Submitted:
16 April 2025
Posted:
17 April 2025
You are already at the latest version
Abstract
This overview explores the complex relationship between environmental factors, particularly obesity, and the timing of puberty, with a focus on how hormonal and genetic interactions are influenced by external conditions. Puberty (Gonadarche) is characterized by the activation of the Hypothalamic - Pituitary - Gonadal (HPG) axis. The onset and progression of puberty vary significantly among individuals, primarily due to genetic factors, with key genes like KISS1 and MKRN3 playing a crucial role. Cohesively, this paper emphasizes that environmental factors, particularly obesity and exposure to endocrine-disrupting chemicals (EDCs) have become a significant facet affecting the timing of puberty. Childhood obesity has significantly risen in recent decades and the age of pubertal onset has declined over the same period. Obesity greatly disrupts hormone regulation in pre-pubertal children. Leptin accelerates the onset of puberty in girls but not in boys. The underlying mechanism is proposed to be the increase in Kiss1/GnRH signalling. On the contrary excess leptin in boys supresses the testosterone production by increasing oestrogen conversion. Low adiponectin in obese girls may contribute to earlier puberty due to reduced inhibition of the Kiss1/GnRH signalling. Low adiponectin in boys is linked to delayed puberty due to its role in maintaining insulin sensitivity and testosterone production. Hyperinsulinemia influences pubertal timing through central and peripheral mechanisms. Insulin acting synergistically with leptin promotes earlier onset of puberty in girls but not in boys. The effects of exposure to certain EDCs, mostly obesogenic that mimic the action of natural hormones on the timing of puberty is still unclear, hence the need for further research on this topic is required. Addressing and preventing obesity in children could potentially mitigate these alterations in pubertal timing.
Keywords:
puberty
; obesity
; endocrine disruptors
; insulin
; adipokines
Introduction
Puberty is defined as the physiological process where a child achieves physical and sexual maturity, and reproductive ability [1]. During puberty, the release of pubertal hormones induces a wide array of changes, encompassing emotional, biological, and physical transformations [1,2]. The release of sex steroids is governed by the hypothalamic-pituitary-gonadal (HPG) axis which is activated after the age of 8 years and 9 years in girls in boys respectively; collectively these changes are known as gonadarche. Prior to gonadarche the reticularis zone within the adrenals is often activated resulting in adrenarche. The activation of the HPG axis is determined by genetic loading and influenced by environmental factors. Certain genes need to be activated or inhibited which leads to the release of certain hormones. The genetic variations among individuals largely account for the significant variability observed in pubertal development [3,4].
The exact timing of puberty remains a mystery and as such, various studies have identified the effect intrinsic genetic pathways have on the onset of puberty. Considering the fact that puberty is also affected by external sources such as environmental factors and nutrition, this paper focuses on the effects that the environment has on pubertal timing [5]. Over the last century the average age that children enter pubertal development puberty has dropped by 3 years and approximately every 25 years this age drops by around 6 – 9 months. This statistic follows secular trends that have been observed in recent changes regarding average childhood Body Mass Index (BMI), showing that, compared to 20 years ago, more children are either on or above the 95th percentile in BMI nowadays, clinically classifying them as obese [6,7]. In addition to the increasing BMI, there has also been an increasing trend in the consumption of endocrine-disrupting-chemicals (EDCs) amongst young children. To that, obesity causes disturbances to the release of pubertal hormones, by disrupting both central and peripheral signalling pathways. This ranges from simple disruptions in the HPG axis, to complex changes to genetic components that regulate the endocrine milieu.
The Timing of Puberty
The onset of the puberty is primarily driven by gonadarche, a critical process involving the reactivation of the hypothalamic-pituitary-gonadal (HPG) axis, which subsequently leads to the production and release of gonadal hormones [5,8]. When the axis is activated. After the activation of this pathway, the hypothalamus starts releasing GnRH in a pulsatile manner which is subsequently detected by the pituitary, prompting it to trigger a hormonal response. This response starts releasing follicle stimulating hormone (FSH) and luteinizing hormone (LH) which in turn, triggers a response in the gonads [9,10]. However, the knock-on effects this has differs between boys and girls. In boys, FSH causes increased sperm production, and LH causes an influx of testosterone. In girls, FSH stimulates an increase in oestrogen, while LH promotes a rise in progesterone [11]. These gonadal hormones in turn exert a negative feedback effect on the hypothalamus and pituitary inhibiting the release of GnRH and Gonadotrophins respectively [12,13]. This collectively regulates the axis via the aforementioned loops which regulate hormonal production during puberty, and thus control the stages it goes through [14]. It is imperative to keep in mind that these endocrinological pathways are affected by a myriad of genetic, endocrinological and, most importantly, environmental factors [1].
Genetic Factors
The onset of puberty varies greatly amongst individuals. This is mainly due to the fact that it is affected by a multitude of factors, one of them being genetic. For this reason, researchers often consider an age range for the onset of puberty rather than a set age. The range is set from 8-13 years and 9-14 years for girls and boys, respectively [1].
To elaborate on the aforementioned genetic factors, certain genes must be activated to initiate essential hormone release, vital to the initiation of puberty. The genes involved are categorized as follows: puberty activating (PA), puberty inhibiting (PI), and dual effect genes. The main PA genes are KISS1, NPY, and LEP, as they play a role in the start of puberty and the stimulation of the HPG axis [15,16].
Cohesively, the main PI genes are GABA, DLK1 and MKRN3, which are all expressed in the hypothalamus, stop puberty from starting until the child is capable of coping with the physiological changes. More specifically, if changes in these genes occur, increasing the level of their expression, then puberty will be delayed. Conversely, in mutations that reduce PI gene expression, puberty is initiated at an earlier age [17]. Out of the PI genes expressed in the hypothalamus, one stands out as the most important, the KISS1 gene which when expressed, signals the hypothalamus to release GnRH. This gene is inhibited by the aforementioned DLK1 gene which stops the KISS1 from binding to its receptors, subsequently halting early onset puberty [18]. To clarify, KISS1 binds to its receptor, KISS1R, also known as GPR54 which are found on GnRH neurons [19]. It should be noted that this process is only possible if the genes MKRN3, DLK1, and GABA are inhibited, which allows for the KISS1 gene to bind to its receptor [20,21,22].
