aphis glycines (Hemiptera) | Cytochrome P450 monooxygenases (CYP450s) | Insect resistance | [49] |
| 11 | Asian citrus psyllid, Diaphorina citri (Hemiptera) | CHC, vATPase-A, Snf7 | Transmembrane system | [50] |
| 12 | Trichogramma dendrolimi (Lepidoptera) | Vitellogenin receptor (VgR) | Female reproductive system | [51] |
| 13 | Domestic silk moth, Bombyx mori (Lepidoptera) | BmToll9-2 | Chitin synthesis | [52] |
| 14 | Silverleaf whitefly, Bemisia tabaci (Hemiptera) | Cysteine protease | Digestive system | [53] |
| 15 | Cowpea weevil, Callosobruchus maculatus (Coleoptera) | Olfactory receptor coreceptor (Cmac\Orco) | Insect sensory system | [53] |
| 16 | White-backed planthopper, S. furcifera (Hemiptera) | β-N-acetylhexosaminidase genes | Insect metamorphosis | [55] |
| 17 | Fall armyworm, S. frugiperda (Lepidoptera) | COPIα, COPIβ, GSTU1 | Insect reproduction | [56] |
| 18 | Desert locust, Schistocerca gregaria (Orthoptera) | Cytochrome P450 | Ecdysteroid pathway | [57] |
| 19 | Red flour beetle, Tribolium castaneum (Coleoptera) | CPAPs | Cuticular proteins | [58] |
| 20 | Chinese white pine beetle, Dendroctonus armandi (Coleoptera) | Aquaporin | Osmoregulation | [59] |
| 21 | Cotton mealybug, P. solenopsis (Hemiptera) | Bursicon, V-ATPase | Cuticle hardening and V-ATPases act as proton pumps | [60] |
| 22 | Kissing bug, Rhodnius prolixus (Hemiptera) | Nitrophorin 2 (NP2) | Anticoagulant and apyrase activities in saliva | [61] |
| 23 | Brown plant hopper, N. lugens (Hemiptera) | NlTPS | Enzymatic activity | [62] |
| 24 | Citrus aphid, Toxoptera citricida (Hemiptera) | TCiCHS | Chitin synthesis | [63] |
| 25 | Potato psyllid, Bactericera cockerelli (Hemiptera) | SUC1, ST4 | Osmoregulatory | [64] |
Table 3.
List of pest species successfully targeted by CUADb.
| Sl. No. | Insects name | Targeted gene(s) | Affected system | References |
|---|---|---|---|---|
| 1 | Euonymous scale, Unaspis euonymi (Hemiptera) | 28S rRNA | Protein biosynthesis | [104,112] |
| 2 | Holly scale, Dynaspidiotus Britannicus (Hemiptera) | 28S rRNA | Protein biosynthesis | [5,104] |
| 3 | Japanese wax scale, Ceroplastes japonicus (Hemiptera) | 28S rRNA | Protein biosynthesis | [103] |
| 4 | Cactus scale, Diaspis echinocacti (Hemiptera) | 28S rRNA | Protein biosynthesis | [113] |
| 5 | Bay sucker, Trioza alacris (Hemiptera) | ITS2 of pre-rRNA and 28S rRNA | Protein biosynthesis | [109] |
| 6 | Cottony cushion scale, Icerya purchase (Hemiptera) | 28S rRNA | Protein biosynthesis | [105] |
| 7 | Chrysanthemum aphid, Macrosiphoniella sanborni (Hemiptera) | ITS2 of pre-rRNA | Protein biosynthesis | [108] |
| 8 | Mealybug, Pseudococcus viburni (Hemiptera) | 5.8S, 18S and 28S rRNA | Protein biosynthesis | [107] |
| 9 | Laureal scale, Aonidia lauri (Hemiptera) | 28S rRNA | Protein biosynthesis | [5] |
| 10 | Soft scale, Coccus hesperidum (Hemiptera) | 28S rRNA | Protein biosynthesis | [102] |
| 11 | Two-spotted spider mite, Tetranychus urticae (Trombidiformes) | ITS2 of pre-rRNA | Protein biosynthesis | [110] |
| 12 | Grey pine aphid, Schizolachnus pineti (Hemiptera) | ITS2 of pre-rRNA | Protein biosynthesis | [13] |
| 13 | Large pine aphid, Cinara pinea (Hemiptera) |
ITS2 of pre-rRNA | Protein biosynthesis | [13] |
| 14 | Pine needle aphid, Eulachnus rileyi (Hemiptera) | ITS2 of pre-rRNA | Protein biosynthesis | [13] |
Table 4.
List of pest species successfully targeted by CRISPR/Cas system in insects.
| Sl. No. | Insect name | Target gene(s) | Affected system | Reference |
|---|---|---|---|---|
| 1 | Mosquito, Anopheles stephensi (Diptera) | Kynurenine hydroxylase | Parasite-resistance | [151] |
| 2 | Fall armyworm, S. frugiperda (Lepidoptera) |
Ebony Doublesex (dsx) (Sfdsx) Antennapedia (Antp) Spermatogenesis-related, tssk2 |
Melanin biosynthesis Sex differentiation Insect thorax and wing development Male reproductive system |
[152-155] |
| 3 | Diamondback moth, P. xylostella (Lepidoptera) |
Yellow Ebony LW-opsin |
Body pigmentation Body pigmentation Efficiency of phototaxis |
[46,156,157] |
| 4 | European bee, Apis mellifera (Hymenoptera) | Amyellow-y | Melanization in cuticle | [158] |
| 5 | Beet armyworm, S. exigua (Lepidoptera) | Desaturase (SexiDES5) | Sex pheromone biosynthesis | [159] |
| 6 | Brown planthopper, N. lugens (Hemiptera) | Cysteine sulfinic acid decarboxylase (CSAD) | Melanin metabolism | [160] |
| 7 | Chickpea pod borer, H. armigera (Lepidoptera) | Wnt1 | Segmentation, appendage development, and pigmentation | [161] |
| 8 | Asian corn borer, Ostrinia furnacalis (Lepidoptera) | Abdominal-A (Abd-A) and Ultrabithorax (Ubx) | Anatomical structure formation | [162] |
| 9 | Black garden ant, Lasius niger (Hymenoptera) | Cinnabar | Eye pigmentation | [163] |
| 10 | Common cutworm, S. litura (Lepidoptera) |
Serine protease 2 Odorant-binding proteins |
Male sterility Perception of a sex pheromone |
[150, 164] |
| 11 | Indian meal moth, Plodia interpunctella (Lepidoptera) | ATP binding cassette (ABC) proteins | Eye pigmentation | [165] |
| 12 | Eggplant shoot and fruit borer, Leucinodes orbonalis (Lepidoptera) |
Tryptophan 2, 3-dioxygenase Vitellogenin (Vg) |
Eye pigmentation Female reproductive system |
[166,167] |
| 13 | Mango fruit fly, B. dorsalis (Diptera) |
White White locus OBP13 |
Eye pigmentation Eye pigmentation Methyl eugenol |
[168-171] |
| 14 | Pomace fly, Drosophila suzukii (Diptera) | Doublesex | Population suppression | [172] |
| 15 | Australian cotton bollworm, H. armigera conferta (Lepidoptera) | Cadherin | Cry1Ac resistance | [173] |
| 16 | Cricket, Gryllus bimaculatus (Orthoptera) | Laccase 2 (Gb-lac2) | Cuticle system pigmentation | [174] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