Adrenarche
Adrenarche refers to the process of maturation of the adrenal glands and the increased production of dehydroepiandrosterone (DHEA) and dehydroepiandrosterone sulphate (DHEAS) [23,24]. The exact underlying mechanism of activation of zona reticularis of the adrenals is still unknown but it could be influenced by the Hypothalamic Pituitary Adrenal (HPA) axis. Androgens in turn activate HPG axis, implying that through an indirect pathway, the HPA axis plays a role in the onset of puberty [25]. Oestradiol enhances the activity of the HPA axis facilitating the activation of the HPG axis, whereas testosterone suppresses the activity of the HPA axis thus indirectly influence the HPG axis [24,26].
Body Weight and Puberty
Childhood Obesity
Obesity is among the most prevalent chronic conditions in children and adolescents. It is typically defined using the Body Mass Index (BMI) considering the individual’s weight and height and age. According to the World Health Organization (WHO), for children aged 5–19, obesity is determined by a BMI-for-age that is more than two standard deviations above the WHO Growth Charts. For children less than 5 years of age, obesity is defined as a weight-for-height greater than three standard deviations above the WHO Child Growth Standards median [27]. According to the United Nations Children’s Fund (UNICEF), the number of children under the age of 5 years that were overweight or obese has increased from 30 million in the year 2000 to 37 million in 2024. The NCD Risk Factor Collaboration (NCD-RisC) further supports this increasing trend as it estimates that the percentage of overweight and obese children aged 5 to 19 years has increased from 4% in 1975 to approximately 20% in 2022 [28]. The underlying causes behind these obesity trends are complex and involve multiple factors, often due intricate interactions between biological, genetic, socioeconomic, environmental, and cultural influences [29].
EDCs and Puberty
In recent years, particular attention has been drawn towards environmental factors such as endocrine-disrupting chemicals (EDCs), which are known for their ability to imitate natural hormones and disrupt hormonal balance, playing a contributing to the modern obesity epidemic [30]. Moreover, the rising prevalence of EDCs, coupled with their potential to interfere with the endocrine system, has sparked interest among researchers investigating the link between EDCs and the altered onset of puberty as observed in recent times.
As mentioned, EDCs have the ability to interfere with the hormonal regulation of puberty through various mechanisms. For example, they may cause epigenetic changes in pubertal genes, act on peripheral tissues like fat cells or adrenal glands, increasing adrenal androgen levels, or influence pathways in the central control systems of puberty, through the HPG axis [31]. In addition, some EDCs have oestrogenic activity and can mimic naturally occurring oestrogens in the body [32]. In fact, a recent study has shown that early pre-pubertal exposure to even very low, environmentally relevant doses of the common EDC, such as Bisphenol A (BPA), can accelerate the onset of puberty, yet decrease reproductive parameters in female mice [33]. Similarly, another EDC that was studied for its ability to influence the reproductive system is the plasticiser, bis (2-ethylhexyl) phthalate (DEHP). A study exposing juvenile rats to DEHP levels comparable to exposure levels in Italian children, during particularly vulnerable peri-pubertal periods, revealed sex-specific metabolic disruptions. In boys, DEHP exposure delayed reproductive development, suggesting anti-androgenic effects, while in girls, the thyroid emerged as a potential target for DEHP toxicity [34,35]. Furthermore, exposing female rats to a mixture of oestrogenic and anti-androgenic EDCs revealed that while the first generation remained largely unaffected, subsequent offspring generations (F2 and F3) experienced significant reproductive issues. These issues included decreased GnRH interpulse intervals, delayed vaginal opening and irregular reproductive cycles [36,37]. Although associations between EDCs and altered pubertal timing have been observed, current literature is limited and inconsistent, preventing conclusive evidence of a causal relationship. Analysis is complicated by the long latency period between EDC exposure and pubertal outcomes, coupled with mixed exposures throughout life and the challenge of distinguishing EDC effects from those of nutrition and adiposity [31]. Further prospective and longitudinal studies are needed to clarify the effects of EDCs on puberty and reproductive health, determine critical exposure periods, and elucidate the underlying mechanisms involved.
Adipokines (Leptin and Adiponectin)
The hormones that are secreted from the adipose tissue have a sexually dimorphic impact on the HPG axis. Adipokines, particularly leptin and adiponectin have been found to interact with the HPG axis by crossing the blood brain barrier (BBB) [38]. They are both secreted from adipose tissue and have a crucial role in maintaining the energy balance. Additionally, they also have different mechanisms by which they exert effects on the HPG axis in opposing aspects. In obesity, since there is an increased amount of adipose tissue there is also an increased level of adipokines [39]. There is a positive association between an early onset of puberty and increased body weight mostly in girls [40,41]. Obesity enhances leptin resistance that precipitates limitations in the effects of leptin, therefore a far greater amount of leptin is required to induce the feeling of satiety [42].
Leptin regulates reproductive function by stimulating the release of gonadotrophins via directly influencing pubertal gene expression [43]. In girls, elevated leptin levels signal excess energy storage, which is crucial for supporting the growth and development associated with puberty [44]. Typically, healthy girls see a rise in leptin before the GnRH pulsatile secretions, which may explain why in girls it causes early puberty. Moreover, leptin also affects the genes associated with initiating puberty. Studies have shown that leptin receptors (LEPR) on the hypothalamus influence KISS1 expression. So far, what has been proven is that a surplus of leptin is associated with an increased expression of KISS1 [45]. This increased expression acts as a signal, initiating pulsatile secretions of GnRH earlier than expected, implying that the increased leptin is a permissive factor in the early onset of puberty [45]. Cohesively, leptin stimulates the production of growth hormone (GH), LH, and FSH via hypothalamic stimulations, subsequently contributing to an earlier onset of puberty.
Furthermore, leptin has been found to affect LH more profoundly than the other hormones as individuals with higher levels of leptin are also associated with having greater levels of LH, this further supports a strong correlation between leptin and LH [46]. Higher levels of LH cause an increased production of oestrogen which also results in an increased release of leptin, feeding a cyclical trend [45]. This process is initiated when oestrogen stimulates the expression of the Ob gene in adipose tissue, piloting an increase in the production and secretion of leptin [47]. Leptin also interacts with both glutamate and GABA genes, albeit to a lesser extent in girls. Moreover, leptin enhances steroidogenesis gene expression, thereby facilitating oestrogen biosynthesis in girls [45]. In conclusion, leptin exerts multiple effects on pubertal girls, primarily by triggering or allowing an earlier than normal onset of puberty. Leptin stimulates KISS1 expression, increasing kisspeptin production, which in turn activates GnRH neurons. In leptin-deficient states (e.g., malnutrition, extreme leanness), KISS1 expression is suppressed, delaying puberty. In obese girls, excess leptin may lead to early KISS1 activation, causing precocious puberty. Furthermore, leptin enhances CYP19A1 (aromatase) expression, promoting oestrogen biosynthesis.
In boys, the relationship between leptin and puberty is more complex and less clearly understood however, boys with obesity enter puberty at a later age [44]. Usually, in boys of normal weight, leptin is reduced before the onset of puberty suggesting that a small drop may be required for pubertal initiation. The excess leptin seen in obese boys affects pubertal onset via genetic pathways and alterations in the hypothalamus [48]. In boys the LEPR that leptin binds onto in the hypothalamus has a substantially different effect on the HPG axis; it causes an increased expression of GABA, leading to a delay in the onset of puberty. This is because a rise in GABA prevents pulsatile releases of GnRH, which is needed for the initiation of puberty [49]. Therefore boys suffering from obesity also have elevated levels of GABA neurotransmitters, which can subsequently delay puberty. In addition to affecting GABA, Leptin also interacts with the expression of glutamate and KISS1. The rise in GABA causes a reduced expression of PA genes, in mechanisms that are not fully understood [50]. Leptin increases aromatase activity leading to higher oestrogen – to -testosterone ratio that may explain the delayed puberty seen in obese boys [51,52]. The effect of leptin on the timing of puberty is represented on Figure 1.
Another adipokine of interest, adiponectin, also plays a role in the onset of puberty. When adiponectin binds to its receptors AdipoR1 and AdipoR2 which are found in the hypothalamus, it exerts an inhibitory effect on the release of kisspeptin facilitating the progression of puberty. This in turn inhibits the release of GnRH delaying the onset of puberty. Analysis has shown that individuals with central obesity experience lower levels of adiponectin in mechanisms that are not fully understood [53]. This might be a result of increased adiponectin resistance via the reduced expression of AdipoR2 receptor [54]. This reduction is secondary to the decrease of adipose tissue that is brought about as a result of; increased insulin resistance, increased secretion of inflammatory cytokines, and dysfunction of adipose tissue. These factors contribute to a high leptin to adiponectin (L/A) ratio in the systemic circulation. A high L/A ratio disrupt the initiation of puberty differently between boys and girls. Adiponectin levels decline significantly upon the onset of puberty, with boys experiencing it far more than girls [55]. Thus when obesity coincides with puberty, it further alters adiponectin levels, which in turn affects pubertal timing.
Obesity is associated with a low-grade chronic inflammatory state, characterized by elevation of pro-inflammatory cytokines, including but not limited to interleuking-6 (IL-6) and tumor necrosis factor alpha (TNFα). This alteration reduced the inhibitory regulation of pubertal initiation, therefore resulting in an earlier onset of puberty for girls [51]. Further, when girls start puberty with an increased BMI, their adiponectin levels are reduced to a further extent. Adiponectin affects puberty by interacting with PA genes, causing a reduction in the bioavailability of kisspeptin to bind on the GPR54 receptors on the hypothalamus. It should be noted, that although its levels are inversely correlated with circulating LH levels, the mechanisms behind this are yet to be fully understood [56,57]. However, what can be concluded based on the interactions that adiponectin has with the HPG axis, is that it does ultimately disrupt the onset of puberty, albeit to a lesser extent. Furthermore, due to the lack of current literature on the matter, more research is needed to understand the specific mechanisms in which adiponectin affects pre-pubertal girls and causes earlier onset puberty.
Cohesively, adiponectin affects boys in a similar manner as it affects girls. Physiologically its levels drop prior to puberty in boys - the drop being much more rapid and extensive than in girls. Therefore, in boys, there is an even further inhibition of the HPG axis from adiponectin, causing delay of puberty. Adiponectin is inversely associated with androgen levels because it increases the levels of sex hormone binding globulin (SHBG), hence decreasing free testosterone [58]. Contrary to girls, the reduction in adiponectin causes delay of puberty as it has a detrimental role in maintaining insulin sensitivity and testosterone production. These correlations guide us in comprehending the mechanisms by which adiponectin affects puberty in obese boys. To elaborate, the rising levels of androgens which start from adrenarche cause a decrease in the release of adiponectin in boys from a younger age than girls [56]. Adiponectin is also involved in the suppression of the secretion of both LH and FSH, influencing gonadal activation. The mechanisms described above result in delayed testicular maturation, concluding to a delay of the onset of puberty. This statistic alone allows for the conclusion that obesity causes earlier onset puberty in boys, presenting a conflicting dilemma. Puberty is a multifactorial process, and studies have shown that obesity causes delay in puberty in boys. The need for further literature and research to be conducted so as to truly grasp the effects that puberty and adiponectin have in boys.
In summary Adiponectin suppresses KISS1 gene expression, which can delay the onset of puberty. In obesity, low adiponectin reduces this inhibition, potentially leading to earlier KISS1 activation and precocious puberty in girls. Adiponectin receptors (AdipoR1, AdipoR2) are expressed in the hypothalamus, where they can influence the secretion GnRH. Lower adiponectin levels, typically observed before puberty, may help release the inhibition on KISS1, allowing puberty to progress. Higher adiponectin levels are often associated with delayed puberty, particularly in conditions like low body fat or chronic illness. The decline of Adiponectin levels seen in girls promotes GnRH and LH/FSH release and subsequent oestrogen production. In boys Adiponectin may modulate testosterone production by influencing Leydig cell function in the testes.
Insulin
Puberty by itself can be characterized by a state of physiological insulin resistance and a reduction in the insulin sensitivity. This means that even in average weight, non-obese, girls and boys there is a change in the amounts of circulating insulin [59]. Obesity is associated with insulin resistance that has a sexually dimorphic effect on the onset of puberty [60].
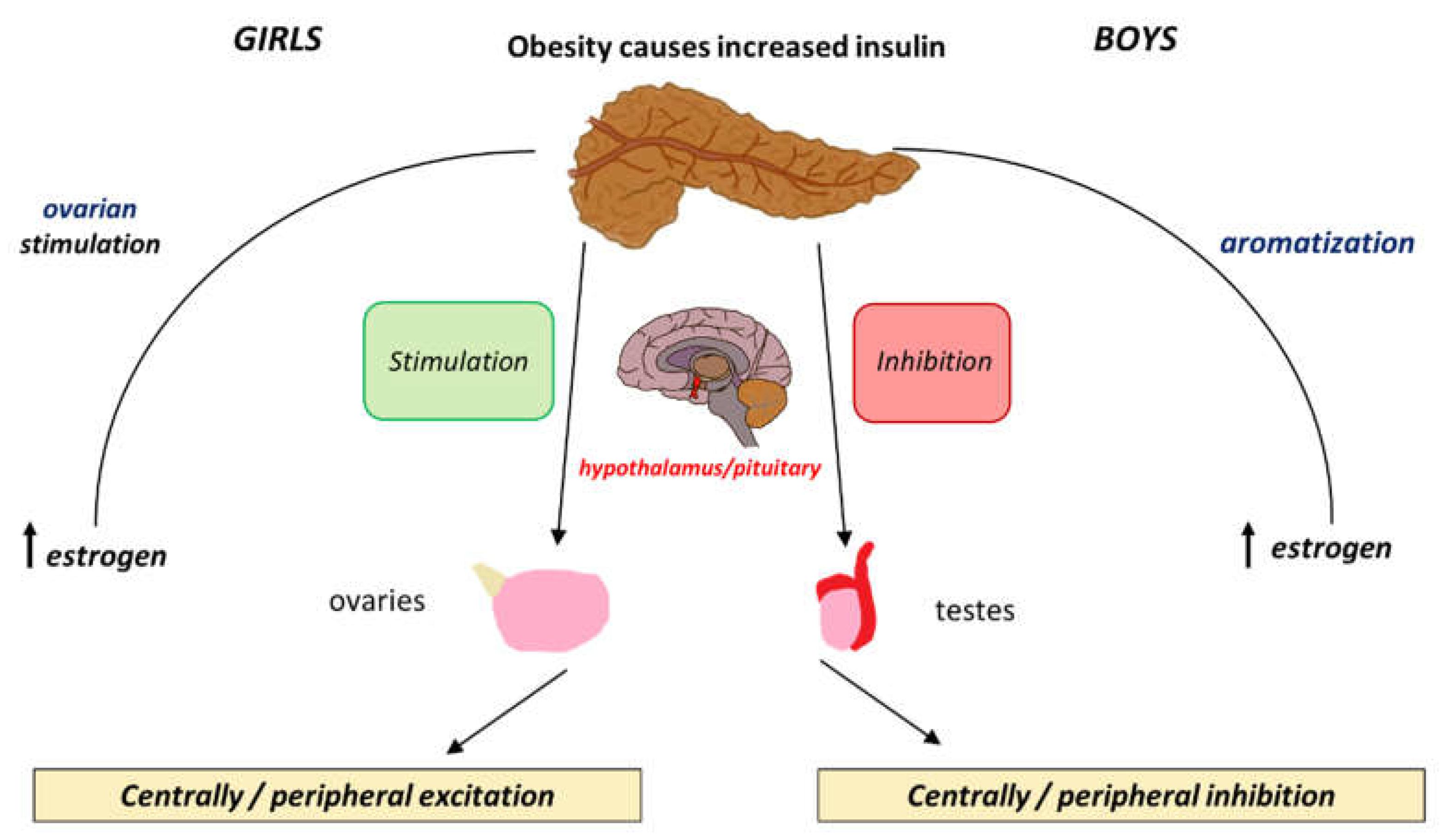
The effects of insulin levels on the HPG axis are similar to those of leptin. Insulin and leptin are both involved in energy homeostasis and metabolism and their relationship becomes particularly relevant during puberty. Insulin promotes leptin production in adipose tissue. Insulin exerts its influence by binding to Insulin receptors (IR) on the hypothalamus and the pituitary facilitating the expression of the KISS1 gene and promoting the pulsatile release of GnRH [61]. Insulin also enhances the production of sex steroids especially oestrogens which are crucial for pubertal changes [62]. The increased level of oestrogens that are synthesized in the adipose tissue seen in obese girls further contributes to the early onset of puberty. The positive feedback mechanisms by which insulin affects both the central and peripheral systems is visually illustrated in Figure 2.
The relationship of insulin and puberty in boys is more complex. Obese boys may experience delayed puberty potentially due to suppression of the HPG axis by insulin resistance. Obese boys have low levels of free testosterone and increased levels of oestrogens due to increased aromatization that acts as an inhibitory mechanism for gonadotropin release. Although there is no clear delay, obese boys tend to show a delay in pubertal onset [63,64,65].
SGA, Obesity and Premature Gonadarche
Small for gestational age (SGA) is defined as a birth weight which is at or below the 3rd percentile in weight, or less than two standard deviations for gestational age [66]. SGA is typically a consequence of inadequate nutrition during foetal development. Over the past decades, studies have demonstrated an association between low birth weight and obesity. SGA infants have a higher risk of developing additional health conditions and experience disrupted growth compared to infants with normal birth weights. Notably, obesity is one of the most well-documented long-term complications of SGA babies, the precise timing of its onset being during childhood years that may affect pubertal onset [67]. This can be attributed to compensatory growth patterns, where SGA children undergo rapid growth to lessen the developmental disparity with their peers. However, this rapid growth can lead to an overcorrection, resulting in a rapid and accelerated weight gain, and therefore obesity [68].
The underlying biological mechanism by which SGA leads to obesity is still unclear [69]. To date, it is believed that the restricted development of SGA newborns alters insulin sensitivity, thus increasing the risk of childhood obesity [70]. This disruption of development is also believed to affect the timing of puberty, leading to an earlier onset in both genders. SGA babies, especially girls experience early adrenarche where the elevated circulating adrenal androgens theoretically trigger the HPG axis [70]. The impact of SGA and obesity on puberty is almost identical in boys and girls as both genders tend to experience earlier onset of puberty. SGA itself is a known risk factor for early-onset obesity that effects pubertal onset. Strategies targeting the management of SGA babies and preventing excess weight gain during childhood years are needed.
Conclusion
This overview investigates the intricate relationship between childhood obesity and the timing of pubertal onset. It provides a comprehensive analysis of how endocrine changes associated with obesity, specifically alterations in adipokines and insulin, influences the timing of puberty in both genders. Adiponectin modulates the expression of puberty-related genes, primarily by regulating KISS1/GnRH signalling and metabolic pathways. Its decline during puberty is necessary for normal pubertal onset, with sex-specific variations in its effects on hormonal pathways. In obese girls, low adiponectin may contribute to earlier puberty, as reduced inhibition on KISS1/GnRH signalling can accelerate pubertal onset. In obese boys, low adiponectin is linked to delayed puberty due to its role in maintaining insulin sensitivity and testosterone production. In addition, excess adiposity increases oestrogen conversion (via aromatase activity), which may suppress testosterone and delay pubertal progression. In obese girls, excess leptin may lead to early KISS1 activation, causing precocious puberty. Furthermore, leptin enhances CYP19A1 expression, promoting oestrogen biosynthesis leading to higher oestrogen – to -testosterone ratio that may explain the delayed puberty seen in obese boys. It can be therefore concluded that adipokines that they are abundant in obese children affect the timing of puberty via central and peripheral metabolic pathways. The understanding of these mechanisms is still unclear hence the need for further research on this topic is highlighted. Addressing and preventing obesity in children could potentially mitigate these alterations in pubertal timing.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
References
- Breehl, L. Physiology, puberty, StatPearls [Internet]. 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK534827/ (accessed on 29 May 2024).
- Vijayakumar, N.; et al. Puberty and the human brain: Insights Into Adolescent Development, Neuroscience and biobehavioral reviews. 2018. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6234123/ (accessed on 29 May 2024).
- Manotas, M.C.; et al. Genetic and epigenetic control of puberty, Sexual development: genetics, molecular biology, evolution, endocrinology, embryology, and pathology of sex determination and differentiation. 2022. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8820423/ (accessed on 29 May 2024).
- Joseph, D.N.; Whirledge, S. Stress and the HPA axis: Balancing homeostasis and fertility, International journal of molecular sciences. 2017. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5666903/#:~:text=As%20part%20of%20the%20physiological,reproductive%20competence%20of%20an%20organism (accessed on 31 May 2024).
- Abreu, A.P.; Kaiser, U.B.; Pubertal development and regulation, The lancet. Diabetes & endocrinology. 2016. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5192018/ (accessed on 31 May 2024).
- Cunningham, S.A.; et al. Changes in the incidence of childhood obesity, Pediatrics. 2022. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9879733/ (accessed on 01 June 2024).
- Eckert-Lind, C.; et al. Worldwide secular trends in age at pubertal onset assessed by breast development among girls: A systematic review and meta-analysis, JAMA pediatrics. 2020. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7042934/ (accessed on 31 May 2024).
- Wohlfahrt-Veje, C.; et al. ‘Pubarche and Gonadarche Onset and Progression Are Differently Associated With Birth Weight and Infancy Growth Patterns’, Journal of the Endocrine Society, 5(8). 2021. Available online: https://doi.org/10.1210/jendso/bvab108 (accessed on 01 June 2024). [CrossRef]
- Beccuti, Guglielmo, and Lucia Ghizzoni. “Normal and Abnormal Puberty.” PubMed, MDText.com, Inc. 2000. Available online: www.ncbi.nlm.nih.gov/books/NBK279024/ (accessed on 01 June 2024).
- Yang, D.; et al. Initiation of the hypothalamic-pituitary-gonadal axis in young girls undergoing central precocious puberty exerts remodeling effects on the prefrontal cortex, Frontiers in psychiatry. 2019. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6524415/ (accessed on 01 June 2024).
- Author links open overlay panelMatteo Spaziani a b et al. Hypothalamo-pituitary axis and puberty, Molecular and Cellular Endocrinology. 2020. Available online: https://www.sciencedirect.com/science/article/pii/S0303720720303968?via%3Dihub (accessed on 01 June 2024).
- Blair, J.A.; et al. Hypothalamic-pituitary-gonadal axis involvement in learning and memory and alzheimer’s disease: More than ‘just’ estrogen, Frontiers in endocrinology. 2015. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4373369/ (accessed on 01 June 2024).
- Guercio, G.; et al. Estrogens in human male gonadotropin secretion and testicular physiology from infancy to late puberty, Frontiers in endocrinology. 2020. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7051936/ (accessed on 01 June 2024).
- Shaw, N.D.; et al. Estrogen negative feedback on gonadotropin secretion: Evidence for a direct pituitary effect in women, The Journal of clinical endocrinology and metabolism. 2010. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2853991/ (accessed on 05 June 2024).
- Manotas, M.C.; et al. Genetic and epigenetic control of puberty, Karger Publishers. 2021. Available online: https://karger.com/sxd/article/16/1/1/829199/Genetic-and-Epigenetic-Control-of-Puberty (accessed on 05 June 2024).
- Chan, Y.-M.; et al. Using kisspeptin to predict pubertal outcomes for youth with pubertal delay, The Journal of clinical endocrinology and metabolism. 2020. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7282711/ (accessed on 05 June 2024).
- Christoforidis, A.; Skordis, N.; Fanis, P.; Dimitriadou, M.; Sevastidou, M.; Phelan, M.M.; Neocleous, V.; Phylactou, L.A. A novel MKRN3 nonsense mutation causing familial central precocious puberty. Endocrine. 2017, 56, 446–449. Available online: https://doi.org/10.1007/s12020-017-1232-6 (accessed on 5 June 2024). [PubMed]
- Howard, S.R. The Genetic Basis of Delayed Puberty. Frontiers in Endocrinology. 2019, 10. Available online: https://doi.org/10.3389/fendo.2019.00423 (accessed on 5 June 2024). [CrossRef]
- Faienza, M.F.; Urbano, F.; Moscogiuri, L.A.; Chiarito, M.; De Santis, S.; Giordano, P. . Genetic, epigenetic and enviromental influencing factors on the regulation of precocious and delayed puberty. Frontiers in Endocrinology. 2022, 13, 1019468. Available online: https://doi.org/10.3389/fendo.2022.1019468 (accessed on 5 June 2024). [CrossRef] [PubMed]
- Pagani, S.; et al. MKRN3 and KISS1R mutations in precocious and early puberty, Italian journal of pediatrics. 2017. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7104496/ (accessed on 05 June 2024).
- Neocleous, V.; Shammas, C.; Phelan, M.M.; Nicolaou, S.; Phylactou, L.A.; Skordis, N. In silico analysis of a novelMKRN3missense mutation in familial central precocious puberty. Clinical Endocrinology 2015, 84, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Fanis, P.; Skordis, N.; Toumba, M.; Papaioannou, N.; Makris, A.; Kyriakou, A.; Neocleous, V.; Phylactou, L.A. Central Precocious Puberty Caused by Novel Mutations in the Promoter and 5′-UTR Region of the Imprinted MKRN3 Gene. Frontiers in Endocrinology 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Rosenfield, R.L. Normal and premature adrenarche, Endocrine reviews. 2021. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8599200/ (accessed on 06 June 2024).
- Sheng, J.A.; et al. The hypothalamic-pituitary-adrenal axis: Development, programming actions of hormones, and maternal-fetal interactions, Frontiers. 2020. Available online: https://www.frontiersin.org/articles/10.3389/fnbeh.2020.601939/full (accessed on 06 June 2024).
- Heck, A.L.; Handa, R.J. Sex differences in the hypothalamic-pituitary-adrenal axis’ response to stress: An important role for gonadal hormones, Neuropsychopharmacology: official publication of the American College of Neuropsychopharmacology. 2019. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6235871/ (accessed on 07 June 2024).
- Stephens, M.A.C.; et al. Hypothalamic-pituitary-adrenal axis response to acute psychosocial stress: Effects of biological sex and circulating sex hormones, Psychoneuroendocrinology. 2016. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4788592/ (accessed on 07 June 2024).
- Obesity and overweight [Internet]. World Health Organization. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 10 August 2024).
- Prevalence of obesity [Internet]. World Obesity Federation. Available online: https://www.worldobesity.org/about/about-obesity/prevalence-of-obesity (accessed on 10 August 2024).
- Tiwari, A.; Daley, S.F.; Balasundaram, P. Obesity in Pediatric Patients [Updated 2023 Mar 8]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2024 Jan-. Available online: https://www.ncbi.nlm.nih.gov/books/NBK570626/.
- Nicolaou, M.; Toumba, M.; Kythreotis, A.; Daher, H.; Skordis, N. Obesogens in adolescence: Challenging Aspects and Prevention Strategies. Children 2024, 11, 602. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Jung, H.W.; Lee, Y.J.; Lee, Y.A. Early-life exposure to endocrine-disrupting chemicals and pubertal development in girls. Annals of Pediatric Endocrinology & Metabolism 2019, 24, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; et al. EDC-2: The endocrine society’s second scientific statement on endocrine-disrupting chemicals. Endocrine Reviews 2015, 36. [Google Scholar] [CrossRef] [PubMed]
- Nah, W.H.; Park, M.J.; Gye, M.C. Effects of early prepubertal exposure to bisphenol A on the onset of puberty, ovarian weights, and estrous cycle in female mice. Clinical and Experimental Reproductive Medicine 2011, 38, 75. [Google Scholar] [CrossRef] [PubMed]
- Tassinari, R.; Tait, S.; Busani, L.; Martinelli, A.; Narciso, L.; Valeri, M.; et al. Metabolic, reproductive and thyroid effects of Bis(2-Ethylhexyl) Phthalate (DEHP) orally administered to male and female juvenile rats at dose levels derived from children biomonitoring study. Toxicology 2021, 449, 152653. [Google Scholar] [CrossRef] [PubMed]
- Shalitin, S.; Gat-Yablonski, G. Associations of obesity with linear growth and puberty. Hormone Research in Paediatrics 2021, 95, 120–36. [Google Scholar] [CrossRef] [PubMed]
- López-Rodríguez, D.; Aylwin, C.F.; Delli, V.; Sevrin, E.; Campanile, M.; Martin, M.; et al. Multi- and transgenerational outcomes of an exposure to a mixture of endocrine-disrupting chemicals (edcs) on puberty and maternal behavior in the female rat. Environmental Health Perspectives 2021, 129. [Google Scholar] [CrossRef] [PubMed]
- Lucaccioni, L.; Trevisani, V.; Marrozzini, L.; Bertoncelli, N.; Predieri, B.; Lugli, L.; et al. Endocrine-disrupting chemicals and their effects during female puberty: A review of current evidence. International Journal of Molecular Sciences 2020, 21, 2078. [Google Scholar] [CrossRef] [PubMed]
- Adamska-Patruno, E.; et al. The relationship between the leptin/ghrelin ratio and meals with various macronutrient contents in men with different nutritional status: A randomized crossover study, Nutrition journal. 2018. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6309055/ (accessed on 10 June 2024).
- Picó, C.; Palou, M.; Pomar, C.A.; Rodríguez, A.M.; Palou, A. Leptin as a key regulator of the adipose organ. Reviews in Endocrine and Metabolic Disorders 2021, 23. Available online: https://doi.org/10.1007/s11154-021-09687-5 (accessed on 10 June 2024). [CrossRef]
- Li, W.; Liu, Q.; Deng, X.; Chen, Y.; Liu, S.; Story, M. Association between Obesity and Puberty Timing: A Systematic Review and Meta-Analysis. International Journal of Environmental Research and Public Health 2017, 14, 1266. [Google Scholar] [CrossRef] [PubMed]
- dDunger, D.B.; Lynn Ahmed, M.; Ong, K.K. Effects of obesity on growth and puberty. Best Practice & Research Clinical Endocrinology & Metabolism 2005, 19, 375–390. Available online: https://doi.org/10.1016/j.beem.2005.04.005 (accessed on 8 April 2025). [CrossRef]
- Myers, M.G.; Leibel, R.L.; Seeley, R.J.; Schwartz, M.W. . Obesity and leptin resistance: distinguishing cause from effect. Trends in Endocrinology & Metabolism 2010, 21, 643–651. Available online: https://doi.org/10.1016/j.tem.2010.08.002 (accessed on 8 April 2025). [CrossRef]
- Cheng, H.L.; Sainsbury, A.; Garden, F.; Sritharan, M.; Paxton, K.; Luscombe, G.; Hawke, C.; Steinbeck, K. Ghrelin and Peptide YY Change During Puberty: Relationships With Adolescent Growth, Development, and Obesity. The Journal of Clinical Endocrinology & Metabolism 2018, 103, 2851–2860. Available online: https://doi.org/10.1210/jc.2017-01825 (accessed on 10 June 2024). [CrossRef]
- Huang, A.; Reinehr, T.; Roth, C.L. Connections between obesity and puberty: Invited by Manuel Tena-Sempere, Cordoba, Current opinion in endocrine and metabolic research. 2020. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7543977/#:~:text=Longitudinal%20studies%20clearly%20demonstrate%20that,puberty%20and%20age%20of%20menarche (accessed on 10 June 2024).
- Shi, L.; Jiang, Z.; Zhang, L. Childhood obesity and central precocious puberty, Frontiers in endocrinology. 2022. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9716129/ (accessed on 10 June 2024).
- Odle, A.K.; et al. Leptin regulation of Gonadotrope gonadotropin-releasing hormone receptors as a metabolic checkpoint and gateway to reproductive competence, Frontiers in endocrinology. 2018. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5760501/ (accessed on 10 June 2024).
- d Machinal-Quélin, F.; Dieudonné, M.-N.; Pecquery, R.; Leneveu, M.-C.; Giudicelli, Y. Direct In Vitro Effects of Androgens and Estrogens on ob Gene Expression and Leptin Secretion in Human Adipose Tissue. Endocrine 2002, 18, 179–184. Available online: https://doi.org/10.1385/endo:18:2:179 (accessed on 8 April 2025). [CrossRef] [PubMed]
- Kaplowitz, P. Delayed puberty in obese boys: Comparison with constitutional delayed puberty and response to testosterone therapy. The Journal of Pediatrics. 1998, 133, 745–749. Available online: https://doi.org/10.1016/s0022-3476(98)70144-1 (accessed on 13 June 2024). [CrossRef]
- Zuure, W.A.; Roberts, A.L.; Quennell, J.H.; Anderson, G.M. Leptin Signaling in GABA Neurons, But Not Glutamate Neurons, Is Required for Reproductive Function. The Journal of Neuroscience. 2013, 33, 17874–17883. Available online: https://doi.org/10.1523/jneurosci.2278-13.2013 (accessed on 13 June 2024). [CrossRef]
- Ahima, R.S. No Kiss1ng by leptin during puberty? Journal of Clinical Investigation. 2011, 121, 34–36. Available online: https://doi.org/10.1172/jci45813 (accessed on 13 June 2024). [CrossRef]
- Nieuwenhuis, D.; Pujol-Gualdo, N.; Arnoldussen, I.A.C.; Kiliaan, A.J. Adipokines: A gear shift in puberty. Obesity Reviews. 2020, 21. Available online: https://doi.org/10.1111/obr.13005 (accessed on 18 June 2024). [CrossRef]
- Song, Y.; Kong, Y.; Xie, X.; Wang, Y.; Wang, N. Association between precocious puberty and obesity risk in children: a systematic review and meta-analysis. Frontiers in Pediatrics. 2023, 11. Available online: https://doi.org/10.3389/fped.2023.1226933 (accessed on 13 June 2024). [CrossRef]
- Ahl, S.; Guenther, M.; Zhao, S.; James, R.; Marks, J.; Szabo, A.; Kidambi, S. Adiponectin Levels Differentiate Metabolically Healthy vs Unhealthy Among Obese and Nonobese White Individuals. The Journal of Clinical Endocrinology & Metabolism. 2015, 100, 4172–4180. Available online: https://doi.org/10.1210/jc.2015-2765 (accessed on 08 April 2024). [CrossRef]
- Atilla Engin 2017. Adiponectin-Resistance in Obesity. Advances in experimental medicine and biology, pp.415–441. Available online: https://doi.org/10.1007/978-3-319-48382-5_18 (accessed on 08 April 2024). [CrossRef]
- Wu, Y.; Zhong, L.; Li, G.; Han, L.; Fu, J.; Li, Y.; Li, L.; Zhang, Q.; Guo, Y.; Xiao, X.; Qi, L.; Li, M.; Gao, S.; Willi Steven, M. Puberty Status Modifies the Effects of Genetic Variants, Lifestyle Factors and Their Interactions on Adiponectin: The BCAMS Study. Frontiers in Endocrinology. 2021, 12, 737459. Available online: https://doi.org/10.3389/fendo.2021.737459 (accessed on 18 June 2024). [CrossRef]
- Xu, L.; Li, M.; Yin, J.; Cheng, H.; Yu, M.; Zhao, X.; Xiao, X.; Mi, J. Change of Body Composition and Adipokines and Their Relationship with Insulin Resistance across Pubertal Development in Obese and Nonobese Chinese Children: The BCAMS Study. International Journal of Endocrinology. 2012, 2012, 389108. Available online: https://doi.org/10.1155/2012/389108 (accessed on 20 June 2024). [CrossRef]
- Barbe, A.; Bongrani, A.; Mellouk, N.; Estienne, A.; Kurowska, P.; Grandhaye, J.; Elfassy, Y.; Levy, R.; Rak, A.; Froment, P.; Dupont, J. Mechanisms of Adiponectin Action in Fertility: An Overview from Gametogenesis to Gestation in Humans and Animal Models in Normal and Pathological Conditions. International Journal of Molecular Sciences. 2019, 20, 1526. Available online: https://doi.org/10.3390/ijms20071526 (accessed on 20 June 2024). [CrossRef]
- Ohman-Hanson, R.A.; Cree-Green, M.; Kelsey, M.M.; Bessesen, D.H.; Sharp, T.A.; Pyle, L.; Pereira, R.I.; Nadeau, K.J. Ethnic and Sex Differences in Adiponectin: From Childhood to Adulthood. The Journal of Clinical Endocrinology & Metabolism. 2016, 101, 4808–4815. Available online: https://doi.org/10.1210/jc.2016-1137 (accessed on 20 July 2024). [CrossRef]
- Kelsey, M.M.; Zeitler, P.S. Insulin Resistance of Puberty. Current Diabetes Reports. 2016, 16. Available online: https://doi.org/10.1007/s11892-016-0751-5 (accessed on 24 June 2024). [CrossRef]
- Gołacki, J.; Matuszek, M.; Matyjaszek-Matuszek, B. Link between Insulin Resistance and Obesity—From Diagnosis to Treatment. Diagnostics. 2022, 12, 1681. Available online: https://doi.org/10.3390/diagnostics12071681 (accessed on 25 June 2024). [CrossRef]
- Saleh, F.L.; Joshi, A.A.; Tal, A.; Xu, P.; Hens, J.R.; Wong, S.L.; Flannery, C.A. Hyperinsulinemia induces early and dyssynchronous puberty in lean female mice. Journal of Endocrinology. 2022, 254, 121–135. Available online: https://doi.org/10.1530/joe-21-0447 (accessed on 27 June 2024). [CrossRef]
- Sliwowska, J.H.; Fergani, C.; Gawałek, M.; Skowronska, B.; Fichna, P.; Lehman, M.N. Insulin: Its role in the central control of reproduction. Physiology & Behavior. 2014, 133, 197–206. Available online: https://doi.org/10.1016/j.physbeh.2014.05.021 (accessed on 28 June 2024). [CrossRef]
- Burt Solorzano, C.M.; McCartney, C.R. Obesity and the pubertal transition in girls and boys. REPRODUCTION. 2010, 140, 399–410. Available online: https://doi.org/10.1530/rep-10-0119 (accessed on 28 June 2024). [CrossRef]
- Mauras, N.; Ross, J.; Mericq, V. Management of Growth Disorders in Puberty: GH, GnRHa, and Aromatase Inhibitors: A Clinical Review. Endocrine Reviews. 2022. Available online: https://doi.org/10.1210/endrev/bnac014 (accessed on 30 June 2024). [CrossRef]
- Nassar, G.N.; Raudales, F.; Leslie, S.W. Physiology, Testosterone [online]. PubMed. 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK526128/ (accessed on 04 July 2024).
- Osuchukwu, O.O.; Reed, D.J. Small for Gestational Age [online]. PubMed. 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK563247/ (accessed on 20 July 2024).
- Hong, Y.H.; Chung, S. Small for gestational age and obesity related comorbidities. Annals of Pediatric Endocrinology & Metabolism. 2018, 23, 4–8. Available online: https://doi.org/10.6065/apem.2018.23.1.4 (accessed on 20 July 2024). [CrossRef]
- Wu, D.; Zhu, J.; Wang, X.; Shi, H.; Huo, Y.; Liu, M.; Sun, F.; Lan, H.; Guo, C.; Liu, H.; Li, T.; Jiang, L.; Hu, X.; Li, T.; Xu, J.; Yao, G.; Zhu, G.; Yu, G.; Chen, J. Rapid BMI Increases and Persistent Obesity in Small-for-Gestational-Age Infants. Frontiers in Pediatrics. 2021a, 9, 625853. Available online: https://doi.org/10.3389/fped.2021.625853 (accessed on 22 July 2024). [CrossRef]
- Nam, H.-K.; Lee, K.-H. Small for gestational age and obesity: epidemiology and general risks. Annals of Pediatric Endocrinology & Metabolism. 2018, 23, 9–13. Available online: https://doi.org/10.6065/apem.2018.23.1.9 (accessed on 22 July 2024). [CrossRef]
- Verkauskiene, R.; Petraitiene, I.; Albertsson Wikland, K. Puberty in children born small for gestational age. Hormone Research in Paediatrics. 2013, 80, 69–77. Available online: https://doi.org/10.1159/000353759 (accessed on 25 July 2024). [CrossRef]
Figure 1.
The release of leptin from adipocyte tissue has different effects on each gender. In boys it causes increased expression of GABA thus decrease expression of GnRH, and in girls it causes an increase in KISSR expression.
Figure 1.
The release of leptin from adipocyte tissue has different effects on each gender. In boys it causes increased expression of GABA thus decrease expression of GnRH, and in girls it causes an increase in KISSR expression.
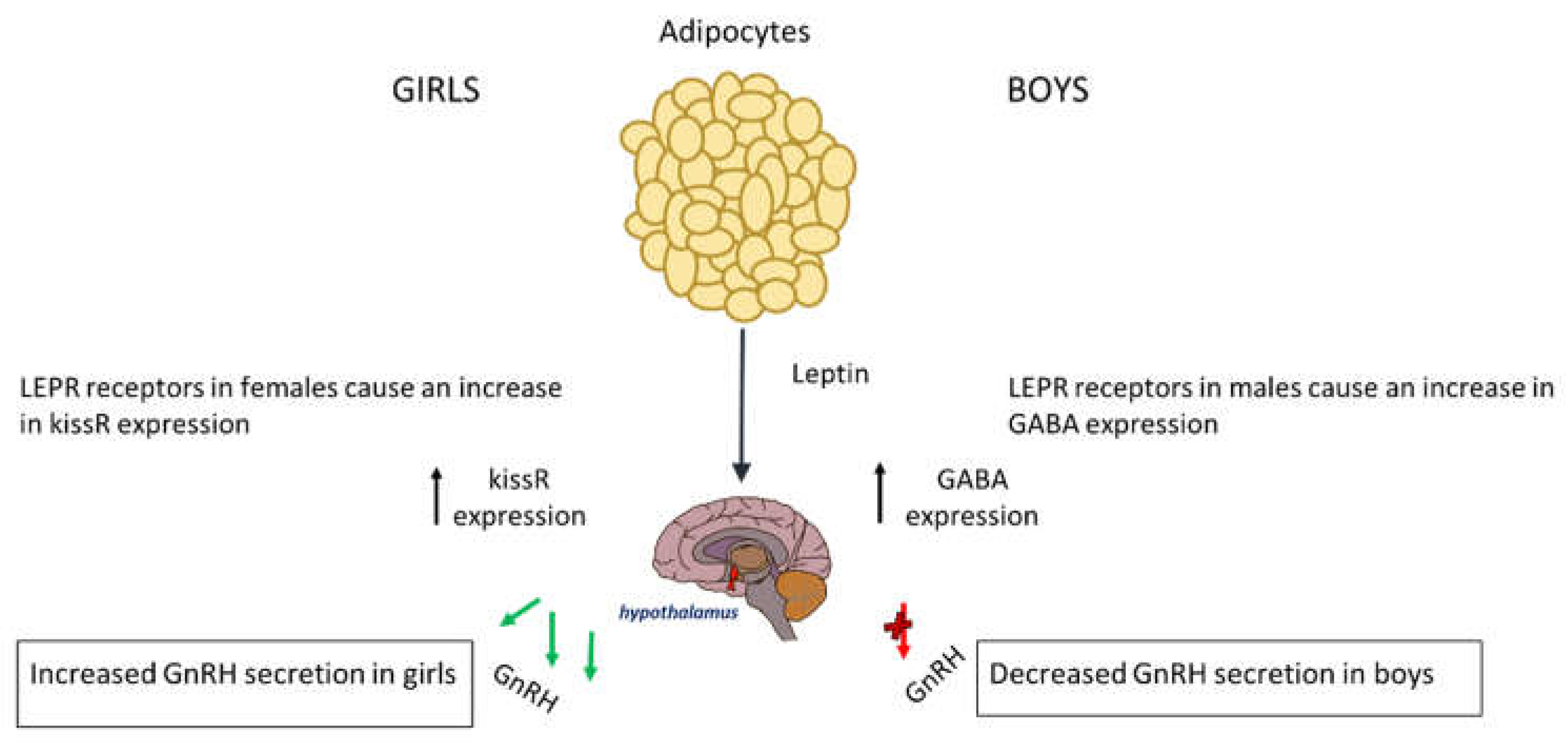
Figure 2.
The differential effects that insulin exerts peripherally and centrally and how these cause positive and negative feedback loop in boys and girls.
Figure 2.
The differential effects that insulin exerts peripherally and centrally and how these cause positive and negative feedback loop in boys and girls.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



